
Study of the interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA
15
181 Biochimica et Biophysica Acta, 606 (1980) 181--195 © Elsevier/North-Holland Biomedical Press BBA 99595 STUDY OF THE INTERACTION OF GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE WITH DNA MANUEL PERUCHO, JOSI~ SALAS and MARIA L. SALAS Centro de Biolog(a Molecular, Consejo Superior de Investigaciones Cient(ficas, Universidad Autdnoma de Madrid, Canto Blanco, Madrid-34 (Spain) (Received May 15th, 1979) Key words: DNA-binding protein; Glyceraldehyde-3-phosphate dehydrogenase; Lactate dehydrogenase Summary Glyceraldehyde-3-phosphate dehydrogenase binds to homologous and heterologous single-stranded but not double-stranded DNA. Binding to RNA, poly(A) and poly(dA-dT) has also been observed. Enzyme binding to these nucleic acids leads to the formation of an insoluble complex which can be sedimented at low speed. The interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA is strongly inhibited by NAD and NADH but not by NADP. Adenine nucleotides, which inhibit the dehydrogenase activity by competing with NAD for its binding site (Yang, S.T. and Deal, W.C., Jr. (1969) Biochemistry 8, 2806-- 2813), also inhibit enzyme binding to DNA, whereas glyceraldehyde-3- phosphate and inorganic phosphate are non-inhibitory. These results suggest that DNA interacts through the NAD binding sites of glyceraldehyde-3- phosphate dehydrogenase. In accordance with this idea, it was found that DNA also binds to lactate dehydrogenase, an enzyme containing a similar dinucleotide binding domain, and that this binding is inhibited by NADH. A study of the base specificity of the DNA-glyceraldehyde-3-phosphate dehydrogenase interaction using dinucleoside monophosphates shows that inhibition of DNA binding by the dinucleotides requires the presence of a 3'-terminal adenosine and is greater when the 5'-terminus contains a pyrimidine instead of a purine. These results suggest that the dinucleotides bind at the NAD site of the dehydrogenase and that the enzyme would interact preferenti- ally with PypA dinucleotides present in the nucleic acid. Abbreviation: HaSV, hamster sarcoma virus; PypA, pyridynyl(3' -* 5')adenosine.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Study of the interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA

181
Biochimica et Biophysica Acta, 606 (1980) 181--195 © Elsevier/North-Holland Biomedical Press
BBA 99595
STUDY OF THE INTERACTION OF GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE WITH DNA
MANUEL PERUCHO, JOSI~ SALAS and MARIA L. SALAS
Centro de Biolog(a Molecular, Consejo Superior de Investigaciones Cient(ficas, Universidad Autdnoma de Madrid, Canto Blanco, Madrid-34 (Spain)
(Received May 15th, 1979)
Key words: DNA-binding protein; Glyceraldehyde-3-phosphate dehydrogenase; Lactate dehydrogenase
Summary
Glyceraldehyde-3-phosphate dehydrogenase binds to homologous and heterologous single-stranded but not double-stranded DNA. Binding to RNA, poly(A) and poly(dA-dT) has also been observed. Enzyme binding to these nucleic acids leads to the formation of an insoluble complex which can be sedimented at low speed.
The interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA is strongly inhibited by NAD and NADH but not by NADP. Adenine nucleotides, which inhibit the dehydrogenase activity by competing with NAD for its binding site (Yang, S.T. and Deal, W.C., Jr. (1969) Biochemistry 8, 2806-- 2813), also inhibit enzyme binding to DNA, whereas glyceraldehyde-3- phosphate and inorganic phosphate are non-inhibitory. These results suggest that DNA interacts through the NAD binding sites of glyceraldehyde-3- phosphate dehydrogenase. In accordance with this idea, it was found that DNA also binds to lactate dehydrogenase, an enzyme containing a similar dinucleotide binding domain, and that this binding is inhibited by NADH.
A study of the base specificity of the DNA-glyceraldehyde-3-phosphate dehydrogenase interaction using dinucleoside monophosphates shows that inhibition of DNA binding by the dinucleotides requires the presence of a 3'-terminal adenosine and is greater when the 5'-terminus contains a pyrimidine instead of a purine. These results suggest that the dinucleotides bind at the NAD site of the dehydrogenase and that the enzyme would interact preferenti- ally with PypA dinucleotides present in the nucleic acid.
Abbrev ia t ion: HaSV, h a m s t e r s a r c o m a virus; PypA, p y r i d y n y l ( 3 ' -* 5 ' )adenos ine .

182
Introduct ion
We have recently shown that the mammalian DNA binding protein P8 is the glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase [1]. Using a Millipore filtration technique it was found that the pure enzyme from hamster fibroblasts and from hamster or rabbit muscle interacts with denatured but not with native DNA. This interaction is strongly inhibited by NAD suggesting that the dinucleotide fold, which is common to dehydrogenases, is involved in DNA binding [ 1 ].
A similar nucleotide binding domain appears to be present in a number of other enzymes, some of which also bind nucleotides (see Ref. 2 for a review). On the basis of these findings, Rossmann et al. [2] proposed that the nucleotide binding unit of these enzymes was derived from a common ancestor. Further- more, they postulate that this basic structure may have also been incorporated, during evolution, into at least some nucleic acid-related proteins for DNA or RNA binding.
It was therefore thought of interest to examine further the characteristics of the interaction between glyceraldehyde-3-phosphate dehydrogenase and DNA, and to investigate whether another enzyme with a similar dinucleotide binding site, such as lactate dehydrogenase, also binds to DNA.
Materials and Methods
[3H]Uridine was obtained from the Radiochemical Center (Amersham). [3H]Poly(dA-dT) and [14C]poly(A) were purchased from Miles Laboratories, Inc. Nicotinamide, nucleotides and dinucleoside monophosphates were from Sigma Chemical Co. Rabbit muscle lactate dehydrogenase was obtained from Boehringer, Mannheim.
14C- or 32p-labeled DNA from bacteriophage ~b29 and RNA polymerase from Bacillus subtilis were kindly donated by Dr. M. Salas. Radioactive DNA from the Syrian hamster fibroblast line NIL-l-hamster sarcoma virus (HaSV) was isolated as indicated before [1], after labeling for 48 h exponentially growing cultures with 2 pCi of [3H]thymidine (20 Ci/mmol) or 0.5 pCi of [14C]- thymidine (2 Ci/mmol)/ml of culture medium. To prepare [3H]RNA from NIL-1-HaSV cells, exponentially growing cultures were labeled for 18 h with 2 pCi of [3H]uridine (29 Ci/mmol)/ml of medium. The RNA was extracted from whole cells by the hot phenol extraction procedure described by Warner et al. [3] as modified by Edmonds and Grace Caramela [4].
All other materials were as described previously [ 1,5].
Assay of glyceraldehyde-3ophosphate dehydrogenase Glyceraldehyde-3-phosphate dehydrogenase activity was determined as
described before [1]. To test the effect of UpA, UpG and ATP on enzyme activity, the same assay conditions were used, except that the pyrophosphate buffer was replaced by 0.1 M Tris-HC1, pH 7, and the concentration of NAD was 0.6 mM.
Purification of glyceraldehyde-3-phosphate dehydrogenase Glyceraldehyde-3-phosphate dehydrogenase from NIL-1-HaSV cells [5] was

183
purified by DNA-cellulose and phosphocellulose chromatography as described previously [1]. To prepare radioactive glyceraldehyde-3-phosphate dehydro- genase, dense NIL-1-HaSV cultures were labeled for 48 h with 2.5 nCi of [3H]- proline (1 Ci/mmol)/ml of culture medium and the enzyme was purified as before. To purify glyceraldehyde-3-phosphate dehydrogenase from Chinese hamster muscle, the homogenate was prepared as described by Velick [6], except that 40 mM Tris-HC1, pH 8.1, 1 mM 2-mercaptoethanol, 0.2 mM EDTA and 10 mM MgCl: was used as extraction buffer. The homogenate was then sonicated, filtered through cheesecloth and centrifuged for 30 min at 20 000 X g. The extract was treated with DNAase and chromatographed on DNA-cellulose and phosphocellulose columns as indicated [1]. These prepara- tions were essentially homogeneous, as judged by dodecyl sulfate gel electro- phoresis [1]. The specific activity of the enzyme from NIL-1-HaSV or hamster muscle was 50--60 units/mg.
Interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA Membrane filtration assay. This assay was performed as described before [1].
Precise conditions are shown in the legends to the tables and figures. Precipitation of the DNA-glyceraldehyde-3-phosphate dehydrogenase com-
plex. Incubations were as described in the membrane filtration assay, using conical 1 ml Sorvall centrifuge tubes. After incubation for 5 min at different temperatures, the mixtures were centrifuged at 8000 rev./min for 10 min in the rotor SS-34 of a Sorvall centrifuge at the incubation temperature. The super- natants were pipet ted on glass fiber discs, Whatman GF/A, 2.4 cm in diameter. The sediments were dissolved by heating for 2 min in 0.1 ml of binding buffer [1] containing 1% sodium dodecyl sulfate and were then adsorbed into What- man GF/A discs. The discs were dried and the radioactivity was counted [5].
Results
Interaction of glyceraldehyde-3-phosphate dehydrogenase with homologous and heterologous DNA
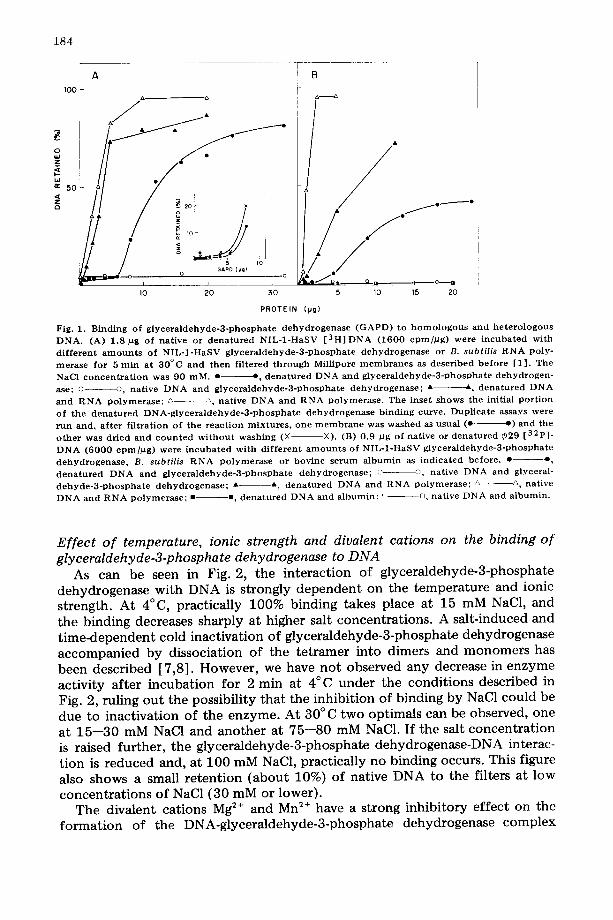
The binding of glyceraldehyde-3-phosphate dehydrogenase from NIL-1- HaSV cells to homologous and heterologous DNA was studied by the Millipore filter technique (Fig. 1). In agreement with previous results [1], the enzyme does no t bind to native NIL-1-HaSV (Fig. 1A) or phage ~b29 (Fig. 1B) DNA. In contrast, glyceraldehyde-3-phosphate dehydrogenase binds to single-stranded homologous (Fig. 1A) and heterologous (Fig. 1B) DNA. When increasing amounts of protein are added to a fixed concentration of DNA, a sigmoidal curve is obtained. The inset in Fig. 1A shows that the shape of the curve is not altered by decreasing the degree of washing of the Millipore filters.
As a comparison, the interaction of highly purified B. subtilis RNA polymer- ase with single-stranded and double-stranded DNA from NIL-1-HaSV and phage ¢29 was examined. As shown in Fig. 1, the polymerase binds to native and denatured DNA from both sources. On the other hand, no binding of native or denatured DNA to the filters is detected with bovine serum "albumin (Fig. 1B).
The rate of binding of glyceraldehyde-3-phosphate dehydrogenase to single- stranded DNA is quite rapid, the interaction being completed in 2 min at 30°C (data not shown).
184
I00 1 A a a
i . / . . 5O
,
° il
o GAPD {pg) o
i l
I 0 20 30
B
o / o ,--------- •
5 10 15 20
PROTEIN (pg)
Fig. 1. B i n d i n g o f g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ( G A P D ) to h o m o l o g o u s and h e t e r o l o g o u s D N A . (A) 1 .8 pg of nat ive or d e n a t u r e d N I L - 1 - H a S V [ 3 H ] D N A ( 1 6 0 0 c p m / p g ) were i n c u b a t e d w i t h
d i f f e r e n t a m o u n t s o f N I L - I - H a S V g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e or B. subtilis R N A poly- • erase for 5 m i n at 3 0 ° C a n d t h e n f i l t e red t h r o u g h Mil l ipore m e m b r a n e s as desc r ibed b e f o r e [1 ] . The
NaCl c o n c e n t r a t i o n was 90 m M . • e , d e n a t u r e d D N A a n d g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n -
ase; o o, nat ive D N A and g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ; A A d e n a t u r e d D N A
a n d R N A p o l y m e r a s e ; ~- ~, na t ive D N A a n d R N A p o l y m e r a s e . The in se t s h o w s the init ial p o r t i o n o f the d e n a t u r e d D N A - g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e b i n d i n g curve . Dup l i ca t e assays were
r u n and , a f t e r f i l t rat ion o f t h e r e a c t i o n m i x t u r e s , o n e m e m b r a n e was w a s h e d as usua l (e- e ) and the o t h e r was d r i ed a n d c o u n t e d w i t h o u t w a s h i n g (× ×). (B) 0 .9 #g of nat ive or d e n a t u r e d ~29 [ 3 2 p ] .
D N A ( 6 0 0 0 e p m / p g ) were i n c u b a t e d w i t h d i f f e r e n t a m o u n t s o f N I L - 1 - H a S V g l y c e r a l d e h y d e - 3 - p h o s p h a t e
d e h y d r o g e n a s e , B. subtilis R N A p o l y m e r a s e or bov ine s e r u m a l b u m i n as i n d i c a t e d be fo re . • o , d e n a t u r e d D N A a n d g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ; ~ ©, nat ive D N A a n d glyceral-
d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ; • A d e n a t u r e d D N A and R N A p o l y m e r a s e ; /, A nat ive D N A and R N A p o l y m e r a s e ; • =, d e n a t u r e d D N A and a l b u m i n ; ~ n nat ive D N A and a l b u m i n .
Effect of temperature, ionic strength and divalent cations on the binding of glyceraldehyde-3-phosphate dehydrogenase to DNA
As can be seen in Fig. 2, the interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA is strongly dependent on the temperature and ionic strength. At 4°C, practically 100% binding takes place at 15 mM NaC1, and the binding decreases sharply at higher salt concentrations. A salt-induced and time-dependent cold inactivation of glyceraldehyde-3-phosphate dehydrogenase accompanied by dissociation of the tetramer into dimers and monomers has been described [7,8] . However, we have not observed any decrease in enzyme activity after incubation for 2 min at 4°C under the conditions described in Fig. 2, ruling out the possibility that the inhibition of binding by NaC1 could be due to inactivation of the enzyme. At 30°C two optimals can be observed, one at 15--30 mM NaC1 and another at 75--80 mM NaC1. If the salt concentration is raised further, the glyceraldehyde-3-phosphate dehydrogenase-DNA interac- tion is reduced and, at 100 mM NaC1, practically no binding occurs. This figure also shows a small retention (about 10%) of native DNA to the filters at low concentrations of NaC1 (30 mM or lower).
The divalent cations Mg 2÷ and Mn 2÷ have a strong inhibitory effect on the formation of the DNA-glyceraldehyde-3-phosphate dehydrogenase complex

185
100 a
Q
\ 25 50 75 100 125 150 175
No CL (raM)
Fig. 2. Ef fec t of ionic s t r eng th a nd t e m p e r a t u r e on the b inding of g ly ce r a id eh y d e -3 -p h o sp h a t e d e h y d r o - genase to DNA. 12 pg of NIL-1 -HaSV g lyce ra idehyde -3 -phospha t e d e h y d r o g e n a s e were i n c u b a t e d wi th 1.6 pg of d e n a t u r e d or na t ive N I L - I - H a S V [ 3 H ] D N A (160 0 cpm/12g) for 2 m in a t 4°C or 30°C an d f i l tered t h r o u g h Millipore m e m b r a n e s as descr ibed [1 ] . The filters were washed wi th b ind ing bu f f e r k e p t at the i n c u b a t i o n t e m p e r a t u r e and con ta in ing the NaCI c o n c e n t r a t i o n of the i n c u b a t i o n mix tu re . • --, d e n a t u r e d D NA and 30°C; o o, na t ive DNA an d 30°C; o D d e n a t u r e d D N A and 4°C.
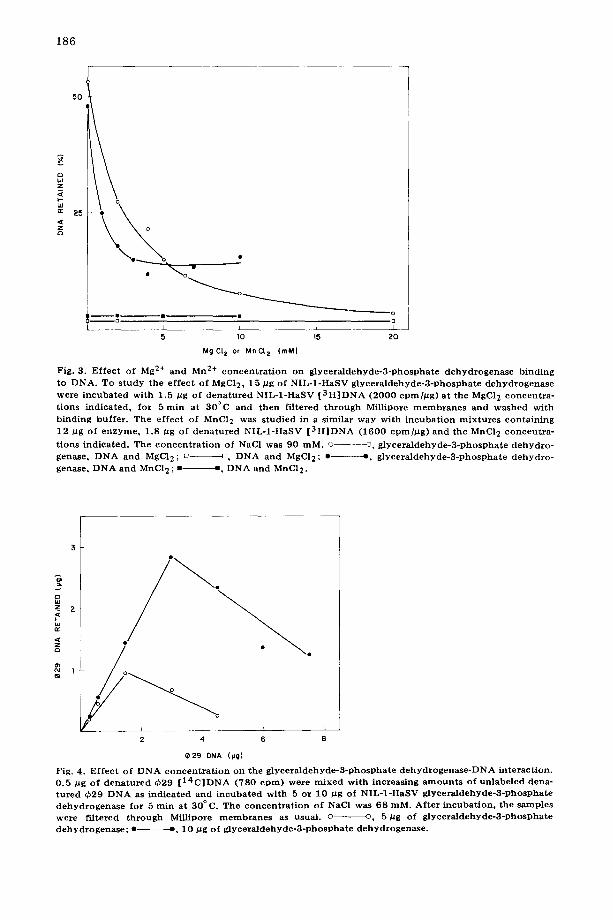
(Fig. 3). About 50% inhibition is obtained at 2 mM MgC12 or 1 mM MnC12. Almost total inhibition is observed at 20 mM MgC12, whereas in the case of Mn 2+, complete inhibition is not reached, and approximately 30% of the DNA- protein complex appears to be resistant to this metal ion.
It should be mentioned that the complex preformed in a 5 min incubation at 30°C and 75 mM NaC1 cannot be destroyed by the addition of 12.5 mM MgC12 or by lowering the temperature at 4°C and continuing the incubation for a further 10 min period (results not shown).
Effect o f DNA concentration on the glyceraldehyde-3-phosphate dehydrogen- ase-DNA interaction
The DNA saturation curves for two different concentrations of glyceralde- hyde-3-phosphate dehydrogenase (5 and 10 pg) are shown in Fig. 4. At both enzyme concentrations retention of DNA on the filter is inhibited at DNA- glyceraldehyde-3-phosphate dehydrogenase ratios above 0.3. From the maximal amount of DNA retained at each protein concentration, and taking into account the molecular weight of single-stranded ~b29 DNA (6 • 106) (Talavera, A., personal communication) and that of native glyceraldehyde-3-phosphate dehydrogenase (145 000) [9,10] it can be calculated that about 150 enzyme molecules are bound by one ¢29 DNA molecule or that one enzyme molecule is bound per 120 nucleotides. In this calculation it has been assumed that all the protein is interacting with the DNA. (However, see later.)
186
4
50
25 •
!
., • • - - o D
I I 5 llO li5 20
MgCt z or MnCt 2 (rnM}
Fig. 3. E f fec t of Mg 2+ and Mn 2+ c o n c e n t r a t i o n on g l y c e r a l d e h y d c - 3 - p h o s p h a t e d e h y d r o g e n a s c b ind ing to DNA. To s t udy the e f fec t of MgC12, 15 #g of N I L - 1 - H a S V g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e were i n c u b a t e d wi th 1.5 #g of d e n a t u r e d N I L - 1 - H a S V [ 3 H ] D N A (2 0 0 0 cpmhug) a t the MgC12 c o n c e n t r a - t ions ind ica ted , for 5 rain at 30°C a nd then f i l tered t h r o u g h Millipore m e m b r a n e s an d w a s h e d wi th b ind ing buf fe r . The e f fec t of MnC12 was s tud ied in a s imilar way wi th i n c u b a t i o n m i x t u r e s con ta in ing 12 /~g of e n z y m e , 1.8 /zg of d e n a t u r e d N I L - 1 - H a S V [ 3 H ] D N A (1 6 0 0 cpm]/Zg) an d the MnCI 2 c o n c e n t r a -
t ions ind ica ted . The c o n c e n t r a t i o n of NaC1 was 90 raM. © - - - - - ~ , g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o - genase, D N A and MgCI2; o D DNA a nd MgC12; • • , g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o - genase, D N A and MnC12 ; • • , D N A a nd MnC12.
Z 2
c3
2 4 6 8
e 2 9 DNA (IJg)
Fig. 4. E f fec t of D N A c o n c e n t r a t i o n on the g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e - D N A in te rac t ion . 0 .5 pg of d e n a t u r e d ~b29 [ 1 4 C ] D N A (780 c p m ) were m i x e d wi th increas ing a m o u n t s of un l abe l ed dena- t u r e d qb29 D N A as ind ica ted a nd i n c u b a t e d wi th 5 or 10 /~g of N I L - 1 - H a S V g ly ce r a ld eh y d e -3 -p h o sPh a t e d e h y d r o g e n a s e for 5 rain at 30 ° C. The c o n c e n t r a t i o n of NaC1 was 68 raM. Af t e r i ncuba t ion , the samples were f i l tered t h r o u g h Millipore m e m b r a n e s as usual , o ©, 5/zg of g l y e e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ; • ¢, 10 / Jg of g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e .

187
Precipitation of the DNA-glyceraldehyde-3-phosphate dehydrogenase complex Generally, the binding of DNA to proteins can be demonstrated by nitro-
cellulose membrane filtration because the proteins and, therefore, the DNA- protein complex, are retained on the filter, while free DNA is not [11,12]. However, when purified 3H-labeled glyceraldehyde-3-phosphate dehydrogenase is passed through a Millipore filter, only about 2--4% binds to the filter. As shown in Table I, the interaction of glyceraldehyde-3-phosphate dehydrogenase with single-stranded [3H]DNA leads to the formation of an insoluble complex which can be sedimented at low speed. No precipitation is detected when DNA is incubated in the absence of protein (Table I) and the protein does not precipitate when incubated without DNA (data not shown). It can be seen in Table I that the cpm recovered in the sediment agree with those retained in the Millipore filter, indicating that the filters are simply acting as sieves which retain the precipitate. No precipitation of native DNA by glyceraldehyde-3- phosphate dehydrogenase has been observed (data not shown).
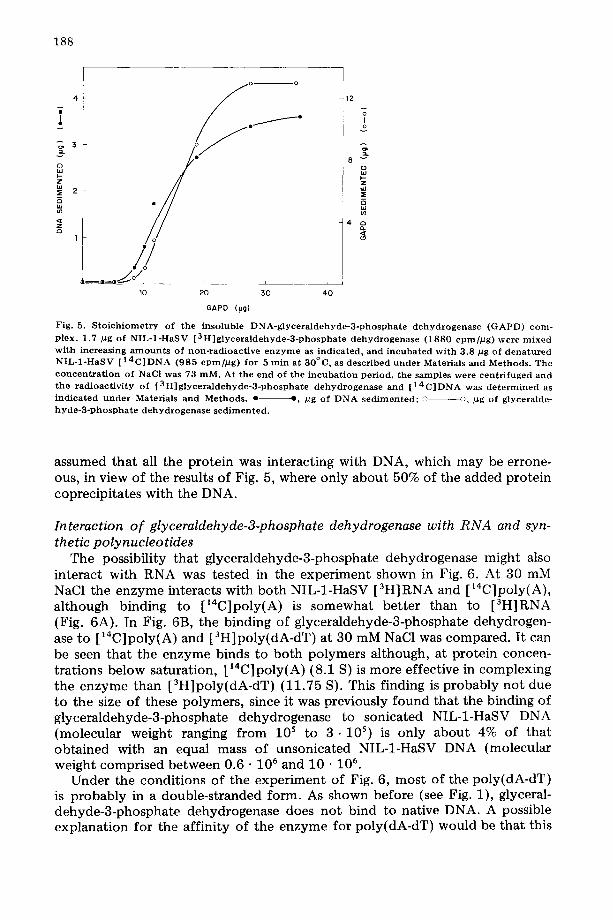
As shown in Fig. 5, glyceraldehyde-3-phosphate dehydrogenase coprecipi- tates with the DNA. In this experiment, a fixed amount of denatured NIL-1- HaSV [14C]DNA was incubated with increasing concentrations of 3H-labeled glyceraldehyde-3-phosphate dehydrogenase and, after centrifuged, the radio- activity corresponding to the DNA and the protein was measured in the sedi- ment. When the amount of DNA or enzyme precipitated is plotted as a func- tion of enzyme concentration, sigmoidal curves are generated, similar to those obtained when DNA binding to glyceraldehyde-3-phosphate dehydrogenase is assayed by the Millipore filtration technique. From the data in Fig. 5, a stoichiometry of about 122 nucleotides per protein molecule can be deter- mined, taking 145 000 for the molecular weight of native glyceraldehyde-3- phosphate dehydrogenase. However, this may be only a maximum value if the DNA can precipitate when not fully saturated with protein. Although the stoichiometry calculated in this experiment agrees with that obtained from the data in Fig. 4, it should be noted that in the experiment of Fig. 4 it was
T A B L E I
P R E C I P I T A T I O N OF D E N A T U R E D DNA BY G L Y C E R A L D E H Y D E - 3 - P H O S P H A T E D E H Y D R O G E N - ASE
1 .75 /~g of d e n a t u r e d NIL-1 -HaSV [ 3 H ] D N A (1600 c p m ] p g ) were incubated w i t h or w i t h o u t 12 pg of NIL-1 -HaSV g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e for 5 m i n as descr ibed b e f o r e [1 ] , at the t em- pe ra tu r e and NaCI c o n c e n t r a t i o n s h o w n . Af ter incubat ion , the m i x t u r e s w e r e e i ther centr i fuged (see Mate- rials and M e t h o d s ) or f i l tered t h r o u g h Millipore m e m b r a n e s [1 ] .
[3 H] DNA in c u b ate d w i t h glyceral- d e h y d e - 3 - p h o s p h a t e d e h yd r oge n ase (cPm)
T e m p e r a t u r e (°C): 30 4 30 NaC1 c o n c e n t r a -
t i o n ( r a M ) : 15 75 15 75 15
[ 3 H ] DNA incubated w i t h o u t glyceral- d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ( c p m )
75 15 75
Supernatant 184 317 218 2706 S e d i m e n t 2447 2395 2715 140 Mill ipore fil ter 2464 2123 2 2 6 0 75
2285 2431 2512 2532 80 53 100 107
- - - - 51 49
188
4 A
I.-
2
co
~3 1
o o
~of f °
12
o
-g 8
bJ
4 2
4O 10 20 3 0
G A P D (lug)
Fig. 5. S t o i c h i o m e t r y of the insoluble D N A - g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ( G A P D ) c o m - plex. 1.7 #g of N I L - 1 - H a S V [ 3 H ] g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e (1880 c p m / p g ) w e r e m i x e d wi th increas ing a m o u n t s of non- rad ioac t ive e n z y m e as ind ica ted , and i n c u b a t e d wi th 3.8 pg of d e n a t u r e d N I L - 1 - H a S V [ 1 4 C ] D N A (985 c p m / p g ) for 5 rain at 30°C, as descr ibed u n d e r Materials and Methods . The c o n c e n t r a t i o n of NaCl was 73 raM. At the end of the i n c u b a t i o n per iod , the samples were cen t r i fuged and t h e r ad ioac t iv i ty of [ 3 H ] g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e and [ 1 4 C ] D N A was d e t e r m i n e d as i nd i ca t ed u n d e r Materials and Methods . e - - e , pg of D N A s e d i m e n t e d ; ~ ---~), ~g of glyceralde- h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e s e d i m e n t e d .
assumed that all the protein was interacting with DNA, which may be errone- ous, in view of the results of Fig. 5, where only about 50% of the added protein coprecipitates with the DNA.
Interaction of glyceraldehyde-3-phosphate dehydrogenase with RNA and syn- the tic poly n ucleo tides
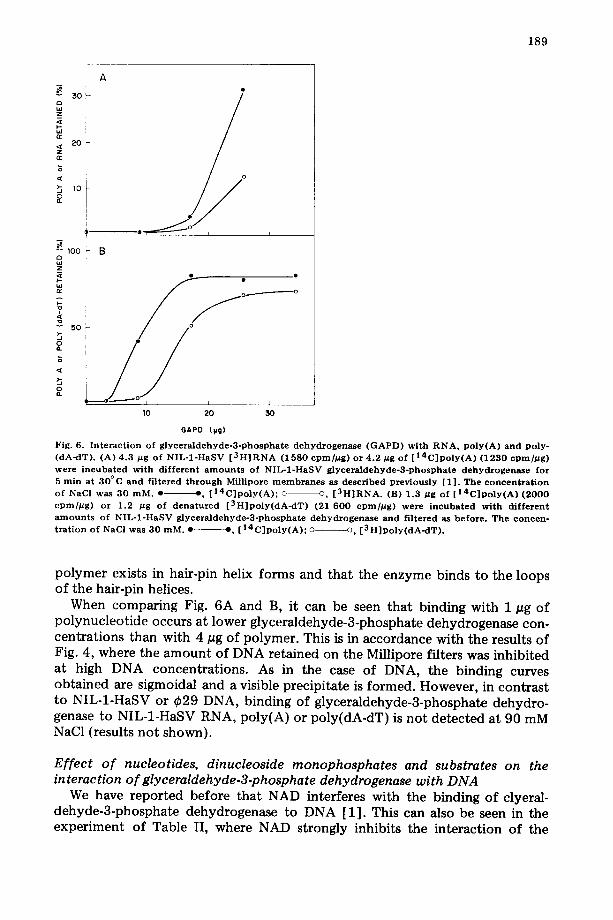
The possibility that glyceraldehyde-3-phosphate dehydrogenase might also interact with RNA was tested in the experiment shown in Fig. 6. At 30 mM NaC1 the enzyme interacts with both NIL-1-HaSV [ 3H] RNA and [ ~4C]poly(A), although binding to [~4C]poly(A) is somewhat bet ter than to [3H]RNA (Fig. 6A). In Fig. 6B, the binding of glyceraldehyde-3-phosphate dehydrogen- ase to [14C]poly(A) and [3H]poly(dA-dT) at 30 mM NaC1 was compared. It can be seen that the enzyme binds to both polymers although, at protein concen- trations below saturation, [14C]poly(A) (8.1 S) is more effective in complexing the enzyme than [3H]poly(dA-dT) (11.75 S). This finding is probably not due to the size of these polymers, since it was previously found that the binding of glyceraldehyde-3-phosphate dehydrogenase to sonicated NIL-1-HaSV DNA (molecular weight ranging from l 0 s to 3 . 1 0 s) is only about 4% of that obtained with an equal mass of unsonicated NIL-1-HaSV DNA (molecular weight comprised between 0.6 • 106 and 10 • 106.
Under the conditions of the experiment of Fig. 6, most of the poly(dA-dT) is probably in a double-stranded form. As shown before (see Fig. 1), glyceral- dehyde-3-phosphate dehydrogenase does not bind to native DNA. A possible explanation for the affinity of the enzyme for poly(dA-dT) would be that this
a)
Z
20
lO
100
Z
50
o
189
10 20 30
GAPD (pg)
O
Fig. 6. I n t e r a c t i o n o f g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e ( G A P D ) w i t h R N A , P o l y ( A ) a n d p o l y - ( d A - d T ) . (A) 4 .3 p g o f N I L - 1 - H a S V [ 3 H ] R N A ( 1 5 8 0 cpmfl~g) o r 4 . 2 ~tg o f [ 1 4 C ] p o l y ( A ) ( 1 2 3 0 c p m / D g ) were i n c u b a t e d w i t h d i f f e r e n t a m o u n t s o f N I L - 1 - H a S V g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e f o r 5 r a in a t 3 0 ° C a n d f i l t e r ed t h r o u g h Mfl l ipore m e m b r a n e s as d e s c r i b e d p r e v i o u s l y [ 1 ] . The c o n c e n t r a t i o n o f NaC] was 3 0 m M . • $, [ 1 4 C ] p o l y ( A ) ; © o, [ 3 H ] R N A . (B) 1 .3 p g o f [ 1 4 C ] p o l y ( A ) ( 2 0 0 0 c p m / # g ) o r 1 .2 /~g o f d e n a t u r e d [ 3 H ] p o l y ( d A - d T ) (21 6 0 0 cpmfl~g) we re i n c u b a t e d w i t h d i f f e r e n t a m o u n t s o f N I L - 1 - H a S V g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e a n d f i l t e red as b e f o r e . T h e c o n c e n - t r a t i o n o f NaCI was 3 0 m M . • --, [ 1 4 C ] p o l y ( A ) ; o o , [ 3 H ] p o l y ( d A - d T ) .
polymer exists in hair-pin helix forms and that the enzyme binds to the loops of the hair-pin helices.
When comparing Fig. 6A and B, it can be seen that binding with 1 pg of polynucleot ide occurs at lower glyceraldehyde-3-phosphate dehydrogenase con- centrations than with 4 #g of polymer. This is in accordance with the results of Fig. 4, where the amount of DNA retained on the Millipore filters was inhibited at high DNA concentrations. As in the case of DNA, the binding curves obtained are sigmoidal and a visible precipitate is formed. However, in contrast to NIL-1-HaSV or ~b29 DNA, binding of glyceraldehyde-3-phosphate dehydro- genase to NIL-1-HaSV RNA, poly(A) or poly(dA-dT) is not detected at 90 mM NaC1 (results not shown).
Effect o f nucleotides, dinucleoside monophosphates and substrates on the interaction o f glyceraldehyde-3-phosphate dehydrogenase with DNA
We have reported before that NAD interferes with the binding of clyeral- dehyde-3-phosphate dehydrogenase to DNA [1]. This can also be seen in the experiment of Table II, where NAD strongly inhibits the interaction of the

1 9 0
T A B L E II
I N H I B I T I O N BY N A D O F T H E B I N D I N G O F G L Y C E R A L D E H Y D E - 3 - P H O S P H A T E D E H Y D R O G E N -
ASE F R O M N I L - 1 - H a S V A N D H A M S T E R M U S C L E TO D N A : C O E N Z Y M E S P E C I F I C I T Y
The b i n d i n g assay was p e r f o r m e d as i n d i c a t e d b e f o r e [1] w i t h S0 m M NaC1, 0 .74 pg o f d e n a t u r e d NIL-1- H a S V [ 1 4 C ] D N A (14 900 c p m / ~ g ) and e i t he r 13.5 pg of N I L - 1 - H a S V or 16 pg of h a m s t e r m u s c l e
g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e . The d i n u c l e o t i d e s were a d d e d at a f inal c o n c e n t r a t i o n of 10
uM. The o rde r of a d d i t i o n of r e a g e n t s was : b i n d i n g b u f f e r , d inuc l e o t i de ( w h e n p re sen t ) , D N A and e n z y m e . I n c u b a t i o n s were for 4 m i n at 30°C . The n u m b e r s in p a r e n t h e s e s r e p r e s e n t the re la t ive a m o u n t
o f D N A r e t a i n e d (%). Data ind ica te [ 14 C ] D N A r e t a i n e d ( c p m ) .
A d d i t i o n s G l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e f r o m :
N I L - 1 - H a S V H a m s t e r musc l e
N o n e 10 127 (100%) 10 905 (100%) N A D 1 980 (20%) 1 486 (14%)
N A D H 1 897 (19%) 2 172 (20%)
N A D P 10 471 (104%) 11 387 (104%)
enzyme from NIL-1-HaSV or hamster muscle with DNA. In this experiment, NAD and the nucleic acid were added before the enzyme, since incubation of glyceraldehyde-3-phosphate dehydrogenase with DNA (4 min at 30 ° C) prior to the addition of NAD protects the enzyme against the effect of the dinucleotide (data not shown). NADH, whose affinity for glyceraldehyde-3-phosphate dehydrogenase is similar to that of NAD [13], also prevents enzyme binding to DNA, whereas NADP, which cannot replace NAD as coenzyme [13], does not inhibit the binding. These results suggest that DNA interacts with glyceral- dehyde-3-phosphate dehydrogenase through the dinucleotide binding fold present in the enzyme.
Adenine nucleotides have been found to inhibit glyceraldehyde-3-phosphate dehydrogenase activity by competing with NAD for its binding site [14,15]. It seemed possible, therefore, that these compounds would also inhibit enzyme binding to DNA. As shown in Table III, ATP, ADP, AMP and ADP-ribose inter- fere with the binding of glyceraldehyde-3-phosphate dehydrogenase to DNA, ATP being the most potent inhibitor. The inhibition appears to be specific for adenine compounds, since 1 mM UMP or CMP have no effect on DNA binding (data not shown).
ATP, like salt (see above), inactivates, at 0°C, glyceraldehyde-3-phosphate dehydrogenase by dissociating the subunits of the tetramer [7,16]. However, the inhibition of binding by ATP is not due to inactivation of the enzyme, since after incubation with 1 mM ATP as indicated in Table III the activity of glyceraldehyde-3-phosphate dehydrogenase does not change.
As can be seen in Table III neither nicotinamide nor NMN have any effect on the interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA, and a combination of NMN with AMP does not cause greater inhibition of binding than AMP alone. These results are consistent with the observation that nicotin- amide compounds do not inhibit the dehydrogenase activity [14].
Glyceraldehyde-3-phosphate and inorganic phosphate are also non-inhibitory of the enzyme-DNA interaction (Table III), indicating that the binding sites for these compounds are not involved in enzyme binding to DNA. The data in

191
T A B L E III
E F F E C T OF N U C L E O T I D E S , G L Y C E R A L D E H Y D E - 3 - P H O S P H A T E A N D I N O R G A N I C P H O S P H A T E ON T H E B I N D I N G OF H A M S T E R M U S C L E G L Y C E R A L D E H Y D E - 3 - P H O S P H A T E D E H Y D R O G E N - ASE TO D N A
0.48 #g o f dena tured NIL-1 -HaSV [ 3 H ] D N A (326 000 c p m / ~ g ) w e r e incuba ted w i t h 19 ~g of ha mster m u s c l e g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e for 4 rain at 3 0 ° C , as descr ibed b e f o r e [1 ]. The NaC1 c o n c e n t r a t i o n w as 80 mM and addi t ions w e r e as ind icated . The e n z y m e was added last. Af ter incubat ion , the samples w e r e f i l tered through Mil l ipore m e m b r a n e s as usual. 125 0 0 0 c p m of [ 3 H ] D N A w e r e b o u n d w i t h o u t added c o m p o u n d s .
Addi t ions Relat ive a m o u n t o f [ 3 H] DN A reta ined (%)
N o n e 100
0.1 m M ATP 100 0 .25 m M A T P 50 I m M A T P 14
0 .25 m M ADP 102 1 m M ADP 36
0 .25 m M AMP 95 0.5 m M AMP 60 1 m M AMP 29
0 .25 m M ADP-ribose 83 1 m M ADP-r ibose 57
1 m M n i c o t i n a m i d e 101 1 m M NMN 97
0.5 m M AMP + 1 m M NMN 76 1 m M g l yce r a l dehyde -3 -phos pha t e 96 5 mM inorganic phosphate 97
Table III add further support to the idea that the interaction of glyceraldehyde- 3-phosphate dehydrogenase with DNA takes place through the NAD binding site on the enzyme.
In the experiments shown in Table IV we have utilized dinucleoside mono- phosphates as model compounds to test the base specificity of the interaction between nucleic acids and glyceraldehyde-3-phosphate dehydrogenase. The results indicate that the inhibition of DNA binding by these compounds depends on their base composit ion. Thus, the presence of a 3'-terminal adeno- sine is a requirement, and the extent of inhibition is greater with dinucleotides containing a pyrimidine instead of a purine at the 5'-terminus.
In agreement with the above findings, 6 mM UpA inhibits 32% of the glyceraldehyde-3-phosphate dehydrogenase activity, in the assay conditions described under Materials and Methods, whereas 6 mM UpG is non-inhibitory. In the same conditions, 6 mM ATP also inhibits 30% of the enzyme activity.
Binding of lactate dehydrogenase to DNA We have investigated whether lactate dehydrogenase, an enzyme containing a
similar dinucleotide binding site [2] , also interacts with DNA. As shown in Fig. 7, this enzyme, like glyceraldehyde-3-phosphate dehydrogenase, binds to denatured but not to native DNA.
The ability of NAD or NADH to inhibit the interaction between lactate

192
T A B L E IV
E F F E C T OF D I N U C L E O S I D E M O N O P H O S P H A T E S ON T H E B I N D I N G OF H A M S T E R M U S C L E G L Y C E R A L D E H Y D E - 3 - P H O S P H A T E D E H Y D R O G E N A S E TO D N A
The b ind ing assay was p e r f o r m e d as desc r ibed be fo re [1 ] . Exp t . 1 : 0 . 4 8 pg of d e n a t u r e d NIL-1 -HaSV [ 3 H ] D N A (326 000 c p m / ~ g ) were i n c u b a t e d wi th 19 pg of g l y c e r a l d e h y d e - 3 - p h o s p h a t e d e h y d r o g e n a s e . Expt . 2 : 1 pg of d e n a t u r e d NIL-1 -HaSV [ 3 H ] D N A (80 898 cpm/Mg) was i n c u b a t e d wi th 9 ~g of e n z y m e . in b o t h e x p e r i m e n t s the NaC1 c o n c e n t r a t i o n was 80 mM an d the d inuc leos ide m o n o p h o s p h a t e s were added to final c o n c e n t r a t i o n of 1.5 raM. The e n z y m e was a lways a d d e d last . Af t e r i n c u b a t i o n for 4 rain at 30°C, the samples were f i l tered t h r o u g h Millipore m e m b r a n e s as usual. Wi thou t addi t ions , 71 400 c p m and 15 067 c p m of [ 3 H ] D N A were b o u n d in Expts . 1 and 2, respec t ive ly .
Add i t ions Relat ive a m o u n t of [ 3 H ] D N A re ta ined (%)
Expt . 1 None 100 UpA 23 CpA 30 ApA 56 GpA 55 G p G 103 UpG 124 UpC 92 GpC 115
Expt . 2 None 100 G p A 41 ApG 86 A p U 85
lO0
5 0
f t e
_ _ _ c
° L
lO 20 30
LACTATE DEHYDROGENASE (~Jg)
40
Fig. 7. Binding of l ac ta te d e h y d r o g e n a s e to DNA. Increas ing a m o u n t s of r abb i t musc le lac ta te d e h y d r o - genase were i n c u b a t e d wi th 0.5 ~g of d e n a t u r e d or na t ive NIL-1 -HaSV [ 1 4 C ] D N A (18 0 0 0 c p m / p g ) for 4 rain a t 30°C and f i l tered t h r o u g h Millipore m e m b r a n e s as ind ica ted be fo re [1 ] . The NaC1 c o n c e n t r a t i o n
was 80 raM. • • d e n a t u r e d D N A ; o ©, na t ive DNA.
193
~Z
== ~z 40
8O
_ _ i i OD5 0.1
NAO
N A D H
.o
I i I
0.15 0 . 2 0 . 2 5
N A D or N A D H ( raM)
// i
1
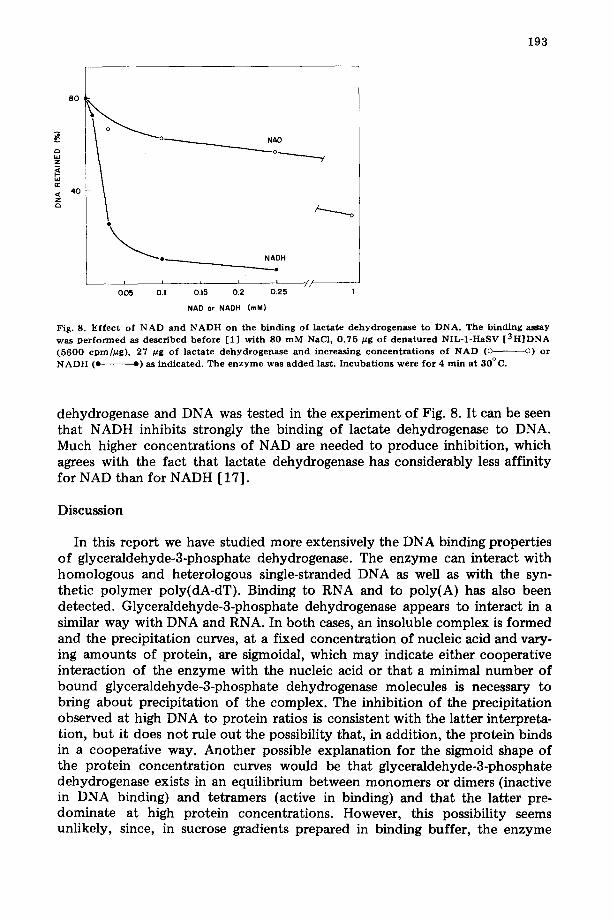
Fig. 8. E f f ec t of N A D and N A D H on the b ind ing of l ac ta te d e h y d r o g e n a s e t o D N A . T h e b i n d i n g a s s a y
was p e r f o r m e d as descr ibed be fo re [1] wi th S0 m M NaC1, 0 .75 #g of d e n a t u r e d NIL-1 -HaSV [ 3 H ] D N A (5600 cpm//~g), 27 btg of l ac ta te d e h y d r o g e n a s e a nd i n c r e a s i n g c o n c e n t r a t i o n s of N A D (o o) or N A D H (e e) as ind ica ted . The e n z y m e was a d d e d last. I n c u b a t i o n s w e r e f o r 4 rain at 30°C.
dehydrogenase and DNA was tested in the experiment of Fig. 8. It can be seen that NADH inhibits strongly the binding of lactate dehydrogenase to DNA. Much higher concentrations of NAD are needed to produce inhibition, which agrees with the fact that lactate dehydrogenase has considerably less affinity for NAD than for NADH [17].
Discussion
In this report we have studied more extensively the DNA binding properties of glyceraldehyde-3-phosphate dehydrogenase. The enzyme can interact with homologous and heterologous single-stranded DNA as well as with the syn- thetic polymer poly(dA-dT). Binding to RNA and to poly(A) has also been detected. Glyceraldehyde-3-phosphate dehydrogenase appears to interact in a similar way with DNA and RNA. In both cases, an insoluble complex is formed and the precipitation curves, at a fixed concentration of nucleic acid and vary- ing amounts of protein, are sigmoidal, which may indicate either cooperative interaction of the enzyme with the nucleic acid or that a minimal number of bound glyceraldehyde-3-phosphate dehydrogenase molecules is necessary to bring about precipitation of the complex. The inhibition of the precipitation observed at high DNA to protein ratios is consistent with the latter interpreta- tion, but it does not rule out the possibility that, in addition, the protein binds in a cooperative way. Another possible explanation for the sigmoid shape of the protein concentration curves would be that glyceraldehyde-3-phosphate dehydrogenase exists in an equilibrium between monomers or dimers (inactive in DNA binding) and tetramers (active in binding) and that the latter pre- dominate at high protein concentrations. However, this possibility seems unlikely, since, in sucrose gradients prepared in binding buffer, the enzyme

194
sediments as a tetramer even at very low protein concentrations (about 10 pg/ ml) (data not shown).
Other studies, not presented here, show that glyceraldehyde-3-phosphate dehydrogenase protects DNA from attack by pancreatic DNAase and prevents the action of eukaryotic or prokaryotic RNA polymerase.
The inhibition of DNA binding by NAD and several adenine nucleotides, but not by glyceraldehyde-3-phosphate and inorganic phosphate, suggest that DNA interacts with glyceraldehyde-3-phosphate dehydrogenase through the NAD binding sites of the enzyme. Both electrostatic and hydrophobic bonds appear to be involved in NAD binding to glyceraldehyde-3-phosphate dehydrogenase [18]. The effect of ionic strength and divalent cations suggests a role for elec- trostatic bonds in the interaction of glyceraldehyde-3-phosphate dehydrogenase with DNA. On the other hand, the fact that the temperature favors this interac- tion and the existence at 30°C of a second optimal at a higher salt concentra- tion suggest that hydrophobic bonds are also involved in the formation of the enzyme-DNA complex. These results are consistent with the suggestion that DNA binds through the dinucleotide fold of glyceraldehyde-3-phosphate dehydrogenase.
In keeping with this view it is shown here that DNA can also bind to lactate dehydrogenase, an enzyme which contains a dinucleotide fold similar to that of glyceraldehyde-3-phosphate dehydrogenase, and that this binding is inhibited by NADH.
The role of the base in the binding interaction was studied by comparing the ability of several dinucleoside monophosphates to inhibit DNA binding to glyceraldehyde-3-phosphate dehydrogenase. The results obtained (see Table IV) are consistent with the idea that these compounds bind at the NAD site of the dehydrogenase and suggest that the enzyme would interact preferentially with PypA dinucleotides present in the nucleic acid.
The characteristics of the interaction between nucleic acids and glyceralde- hyde-3-phosphate dehydrogenase or lactate dehydrogenase may provide a use- ful model for the behavior of biologically significant nucleic acid binding pro- teins. In connect ion with this, it is interesting to mention that, as pointed out by Wyckoff et al. [19] and Wodak et ah [20], the complexes of dinucleoside monophosphates with pancreatic ribonuclease S have several features in common with those of NAD with glyceraldehyde-3-phosphate dehydrogenase and other dehydrogenases [2].
Finally, it should be noted that the dinucleoside monophosphate compounds could be useful to further explore the properties of the coenzyme binding domain of dehydrogenases.
Acknowledgements
We thank Professors E. Vifiuela and M. Salas for the critical reading of the manuscript. This investigation has been aided by grants from the Comisi6n Asesora para el Desarrollo de la Investigaci6n Cientffica y T6cnica and Comi- si6n Administradora del Descuento Complementario (I.N.P.).

195
References
1 Perucho, M., Salas, J. and Salas, M.L. (1977) Eur. J. Biochem. 81, 557--562 2 Rossman, M.G., IAljas, A., Br~nden, C.I. and Bansrzak, L.J. (1975) in The Enzymes (Boyer, P,D., ed.),
Vol. 11, pp. 61--102, Academic Press, New York 3 Warner, J.R., Soeiro, R., Birnboim, H.C., Girard, M. and Darnell, J.E. (1966) J. Mol. Biol. 19, 349--
361 4 Edmonds, M. and Grace Caramela, M. (1969) J. Biol. Chem. 244, 1314--1324 5 Melero, J.A., Salas, M.L., Salas, J. and Macpherson, I.A. (1975) J. Biol. Chem. 250, 3683--3689 6 Velick, S.F. (1955) Methods Enzymol. 1 ,401- -406 7 Constantinides, S.M. and Deal, W.C., Jr. (1970) J. Biol. Chem. 245, 246--253 8 Nagradova, N.K., Muronetz, V.I., Grozdova, I.D. and Golovina, T.O. (1975) Biochim. Biophys. Acta
377, 15--25 9 Harris, J.I. and Perham, R.N. (1965) J. Mol. Biol. 13, 876--884
10 Harrington, W.F. and Karr, G.M. (1965) J. Mol. Biol. 13, 885--893 11 Riggs, A.D., Suzuke, H. and Bourgeois, S. (1970) J. Mol. Biol. 48, 67--83 12 Hinkle, D.C. and Chamberlin, M.J. (1972) J. Mol. Biol. 70, 157--185 13 Colowick, S.P., van Eeys, J. and Park, J.H. (1966) in Comprehensive Biochemistry (Florkin, M. and
Stotz, E.H., eds.), Vol. 14, Pp. 1---98, Elsevier, Amsterdam 14 Yang, S.T. and Deal, W.C., Jr. (1969) Biochemistry 8, 2806--2813 15 Oguehi, M., Meriwether, B.P. and Park, J.H. (1973) J. Biol. Chem. 248, 5562--5570 16 Constantinides, S.M. and Deal, W.C., Jr. (1969) J. Biol. Chem. 244, 5695--5702 17 Holbrook, J.J., Liljas, A., Steindel, S.J. and Rossmann, M.G. (1975) in The Enzymes (Boyer, P.D.,
ed.), Vol. 11, pp. 191--292, Academic Press, New York 18 Harris, J.I. and Waters, M. (1976) in The Enzymes (Boyer, P.D., ed.), Vol. 13, pp. 1--49, Academic
Press, New York 19 Wyekoff, H.W., Carlson, W. and Wodak, S.J. (1977) in Nucleic Acid-Protein Recognit ion (Vogel, H.J.,
ed.), pp. 569--580, Academic Press, New York 20 Wodak, S.Y., Liu, M.Y. and Wyckoff, H.W. (1977) J. Mol. Biol. 116, 855--875




















