
Subtype-specific alterations of the Wnt signaling pathway in breast cancer: clinical and prognostic...
11
Subtype-specific alterations of the Wnt signaling pathway in breast cancer: Clinical and prognostic significance Nupur Mukherjee, 1 Nilanjana Bhattacharya, 1 Neyaz Alam, 2 Anup Roy, 3 Susanta Roychoudhury 4 and Chinmay K. Panda 1,5 Departments of 1 Oncogene Regulation and 2 Surgical Oncology, Chittaranjan National Cancer Institute, Kolkata; 3 Department of Health and Family Welfare, Government of West Bengal, Swasthya Bhavan, Kolkata; 4 Molecular and Human Genetics Division, Indian Institute of Chemical Biology, Kolkata, India (Received June 19, 2011 ⁄ Revised October 5, 2011 ⁄ Accepted October 17, 2011 ⁄ Accepted manuscript online October 25, 2011 ⁄ Article first published online December 7, 2011) The aim of the study is to understand the importance of the Wnt ⁄ b-catenin pathway in the development of breast cancer (BC) and its association with different clinicopathological parameters. Alterations (deletion ⁄ methylation ⁄ expression) of some Wnt ⁄ b-catenin pathway antagonists like APC, SFRP1 ⁄ 2, CDH1 and acti- vator b-catenin (CTNNB1) were analyzed in primary BC in Indian patients. High frequencies (65–70%) of overall alterations (dele- tion ⁄ methylation) of the antagonists were seen in the BC samples. Also, 99% (156 ⁄ 158) of the samples showed alterations in any one of the genes, indicating the importance of this pathway in the development of this tumor. Co-alterations of these genes were observed in 30% of samples, with significantly high alterations in late-onset (37%) and estrogen receptor (ER)) ⁄ progesterone recep- tor (PR)) (37%) BC compared with early onset (21%) and ER ⁄ PR+ (18%) BC samples, respectively. Significantly high (P-value = 0.001–0.02) alterations of APC and CDH1 genes were seen in ER) ⁄ PR) BC compared with ER ⁄ PR+ BC. Immunohistochemical analysis showed reduced expression of the Wnt antagonists in BC concordant with their molecular alterations. Nuclear localization of b-catenin showed significant association with alterations in the antagonists and was also significantly high in the ER) ⁄ PR) BC samples. Alterations of SFRP2 coupled with a late clinical stage and low ⁄ nulliparity predicted the worst prognosis in BC patients. Therefore, the present study suggests that cumulative alterations in more than one Wnt antagonist along with increased nuclear accumulation of b-catenin play an important role in the develop- ment of BC and have significant clinical as well as prognostic importance. (Cancer Sci 2012; 103: 210–220) B reast cancer (BC) is a frequently occurring cancer in women worldwide and accounts for 25% of cancer cases in the Eastern Indian population, especially in urban areas. (1) There are several known risk factors for BC. (2) In general, younger women with BC (age £ 40 years) exhibit more aggressive path- ological features and lower survival rates compared with older women with BC. (3) The molecular subtypes of BC, which usu- ally lack expression of the estrogen receptor (ER) and progester- one receptor (PR), also show the worst prognosis of BC. (4) It has been suggested that ER and PR play important roles in regulat- ing the mammary stem cell population and its development. (5) The stem cell population is regulated by a number of self- renewal pathways, (6) among which the Wnt ⁄ b-catenin pathway is quite important due to its cross-talk with the ER signaling pathway (7) and its alterations in several carcinomas including BC. (8) There are several antagonists of the Wnt ⁄ b-catenin path- way like SFRP1, SFRP2, APC and CDH1. (9,10) In different ways, these antagonists regulate the cytoplasmic and nuclear accumulation of b-catenin, the signal transducer of this path- way. (8,11) Accumulation of data shows varying frequencies of deletion (23–73%) and frequent promoter methylation in the Wnt anta- gonists such as SFRP1 ⁄ SFRP2, (9,12,13) APC (10,14–17) and CDH1 (10,18–20) were present in BC. Moreover, loss of b-catenin membranous expression and nuclear accumulation of b-catenin have been reported to be associated with ER) and reduced CDH1 expression, (21,22) along with aggressive tumor phenotype and poor patient outcome in BC. (22,23) It has been reported that ERa could interact with b-catenin (CTNNB1) to activate differ- ent target genes in the nucleus. (24) In addition, there are several conflicting reports concerning the association of Wnt antagonists and b-catenin with conventional clinicopathological parameters of BC such as ER ⁄ PR status and clinical stage. (16,20,25,26) How- ever, the molecular changes and mechanisms that regulate the development and progression of BC remain unclear. Thus, to understand the importance of the Wnt ⁄ b-catenin pathway in the development of BC and its association with dif- ferent clinicopathological parameters, it is pertinent to analyze alterations of the important antagonists of this pathway and b- catenin in the same set of samples. In the present study, altera- tions (deletion ⁄ methylation) of SFRP1 ⁄ 2, APC and CDH1 were analyzed in 158 primary BC samples from Indian patients. Also, to understand the effect of alterations of these genes on b-catenin metabolism the expression of SFRP1, APC and CDH1 along with b-catenin were analyzed immunohisto- chemically in 115 primary BC samples. Our data suggest dif- ferential alterations of Wnt antagonists and nuclear b-catenin expression in ER ⁄ PR (+ ⁄ )) and early ⁄ late onset BC, suggest- ing differences in the molecular pathogenesis of the different subtypes. Materials and Methods Sample collection and clinical data. One hundred and fifty- eight invasive BC samples and their corresponding normal tis- sues or peripheral blood leukocytes were randomly collected from 155 unrelated and untreated BC patients undergoing sur- gery at the hospital section of Chittaranjan National Cancer Institute (CNCI), Kolkata, India, during the period 1999–2008. Informed consent from patients and approval from the Research Ethics Committee of the institute were obtained. All tumors were staged according to the International Union against Cancer (UICC) tumor–node–metastasis (TNM) classification. (27) Details regarding the work plan for the BC samples are presented in 5 To whom correspondence should be addressed. E-mail: [email protected] Cancer Sci | February 2012 | vol. 103 | no. 2 | 210–220 doi: 10.1111/j.1349-7006.2011.02131.x ª 2011 Japanese Cancer Association
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Subtype-specific alterations of the Wnt signaling pathway in breast cancer: clinical and prognostic...

B
Subtype-specific alterations of the Wnt signalingpathway in breast cancer: Clinical and prognosticsignificanceNupur Mukherjee,1 Nilanjana Bhattacharya,1 Neyaz Alam,2 Anup Roy,3 Susanta Roychoudhury4 andChinmay K. Panda1,5
Departments of 1Oncogene Regulation and 2Surgical Oncology, Chittaranjan National Cancer Institute, Kolkata; 3Department of Health and FamilyWelfare, Government of West Bengal, Swasthya Bhavan, Kolkata; 4Molecular and Human Genetics Division, Indian Institute of Chemical Biology, Kolkata,India
(Received June 19, 2011 ⁄ Revised October 5, 2011 ⁄ Accepted October 17, 2011 ⁄ Accepted manuscript online October 25, 2011 ⁄ Article first published online December 7, 2011)
The aim of the study is to understand the importance of theWnt ⁄ b-catenin pathway in the development of breast cancer (BC)and its association with different clinicopathological parameters.Alterations (deletion ⁄ methylation ⁄ expression) of some Wnt ⁄b-catenin pathway antagonists like APC, SFRP1 ⁄ 2, CDH1 and acti-vator b-catenin (CTNNB1) were analyzed in primary BC in Indianpatients. High frequencies (65–70%) of overall alterations (dele-tion ⁄ methylation) of the antagonists were seen in the BC samples.Also, 99% (156 ⁄ 158) of the samples showed alterations in any oneof the genes, indicating the importance of this pathway in thedevelopment of this tumor. Co-alterations of these genes wereobserved in 30% of samples, with significantly high alterations inlate-onset (37%) and estrogen receptor (ER)) ⁄ progesterone recep-tor (PR)) (37%) BC compared with early onset (21%) and ER ⁄ PR+(18%) BC samples, respectively. Significantly high (P-value =0.001–0.02) alterations of APC and CDH1 genes were seen inER) ⁄ PR) BC compared with ER ⁄ PR+ BC. Immunohistochemicalanalysis showed reduced expression of the Wnt antagonists in BCconcordant with their molecular alterations. Nuclear localizationof b-catenin showed significant association with alterations in theantagonists and was also significantly high in the ER) ⁄ PR) BCsamples. Alterations of SFRP2 coupled with a late clinical stageand low ⁄ nulliparity predicted the worst prognosis in BC patients.Therefore, the present study suggests that cumulative alterationsin more than one Wnt antagonist along with increased nuclearaccumulation of b-catenin play an important role in the develop-ment of BC and have significant clinical as well as prognosticimportance. (Cancer Sci 2012; 103: 210–220)
reast cancer (BC) is a frequently occurring cancer inwomen worldwide and accounts for 25% of cancer cases in
the Eastern Indian population, especially in urban areas.(1) Thereare several known risk factors for BC.(2) In general, youngerwomen with BC (age £ 40 years) exhibit more aggressive path-ological features and lower survival rates compared with olderwomen with BC.(3) The molecular subtypes of BC, which usu-ally lack expression of the estrogen receptor (ER) and progester-one receptor (PR), also show the worst prognosis of BC.(4) It hasbeen suggested that ER and PR play important roles in regulat-ing the mammary stem cell population and its development.(5)
The stem cell population is regulated by a number of self-renewal pathways,(6) among which the Wnt ⁄ b-catenin pathwayis quite important due to its cross-talk with the ER signalingpathway(7) and its alterations in several carcinomas includingBC.(8) There are several antagonists of the Wnt ⁄ b-catenin path-way like SFRP1, SFRP2, APC and CDH1.(9,10) In differentways, these antagonists regulate the cytoplasmic and nuclear
Cancer Sci | February 2012 | vol. 103 | no. 2 | 210–220
accumulation of b-catenin, the signal transducer of this path-way.(8,11)
Accumulation of data shows varying frequencies of deletion(23–73%) and frequent promoter methylation in the Wnt anta-gonists such as SFRP1 ⁄ SFRP2,(9,12,13) APC(10,14–17) andCDH1(10,18–20) were present in BC. Moreover, loss of b-cateninmembranous expression and nuclear accumulation of b-cateninhave been reported to be associated with ER) and reducedCDH1 expression,(21,22) along with aggressive tumor phenotypeand poor patient outcome in BC.(22,23) It has been reported thatERa could interact with b-catenin (CTNNB1) to activate differ-ent target genes in the nucleus.(24) In addition, there are severalconflicting reports concerning the association of Wnt antagonistsand b-catenin with conventional clinicopathological parametersof BC such as ER ⁄ PR status and clinical stage.(16,20,25,26) How-ever, the molecular changes and mechanisms that regulate thedevelopment and progression of BC remain unclear.
Thus, to understand the importance of the Wnt ⁄ b-cateninpathway in the development of BC and its association with dif-ferent clinicopathological parameters, it is pertinent to analyzealterations of the important antagonists of this pathway and b-catenin in the same set of samples. In the present study, altera-tions (deletion ⁄ methylation) of SFRP1 ⁄ 2, APC and CDH1were analyzed in 158 primary BC samples from Indianpatients. Also, to understand the effect of alterations of thesegenes on b-catenin metabolism the expression of SFRP1, APCand CDH1 along with b-catenin were analyzed immunohisto-chemically in 115 primary BC samples. Our data suggest dif-ferential alterations of Wnt antagonists and nuclear b-cateninexpression in ER ⁄ PR (+ ⁄ )) and early ⁄ late onset BC, suggest-ing differences in the molecular pathogenesis of the differentsubtypes.
Materials and Methods
Sample collection and clinical data. One hundred and fifty-eight invasive BC samples and their corresponding normal tis-sues or peripheral blood leukocytes were randomly collectedfrom 155 unrelated and untreated BC patients undergoing sur-gery at the hospital section of Chittaranjan National CancerInstitute (CNCI), Kolkata, India, during the period 1999–2008.Informed consent from patients and approval from the ResearchEthics Committee of the institute were obtained. All tumorswere staged according to the International Union against Cancer(UICC) tumor–node–metastasis (TNM) classification.(27) Detailsregarding the work plan for the BC samples are presented in
5To whom correspondence should be addressed. E-mail: [email protected]
doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association

Table 1. Clinicopathological features of breast cancer (BC) patients
Clinical features n (%)Mean
age (years)
ER and PR
negative
status
(n = 114)P-value
) ⁄ ) + %
Age (years)
£40 67 (42) 34 ± 6 32 11 72 0.171
>40 91 (58) 53 ± 8 44 27 58
Sex
Female 153 (98) 44 ± 10 71 38 65 0.95
Male 3 (2) 50 ± 5 2 1 67
Menopausal status
Premenopausal 79 (50) 37 ± 6 32 16 67 0.74
Postmenopausal 79 (50) 51 ± 7 42 24 64
Parity
P £ 2 68 (49) 42 ± 13 34 15 69 0.84
P > 2 71 (51) 46 ± 10 25 10 71
Familial BC
Familial BC+ 15 (10) 38 ± 6 5 3 62 0.84
Familial BC) 140 (90) 43 ± 9 70 36 66
Tumor grade
Grade I 18 (11) 42 ± 12 5 7 42 0.003*
Grade II 91 (57) 43 ± 11 37 21 34
Grade III 49 (31) 49 ± 12 26 4 87
Stage
Stage I and II 60 (38) 46 ± 13 19 24 44 0.0001*
Stage III and IV 98 (62) 43 ± 10 57 14 80
Lymph node (LN)
LN+ 104 (66) 45 ± 11 50 18 73 0.07
LN) 54 (44) 44 ± 12 19 15 56
*Statistically significant (P < 0.05). ER, estrogen receptor;PR, progesterone receptor.
(A)
(B
(a)
(d)
Fig. 1. Deletion status of the Wnt antagonists inbreast cancer (BC). (A) Representative autoradiographshowing: (a) loss of heterozygosity (LOH); (b) MA-1,microsatellite size alteration of one allele; (c)LOH + MA, loss of one allele and size alteration ofthe other; (d,e) homozygous ⁄ hemizygous deletion(HD ⁄ HED); and (f) LOH analysis of exonic markers.Arrow indicates loss of corresponding alleles. N,normal DNA; T, tumor DNA. (B) Pattern of deletionof Wnt ⁄ b-catenin antagonists in early ⁄ late-onset BCand estrogen receptor ⁄ progesterone receptor(ER ⁄ PR) (+ ⁄ )) BC. *Statistically significant differencein the deletion frequency of Wnt antagonistsbetween ER ⁄ PR (+ ⁄ )) BC.
Mukherjee et al.
Data S1 and Figure S1. Detailed clinicopathological histories ofthe patients are presented in Table 1.
Microdissection and DNA extraction. The procedure isdescribed in detail in Data S1. High-molecular-weight DNAwas extracted according to the standard procedure.(28)
Deletion analysis. Deletion of SFRP1 ⁄ 2, APC and CDH1genes were analyzed using microsatellite and restriction frag-ment length polymorphism markers located in or around thesegenes (Table S1) according to the method previouslydescribed(1,29) (Data S1).
Promoter methylation analysis. The promoter methylationstatus of APC, SFRP1, SFRP2 and CDH1 genes were deter-mined by methylation-sensitive restriction analysis(30) (MSRA)using methyl-sensitive restriction enzyme HhaI with recognitionsite GCGC. Promoter regions of the genes harbored at least oneHhaI site. Details of primer sequences are listed in Table S1.The 445-bp fragment of b-3A adaptin gene (K1) and 229-bpfragment of RARb2 exon-1 (K2) was used as digestion andintegrity controls, respectively (see Data S1 and Table S1 fordetails of PCR conditions).
Immunohistochemical analysis. Of the 158 BC samples, 114paraffin-embedded BC tissue sections were randomly selectedfor immunohistochemical assay to analyze the expression statusof ER and PR in BC.(31) To verify the impact of gene alterationsat the protein level, the expression status of APC, SFRP1,CDH1 and b-catenin were detected immunohistochemically inparaffin-embedded sections of five normal breast tissues and115 BC samples(32) (randomly selected from 158 BC samplesincluded in the present study). For color development, diam-inobenzidine (sc-24982, SantaCruz Biotechnology, Santa Cruz,CA, USA) was used, with hematoxylin as a counterstain. Theantibodies, dilutions and antigen retrieval methods used aresummarized in Table S2 (see Data S1 for details).
Statistical analysis. Probability value (P-value) £ 0.05 wasconsidered statistically significant. All statistical analysis was
)
(b) (c)
(e) (f)
Cancer Sci | February 2012 | vol. 103 | no. 2 | 211ªª 2011 Japanese Cancer Association

performed using statistical programs EpiInfo (version 6.04b,1997; Centers for Disease Control and Prevention, Atlanta, GA,USA) and SPSS 10.0 (SPSS Inc. Chicago, IL, USA) (see DataS1 for details).
Results
ER ⁄ PR status of the BC samples. Positive staining withER ⁄ PR was localized to the nuclei of BC cells. Also, hetero-geneity of staining was frequently observed. The majority (66%;76 ⁄ 114) of the BC samples showed no expression of ER and PR(Fig. S2). The ER and PR negative status of the samples showeda significant association with late tumor stage (P = 0.003) andincreased grade (P = 0.004) in BC patients, along with border-
(A)
(C)
(B)
Fig. 3. Pattern of overall alterations (deletion ⁄ methylation) of differentER ⁄ PR (+ ⁄ )) BC. # and *Significant difference (P < 0.05) in the two subsets
212
line significance with nodes at pathology (P = 0.07) (Table 1).However, no such correlation was seen with age of onset, meno-pausal status and parity.
Deletion analysis of the candidate Wnt ⁄ b-cateninantagonists. After excluding non-informative tumor samples(35 of 158 at SFRP1, 14 of 158 at APC and 79 of 158 at theCDH1 locus), deletion analysis of the Wnt ⁄ b-catenin antago-nists revealed the highest deletion frequency in SFRP1 (29%,36 ⁄ 123) followed by APC (26%, 38 ⁄ 144), CDH1 (23%, 18 ⁄ 79)and SFRP2 (21%, 33 ⁄ 158) (Fig. 1). No significant differenceswere observed in the deletion frequencies of these genesbetween early ⁄ late-onset BC. However, significantly higherdeletion frequencies of APC and CDH1 were observed inER) ⁄ PR) BC than ER ⁄ PR+ BC samples (P = 0.029, 0.048,
Fig. 2. Promoter methylation status of Wntantagonists analyzed by methylation-sensitiverestriction analysis. (A) Schematic representation ofpromoter regions of candidate genes revealingdistribution of HhaI (GCGC: arrowhead) restrictionsites. ( fi ), Location of methylation primers; +1,transcription start site. (B) Methylation status ofantagonists in breast cancer (BC). H, HhaI digestedDNA; U, undigested DNA; K1 and K2, controls forDNA digestion and integrity check, respectively. (C)Pattern of promoter methylation of different Wntantagonists in early ⁄ late-onset BC and ER ⁄ PR (+ ⁄ ))BC. *Significant difference in methylation fre-quency of the genes between ER ⁄ PR (+ ⁄ )) BC.
Wnt ⁄ b-catenin antagonists in early ⁄ late-onset breast cancer (BC) andof BC; A, alteration.
doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association
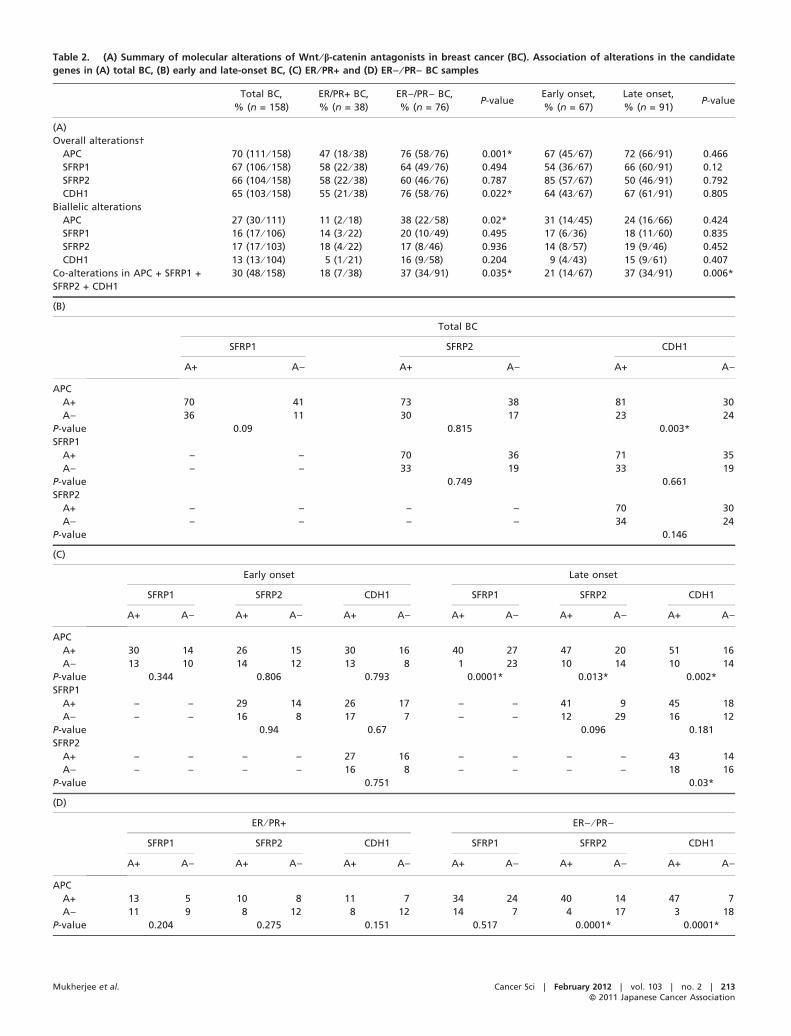
Table 2. (A) Summary of molecular alterations of Wnt ⁄ b-catenin antagonists in breast cancer (BC). Association of alterations in the candidate
genes in (A) total BC, (B) early and late-onset BC, (C) ER ⁄ PR+ and (D) ER) ⁄ PR) BC samples
Total BC,
% (n = 158)
ER/PR+ BC,
% (n = 38)
ER)/PR) BC,
% (n = 76)P-value
Early onset,
% (n = 67)
Late onset,
% (n = 91)P-value
(A)
Overall alterations†
APC 70 (111 ⁄ 158) 47 (18 ⁄ 38) 76 (58 ⁄ 76) 0.001* 67 (45 ⁄ 67) 72 (66 ⁄ 91) 0.466
SFRP1 67 (106 ⁄ 158) 58 (22 ⁄ 38) 64 (49 ⁄ 76) 0.494 54 (36 ⁄ 67) 66 (60 ⁄ 91) 0.12
SFRP2 66 (104 ⁄ 158) 58 (22 ⁄ 38) 60 (46 ⁄ 76) 0.787 85 (57 ⁄ 67) 50 (46 ⁄ 91) 0.792
CDH1 65 (103 ⁄ 158) 55 (21 ⁄ 38) 76 (58 ⁄ 76) 0.022* 64 (43 ⁄ 67) 67 (61 ⁄ 91) 0.805
Biallelic alterations
APC 27 (30 ⁄ 111) 11 (2 ⁄ 18) 38 (22 ⁄ 58) 0.02* 31 (14 ⁄ 45) 24 (16 ⁄ 66) 0.424
SFRP1 16 (17 ⁄ 106) 14 (3 ⁄ 22) 20 (10 ⁄ 49) 0.495 17 (6 ⁄ 36) 18 (11 ⁄ 60) 0.835
SFRP2 17 (17 ⁄ 103) 18 (4 ⁄ 22) 17 (8 ⁄ 46) 0.936 14 (8 ⁄ 57) 19 (9 ⁄ 46) 0.452
CDH1 13 (13 ⁄ 104) 5 (1 ⁄ 21) 16 (9 ⁄ 58) 0.204 9 (4 ⁄ 43) 15 (9 ⁄ 61) 0.407
Co-alterations in APC + SFRP1 +
SFRP2 + CDH1
30 (48 ⁄ 158) 18 (7 ⁄ 38) 37 (34 ⁄ 91) 0.035* 21 (14 ⁄ 67) 37 (34 ⁄ 91) 0.006*
(B)
Total BC
SFRP1 SFRP2 CDH1
A+ A) A+ A) A+ A)
APC
A+ 70 41 73 38 81 30
A) 36 11 30 17 23 24
P-value 0.09 0.815 0.003*
SFRP1
A+ – – 70 36 71 35
A) – – 33 19 33 19
P-value 0.749 0.661
SFRP2
A+ – – – – 70 30
A) – – – – 34 24
P-value 0.146
(C)
Early onset Late onset
SFRP1 SFRP2 CDH1 SFRP1 SFRP2 CDH1
A+ A) A+ A) A+ A) A+ A) A+ A) A+ A)
APC
A+ 30 14 26 15 30 16 40 27 47 20 51 16
A) 13 10 14 12 13 8 1 23 10 14 10 14
P-value 0.344 0.806 0.793 0.0001* 0.013* 0.002*
SFRP1
A+ – – 29 14 26 17 – – 41 9 45 18
A) – – 16 8 17 7 – – 12 29 16 12
P-value 0.94 0.67 0.096 0.181
SFRP2
A+ – – – – 27 16 – – – – 43 14
A) – – – – 16 8 – – – – 18 16
P-value 0.751 0.03*
(D)
ER ⁄ PR+ ER) ⁄ PR)
SFRP1 SFRP2 CDH1 SFRP1 SFRP2 CDH1
A+ A) A+ A) A+ A) A+ A) A+ A) A+ A)
APC
A+ 13 5 10 8 11 7 34 24 40 14 47 7
A) 11 9 8 12 8 12 14 7 4 17 3 18
P-value 0.204 0.275 0.151 0.517 0.0001* 0.0001*
Mukherjee et al. Cancer Sci | February 2012 | vol. 103 | no. 2 | 213ªª 2011 Japanese Cancer Association

Table 2. (continued)
ER ⁄ PR+ ER) ⁄ PR)
SFRP1 SFRP2 CDH1 SFRP1 SFRP2 CDH1
A+ A) A+ A) A+ A) A+ A) A+ A) A+ A)
SFRP1
A+ – – 11 13 14 10 – – 32 13 35 13
A) – – 7 7 5 9 – – 17 11 15 12
P-value 0.959 0.128 0.507 0.125
SFRP2
A+ – – – – 11 7 – – – – 37 11
A) – – – – 8 12 – – – – 18 9
P-value 0.152 0.847
*Statistically significant (P < 0.05). †Deletion ⁄ methylation. A+, alteration positive; A), alteration negative; ER, estrogen receptor;PR, progesterone receptor.
(A) (B) (C)
(D) (E) (F)
(J) (K) (L)
(G) (H) (I)
Fig. 4. Representative immunohistochemical staining patterns of CDH1, SFRP1, APC and b-catenin proteins in breast tissues. (A,D,G,J) Normalbreast tissue showing the expression pattern of CDH1, SFRP1, APC and b-catenin. (B,C) Reduced ⁄ negative expression of CDH1 in breast cancer(BC). (E,F) Reduced ⁄ negative, predominantly cytoplasmic expression of SFRP1 in BC. (H,I) Reduced ⁄ negative cytoplasmic expression along withnuclear expression of APC in BC. (K,L) Intense nuclear and ⁄ or cytoplasmic staining of b-catenin in BC. Arrows indicate membrane ⁄ cytoplasmicexpression. Arrowheads indicate nuclear expression pattern of these genes in primary BC. Bar, 25 lm.
214 doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association

Table 3. Expression status of the Wnt ⁄ b-catenin pathway-associated genes in breast cancer (BC) samples
Expression statusTotal BC,
% (n = 115)
ER/PR+ BC,
% (n = 38)
ER)/PR) BC,
% (n = 76)P-value
Early onset,
% (n = 49)
Late-onset,
% (n = 66)P-value
CDH1 (mem ⁄ cyto)† 58 (67 ⁄ 115) 26 (10 ⁄ 38) 74 (56 ⁄ 76) 0.000001* 63 (31 ⁄ 49) 61 (40 ⁄ 66) 0.771
SFRP1 (cyto)† 43 (49 ⁄ 115) 29 (11 ⁄ 38) 50 (38 ⁄ 76) 0.032* 51 (25 ⁄ 49) 41 (27 ⁄ 66) 0.281
APC (cyto)† 64 (74 ⁄ 115) 34 (13 ⁄ 38) 80 (61 ⁄ 76) 0.000001* 61 (30 ⁄ 49) 67 (44 ⁄ 66) 0.546
APC (nuc)‡ 33 (38 ⁄ 115) 18 (7 ⁄ 38) 29 (22 ⁄ 76) 0.223 26 (13 ⁄ 49) 21 (14 ⁄ 66) 0.169
b-catenin (cyto)† 54 (62 ⁄ 115) 66 (25 ⁄ 38) 49 (37 ⁄ 76) 0.33 53 (26 ⁄ 49) 54 (36 ⁄ 66) 0.874
b-catenin (nuc)§ 47 (54 ⁄ 115) 16 (6 ⁄ 38) 63 (48 ⁄ 76) 0.000001* 53 (26 ⁄ 49) 42 (28 ⁄ 66) 0.252
*Statistically significant (P < 0.05). †Reduced expression () ⁄ +). ‡Positive expression of APC in nucleus. §High (+++) nuclear expression ofb-catenin. cyto, cytoplasmic expression; ER, estrogen receptor; mem, membrane expression; nuc, nuclear expression.
0.023, respectively). Two samples showed homozygous dele-tions in the SFRP2 locus (#4187, #5051). A low frequency ofmicrosatellite alterations (MA) (1–6%) and biallelic alterations(LMA) (3–5%) were observed in the samples (Table S3).
Overall 44% (69 ⁄ 158) of the samples harbored deletions in atleast one of the antagonists and 6 (4%) samples harbored dele-tions in all genes. Deletion of these genes showed a significantassociation with each other in the overall BC samples. A similartrend was observed in early ⁄ late-onset BC and ER) ⁄ PR) BC(Table S4).
Promoter methylation analysis of candidate Wnt ⁄ b-cateninantagonists. The differential methylation pattern in promoterregions of the antagonists was seen in the following order: APC(65%, 103 ⁄ 158) > CDH1 (63%, 100 ⁄ 158) > SFRP1 (56%,85 ⁄ 158) > SFRP2 (54%, 82 ⁄ 158). No significant difference wasobserved in the promoter methylation frequencies of the candi-date genes in the early ⁄ late-onset BC samples. However, asignificant increase in promoter methylation was seen in APCand CDH1 in the ER) ⁄ PR) BC samples compared with theER ⁄ PR+ BC samples (P = 0.0008, 0.042) (Fig. 2).
Simultaneous promoter methylation in the four genes wasseen in 18% (28 ⁄ 158) of the total BC samples (Table S3). Sig-nificant association was observed among promoter methylationof APC and SFRP2 with CDH1, and SFRP1 with SFRP2 in thetotal BC. A differential pattern of correlation was observed inthese genes in early ⁄ late-onset BC and in the ER ⁄ PR(+ ⁄ )) BCsamples (P = 0.001–0.03) (Table S5).
Overall alterations of the Wnt ⁄ b-catenin antagonists. Overallalterations (deletion ⁄ methylation) of the Wnt ⁄ b-catenin antago-nists were seen in the following order: APC (70%, 111 ⁄158) > SFRP1 (67%, 106 ⁄ 158) > CDH1 (66%, 104 ⁄ 158) >SFRP2 (65%, 103 ⁄ 158) (Fig. 3). Alterations in any one of thesegenes were seen in approximately 99% (157 ⁄ 158) of the BCsamples indicating the importance of these genes in the develop-ment of this tumor.
Biallelic alterations of these genes by deletion and methyla-tion were observed in the following order: APC (27%,30 ⁄ 111) > SFRP2 (18%, 17 ⁄ 103) > SFRP1 (20%, 17 ⁄ 106)> CDH1 (13%, 13 ⁄ 104). Co-alterations of these genes wereobserved in 30% of the BC samples with significantly high alter-ations in the late-onset (37%) and ER) ⁄ PR) (37%) BC samplescompared with early onset (21%) and ER ⁄ PR+ (18%) BC. Sig-nificantly higher alterations of APC and CDH1 were observed inER) ⁄ PR) BC compared with ER ⁄ PR+ BC samples, but no suchdifference was seen with age of onset of the disease (Table 2A).
Interestingly, in the absence of APC alterations, significantlyhigh alterations of the other three genes were observed in theER ⁄ PR+ BC samples compared with the ER) ⁄ PR) BC sam-ples. A similar phenomenon was also evident in the sampleswithout CDH1 alterations, but no such association was seen inthe samples without SFRP1 ⁄ 2 alterations. Also, 13% of the sam-ples (23 ⁄ 158) showed alterations in SFRP1 ⁄ 2 only and wereprevalent in the ER ⁄ PR+ BC samples (Fig. 3).
Mukherjee et al.
Chi-squared analysis of the alterations of the genes revealedalterations of APC were significantly associated with CDH1(P = 0.003) in the BC samples (Table 2B). However, differen-tial association among alterations of these genes was observedin early ⁄ late-onset BC and ER ⁄ PR (+ ⁄ )) BC (Table 2C,D).
Immunohistochemical analysis. In normal breast tissue, pre-dominant membrane expression of CDH1, b-catenin and SFRP1was observed in luminal epithelial and myoepithelial cells(Fig. 4). In contrast, cytoplasmic expression of APC and SFRP1was evident in both luminal and myoepithelial cells, while cyto-plasmic expression of b-catenin was present in luminal epithe-lial cells only.
In the BC samples, reduced or absence of expression ofCDH1, SFRP1 and APC proteins was seen in 58% (67 ⁄ 115),43% (49 ⁄ 115) and 64% (74 ⁄ 115) of the samples, respectively,while overexpression of b-catenin in the nucleus and cytoplasmwas seen in 47% (54 ⁄ 115) and 54% (62 ⁄ 115) of the samples,respectively (Table S3). In addition to reduced cytoplasmicexpression of APC seen in the breast tissue samples, nuclearexpression of APC was evident in 33% (38 ⁄ 115) of the BC sam-ples (Table 3). Expression of APC, SFRP1 and CDH1 proteinsshowed concordance with their molecular alterations in the sam-ples. A statistically significant correlation was observed betweenb-catenin nuclear staining and alterations in APC, SFRP2 andCDH1 genes (P = 0.013–0.048) in the BC samples (Table 4).Interestingly, in the ER) ⁄ PR) BC samples, significantly lowexpression of CDH1, SFRP1 and APC along with a significantincrease in nuclear expression of b-catenin were observed(P = 0.000001–0.032), pointing towards differences in themolecular pathogenesis of the ER ⁄ PR+ and ER) ⁄ PR) BC sam-ples (Table 3).
Clinicopathological association and patient survival. Alterations(deletion ⁄ methylation) of these genes were comparatively high(63–68%) in stage I and II tumor samples and became compara-ble in the later stages (stages III and IV) (60–71%). A similartrend was also evident in tumor grades, except for significantlyhigh alterations of CDH1 in higher grades of BC (P = 0.029).Alterations of these genes showed no association with the lymphnode status of the patients (Table S6; Fig. S3).
Significant association was observed in promoter methylationof SFRP2 and CDH1 with a poor patient outcome for BCpatients. The BC patients with alterations in CDH1, SFRP2 orAPC showed significantly poor overall survival (P = 0.002–0.052) (Fig. 5). Importantly, significantly poor survival was seenin the ER) ⁄ PR) BC patients harboring alterations in any of theWnt pathway antagonists (Fig. 5H). Interestingly, BC patientswith high nuclear b-catenin expression in the samples alsoshowed significantly poor overall survival (Fig. 5G).
The univariate Cox model (Table 5) showed a significantassociation (P = 0.00026–0.0065) with alterations in SFRP2 andCDH1 genes, negative ER ⁄ PR status, positive lymph node sta-tus, late stage (III and IV) and low parity (£2) with the overallsurvival of BC patients (Table 5). However, Cox multivariate
Cancer Sci | February 2012 | vol. 103 | no. 2 | 215ªª 2011 Japanese Cancer Association

Tab
le4.
Co
rrela
tio
nb
etw
een
the
gen
eti
cst
atu
san
dp
rote
inexp
ress
ion
of
APC
,SFR
P1
an
dC
DH
1g
en
es
inb
reast
can
cer
CD
H1
(mem
⁄cyt
o)†
SFR
P1
(cyt
o)†
APC
(cyt
o)†
APC
(nu
c)†
b-c
ate
nin
(cyt
o)†
b-c
ate
nin
(nu
c)†
)⁄+
++
⁄nP-v
alu
e)
⁄++
+⁄n
P-v
alu
e)
⁄++
+⁄n
P-v
alu
e)
⁄++
+⁄+
++
P-v
alu
e)
⁄++
+⁄+
++
P-v
alu
e)
⁄++
++
+P-v
alu
e
CD
H1
alt
era
tio
ns
Po
siti
ve53
26
0.0
04*
37
42
0.1
74
57
22
0.0
1*
55
24
0.1
41
36
43
0.0
07*
37
42
0.0
48*
Neg
ati
ve14
22
12
24
17
19
20
16
26
10
24
12
SFR
P1
alt
era
tio
ns
Po
siti
ve42
29
0.8
04
38
33
0.0
02*
50
21
0.1
51
48
21
0.2
339
32
0.7
81
38
33
0.8
96
Neg
ati
ve25
19
11
33
24
18
27
19
23
21
23
21
SFR
P2
alt
era
tio
ns
Po
siti
ve43
25
0.1
93
32
36
0.2
45
47
21
0.2
49
19
0.1
15
34
34
0.0
86
30
38
0.0
21*
Neg
ati
ve24
23
17
30
27
20
26
19
28
19
31
16
APC
alt
era
tio
ns
Po
siti
ve52
24
0.0
02*
40
36
0.0
02*
59
18
0.0
001*
59
17
0.6
95
34
42
0.0
05*
34
42
0.0
13*
Neg
ati
ve15
24
930
15
23
29
10
28
11
27
12
*St
ati
stic
all
ysi
gn
ifica
nt
(P<
0.0
5).
†In
the
case
of
CD
H1,
SFR
P1,
APC
an
db-c
ate
nin
(cyt
op
lasm
),n
eg
ati
ve⁄lo
wexp
ress
ion
of
pro
tein
sw
as
con
sid
ere
das
an
eve
nt
wh
ile
hig
hexp
ress
ion
of
nu
clear
b-c
ate
nin
was
con
sid
ere
das
an
eve
nt.
)⁄+
,n
eg
ati
ve⁄lo
wexp
ress
ion
;+
+⁄+
++
⁄n,
mo
dera
te⁄h
igh
⁄no
rmal
exp
ress
ion
;n
,n
orm
al
exp
ress
ion
com
pare
dw
ith
no
rmal
bre
ast
tiss
ue.
cyto
,cy
top
lasm
ic;
mem
,m
em
bra
ne
exp
ress
ion
;n
uc,
nu
clear
exp
ress
ion
.
216
analysis revealed that SFRP2 alterations, negative ER ⁄ PRstatus, positive lymph node status, low ⁄ nulliparity (parity £2)and late tumor stage were significantly associated with overallsurvival of BC patients (P = 0.0133–0.038) (Table 5).
Discussion
The aim of the present study was to understand the importance ofthe Wnt ⁄ b-catenin pathway in the development of BC. To begin,alterations of some antagonists of the Wnt ⁄ b-catenin pathway,namely APC, SFRP1, SFRP2 and CDH1, were analyzed in pri-mary BC of Indian patients. Alterations of these genes were thencorrelated with different clinicopathological parameters such asage of onset, ER ⁄ PR expression status, nodes at pathology andparity. The majority of samples (67%) was ER) ⁄ PR) andshowed a positive correlation with high tumor grade and stage ofBC. A similar expression pattern of ER and PR has been reportedin Indian patients(33,34) unlike European and North Americanpopulations.(33) This could be due to differences in etio-logical factors and probably ethnicity as suggested by otherinvestigators.(5,34) Association of ER) ⁄ PR) status of BC with ahigher tumor grade in ductal carcinoma of the breast has alsobeen reported.(35)
The majority of samples (99%; 157 ⁄ 158) showed genetic andepigenetic alterations (deletion ⁄ methylation) in at least one ofthe antagonists of the Wnt ⁄ b-catenin pathway. This suggeststhat alterations in different antagonists might act in a comple-mentary fashion to make comparable contributions to the pro-cess of breast carcinogenesis. A comparatively low frequency ofdeletion (21–26%) compared with methylation (65–67%) wasobserved in the Wnt antagonists in the samples, suggesting epi-genetic alteration as the primary mechanism of inactivation ofthese genes in BC. Deletions of these genes showed a significantassociation with each other in the total BC samples, but showeddifferential association either in early ⁄ late-onset BC or inER ⁄ PR() ⁄ +) BC samples, indicating differences in the subtype-specific deletion pattern. A variable frequency of deletion(23–73%) in APC(10,15–17) and CDH1(10,18,19) genes has beenreported by other investigators in BC, as well as in a variety ofother human cancers.(36–38) This difference might be due to dif-ferences in sample size, methodology used in the analysis andethnicity of BC patients. In addition, deletions in APC andSFRP2 associated with a poor overall survival of BC patientssuggest their prognostic importance.
Like deletion, methylation of these genes showed a significantassociation with each other, suggesting the necessity of cumula-tive epigenetic hits for this tumorigenesis. Moreover, differentialassociation in methylation of these genes was observed in eitherearly ⁄ late onset of the disease or ER ⁄ PR (+ ⁄ )) status of thesamples. Varying frequencies of methylation in APC (36–71%),SFRP1 (40–61%), SFRP2 (77–83%) and CDH1 (41–72%) havebeen reported in BC by other investigators.(11,13,16–19) Like ourdata, association of CDH1 methylation with APC methylationand reduced ER expression has been reported in BC.(10,20,39)
However, some investigators(16,25) did not find any associationbetween methylation of APC and SFRP2 with the ER ⁄ PR statusof the BC samples. The prognostic significance of CDH1,SFRP1 and APC methylation was also reported in BC and othercancers,(12,40,41) while promoter methylation of SFRP2 wasreported to show no correlation with clinical patient outcome inBC.(25)
Besides, a significant difference in co-alterations (dele-tion ⁄ methylation) of these genes was observed in ER) ⁄ PR) BCcompared with early onset and ER ⁄ PR+ BC samples, suggestingdifferences in the molecular pathogenesis of the subtypes. Thiswas also evident from the significant association of APC ⁄ CDH1alterations with ER ⁄ PR) BC samples and the prevalence ofSFRP1 ⁄ 2 alterations alone in ER ⁄ PR+ BC samples. Similar dif-
doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association

Fig. 5. Kaplan–Meier analysis of survival (up to 5 years) of breast cancer (BC) patients. Molecular alterations (deletion and methylation) of APC,SFRP1 ⁄ 2 and CDH1 genes and nuclear accumulation of b-catenin (CTNNB1) were significantly associated with poor overall survival (OS) (A–G).Survival of BC patients with ⁄ without alteration of Wnt antagonists stratified for estrogen receptor ⁄ progesterone receptor (ER ⁄ PR) status (H). A,alteration; n, sample size.
ferences in the molecular pathogenesis of the different subtypesof BC were observed in the Chi-squared analysis of alterationsof these genes.
In the present study we did not find any homozygous dele-tion of these genes, but their bi-allele alterations were evidentin 13–27% of samples, suggesting these as candidate tumorsuppressor genes (TSGs) according to the Knudson’s two-hithypothesis,(42) although other types of gene inactivation mech-anisms such as mutation and splice variations of these genescannot be ruled out in this tumor.(43,44)
Clinicopathological analysis suggests that alterations of theWnt antagonists are needed for early invasiveness of the disease
Mukherjee et al.
and have prognostic importance in either BC or ER) ⁄ PR) BCpatients. Moreover, association of CDH1 alterations and higherhistological grades of BC suggest association with tumor pro-gression. In addition, univariate analysis suggests thatER) ⁄ PR) patients with parity £2, high clinical stage and nodesat pathology have a worst prognosis having alterations of any ofthe APC ⁄ CDH1 ⁄ SFRP2 genes. In contrast, in multivariate anal-ysis, SFRP2 alterations are suggested to be an important riskfactor in patients with parity £2 and higher histological grades.The reason for the association of SFRP2 alterations with theabove etiological factors in BC is not clear. Similar to our find-ings, Hill et al.(45) have reported a significant association
Cancer Sci | February 2012 | vol. 103 | no. 2 | 217ªª 2011 Japanese Cancer Association
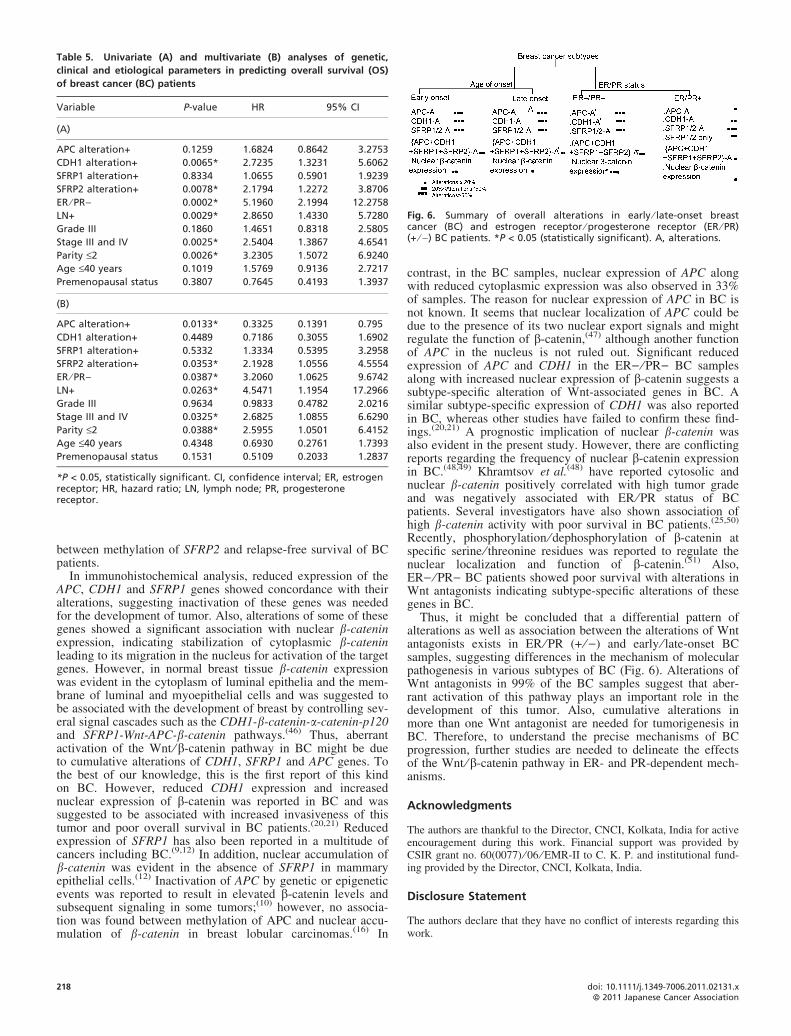
Fig. 6. Summary of overall alterations in early ⁄ late-onset breastcancer (BC) and estrogen receptor ⁄ progesterone receptor (ER ⁄ PR)(+ ⁄ )) BC patients. *P < 0.05 (statistically significant). A, alterations.
Table 5. Univariate (A) and multivariate (B) analyses of genetic,
clinical and etiological parameters in predicting overall survival (OS)
of breast cancer (BC) patients
Variable P-value HR 95% CI
(A)
APC alteration+ 0.1259 1.6824 0.8642 3.2753
CDH1 alteration+ 0.0065* 2.7235 1.3231 5.6062
SFRP1 alteration+ 0.8334 1.0655 0.5901 1.9239
SFRP2 alteration+ 0.0078* 2.1794 1.2272 3.8706
ER ⁄ PR) 0.0002* 5.1960 2.1994 12.2758
LN+ 0.0029* 2.8650 1.4330 5.7280
Grade III 0.1860 1.4651 0.8318 2.5805
Stage III and IV 0.0025* 2.5404 1.3867 4.6541
Parity £2 0.0026* 3.2305 1.5072 6.9240
Age £40 years 0.1019 1.5769 0.9136 2.7217
Premenopausal status 0.3807 0.7645 0.4193 1.3937
(B)
APC alteration+ 0.0133* 0.3325 0.1391 0.795
CDH1 alteration+ 0.4489 0.7186 0.3055 1.6902
SFRP1 alteration+ 0.5332 1.3334 0.5395 3.2958
SFRP2 alteration+ 0.0353* 2.1928 1.0556 4.5554
ER ⁄ PR) 0.0387* 3.2060 1.0625 9.6742
LN+ 0.0263* 4.5471 1.1954 17.2966
Grade III 0.9634 0.9833 0.4782 2.0216
Stage III and IV 0.0325* 2.6825 1.0855 6.6290
Parity £2 0.0388* 2.5955 1.0501 6.4152
Age £40 years 0.4348 0.6930 0.2761 1.7393
Premenopausal status 0.1531 0.5109 0.2033 1.2837
*P < 0.05, statistically significant. CI, confidence interval; ER, estrogenreceptor; HR, hazard ratio; LN, lymph node; PR, progesteronereceptor.
between methylation of SFRP2 and relapse-free survival of BCpatients.
In immunohistochemical analysis, reduced expression of theAPC, CDH1 and SFRP1 genes showed concordance with theiralterations, suggesting inactivation of these genes was neededfor the development of tumor. Also, alterations of some of thesegenes showed a significant association with nuclear b-cateninexpression, indicating stabilization of cytoplasmic b-cateninleading to its migration in the nucleus for activation of the targetgenes. However, in normal breast tissue b-catenin expressionwas evident in the cytoplasm of luminal epithelia and the mem-brane of luminal and myoepithelial cells and was suggested tobe associated with the development of breast by controlling sev-eral signal cascades such as the CDH1-b-catenin-a-catenin-p120and SFRP1-Wnt-APC-b-catenin pathways.(46) Thus, aberrantactivation of the Wnt ⁄ b-catenin pathway in BC might be dueto cumulative alterations of CDH1, SFRP1 and APC genes. Tothe best of our knowledge, this is the first report of this kindon BC. However, reduced CDH1 expression and increasednuclear expression of b-catenin was reported in BC and wassuggested to be associated with increased invasiveness of thistumor and poor overall survival in BC patients.(20,21) Reducedexpression of SFRP1 has also been reported in a multitude ofcancers including BC.(9,12) In addition, nuclear accumulation ofb-catenin was evident in the absence of SFRP1 in mammaryepithelial cells.(12) Inactivation of APC by genetic or epigeneticevents was reported to result in elevated b-catenin levels andsubsequent signaling in some tumors;(10) however, no associa-tion was found between methylation of APC and nuclear accu-mulation of b-catenin in breast lobular carcinomas.(16) In
218
contrast, in the BC samples, nuclear expression of APC alongwith reduced cytoplasmic expression was also observed in 33%of samples. The reason for nuclear expression of APC in BC isnot known. It seems that nuclear localization of APC could bedue to the presence of its two nuclear export signals and mightregulate the function of b-catenin,(47) although another functionof APC in the nucleus is not ruled out. Significant reducedexpression of APC and CDH1 in the ER) ⁄ PR) BC samplesalong with increased nuclear expression of b-catenin suggests asubtype-specific alteration of Wnt-associated genes in BC. Asimilar subtype-specific expression of CDH1 was also reportedin BC, whereas other studies have failed to confirm these find-ings.(20,21) A prognostic implication of nuclear b-catenin wasalso evident in the present study. However, there are conflictingreports regarding the frequency of nuclear b-catenin expressionin BC.(48,49) Khramtsov et al.(48) have reported cytosolic andnuclear b-catenin positively correlated with high tumor gradeand was negatively associated with ER ⁄ PR status of BCpatients. Several investigators have also shown association ofhigh b-catenin activity with poor survival in BC patients.(25,50)
Recently, phosphorylation ⁄ dephosphorylation of b-catenin atspecific serine ⁄ threonine residues was reported to regulate thenuclear localization and function of b-catenin.(51) Also,ER) ⁄ PR) BC patients showed poor survival with alterations inWnt antagonists indicating subtype-specific alterations of thesegenes in BC.
Thus, it might be concluded that a differential pattern ofalterations as well as association between the alterations of Wntantagonists exists in ER ⁄ PR (+ ⁄ )) and early ⁄ late-onset BCsamples, suggesting differences in the mechanism of molecularpathogenesis in various subtypes of BC (Fig. 6). Alterations ofWnt antagonists in 99% of the BC samples suggest that aber-rant activation of this pathway plays an important role in thedevelopment of this tumor. Also, cumulative alterations inmore than one Wnt antagonist are needed for tumorigenesis inBC. Therefore, to understand the precise mechanisms of BCprogression, further studies are needed to delineate the effectsof the Wnt ⁄ b-catenin pathway in ER- and PR-dependent mech-anisms.
Acknowledgments
The authors are thankful to the Director, CNCI, Kolkata, India for activeencouragement during this work. Financial support was provided byCSIR grant no. 60(0077) ⁄ 06 ⁄ EMR-II to C. K. P. and institutional fund-ing provided by the Director, CNCI, Kolkata, India.
Disclosure Statement
The authors declare that they have no conflict of interests regarding thiswork.
doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association

References
1 Sinha S, Chunder N, Mukherjee N et al. Frequent deletion and methylation inSH3GL2 and CDKN2A loci are associated with early- and late-onset breastcarcinoma. Ann Surg Oncol 2008; 15: 1070–80.
2 Nkondjock A, Ghadirian P. Risk factors and risk reduction of breast cancer.Med Sci (Paris) 2005; 21: 175–80.
3 Chunder N, Mandal S, Basu D et al. Deletion mapping of chromosome 1 inearly onset and late onset breast tumors–a comparative study in eastern India.Pathol Res Pract 2003; 99: 313–21.
4 Alquobali F, Miller SA, Muhie S, Day A, Jett M, Hammamieh R. Estrogenreceptor dependent genomic expression profiles in breast cancer cells inresponse to fatty acids. J Carcinog 2010; 8: 17.
5 Polyak K. Breast cancer: origins and evolution. J Clin Invest 2007; 117:3155–63.
6 Behbod F, Rosen JM. Will cancer stem cells provide new therapeutic targets?Carcinogenesis 2004; 26: 703–11.
7 Kouzmenko AP, Takeyama K, Ito S et al. Wnt ⁄ b-catenin and estrogensignaling converge in vivo. J Biol Chem 2004; 279: 40255–8.
8 Collu GM, Meurette O, Brennan K. Is there more to Wnt signaling in breastcancer than stabilization of beta-catenin? Breast Cancer Res 2009; 11: 105.
9 Bovolenta P, Esteve P, Ruiz JM, Cisneros E, Lopez-Rios J. Beyond Wntinhibition: new functions of secreted Frizzled-related proteins in developmentand disease. J Cell Sci 2008; 121: 737–46.
10 Sarrio D, Moreno-Bueno G, Hardisson D et al. Epigenetic and geneticalterations of APC and CDH1 genes in lobular breast cancer: relationshipswith abnormal E-cadherin and catenin expression and microsatelliteinstability. Int J Cancer 2003; 106: 208–15.
11 Aoki K, Taketo MM. Adenomatous polyposis coli (APC): a multi-functionaltumor suppressor gene. J Cell Sci 2007; 120: 3327–35.
12 Suzuki H, Toyota M, Caraway H et al. Frequent epigenetic inactivation ofWnt antagonist genes in breast cancer. Br J Cancer 2008; 98: 1147–56.
13 Gauger KJ, Hugh JM, Troester MA, Schneider SS. Down-regulation of sfrp1in a mammary epithelial cell line promotes the development of acd44high ⁄ cd24low population which is invasive and resistant to anoikis.Cancer Cell Int 2009; 9: 11.
14 Medeiros AC, Nagai MA, Neto MM, Brentani RR. Loss of heterozygosityaffecting the APC and MCC genetic loci in patients with primary breastcarcinomas. Cancer Epidemiol Biomarkers Prev 1994; 3: 331–3.
15 Van der Auwera I, Van Laere SJ, Van den Bosch SM et al. Aberrantmethylation of the adenomatous polyposis coli (APC) gene promoter isassociated with the inflammatory breast cancer phenotype. Br J Cancer 2008;99: 1735–42.
16 Jin Z, Tamura G, Tsuchiya T et al. Adenomatous polyposis coli (APC) genepromoter hypermethylation in primary breast cancers. Br J Cancer 2001;85(1): 69–73.
17 Virmani AK, Rathi A, Sathyanarayana UG et al. Aberrant methylation of theadenomatous polyposis coli (APC) gene promoter 1A in breast and lungcarcinomas. Clin Cancer Res 2001; 7: 1998–2004.
18 Kouvaraki M, Gorgoulis VG, Rassidakis GZ et al. Alterations of the 16q22.1and 16q24.3 chromosomal loci in sporadic invasive breast carcinomas:correlation with proliferative activity, ploidy and hormonal status of thetumors. Anticancer Res 2001; 21: 991–9.
19 Cowin P, Rowlands TM, Hatsell SJ. Cadherins and catenins in breast cancer.Curr Opin Cell Biol 2005; 17: 499–508.
20 Caldeira JRF, Prando EC, Quevedo FC, Neto FAM, Rainho CA, Rogatto SR.CDH1 promoter hypermethylation and E-cadherin protein expression ininfiltrating breast cancer. BMC Cancer 2006; 6: 48.
21 Kashiwagi S, Yashiro M, Takashima T et al. Significance of E-cadherinexpression in triple-negative breast cancer. Br J Cancer 2010; 103: 249–55.
22 Geyer FC, Lacroix-Triki M, Savage K et al. b-Catenin pathway activation inbreast cancer is associated with triple-negative phenotype but not withCTNNB1 mutation. Mod Pathol 2011; 24: 209–31.
23 Nakopoulou L, Mylona E, Papadaki I et al. Study of phospho-beta-cateninsubcellular distribution in invasive breast carcinomas in relation to theirphenotype and the clinical outcome. Mod Pathol 2006; 19: 556–63.
24 Shi B, Liang J, Yang X et al. Integration of estrogen and Wnt signalingcircuits by the polycomb group protein EZH2 in breast cancer cells. Mol CellBiol 2007; 27: 5105–19.
25 Veeck J, Noetzel E, Bektas N et al. Promoter hypermethylation of the SFRP2gene is a high-frequent alteration and tumor-specific epigenetic marker inhuman breast cancer. Mol Cancer 2008; 7: 83.
Mukherjee et al.
26 Veeck J, Niederacher D, An H et al. Aberrant methylation of the Wntantagonist SFRP1 in breast cancer is associated with unfavorable prognosis.Oncogene 2006; 25: 3479–88.
27 Sobin LH, Wittekind C. TNM Classification of Malignant Tumors, 6th edn.New York: Wiley, 2002.
28 Dasgupta S, Mukherjee N, Roy S et al. Mapping of candidate tumorsuppressor genes’ loci on human chromosome 3 in head and neck squamouscell carcinoma of Indian patient population. Oral Oncol 2002; 38: 6–15.
29 Singh RK, Indra D, Mitra S et al. Deletions in chromosome 4 differentiallyassociated with the development of cervical cancer: evidence of slit2 as acandidate tumor suppressor gene. Hum Genet 2007; 122: 71–8.
30 Loginov VI, Maliukova AV, Seregin Iu A et al. Methylation of the promoterregion of the RASSF1A gene, a candidate tumor suppressor, in primaryepithelial tumors. Mol Biol (Mosk) 2004; 38: 654–67.
31 Reiner A, Neumeister B, Spona J, Reiner G, Schemper M, Jakesz R.Immunocytochemical localization of estrogen and progesterone receptor andprognosis in human primary breast cancer. Cancer Res 1990; 50: 7057–61.
32 Perrone F, Suardi S, Pastore E et al. Molecular and cytogenetic subgroups oforopharyngeal squamous cell carcinoma. Clin Cancer Res 2006; 12: 6643–51.
33 Dutta V, Chopra GS, Sahai K, Nema SK. Hormone receptors, Her-2 ⁄ Neu andchromosomal aberrations in breast cancer. MJAFI 2008; 64: 11–5.
34 Dey S, Boffetta P, Mathews A, Brennan P, Soliman A, Mathew A. Riskfactors according to estrogen receptor status of breast cancer patients inTrivandrum, South India. Int J Cancer 2009; 125: 1663–70.
35 Desai SB, Moonim MT, Gill AK, Punia RS, Naresh KN, Chinoy RF.Hormone receptor status of breast cancer in India: a study of 798 tumours.Breast 2000; 9: 267–70.
36 Sud R, Talbot IC, Delhanty JD. Infrequent alterations of the APC and MCCgenes in gastric cancers from British patients. Br J Cancer 1996; 74: 1104–8.
37 Stoehr R, Wissmann C, Suzuki H et al. Deletions of chromosome 8p and lossof sFRP1 expression are progression markers of papillary bladder cancer. LabInvest 2004; 84: 465–78.
38 Huiping C, Kristjansdottir S, Jonasson JG, Magnusson J, Egilsson V,Ingvarsson S. Alterations of E-cadherin and beta-catenin in gastric cancer.BMC Cancer 2001; 1: 16.
39 Nass SJ, Herman JG, Gabrielson E et al. Aberant methylation of the estrogenreceptor and E-cadherin 5¢ CpG islands increases with malignant progressionin human breast cancer. Cancer Res 2000; 60: 4346–8.
40 Henrique R, Ribeiro FR, Fonseca D et al. High promoter methylation levels ofAPC predict poor prognosis in sextant biopsies from prostate cancer patients.Clin Cancer Res 2007; 13: 6122–9.
41 Dahl E, Wiesmann F, Woenckhaus M et al. Frequent loss of SFRP1expression in multiple human solid tumours: association with aberrantpromoter methylation in renal cell carcinoma. Oncogene 2007; 26: 5680–91.
42 Knudson AG. Two genetic hits (more or less) to cancer. Nat Rev Cancer2001; 1: 157–62.
43 Segditsas S, Rowan AJ, Howarth K et al. APC and the three-hit hypothesis.Oncogene 2009; 28(1): 146–55.
44 Fostira F, Yannoukakos D. A distinct mutation on the alternative splice site ofAPC exon 9 results in attenuated familial adenomatous polyposis phenotype.Fam Cancer 2010; 9: 395–400.
45 Hill VK, Ricketts C, Bieche I et al. Genome-wide DNA methylation profilingof CpG islands in breast cancer identifies novel genes associated withtumorigenicity. Cancer Res 2011; 71: 2988–99.
46 Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherinpathways. Science 2004; 303: 1483–7.
47 Zhang F, White RL, Neufeld KL. Cell density and phosphorylation control thesubcellular localization of adenomatous polyposis coli protein. Mol Cell Biol2001; 21: 8143–56.
48 Khramtsov AI, Khramtsova GF, Tretiakova M, Huo D, Olopade OI, Goss KH.Wnt ⁄ beta-catenin pathway activation is enriched in basal-like breast cancersand predicts poor outcome. Am J Pathol 2010; 176: 2911–20.
49 Ryo A, Nakamura M, Wulf G, Liou YC, Lu KP. Pin1 regulates turnover andsubcellular localization of beta-catenin by inhibiting its interaction with APC.Nat Cell Biol 2001; 3: 793–801.
50 Lopez-Knowles E, Zardawi SJ, McNeil CM et al. Cytoplasmic localization ofbeta-catenin is a marker of poor outcome in breast cancer patients. CancerEpidemiol Biomarkers Prev 2010; 19(1): 301–9.
51 Maher MT, Mo R, Flozak AS, Peled ON, Gottardi CJ. Beta-cateninphosphorylated at serine 45 is spatially uncoupled from beta-cateninphosphorylated in the GSK3 domain: implications for signaling. PLoS ONE2010; 5: e10184.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Fig. S1. Flow chart demonstrating sample utilization for different analysis.
Cancer Sci | February 2012 | vol. 103 | no. 2 | 219ªª 2011 Japanese Cancer Association

Fig. S2. Expression pattern of estrogen and progesterone receptors in breast cancer of Indian patients.
Fig. S3. Alteration patterns of Wnt ⁄ b-catenin antagonists in stage I and II ⁄ III and IV and Grade I and I ⁄ III breast cancer.
Table S1. Summary of oligonucleotides.
Table S2. Antibodies and conditions used for immunohistochemical analyses.
Table S3. Alteration patterns of Wnt ⁄ b-catenin antagonists and b-catenin in breast cancer.
Table S4. Association of deletion between Wnt ⁄ b-catenin antagonists in breast cancer samples.
Table S5. Association of promoter methylation between Wnt ⁄ b-catenin antagonists in breast cancer.
Table S6. Clinicopathological correlation of the alterations (deletion and methylation) of Wnt ⁄ b-catenin antagonists in breast cancer.
Data S1. Additional methods including: sample collection and clinical data; microdissection and DNA extraction; deletion analysis; promotermethylation analysis; expression analysis by immunohistochemistry; and statistical analysis.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries(other than missing material) should be directed to the corresponding author for the article.
220 doi: 10.1111/j.1349-7006.2011.02131.xªª 2011 Japanese Cancer Association




















