
Crystal structure of the human beta2 adrenergic G-protein-coupled receptor
Upload
independentCategory
view
4download
0
doi:10.1152/ajpheart.00112.2010 298:1797-1806, 2010. First published Mar 26, 2010;Am J Physiol Heart Circ Physiol
Ravi Goyal, Ashwani Mittal, Nina Chu, Lubo Zhang and Lawrence D. Longo
You might find this additional information useful...
55 articles, 29 of which you can access free at: This article cites http://ajpheart.physiology.org/cgi/content/full/298/6/H1797#BIBL
including high-resolution figures, can be found at: Updated information and services http://ajpheart.physiology.org/cgi/content/full/298/6/H1797
can be found at: AJP - Heart and Circulatory Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpheart
This information is current as of June 18, 2010 .
http://www.the-aps.org/.ISSN: 0363-6135, ESSN: 1522-1539. Visit our website at Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the American Physiological Society. intact animal to the cellular, subcellular, and molecular levels. It is published 12 times a year (monthly) by the Americanlymphatics, including experimental and theoretical studies of cardiovascular function at all levels of organization ranging from the
publishes original investigations on the physiology of the heart, blood vessels, andAJP - Heart and Circulatory Physiology
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

�1-Adrenergic receptor subtype function in fetal and adult cerebral arteries
Ravi Goyal,1 Ashwani Mittal,1 Nina Chu,1 Lubo Zhang,1 and Lawrence D. Longo1,2
1Center for Perinatal Biology, Department of Physiology and Pharmacology, and 2Department of Obstetrics and Gynecology,School of Medicine, Loma Linda University, Loma Linda, California
Submitted 3 February 2010; accepted in final form 24 March 2010
Goyal R, Mittal A, Chu N, Zhang L, Longo LD. �1-Adrenergicreceptor subtype function in fetal and adult cerebral arteries. Am JPhysiol Heart Circ Physiol 298: H1797–H1806, 2010. First publishedMarch 26, 2010; doi:10.1152/ajpheart.00112.2010.—In the develop-ing fetus, cerebral artery (CA) contractility demonstrates significantfunctional differences from that of the adult. This may be a conse-quence of differential activities of �1-adrenergic receptor (�1-AR)subtypes. Thus we tested the hypothesis that maturational differencesin adrenergic-mediated CA contractility are, in part, a consequence ofdifferential expression and/or activities of �1-AR subtypes. In CAfrom fetal (�140 days) and nonpregnant adult sheep, we used wiremyography and imaging, with simultaneous measurement of tensionand intracellular Ca2� concentration ([Ca2�]i), radioimmunoassay,and Western immunoblots to examine phenylephrine (Phe)-inducedcontractile responses. The �1A-AR antagonists (5-MU and WB-4101)completely inhibited Phe-induced contraction in adult but not fetalCA; however, [Ca2�]i increase was reduced significantly in both agegroups. The �1D-AR antagonist (BMY-7378) blocked both Phe-induced contractions and Ca2� responses to a significantly greaterextent in adult compared with fetal CA. In both age groups, inhibitionof �1A-AR and �1B-AR, but not �1D-AR, significantly reducedinositol 1,4,5-trisphosphate responses to Phe. Western immunoblotsdemonstrated that the �1-AR subtype expression was only �20% infetal CA compared with the adult. Moreover, in fetal CA, the �1D-ARwas expressed significantly greater than the other two subtypes. Also,in fetal but not adult CA, Phe induced a significant increase inactivated ERK1/2; this increase in phosphorylated ERK was blockedby �1B-AR (CEC) and �1D-AR (BMY-7378) inhibitors, but not by�1A-AR inhibitors (5-MU or WB-4101). In conclusion, in the fetalCA, �1B-AR and �1D-AR subtypes play a key role in contractileresponse as well as in ERK activation. We speculate that in fetal CA�1B-AR and �1D-AR subtypes may be a critical factor associated withcerebrovascular growth and function.
vascular smooth muscle; development; maturation
THE ADRENERGIC PATHWAY is one of the main regulators ofcerebrovascular tone and cerebral blood flow (CBF) (33),mediating the majority of its effects through �-adrenergicreceptors (�-AR). In previous studies in fetal and adult ovinecerebral artery (CA), we have shown that �1-AR play a keyrole in contractile response (27, 28). In addition, �2-AR appearto be chiefly prejunctional, with the fetal CA having a signif-icant component of postjunctional �2-AR (1). However, in thecerebral vasculature of most species studied several reportsindicate that �1-AR are also abundant (6, 21, 41, 48), whichsuggests that cerebral vascular tone, and thus CBF, is regu-lated, in part, by activation/inhibition of these receptors. �1-ARbelong to the G protein-coupled receptor (GPCR) superfamilythat bind specifically to norepinephrine (NE) and epinephrine,and thus mediate sympathetic nervous system responses in
vascular smooth muscle cells (SMCs). �1-AR stimulation leadsto increase in inositol 1,4,5-trisphosphate [Ins(1,4,5)P3], whichin turn increases intracellular Ca2� concentration ([Ca2�]i) andacts as a major signaling pathway (Ca2�-dependent mecha-nism) mediating vascular contraction (5, 27). Other signalingpathways coupled to �1-AR include activation of protein ki-nase C (PKC) and extracellular signal-regulated kinases 1 and2 (ERK1/2) (Ca2� sensitization/independent mechanism) me-diating vascular contraction (4, 22). Our previous studies (51,52) have shown that �1-AR-mediated contraction of CA occursby both Ca2�-dependent and Ca2�-independent mechanisms.We also have shown that in ovine CA adrenergic-mediatedvasoconstrictor responses differ significantly in fetus and adult(27, 28, 51, 52). Recently, we have shown (8) significantchanges in the Ca2� sensitization pathway (PKC and ERK1/2)with developmental maturation from fetus to adult.
Radioligand binding and molecular cloning in several spe-cies have demonstrated that the �1-AR family has three struc-turally distinct subtypes (�1A, �1B, �1D), which are widelyexpressed in tissues and have different pharmacological prop-erties and amino acid sequences (36, 53). Although the three�1-adrenoceptor subtypes have been reported in various celltypes, little is known about their expression, physiologicalfunctions, or downstream pathways in CA (20, 22, 53). Ofnote, in fetal and adult CA we have demonstrated differingroles of NE (�1- and �2-AR agonist) and phenylephrine (Phe;�1-AR agonist) in contractility and downstream activation ofPKC and ERKs (8, 29, 51, 52). Thus the question arises as tothe role of the specific �1-AR subtypes and the role of down-stream effectors in �-adrenergic-mediated contraction in CA,and their changes with maturation. In the present study, wetested the hypothesis that in CA differential expression andactivation of �1-AR subtypes (�1A-, �1B-, and �1D-AR), infetus compared with adult differentially activate downstreamsignaling pathway responsible for contractile response and/orgrowth and development. The present study provides a deeperunderstanding of CBF regulation during fetal and adult life andprovides more specific targets for treatment of conditions suchas dysregulation of CBF.
METHODS
Experimental animals and tissues. All experimental procedureswere performed within the regulations of the Animal Welfare Act, theNational Institutes of Health Guide for the Care and Use of Labora-tory Animals, and the “Guiding Principles in the Care and Use ofAnimals” approved by the Council of the American PhysiologicalSociety and were approved by the Animal Care and Use Committee ofLoma Linda University. For these studies, we used CA from near-term fetal (�140 days) and nonpregnant adult (18–24 mo) sheepobtained from Nebeker Ranch (Lancaster, CA). For each experimentfive or more animals were used; in case of fetal twins n was countedas 1. Pregnant and nonpregnant ewes were anesthetized with thiopen-tal sodium (10 mg/kg iv), and anesthesia was maintained with inha-
Address for reprint requests and other correspondence: L. D. Longo, Centerfor Perinatal Biology, Loma Linda Univ., School of Medicine, Loma Linda,CA 92350 (e-mail: [email protected]).
Am J Physiol Heart Circ Physiol 298: H1797–H1806, 2010.First published March 26, 2010; doi:10.1152/ajpheart.00112.2010.
0363-6135/10 Copyright © 2010 the American Physiological Societyhttp://www.ajpheart.org H1797
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

lation of 1% isoflurane in oxygen throughout surgery. After the fetuswas delivered by hysterotomy, the fetuses and ewes were killed withan overdose of the proprietary euthanasia solution Euthasol (pento-barbital sodium 100 mg/kg and phenytoin sodium 10 mg/kg; Virbac,Fort Worth, TX). Studies were performed in isolated cerebral vesselscleaned of adipose and connective tissue. To avoid the complicationsof endothelium-mediated effects, we removed the endothelium bycarefully inserting a small wire three times, as previously described(26, 28). The vessels were used immediately for experiments.
Simultaneous measurement of [Ca2�]i and tension. We have de-scribed this technique in several reports (8, 24–26, 29). Briefly, fetalor adult middle CA rings of 2-mm length were incubated at 25°C for40 min with the acetoxymethyl ester of fura-2 (fura-2 AM; MolecularProbes, Eugene, OR), a fluorescent Ca2� indicator (24, 25, 26). Afterloading the dye, we mounted the arterial segments in a Jasco CAF-110, an intracellular Ca2� analyzer (Jasco, Easton, MD). We thenstabilized CA rings at 38°C for 40 min in an oxygenated standardKrebs solution. The bath chambers were continuously bubbled with95% O2-5% CO2, and all experiments were conducted at 38°C (coretemperature of sheep). After stabilization, on the basis of our previousstudies, the optimum resting tension was 0.6 g for fetal and 0.7 g foradult CA, because at these tensions the contractility response to 125mM KCl was maximum (28, 39). We then stimulated the isolated CArings from fetal and adult sheep with 125 mM KCl, and after thecontractile force plateaued 100 �M acetylcholine (ACh) was applied.The arteries that relaxed in the presence of ACh were discarded fromthe study, allowing for the evaluation of the relative quantity ofcontractile smooth muscle (34) and to determine the status of endo-thelium disruption (10). The contractile force due to 125 mM KCl wasmeasured as grams (g) of tension, before stimulation of the testcompound for each arterial segment, and was used to normalize thearterial contraction with other agonists and antagonists for variation inthe smooth muscle mass (26). For all vessels we evaluated thecontractile response for tension and fluorescence ratio by measuringthe maximum peak height and expressing it in both absolute terms andas percent Kmax (a measure of “efficacy”), and calculated pD2 (neg-ative logarithm of EC50, or half-maximal concentration, for NE or Pheand an index of tissue “sensitivity” or “potency”) (12, 26). As we havepreviously reported, although in absolute terms the maximal values offetal CA K�- and NE-induced tension are 20–30% less than those ofthe adult (25, 26), expressing these in terms of Kmax helps tonormalize the data and does not alter the interpretation of the results.In arteries used for response to a given agonist, i.e., NE (a nonselec-tive agonist for both �1- and �2-AR) or Phe (a selective agonist for�1-AR) and/or selective �1-AR (subtype) antagonists, we added theantagonist for 20 min before administration of NE or Phe. Unlessotherwise noted, all chemicals were obtained from Sigma Chemical(St. Louis, MO).
Role of �1-AR subtype blockers. Three pharmacologically distinct�1-AR subtypes can be distinguished with competitive antagonists andalkylating agents. The �1A-AR is shown to be inhibited by 2-(2,6-dimethoxyphenoxyethyl)aminomethyl-1,4-benzodioxane hydrochloride(WB-4101) (38), phentolamine (14), and 5-methyl-6[[3-[4-(2-methoxyphenyl)-1-piperazinyl]propyl]amino]-1,3-dimethyluracil (5-MU). Moreover, WB-4101 is a noncompetitive antagonist, whereas5-MU is a competitive antagonist and is more selective than otherpharmacological agents for �1A-AR (9, 11, 17, 18). Furthermore, studieshave demonstrated that the �1B-AR subtype is selectively inhibited bychlorethylclonidine (CEC) (14, 15, 35, 37). Other studies have estab-lished that 8-(2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl)-8-azaspiro(4,5)decane-7,9-dione (BMY-7378) selectively inhibits �1D-ARsubtype (7, 23, 40, 42). All of these antagonists are well characterized,with known IC50 for vascular adrenergic receptors. Therefore, on thebasis of these studies, we chose 5-MU and WB-4101 for �1A-AR, CECfor �1B-AR, and BMY-7378 for �1D-AR inhibition in the present study.First, using the fixed agonist, we performed dose-response curves withspecific �1-AR subtype antagonists to determine the pIC50 in sheep CA
(Table 1). Subsequently, to determine the role of the several subtypes inPhe-induced tension and [Ca2�]i, and the extent to which the selective�1-AR blockers inhibited the responses to Phe, we first measuredcontrol vascular tension and [Ca2�]i responses to 10�9–10�2 M Phe.Then after washout and 40 min reequilibration, we repeated the Phedose-response curve in the presence of an appropriate concentrationone of the �1-AR subtype blockers. For each agent we determined thedissociation constant KB.
Ins(1,4,5)P3 quantification. From each fetal and adult animal,cleaned CA was cut into segments weighing �20 mg (wet wt), placedin Krebs buffer, and bubbled with 95% O2-5% CO2 for 0.5 h. Weexposed the arteries to various treatments outlined above (Phe stim-ulation with selective �1-AR subtype inhibition) at 38°C according tothe protocol. Reactions were terminated by freezing in liquid N2, thefrozen samples were homogenized in 2 ml of iced 16% trichloroaceticacid with a glass grinder and centrifuged for 30 min at 1,500 g, thesupernatant was transferred, and the pellet was resuspended in 1 MNaOH for protein measurements (2). The supernatant was washed for30 s with H2O-saturated ether (5� volume, 2 times). After etherevaporation, we determined Ins(1,4,5)P3 mass by competitive ligandbinding assay in which a radioactive ligand competes with a nonra-dioactive ligand for a fixed number of receptor binding sites, as wehave described previously (27). [3H]Ins(1,4,5)P3 assay kits wereobtained from DuPont (Boston, MA); intra- and interassay coeffi-cients of variation were 7% and 9%, respectively.
Immunoblot of �1-AR subtypes. Isolated CA from fetal and adultsheep were cleaned of adventitia, and endothelium was denuded.These arteries were homogenized with a tissue grinder in ice-cold celllysis buffer (Cell Signaling Technology, Danvers, MA) as we havedescribed previously (29, 51). Protein concentrations were measuredwith a protein assay kit (Bio-Rad Laboratories, Hercules, CA), andbovine serum albumin (BSA) was used as a reference protein (2). TheMini Trans-Blot Electrophoretic Transfer Cell System (Bio-Rad Lab-oratories) was used to transfer proteins from the gel to a nitrocellulosemembrane at 100 V for 3 h. We then performed an overnightincubation of subtype-specific primary antibodies (1:500 dilution) for�1A-, �1B-, and �1D-AR (Santa Cruz Biotechnology, Santa Cruz, CA).We used �-actin as an internal control for equal protein loading aswell as the blocking peptide for each subtype-specific antibody as anegative control (Santa Cruz Biotechnology). The membrane was thenincubated in chemiluminescence luminol reagent (Pierce, Rockford,IL) for 1 min, and the protein band was detected with the AlphaInnotech Chemiluminescent imaging system (San Leandro, CA).After protein detection, the membrane was striped with strip buffer(Pierce) and immunoblotted by anti-�-actin antibody.
Immunoblotting of ERK1/2. After tension measurement, fetal andadult sheep CA were frozen rapidly in liquid N2. Frozen samples were
Table 1. pIC50 values of �1-adrenergic receptor subtypeinhibitors
Inhibitor Parameter Measured
IC50 Values
Adult Fetus
5-MU Contractility 7.4 � 0.1 7.7 � 0.1Ca2� signaling 7.3 � 0.1 7.7 � 0.1
WB-4101 Contractility 6.9 � 0.1 7.2 � 0.1Ca2� signaling 8.1 � 0.1 6.7 � 0.1
CEC Contractility 7.3 � 0.1 6.4 � 0.1Ca2� signaling 8.1 � 0.1 6.4 � 0.1
BMY-7378 Contractility 6.5 � 0.1 6.8 � 0.1Ca2� signaling 6.6 � 0.1 7.1 � 0.1
Values are means � SE. 5-MU, 5-methyl-6[[3-[4-(2-methoxyphenyl)-1-piperazinyl]propyl]amino]-1,3-dimethyluracil; WB-4101, 2-(2,6-dimethoxy-phenoxyethyl)aminomethyl-1,4-benzodioxane hydrochloride; CEC, chloreth-ylclonidine; BMY-7378, 8-(2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl)-8-azaspiro(4,5)decane-7,9-dione; pIC50, negative logarithm of 50% inhibitionconcentration values. (n � 4 to 5.) See METHODS for details.
H1798 �1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

homogenized in the 1� cell lysis buffer (Cell Signaling Technology)containing 1� phosphatase inhibitor cocktail and 1 mM phenylmeth-ylsulfonyl fluoride. Nuclei and debris were pelleted by centrifugationat 1,000 g for 10 min. The supernatant was collected and stored at�80°C. SDS-gel and Western immunoblots were performed, as men-tioned above for �1-AR immunoblot by using phospho- and total-ERK1/2 antibody (Cell Signaling Technology), as we have describedpreviously (8).
Statistical analysis. All values were calculated as means � SE. Inall cases, n values refer to the number of vessels (which correspondsto the number of animals and is included in Figs. 1–6) for a particularstudy. Because of the nature of these studies, we used severalstatistical tests to examine for significant differences. For testingdifferences between two groups, we used a simple unpaired Student’st-test. For multiple comparisons, we used one-way and two-wayanalysis of variance (ANOVA; vessel, age) coupled with Duncan’smultiple range test. Where appropriate, we used ANOVA with re-peated measures (Prism, GraphPad Software, La Jolla, CA). A P valueof 0.05 was considered significant.
RESULTS
Norepinephrine-induced CA contraction through �1-AR. Toestablish the extent to which NE mediates its effects through�1-AR in both fetal and adult CA, we measured NE-inducedcontractile and [Ca2�]i responses in the presence and absenceof the �1-AR antagonist prazosin (selective �1-AR blocker).As seen in Fig. 1, in the presence of prazosin (10�8 M), the pD2
values for NE responses of tension and fluorescence ratio forboth adult and fetal vessels were significantly decreased by 1log unit. Also, as shown in Table 2, in both fetal and adult CA,the dissociation constants (KB values) for prazosin-mediatedinhibition of NE-induced vascular contraction and [Ca2�]i
responses were similar, suggesting that both events were me-diated by the �1-AR.
Role of �1-AR subtypes in cerebrovascular contraction and[Ca2�]i responses. We measured the vascular tension and[Ca2�]i responses in the presence of selective inhibitors foreach of these subtypes. We first conducted �1-AR selectiveantagonist dose-response curves with 5-MU, WB-4101, CEC,and BMY-7378 in the presence of 10�2 M Phe. Table 1presents the pIC50 values of the several �1-AR subtype antag-onists. We then quantified �1-AR selective agonist Phe (10�9–10�2 M)-induced contraction and [Ca2�]i responses in thepresence and absence of �1A-AR antagonist 5-MU (10�8 M).As shown in Fig. 2, in the presence of 5-MU both fetal andadult CA showed a significant right shift of the Phe dose-response curve for contraction (Fig. 2, A and D), as well asincreases in [Ca2�]i (Fig. 2, B and E; see Table 2 for KB valuesand Fig. 2 for pD2 values). This demonstrates that in both fetaland adult CA �1A-AR play a significant role in Phe-inducedcontractile response, acting through a Ca2�-dependent path-way. Moreover, 5-MU had a greater effect in fetal CA com-pared with that in adult CA. To confirm this possibility further,we used a different �1A-AR antagonist, WB-4101 (10�7 M), totest Phe-induced tension and [Ca2�]i responses in anotherseries of CA. As shown in Fig. 2F for adult CA, the tensionresponses were similar to those in the presence of 5-MU (seeTable 2 for KB values). For fetal CA (Fig. 2C), the KB value forthe effect of WB-4101 on tension was similar to that of adultCA (Table 2).
To examine the effect of �1B-AR inhibition on Phe concen-tration-response curves, we performed a Phe dose-responsecurve in the presence and absence of the �1B-AR antagonistCEC (10�5 M). In adult CA, in the presence of CEC (10�5 M)both tension (Fig. 3C) and [Ca2�]i (Fig. 3D) were significantlyright-shifted. In contrast, in fetal CA 10�5 M CEC had nosignificant effect on the Phe-induced vascular tension (Fig. 3A)
Fig. 1. Norepinephrine (NE) dose-responserelations for fetal and adult cerebral arteries(CA) in the presence or absence of the �1-adrenergic receptor (�1-AR) antagonist pra-zosin. A and C: vascular tension (g) for fetalCA (A) and adult CA (C) in response to NEunder control conditions and after adminis-tration of 10�8 M prazosin (n � 4). B andD: fluorescence ratio (F340/380), i.e., intracel-lular Ca2� concentration ([Ca2�]i), for fetalCA (B) and adult CA (D) in response to NEunder control conditions and after additionof 10�8 M prazosin. pD2, negative logarithmof EC50. (n � 5 each.)
H1799�1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

and only modestly right-shifted the Phe dose response for[Ca2�]i (Fig. 3B); see Table 2 for CEC KB values. Thus in fetalCA, �1B-AR has significantly less role in Phe-induced contrac-tile response compared with adult CA.
To determine the extent to which �1D-AR inhibition alteredPhe-induced tension and [Ca2�]i responses, we measured thesevariables in response to increasing doses of Phe (10�9–10�2
M) in the absence or presence of the �1D-AR antagonistBMY-7378 (10�7 M). As shown in Fig. 4C, in response to
increasing Phe alone, adult CA showed a typical increase invascular tension. After �1D-AR inhibition, however, the max-imal Phe-induced contractile response and [Ca2�]i was atten-uated compared with control (P 0.01 for each; Fig. 4, C andD). As shown in Fig. 4A, in response to increasing Phe alone,fetal CA showed a typical contraction. In the presence of�1D-AR antagonist BMY-7378, the Phe-induced contractileand [Ca2�]i responses were decreased, but significantly less sothan those in adult (see Table 2 for KB values for BMY-7378).
Table 2. Dissociation constants (KB values) of antagonist inhibition
Receptor Drug Response Adult, M Fetus, M
�1 Prazosin (10�8 M) Contraction 6.4 � 10�10 1.1 � 10�10
[Ca2�]i 1.1 � 10�10 1.6 � 10�10
�1A 5-MU (10�8 M) Contraction 1.1 � 10�9 0.2 � 10�9
[Ca2�]i 1.1 � 10�9 0.7 � 10�9
�1B CEC (10�5 M) Contraction 5.3 � 10�10 No significant inhibition[Ca2�]i 0.8 � 10�10
�1D BMY-7378 (10�7 M) Contraction 6.7 � 10�9 No significant inhibition[Ca2�]i 25 � 10�9
[Ca2�]i, intracellular Ca2� concentration. See METHODS for details.
Fig. 2. Fetal and adult CA tension and [Ca2�]i
responses to phenylephrine (Phe) under con-trol conditions and in the presence of �1A-ARinhibition by 5-methyl-6[[3-[4-(2-methoxy-phenyl)-1-piperazinyl]propyl]amino]-1,3-dimethyluracil (5-MU, 10�8 M). A andD: vascular tensions (g) for fetal CA (A) andadult CA (D) in response to Phe alone and inthe presence of 10�8 M 5-MU (n � 4). B andE: fluorescence ratio (F340/380), i.e., [Ca2�]i forfetal CA (B) and adult CA (E) in response toPhe alone or after 5-MU. C and F: fetal (C)and adult (F) CA tension responses to Pheunder control conditions and in the presence of�1A-AR inhibition by 2-(2,6-dimethoxyphe-noxyethyl)aminomethyl-1,4-benzodioxane hy-drochloride (WB-4101). (n � 4 to 5 each.)
H1800 �1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

Protein abundance of �1-AR subtypes. Figure 5A illustratesthe �1-AR profiles from whole cell extracts that were subjectedto Western immunoblot analysis using enhanced chemilumi-nescence substrate, as described in METHODS. As shown, adultovine CA express relatively abundant amounts of each of thesubtype �1A-, �1B-, and �1D-AR. In contrast, near-term fetal
CA demonstrated low levels of the three �1-AR subtypes.Immunoblots of �-actin are shown as an internal control. Ofinterest, in fetal CA the �1D-AR subtype is expressed to arelatively greater degree than the other two subtypes (Fig. 5A).
�1-AR subtypes and Ins(1,4,5)P3 responses. To examine thedownstream signaling of the several �1-AR subtypes, we
Fig. 3. Adult and fetal CA tension and[Ca2�]i responses to Phe under control con-ditions and in the presence of �1B-AR inhi-bition by chlorethylclonidine (CEC). A andC: vascular tensions (g) for fetal (A) andadult (C) CA in response to Phe alone and inthe presence of 10�5 M CEC (n � 4 each).B and D: fluorescence ratio (F340/380) forfetal (B) and adult (D) in response to Phealone or after 10�5 M CEC.
Fig. 4. Fetal and adult CA tension and [Ca2�]i
responses to Phe under control conditions andin the presence of �1D-AR inhibition by 8-(2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl)-8-azaspiro(4,5)decane-7,9-dione (BMY-7378;BMY). A and C: vascular tension (g) for fetal(A) and adult (C) in response to Phe alone andin the presence of 10�7 M BMY-7378 (n � 4each). B and D: fluorescence ratio (F340/380) forfetal (B) and adult (D) CA in response to Phealone or after 10�7 M BMY-7378 (n � 4each).
H1801�1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from
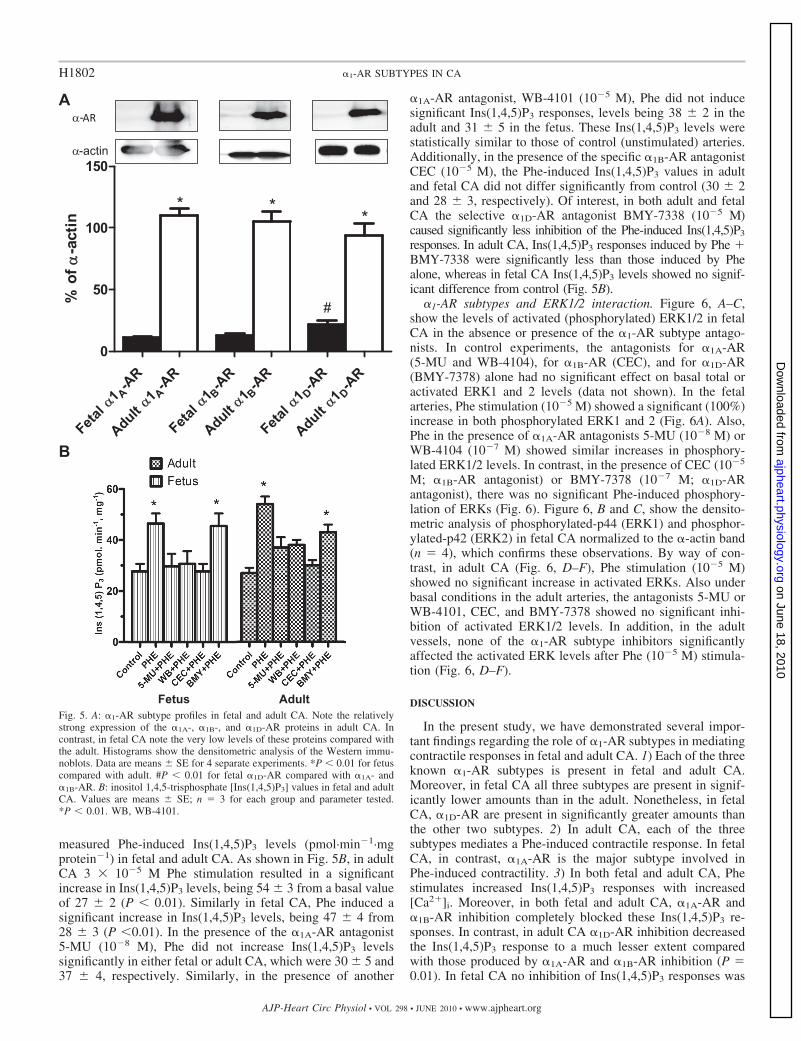
measured Phe-induced Ins(1,4,5)P3 levels (pmol·min�1·mgprotein�1) in fetal and adult CA. As shown in Fig. 5B, in adultCA 3 � 10�5 M Phe stimulation resulted in a significantincrease in Ins(1,4,5)P3 levels, being 54 � 3 from a basal valueof 27 � 2 (P 0.01). Similarly in fetal CA, Phe induced asignificant increase in Ins(1,4,5)P3 levels, being 47 � 4 from28 � 3 (P 0.01). In the presence of the �1A-AR antagonist5-MU (10�8 M), Phe did not increase Ins(1,4,5)P3 levelssignificantly in either fetal or adult CA, which were 30 � 5 and37 � 4, respectively. Similarly, in the presence of another
�1A-AR antagonist, WB-4101 (10�5 M), Phe did not inducesignificant Ins(1,4,5)P3 responses, levels being 38 � 2 in theadult and 31 � 5 in the fetus. These Ins(1,4,5)P3 levels werestatistically similar to those of control (unstimulated) arteries.Additionally, in the presence of the specific �1B-AR antagonistCEC (10�5 M), the Phe-induced Ins(1,4,5)P3 values in adultand fetal CA did not differ significantly from control (30 � 2and 28 � 3, respectively). Of interest, in both adult and fetalCA the selective �1D-AR antagonist BMY-7338 (10�5 M)caused significantly less inhibition of the Phe-induced Ins(1,4,5)P3
responses. In adult CA, Ins(1,4,5)P3 responses induced by Phe �BMY-7338 were significantly less than those induced by Phealone, whereas in fetal CA Ins(1,4,5)P3 levels showed no signif-icant difference from control (Fig. 5B).
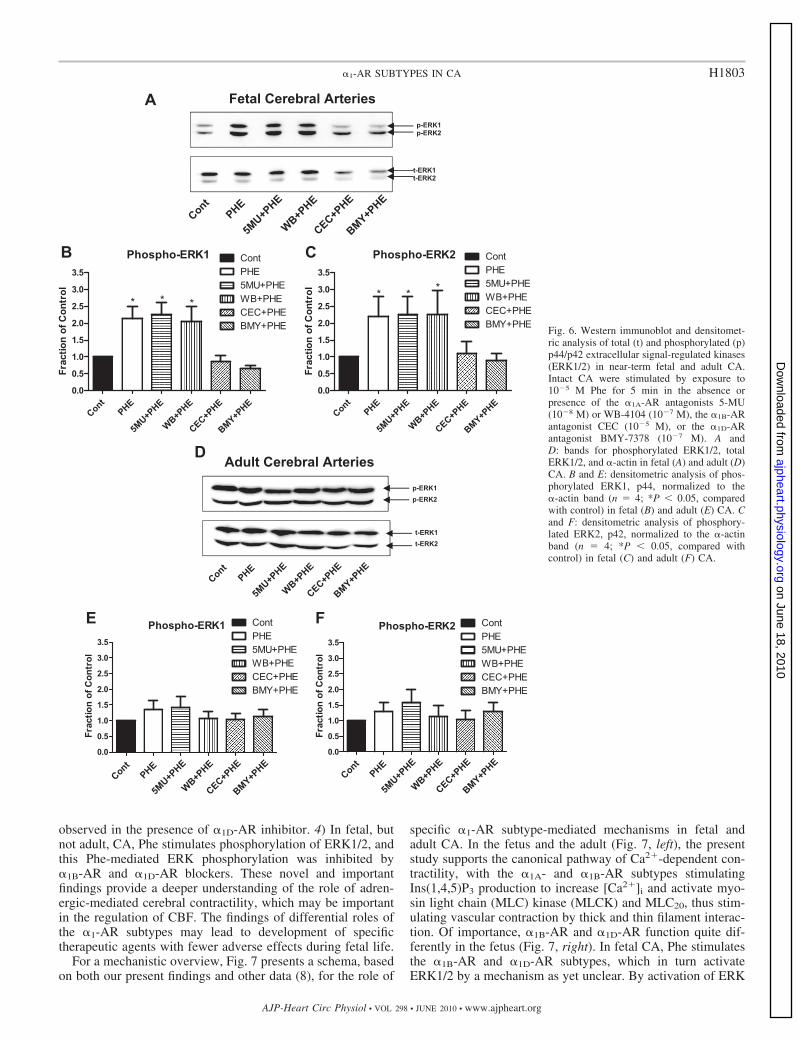
�1-AR subtypes and ERK1/2 interaction. Figure 6, A–C,show the levels of activated (phosphorylated) ERK1/2 in fetalCA in the absence or presence of the �1-AR subtype antago-nists. In control experiments, the antagonists for �1A-AR(5-MU and WB-4104), for �1B-AR (CEC), and for �1D-AR(BMY-7378) alone had no significant effect on basal total oractivated ERK1 and 2 levels (data not shown). In the fetalarteries, Phe stimulation (10�5 M) showed a significant (100%)increase in both phosphorylated ERK1 and 2 (Fig. 6A). Also,Phe in the presence of �1A-AR antagonists 5-MU (10�8 M) orWB-4104 (10�7 M) showed similar increases in phosphory-lated ERK1/2 levels. In contrast, in the presence of CEC (10�5
M; �1B-AR antagonist) or BMY-7378 (10�7 M; �1D-ARantagonist), there was no significant Phe-induced phosphory-lation of ERKs (Fig. 6). Figure 6, B and C, show the densito-metric analysis of phosphorylated-p44 (ERK1) and phosphor-ylated-p42 (ERK2) in fetal CA normalized to the �-actin band(n � 4), which confirms these observations. By way of con-trast, in adult CA (Fig. 6, D–F), Phe stimulation (10�5 M)showed no significant increase in activated ERKs. Also underbasal conditions in the adult arteries, the antagonists 5-MU orWB-4101, CEC, and BMY-7378 showed no significant inhi-bition of activated ERK1/2 levels. In addition, in the adultvessels, none of the �1-AR subtype inhibitors significantlyaffected the activated ERK levels after Phe (10�5 M) stimula-tion (Fig. 6, D–F).
DISCUSSION
In the present study, we have demonstrated several impor-tant findings regarding the role of �1-AR subtypes in mediatingcontractile responses in fetal and adult CA. 1) Each of the threeknown �1-AR subtypes is present in fetal and adult CA.Moreover, in fetal CA all three subtypes are present in signif-icantly lower amounts than in the adult. Nonetheless, in fetalCA, �1D-AR are present in significantly greater amounts thanthe other two subtypes. 2) In adult CA, each of the threesubtypes mediates a Phe-induced contractile response. In fetalCA, in contrast, �1A-AR is the major subtype involved inPhe-induced contractility. 3) In both fetal and adult CA, Phestimulates increased Ins(1,4,5)P3 responses with increased[Ca2�]i. Moreover, in both fetal and adult CA, �1A-AR and�1B-AR inhibition completely blocked these Ins(1,4,5)P3 re-sponses. In contrast, in adult CA �1D-AR inhibition decreasedthe Ins(1,4,5)P3 response to a much lesser extent comparedwith those produced by �1A-AR and �1B-AR inhibition (P �0.01). In fetal CA no inhibition of Ins(1,4,5)P3 responses was
Fig. 5. A: �1-AR subtype profiles in fetal and adult CA. Note the relativelystrong expression of the �1A-, �1B-, and �1D-AR proteins in adult CA. Incontrast, in fetal CA note the very low levels of these proteins compared withthe adult. Histograms show the densitometric analysis of the Western immu-noblots. Data are means � SE for 4 separate experiments. *P 0.01 for fetuscompared with adult. #P 0.01 for fetal �1D-AR compared with �1A- and�1B-AR. B: inositol 1,4,5-trisphosphate [Ins(1,4,5)P3] values in fetal and adultCA. Values are means � SE; n � 3 for each group and parameter tested.*P 0.01. WB, WB-4101.
H1802 �1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from
observed in the presence of �1D-AR inhibitor. 4) In fetal, butnot adult, CA, Phe stimulates phosphorylation of ERK1/2, andthis Phe-mediated ERK phosphorylation was inhibited by�1B-AR and �1D-AR blockers. These novel and importantfindings provide a deeper understanding of the role of adren-ergic-mediated cerebral contractility, which may be importantin the regulation of CBF. The findings of differential roles ofthe �1-AR subtypes may lead to development of specifictherapeutic agents with fewer adverse effects during fetal life.
For a mechanistic overview, Fig. 7 presents a schema, basedon both our present findings and other data (8), for the role of
specific �1-AR subtype-mediated mechanisms in fetal andadult CA. In the fetus and the adult (Fig. 7, left), the presentstudy supports the canonical pathway of Ca2�-dependent con-tractility, with the �1A- and �1B-AR subtypes stimulatingIns(1,4,5)P3 production to increase [Ca2�]i and activate myo-sin light chain (MLC) kinase (MLCK) and MLC20, thus stim-ulating vascular contraction by thick and thin filament interac-tion. Of importance, �1B-AR and �1D-AR function quite dif-ferently in the fetus (Fig. 7, right). In fetal CA, Phe stimulatesthe �1B-AR and �1D-AR subtypes, which in turn activateERK1/2 by a mechanism as yet unclear. By activation of ERK
Fig. 6. Western immunoblot and densitomet-ric analysis of total (t) and phosphorylated (p)p44/p42 extracellular signal-regulated kinases(ERK1/2) in near-term fetal and adult CA.Intact CA were stimulated by exposure to10�5 M Phe for 5 min in the absence orpresence of the �1A-AR antagonists 5-MU(10�8 M) or WB-4104 (10�7 M), the �1B-ARantagonist CEC (10�5 M), or the �1D-ARantagonist BMY-7378 (10�7 M). A andD: bands for phosphorylated ERK1/2, totalERK1/2, and �-actin in fetal (A) and adult (D)CA. B and E: densitometric analysis of phos-phorylated ERK1, p44, normalized to the�-actin band (n � 4; *P 0.05, comparedwith control) in fetal (B) and adult (E) CA. Cand F: densitometric analysis of phosphory-lated ERK2, p42, normalized to the �-actinband (n � 4; *P 0.05, compared withcontrol) in fetal (C) and adult (F) CA.
H1803�1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

directly (with or without involvement of other kinases),�1B-AR and �1D-AR in the fetus may play a critical role invascular development by stimulating maturation and growth ofSMCs. In addition, the �1B-AR subtype-mediated productionof diacylglycerol (DAG) activates PKC, which leads to phos-phorylation in MAPK and ERK1/2. Thus the several �1-ARsubtypes function in a very different manner in the fetuscompared with the adult. A grasp of these interactions, alongwith subtype-specific information, is essential for understand-ing the regulation of cerebrovascular tone and CBF. This isimportant, as the unique features of these subtypes may play arole in dysregulation of CBF in the fetus and prematurenewborn, as well as in the adult. Quite obviously, we yet havemuch to learn in regard to �1-AR subtypes and their changingrole with maturation. A caveat of these studies concerns therelative selectivity of the purported pharmacological antago-nists for the several �1-AR subtypes (53), because subtype-selective agents may not be overly selective and some interactwith other adrenergic and nonadrenergic receptors. Both�1A-AR antagonists, 5-MU (11, 31) and WB-4101 (38), arerelatively selective, although they differ in action. In turn, thealkylating agent CEC (16) and BMY-7378 (7) have beendemonstrated to be quite selective antagonists for �1B-AR and�1D-AR, respectively.
�1-AR subtype expression and vascular reactivity. The In-ternational Union of Pharmacology has designated �1-ARsubtypes as �1A/a, �1B/b, and �1D/d, with the uppercase lettersrepresenting the pharmacologically identified subtypes and thelowercase letters representing the subtype molecular clones(19). Important mediators of sympathetic nervous response, the�1-AR are widely distributed throughout many cell types, andin SMCs regulate vascular tone and blood flow (53). In SMCs,the �1-AR subtypes and their levels vary considerably in theirdistribution in various circulatory beds, in different-size ves-
sels of a given vascular bed, and in different species (16, 36,41, 48). The present findings of a paucity of expression of theseveral �1-AR subtypes in the fetal CA agrees with the conceptof an increase in �1-AR density with development (13, 32). Forinstance, in rats with aging from 1 mo to 24 mo, both �1B-ARmRNA and protein levels of �1A-AR increased, while �1D-ARlevels remained constant (13). Moreover, in another report,after 10 wk of age �1D-AR have been reported to decrease, sothat by 12 mo of age vasoconstrictor response to the �1A-ARsubtype was predominant (32). Also, in comparing 7-day-oldand adult rats, each of the three subtypes increased in braintissue, �1B-AR increased in heart but decreased in liver, andnone of the subtypes changed significantly in kidney (43).
�1-AR subtypes and intracellular signaling. Differences in�1-AR subtype expression levels with development would beanticipated to have important physiological significance. More-over, coupling of different �1-AR with different signal trans-duction pathways increases such significance. Relatively fewstudies have examined these relations in the vascular smoothmuscle, and none to our knowledge in CA, much less compar-ing the immature organism with the adult. In adult but not infetal CA, it has been well documented that the three �1-ARsubtypes, members of the Gq/11 protein-coupled receptors,activate PLC-� to hydrolyze phosphatidylinositol 4,5-bisphos-phate to Ins(1,4,5)P3 and DAG, the latter of which, in turn,activates PKC. �1D-AR inhibition appears to cause modestdecrease in Phe-induced Ins(1,4,5)P3 responses, however, thecontractility and Ca2� responses being inhibited to a significantextent. �1D-AR may be mediating these effects through othersignal transduction pathways such as RhoA/Rho kinase and/oractivation of L-type Ca2� channels and so forth.
In fetal but not adult CA we also observed a pronouncedincrease in ERK phosphorylation with Phe, which was inhib-ited by �1B-AR and �1D-AR antagonists. Of interest, recentlywe have shown (8) that PKC also differentially activates ERKin fetal and adult CA. Other reports indicate that GPCR such as�1-AR acting through Gq/11 also can activate the MAPKpathway via PKC and/or the Ras/Raf pathway by adapterproteins (30, 54). In 1- to 2-day-old rat cardiac myocytes,MAPK phosphorylation appeared to be mediated by the�1B-AR (45). To our knowledge, no studies in cerebrovascularSMCs have examined the relation of �1-AR subtypes to acti-vation of the MAPK cascade and downstream effects of cellgrowth and differentiation, and this requires further investiga-tion. In cultured rat aortic SMCs, protein synthesis and hyper-trophy appeared to be mediated by the prominent �1B-AR (3).Similarly in rabbit aortas, �1B-AR stimulated protein synthesis(44). Along this line, in cultured rat thoracic aorta SMCs stablytransfected with full-length �1-AR cDNAs, the �1D-AR ap-peared to couple to MAPK-mediated protein synthesis and cellgrowth, and these responses could be inhibited by the MEKinhibitor PD-98059 (46).
Perspectives and significance. The present study extends ourunderstanding of the adrenergic receptor pathway in CA con-tractility. It is a logical extension of our previous findings andillustrates a small facet of this complexity, e.g., that having todo with developmental maturation. The study provides detailsregarding maturational shift in expression and functions of�1-AR subtypes, as well as coupling with downstream path-ways. However, this study makes more apparent the complex-ity of intracellular signaling pathways and raises a number of
Fig. 7. Proposed signal transduction pathways for specific �1-AR subtype-mediated Ca2�-dependent and Ca2�-independent smooth muscle cell (SMC)contraction in adult and fetal CA. Left: �1A-AR and �1B-AR subtypes mediateproduction of Ins(1,4,5)P3, with the result of increasing intracellular Ca2�
release, and effect contraction in the Ca2�-dependent pathway in both fetusand adult. In addition, �1D-AR with dashed line indicates activation ofIns(1,4,5)P3 pathway only in adult. Right: fetus, in contrast to adult, �1B- and�1D-AR activate PKC, which in turn activates the extracellular regulatedkinases ERK1/2 to stimulate gene expression to result in SMC differentiationand vascular growth.
H1804 �1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

new questions. For instance, what is the functional significanceof �1B- and �1D-AR during fetal life? To what extent do the Gprotein coupling pathways differ in the maturing fetus fromthose of the adult? During fetal life, to what extent is ERKactivation responsible for contractility and/or vascular growth?What are the pathways involved in �1D-AR-mediated contrac-tile responses in adult CA? To what extent do �1-AR mediateangiogenesis in the adult? If not via MAPK, then what are thepathways involved? Insight into these pathways and physio-logical mechanisms may prove of great value in developingtherapeutic interventions in preventing and/or ameliorating thesequelae of functional dysregulation of CBF, such as occur inthe developing fetus and newborn infant.
ACKNOWLEDGMENTS
We thank Wen Long and Yu Zhao for technical assistance and BrendaKreutzer for preparing the manuscript.
GRANTS
This work was supported by National Institutes of Health Grant HD/HL-03807 to L. D. Longo.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
REFERENCES
1. Bishai JM, Penninga L, Nijland R, Meulenaar R, Gheorghe CP, ZhaoY, Buchholz JN, Zhang L, Longo LD. Pre- and postjunctional alpha2-adrenergic receptors in fetal and adult ovine cerebral arteries. Am J PhysiolRegul Integr Comp Physiol 282: R1654–R1662, 2002.
2. Bradford MM. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72: 248–254, 1976.
3. Chen L, Xin X, Eckhart AD, Yang N, Faber JE. Regulation of vascularsmooth muscle growth by alpha1-adrenoreceptor subtypes in vitro and insitu. J Biol Chem 270: 30980–30988, 1995.
4. Cobb MH, Robbins DJ, Boulton TG. ERKs, extracellular signal-regu-lated MAP-2 kinases. Curr Opin Cell Biol 3: 1025–1032, 1991.
5. Exton JH. Regulation of phosphoinositide phospholipases by hormones,neurotransmitters, and other agonists linked to G proteins. Annu RevPharmacol Toxicol 36: 481–509, 1996.
6. Fagura MS, Lydford SJ, Dougall IG. Pharmacological classification ofalpha1-adrenoceptors mediating contractions of rabbit isolated ear artery:comparison with rat isolated thoracic aorta. Br J Pharmacol 120: 247–258,1997.
7. Goetz AS, King HK, Ward SD, True TA, Rimele TJ, Saussy DL Jr.BMY 7378 is a selective antagonist of the D subtype of alpha1-adreno-ceptors. Eur J Pharmacol 272: R5–R6, 1995.
8. Goyal R, Mittal A, Chu N, Shi L, Zhang L, Longo LD. Maturation andthe role of PKC-mediated contractility in ovine cerebral arteries. Am JPhysiol Heart Circ Physiol 297: H2242–H2252, 2009.
9. Graziadei I, Zernig G, Boer R, Glossman H. Stereoselective binding ofniguldipine enantiomers to alpha1A-adrenoceptors labeled with [3H]5-methyl-urapidil. Eur J Pharmacol 172: 329–337, 1989.
10. Greenberg B, Kishiyama S. Endothelium-dependent and -independentresponses to severe hypoxia in rat pulmonary artery. Am J Physiol HeartCirc Physiol 265: H1712–H1720, 1993.
11. Gross G, Hanft G, Rugevics C. 5-Methyl-urapidil discriminates betweensubtypes of the alpha1-adrenoceptor. Eur J Pharmacol 151: 333–335,1988.
12. Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2�
indicators with greatly improved fluorescence properties. J Biol Chem260: 3440–3450, 1985.
13. Gurdal H, Tilakaratne N, Brown RD, Fonseca M, Friedman E,Johnson MD. The expression of alpha1 adrenoceptor subtypes changeswith age in the rat aorta. J Pharmacol Exp Ther 275: 1656–1662, 1995.
14. Han C, Abel PW, Minneman KP. Alpha1-adrenoceptor subtypes linkedto different mechanisms for increasing intracellular Ca2� in smoothmuscle. Nature 329: 333–335, 1987.
15. Han C, Abel PW, Minneman KP. Heterogeneity of alpha1-adrenergicreceptors revealed by chlorethylclonidine. Mol Pharmacol 32: 505–510,1987.
16. Han C, Li J, Minneman KP. Subtypes of alpha1-adrenoceptors in ratblood vessels. Eur J Pharmacol 190: 97–104, 1990.
17. Hanft G, Gross G. Subclassification of alpha 1-adrenoceptor recognitionsites by urapidil derivatives and other selective antagonists. Br J Phar-macol 97: 691–700, 1989.
18. Hanft G, Gross G, Beckeringh JJ, Korstanje C. Alpha1-adrenoceptors:the ability of various agonists and antagonists to discriminate between twodistinct [3H]prazosin binding sites. J Pharm Pharmacol 41: 714–716,1989.
19. Hieble JP, Bylund DB, Clarke DE, Eikenburg DC, Langer SZ,Lefkowitz RJ, Minneman KP, Ruffolo RR Jr. International Union ofPharmacology. X. Recommendation for nomenclature of alpha1-adreno-ceptors: consensus update. Pharmacol Rev 47: 267–270, 1995.
20. Hirasawa A, Awaji T, Xu Z, Shinoura H, Tsujimoto G. Regulation ofsubcellular localization of alpha1-adrenoceptor subtypes. Life Sci 68:2259–2267, 2001.
21. Hussain MB, Marshall I. Characterization of alpha1-adrenoceptor sub-types mediating contractions to phenylephrine in rat thoracic aorta, mes-enteric artery and pulmonary artery. Br J Pharmacol 122: 849–858, 1997.
22. Jiao X, Gonzalez-Cabrera PJ, Xiao L, Bradley ME, Abel PW, JeffriesWB. Tonic inhibitory role for cAMP in alpha1a-adrenergic receptorcoupling to extracellular signal-regulated kinases 1/2. J Pharmacol ExpTher 303: 247–256, 2002.
23. Kenny BA, Chalmers DH, Philpott PC, Naylor AM. Characterization ofan alpha1D-adrenoceptor mediating the contractile response of rat aorta tonoradrenaline. Br J Pharmacol 115: 981–986, 1995.
24. Long W, Zhang L, Longo LD. Cerebral artery KATP- and KCa-channelactivity and contractility: changes with development. Am J Physiol RegulIntegr Comp Physiol 279: R2004–R2014, 2000.
25. Long W, Zhang L, Longo LD. Cerebral artery sarcoplasmic reticulumCa2� stores and contractility: changes with development. Am J PhysiolRegul Integr Comp Physiol 279: R860–R873, 2000.
26. Long W, Zhao Y, Zhang L, Longo LD. Role of Ca2� channels inNE-induced increase in [Ca2�]i and tension in fetal and adult cerebralarteries. Am J Physiol Regul Integr Comp Physiol 277: R286–R294, 1999.
27. Longo LD, Ueno N, Zhao Y, Pearce WJ, Zhang L. Developmentalchanges in alpha1-adrenergic receptors, IP3 responses, and NE-inducedcontraction in cerebral arteries. Am J Physiol Heart Circ Physiol 271:H2313–H2319, 1996.
28. Longo LD, Ueno N, Zhao Y, Zhang L, Pearce WJ. NE-inducedcontraction, alpha1-adrenergic receptors, and Ins(1,4,5)P3 responses incerebral arteries. Am J Physiol Heart Circ Physiol 270: H915–H923, 1996.
29. Longo LD, Zhao Y, Long W, Miguel C, Windemuth RS, CantwellAM, Nanyonga AT, Saito T, Zhang L. Dual role of PKC in modulatingpharmacomechanical coupling in fetal and adult cerebral arteries. Am JPhysiol Regul Integr Comp Physiol 279: R1419–R1429, 2000.
30. Lopez-Ilasaca M, Crespo P, Pellici PG, Gutkind JS, Wetzker R.Linkage of G protein-coupled receptors to the MAPK signaling pathwaythrough PI 3-kinase gamma. Science 275: 394–397, 1997.
31. Low AM, Lu-Chao H, Wang YF, Brown RD, Kwan CY, Daniel EE.Pharmacological and immunocytochemical characterization of subtypes ofalpha-1 adrenoceptors in dog aorta. J Pharmacol Exp Ther 285: 894–901,1998.
32. Lu ZZ, Zhang YY, Han QD. Comparison of functional alpha1-adreno-ceptor subtypes in aortae between 12 month- and 10 week-old rats. ShengLi Xue Bao 49: 414–418, 1997.
33. McCalden TA. Sympathetic control of the cerebral circulation. J AutonPharmacol 1: 421–431, 1981.
34. McDonald TF, Pelzer S, Trautwein W, Pelzer DJ. Regulation andmodulation of calcium channels in cardiac, skeletal, and smooth musclecells. Physiol Rev 74: 365–507, 1994.
35. Minneman KP, Atkinson B. Interaction of subtype-selective antagonistswith alpha1-adrenergic receptor-mediated second messenger responses inrat brain. Mol Pharmacol 40: 523–530, 1991.
36. Minneman KP, Esbenshade TA. Alpha1-adrenergic receptor subtypes.Annu Rev Pharmacol Toxicol 34: 117–133, 1994.
37. Minneman KP, Han C, Abel PW. Comparison of alpha1-adrenergicreceptor subtypes distinguished by chlorethylclonidine and WB 4101. MolPharmacol 33: 509–514, 1988.
H1805�1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from

38. Morrow AL, Creese I. Characterization of alpha1-adrenergic receptorsubtypes in rat brain: a reevaluation of [3H]WB4104 and [3H]prazosinbinding. Mol Pharmacol 29: 321–330, 1986.
39. Pearce WJ, Hull AD, Long DM, Longo LD. Developmental changes inovine cerebral artery composition and reactivity. Am J Physiol RegulIntegr Comp Physiol 261: R458–R465, 1991.
40. Piascik MT, Guarino RD, Smith MS, Soltis EE, Saussy DL Jr, PerezDM. The specific contribution of the novel alpha-1D adrenoceptor to thecontraction of vascular smooth muscle. J Pharmacol Exp Ther 275:1583–1589, 1995.
41. Rudner XL, Berkowitz DE, Booth JV, Funk BL, Cozart KL, D’AmicoEB, El-Moalem H, Page SO, Richardson CD, Winters B, Marucci L,Schwinn DA. Subtype specific regulation of human vascular alpha1-adrenergic receptors by vessel bed and age. Circulation 100: 2336–2343,1999.
42. Saussy DL Jr, Goetz AS, Queen KL, King HK, Lutz MW, Rimele TJ.Structure activity relationships of a series of buspirone analogs at alpha-1adrenoceptors: further evidence that rat aorta alpha-1 adrenoceptors are ofthe alpha-1D-subtype. J Pharmacol Exp Ther 278: 136–144, 1996.
43. Shen H, Peri KG, Deng XF, Chemtob S, Varma DR. Distribution ofalpha1-adrenoceptor subtype proteins in different tissues of neonatal andadult rats. Can J Physiol Pharmacol 78: 237–243, 2000.
44. Siwik DA, Brown RD. Regulation of protein synthesis by alpha1-adrenergic receptor subtypes in cultured rabbit aortic vascular smoothmuscle cells. J Cardiovasc Pharmacol 27: 508–518, 1996.
45. Wenham D, Rahmatullah RJ, Rahmatullah M, Hansen CA, RobishawJD. Differential coupling of alpha1-adrenoreceptor subtypes to phospho-lipase C and mitogen activated protein kinase in neonatal rat cardiacmyocytes. Eur J Pharmacol 339: 77–86, 1997.
46. Xin X, Yang N, Eckhart AD, Faber JE. Alpha1D-adrenergic receptorsand mitogen-activated protein kinase mediate increased protein synthesisby arterial smooth muscle. Mol Pharmacol 51: 764–775, 1997.
47. Yamamoto Y, Koike K. Characterization of alpha1-adrenoceptor-medi-ated contraction in the mouse thoracic aorta. Eur J Pharmacol 424:131–140, 2001.
48. Yokoo H, Kobayashi H, Minami S, Shiraishi S, Yamamoto R,Yanagita T, Tsuchiya K, Mohri M, Wada A. Alpha1-adrenergic recep-tor subtypes in rat cerebral microvessels. Brain Res 878: 183–187, 2000.
49. Zhang H, Xiao D, Longo LD, Zhang L. Regulation of alpha1-adreno-ceptor-mediated contractions of uterine arteries by PKC: effect of preg-nancy. Am J Physiol Heart Circ Physiol 291: H2282–H2289, 2006.
50. Zhang L, Pearce WJ, Longo LD. Noradrenaline-mediated contractionsof ovine uterine artery: role of inositol 1,4,5-trisphosphate. Eur J Phar-macol 289: 375–382, 1995.
51. Zhao Y, Long W, Zhang L, Longo LD. Extracellular signal-regulatedkinases and contractile responses in ovine adult and fetal cerebral arteries.J Physiol 551: 691–703, 2003.
52. Zhao Y, Zhang L, Longo LD. PKC-induced ERK1/2 interactions anddownstream effectors in ovine cerebral arteries. Am J Physiol Regul IntegrComp Physiol 289: R164–R171, 2005.
53. Zhong H, Minneman KP. Alpha1-adrenoceptor subtypes. Eur J Phar-macol 375: 261–276, 1999.
54. Zhong H, Minneman KP. Differential activation of mitogen-activatedprotein kinase pathways in PC12 cells by closely related alpha1-adrenergicreceptor subtypes. J Neurochem 72: 2388–2396, 1999.
55. Zhou L, Zhao Y, Nijland R, Zhang L, Longo LD. Ins(1,4,5)P3 receptorsin cerebral arteries: changes with development and high-altitude hypoxia.Am J Physiol Regul Integr Comp Physiol 272: R1954–R1959, 1997.
H1806 �1-AR SUBTYPES IN CA
AJP-Heart Circ Physiol • VOL 298 • JUNE 2010 • www.ajpheart.org
on June 18, 2010 ajpheart.physiology.org
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















