
A TCP domain transcription factor controls flower type ... - PNAS
6
A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence Suvi K. Broholm*, Sari Ta ¨ htiharju*, Roosa A. E. Laitinen † , Victor A. Albert ‡ , Teemu H. Teeri*, and Paula Elomaa* § *Department of Applied Biology, P.O. Box 27, University of Helsinki, Helsinki FIN-00014, Finland; † Max Planck Institute for Developmental Biology, Spemannstrasse 37-39, D-72076 Tu ¨ bingen, Germany; and ‡ Department of Biological Sciences, State University of New York, Buffalo, NY 14260 Edited by M. T. Clegg, University of California, Irvine, CA, and approved April 4, 2008 (received for review February 11, 2008) Several key processes in plant development are regulated by TCP transcription factors. CYCLOIDEA-like (CYC-like) TCP domain pro- teins have been shown to control flower symmetry in distantly related plant lineages. Gerbera hybrida, a member of one of the largest clades of angiosperms, the sunflower family (Asteraceae), is an interesting model for developmental studies because its elaborate inflorescence comprises different types of flowers that have specialized structures and functions. The morphological dif- ferentiation of flower types involves gradual changes in flower size and symmetry that follow the radial organization of the densely packed inflorescence. Differences in the degree of petal fusion further define the distinct shapes of the Gerbera flower types. To study the role of TCP transcription factors during speci- fication of this complex inflorescence organization, we character- ized the CYC-like homolog GhCYC2 from Gerbera. The expression of GhCYC2 follows a gradient along the radial axis of the inflo- rescence. GhCYC2 is expressed in the marginal, bilaterally symmet- rical ray flowers but not in the centermost disk flowers, which are nearly radially symmetrical and have significantly less fused petals. Overexpression of GhCYC2 causes disk flowers to obtain morphol- ogies similar to ray flowers. Both expression patterns and trans- genic phenotypes suggest that GhCYC2 is involved in differentia- tion among Gerbera flower types, providing the first molecular evidence that CYC-like TCP factors take part in defining the com- plex inflorescence structure of the Asteraceae, a major determinant of the family’s evolutionary success. CYCLOIDEA flower development evo-devo organ fusion T he enormous variety of flowers makes them fascinating targets for comparative developmental genetic studies. Spe- cies of the sunf lower family (Asteraceae), such as the cut-f lower crop Gerbera hybrida, present a unique and challenging benefit: more than one f loral phenotype can be analyzed within the same genotype. A typical Asteraceae inflorescence consists of mor- phologically and functionally differentiated flowers packed into a condensed inflorescence (the capitulum) that resembles (as a pseudanthium) a single large flower. This inflorescence com- plexity, not shared by other model species used for flower developmental research such as Arabidopsis, Antirrhinum, or Petunia, has apparently proved to be evolutionarily successful because Asteraceae is one of the largest families of flowering plants with 23,000 species (1). Gerbera bears three f lower types (ray, trans, and disk) that are morphologically similar during their early development. One of the most prominent developmental differences, which becomes established early in f loral ontogeny, is a gradual change of f lower symmetry that follows the radial organization of the capitulum (2). The marginal ray flowers are bilaterally symmetrical (zygo- morphic). Three of the five petals are fused ventrally to form the large, elongate ligule, whereas the remaining two (dorsal) petals remain rudimentary. Disk f lowers at the center of the capitulum have shorter and less fused petals, and they eventually become radially symmetrical (actinomorphic) with separate petals at the inf lorescence apex. Trans f lowers, which occupy an intermediate radial position between ray and disk flowers, are strongly zygomorphic, yet smaller in size in comparison with ray f lowers. Stamen development also differs among individual f lower types (3). In the female ray and trans flowers, stamen development arrests, resulting in rudimentary staminodia (4), whereas in the disk f lowers, anthers develop fully and form a postgenitally fused structure that covers the carpel. Despite continued efforts, the molecular basis for f lower type differentiation in Asteraceae remains unclear. Microarray com- parison of developing Gerbera ray and disk f lower primordia has identified a number of genes that are differentially expressed in individual flower types (3). We have shown that many MADS box genes encoding known regulators of flower organ develop- ment are expressed gradientially along the capitulum radius. While this may suggest that different MADS protein complexes participate in developmental regulation of individual flower types, transgenic experiments show that such complexes are not independently sufficient (4, 5). Indeed, classical genetics has shown that the presence or absence of ray f lowers in Asteraceae capitula appears to be under control of one or two major and several modifier genes (reviewed in ref. 6). Berti et al. (7) have characterized two sunflower mutants in which flower type identity (and consequently symmetry) has changed. In the chrysanthemoides (chry) mutant, all f lowers in the inf lorescences are ray-like (zygomorphic), whereas in the tubular ray flower (turf ) mutant, all f lower types resemble radially symmetrical disk flowers. In Senecio, capitulum organization is principally con- trolled by the RAY locus, with radiate inflorescences (ray plus disk) dominant over discoid (disk only) (8 –10). For both Senecio and sunflower, hypotheses have been constructed whereby the RAY (6, 11), CHRY and TURF loci (7) could encode homologs of the Antirrhinum floral symmetry gene CYCLOIDEA (12). However, these intriguing postulates remain unevaluated by molecular evidence. CYCLOIDEA-like genes have been reported to be involved in flower symmetry regulation in various plant species. CYCLOIDEA (CYC) of Antirrhinum was the first gene isolated, and thereafter the most extensively studied (12, 13). It belongs to the plant-specific gene family encoding TCP transcription factors, which share a conserved basic helix–loop–helix TCP Author contributions: S.K.B., R.A.E.L., T.H.T., and P.E. designed research; S.K.B., S.T., R.A.E.L., V.A.A., and P.E. performed research; S.K.B., V.A.A., and P.E. analyzed data; and S.K.B., S.T., V.A.A., T.H.T., and P.E. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Data deposition: The sequences reported in this paper have been deposited in the GenBank database [accession nos. EU429302 (GhCYC1), EU429303 (GhCYC2), EU429304 (GhCYC3), and EU429305 (GhCYC4)]. § To whom correspondence should be addressed. E-mail: paula.elomaa@helsinki.fi. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0801359105/DCSupplemental. © 2008 by The National Academy of Sciences of the USA www.pnas.orgcgidoi10.1073pnas.0801359105 PNAS July 1, 2008 vol. 105 no. 26 9117–9122 PLANT BIOLOGY Downloaded by guest on February 13, 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of A TCP domain transcription factor controls flower type ... - PNAS

A TCP domain transcription factor controls flowertype specification along the radial axis of theGerbera (Asteraceae) inflorescenceSuvi K. Broholm*, Sari Tahtiharju*, Roosa A. E. Laitinen†, Victor A. Albert‡, Teemu H. Teeri*, and Paula Elomaa*§
*Department of Applied Biology, P.O. Box 27, University of Helsinki, Helsinki FIN-00014, Finland; †Max Planck Institute for Developmental Biology,Spemannstrasse 37-39, D-72076 Tubingen, Germany; and ‡Department of Biological Sciences, State University of New York, Buffalo, NY 14260
Edited by M. T. Clegg, University of California, Irvine, CA, and approved April 4, 2008 (received for review February 11, 2008)
Several key processes in plant development are regulated by TCPtranscription factors. CYCLOIDEA-like (CYC-like) TCP domain pro-teins have been shown to control flower symmetry in distantlyrelated plant lineages. Gerbera hybrida, a member of one of thelargest clades of angiosperms, the sunflower family (Asteraceae),is an interesting model for developmental studies because itselaborate inflorescence comprises different types of flowers thathave specialized structures and functions. The morphological dif-ferentiation of flower types involves gradual changes in flowersize and symmetry that follow the radial organization of thedensely packed inflorescence. Differences in the degree of petalfusion further define the distinct shapes of the Gerbera flowertypes. To study the role of TCP transcription factors during speci-fication of this complex inflorescence organization, we character-ized the CYC-like homolog GhCYC2 from Gerbera. The expressionof GhCYC2 follows a gradient along the radial axis of the inflo-rescence. GhCYC2 is expressed in the marginal, bilaterally symmet-rical ray flowers but not in the centermost disk flowers, which arenearly radially symmetrical and have significantly less fused petals.Overexpression of GhCYC2 causes disk flowers to obtain morphol-ogies similar to ray flowers. Both expression patterns and trans-genic phenotypes suggest that GhCYC2 is involved in differentia-tion among Gerbera flower types, providing the first molecularevidence that CYC-like TCP factors take part in defining the com-plex inflorescence structure of the Asteraceae, a major determinantof the family’s evolutionary success.
CYCLOIDEA � flower development � evo-devo � organ fusion
The enormous variety of flowers makes them fascinatingtargets for comparative developmental genetic studies. Spe-
cies of the sunflower family (Asteraceae), such as the cut-f lowercrop Gerbera hybrida, present a unique and challenging benefit:more than one floral phenotype can be analyzed within the samegenotype. A typical Asteraceae inflorescence consists of mor-phologically and functionally differentiated flowers packed intoa condensed inflorescence (the capitulum) that resembles (as apseudanthium) a single large flower. This inflorescence com-plexity, not shared by other model species used for flowerdevelopmental research such as Arabidopsis, Antirrhinum, orPetunia, has apparently proved to be evolutionarily successfulbecause Asteraceae is one of the largest families of floweringplants with �23,000 species (1).
Gerbera bears three flower types (ray, trans, and disk) that aremorphologically similar during their early development. One ofthe most prominent developmental differences, which becomesestablished early in floral ontogeny, is a gradual change of flowersymmetry that follows the radial organization of the capitulum(2). The marginal ray flowers are bilaterally symmetrical (zygo-morphic). Three of the five petals are fused ventrally to form thelarge, elongate ligule, whereas the remaining two (dorsal) petalsremain rudimentary. Disk flowers at the center of the capitulumhave shorter and less fused petals, and they eventually becomeradially symmetrical (actinomorphic) with separate petals at the
inflorescence apex. Trans flowers, which occupy an intermediateradial position between ray and disk flowers, are stronglyzygomorphic, yet smaller in size in comparison with ray flowers.Stamen development also differs among individual f lower types(3). In the female ray and trans flowers, stamen developmentarrests, resulting in rudimentary staminodia (4), whereas in thedisk flowers, anthers develop fully and form a postgenitally fusedstructure that covers the carpel.
Despite continued efforts, the molecular basis for flower typedifferentiation in Asteraceae remains unclear. Microarray com-parison of developing Gerbera ray and disk flower primordia hasidentified a number of genes that are differentially expressed inindividual f lower types (3). We have shown that many MADSbox genes encoding known regulators of flower organ develop-ment are expressed gradientially along the capitulum radius.While this may suggest that different MADS protein complexesparticipate in developmental regulation of individual f lowertypes, transgenic experiments show that such complexes are notindependently sufficient (4, 5). Indeed, classical genetics hasshown that the presence or absence of ray flowers in Asteraceaecapitula appears to be under control of one or two major andseveral modifier genes (reviewed in ref. 6). Berti et al. (7) havecharacterized two sunflower mutants in which flower typeidentity (and consequently symmetry) has changed. In thechrysanthemoides (chry) mutant, all f lowers in the inflorescencesare ray-like (zygomorphic), whereas in the tubular ray flower(turf ) mutant, all f lower types resemble radially symmetrical diskflowers. In Senecio, capitulum organization is principally con-trolled by the RAY locus, with radiate inflorescences (ray plusdisk) dominant over discoid (disk only) (8–10). For both Senecioand sunflower, hypotheses have been constructed whereby theRAY (6, 11), CHRY and TURF loci (7) could encode homologsof the Antirrhinum f loral symmetry gene CYCLOIDEA (12).However, these intriguing postulates remain unevaluated bymolecular evidence.
CYCLOIDEA-like genes have been reported to be involvedin f lower symmetry regulation in various plant species.CYCLOIDEA (CYC) of Antirrhinum was the first gene isolated,and thereafter the most extensively studied (12, 13). It belongsto the plant-specific gene family encoding TCP transcriptionfactors, which share a conserved basic helix–loop–helix TCP
Author contributions: S.K.B., R.A.E.L., T.H.T., and P.E. designed research; S.K.B., S.T.,R.A.E.L., V.A.A., and P.E. performed research; S.K.B., V.A.A., and P.E. analyzed data; andS.K.B., S.T., V.A.A., T.H.T., and P.E. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The sequences reported in this paper have been deposited in the GenBankdatabase [accession nos. EU429302 (GhCYC1), EU429303 (GhCYC2), EU429304 (GhCYC3),and EU429305 (GhCYC4)].
§To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0801359105/DCSupplemental.
© 2008 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0801359105 PNAS � July 1, 2008 � vol. 105 � no. 26 � 9117–9122
PLA
NT
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2

domain. The name TCP derives from the three founding mem-bers of the family, TEOSINTE BRANCHED1 (TB1) of maize,CYC, and PROLIFERATING CELL FACTOR (PCF) of rice,all of which control meristem growth by affecting cell prolifer-ation (14). Phylogenetic analysis based on the TCP domain hasuncovered two subfamilies; PCF proteins form one clade (classI) whereas CYC/TB1 and CINCINNATA (CIN) group togetherto form the class II TCP proteins (15). The CYC/TB1 group isalso called the ECE clade, within which three subclades, CYC1,-2, and -3, have been identified in core eudicots (16). TB1 hasplayed a major role in the evolution of maize from its wildancestor teosinte. It acts as a repressor of growth by suppressingaxillary branching, and thus promotes apical dominance indomesticated maize (17). TB1 homologs have also been shownto prevent the growth of axillary buds in rice and Arabidopsis (15,18). The two TCP genes CYC and DICHOTOMA (DICH) inAntirrhinum regulate flower symmetry by modulating cell-division related gene expression (12–14, 19). The wild-typeflowers of Antirrhinum have an axis of dorsoventral asymmetrywith distinct dorsal, lateral, and ventral organ types. The expres-sion of CYC and DICH is restricted to the dorsal domain of theflowers to establish bilateral petal symmetry and to arrest thedevelopment of the dorsalmost stamen (12, 13). CIN arreststhe growth of Antirrhinum leaf margins but also affects differ-entiation of epidermal cells and growth of petal lobes througheffects on cell proliferation (20).
In Gerbera, specification of flower types involves differentialdevelopment of petals and stamens, probably through distinctcell division and elongation as well as organ fusion events. TCPtranscription factors have previously been connected with allthree of these processes, making them good candidates forregulators of the distinct morphologies of Gerbera f lower types.We have isolated several TCP transcription factor genes ascandidates from Gerbera. We show that one of these proteins,GhCYC2, has a symmetry function different from that of classicCYC-like genes. Instead of regulating the dorsoventral symmetryof individual f lowers, GhCYC2 participates in the control of theidentity and radial extent of flower types in Gerbera inflores-cences. Moreover, GhCYC2 plays an important and unique rolein organ fusion that further differentiates Gerbera f lower types.
ResultsIsolation of Gerbera CYC Homologs. To study the role of TCPdomain transcription factors during Gerbera f lower develop-ment, cDNA clones encoding these proteins were isolated fromyoung developing Gerbera inflorescences. The inferred aminoacid sequences of the four cDNAs, GhCYC1, GhCYC2, Gh-CYC3, and GhCYC4 (Gerbera hybrida CYCLOIDEA-like 1, 2, 3,and 4), exhibited the conserved TCP and R domains typical forthe CYC/TB1 subfamily. Low stringency hybridization of Ger-bera genomic DNA with a full-length GhCYC2 probe suggestedthe presence of a small gene family [supporting information (SI)Fig. S1]. Although we cannot exclude possible allelism, thisinference was supported by isolation of 4 distinct sequencefragments (data not shown) in addition to the four full-lengthsequences presented in this study. The alignment of full-lengthGhCYC cDNAs (Fig. S2) showed that the inferred GhCYC1amino acid sequence is markedly different from the GhCYC2–4sequences outside the highly conserved regions. Motif Scan (21)analysis of GhCYC amino acid sequences predicted that Gh-CYC2, 3 and 4 have putative bipartite nuclear localizationsignals, whereas GhCYC1 does not. Furthermore, GhCYC1 waspredicted by TargetP (22) to be targeted to the chloroplast,suggesting a different role for this Gerbera TCP.
Phylogenetic Analysis of Gerbera TCP Domain Factors. We per-formed phylogenetic analysis on a selected set of class II TCPfactors from various plant species (Table S1) to explore the
evolutionary relationships of the Gerbera TCP genes. Nucleotidesequences encoding the conserved TCP and R domains werealigned, and parsimony and maximum likelihood analyses wereperformed on these data and the inferred amino acid alignment.Robustness of results was assessed using jacknife and bootstrapresampling, respectively (Fig. S3). In most analyses, CIN-likegenes from Antirrhinum and Arabidopsis grouped together withsupport. CYC-like genes from Lamiales, Fabales, and Asteraceaewere divided into separate, well supported groups. The greatergenetic similarity within rather than among these groups sug-gests that many functionally important duplication events in theECE gene subfamily occurred after the divergence of the Lamia-les, Fabales, and Asteraceae lineages, respectively. The results ofHowarth and Donoghue (16) are similar in this respect in thatgroupings within subclades CYC1, CYC2, and CYC3 are largelyconsistent with organismal phylogeny. Given this concurrence,we tested the phylogenetic distribution of our gene data whencombined with many of the less-complete Dipsacales and Aste-rales sequences published by the former authors, and all wellsupported relationships were consistent with the results reportedhere (data not shown). According to our trees and their under-lying nucleotide alignment, GhCYC1 is deviant from the otherthree GhCYC genes, which are quite similar to each other (Fig.S2). In addition to its sister gene HpTCP from sunflower, in themost-parsimonious and maximum likelihood trees based onnucleotide data, GhCYC1 groups (without statistical support)with Arabidopsis AtBRC1 and Lotus LjCYC5 (data not shown).AtBRC1 and AtBRC2 have been shown to be functionallyrelated to maize TB1 (15). However, in our analyses, support forparticular gene groupings with TB1 is lacking.
The Expression of GhCYC2 Follows the Zonal Organization of theCapitulum. RNA-blot analysis showed that GhCYC2 is expressedonly in floral tissues. We could not detect expression in vege-tative organs such as leaves, roots, or floral stems (scapes).Expression was strongest in young inflorescences, and in petals,whereas weaker expression was observed in carpels (stigma andstyle), in the ovary and in involucral bracts (the leaf-like struc-tures that tightly surround the reproductive centers of capitula)(Fig. 1). We have recently demonstrated that at f loral primor-dium stage 3, the ray and disk flowers of Gerbera are morpho-logically similar to each other, whereas at later stages (after stage4) morphological differentiation of the flower types is visible(Fig. 2 B–E) (3). Therefore, a more detailed analysis of GhCYC2expression in young developing capitula was performed onseparately dissected ray and disk flower primordia using bothRNA-blots (Fig. 2 A) and quantitative PCR (data not shown).The results affirmed that at early developmental stages 3 and 5,GhCYC2 is expressed only in ray flower primordia and not indisk flower primordia located in the center of the capitulum. Inlater developmental stages 6 and 7, GhCYC2 expression was stilllacking from the centermost disk flowers, although present inouter disk flowers and in ray flowers (Fig. 2 A).
Fig. 1. Expression of GhCYC2 in various Gerbera tissues. GhCYC2 showedstrongest expression in young inflorescences, petals, and carpel (stigma andstyle). Weaker expression was also seen in bracts and ovary. The lower blotshows ethidium bromide-stained ribosomal RNA bands to control for RNAloading.
9118 � www.pnas.org�cgi�doi�10.1073�pnas.0801359105 Broholm et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2
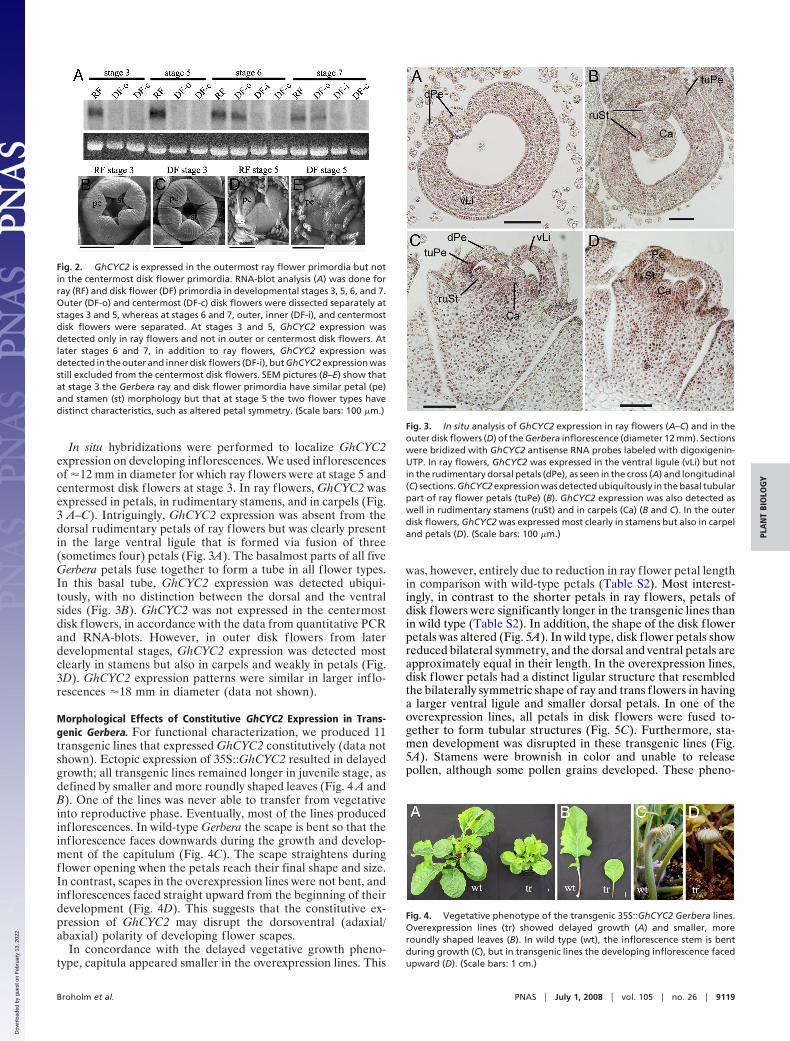
In situ hybridizations were performed to localize GhCYC2expression on developing inflorescences. We used inflorescencesof �12 mm in diameter for which ray flowers were at stage 5 andcentermost disk flowers at stage 3. In ray flowers, GhCYC2 wasexpressed in petals, in rudimentary stamens, and in carpels (Fig.3 A–C). Intriguingly, GhCYC2 expression was absent from thedorsal rudimentary petals of ray flowers but was clearly presentin the large ventral ligule that is formed via fusion of three(sometimes four) petals (Fig. 3A). The basalmost parts of all fiveGerbera petals fuse together to form a tube in all f lower types.In this basal tube, GhCYC2 expression was detected ubiqui-tously, with no distinction between the dorsal and the ventralsides (Fig. 3B). GhCYC2 was not expressed in the centermostdisk flowers, in accordance with the data from quantitative PCRand RNA-blots. However, in outer disk flowers from laterdevelopmental stages, GhCYC2 expression was detected mostclearly in stamens but also in carpels and weakly in petals (Fig.3D). GhCYC2 expression patterns were similar in larger inflo-rescences �18 mm in diameter (data not shown).
Morphological Effects of Constitutive GhCYC2 Expression in Trans-genic Gerbera. For functional characterization, we produced 11transgenic lines that expressed GhCYC2 constitutively (data notshown). Ectopic expression of 35S::GhCYC2 resulted in delayedgrowth; all transgenic lines remained longer in juvenile stage, asdefined by smaller and more roundly shaped leaves (Fig. 4 A andB). One of the lines was never able to transfer from vegetativeinto reproductive phase. Eventually, most of the lines producedinflorescences. In wild-type Gerbera the scape is bent so that theinflorescence faces downwards during the growth and develop-ment of the capitulum (Fig. 4C). The scape straightens duringflower opening when the petals reach their final shape and size.In contrast, scapes in the overexpression lines were not bent, andinflorescences faced straight upward from the beginning of theirdevelopment (Fig. 4D). This suggests that the constitutive ex-pression of GhCYC2 may disrupt the dorsoventral (adaxial/abaxial) polarity of developing flower scapes.
In concordance with the delayed vegetative growth pheno-type, capitula appeared smaller in the overexpression lines. This
was, however, entirely due to reduction in ray flower petal lengthin comparison with wild-type petals (Table S2). Most interest-ingly, in contrast to the shorter petals in ray flowers, petals ofdisk flowers were significantly longer in the transgenic lines thanin wild type (Table S2). In addition, the shape of the disk flowerpetals was altered (Fig. 5A). In wild type, disk flower petals showreduced bilateral symmetry, and the dorsal and ventral petals areapproximately equal in their length. In the overexpression lines,disk flower petals had a distinct ligular structure that resembledthe bilaterally symmetric shape of ray and trans flowers in havinga larger ventral ligule and smaller dorsal petals. In one of theoverexpression lines, all petals in disk flowers were fused to-gether to form tubular structures (Fig. 5C). Furthermore, sta-men development was disrupted in these transgenic lines (Fig.5A). Stamens were brownish in color and unable to releasepollen, although some pollen grains developed. These pheno-
Fig. 2. GhCYC2 is expressed in the outermost ray flower primordia but notin the centermost disk flower primordia. RNA-blot analysis (A) was done forray (RF) and disk flower (DF) primordia in developmental stages 3, 5, 6, and 7.Outer (DF-o) and centermost (DF-c) disk flowers were dissected separately atstages 3 and 5, whereas at stages 6 and 7, outer, inner (DF-i), and centermostdisk flowers were separated. At stages 3 and 5, GhCYC2 expression wasdetected only in ray flowers and not in outer or centermost disk flowers. Atlater stages 6 and 7, in addition to ray flowers, GhCYC2 expression wasdetected in the outer and inner disk flowers (DF-i), but GhCYC2 expression wasstill excluded from the centermost disk flowers. SEM pictures (B–E) show thatat stage 3 the Gerbera ray and disk flower primordia have similar petal (pe)and stamen (st) morphology but that at stage 5 the two flower types havedistinct characteristics, such as altered petal symmetry. (Scale bars: 100 �m.)
Fig. 3. In situ analysis of GhCYC2 expression in ray flowers (A–C) and in theouter disk flowers (D) of the Gerbera inflorescence (diameter 12 mm). Sectionswere bridized with GhCYC2 antisense RNA probes labeled with digoxigenin-UTP. In ray flowers, GhCYC2 was expressed in the ventral ligule (vLi) but notin the rudimentary dorsal petals (dPe), as seen in the cross (A) and longitudinal(C) sections. GhCYC2 expression was detected ubiquitously in the basal tubularpart of ray flower petals (tuPe) (B). GhCYC2 expression was also detected aswell in rudimentary stamens (ruSt) and in carpels (Ca) (B and C). In the outerdisk flowers, GhCYC2 was expressed most clearly in stamens but also in carpeland petals (D). (Scale bars: 100 �m.)
Fig. 4. Vegetative phenotype of the transgenic 35S::GhCYC2 Gerbera lines.Overexpression lines (tr) showed delayed growth (A) and smaller, moreroundly shaped leaves (B). In wild type (wt), the inflorescence stem is bentduring growth (C), but in transgenic lines the developing inflorescence facedupward (D). (Scale bars: 1 cm.)
Broholm et al. PNAS � July 1, 2008 � vol. 105 � no. 26 � 9119
PLA
NT
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2

typic changes imply that constitutive expression of GhCYC2caused disk flowers to obtain ray-like flower characteristics suchas enlarged more markedly fused petals and disrupted stamendevelopment.
Effects of Reduced GhCYC2 Expression in Transgenic Gerbera. Weobtained one transgenic line with cosuppressed GhCYC2 ex-pression. In this line, the expression of GhCYC2 could not bedetected in RNA blots (Fig. S4). Transformation of the GhCYC2cDNA in antisense orientation resulted in an additional line thatshowed similar phenotypic changes as the cosuppression line.This line accumulated very high amounts of antisense transcriptsnearly equal to the size of the endogenous transcript correspond-ing to GhCYC2. Hence, we could not convincingly detectwhether the endogenous GhCYC2 expression was silenced (Fig.S4). However, in both lines, we observed flower-type specificalterations. The length of trans flower petals was shorter than inwild type (Fig. 6 and Table S2). In other flower types the lengthsof petals did not differ significantly from wild type. We alsodetected occasional splitting of the trans flower ligules into fiveto eight separate petals (data not shown).
DiscussionGerbera TCP Factors Form a Clade Independent from AntirrhinumCYC/DICH and Functionally Characterized Genes from Legumes andCrucifers. Patterns of homology apparent in our phylogeneticanalyses of TCP genes are important for understanding GhCYC2function. Similar to what has been observed previously forCYC-like genes in legumes (23), GhCYC2 represents one prod-uct of several duplication events that occurred within Asteraceae(or perhaps within the larger clade that contains both Asteraceaeand Dipsacales). Floral zygomorphy in Lotus and Antirrhinumappears to have evolved independently, because members of twodistinct gene clades perform dorsalizing functions within discretelineages of plants, the rosids and asterids (respectively), whichare very likely to be primitively actinomorphic (see, e.g., ref. 24).Likewise, bilateral symmetry in the Arabidopsis relative Iberis,which is distinguished by expression timing effects in its CYC-likegene IaTCP1, may also represent an independent development
in yet another rosid lineage (25). Given this diversity of dem-onstrated CYC-like protein function in eudicots, it is not sur-prising that duplication and divergence has led to further noveltyin Gerbera, and perhaps also within other members of theAsteraceae CYC-like gene clade.
GhCYC2 Affects Growth in Transgenic Gerbera. In general, TCPdomain regulatory proteins have been shown to affect keydevelopmental processes to produce morphological novelties inplants. They function primarily by modulating cell growth andproliferation. We have shown that Gerbera GhCYC2 also reg-ulates growth. Overexpression of GhCYC2 in transgenic Gerberaresulted in slower vegetative growth and reduced inflorescencesize. Importantly, the effect of GhCYC2 overexpression on petalgrowth varied among the different flower types. In ray flowers,petals were shorter (thus the entire inflorescence appearedsmaller), whereas in disk flowers petals were longer comparedwith wild type. GhCYC2 is therefore able to either reduce orpromote growth depending on the site of its expression incapitula. Such opposite effects on growth have been describedfor other TCP transcription factors but not at the inflorescencelevel. For example in Antirrhinum, CYC reduces growth in thedorsalmost stamen but promotes the growth of dorsal petals(12). Heterologous expression of CYC in Arabidopsis causesreduced vegetative growth but increased growth of petals. Thedecreased leaf size in these plants was due to reduction in bothcell proliferation and cell expansion, whereas enlarged petalsresulted from increased cell expansion (26). Furthermore, theclass II TCP factor CIN reduces growth in Antirrhinum leaves butpromotes the growth of petals (20).
GhCYC2 Expression Is Primarily Ventral and Correlates with OrganFusion in Different Flower Types. We found differences between theexpression patterns of GhCYC2 and other CYC/TB1-like genesknown to be involved in the regulation of floral symmetry.CYC-like genes are expressed in the dorsal parts of the flower toestablish bilateral symmetry in the Lamiales and Fabales speciesstudied so far (12–14, 23, 27, 28). In Gerbera ray flowers, theexpression of GhCYC2 was specifically excluded from the dorsalrudimentary petals, suggesting functional deviation from Lamia-les and Fabaceae. A similarly ventral expression pattern was
Fig. 5. The effect of GhCYC2 overexpression on Gerbera disk flowers.Transgenic disk flowers (tr df) had a clearly distinct phenotype compared withwild-type disk flowers (wt df). (A) Disk flower petals were longer and petalshad a more pronounced ligular structure. Development of disk flower stamens(St) was disrupted. At the same developmental stages, wild-type disk flowersrelease pollen (B), but no pollen was seen in the 35S::GhCYC2 lines (A and C).In one of the overexpression lines, disk flower petals were fused together toform tubular structures (C). (Scale bar: 0.5 cm.)
Fig. 6. The effect of suppressed GhCYC2 expression on Gerbera inflores-cence phenotype. In nontransformed Terra Regina (A and B), trans flowers (wttf) are longer than in the transgenic lines (A and C). Only the length of thedistal ligule and not the basal petal tube differs between wild-type andtransgenic trans flowers (tr tf) (A). (Scale bar: 0.5 cm.)
9120 � www.pnas.org�cgi�doi�10.1073�pnas.0801359105 Broholm et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2

documented for a CYC3-subclade gene from Lonicera (Dip-sacales), whereas a CYC2-subclade gene expressed dorsally (16).However, preliminary phylogenetic analyses including some ofthese authors’ shorter sequences suggest that GhCYC2 belongsto the CYC2 subclade (data not shown). If so, ECE CYC-likegenes have undergone functional divergence within subclades aswell as within organismal groups.
Intriguingly, GhCYC2 expression correlated with organ fusionin petals. In ray flowers, GhCYC2 expression was uniform in thebasal petal tube and in the fused ligule but was lacking from therudimentary dorsal petals, which remain unfused. At the inflo-rescence level, GhCYC2 was specifically expressed in the bilat-erally symmetrical ray flowers with their more markedly fusedpetals, whereas the lack of GhCYC2 expression in central diskflowers correlated with radial symmetry and decreased petalfusion. Overexpression phenotypes indicate that GhCYC2 notonly promotes the growth of ligules in disk flowers but is alsocapable of generating fully tubular disk flower corollas. Indeed,all three Gerbera f lower types can become fully tubular, asobserved when a CYC-like gene from Senecio is overexpressedin Gerbera (P.E. and S.K.B., unpublished results). We alsoobserved frequent splitting of trans flower petals in the trans-genic Gerbera line with reduced GhCYC2 expression, which inaddition to the expression pattern of GhCYC2, provides furthersupport for the role of Asteraceae TCP factors in promotingorgan fusion.
Previous studies have connected TCP domain transcriptionfactors with regulation of organ boundaries. In Arabidopsis,CIN-like proteins negatively regulate boundary-specific NACdomain transcription factors (CUP-SHAPED COTYLEDON(CUC) 1, 2, 3) (29). Overexpression of CIN-like genes causedsuppression of CUC expression, which resulted in fusion ofcotyledons (29, 30). In Antirrhinum, the PCF-like TCP proteinTIC (TCP-Interacting with CUP) has been shown to interactwith a NAC domain protein that regulates organ fusion (31).Thus, NAC domain proteins have a clear connection with TCPdomain proteins, both as downstream targets and as interactionpartners.
GhCYC2 Plays a Major Role in Differentiating Gerbera Flower Types.The experimental evidence shown here suggests that GhCYC2is involved in differentiation between Gerbera f lower types.GhCYC2 is specifically expressed in the marginal ray flowerprimordia. Based on transgenic phenotypes, the absence ofGhCYC2 expression in the central disk flower primordia appearsto be crucial for proper flower type specification. Overexpres-sion of GhCYC2 causes disk flowers to obtain characteristicstypical for ray flowers, including enlarged ventral petals anddisrupted stamen development. These Gerbera lines resemblethe semidominant chry mutant of sunflower (7), in which diskflowers also obtain ray-f lower like traits. In chry mutants, theform of disk flower petals has shifted toward bilateral symmetry,and stamens are smaller with disrupted pollen production. Thesimilarities of the two phenotypes in these closely related speciesmakes it tempting to speculate that the semidominant chrymutant could result from overexpression of a GhCYC2 orthologin sunflower.
In Gerbera, some cultivars resemble the transgenic GhCYC2lines and the sunflower chry mutants. The ‘‘crested’’ trait ischaracterized by enlarged trans flowers or enlarged trans anddisk flowers that are male-sterile and defined by a single locuscontaining three alleles with semidominant relationships(crested � CRESTED � CRESTEDD) (32). In our overexpres-sion lines, the increase in petal length and reduction in stamenfunctionality in disk flowers implies similarity with the crestedtrait. Moreover, the recessive homozygotes of crested are char-acterized by short trans flowers resembling the transgenic linewith suppressed GhCYC2 expression. However, because Gerbera
cultivars showing the crested phenotype do not display GhCYC2expression differences compared with semicrested cultivars(such as Terra Regina; used here) or cultivars having short transflowers (S.K.B., unpublished results), we postulate that GhCYC2is not sufficient for complete transformation of flower types.
The transgenic Gerbera line with suppressed GhCYC2 expres-sion did not show alterations in ray flower identity. This againimplies that GhCYC2 is not sufficient for regulating differenti-ation between the rayed and nonrayed form of Asteraceaeinflorescences but that it takes part in this process, most likely asa modifier gene. Genetic analyses performed so far have sug-gested that in addition to the major loci (RAY/turf/CHRY/CRESTED), an unknown number of modifier genes are alsoinvolved. The Gerbera GhCYC3 and GhCYC4 genes show ray-flower specific expression similar to GhCYC2 (S.T. and S.K.B.,unpublished results). Their expression was not reduced in thetransgenic lines with suppressed GhCYC2 expression. Therefore,the absence of altered ray flower phenotype in GhCYC2 sup-pressed lines might be due to redundant functions of theseclosely related genes. Functional analyses of GhCYC3 andGhCYC4 are currently underway.
Our data provides the first molecular evidence that CYC-likeTCP factors are involved in defining identities of different flowertypes within a single genotype. The expression of GhCYC2 showsclear radial specificity along the axis of the Gerbera capitulum.In previous work we showed that several MADS box genes alsodisplay radial expression differences among developing flowerprimordia (3). As such, it seems likely that both TCP and MADSdomain proteins participate in regulating the developmentalspecification of Gerbera f lower types through response to earlyacting signals at the inflorescence level. A gradient of a cellnonautonomously moving substance, such as a phytohormone ora transcription factor, could induce differentiation of the distinctf lower morphologies by affecting putative downstream targetssuch as TCP and MADS domain proteins.
ConclusionsAsteraceae (1), Fabales, and Orchidaceae (33) are the threelargest lineages of flowering plants, and each are defined bysuites of traits that have lent themselves to extensive evolutionarymodification. For Asteraceae, the capitulate inflorescence withits capacity to produce different flower types has been itsprincipal innovation. Through analysis of a CYC-like TCP-encoding gene in Gerbera, we have shown that gene duplicationand divergence, a frequent theme in the evolution of organismalnovelty (34–36), has been one major factor in establishing theuniqueness and success of the sunflower family.
Materials and MethodsPlant Material. Gerbera hybrida (Asteraceae) variety Terra Regina (TerraNigra) was grown under standard greenhouse conditions. The developmentalstages for inflorescences are described in ref. 37 and for the young developingfloral primordia in ref. 3.
Isolation of Gerbera TCP Domain Factors. Gerbera TCP domain factors wereamplified from cDNA prepared from young developing inflorescences ofGerbera by PCR using two different pairs of degenerate primers. The firstdegenerate primer pair was designed from the conserved TCP domain andcorresponded to the amino acid sequences KKDRHSKI and ERTKEK. Poly(A)-RNA (450 ng) was used as a template for cDNA synthesis (Boehringer first-strand cDNA kit). PCR conditions were the following: 94°C for 75 sec, 46°C for2 min, and 72°C for 3 min for 30 cycles. Two different fragments that showedhigh similarity with CYC-like genes were obtained by using this primer pair.For the second round of PCR screening, degenerate primers were designed byusing the CODEHOP strategy (38) and were optimized for codon usage.Primers corresponded to the amino acid sequences DLQDMLGFDK andARARARERTKEK. cDNA synthesis was performed from poly(A)-RNA by usingSuperScript III reverse transcriptase (Invitrogen). Phusion DNA polymerase wasused for PCR reactions (Finnzymes). The full-length cDNAs were amplified byusing the SMART RACE cDNA amplification kit (Clontech). The TCP mRNA
Broholm et al. PNAS � July 1, 2008 � vol. 105 � no. 26 � 9121
PLA
NT
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2

coding sequences have been deposited in the GenBank database (accessionnumbers EU429302–EU429305).
Phylogenetic Analyses. Phylogenetic analyses were performed on correspond-ing nucleotide and amino acid alignments for the TCP and R domains ofselected TCP factors. Two phylogeny reconstruction methods were used:parsimony and maximum likelihood (see SI Text for full details).
RNA Blot, in Situ Hybridizations, and Scanning Electron Microscopy (SEM). TotalRNA was isolated by using TRIzol reagent (Invitrogen). Ten micrograms oftotal RNA was loaded on RNA blots and hybridized by using 32P-labeled DNAprobes. For a gene-specific probe, a 253-bp fragment from the 3� end of thecDNA was used. The blots were washed with 0.5� SSC and 0.1% (wt/vol) SDSat 58°C. In situ hybridization analysis using full-length GhCYC2 probe wasperformed as in ref. 39, with the following exceptions: probe concentrationwas 0.7 �g/ml/kb and detection time was 40–45 h. SEM analysis of the Gerberaflower primordia was performed as described in ref. 5.
Plant Transformation and Analysis of Transgenic Lines. The full-length GhCYC2cDNA was transformed under the CaMV 35S promoter into Gerbera varietyTerra Regina as reported in refs. 39 and 40. Differences in the length of petalsbetween wild-type variety Terra Regina and transgenic lines were tested byusing the Mann–Whitney rank-sum test. Flowers were counted, and thelength of petals was measured from five inflorescences of each line. Onaverage, for ray flowers n � 63 per line, for trans flowers n � 213 per line, andfor disk flowers n � 310 per line.
ACKNOWLEDGMENTS. We thank E. Coen (John Innes Centre, Norwich, UK)for providing the Senecio squalidus cDNA (plasmid pJAM2176); the Elec-tron Microscopy Unit of the Institute of Biotechnology, University ofHelsinki, for providing laboratory facilities; Eija Takala and Anu Rokkanenfor excellent technical assistance; and Sanna Peltola for taking care of theplants in the greenhouse. This work was supported by Academy of FinlandGrant 115849 (to P.E.). S.K.B. is supported by the Viikki Graduate School inBiosciences.
1. Bremer K (1994) Asteraceae: Cladistics and Classification (Timber Press, Portland, OR).2. Teeri TH, et al. (2006) Floral developmental genetics of Gerbera (Asteraceae). Devel-
opmental Genetics of the Flower, Advances in Botanical Research, eds Soltis DE,Leebens-Mack JH Soltis PS (Elsevier, New York), pp 324–351.
3. Laitinen RA, Broholm S, Albert VA, Teeri TH, Elomaa P (2006) Patterns of MADS-boxgene expression mark flower-type development in Gerbera hybrida (Asteraceae). BMCPlant Biol 6:11.
4. Kotilainen M, et al. (2000) GRCD1, an AGL2-like MADS box gene, participates in the Cfunction during stamen development in Gerbera hybrida. Plant Cell 12:1893–1902.
5. Uimari A, et al. (2004) Integration of reproductive meristem fates by a SEPALLATA-likeMADS-box gene. Proc Natl Acad Sci USA 101:15817–15822.
6. Gillies ACM, Cubas P, Coen ES, Abbott RJ (2002) Making rays in the Asteraceae: Geneticsand evolution of radiate versus discoid flower heads. Developmental Genetics andPlant Evolution, eds Cronk QCB, Bateman RM Hawkins JA (Taylor & Francis, London),pp 233–246.
7. Berti F, Fambrini M, Turi M, Bertini D, Pugliesi C (2005) Mutations of corolla symmetryaffect carpel and stamen development in Helianthus annuus. Can J Bot 83:1065–1072.
8. Abbott RJ, Ashton PA, Forbes DG (1992) Introgressive origin of the radiate groundselSenecio vulgaris var hibernicus Syme: Aat-3 evidence. Heredity 68:425–435.
9. Comes H (1998) Major gene effects during weed evolution: Phenotypic characterscosegregate with alleles at the ray floret locus in Senecio vulgaris L. (Asteraceae).J Hered 89:54–61.
10. Andersson S (2001) The genetic basis of floral variation in Senecio jacobaea (Aster-aceae). J Hered 92:409–414.
11. Coen ES, Nugent JM (1994) Evolution of flowers and inflorescences. DevelopmentSuppl:107–116.
12. Luo D, Carpenter R, Vincent C, Copsey L, Coen E (1996) Origin of floral asymmetry inAntirrhinum. Nature 383:794–799.
13. Luo D, et al. (1999) Control of organ asymmetry in flowers of Antirrhinum. Cell99:367–376.
14. Cubas P, Lauter N, Doebley J, Coen E (1999) The TCP domain: A motif found in proteinsregulating plant growth and development. Plant J 18:215–222.
15. Aguilar-Martinez JA, Poza-Carrion C, Cubas P (2007) Arabidopsis BRANCHED1 acts asan integrator of branching signals within axillary buds. Plant Cell 19:458–472.
16. Howarth DG, Donoghue MJ (2006) Phylogenetic analysis of the ‘‘ECE’’ (CYC/TB1) cladereveals duplications predating the core eudicots. Proc Natl Acad Sci USA 103:9101–9106.
17. Doebley J, Stec A, Hubbard L (1997) The evolution of apical dominance in maize. Nature386:485–488.
18. Takeda T, et al. (2003) The OsTB1 gene negatively regulates lateral branching in rice.Plant J 33:513–520.
19. Gaudin V, et al. (2000) The expression of D-cyclin genes defines distinct developmentalzones in snapdragon apical meristems and is locally regulated by the CYCLOIDEA gene.Plant Physiol 122:1137–1148.
20. Crawford BC, Nath U, Carpenter R, Coen ES (2004) CINCINNATA controls both celldifferentiation and growth in petal lobes and leaves of Antirrhinum. Plant Physiol135:244–253.
21. Pagni M, et al. (2004) MyHits: A new interactive resource for protein annotation anddomain identification. Nucleic Acids Res 32:W332–W335.
22. Emanuelsson O, Nielsen H, Brunak S, von Heijne G (2000) Predicting subcellularlocalization of proteins based on their N-terminal amino acid sequence. J Mol Biol300:1005–1016.
23. Feng X, et al. (2006) Control of petal shape and floral zygomorphy in Lotus japonicus.Proc Natl Acad Sci USA 103:4970–4975.
24. Donoghue MJ, Ree RH, Baum DA (1998) Phylogeny and the evolution of flowersymmetry in the Asteridae. Trends Plants Sci 3:311–317.
25. Busch A, Zachgo S (2007) Control of corolla monosymmetry in the Brassicaceae Iberisamara. Proc Natl Acad Sci USA 104:16714–16719.
26. Costa MM, Fox S, Hanna AI, Baxter C, Coen E (2005) Evolution of regulatory interactionscontrolling floral asymmetry. Development 132:5093–5101.
27. Hileman LC, Kramer EM, Baum DA (2003) Differential regulation of symmetry genesand the evolution of floral morphologies. Proc Natl Acad Sci USA 100:12814–12819.
28. Citerne HL, Pennington RT, Cronk QC (2006) An apparent reversal in floral symmetryin the legume Cadia is a homeotic transformation. Proc Natl Acad Sci USA 103:12017–12020.
29. Koyama T, Furutani M, Tasaka M, Ohme-Takagi M (2007) TCP transcription factorscontrol the morphology of shoot lateral organs via negative regulation of the expres-sion of boundary-specific genes in Arabidopsis. Plant Cell 19:473–484.
30. Palatnik JF, et al. (2003) Control of leaf morphogenesis by microRNAs. Nature 425:257–263.
31. Weir I, et al. (2004) CUPULIFORMIS establishes lateral organ boundaries in Antirrhi-num. Development 131:915–922.
32. Kloos WE, George CG, Sorge LK (2004) Inheritance of the flower types of Gerberahybrida. J Am Soc Hort Sci 129:802–810.
33. Endress PK (1994) Diversity and Evolutionary Biology of Flowers (Cambridge Univ Press,Cambridge, UK).
34. Hoekstra HE, Coyne JA (2007) The locus of evolution: evo devo and the genetics ofadaptation. Evolution 61:995–1016.
35. Roth C, et al. (2007) Evolution after gene duplication: Models, mechanisms, sequences,systems, and organisms. J Exp Zool B Mol Dev Evol 308:58–73.
36. Baxter CE, Costa MM, Coen ES (2007) Diversification and co-option of RAD-like genesin the evolution of floral asymmetry. Plant J 52:105–113.
37. Helariutta Y, Elomaa P, Kotilainen M, Seppanen P, Teeri TH (1993) Cloning of cDNAcoding for dihydroflavonol-4-reductase (DFR) and characterization of dfr expression inthe corollas of Gerbera hybrida var. Regina (Compositae). Plant Mol Biol 22:183–193.
38. Morant M, Hehn A, Werck-Reichhart D (2002) Conservation and diversity of genefamilies explored using the CODEHOP strategy in higher plants. BMC Plant Biol 2:7.
39. Elomaa P, et al. (2003) Activation of anthocyanin biosynthesis in Gerbera hybrida(Asteraceae) suggests conserved protein–protein and protein–promoter interactionsbetween the anciently diverged monocots and eudicots. Plant Physiol 133:1831–1842.
40. Elomaa P, Teeri TH (2001) Transgenic Gerbera. Biotechnology in Agriculture andForestry, Transgenic Crops III, ed YPS Bajaj (Springer, Berlin), pp 139–154.
9122 � www.pnas.org�cgi�doi�10.1073�pnas.0801359105 Broholm et al.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 13
, 202
2




















