
Subcutaneous Allergic Sensitization to Protease Allergen Is ...
12
of January 11, 2022. This information is current as Subcutaneous and Intranasal Routes but Not IL-33: Distinct Mechanisms between Cells Protease Allergen Is Dependent on Mast Subcutaneous Allergic Sensitization to Ikeda, Hideoki Ogawa, Ko Okumura and Toshiro Takai Saito, Susumu Nakae, Hajime Suto, Saori Ichikawa, Shigaku Hirono Ochi, Natsuko Maruyama, Akira Matsuda, Hirohisa Seiji Kamijo, Mayu Suzuki, Mutsuko Hara, Sakiko Shimura, ol.1500717 http://www.jimmunol.org/content/early/2016/03/18/jimmun published online 21 March 2016 J Immunol Material Supplementary 7.DCSupplemental http://www.jimmunol.org/content/suppl/2016/03/18/jimmunol.150071 average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2016 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on January 11, 2022 http://www.jimmunol.org/ Downloaded from by guest on January 11, 2022 http://www.jimmunol.org/ Downloaded from
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Subcutaneous Allergic Sensitization to Protease Allergen Is ...

of January 11, 2022.This information is current as
Subcutaneous and Intranasal Routesbut Not IL-33: Distinct Mechanisms between
CellsProtease Allergen Is Dependent on Mast Subcutaneous Allergic Sensitization to
Ikeda, Hideoki Ogawa, Ko Okumura and Toshiro TakaiSaito, Susumu Nakae, Hajime Suto, Saori Ichikawa, ShigakuHirono Ochi, Natsuko Maruyama, Akira Matsuda, Hirohisa Seiji Kamijo, Mayu Suzuki, Mutsuko Hara, Sakiko Shimura,
ol.1500717http://www.jimmunol.org/content/early/2016/03/18/jimmun
published online 21 March 2016J Immunol
MaterialSupplementary
7.DCSupplementalhttp://www.jimmunol.org/content/suppl/2016/03/18/jimmunol.150071
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2016 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Subcutaneous Allergic Sensitization to Protease Allergen IsDependent on Mast Cells but Not IL-33: Distinct Mechanismsbetween Subcutaneous and Intranasal Routes
Seiji Kamijo,* Mayu Suzuki,*,† Mutsuko Hara,* Sakiko Shimura,*,‡ Hirono Ochi,*,‡
Natsuko Maruyama,*,‡ Akira Matsuda,x Hirohisa Saito,{ Susumu Nakae,‖,#
Hajime Suto,*,‡ Saori Ichikawa,† Shigaku Ikeda,*,‡ Hideoki Ogawa,* Ko Okumura,* and
Toshiro Takai*
Protease activity of papain, a plant-derived occupational allergen homologous to mite major allergens, is essential to IgE/IgG1 pro-
duction and lung eosinophilia induced by intranasal papain administration in mice, and IL-33 contributes to these responses. In this
work, we investigate skin and Ab responses induced by s.c. papain administration into ear lobes and responses induced by subsequent
airway challenge with papain. Subcutaneous papain injection induced swelling associated with increased epidermal thickness, dermal
inflammation, serum IgE/IgG1 responses, and Th2 cytokine production in draining lymph node cells restimulated in vitro. These
responses were markedly less upon s.c. administration of protease inhibitor-treated papain. Results obtained by using mast cell–
deficient mice and reconstitution of tissue mast cells suggested the contribution of mast cells to papain-specific IgE/IgG1 responses
and eosinophil infiltration. The responses were equivalent between wild-type and IL-332/2 mice. After the subsequent airway
challenge, the s.c. presensitized wild-type mice showed more severe lung eosinophilia than those without the presensitization. The
presensitized IL-332/2 mice showed modest lung eosinophilia, which was absent without the presensitization, but its severity
and IgE boost by the airway challenge were markedly less than the presensitized wild-type mice, in which protease activity of
inhaled papain contributed to the responses. The results suggest that mechanisms for the protease-dependent sensitization
differ between skin and airway and that cooperation of mast cell–dependent, IL-33–independent initial sensitization via skin
and protease-induced, IL-33–mediated mechanism in re-exposure via airway to protease allergens maximizes the magnitude of
the transition from skin inflammation to asthma in natural history of progression of allergic diseases. The Journal of Immu-
nology, 2016, 196: 000–000.
Various factors found in the environment have beenreported as triggers of allergic sensitization, which ischaracterized by Th2-dominant immune response and
induction of allergen-specific IgE (1, 2). House dust mites (HDM),insects, pollen, fungi, bacterial components in house dust, animal
dander, and enzymes used in the food industry are examples ofallergenic environmental factors. One of the important proper-
ties of these allergenic environmental factors is protease activ-
ity (1–3). The protease activity of HDM allergens has been
shown to cause barrier dysfunction, to induce the production of
proinflammatory cytokines in epithelial cells and keratinocytes,
to cleave various molecules, to modulate functions of various
cell types, and to induce Th2 responses and IgE production (3–
10). HDM group 1 allergens, Der f 1 and Der p 1, and papain, a
papaya fruit–derived occupational allergen, belong to the same
family of cysteine proteases, family C1 of clan CA (5, 11, 12).
Protease activity of papain (10, 13) and recombinant Der f 1
(rDer f 1) (10) is essential to airway eosinophilia and serum
IgE/IgG1 responses induced by inhalation of the protease al-
lergens. In recent studies, papain has been used as a model
protease allergen mimicking those contained in allergen sources
(10, 13–16).Protease allergens stimulate various cell types (3, 4, 17). Mast
cell deficiency did not reduce the IgE/IgG1 responses and allergic
airway inflammation induced by papain (10). Some murine
models suggested contribution of basophils in papain- or HDM-
induced allergic responses in vivo (15, 18, 19), although still
controversial (10, 18, 20).An epithelial cell–derived IL-1 family cytokine, IL-33, is
implicated in allergic and type 2 immune responses in various
models (10, 21–27). IL-33–mediated innate immunity is crit-
ical for the lung eosinophilia induced by high-dose, 3-day in-
*Atopy (Allergy) Research Center, Juntendo University Graduate School of Medi-cine, Tokyo 113-8421, Japan; †Department of Materials and Biological Sciences,Faculty of Science, Japan Women’s University, Tokyo 112-8681, Japan;‡Department of Dermatology and Allergology, Juntendo University Graduate Schoolof Medicine, Tokyo 113-8421, Japan; xDepartment of Ophthalmology, Juntendo Uni-versity Graduate School of Medicine, Tokyo 113-8421, Japan; {Department of Al-lergy and Immunology, National Research Institute for Child Health andDevelopment, Tokyo 157-8535, Japan; ‖Laboratory of Systems Biology, Center forExperimental Medicine and Systems Biology, The Institute of Medical Science, TheUniversity of Tokyo, Tokyo 108-8639, Japan; and #Precursory Research for Embry-onic Science and Technology, Japan Science and Technology Agency, Saitama 332-0012, Japan
Received for publication June 5, 2015. Accepted for publication February 19, 2016.
This work was supported by the Ministry of Education, Culture, Sports, Science, andTechnology (MEXT)-supported Program for Strategic Research Foundation at Pri-vate Universities and Grants-in-Aid for Scientific Research from MEXT.
Address correspondence and reprint requests to Dr. Toshiro Takai, Atopy (Allergy)Research Center, Juntendo University Graduate School of Medicine, 2-1-1 Hongo,Bunkyo-ku, Tokyo 113-8421, Japan. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: AD, atopic dermatitis; BAL, bronchial alveolarlavage; BMMC, bone marrow–derived mast cell; DLN, draining lymph node; HDM,house dust mite; ILC2, group 2 innate lymphoid cell; i.n., intranasal; rDer f 1,recombinant Der f 1; TSLP, thymic stromal lymphopoietin.
Copyright� 2016 by The American Association of Immunologists, Inc. 0022-1767/16/$30.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1500717
Published March 21, 2016, doi:10.4049/jimmunol.1500717 by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
halation of papain (22). Serum Ab production and lung eosinophiliain another model of lower-dose papain inhalation for a longer pe-riod than a week are dependent on IL-33 and protease activity, inwhich the cooperation of IL-33–mediated innate responses andadaptive immune cells is required for robust eosinophilic lung in-flammation (10). Halim et al. (28, 29) revealed that the IL-33–group2 innate lymphoid cell (ILC2) axis critically contributes to theinnate-type (22, 28) and acquired-type lung eosinophilia and initi-ation of Th2 differentiation and Ag-specific IgE/IgG1 production(10, 29). In contrast, the roles of IL-33 and IL-33–responsive ILC2sin response to cutaneous administration of protease allergen areunknown, although several lines of evidence suggest the importanceof IL-33 and/or ILC2s in allergic skin diseases, such as atopicdermatitis (AD), and the corresponding animal models (27, 30–33).“Atopic march” is a term defined as the natural progression of
allergic diseases, in which development of atopic dermatitis pre-dates development of food allergy, asthma, or allergic rhinitis,suggesting that skin, especially skin with defective barrier functionor eczema, is the initial site of allergic sensitization in the naturalexposure to environmental allergens (34–36). In the current study,we investigated whether mast cell deficiency and IL-33 deficiencyaffect skin inflammation and serum Ab responses induced by s.c.injection of papain into ear lobes. Furthermore, we examined thecontribution of the s.c. presensitization to papain and IL-33 in an-other model, in which mice s.c. administered papain to induce skininflammation were then exposed to a threshold dose of papain viaairway to induce allergic airway inflammation.
Materials and MethodsMice
Seven- to ten-week-old female C57BL/6N and BALB/c mice pur-chased from Charles River Japan (Yokohama, Japan), WBB6F1-+/+ andWBB6F1-W/Wv mice purchased from Japan SLC (Shizuoka, Japan),and IL-332/2 mice (C57BL/6N background) (22) were maintained in aspecific pathogen-free animal facility at Juntendo University and used inaccordance with the guidelines of the institutional committee on animalexperiments.
Reagents
Papain was purchased from Calbiochem (San Diego, CA). To preparepapain treated with E64 (Peptide Institute, Osaka, Japan), similar to ourprevious studies (10, 16), papain was reacted with an excess of E64after the addition of L-cysteine and, after the reaction, L-cysteine andunbound free E64 were removed by dialysis (E64-papain). Reductionsin the catalytic cysteine residue of papain-like cysteine proteases bythe addition of a reducing reagent such as L-cysteine are necessary forE64 to bind covalently and irreversibly to papain. In some experiments(Figs. 2, 8), papain, which was incubated similarly to E64-treated papain(but without the addition of E64), dialyzed, and reoxidized, was preparedand used for comparisons with E64-papain. The Ags were administeredto mice without being treated with reducing reagents just before theadministration.
Subcutaneous administration of papain to mice
Mice were lightly anesthetized with an i.p. injection of pentobarbital(Somnopentyl; Kyoritsu Pharmaceutical, Tokyo, Japan), and the indicatedamount of papain or E64-papain (10) dissolved in 10 ml PBS was s.c.injected into ear skin. Ear thickness was measured from day 0 until day
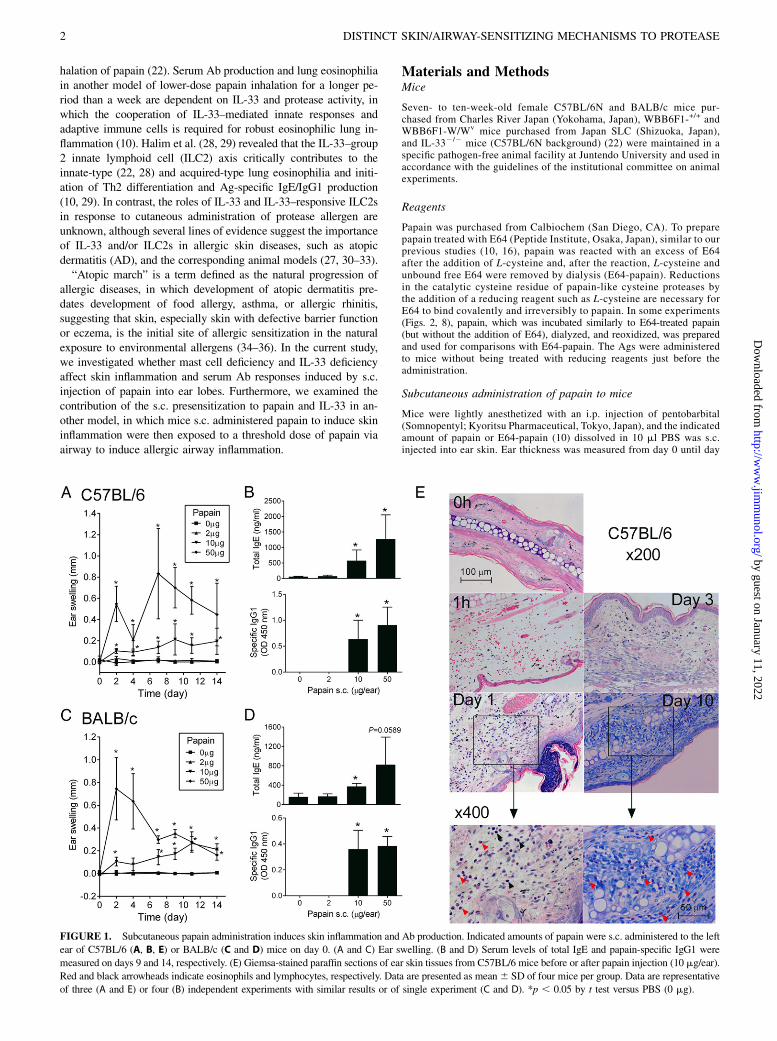
FIGURE 1. Subcutaneous papain administration induces skin inflammation and Ab production. Indicated amounts of papain were s.c. administered to the left
ear of C57BL/6 (A, B, E) or BALB/c (C and D) mice on day 0. (A and C) Ear swelling. (B and D) Serum levels of total IgE and papain-specific IgG1 were
measured on days 9 and 14, respectively. (E) Giemsa-stained paraffin sections of ear skin tissues from C57BL/6 mice before or after papain injection (10 mg/ear).
Red and black arrowheads indicate eosinophils and lymphocytes, respectively. Data are presented as mean6 SD of four mice per group. Data are representative
of three (A and E) or four (B) independent experiments with similar results or of single experiment (C and D). *p , 0.05 by t test versus PBS (0 mg).
2 DISTINCT SKIN/AIRWAY-SENSITIZING MECHANISMS TO PROTEASE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
14 using dial thickness gauge (G-1A; Ozaki, Tokyo, Japan). Serumsamples were collected on days 9–14. Specimens of ear skin tissue, sixear lobes from three mice at each time point, were collected (on day 14,unless otherwise described), fixed with 20% formalin neutral buffered
solution, and embedded in paraffin. Paraffin-embedded sections werestained with H&E or Giemsa staining for histological analyses. Somespecimens of ear skin tissue were used to extract total RNA for geneexpression analyses.
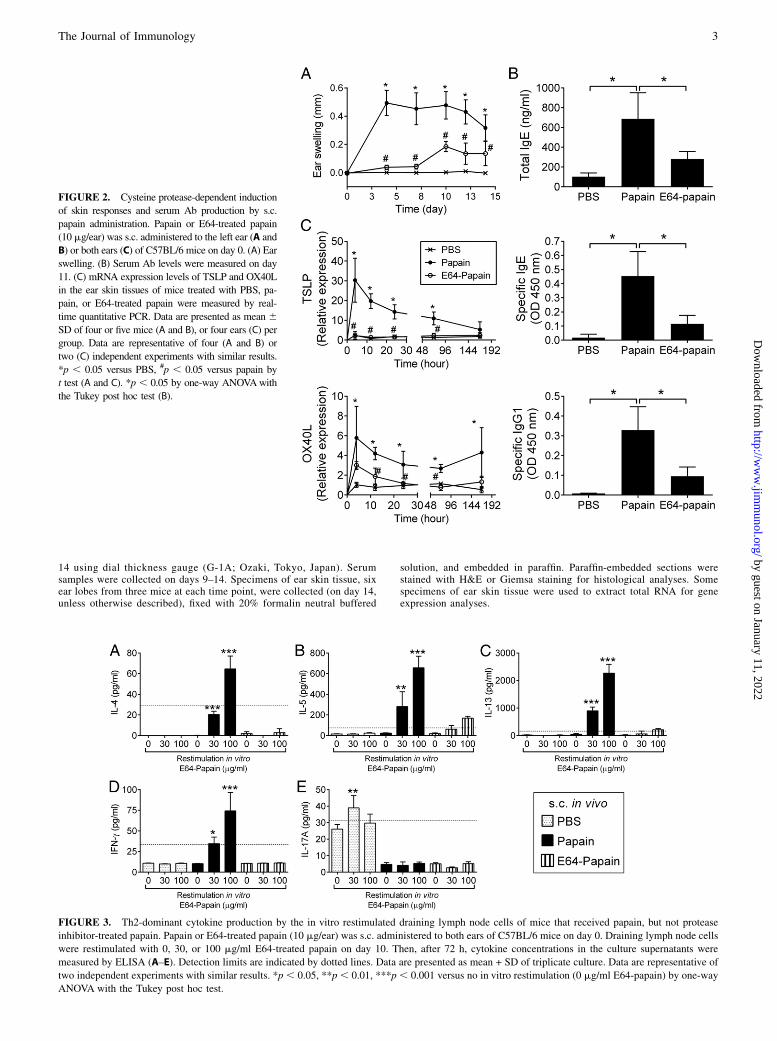
FIGURE 2. Cysteine protease-dependent induction
of skin responses and serum Ab production by s.c.
papain administration. Papain or E64-treated papain
(10 mg/ear) was s.c. administered to the left ear (A and
B) or both ears (C) of C57BL/6 mice on day 0. (A) Ear
swelling. (B) Serum Ab levels were measured on day
11. (C) mRNA expression levels of TSLP and OX40L
in the ear skin tissues of mice treated with PBS, pa-
pain, or E64-treated papain were measured by real-
time quantitative PCR. Data are presented as mean 6SD of four or five mice (A and B), or four ears (C) per
group. Data are representative of four (A and B) or
two (C) independent experiments with similar results.
*p , 0.05 versus PBS, #p , 0.05 versus papain by
t test (A and C). *p , 0.05 by one-way ANOVAwith
the Tukey post hoc test (B).
FIGURE 3. Th2-dominant cytokine production by the in vitro restimulated draining lymph node cells of mice that received papain, but not protease
inhibitor-treated papain. Papain or E64-treated papain (10 mg/ear) was s.c. administered to both ears of C57BL/6 mice on day 0. Draining lymph node cells
were restimulated with 0, 30, or 100 mg/ml E64-treated papain on day 10. Then, after 72 h, cytokine concentrations in the culture supernatants were
measured by ELISA (A–E). Detection limits are indicated by dotted lines. Data are presented as mean + SD of triplicate culture. Data are representative of
two independent experiments with similar results. *p , 0.05, **p , 0.01, ***p , 0.001 versus no in vitro restimulation (0 mg/ml E64-papain) by one-way
ANOVA with the Tukey post hoc test.
The Journal of Immunology 3
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
Histology
Photographs of the papain-injected sites in the Giemsa-stained paraffin sectionswere taken under microscopy. Each photograph was separated into several0.01-mm2 (100 mm 3 100 mm) areas, and eosinophils within each 0.01-mm2
area were counted. The total eosinophil number in one or more photographswas divided by the total counted area (unit: mm2) in the same photograph(s)and indicated as eosinophils/mm2. These histological analyses were per-formed by observers, including one who knows the experimental treatmentand additional one or two who were blind to the experimental treatment.
Generation of bone marrow–derived mast cell andreconstitution of dermal mast cells
Bone marrow–derived mast cells (BMMCs) were generated by culturing bonemarrow cells from WBB6F1-+/+ mice in RPMI 1640 media supplementedwith 10% heat-inactivated FCS, 0.1 mM 2-ME, 0.01 mM MEM–nonessentialamino acids, 100 U/ml penicillin, 0.1 mg/ml streptomycin, 10 ng/ml murineIL-3, and 10 ng/ml murine stem cell factor. After 5–8 wk of culture, cellssuspended in saline were s.c. injected (1 3 106 cells/20 ml saline/ear) to bothears of W/Wv mice. Then 7.5 wk after the s.c. injection of BMMCs, micewere treated s.c. with papain (10 mg/10 ml PBS/ear, day 0) in both ears.
Restimulation of draining lymph node cells
Cervical lymph nodes were collected 10 d after the s.c. injection of PBS-,papain-, or E64-papain. Single-cell suspensions (2.5 3 106 cells/ml) wereprepared from the draining lymph nodes (DLNs) and restimulated withE64-papain (0, 30, or 100 mg/ml) in 96-well cell culture plates (200 ml/well)for 72 h, and culture supernatants were collected for the measurement ofcytokines.
Cytokine ELISA
After restimulation for 72 h, culture supernatants were recovered bycentrifugation at 310 3 g for 5 min. Cytokine concentrations were mea-sured with ELISA kits (Quantikine or DuoSet; R&D Systems, Minneap-olis, MN). Lower detection limits were 31.25 pg/ml for IL-4 and IL-17A,125 pg/ml for IL-13, and 62.5 pg/ml for IL-5 and IFN-g.
Intranasal administration of papain to mice
Mice were lightly anesthetized with an i.p. injection of pentobarbital andallowed to inhale 40 ml papain following alternate application to both nareswith a pipette. The intranasal (i.n.) administration of papain was performed
twice (days 14 and 21). Four days after the last i.n. administration, sera andbronchial alveolar lavage (BAL) fluid cells were collected.
Bronchial alveolar lavage
Four days after the last i.n. administration, mice were terminally anes-thetized, the tracheas were cannulated, and internal airspaces were lavagedwith 1000 ml PBS with 5% FCS. Fluids were centrifuged at 1200 3 g, andthe pellets were recovered for cellular analysis. Specimens were preparedon glass slides by Cytospin 4 (Thermo Shandon, Cheshire, U.K.), followedby Diff-Quick (Sysmex, Kobe, Japan) staining. Differential cell countswere performed with a minimum of 200 cells.
ELISA for serum total IgE and papain-specific Abs
Serum total IgE was measured by a sandwich ELISA, as describedpreviously (10). Papain-specific Abs were detected on plates coatedwith papain (30 mg/ml for Figs. 1, 2 and 5 mg/ml for Figs. 4–8) andblocked with BlockAce (Snow Bland, Sapporo, Japan), and the plateswere developed with HRP-conjugated Abs specific to the murine IgE(biotin-conjugated anti-murine IgE plus HRP-conjugated streptavidinfor Figs. 4–8) and IgG subclasses, as described previously (10, 37).Sera and detection Abs were diluted with Solution 1 and 2 ofCanGetSignal (TOYOBO, Osaka, Japan), respectively. Serum dilutionswere 1/200 (1/40 for Figs. 6, 7) and 1/5,000 (1/10,000 for Fig. 1) fordetecting allergen-specific IgE and IgG1, respectively. Serum sampleswere serially diluted from 1/10 until 1/107 for titration. For detectingpapain-specific IgE, incubation with serum or a detection Ab was for15 h at 4˚C or for 2 h at room temperature, respectively. For detectingpapain-specific IgGs, incubation with serum or a detection Ab was for1.5 h at 37˚C.
Quantitative PCR
Ear lobes were excised and homogenized in QIAzol Lysis Reagent (Qiagen,Clifton Hill, Australia) by using TissueLyser II (Qiagen). Total RNAwas extracted from the homogenized skin tissue specimens by using aRNeasy Plus Micro Kit (Qiagen). First-strand cDNAwas synthesized fromtotal RNA by using a High Capacity cDNA Reverse Transcription Kit(Applied Biosystems, Foster City, CA). Real-time quantitative PCR wasperformed with the TaqMan method using an ABI 7500 system (AppliedBiosystems). The mRNA levels of the target gene were normalized to thegene expression levels of GAPDH and are shown as relative expressionlevels to the control group.
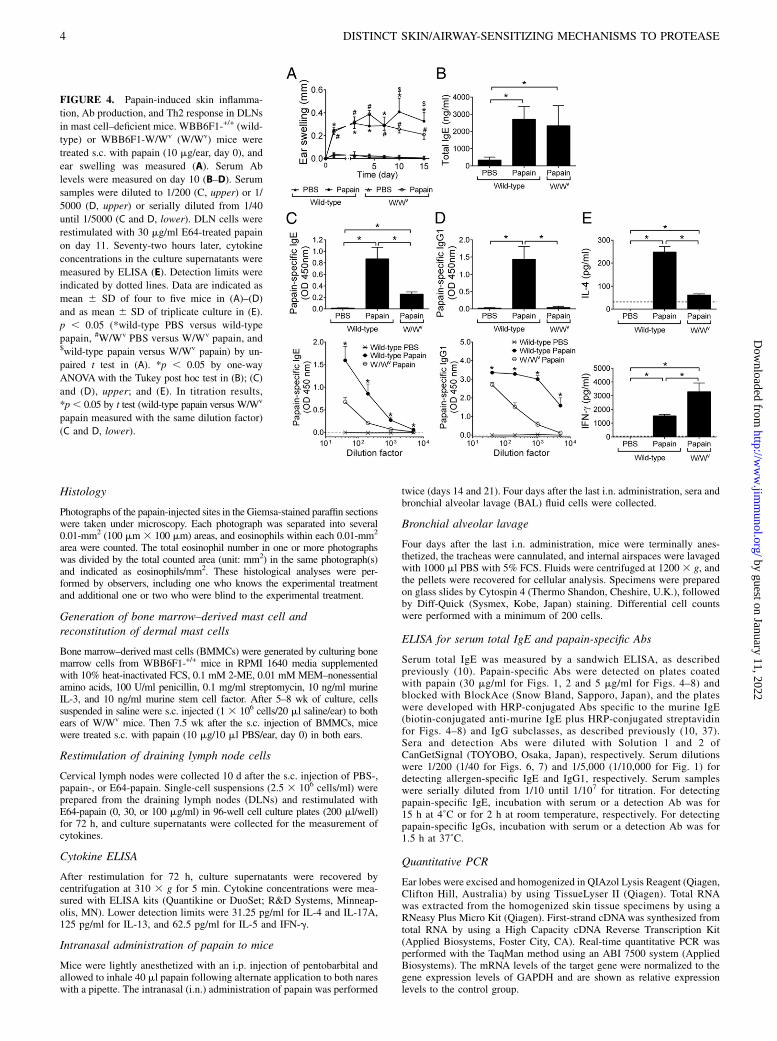
FIGURE 4. Papain-induced skin inflamma-
tion, Ab production, and Th2 response in DLNs
in mast cell–deficient mice. WBB6F1-+/+ (wild-
type) or WBB6F1-W/Wv (W/Wv) mice were
treated s.c. with papain (10 mg/ear, day 0), and
ear swelling was measured (A). Serum Ab
levels were measured on day 10 (B–D). Serum
samples were diluted to 1/200 (C, upper) or 1/
5000 (D, upper) or serially diluted from 1/40
until 1/5000 (C and D, lower). DLN cells were
restimulated with 30 mg/ml E64-treated papain
on day 11. Seventy-two hours later, cytokine
concentrations in the culture supernatants were
measured by ELISA (E). Detection limits were
indicated by dotted lines. Data are indicated as
mean 6 SD of four to five mice in (A)–(D)
and as mean 6 SD of triplicate culture in (E).
p , 0.05 (*wild-type PBS versus wild-type
papain, #W/Wv PBS versus W/Wv papain, and$wild-type papain versus W/Wv papain) by un-
paired t test in (A). *p , 0.05 by one-way
ANOVAwith the Tukey post hoc test in (B); (C)
and (D), upper; and (E). In titration results,
*p, 0.05 by t test (wild-type papain versus W/Wv
papain measured with the same dilution factor)
(C and D, lower).
4 DISTINCT SKIN/AIRWAY-SENSITIZING MECHANISMS TO PROTEASE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
Statistical analysis
One-way ANOVA with the Tukey post hoc test, unpaired t test, or Mann–Whitney U test was used as indicated in the figure legends. A p value,0.05 was regarded as statistically significant.
ResultsSubcutaneous administration of papain-induced skininflammation and serum IgE and IgG1 production
We previously reported the cysteine protease-dependent inductionof serum IgE/IgG1 production and allergic airway inflammation byrepeated inhalation of papain (10). In the current study, we testedthe induction of skin inflammation by s.c. administered papain(Fig. 1). The s.c. injection of 10–50 mg/10 ml/ear papain resultedin ear swelling that started 1 h after the injection and lasted at least14 d (Fig. 1A, 1C) and induced production of total IgE andpapain-specific IgG1 in the serum (Fig. 1B, 1D). Injection of 50mg papain resulted in the formation of scabs after the initial er-ythematous swelling subsided, whereas injection of 10 mg papaincaused less severe or no scab formation (data not shown). The first
histological manifestation was hemorrhage in the dermis, whichwas observed 1 h after 10 mg papain injection (Fig. 1E, 1 h).Moderate cellular infiltration including lymphocytes and eosino-phils was observed on day 1 (Fig. 1E, day 1). Massive eosinophilicinfiltration was found on day 10, after the initial moderate cellularinfiltration subsided on day 3 (Fig. 1E, days 10 and 3).
Subcutaneous administration of papain induced the skinresponses and Ab production in a protease-dependent manner
To test the contribution of the cysteine protease activity of s.c.administered papain to the skin inflammation and Ab responses, weinjected papain that had been treated with a cysteine protease-specific inhibitor, E64. The treatment of papain with E64 signifi-cantly reduced the papain-induced ear swelling (Fig. 2A) andproduction of total IgE and papain-specific IgE and IgG1 (Fig. 2B).Tang et al. (18) reported that s.c. injection of papain (50 mg)
leads to the expression of thymic stromal lymphopoietin (TSLP), aTh2-inducing cytokine (38, 39), in the ear skin. We confirmed theTSLP mRNA expression in the s.c. papain (10 mg/ear)-injected ear
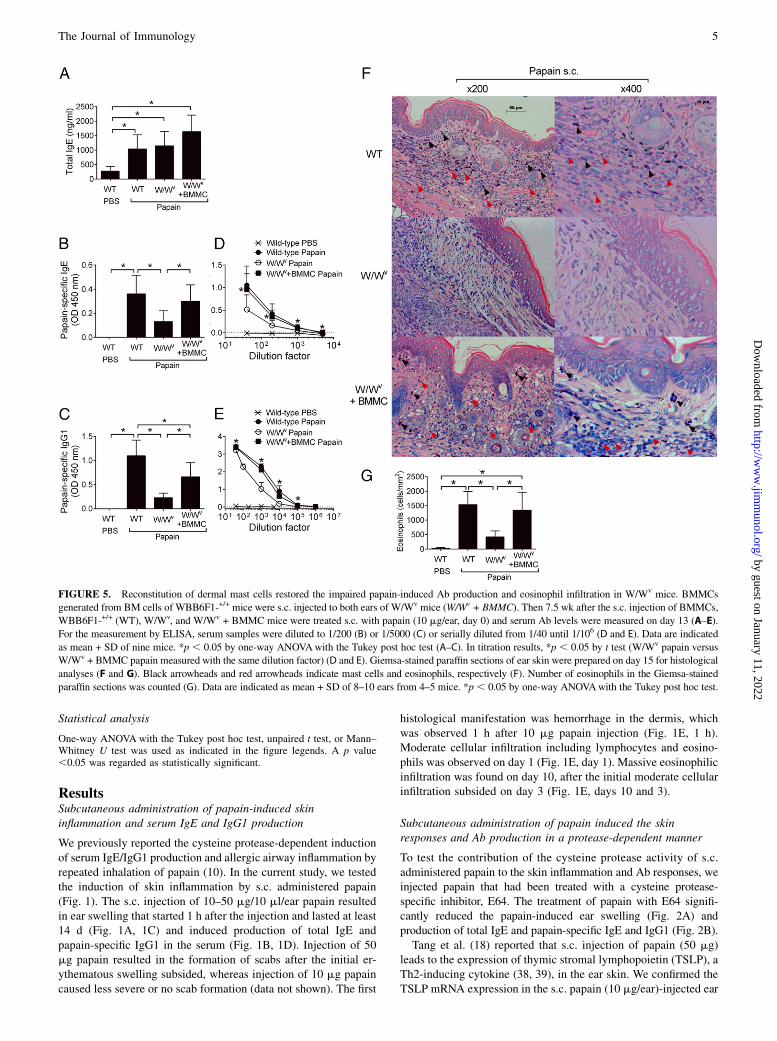
FIGURE 5. Reconstitution of dermal mast cells restored the impaired papain-induced Ab production and eosinophil infiltration in W/Wv mice. BMMCs
generated from BM cells of WBB6F1-+/+ mice were s.c. injected to both ears of W/Wv mice (W/Wv + BMMC). Then 7.5 wk after the s.c. injection of BMMCs,
WBB6F1-+/+ (WT), W/Wv, and W/Wv + BMMC mice were treated s.c. with papain (10 mg/ear, day 0) and serum Ab levels were measured on day 13 (A–E).
For the measurement by ELISA, serum samples were diluted to 1/200 (B) or 1/5000 (C) or serially diluted from 1/40 until 1/106 (D and E). Data are indicated
as mean + SD of nine mice. *p , 0.05 by one-way ANOVAwith the Tukey post hoc test (A–C). In titration results, *p , 0.05 by t test (W/Wv papain versus
W/Wv + BMMC papain measured with the same dilution factor) (D and E). Giemsa-stained paraffin sections of ear skin were prepared on day 15 for histological
analyses (F and G). Black arrowheads and red arrowheads indicate mast cells and eosinophils, respectively (F). Number of eosinophils in the Giemsa-stained
paraffin sections was counted (G). Data are indicated as mean + SD of 8–10 ears from 4–5 mice. *p , 0.05 by one-way ANOVAwith the Tukey post hoc test.
The Journal of Immunology 5
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
tissues, which was induced 4 h after the injection and lasted for atleast 72 h (Fig. 2C). We analyzed gene expression of OX40L, whichis induced in TSLP-stimulated dendritic cells to differentiate naiveT cells to inflammatory Th2 cells (38), in the ear skin tissues. Theelevated expression of OX40L mRNA was observed from 4 h andlasted for at least 7 d after the s.c. papain injection (Fig. 2C). Theelevations of TSLP and OX40L mRNA expression were not ob-served in E64-papain–injected mice (Fig. 2C).The results showed that the cysteine protease activity of
papain induces the skin responses associated with upregulationof TSLP and OX40L gene expression in the skin and Ab re-sponses.
Subcutaneous administration of papain induced Th2-dominantdifferentiation in DLN cells in a protease-dependent manner
We examined the cytokine production by DLN cells from papain-injected mice (Fig. 3). We restimulated DLN cells with E64-papain, but not intact papain to exclude the potential activation ofT cells via the protease activity in vitro. When restimulated, DLNcells from papain-injected mice produced IL-4, IL-5, and IL-13(Fig. 3A–C). A low concentration of IFN-g was produced byDLN cells from papain-injected mice (Fig. 3D). The Th2/Th1 cy-tokine production in response to in vitro restimulation was absent ormarkedly reduced in the DLN cells from E64-papain–injected mice(Fig. 3). The results demonstrated that the protease activity of s.c.administered papain was necessary for the Th2-dominant cytokineproduction profile in the DLNs.
Mast cell–deficient mice showed reduced serumpapain-specific IgE and IgG1 production and eosinophilinfiltration in the papain-induced skin inflammation model
Next, we examined the contribution of mast cells to papain-induced skin inflammation and serum Ab responses (Fig. 4). Earswelling was equally induced by s.c. injected papain both in mastcell–sufficient WBB6F1-+/+ (wild-type) and mast cell–deficientWBB6F1-W/Wv (W/Wv) mice during the first week after injection(Fig. 4A). The swelling of papain-injected ears was less in W/Wv
mice than wild-type mice from days 10 to 15 (Fig. 4A). Althoughserum levels of total IgE were not affected by the mast celldeficiency (Fig. 4B), reduced production of papain-specific IgE(Fig. 4C) and IgG1 (Fig. 4D) was observed in W/Wv mice. Uponrestimulation with E64-papain in vitro, DLN cells from papain-injected W/Wv mice showed significantly lower IL-4 productionand higher IFN-g production compared with DLN cells frompapain-injected wild-type mice (Fig. 4E). Reduced infiltrationof eosinophils into dermis was also observed in W/Wv mice(Fig. 5F, 5G).Then, we reconstituted mast cells in the dermal tissue of W/Wv
mice by s.c. injection of BMMCs to the ear lobes and testedthe effect of papain injection on the serum Ab responses andeosinophil infiltration (Fig. 5). Whereas serum levels of totalIgE were similar among the papain-injected wild-type, W/Wv,and BMMC-reconstituted W/Wv mice (Fig. 5A), the reductionof papain-specific IgE (Fig. 5B, 5D), IgG1 (Fig. 5C, 5E), andeosinophil infiltration (Fig. 5F, 5G) in W/Wv mice was restored by
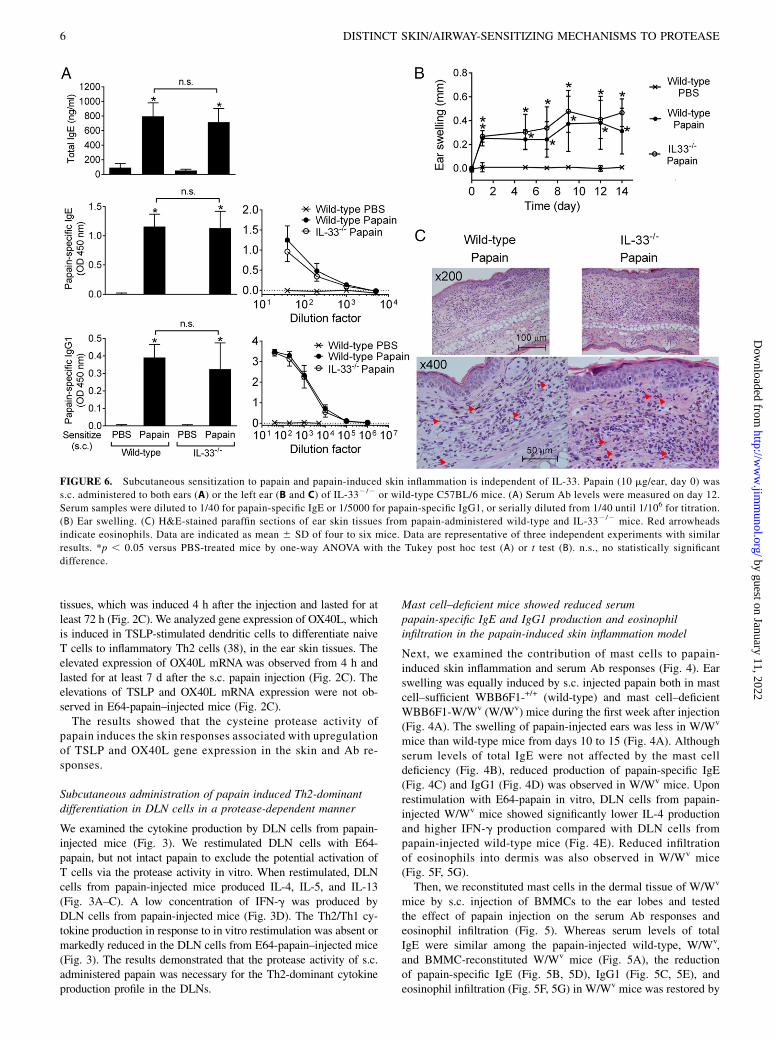
FIGURE 6. Subcutaneous sensitization to papain and papain-induced skin inflammation is independent of IL-33. Papain (10 mg/ear, day 0) was
s.c. administered to both ears (A) or the left ear (B and C) of IL-332/2 or wild-type C57BL/6 mice. (A) Serum Ab levels were measured on day 12.
Serum samples were diluted to 1/40 for papain-specific IgE or 1/5000 for papain-specific IgG1, or serially diluted from 1/40 until 1/106 for titration.
(B) Ear swelling. (C) H&E-stained paraffin sections of ear skin tissues from papain-administered wild-type and IL-332/2 mice. Red arrowheads
indicate eosinophils. Data are indicated as mean 6 SD of four to six mice. Data are representative of three independent experiments with similar
results. *p , 0.05 versus PBS-treated mice by one-way ANOVA with the Tukey post hoc test (A) or t test (B). n.s., no statistically significant
difference.
6 DISTINCT SKIN/AIRWAY-SENSITIZING MECHANISMS TO PROTEASE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
the reconstitution of mast cells in the dermal tissue. These resultsshowed that mast cells contribute profoundly to the induction ofpapain-specific Abs and eosinophil infiltration in the papain-induced skin inflammation model.
Subcutaneous administration of papain induced skininflammation and serum IgE/IgG1 responses in an IL-33–independent manner
As precedent studies demonstrated the contribution of IL-33 toallergic airway inflammation (10, 22) and serum IgE/IgG1 re-sponses induced by papain inhalation (10), we next examined theinvolvement of IL-33 in the skin inflammation and Ab responsesinduced by s.c. papain administration (Fig. 6). Total IgE andpapain-specific IgE and IgG1 were induced equivalently betweenwild-type and IL-332/2 mice on day 12 after the s.c. papain ad-ministration (Fig. 6A). There were no differences between wild-type and IL-332/2 mice in the swelling of papain-injected ears,cellular infiltration to the dermis, or epidermal hyperplasia(Fig. 6B, 6C). Thus, IL-33 is not required for the skin inflam-mation or allergic sensitization to papain that penetrated the epi-dermal barrier.
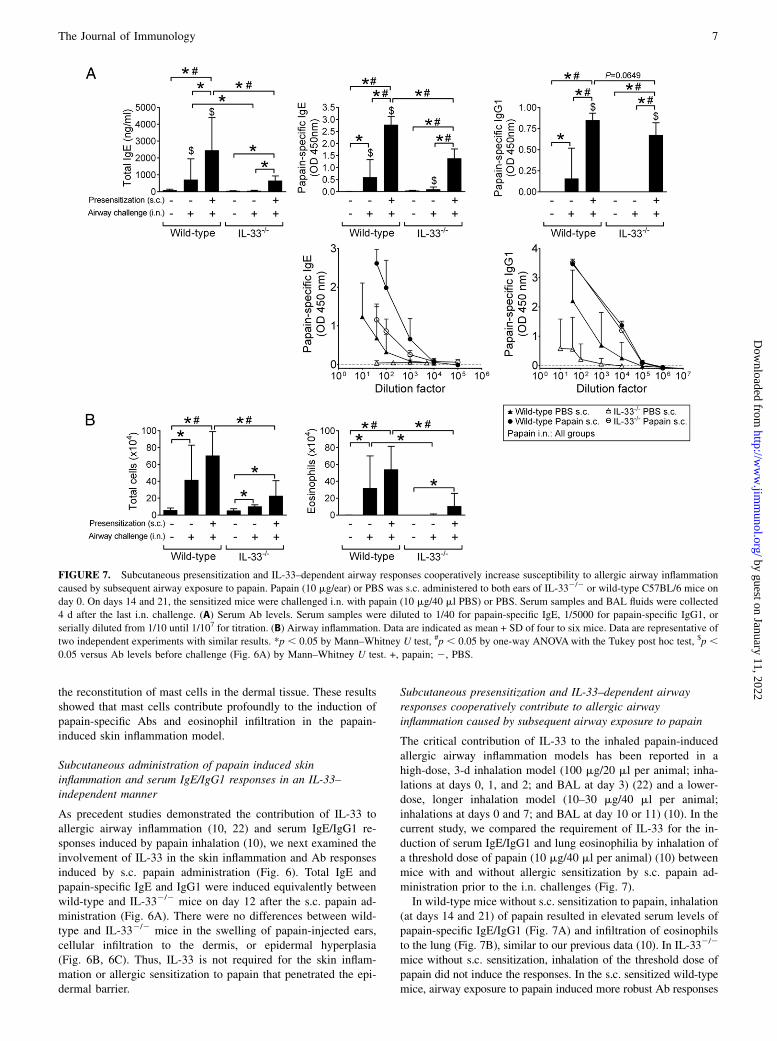
Subcutaneous presensitization and IL-33–dependent airwayresponses cooperatively contribute to allergic airwayinflammation caused by subsequent airway exposure to papain
The critical contribution of IL-33 to the inhaled papain-inducedallergic airway inflammation models has been reported in ahigh-dose, 3-d inhalation model (100 mg/20 ml per animal; inha-lations at days 0, 1, and 2; and BAL at day 3) (22) and a lower-dose, longer inhalation model (10–30 mg/40 ml per animal;inhalations at days 0 and 7; and BAL at day 10 or 11) (10). In thecurrent study, we compared the requirement of IL-33 for the in-duction of serum IgE/IgG1 and lung eosinophilia by inhalation ofa threshold dose of papain (10 mg/40 ml per animal) (10) betweenmice with and without allergic sensitization by s.c. papain ad-ministration prior to the i.n. challenges (Fig. 7).In wild-type mice without s.c. sensitization to papain, inhalation
(at days 14 and 21) of papain resulted in elevated serum levels ofpapain-specific IgE/IgG1 (Fig. 7A) and infiltration of eosinophilsto the lung (Fig. 7B), similar to our previous data (10). In IL-332/2
mice without s.c. sensitization, inhalation of the threshold dose ofpapain did not induce the responses. In the s.c. sensitized wild-typemice, airway exposure to papain induced more robust Ab responses
FIGURE 7. Subcutaneous presensitization and IL-33–dependent airway responses cooperatively increase susceptibility to allergic airway inflammation
caused by subsequent airway exposure to papain. Papain (10 mg/ear) or PBS was s.c. administered to both ears of IL-332/2 or wild-type C57BL/6 mice on
day 0. On days 14 and 21, the sensitized mice were challenged i.n. with papain (10 mg/40 ml PBS) or PBS. Serum samples and BAL fluids were collected
4 d after the last i.n. challenge. (A) Serum Ab levels. Serum samples were diluted to 1/40 for papain-specific IgE, 1/5000 for papain-specific IgG1, or
serially diluted from 1/10 until 1/107 for titration. (B) Airway inflammation. Data are indicated as mean + SD of four to six mice. Data are representative of
two independent experiments with similar results. *p , 0.05 by Mann–Whitney U test, #p , 0.05 by one-way ANOVAwith the Tukey post hoc test, $p ,0.05 versus Ab levels before challenge (Fig. 6A) by Mann–Whitney U test. +, papain; 2, PBS.
The Journal of Immunology 7
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
and lung eosinophilia than in mice without s.c. papain injection.The s.c. sensitized IL-332/2 mice showed lower, but still sig-nificant lung eosinophilia upon airway exposure to papain(Fig. 7B). The airway exposure to papain boosted total IgE andpapain-specific IgE in the s.c. sensitized wild-type, but not IL-332/2 mice (Fig. 7A compared with Fig. 6A). Papain-specificIgG1 was boosted in both of the s.c. sensitized wild-type and IL-332/2 mice. Upon the airway exposure to PBS, no lung eosin-ophilia was observed in the s.c. sensitized mice (data notshown).We examined dependency of the responses upon the i.n.
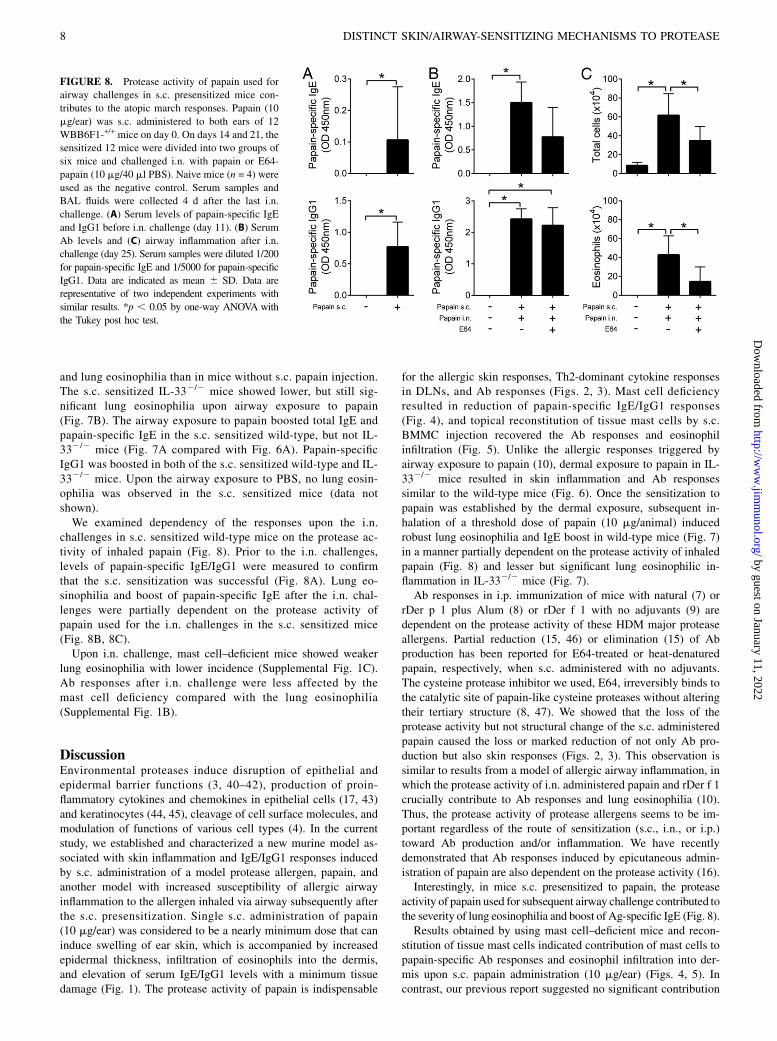
challenges in s.c. sensitized wild-type mice on the protease ac-tivity of inhaled papain (Fig. 8). Prior to the i.n. challenges,levels of papain-specific IgE/IgG1 were measured to confirmthat the s.c. sensitization was successful (Fig. 8A). Lung eo-sinophilia and boost of papain-specific IgE after the i.n. chal-lenges were partially dependent on the protease activity ofpapain used for the i.n. challenges in the s.c. sensitized mice(Fig. 8B, 8C).Upon i.n. challenge, mast cell–deficient mice showed weaker
lung eosinophilia with lower incidence (Supplemental Fig. 1C).Ab responses after i.n. challenge were less affected by themast cell deficiency compared with the lung eosinophilia(Supplemental Fig. 1B).
DiscussionEnvironmental proteases induce disruption of epithelial andepidermal barrier functions (3, 40–42), production of proin-flammatory cytokines and chemokines in epithelial cells (17, 43)and keratinocytes (44, 45), cleavage of cell surface molecules, andmodulation of functions of various cell types (4). In the currentstudy, we established and characterized a new murine model as-sociated with skin inflammation and IgE/IgG1 responses inducedby s.c. administration of a model protease allergen, papain, andanother model with increased susceptibility of allergic airwayinflammation to the allergen inhaled via airway subsequently afterthe s.c. presensitization. Single s.c. administration of papain(10 mg/ear) was considered to be a nearly minimum dose that caninduce swelling of ear skin, which is accompanied by increasedepidermal thickness, infiltration of eosinophils into the dermis,and elevation of serum IgE/IgG1 levels with a minimum tissuedamage (Fig. 1). The protease activity of papain is indispensable
for the allergic skin responses, Th2-dominant cytokine responsesin DLNs, and Ab responses (Figs. 2, 3). Mast cell deficiencyresulted in reduction of papain-specific IgE/IgG1 responses(Fig. 4), and topical reconstitution of tissue mast cells by s.c.BMMC injection recovered the Ab responses and eosinophilinfiltration (Fig. 5). Unlike the allergic responses triggered byairway exposure to papain (10), dermal exposure to papain in IL-332/2 mice resulted in skin inflammation and Ab responsessimilar to the wild-type mice (Fig. 6). Once the sensitization topapain was established by the dermal exposure, subsequent in-halation of a threshold dose of papain (10 mg/animal) inducedrobust lung eosinophilia and IgE boost in wild-type mice (Fig. 7)in a manner partially dependent on the protease activity of inhaledpapain (Fig. 8) and lesser but significant lung eosinophilic in-flammation in IL-332/2 mice (Fig. 7).Ab responses in i.p. immunization of mice with natural (7) or
rDer p 1 plus Alum (8) or rDer f 1 with no adjuvants (9) aredependent on the protease activity of these HDM major proteaseallergens. Partial reduction (15, 46) or elimination (15) of Abproduction has been reported for E64-treated or heat-denaturedpapain, respectively, when s.c. administered with no adjuvants.The cysteine protease inhibitor we used, E64, irreversibly binds tothe catalytic site of papain-like cysteine proteases without alteringtheir tertiary structure (8, 47). We showed that the loss of theprotease activity but not structural change of the s.c. administeredpapain caused the loss or marked reduction of not only Ab pro-duction but also skin responses (Figs. 2, 3). This observation issimilar to results from a model of allergic airway inflammation, inwhich the protease activity of i.n. administered papain and rDer f 1crucially contribute to Ab responses and lung eosinophilia (10).Thus, the protease activity of protease allergens seems to be im-portant regardless of the route of sensitization (s.c., i.n., or i.p.)toward Ab production and/or inflammation. We have recentlydemonstrated that Ab responses induced by epicutaneous admin-istration of papain are also dependent on the protease activity (16).Interestingly, in mice s.c. presensitized to papain, the protease
activity of papain used for subsequent airway challenge contributed tothe severity of lung eosinophilia and boost of Ag-specific IgE (Fig. 8).Results obtained by using mast cell–deficient mice and recon-
stitution of tissue mast cells indicated contribution of mast cells topapain-specific Ab responses and eosinophil infiltration into der-mis upon s.c. papain administration (10 mg/ear) (Figs. 4, 5). Incontrast, our previous report suggested no significant contribution
FIGURE 8. Protease activity of papain used for
airway challenges in s.c. presensitized mice con-
tributes to the atopic march responses. Papain (10
mg/ear) was s.c. administered to both ears of 12
WBB6F1-+/+ mice on day 0. On days 14 and 21, the
sensitized 12 mice were divided into two groups of
six mice and challenged i.n. with papain or E64-
papain (10 mg/40 ml PBS). Naive mice (n = 4) were
used as the negative control. Serum samples and
BAL fluids were collected 4 d after the last i.n.
challenge. (A) Serum levels of papain-specific IgE
and IgG1 before i.n. challenge (day 11). (B) Serum
Ab levels and (C) airway inflammation after i.n.
challenge (day 25). Serum samples were diluted 1/200
for papain-specific IgE and 1/5000 for papain-specific
IgG1. Data are indicated as mean 6 SD. Data are
representative of two independent experiments with
similar results. *p , 0.05 by one-way ANOVA with
the Tukey post hoc test.
8 DISTINCT SKIN/AIRWAY-SENSITIZING MECHANISMS TO PROTEASE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

of mast cells to IgE/IgG1 responses and lung eosinophilia uponi.n. papain administration (10). These results suggest the contri-bution of mast cells to allergic sensitization to protease allergensadministered via s.c. but not i.n. route. No decrease of serum totalIgE by the mast cell deficiency (Figs. 4B, 5A) could be attribut-able to the fact that papain-specific IgE was induced in mast cell–deficient mice, although at lower levels than wild-type mice, andIgE harbored on the high-affinity IgE receptor expressed on thesurface of mast cells in wild-type mice should exist as free IgE inserum in the mast cell–deficient mice. Upon subsequent i.n.challenge, mast cell–deficient mice showed weaker lung eosino-philia (Supplemental Fig. 1C). To our knowledge, this is the firstdemonstration that Ab responses against protein Ags were reducedin mast cell–deficient mice, although the mechanism is yet to beinvestigated. Recent reports showed contribution of mast cells toskin inflammation in mice (48, 49), but did not describe contri-bution of mast cells to Ag-specific Ab responses.IL-33 was not necessary for the skin responses and Ab responses
upon s.c. administration to ear skin (Fig. 6), in contrast to airwayinflammation induced by exposure of airway mucosa to papain,in which the indispensable contribution of IL-33 (10, 22) and IL-33–activated ILC2s (28, 29) has been reported; these results alsosuggest the differential contribution of IL-33 to allergic reactionsin the skin and airway. Similar to our results using papain withoutany adjuvant, Savinko et al. (50) reported that mice lacking asubunit of the receptor for IL-33, ST2, that were epicutaneouslysensitized with OVA in the presence of a superantigen, staphylo-coccal enterotoxin B, as an adjuvant and subsequently i.n. chal-lenged with OVA alone, showed decreased numbers of eosinophilsin the airway, whereas dermal eosinophil numbers were equal tothose in the epicutaneously sensitized wild-type mice. However,expression of IL-33 and ST2 (30, 31) and presence of ILC2s (32,33) in the skin tissue of AD patients and association of single-nucleotide polymorphisms in the ST2 gene with AD (30) havebeen reported, and skin-specific overexpression of IL-33 in miceresulted in activation of ILC2s and spontaneous onset of AD-likeinflammation (27), suggesting possible involvement of the IL-33–ST2 interaction and ILC2s in the pathogenesis of AD. Although acontribution of IL-33 to skin inflammation and Ab production wasnot observed in our model, suggesting that IL-33 does not con-tribute to the initial dermal sensitization phase, we cannot excludethe possibility of a contribution of IL-33 to the pathogenesis indifferent experimental settings such as those associated with moreexacerbated or chronic symptoms.The term “atopic march” refers to the natural history of allergic
diseases, which is characterized by a typical sequence of IgEresponses and clinical symptoms that appear early in life, persistover years, and often remit spontaneously with age. Generally, theearliest clinical sign in the atopic march is AD, and the onset offood allergy, atopic rhinitis, or asthma becomes prominent later asthe initial signs of AD subside. Based on the sequence of theaffected tissues, it is proposed that skin, especially skin withdefective barrier function (51, 52) or eczema (36), is the initial siteof allergic sensitization to allergens in the natural history ofallergic diseases (34, 35, 53). In our murine model, in which mices.c. administered papain were then exposed to a threshold dose ofpapain via airway (10 mg), mice with a history of allergic dermalsensitization have an increased susceptibility to allergic airwayinflammation (Fig. 7). The protease activity of papain used forthe i.n. challenges (Fig. 8) and IL-33 (Fig. 7) contributes to theatopic march responses. Two distinct pathways cooperativelycontributed to the induction of maximum IgE response and lungeosinophilia in this s.c. sensitization–i.n. challenge model, that is,s.c. presensitization-dependent, IL-33–independent pathway and
airway protease–induced, IL-33–mediated pathway. It is likelythat the first pathway involves memory Th2 responses and Ab-mediated responses and that the second pathway is mediated byIL-33–responsive cells in the airway, which include ILC2s and/orother cell types (21, 54).Ab production and eosinophil infiltration into dermis were de-
pendent on mast cells, but not on IL-33 in the s.c. model (Figs. 4–6), whereas IL-33 contributed to Ab responses and lung eosino-philia (10) (Fig. 7) and mast cell did not significantly contribute tothe responses (10) in i.n. models. The main role of IL-33 in the i.n.models is considered to stimulate ST2-expressing ILC2s to releaseabundant IL-5 and IL-13, both of which cooperating with Th2-derived ones cause lung eosinophilia, and the latter of which, IL-13, stimulates lung dendritic cells to initiate Th2 differentiation,leading to the IgE/IgG1 responses (10, 29). Hara et al. (55) sug-gested that uric acid is a key sensor molecule to alarm the proteaseinhalation or tissue damage by induction of IL-33 release. In thes.c. model, how mast cells contribute to the Ab and eosinophilresponses, possible contribution of ILC2s activated with othercytokines than IL-33 (32), mediators inducing the responses,sensor molecules for the protease penetration, and so on are yetto be investigated in future studies.In summary, we showed that s.c. administration of protease
allergen causes skin inflammation, sensitization with IgE/IgG1production, and Th2 differentiation in a manner dependent onthe protease activity of the allergen (Figs. 1–3) similarly to anallergic airway inflammation model. However, the responsiblemechanisms seem to be different between the s.c. and i.n. sensi-tizations, that is, IL-33 is dispensable and Ab production andeosinophil infiltration are dependent on mast cells in the s.c.model (Figs. 4–6), but not in the i.n. model (10). Furthermore, thedermal sensitization–airway challenge model with protease aller-gen enables the robust lung eosinophilia and IgE boost upon i.n.challenge with a threshold dose of the allergen in wild-type, butnot IL-332/2 mice (Fig. 7), and the protease activity of the inhaledAg contributes to the responses (Fig. 8), suggesting that both thedermal sensitization prior to the airway exposure and protease-induced, IL-33–mediated responses upon the airway exposurecontribute to the transition from skin inflammation to asthma andthat the combination of both maximizes the responses. This studyprovides a concept of allergic sensitization that mammals cansense exogenous proteases as innate danger signals regardless ofthe sensitization route, but via distinct ways according to thesensitization route. Cooperation of initial sensitization via skinand re-exposure via airway to the protease allergen maximizesthe magnitude of the atopic march, natural history of progressionof allergic diseases. Based on clinical studies about the treatmentof human atopic diseases, it is proposed that early treatment ofchildren with AD will prevent the subsequent development of theatopic march (34, 35). Unknown mechanisms behind the mastcell–dependent, IL-33–independent skin inflammation and Abresponses in the s.c. model and investigations in epicutaneoussensitization models (16) should be addressed in future studies.
AcknowledgmentsWe thankMichiyoMatsumoto for secretarial assistance; Tomoko Tokura for
technical assistance; Hideo Iida, Hiroko Ushio, and Nobuhiro Nakano for
comments; Hiroshi Kawai, Toyoko Hidano, and Takatoshi Kuhara for an-
imal care; and Noriyoshi Sueyoshi, Shinji Nakamura, Yuko Kojima, Atsushi
Furuhata, Yasuko Toi, Katsumi Miyahara, and Mio Miyazawa for specimen
preparation for histology.
DisclosuresThe authors have no financial conflicts of interest.
The Journal of Immunology 9
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

References1. Pulendran, B., and D. Artis. 2012. New paradigms in type 2 immunity. Science
337: 431–435.2. Palm, N. W., R. K. Rosenstein, and R. Medzhitov. 2012. Allergic host defences.
Nature 484: 465–472.3. Takai, T., and S. Ikeda. 2011. Barrier dysfunction caused by environmental
proteases in the pathogenesis of allergic diseases. Allergol. Int. 60: 25–35.4. Shakib, F., A. M. Ghaemmaghami, and H. F. Sewell. 2008. The molecular basis
of allergenicity. Trends Immunol. 29: 633–642.5. Takai, T. 2013. Mite endopeptidase 1. In Handbook of Proteolytic Enzymes, 3rd
Ed. N. D. Rawlings, and G. S. Salvesen, eds. Academic Press, Oxford, p. 1957–1963.
6. Takai, T. 2013. Serine endopeptidase allergens from Dermatophagoides species.In Handbook of Proteolytic Enzymes, 3rd Ed. N. D. Rawlings, andG. S. Salvesen, eds. Academic Press, Oxford, p. 3055–3060.
7. Gough, L., O. Schulz, H. F. Sewell, and F. Shakib. 1999. The cysteine proteaseactivity of the major dust mite allergen Der p 1 selectively enhances the im-munoglobulin E antibody response. J. Exp. Med. 190: 1897–1902.
8. Kikuchi, Y., T. Takai, T. Kuhara, M. Ota, T. Kato, H. Hatanaka, S. Ichikawa,T. Tokura, H. Akiba, K. Mitsuishi, et al. 2006. Crucial commitment of proteo-lytic activity of a purified recombinant major house dust mite allergen Der p1 tosensitization toward IgE and IgG responses. J. Immunol. 177: 1609–1617.
9. Takai, T., T. Kato, H. Hatanaka, K. Inui, T. Nakazawa, S. Ichikawa, K. Mitsuishi,H. Ogawa, and K. Okumura. 2009. Modulation of allergenicity of major housedust mite allergens Der f 1 and Der p 1 by interaction with an endogenous ligand.J. Immunol. 183: 7958–7965.
10. Kamijo, S., H. Takeda, T. Tokura, M. Suzuki, K. Inui, M. Hara, H. Matsuda,A. Matsuda, K. Oboki, T. Ohno, et al. 2013. IL-33-mediated innate response andadaptive immune cells contribute to maximum responses of protease allergen-induced allergic airway inflammation. J. Immunol. 190: 4489–4499.
11. Thomas, W. R. 2015. Hierarchy and molecular properties of house dust miteallergens. Allergol. Int. 64: 304–311.
12. Chua, K. Y., G. A. Stewart, W. R. Thomas, R. J. Simpson, R. J. Dilworth,T. M. Plozza, and K. J. Turner. 1988. Sequence analysis of cDNA coding for amajor house dust mite allergen, Der p 1: homology with cysteine proteases. J.Exp. Med. 167: 175–182.
13. Cunningham, P. T., C. E. Elliot, J. C. Lenzo, A. G. Jarnicki, A. N. Larcombe,G. R. Zosky, P. G. Holt, and W. R. Thomas. 2012. Sensitizing and Th2 adjuvantactivity of cysteine protease allergens. Int. Arch. Allergy Immunol. 158: 347–358.
14. McGlade, J. P., S. Gorman, J. C. Lenzo, J. W. Tan, T. Watanabe, J. J. Finlay-Jones, W. R. Thomas, and P. H. Hart. 2007. Effect of both ultraviolet B irradi-ation and histamine receptor function on allergic responses to an inhaled antigen.J. Immunol. 178: 2794–2802.
15. Sokol, C. L., G. M. Barton, A. G. Farr, and R. Medzhitov. 2008. A mechanismfor the initiation of allergen-induced T helper type 2 responses. Nat. Immunol. 9:310–318.
16. Iida, H., T. Takai, Y. Hirasawa, S. Kamijo, S. Shimura, H. Ochi, I. Nishioka,N. Maruyama, H. Ogawa, K. Okumura, and S. Ikeda. 2014. Epicutaneous ad-ministration of papain induces IgE and IgG responses in a cysteine proteaseactivity-dependent manner. Allergol. Int. 63: 219–226.
17. Kouzaki, H., S. M. O’Grady, C. B. Lawrence, and H. Kita. 2009. Proteases in-duce production of thymic stromal lymphopoietin by airway epithelial cellsthrough protease-activated receptor-2. J. Immunol. 183: 1427–1434.
18. Tang, H., W. Cao, S. P. Kasturi, R. Ravindran, H. I. Nakaya, K. Kundu,N. Murthy, T. B. Kepler, B. Malissen, and B. Pulendran. 2010. The T helper type2 response to cysteine proteases requires dendritic cell-basophil cooperation viaROS-mediated signaling. Nat. Immunol. 11: 608–617.
19. Motomura, Y., H. Morita, K. Moro, S. Nakae, D. Artis, T. A. Endo, Y. Kuroki,O. Ohara, S. Koyasu, and M. Kubo. 2014. Basophil-derived interleukin-4 con-trols the function of natural helper cells, a member of ILC2s, in lung inflam-mation. Immunity 40: 758–771.
20. Hammad, H., M. Plantinga, K. Deswarte, P. Pouliot, M. A. Willart, M. Kool,F. Muskens, and B. N. Lambrecht. 2010. Inflammatory dendritic cells–not ba-sophils–are necessary and sufficient for induction of Th2 immunity to inhaledhouse dust mite allergen. J. Exp. Med. 207: 2097–2111.
21. Cayrol, C., and J. P. Girard. 2014. IL-33: an alarmin cytokine with crucial rolesin innate immunity, inflammation and allergy. Curr. Opin. Immunol. 31: 31–37.
22. Oboki, K., T. Ohno, N. Kajiwara, K. Arae, H. Morita, A. Ishii, A. Nambu,T. Abe, H. Kiyonari, K. Matsumoto, et al. 2010. IL-33 is a crucial amplifier ofinnate rather than acquired immunity. Proc. Natl. Acad. Sci. USA 107: 18581–18586.
23. Kouzaki, H., K. Iijima, T. Kobayashi, S. M. O’Grady, and H. Kita. 2011. Thedanger signal, extracellular ATP, is a sensor for an airborne allergen and triggersIL-33 release and innate Th2-type responses. J. Immunol. 186: 4375–4387.
24. Haenuki, Y., K. Matsushita, S. Futatsugi-Yumikura, K. J. Ishii, T. Kawagoe,Y. Imoto, S. Fujieda, M. Yasuda, Y. Hisa, S. Akira, et al. 2012. A critical role ofIL-33 in experimental allergic rhinitis. J. Allergy Clin. Immunol. 130: 184–194.e11. doi:10.1016/j.jaci.2012.02.013
25. Albacker, L. A., V. Chaudhary, Y. J. Chang, H. Y. Kim, Y. T. Chuang,M. Pichavant, R. H. DeKruyff, P. B. Savage, and D. T. Umetsu. 2013. Invariantnatural killer T cells recognize a fungal glycosphingolipid that can induce airwayhyperreactivity. Nat. Med. 19: 1297–1304.
26. Yasuda, K., T. Muto, T. Kawagoe, M. Matsumoto, Y. Sasaki, K. Matsushita,Y. Taki, S. Futatsugi-Yumikura, H. Tsutsui, K. J. Ishii, et al. 2012. Contributionof IL-33-activated type II innate lymphoid cells to pulmonary eosinophilia inintestinal nematode-infected mice. Proc. Natl. Acad. Sci. USA 109: 3451–3456.
27. Imai, Y., K. Yasuda, Y. Sakaguchi, T. Haneda, H. Mizutani, T. Yoshimoto,K. Nakanishi, and K. Yamanishi. 2013. Skin-specific expression of IL-33 acti-vates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflam-mation in mice. Proc. Natl. Acad. Sci. USA 110: 13921–13926.
28. Halim, T. Y., R. H. Krauss, A. C. Sun, and F. Takei. 2012. Lung natural helpercells are a critical source of Th2 cell-type cytokines in protease allergen-inducedairway inflammation. Immunity 36: 451–463.
29. Halim, T. Y., C. A. Steer, L. Matha, M. J. Gold, I. Martinez-Gonzalez,K. M. McNagny, A. N. McKenzie, and F. Takei. 2014. Group 2 innate lymphoidcells are critical for the initiation of adaptive T helper 2 cell-mediated allergiclung inflammation. Immunity 40: 425–435.
30. Shimizu, M., A. Matsuda, K. Yanagisawa, T. Hirota, M. Akahoshi, N. Inomata,K. Ebe, K. Tanaka, H. Sugiura, K. Nakashima, et al. 2005. Functional SNPs inthe distal promoter of the ST2 gene are associated with atopic dermatitis. Hum.Mol. Genet. 14: 2919–2927.
31. Savinko, T., S. Matikainen, U. Saarialho-Kere, M. Lehto, G. Wang,S. Lehtimaki, P. Karisola, T. Reunala, H. Wolff, A. Lauerma, and H. Alenius.2012. IL-33 and ST2 in atopic dermatitis: expression profiles and modulation bytriggering factors. J. Invest. Dermatol. 132: 1392–1400.
32. Kim, B. S., M. C. Siracusa, S. A. Saenz, M. Noti, L. A. Monticelli,G. F. Sonnenberg, M. R. Hepworth, A. S. Van Voorhees, M. R. Comeau, andD. Artis. 2013. TSLP elicits IL-33-independent innate lymphoid cell responsesto promote skin inflammation. Sci. Transl. Med. 5: 170ra16.
33. Salimi, M., J. L. Barlow, S. P. Saunders, L. Xue, D. Gutowska-Owsiak, X. Wang,L. C. Huang, D. Johnson, S. T. Scanlon, A. N. McKenzie, et al. 2013. A role forIL-25 and IL-33-driven type-2 innate lymphoid cells in atopic dermatitis. J. Exp.Med. 210: 2939–2950.
34. Dharmage, S. C., A. J. Lowe, M. C. Matheson, J. A. Burgess, K. J. Allen, andM. J. Abramson. 2014. Atopic dermatitis and the atopic march revisited. Allergy69: 17–27.
35. Spergel, J. M., and A. S. Paller. 2003. Atopic dermatitis and the atopic march. J.Allergy Clin. Immunol. 112: S118–S127.
36. Matsumoto, K., and H. Saito. 2013. Epicutaneous immunity and onset of allergicdiseases: per-“eczema”tous sensitization drives the allergy march. Allergol. Int.62: 291–296.
37. Kamijo, S., T. Takai, T. Kuhara, T. Tokura, H. Ushio, M. Ota, N. Harada,H. Ogawa, and K. Okumura. 2009. Cupressaceae pollen grains modulate den-dritic cell response and exhibit IgE-inducing adjuvant activity in vivo. J.Immunol. 183: 6087–6094.
38. Ziegler, S. F., and D. Artis. 2010. Sensing the outside world: TSLP regulatesbarrier immunity. Nat. Immunol. 11: 289–293.
39. Takai, T. 2012. TSLP expression: cellular sources, triggers, and regulatorymechanisms. Allergol. Int. 61: 3–17.
40. Wan, H., H. L. Winton, C. Soeller, E. R. Tovey, D. C. Gruenert, P. J. Thompson,G. A. Stewart, G. W. Taylor, D. R. Garrod, M. B. Cannell, and C. Robinson.1999. Der p 1 facilitates transepithelial allergen delivery by disruption of tightjunctions. J. Clin. Invest. 104: 123–133.
41. Nakamura, T., Y. Hirasawa, T. Takai, K. Mitsuishi, M. Okuda, T. Kato,K. Okumura, S. Ikeda, and H. Ogawa. 2006. Reduction of skin barrier functionby proteolytic activity of a recombinant house dust mite major allergen Der f 1.J. Invest. Dermatol. 126: 2719–2723.
42. Hirasawa, Y., T. Takai, T. Nakamura, K. Mitsuishi, H. Gunawan, H. Suto,T. Ogawa, X. L. Wang, S. Ikeda, K. Okumura, and H. Ogawa. 2010. Staphy-lococcus aureus extracellular protease causes epidermal barrier dysfunction. J.Invest. Dermatol. 130: 614–617.
43. Adam, E., K. K. Hansen, O. Astudillo Fernandez, L. Coulon, F. Bex, X. Duhant,E. Jaumotte, M. D. Hollenberg, and A. Jacquet. 2006. The house dust mite al-lergen Der p 1, unlike Der p 3, stimulates the expression of interleukin-8 inhuman airway epithelial cells via a proteinase-activated receptor-2-independentmechanism. J. Biol. Chem. 281: 6910–6923.
44. Kato, T., T. Takai, K. Mitsuishi, K. Okumura, and H. Ogawa. 2005. Cystatin Ainhibits IL-8 production by keratinocytes stimulated with Der p 1 and Der f 1:biochemical skin barrier against mite cysteine proteases. J. Allergy Clin.Immunol. 116: 169–176.
45. Kato, T., T. Takai, T. Fujimura, H. Matsuoka, T. Ogawa, K. Murayama, A. Ishii,S. Ikeda, K. Okumura, and H. Ogawa. 2009. Mite serine protease activatesprotease-activated receptor-2 and induces cytokine release in human keratino-cytes. Allergy 64: 1366–1374.
46. Chambers, L., A. Brown, D. I. Pritchard, S. Sreedharan, K. Brocklehurst, andN. A. Kalsheker. 1998. Enzymatically active papain preferentially induces anallergic response in mice. Biochem. Biophys. Res. Commun. 253: 837–840.
47. Varughese, K. I., F. R. Ahmed, P. R. Carey, S. Hasnain, C. P. Huber, andA. C. Storer. 1989. Crystal structure of a papain-E-64 complex. Biochemistry 28:1330–1332.
48. Ando, T., K. Matsumoto, S. Namiranian, H. Yamashita, H. Glatthorn,M. Kimura, B. R. Dolan, J. J. Lee, S. J. Galli, Y. Kawakami, et al. 2013. Mastcells are required for full expression of allergen/SEB-induced skin inflammation.J. Invest. Dermatol. 133: 2695–2705.
49. Ando, T., W. Xiao, P. Gao, S. Namiranian, K. Matsumoto, Y. Tomimori,H. Hong, H. Yamashita, M. Kimura, J. Kashiwakura, et al. 2014. Critical role formast cell Stat5 activity in skin inflammation. Cell Reports 6: 366–376.
50. Savinko, T., P. Karisola, S. Lehtimaki, A. M. Lappetelainen, R. Haapakoski,H. Wolff, A. Lauerma, and H. Alenius. 2013. ST2 regulates allergic airwayinflammation and T-cell polarization in epicutaneously sensitized mice. J. Invest.Dermatol. 133: 2522–2529.
51. Palmer, C. N., A. D. Irvine, A. Terron-Kwiatkowski, Y. Zhao, H. Liao, S. P. Lee,D. R. Goudie, A. Sandilands, L. E. Campbell, F. J. Smith, et al. 2006. Common
10 DISTINCT SKIN/AIRWAY-SENSITIZING MECHANISMS TO PROTEASE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

loss-of-function variants of the epidermal barrier protein filaggrin are a majorpredisposing factor for atopic dermatitis. Nat. Genet. 38: 441–446.
52. Horimukai, K., K. Morita, M. Narita, M. Kondo, H. Kitazawa, M. Nozaki,Y. Shigematsu, K. Yoshida, H. Niizeki, K. Motomura, et al. 2014. Application ofmoisturizer to neonates prevents development of atopic dermatitis. J. AllergyClin. Immunol. 134: 824-830.e6. doi:10.1016/j.jaci.2014.07.060
53. Lack, G. 2008. Epidemiologic risks for food allergy. J. Allergy Clin. Immunol.121: 1331–1336.
54. Endo, Y., K. Hirahara, T. Iinuma, K. Shinoda, D. J. Tumes, H. K. Asou,N. Matsugae, K. Obata-Ninomiya, H. Yamamoto, S. Motohashi, et al. 2015. Theinterleukin-33-p38 kinase axis confers memory T helper 2 cell pathogenicity inthe airway. Immunity 42: 294–308.
55. Hara, K., K. Iijima, M. K. Elias, S. Seno, I. Tojima, T. Kobayashi,G. M. Kephart, M. Kurabayashi, and H. Kita. 2014. Airway uric acid is a sensorof inhaled protease allergens and initiates type 2 immune responses in respira-tory mucosa. J. Immunol. 192: 4032–4042.
The Journal of Immunology 11
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from




















