
Involvement of Lysophosphatidic Acid, Sphingosine 1-Phosphate and Ceramide 1-Phosphate in the...
11
ORIGINAL PAPER Involvement of Lysophosphatidic Acid, Sphingosine 1-Phosphate and Ceramide 1-Phosphate in the Metabolization of Phosphatidic Acid by Lipid Phosphate Phosphatases in Bovine Rod Outer Segments Susana J. Pasquare ´ Gabriela A. Salvador Norma Maria Giusto Accepted: 12 December 2008 / Published online: 21 February 2008 Ó Springer Science+Business Media, LLC 2008 Abstract The aim of the present research was to evaluate the generation of [2- 3 H]diacylglycerol ([2- 3 H]DAG) from [2- 3 H]-Phosphatidic acid ([2- 3 H]PA) by lipid phosphate phosphatases (LPPs) at different concentrations of lyso- phosphatidic acid (LPA), sphingosine 1-phosphate (S1P), and ceramide 1-phosphate (C1P) in purified ROS obtained from dark-adapted retinas (DROS) or light-adapted retinas (BLROS) as well as in ROS membrane preparations depleted of soluble and peripheral proteins. Western blot analysis revealed the presence of LPP3 exclusively in all membrane preparations. Immunoblots of entire ROS and depleted ROS did not show dark–light differences in LPP3 levels. LPPs activities were diminished by 53% in BLROS with respect to DROS. The major competitive effect on PA hydrolysis was exerted by LPA and S1P in DROS and by C1P in BLROS. LPPs activities in depleted ROS were similar to the activity observed in entire DROS and BLROS, respectively. LPA, S1P and C1P competed at different extent in depleted DROS and BLROS. Sphingo- sine and ceramide inhibited LPPs activities in entire and depleted DROS. Ceramide also inhibited LPPs activities in entire and in depleted BLROS. Our findings are indicative of a different degree of competition between PA and LPA, S1P and C1P by LPPs depending on the illumination state of the retina. Keywords Rod outer segments Lipid phosphate phosphatases Phosphatidic acid Diacylglycerol Abbreviations C1P Ceramide 1-phosphate DAG Diacylglycerol DTT Dithiotreitol EDTA Ethylenediaminetetraacetic acid EGTA Ethylene glycol bis (b-aminoethyl ether)- N,N,N 0 ,N 0 -tetra acetic acid LPA Lysophosphatidic acid LPPs Lipid phosphate phosphatases NEM N-ethylmaleimide PA Phosphatidic acid PAP2 NEM-insensitive phosphatidate phosphohidrolase PC Phosphatidylcholine PMSF Phenylmethylsulfonylfluoride S1P Sphingosine 1-phosphate TLC Thin layer chromatography Introduction Lipid phosphate monoesters, including phosphatidic acid (PA), lysophosphatidic acid (LPA), sphingosine 1-phos- phate (S1P) and ceramide 1-phosphate (C1P), are intermediaries in phospho- and sphyngolipid biosynthesis and they also play important roles in intra- and extracel- lular signaling. Dephosphorylation of these lipids terminates their signaling actions and generates products with additional biological activities or metabolic fates [1]. The key enzymes responsible for the dephosphorylation of these lipid phosphate substrates are termed lipid phosphate phosphatases (LPPs). The latter display isoforms and cell specific localization patterns which are distributed between endomembrane compartments and the plasma membrane. The role of LPPs in intracellular lipid metabolism and in the regulation of both intra- and extracellular signaling S. J. Pasquare ´ G. A. Salvador N. M. Giusto (&) Instituto de Investigaciones Bioquı ´micas de Bahı ´a Blanca, Universidad Nacional del Sur and Consejo Nacional de Investigaciones Cientı ´ficas y Te ´cnicas (CONICET), C.C. 857, B8000FWB Bahia Blanca, Buenos Aires, Argentina e-mail: [email protected] 123 Neurochem Res (2008) 33:1205–1215 DOI 10.1007/s11064-007-9569-5
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Involvement of Lysophosphatidic Acid, Sphingosine 1-Phosphate and Ceramide 1-Phosphate in the...

ORIGINAL PAPER
Involvement of Lysophosphatidic Acid, Sphingosine 1-Phosphateand Ceramide 1-Phosphate in the Metabolization of PhosphatidicAcid by Lipid Phosphate Phosphatases in Bovine Rod OuterSegments
Susana J. Pasquare Æ Gabriela A. Salvador ÆNorma Maria Giusto
Accepted: 12 December 2008 / Published online: 21 February 2008
� Springer Science+Business Media, LLC 2008
Abstract The aim of the present research was to evaluate
the generation of [2-3H]diacylglycerol ([2-3H]DAG) from
[2-3H]-Phosphatidic acid ([2-3H]PA) by lipid phosphate
phosphatases (LPPs) at different concentrations of lyso-
phosphatidic acid (LPA), sphingosine 1-phosphate (S1P),
and ceramide 1-phosphate (C1P) in purified ROS obtained
from dark-adapted retinas (DROS) or light-adapted retinas
(BLROS) as well as in ROS membrane preparations
depleted of soluble and peripheral proteins. Western blot
analysis revealed the presence of LPP3 exclusively in all
membrane preparations. Immunoblots of entire ROS and
depleted ROS did not show dark–light differences in LPP3
levels. LPPs activities were diminished by 53% in BLROS
with respect to DROS. The major competitive effect on PA
hydrolysis was exerted by LPA and S1P in DROS and by
C1P in BLROS. LPPs activities in depleted ROS were
similar to the activity observed in entire DROS and
BLROS, respectively. LPA, S1P and C1P competed at
different extent in depleted DROS and BLROS. Sphingo-
sine and ceramide inhibited LPPs activities in entire and
depleted DROS. Ceramide also inhibited LPPs activities in
entire and in depleted BLROS. Our findings are indicative
of a different degree of competition between PA and LPA,
S1P and C1P by LPPs depending on the illumination state
of the retina.
Keywords Rod outer segments � Lipid phosphate
phosphatases � Phosphatidic acid � Diacylglycerol
Abbreviations
C1P Ceramide 1-phosphate
DAG Diacylglycerol
DTT Dithiotreitol
EDTA Ethylenediaminetetraacetic acid
EGTA Ethylene glycol bis (b-aminoethyl ether)-
N,N,N0,N0-tetra acetic acid
LPA Lysophosphatidic acid
LPPs Lipid phosphate phosphatases
NEM N-ethylmaleimide
PA Phosphatidic acid
PAP2 NEM-insensitive phosphatidate phosphohidrolase
PC Phosphatidylcholine
PMSF Phenylmethylsulfonylfluoride
S1P Sphingosine 1-phosphate
TLC Thin layer chromatography
Introduction
Lipid phosphate monoesters, including phosphatidic acid
(PA), lysophosphatidic acid (LPA), sphingosine 1-phos-
phate (S1P) and ceramide 1-phosphate (C1P), are
intermediaries in phospho- and sphyngolipid biosynthesis
and they also play important roles in intra- and extracel-
lular signaling. Dephosphorylation of these lipids
terminates their signaling actions and generates products
with additional biological activities or metabolic fates [1].
The key enzymes responsible for the dephosphorylation of
these lipid phosphate substrates are termed lipid phosphate
phosphatases (LPPs). The latter display isoforms and cell
specific localization patterns which are distributed between
endomembrane compartments and the plasma membrane.
The role of LPPs in intracellular lipid metabolism and in
the regulation of both intra- and extracellular signaling
S. J. Pasquare � G. A. Salvador � N. M. Giusto (&)
Instituto de Investigaciones Bioquımicas de Bahıa Blanca,
Universidad Nacional del Sur and Consejo Nacional de
Investigaciones Cientıficas y Tecnicas (CONICET), C.C. 857,
B8000FWB Bahia Blanca, Buenos Aires, Argentina
e-mail: [email protected]
123
Neurochem Res (2008) 33:1205–1215
DOI 10.1007/s11064-007-9569-5

pathways that control different cellular functions have been
described [2–5].
Rod outer segments (ROS) have the ability to adapt the
sensitivity and speed of their responses to ever changing
conditions of ambient illumination. Recent evidence has
demonstrated that a major contributor to this adaptation is
the light-driven translocation of key signaling proteins into
and out of ROS, which constitute the cellular place where
phototransduction occurs [6]. It has also been reported that
transducin, arrestin and recoverin [7–10] are proteins
involved in this mechanism. Previous studies revealed the
presence of phosphatidate phosphohydrolase 2 (PAP2,
renamed as LPPs) and its regulation in isolated ROS from
bovine retina [11–14]. In this work, the generation of
[2-3H]DAG from [2-3H]PA was analyzed either in the
absence or in the presence of LPA, S1P or C1P, all of
which are alternative substrates for LPPs. The presence of
LPP3 isoform in isolated ROS was also determined by
western blot.
It has been extensively reported that the activity of enzymes
involved in ROS phospholipid turnover such as phospholipase
C [15, 16], phospholipase A2 [17], phosphatidylethanolamine
N-methyltransferase [18], diacylglycerol kinase [19], PAP2
[14], phosphoinositide-3-kinase [20, 21], and phospholipase
D [22] is modulated by light. This fact and the evidence that
light induces a redistribution of proteins involved in photo-
transduction open an attractive field for the study of PA
hydrolysis by LPPs under illumination conditions. We have
also characterized LPP activities in the presence of [2-3H]PA
co-incubated with LPA, or S1P or C1P in ROS prepared from
dark-adapted retinas (DROS) as well as in ROS prepared
from light-adapted retinas (BLROS). Our results provide
evidence that PA is differently hydrolyzed by LPPs in the
presence of LPA, S1P, and C1P. Light participation in the
competition of LPA, S1P and C1P on PA metabolization is
also demonstrated.
Experimental Procedure
Materials
[2-3H] Glycerol (200 mCi/mmol) was obtained from New
England Nuclear-Dupont, Boston, MA. Sphingosine 1-
phosphate, ceramide 1-phosphate from bovine brain, oleoyl-
L-a-lysophosphatidic acid, D-sphingosine, and non-hydroxy
fatty acid ceramide from bovine brain were obtained from
Sigma-Aldrich, St. Louis, MO, USA. Monoclonal antibody
against rhodopsin, Rho4D2, was generously supplied by
Robert Molday from the University of British Columbia,
Vancouver, Canada. The secondary antibody used by the
rhodopsin immunoblots was HRP-conjugated anti-mouse.
Anti-PAP2B (anti-LPP3) was from Upsate biotechnology
(catalog number 07-142). The secondary antibody used for
anti-PAP2b detection was HRP-conjugated anti-rabbit. All
the other chemicals were of the highest purity available.
Dark and Light Adaptation Procedure
Bovine eyes were obtained from a local abattoir, placed on
ice within 10 min of the animal’s death, and they were kept
in darkness for 2 h. Retinas were dissected from the eyes
after dark- or light adaptation. Dark-adapted bovine ROS
(DROS) and light-adapted bovine ROS (LROS) were pre-
pared under dim red light from DROS. The bleaching
process was carried out after removing the cornea and the
lens from the eyeballs and by exposing them to light
(300 W) at 30 cm for 30 min at room temperature [20].
Controls were performed under the same conditions but
under darkness (DROS).
Rod Outer Segment Isolation
The subsequent procedures for ROS preparation were
conducted under dim red light for DROS and LROS, and
under room light for BLROS and they were carried out at
2–4�C. To isolate ROS, retinas were removed and shaken
twice in a 40% sucrose solution containing 1 mM MgCl2,
1 mM DTT, 0.1 mM PMSF, 1 lg/ml aprotinin, and 2 lg/
ml leupeptin in 70 mM sodium phosphate buffer (pH 7.2).
The remains of retinas were sedimented at 2,200g for
4 min and the supernatants containing ROS were diluted
1:2 with sucrose-free buffer and then centrifuged during
30 min at 35,300g. ROS were purified by a discontinuous
gradient of sucrose [23], yielding: (i) a ROS band (Band I)
retained at the 0.84/1.00 M density interface; (ii) a band
(Band II) retained at the 1.00/1.14 M density interface of
broken ROS contaminated with mitochondria and RIS, and
(iii) a pellet composed of non-ROS membranes. The purity
of ROS membrane preparations was monitored by electron
microscopy. Electron micrographs from purified ROS
(Band I) showed intact ROS with their typical structures
and no other membrane material was observed (data not
shown). The purity of Band I was also controlled by the
determination of the absorbance ratio at 278 and 500 nm of
solubilized membranes in 70 mM potassium phosphate
buffer (pH 7) containing 1% emulphogene. Values of
2.3 ± 0.2 were typically obtained for this ratio. In addition,
the purity of membranes was checked by sodium dodecyl
sulfate–polyacrylamide gel electrophoresis [24]. Even in
overloaded gels (80 lg of ROS protein), rhodopsin com-
prised 85–90% of photoreceptor integral membrane
proteins. Moreover, thin-layer chromatography of
1206 Neurochem Res (2008) 33:1205–1215
123

photoreceptor membrane lipids in overloaded plates
showed no cardiolipin, thus suggesting non-detectable
contamination with mitochondria. Instead, an enrichment
of long-chain polyunsaturated fatty acids esterified to di-
polyunsaturated molecular species of PC was observed.
This enrichment, which is characteristic of bovine ROS
[25]. Purity of ROS populations, particularly batches, was
determined by assaying marker enzymes activities in all
fractions of the gradients. NADPH-cytochrome c-reductase
(microsomal marker) and cytochrome c-oxidase (mito-
chondrial marker) activities were measured in Band I, Band
II and pellet of the gradient. Cytochrome c-oxidase was
more enriched in Band II and pellet whereas the micro-
somal marker activity was very low in Band I and
evidenced the highest activity in the pellet. These results
led us to confirm that Band I (purified ROS) contamination
with microsomes or mitochondria was lower than 5% [26,
27].
Soluble and Peripheral Protein Extraction from ROS
with Low Ionic Strength Buffer
ROS pellets from DROS or BLROS were resuspended (1 mg
protein per ml) in low ionic strength buffer prepared with
5 mM Tris–HCl (pH 7.4), containing 0.5 mM MgCl2, 1 mM
DTT, 0.1 mM PMSF, 1 lg/ml aprotinin, 2 lg/ml leupeptin
and 1 lg/ml of pepstatin, followed by 15 passages through a
G25 5/8 needle. The membrane suspensions were centri-
fuged at 35,300g for 30 min. The supernatants were removed
and centrifuged again in order to ensure that all particulate
material was sedimented and membrane pellets were
extensively washed. Both membranes and the clear super-
natants were analyzed to determine the polypeptide
composition, the presence of LPP3 and the activity of LPPs.
Preparation of Radioactive 1,2-diacyl-sn-glycerol-3-
phosphate
Radioactive phosphatidic acid was obtained by the action
of phospholipase D on [2-3H]-phosphatidylcholine [28].
[2-3H]-phosphatidylcholine was synthesized from bovine
retinas incubated with [2-3H] glycerol (200 mCi/mmol) as
previously described [11]. Lipids were extracted from the
tissue following Folch et al. procedure [29]. [2-3H]-Phos-
phatidylcholine was isolated by mono-dimensional TLC
and eluted there from [30]. [2-3H]PA, the hydrolysis
product of [2-3H]-PC, was purified by one-dimensional
TLC on silica gel H developed with chloroform/methanol/
acetic acid/acetone/water (9:3:3:12:1.5, v/v). The substrate
was eluted from silica gel with neutral solvents to avoid the
formation of lysophosphatidic acid. It was subsequently
converted into free acid by washing it twice using an upper
phase containing 0.1 M sulfuric acid and then an upper
phase containing water. Radioactivity and phosphorous
content [31] were measured to determine specific radio-
activity. [2-3H]PA with a specific radioactivity of 0.1–
0.2 lCi/lmol was obtained.
Determination of LPPs Activities
For the determination of LPPs activities the assay contained
50 mM Tris–maleate buffer, pH 6.5, 1 mM DTT, 1 mM
EDTA plus 1 mM ethylene glycol-bis(b-aminoethylether
N,N,N0,N-tetraacetic acid (EGTA), 4.2 mM NEM and
100 lg of ROS membrane proteins in a volume of 200 ll.
The reaction was started by the addition of 100 lM of
[2-3H]-phosphatidate/Triton X-100 mixed micelles in a
constant 1:50 molar ratio of lipid to Triton X-100 [32].
[2-3H]-phosphatidate was dried under a stream of nitrogen
and resuspended in 30 ll of buffer assay containing Triton
X-100, this aqueous microdispersion was sonicated in a
sonication tip until clarity. The effect of the alternative
substrates on PA metabolization by LPP activities, was
evaluated using 100 lM [2-3H]PA/Triton X-100 mixed
micelles in the presence of LPA, S1P or C1P (previously
resuspended in the buffer assay containing Triton X-100) at
the indicated concentrations [32, 33]. Sphingosine and cer-
amide were solubilized in 0.1% of dimethyl sulfoxide
(DMSO) as vehicle; the results obtained were compared
against controls performed with the vehicle. The assays for
the determination of LPPs were conducted at 37�C for
30 min under dim red light (DROS), under room light
(LROS) or under 300 W light (BLROS). The enzymatic
assay was stopped by adding chloroform/methanol (2:1, v/
v). Blanks were prepared identically, except that membranes
were boiled for 5 min before being used. PA products gen-
erated by LPP activities were isolated and measured as
described below. The enzymatic activity was expressed as
the sum of nmol of [2-3H] diacylglycerol and [2-3H] mon-
oacylglycerol 9 (h mg protein)-1.
Extraction and Isolation of Lipids
Lipids were extracted with chloroform/methanol (2:1, v/v)
and washed with 0.2 volumes of CaCl2 (0.05%) [32]. Neutral
lipids were separated by gradient-thickness thin-layer chro-
matography on silica gel G [34] and developed with hexane/
diethyl ether/acetic acid (35:65:1, v/v). To separate mono-
acylglycerol (MAG) from phospholipids, the chromatogram
was rechromatographed up to the middle of the plate by using
hexane/diethyl ether/acetic acid (20:80:2.3, v/v) as devel-
oping solvent. Once the chromatogram was developed,
Neurochem Res (2008) 33:1205–1215 1207
123

[2-3H] PA and phospholipids were retained at the spotting
site. Lipids were visualized by exposure of the chromato-
grams to iodine vapors, scraped off the plate, and quantified
by liquid scintillation spectroscopy.
SDS–PAGE and Immunoblot Analysis
SDS–PAGE was performed using 7.5% or 10% gels
according to Laemmli [24]. Resolved proteins were trans-
ferred to immobilon P membranes using a Mini Trans-Blot
cell electro blotter (BIO-RAD Life Science Group, Cali-
fornia) for 1 h. Membranes were blocked overnight with
Tris-buffered saline (20 mM Tris–HCl, 300 mM NaCl) pH
7.5, containing 0.1% Tween 20 and 5% crystalline grade
bovine serum albumin (BSA). Incubations with primary
antiserum were performed at room temperature for 2–3 h.
Immunoreactions were detected by means of either horse-
radish peroxidase conjugated to goat antirabbit or goat anti-
mouse IgG followed by enhanced chemiluminescence
substrates (ECL; Amersham Biosciences, Inc.). In some
experiments, immunoblots were stripped by incubation in
200 mM Tris–HCl buffer, pH 6.7, containing 100 mM b-
mercaptoethanol, 2% sodium dodecyl sulfate (SDS) for 1 h
at 50�C with gentle agitation. Blots were reblocked with
5% BSA in Tris-buffered saline and probed as described
above. Immunoreactive bands were quantified using image
analysis software (Image J, a freely available application in
the public domain for image analysis and processing,
developed and maintained by Wayne Rasband at the
Research Services Branch, National Institute of Mental
Health, Bethesda, Maryland, USA).
Other Methods
Lipid phosphorus and protein were determined as described
elsewhere [31, 35].
Statistical Analysis
Statistical analysis was done using Student’s t-test with the
values representing the mean ± SD of the total number of
samples indicated in each figure legend.
Results
Hydrolysis of [2-3H]-phosphatidate by LPPs in DROS
and BLROS as a Function of LPA, S1P and C1P
Concentrations
The DAG formed by LPP activities is partially hydrolyzed
by diacylglycerol lipase (DAGL), yielding MAG. DAGL is
coupled to LPPs and it seems that these enzyme activities
are working in a sort of enzyme complex. Taking this into
account the DAG generation and its partial degradation by
DAGL immediately occurs. This fact has been extensively
described in our laboratory [11, 12]. For this reason the
most accurate from to express LPP activity is as the sum of
nmol of [2-3H] diacylglycerol and [2-3H] monoacylglyc-
erol 9 (h mg protein)-1.
Data presented in Fig. 1 show the rate of DAG forma-
tion from [2-3H]-phosphatidate (PA) in the presence of the
alternative substrates using two ROS populations: DROS
(square symbols) and BLROS (circle symbols). When the
light effect was evaluated on LPP activities using only
[2-3H]PA as substrate, an inhibition of 53% was observed
in BLROS with respect to DROS (Fig. 1). LPA and S1P
significantly decreased DAG (P \ 0.025) production from
[2-3H]PA in a concentration-dependent manner in DROS
(Fig. 1, square symbols). LPA diminished DAG formation
by 15% at concentrations ranging from 10 to 50 lM and by
29% at the highest concentration of LPA assayed (200 lM)
(Fig. 1A). MAG generation from LPA could also be cat-
alyzed by LPA phosphohydrolase. The presence of this
enzyme activity has not been described in isolated ROS.
Moreover, in our experiments with non-labeled LPA it is
not possible to evaluate the presence of this enzyme.
An important diminution in DAG production was
observed at low concentrations of S1P reaching 27% at
50 lM of S1P (Fig. 1B). C1P exerted an inhibitory effect
(30%) on DAG formation at concentrations higher than
100 lM (Fig. 1C).
In order to determine the effect of light (Fig. 1, circle
symbols) in the competition of LPA, S1P and C1P in PA
hydrolysis, LPP activities were also evaluated in BLROS in
the presence of increasing concentrations of the alternative
substrates for the enzyme. In BLROS a 22% decrease in
DAG formation was observed at 100 lM of LPA (Fig. 1
A). In BLROS PA dephosphorylation was diminished by
19% at 20 lM of S1P whereas at 100 lM of S1P, DAG
levels returned to basal levels (Fig. 1B). On the other hand,
C1P diminished DAG production at all the concentrations
assayed, being highest (41%) at 200 lM (Fig. 1C).
Hydrolysis of [2-3H]-phosphatidate by LPPs in LROS
and BLROS in the Presence of LPA, S1P and C1P
For the experiments aimed at the description of LPP
activities under different illumination conditions we used
three distinct ROS populations: (i) DROS: obtained from
dark-adapted retinas and purified under dim red light, (ii)
LROS: obtained from DROS and exposed to room light for
the enzyme assays, (iii) BLROS: obtained from light-
adapted retinas and purified under room light.
1208 Neurochem Res (2008) 33:1205–1215
123

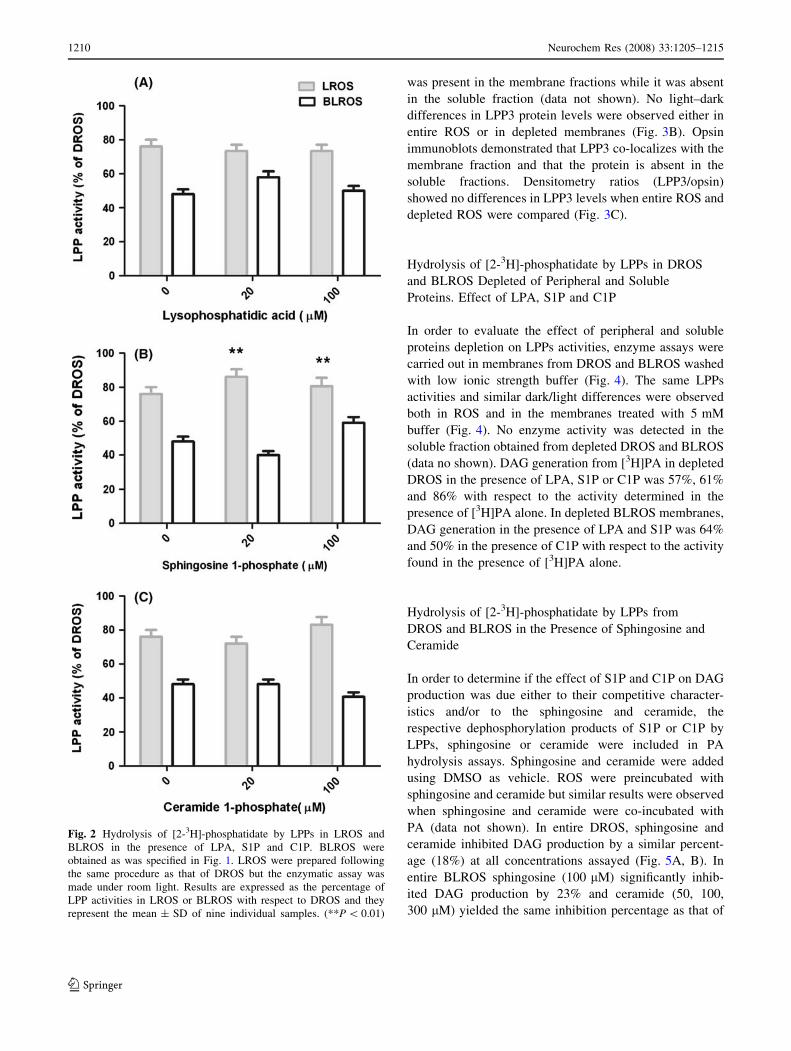
Figure 2 shows the percentage of LPPs activities in LROS
and BLROS with respect to the activities determined in
DROS (considered as 100%) in the absence or in the pres-
ence of 20 or 100 lM of LPA, S1P or C1P. LROS were
obtained, exposing DROS to room light. Under these
experimental conditions an inhibition of 24% in LPPs
activities with respect to DROS was observed (Fig. 2). LPA
and C1P did not modify LPPs activities from LROS
(Fig. 2A, C), whereas in the presence of S1P, LPPs activities
were stimulated by 13% (Fig. 2B). A differential effect of
alternative substrates in PA hydrolysis pattern was observed
between LROS and BLROS. In all cases, LPPs activities
were higher in LROS than in BLROS. Based on the results
shown in Fig. 2 a ratio between the percentages of LPPs
activities from LROS and BLROS was determined. This
ratio was 1.6 in the presence of PA as unique substrate, 1.3 in
the presence of LPA (20, 100 lM), 2.2 and 1.3 in the pres-
ence of either 20 or 100 lM of S1P, respectively, and it was 1
in the presence of 100 lM of C1P.
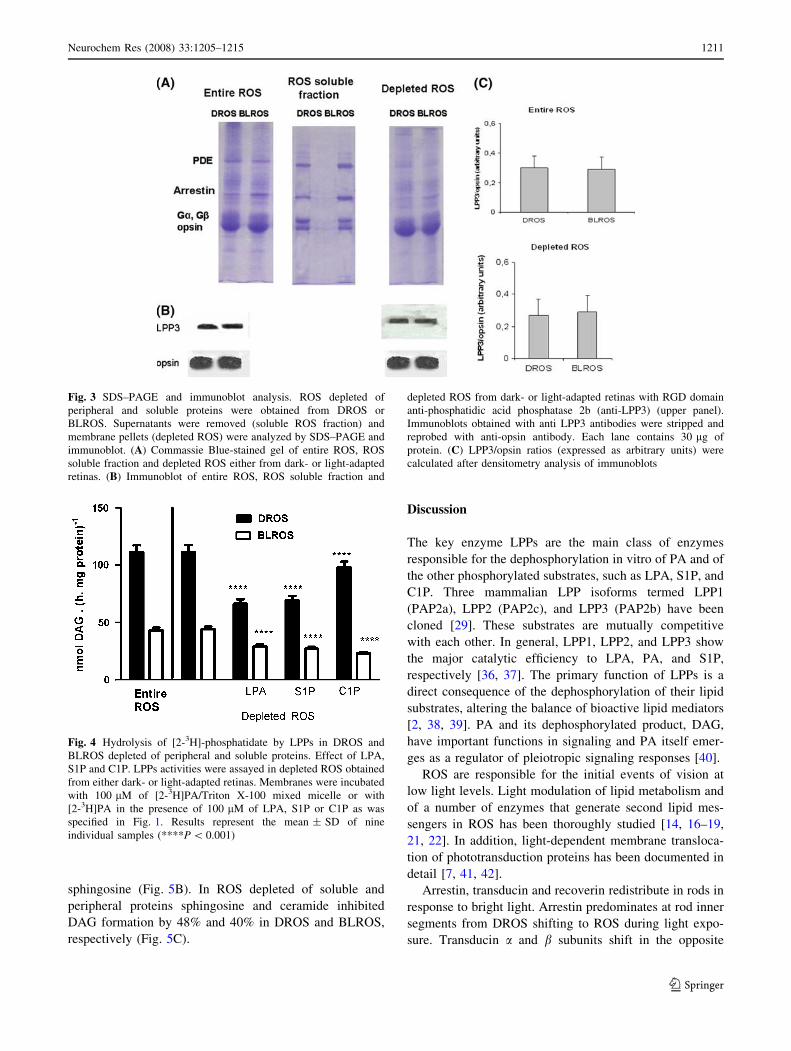
SDS–PAGE and Immunoblot Analysis
We determined the protein composition of entire DROS
and BLROS preparation and of DROS and BLROS subject
to successive washes with low ionic strength buffers. Low
ionic buffer treatment renders a ROS membrane prepara-
tion depleted of soluble and peripheral proteins. Both the
membrane and soluble fractions obtained there from were
used for SDS–PAGE analysis. The levels of arrestin were
higher in entire BLROS than in DROS (Fig. 3A). The
major polypeptides present in the ROS soluble fraction
(supernatant obtained after washing ROS preparations with
low ionic buffer) were a and b subunits of transducin (Ga,
Gb), arrestin and phosphodiesterase (PDE) (Fig. 3A). The
effect of GTP concentrations, and the rate of GTP hydro-
lysis participates in Gt translocation [6]. In our
experimental setup it seems that the initial concentration of
GTP at the moment of ROS isolation is enough for
allowing a significant light-mediated translocation of Gt.
Immunoblot analysis using RGD domain anti-phosphatidic
acid phosphatase 2b (anti-LPP3) revealed that this isoform
Fig. 1 Hydrolysis of [2-3H]-phosphatidate (PA) by LPPs in DROS
and BLROS as a function of lysophosphatidic acid (LPA), sphingo-
sine 1-phosphate (S1P) and ceramide 1-phosphate (C1P)
concentrations. LPPs activities were determined using, as an enzyme
source, either purified ROS from dark-adapted retinas (DROS) or
bleached ROS (BLROS) from DROS whose eye cups were exposed
to room light (300 W at 30 cm) for 30 min as specified in
Experimental Procedure. The effect of the alternative substrates on
PA hydrolysis by LPP activities, was evaluated using 100 lM
[2-3H]PA/Triton X-100 mixed micelles in the presence of LPA (A),
S1P (B) or C1P (C) at the indicated concentrations. The enzymatic
assay was made under dim red light for DROS or under 300 W light
for BLROS. Incubation products were subsequently extracted and
separated by gradient-thickness thin-layer chromatography and visu-
alized after exposure to iodine vapor. The bands corresponding to PA,
DAG and MAG were scraped and quantitated by liquid scintillation
spectroscopy. Results represent the mean ± SD of nine individual
samples. (*P \ 0.025)
b
Neurochem Res (2008) 33:1205–1215 1209
123
was present in the membrane fractions while it was absent
in the soluble fraction (data not shown). No light–dark
differences in LPP3 protein levels were observed either in
entire ROS or in depleted membranes (Fig. 3B). Opsin
immunoblots demonstrated that LPP3 co-localizes with the
membrane fraction and that the protein is absent in the
soluble fractions. Densitometry ratios (LPP3/opsin)
showed no differences in LPP3 levels when entire ROS and
depleted ROS were compared (Fig. 3C).
Hydrolysis of [2-3H]-phosphatidate by LPPs in DROS
and BLROS Depleted of Peripheral and Soluble
Proteins. Effect of LPA, S1P and C1P
In order to evaluate the effect of peripheral and soluble
proteins depletion on LPPs activities, enzyme assays were
carried out in membranes from DROS and BLROS washed
with low ionic strength buffer (Fig. 4). The same LPPs
activities and similar dark/light differences were observed
both in ROS and in the membranes treated with 5 mM
buffer (Fig. 4). No enzyme activity was detected in the
soluble fraction obtained from depleted DROS and BLROS
(data no shown). DAG generation from [3H]PA in depleted
DROS in the presence of LPA, S1P or C1P was 57%, 61%
and 86% with respect to the activity determined in the
presence of [3H]PA alone. In depleted BLROS membranes,
DAG generation in the presence of LPA and S1P was 64%
and 50% in the presence of C1P with respect to the activity
found in the presence of [3H]PA alone.
Hydrolysis of [2-3H]-phosphatidate by LPPs from
DROS and BLROS in the Presence of Sphingosine and
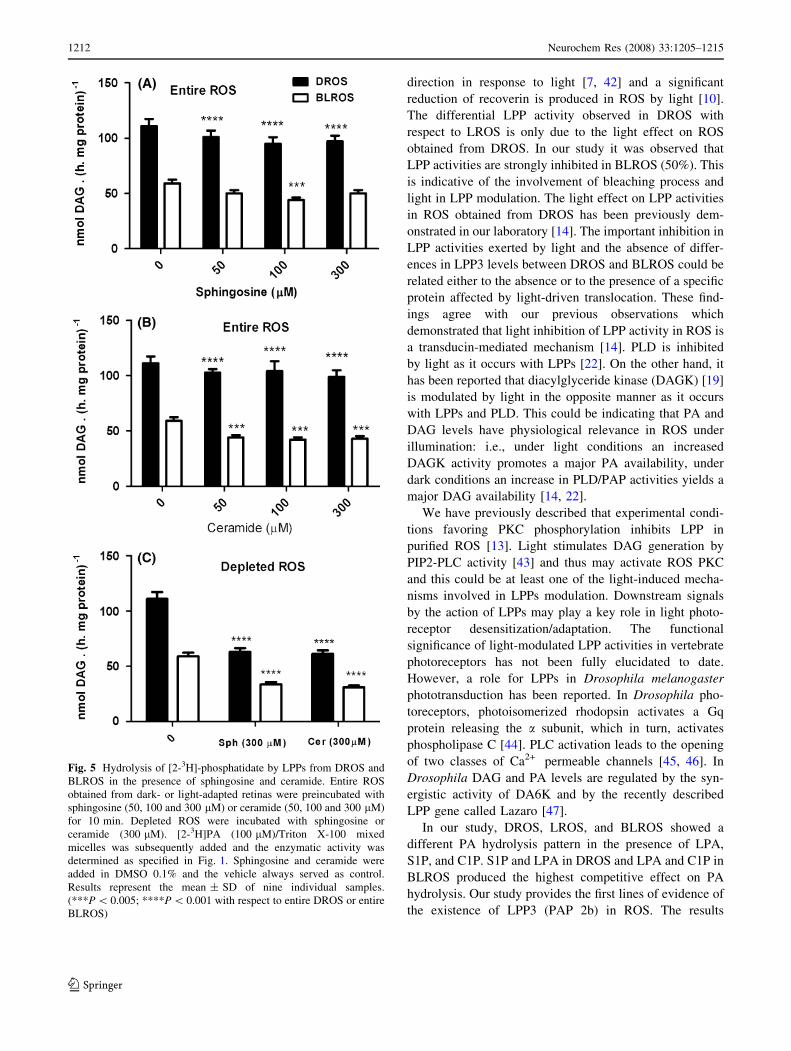
Ceramide
In order to determine if the effect of S1P and C1P on DAG
production was due either to their competitive character-
istics and/or to the sphingosine and ceramide, the
respective dephosphorylation products of S1P or C1P by
LPPs, sphingosine or ceramide were included in PA
hydrolysis assays. Sphingosine and ceramide were added
using DMSO as vehicle. ROS were preincubated with
sphingosine and ceramide but similar results were observed
when sphingosine and ceramide were co-incubated with
PA (data not shown). In entire DROS, sphingosine and
ceramide inhibited DAG production by a similar percent-
age (18%) at all concentrations assayed (Fig. 5A, B). In
entire BLROS sphingosine (100 lM) significantly inhib-
ited DAG production by 23% and ceramide (50, 100,
300 lM) yielded the same inhibition percentage as that of
Fig. 2 Hydrolysis of [2-3H]-phosphatidate by LPPs in LROS and
BLROS in the presence of LPA, S1P and C1P. BLROS were
obtained as was specified in Fig. 1. LROS were prepared following
the same procedure as that of DROS but the enzymatic assay was
made under room light. Results are expressed as the percentage of
LPP activities in LROS or BLROS with respect to DROS and they
represent the mean ± SD of nine individual samples. (**P \ 0.01)
1210 Neurochem Res (2008) 33:1205–1215
123
sphingosine (Fig. 5B). In ROS depleted of soluble and
peripheral proteins sphingosine and ceramide inhibited
DAG formation by 48% and 40% in DROS and BLROS,
respectively (Fig. 5C).
Discussion
The key enzyme LPPs are the main class of enzymes
responsible for the dephosphorylation in vitro of PA and of
the other phosphorylated substrates, such as LPA, S1P, and
C1P. Three mammalian LPP isoforms termed LPP1
(PAP2a), LPP2 (PAP2c), and LPP3 (PAP2b) have been
cloned [29]. These substrates are mutually competitive
with each other. In general, LPP1, LPP2, and LPP3 show
the major catalytic efficiency to LPA, PA, and S1P,
respectively [36, 37]. The primary function of LPPs is a
direct consequence of the dephosphorylation of their lipid
substrates, altering the balance of bioactive lipid mediators
[2, 38, 39]. PA and its dephosphorylated product, DAG,
have important functions in signaling and PA itself emer-
ges as a regulator of pleiotropic signaling responses [40].
ROS are responsible for the initial events of vision at
low light levels. Light modulation of lipid metabolism and
of a number of enzymes that generate second lipid mes-
sengers in ROS has been thoroughly studied [14, 16–19,
21, 22]. In addition, light-dependent membrane transloca-
tion of phototransduction proteins has been documented in
detail [7, 41, 42].
Arrestin, transducin and recoverin redistribute in rods in
response to bright light. Arrestin predominates at rod inner
segments from DROS shifting to ROS during light expo-
sure. Transducin a and b subunits shift in the opposite
Fig. 3 SDS–PAGE and immunoblot analysis. ROS depleted of
peripheral and soluble proteins were obtained from DROS or
BLROS. Supernatants were removed (soluble ROS fraction) and
membrane pellets (depleted ROS) were analyzed by SDS–PAGE and
immunoblot. (A) Commassie Blue-stained gel of entire ROS, ROS
soluble fraction and depleted ROS either from dark- or light-adapted
retinas. (B) Immunoblot of entire ROS, ROS soluble fraction and
depleted ROS from dark- or light-adapted retinas with RGD domain
anti-phosphatidic acid phosphatase 2b (anti-LPP3) (upper panel).
Immunoblots obtained with anti LPP3 antibodies were stripped and
reprobed with anti-opsin antibody. Each lane contains 30 lg of
protein. (C) LPP3/opsin ratios (expressed as arbitrary units) were
calculated after densitometry analysis of immunoblots
Fig. 4 Hydrolysis of [2-3H]-phosphatidate by LPPs in DROS and
BLROS depleted of peripheral and soluble proteins. Effect of LPA,
S1P and C1P. LPPs activities were assayed in depleted ROS obtained
from either dark- or light-adapted retinas. Membranes were incubated
with 100 lM of [2-3H]PA/Triton X-100 mixed micelle or with
[2-3H]PA in the presence of 100 lM of LPA, S1P or C1P as was
specified in Fig. 1. Results represent the mean ± SD of nine
individual samples (****P \ 0.001)
Neurochem Res (2008) 33:1205–1215 1211
123
direction in response to light [7, 42] and a significant
reduction of recoverin is produced in ROS by light [10].
The differential LPP activity observed in DROS with
respect to LROS is only due to the light effect on ROS
obtained from DROS. In our study it was observed that
LPP activities are strongly inhibited in BLROS (50%). This
is indicative of the involvement of bleaching process and
light in LPP modulation. The light effect on LPP activities
in ROS obtained from DROS has been previously dem-
onstrated in our laboratory [14]. The important inhibition in
LPP activities exerted by light and the absence of differ-
ences in LPP3 levels between DROS and BLROS could be
related either to the absence or to the presence of a specific
protein affected by light-driven translocation. These find-
ings agree with our previous observations which
demonstrated that light inhibition of LPP activity in ROS is
a transducin-mediated mechanism [14]. PLD is inhibited
by light as it occurs with LPPs [22]. On the other hand, it
has been reported that diacylglyceride kinase (DAGK) [19]
is modulated by light in the opposite manner as it occurs
with LPPs and PLD. This could be indicating that PA and
DAG levels have physiological relevance in ROS under
illumination: i.e., under light conditions an increased
DAGK activity promotes a major PA availability, under
dark conditions an increase in PLD/PAP activities yields a
major DAG availability [14, 22].
We have previously described that experimental condi-
tions favoring PKC phosphorylation inhibits LPP in
purified ROS [13]. Light stimulates DAG generation by
PIP2-PLC activity [43] and thus may activate ROS PKC
and this could be at least one of the light-induced mecha-
nisms involved in LPPs modulation. Downstream signals
by the action of LPPs may play a key role in light photo-
receptor desensitization/adaptation. The functional
significance of light-modulated LPP activities in vertebrate
photoreceptors has not been fully elucidated to date.
However, a role for LPPs in Drosophila melanogaster
phototransduction has been reported. In Drosophila pho-
toreceptors, photoisomerized rhodopsin activates a Gq
protein releasing the a subunit, which in turn, activates
phospholipase C [44]. PLC activation leads to the opening
of two classes of Ca2+ permeable channels [45, 46]. In
Drosophila DAG and PA levels are regulated by the syn-
ergistic activity of DA6K and by the recently described
LPP gene called Lazaro [47].
In our study, DROS, LROS, and BLROS showed a
different PA hydrolysis pattern in the presence of LPA,
S1P, and C1P. S1P and LPA in DROS and LPA and C1P in
BLROS produced the highest competitive effect on PA
hydrolysis. Our study provides the first lines of evidence of
the existence of LPP3 (PAP 2b) in ROS. The results
Fig. 5 Hydrolysis of [2-3H]-phosphatidate by LPPs from DROS and
BLROS in the presence of sphingosine and ceramide. Entire ROS
obtained from dark- or light-adapted retinas were preincubated with
sphingosine (50, 100 and 300 lM) or ceramide (50, 100 and 300 lM)
for 10 min. Depleted ROS were incubated with sphingosine or
ceramide (300 lM). [2-3H]PA (100 lM)/Triton X-100 mixed
micelles was subsequently added and the enzymatic activity was
determined as specified in Fig. 1. Sphingosine and ceramide were
added in DMSO 0.1% and the vehicle always served as control.
Results represent the mean ± SD of nine individual samples.
(***P \ 0.005; ****P \ 0.001 with respect to entire DROS or entire
BLROS)
1212 Neurochem Res (2008) 33:1205–1215
123

observed in the presence of S1P reinforced the existence of
LPP3 in ROS [29]. In view of the results obtained with
LPA we cannot discard the presence of LPP1 and LPP2
besides LPP3 in purified ROS. LPP3 has been localized
with PLD in caveolin-enriched detergent-resistant micro-
domains (DRMs) where LPP3 metabolizes phospholipase
D2-derived PA [48, 49]. The presence of PLD activity [48]
and the existence of DRMs [50] in purified ROS, reinforce
a physiological role for LPP3 in this system.
Previous observations suggest that LPP3 protein regu-
lates cell–cell interactions and act as an ecto-enzyme [3,
51]. It was previously demonstrated that LPP from ROS is
mainly located in the disk membrane [11]. For this reason
ecto- LPP activity could provide lipid messengers in the
extracellular medium inside the ROS disks. The pattern
observed in the presence of S1P, mainly in DROS, could be
either a consequence of a competition with PA by LPPs or
it could be due to the formation of sphingosine from S1P,
which may inhibit PA hydrolysis. This possibility was
corroborated by our observations of the sphingosine effect
on DAG generation (Fig. 4). In this respect, it has been
reported that sphingosine not only inhibits DAG formation
but also stimulates PA formation, inhibiting LPPs and
stimulating PLD and DAGK [52, 53].
C1P is a potent inhibitor of protein phosphatases (PP)
and PPs have been implicated in the inhibition of LPPs in
isolated ROS [13, 54, 55]. Another possibility is that C1P
exerts a direct action on LPPs. It has been reported that
C1P is required for the activation and translocation of other
enzymes involved in lipid metabolism such as cytosolic
phospholipase A2 [56]. An inhibition in DAG production
from PA by ceramide was also observed. Ceramide itself is
an important second messenger in various stress responses
and there are several candidates for ceramide-regulated
enzymes [57]. Increased intracellular ceramide levels have
been involved in the activation of photoreceptor apoptosis
[58]. Furthermore, it has been suggested that LPP2 and
LPP3 play an important role in apoptotic processes. This is
supported by the fact that DAG and sphingosine, the
products of LPPs, are involved in the apoptosis induction
[39] while S1P and LPA have anti-apoptotic roles [59].
Our results regarding basal LPP activities under dark
and light conditions showed no differences between entire
ROS and ROS depleted of soluble and peripheral proteins,
thus corroborating the fact that LPPs are membrane-asso-
ciated proteins. On the contrary, it has been reported that
DAGK exhibits a different activity in ROS depleted of
soluble and peripheral proteins with respect to intact ROS
[60]. The effects of LPP alternative substrates obtained in
depleted ROS with respect to that observed in entire ROS
could be indicating the involvement of soluble and/or
peripheral proteins in the regulation of LPPs (Fig. 4). The
presence of sphingosine and ceramide strongly inhibited
LPPs activities in depleted DROS, being the activity levels
similar to those found in depleted BLROS (in the absence
of sphingosine or ceramide) (Fig. 5C).
Our findings indicate that the competition between PA
and LPA, or S1P or C1P by the active site of LPPs is
modulated by ROS illumination state and by ROS protein
association/dissociation. The role of LPPs in signaling
events and the fact that the products generated by these
enzymes control the cell-death decisions in photoreceptor
cells, open an interesting field to further studies about the
regulation of LPPs by light in these cells.
Acknowledgments This work was supported by the Consejo Nac-
ional de Investigaciones Cientıficas y Tecnicas (CONICET), the
Agencia Nacional de Promocion Cientıfica y Tecnologica (FONCyT)
and the Secretarıa General de Ciencia y Tecnologıa, Universidad
Nacional del Sur, Argentina. N.M.Giusto and G.A. Salvador are
research members of CONICET.
References
1. McDermott MI, Sigal YJ, Crump JS et al (2006) Enzymatic
analysis of lipid phosphate phosphatases. Methods 39:169–179
2. Brindley DN, Waggoner DW (1998) Mammalian lipid phosphate
phosphohydrolases. J Biol Chem 273:24281–24284
3. Escalante-Alcalde D, Hernandez L, Le Stunff H et al (2003) The
lipid phosphatase LPP3 regulates extra-embryonic vasculogene-
sis and axis patterning. Development 130:4623–4637
4. Renault AD, Sigal YJ, Morris AJ et al (2004) Soma-germ line
competition for lipid phosphate uptake regulates germ cell
migration and survival. Science 305:1963–1966
5. Pyne S, Kong KC, Darroch PI (2004) Lysophosphatidic acid and
sphingosine 1-phosphate biology: the role of lipid phosphate
phosphatases. Semin Cell Dev Biol 15:491–501
6. Sokolov M, Lyubarsky AL, Strissel KJ et al (2002) Massive light-
driven translocation of transducin between the two major com-
partments of rod cells: a novel mechanism of light adaptation.
Neuron 34:95–106
7. Philp NJ, Chang W, Long K (1987) Light-stimulated protein
movement in rod photoreceptor cells of the rat retina. FEBS Lett
225:127–132
8. Broekhuyse RM, Janssen AP, Tolhuizen EF (1987) Effect of
light-adaptation on the binding of 48-kDa protein (S-antigen) to
photoreceptor cell membranes. Curr Eye Res 6:607–610
9. Kerov V, Chen D, Moussaif M et al (2005) Transducin activation
state controls its light-dependent translocation in rod photore-
ceptors. J Biol Chem 280:41069–41076
10. Strissel KJ, Lishko PV, Trieu LH et al (2005) Recoverin under-
goes light-dependent intracellular translocation in rod
photoreceptors. J Biol Chem 280:29250–29255
11. Pasquare de Garcıa SJ, Giusto NM (1986) Phosphatidate phos-
phatase activity in isolated rod outer segment from bovine retina.
Biochim Biophys Acta 875:195–202
12. Pasquare SJ, Giusto NM (1993) Differential properties of phos-
phatidate phosphohydrolase and diacylgliceride lipase activities
in retinal subcellular fractions and rod outer segments. Comp
Biochem Physiol 104:141–148
13. Roque ME, Pasquare SJ, Castagnet PI et al (1998) Can phos-
phorylation and dephosphorylation of rod outer segment
membranes affect phosphatidate phosphohydrolase and diacyl-
glycerol lipase activities? Comp Biochem Physiol B 119:85–93
Neurochem Res (2008) 33:1205–1215 1213
123

14. Pasquare SJ, Salvador GA, Roque ME et al (2000) Effect of light
on phosphatidate phosphohydrolase activity of retina rod outer
segments. The role of transducin. Arch Biochem Biophys
379:299–306
15. Giusto NM, Pasquare SJ, Salvador GA et al (2000) Lipid
metabolism in vertebrate retinal rod outer segments. Progr Lipid
Res 39:315–391
16. Ghalayini AJ, Anderson RE (1992) Activation of bovine rod
outer segment phospholipase C by arrestin. J Biol Chem
267:17977–17982
17. Castagnet PI, Giusto NM (1993) Properties of phospholipase A2
activity from bovine retinal rod outer segments. Exp Eye Res
56:709–719
18. Roque ME, Salvador GA, Giusto NM (1999) Light activation of
phosphatidylethanolamine N-methyltransferase in rod outer seg-
ments and its modulation by association states of transducin. Exp
Eye Res 69:555–562
19. Huang Z, Ghalayini A, Guo XX et al (2000) Light-mediated
activation of diacylglycerol kinase in rat and bovine rod outer
segments. J Neurochem 75:355–362
20. Guo X, Ghalayini A, Chen H et al (1997) Phosphatidylinositol 3-
kinase in bovine photoreceptor rod outer segments. Invest Oph-
thalmol Vis Sci 38:1873–1882
21. Rajala RV, McClellan ME, Ash JD et al (2002) In vivo regulation
of phosphoinositide 3-kinase in retina through light-induced
tyrosine phosphorylation of the insulin receptor beta-subunit. J
Biol Chem 277:43319–43326
22. Salvador GA, Giusto NM (2006) Phospholipase D from photore-
ceptor rod outer segments is a downstream effector of RhoA:
evidence of a light-dependent mechanism. Exp Eye Res 83:202–211
23. Kuhn H (1982) Light-regulated binding of proteins to photore-
ceptor membranes and its use for the purification of several rod
cell proteins. Methods Enzymol 81:556–564
24. Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685
25. Aveldano MI (1987) A novel group of very long chain polyenoic
fatty acids in dipolyunsaturated phosphatidylcholines from ver-
tebrate retina. J Biol Chem 262:1172–1179
26. Hodges TK, Leonard RT (1974) Purification of a plasma mem-
brane-bound adenosine triphosphatase from plant roots. Methods
Enzymol 32:392–406
27. Roque ME, Giusto NM (1995) Phosphatidylethanolamine N-
methyltransferase activity in isolated rod outer segments from
bovine retina. Exp Eye Res 60:631–643
28. Kates M, Sastry PS (1969) Phospholipase D. Methods Enzymol
14:197–203
29. Folch J, Lees M, Sloane Stanley GH (1957) A Simple method for
the isolation and purification of total lipids from Animal Tissues.
J Biol Chem 226:497–509
30. Arvidson GAE (1968) Structural and metabolic heterogeneity of
rat liver glycerophosphatides. Eur J Biochem 4:478–486
31. Rouser G, Fleischer S, Yamamoto A (1970) Two dimensional
thin-layer chromatographic separation of polar lipids and deter-
mination of phospholipids by phosphorus analysis of spots.
Lipids 5:494–496
32. Hooks SB, Ragan SP, Lynch KR (1998) Identification of a Novel
Human Phosphatidic Acid Phosphatase Type 2 Isoform. FEBS
Lett 427:188–192
33. Roberts R, Sciorra VA, Morris AJ (1998) Human type 2 phos-
phatidic acid phosphohydrolases. Substrate specificity of the type
2a, 2b, and 2c enzymes and cell surface activity of the 2a isoform.
J Biol Chem 273:22059–22067
34. Giusto NM, Bazan NG (1979) Phospholipids and acylglycerols
biosynthesis and 14C production from [14C]glycerol in the bovine
retina: the effects of incubation time, oxygen and glucose, Exp.
Eye Res 29:155–168
35. Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein–dye binding. Anal Biochem 72:248–254
36. Alderton F, Darroch P, Sambi B et al (2001) G-protein-coupled
receptor stimulation of the p42/p44 mitogen-activated protein
kinase pathway is attenuated by lipid phosphate phosphatases 1,
1a, and 2 in human embryonic kidney 293 cells. J Biol Chem
276:13452–13460
37. Tanyi JL, Morris AJ, Wolf JK et al (2003) The human lipid
phosphate phosphatase-3 decreases the growth, survival, and
tumorigenesis of ovarian cancer cells: validation of the lyso-
phosphatidic acid signaling cascade as a target for therapy in
ovarian cancer. Cancer Res 63:1073–1082
38. Brindley DN (2004) Lipid phosphate phosphatases and related
proteins: signaling functions in development, cell division, and
cancer. J Cell Biochem 92:900–912
39. Pyne S, Long JS, Ktistakis NT et al (2005) Lipid phosphate
phosphatases and lipid phosphate signaling. Biochem Soc Trans
33:1370–1374
40. Carman GM, Han GS (2006) Roles of phosphatidate phos-
phatase enzymes in lipid metabolism. Trends Biochem Sci
31:694–699
41. Mangini NJ, Pepperberg DR (1988) Immunolocalization of 48 K
in rod photoreceptors. Light and ATP increase OS labeling.
Invest Ophthalmol Vis Sci 29:1221–1234
42. Whelan JP, McGinnis JF (1988) Light-dependent subcellular
movement of photoreceptor proteins. J Neurosci Res 20:263–270
43. Ghalayini AJ, Weber NR, Rundle DR et al (1998) Phospholipase
C gamma1 in bovine rod outer segments: immunolocalization
and light-dependent binding to membranes. J Neurochem
70:171–178
44. Bloomquist BT, Shortridge RD, Schneuwly S et al (1988) Iso-
lation of putative phospholipase C gene of Drosophila, norpA and
its role in phototransduction. Cell 54:723–733
45. Reuss H, Mojet MH, Chyb S et al (1997) In vivo analysis of the
Drosophila light-sensitive channels, TRP and TRPL. Neuron
19:1249–1259
46. Raghu P, Colley NJ, Webel R (2000) Normal phototransduction
in Drosophila photoreceptors lacking an InsP(3) receptor gene.
Mol Cell Neurosci 15:429–445
47. Garcia-Murillas I, Pettitt, Macdonald E et al (2006) Lazaroencodes a lipid phosphate phosphohydrolase that regulates
phosphatidilinositol turnover during Drosophila phototransduc-
tion. Neuron 49:533–546
48. Sciorra VA, Morris AJ (1999) Sequential actions of phospholi-
pase D and phosphatidic acid phosphohydrolase 2b generate
diglyceride in mammalian cells. Mol Biol Cell 10:3863–3876
49. Salvador GA, Giusto NM (1998) Characterization of phospholi-
pase D activity in bovine photoreceptor membranes. Lipids
33:853–860
50. Seno K, Kishimoto M, Abe M et al (2001) Light- and guanosine
50-3-O-(thio)triphosphate-sensitive localization of a G protein and
its effector on detergent-resistant membrane rafts in rod photo-
receptor outer segments. J Biol Chem 276:20813–20816
51. Humtsoe JO, Feng S, Thakker GD et al (2003) Regulation of
cell–cell interactions by phosphatidic acid phosphatase 2b/VCIP.
EMBO J 22:1539–1554
52. Waggoner DW, Xu J, Singh I et al (1999) Structural organization
of mammalian lipid phosphate phosphatases: implications for
signal transduction. Biochim Biophys Acta 1439:299–316
53. Alessenko AV (1998) Functions of sphingosine in cell prolifer-
ation and death. Biochemistry (Mosc) 63:62–68
54. Chalfant CE (2004) Ceramide kinase. In: Haldar D, Das KS (eds)
Lipids: Sphingolipid Metabolizing Enzymes. Kerala India,
Research Signpost, pp 17–31
1214 Neurochem Res (2008) 33:1205–1215
123

55. Palczewski K, Hargrave PA, McDowell JH et al (1989) The
catalytic subunit of phosphatase 2A dephosphorylates phosp-
hoopsin. Biochemistry 28:415–419
56. Chalfant CE, Spiegel S (2005) Sphingosine 1-phosphate and
ceramide 1-phosphate: expanding roles in cell signaling. J Cell
Sci 118:4605–4612
57. Hannun YA, Obeid LM (2002) The Ceramide-centric universe of
lipid-mediated cell regulation: stress encounters of the lipid kind.
J Biol Chem 277:25847–25850
58. German OL, Miranda GE, Abrahan CE et al (2006) Ceramide is a
mediator of apoptosis in retina photoreceptors. Invest Ophthalmol
Vis Sci 47:1658–1668
59. Grey A, Chen Q, Callon K et al (2002) The phospholipids
sphingosine-1-phosphate and lysophosphatidic acid prevent
apoptosis in osteoblastic cells via a signaling pathway involving
G(i) proteins and phosphatidylinositol-3 kinase. Endocrinology
143:4755–4763
60. Ilincheta de Boschero MG, Giusto NM (1992) Phosphatidic acid
and polyphosphoinositide metabolism in rod outer segments.
Differential role of soluble and peripheral proteins. Biochim
Biophys Acta 1127:105–115
Neurochem Res (2008) 33:1205–1215 1215
123









![Phosphoinositides and phosphatidic acid regulate pollen tube growth and reorientation through modulation of [Ca2+]c and membrane secretion](https://static.fdokumen.com/doc/165x107/63218d798a1d893baa0d20bd/phosphoinositides-and-phosphatidic-acid-regulate-pollen-tube-growth-and-reorientation.jpg)










