
Structure-function relationships in Drosophila melanogaster alcohol dehydrogenase allozymes ADHS,...
10
Eur. J. Biochem. 267, 3613–3622 (2000) q FEBS 2000 Structure–function relationships in Drosophila melanogaster alcohol dehydrogenase allozymes ADH S , ADH F and ADH UF , and distantly related forms Jordi Benach 1, *, Sı ´lvia Atrian 2, *, Joan Fibla 3 , Roser Gonza ` lez-Duarte 2 and Rudolf Ladenstein 1 1 Center for Structural Biochemistry, Karolinska Institutet, Huddinge, Sweden; 2 Departament de Gene `tica, Universitat de Barcelona, Spain; 3 Departament de Cie `ncies Me `diques Ba `siques, Universitat de Lleida, Spain Drosophila melanogaster alcohol dehydrogenase (ADH), a paradigm for gene–enzyme molecular evolution and natural selection studies, presents three main alleloforms (ADH S , ADH F and ADH UF ) differing by one or two substitutions that render different biochemical properties to the allelozymes. A three-dimensional molecular model of the three allozymes was built by homology modeling using as a template the available crystal structure of the orthologous D. lebanonensis ADH, which shares a sequence identity of 82.2%. Comparison between D. lebanonensis and D. melanogaster structures showed that there is almost no amino-acid change near the substrate or coenzyme binding sites and that the hydrophobic active site cavity is strictly conserved. Nevertheless, substitutions are not distributed at random in nonconstricted positions, or located in external loops, but they appear clustered mainly in secondary structure elements. From comparisons between D. melanogaster allozymes and with D. simulans, a very closely related species, a model based on changes in the electrostatic potential distribution is presented to explain their differential behavior. The depth of knowledge on Drosophila ADH genetics and kinetics, together with the recently obtained structural information, could provide a better understanding of the mechanisms underlying molecular evolution and population genetics. Keywords: alcohol dehydrogenase; Drosophila melanogaster ; 3D-model structure; molecular evolution; natural polymorphism. Drosophila alcohol dehydrogenase (ADH) has been the subject of intense research during the last 35 years, and has illuminated important debates on molecular biology, evolution, ecology and population genetics [1–4]. However, some of the main issues raised long ago remain unanswered, in particular, those which imply deep knowledge on the molecular structure and reaction mechanism and could be the basis of the driving forces of natural selection and adaptation to alcohol-rich environments. Drosophila ADH (EC 1.1.1.1) belongs to a broad and heterogeneous family of alcohol dehydrogenases, named short-chain dehydrogenases/reductases (SDR) [5], and is the only member that utilizes small alcohols as substrates. All the others are mostly steroid and prostaglandin dehydrogenases/ reductases, of prokaryotic and mammalian origin. Common ethanol-oxidizing enzymes belong to the evolutionary non- related medium-chain dehydrogenases/reductase (MDR) family, reported long ago in mammals, first in horse liver and later in many other organisms [6]. Therefore, neither the distant MDR members nor the closer SDR relatives are good models to assess the structure–function relationship of the Drosophila enzyme. A recent breakthrough to this impasse has been the elucidation of both the crystal structure and the reaction mechanism of Drosophila lebanonensis ADH [7,8]. From these data, many questions regarding structure–function relationships could now be approached, not only in this species, but also in the paradigmatic D. melanogaster. Substantial differences in ADH activity have been reported among Drosophila species and also in different allomorphs of the same species. They have been related to ethanol tolerance, thus alcohol-rich ecological niche exploitation, and finally Drosophila speciation. Natural population studies showed that the three most frequent D. melanogaster Adh alleles, Adh UF , Adh F and Adh S exhibit distinct geographic distribution [9], but no molecular explanation could account for this fact. Few amino-acid substitutions are involved in this polymorphism with respect to ADH S : ADH F has only a Lys192Thr replacement, and ADH UF has two additional replacements, Asn8Ala and Ala45Asp. Another highly studied species was D. simulans, very closely related to D. melanogaster, but with a poor capacity to exploit ethanol-rich environments [10]. Regarding D. melanogaster ADH S , it differs only in positions 1 (Ala1Ser) and 82 (Lys82Gln). Although in the absence of the tertiary structure the effects of amino-acid substitutions had been shown kinetically [11–15], it was not possible to assess how they could alter the overall conformation or modulate the activity by providing new chemical interactions. To address all these questions, a molecular model of the D. melanogaster ADH S enzyme based on the tertiary structure of the D. lebanonensis enzyme was constructed. The high sequence identity (82.3%) and homology (87.4%) between these two Correspondence to: R. Ladenstein, Center for Structural Biochemistry, Karolinska Institutet-NOVUM, Huddinge, Sweden. Fax: 1 46 8 60 89 290, Tel.: 1 46 8 60 89 222, E-mail: [email protected] Abbreviations: ADH, alchol dehydrogenase; DADH, Drosophila alcohol dehydrogenase; MDR, medium-chain dehydrogenase/reductase SDR, short-chain dehydrogenase/reductase. Enzymes: alcohol dehydrogenase (EC 1.1.1.1). *Note: J. Benach and S. Atrian contributed equally to this study. (Received 18 February 2000, revised 29 March 2000, accepted 3 April 2000)
-
Upload
damiavericat -
Category
Documents
-
view
0 -
download
0
Transcript of Structure-function relationships in Drosophila melanogaster alcohol dehydrogenase allozymes ADHS,...

Eur. J. Biochem. 267, 3613±3622 (2000) q FEBS 2000
Structure±function relationships in Drosophila melanogaster alcoholdehydrogenase allozymes ADHS, ADHF and ADHUF, and distantlyrelated forms
Jordi Benach1,*, SõÂlvia Atrian2,*, Joan Fibla3, Roser GonzaÁ lez-Duarte2 and Rudolf Ladenstein1
1Center for Structural Biochemistry, Karolinska Institutet, Huddinge, Sweden; 2Departament de GeneÁtica, Universitat de Barcelona, Spain;3Departament de CieÁncies MeÁdiques BaÁsiques, Universitat de Lleida, Spain
Drosophila melanogaster alcohol dehydrogenase (ADH), a paradigm for gene±enzyme molecular evolution and
natural selection studies, presents three main alleloforms (ADHS, ADHF and ADHUF) differing by one or two
substitutions that render different biochemical properties to the allelozymes. A three-dimensional molecular
model of the three allozymes was built by homology modeling using as a template the available crystal structure
of the orthologous D. lebanonensis ADH, which shares a sequence identity of 82.2%. Comparison between
D. lebanonensis and D. melanogaster structures showed that there is almost no amino-acid change near the
substrate or coenzyme binding sites and that the hydrophobic active site cavity is strictly conserved. Nevertheless,
substitutions are not distributed at random in nonconstricted positions, or located in external loops, but they
appear clustered mainly in secondary structure elements. From comparisons between D. melanogaster allozymes
and with D. simulans, a very closely related species, a model based on changes in the electrostatic potential
distribution is presented to explain their differential behavior. The depth of knowledge on Drosophila ADH
genetics and kinetics, together with the recently obtained structural information, could provide a better
understanding of the mechanisms underlying molecular evolution and population genetics.
Keywords: alcohol dehydrogenase; Drosophila melanogaster ; 3D-model structure; molecular evolution; natural
polymorphism.
Drosophila alcohol dehydrogenase (ADH) has been the subjectof intense research during the last 35 years, and has illuminatedimportant debates on molecular biology, evolution, ecology andpopulation genetics [1±4]. However, some of the main issuesraised long ago remain unanswered, in particular, those whichimply deep knowledge on the molecular structure and reactionmechanism and could be the basis of the driving forces ofnatural selection and adaptation to alcohol-rich environments.
Drosophila ADH (EC 1.1.1.1) belongs to a broad andheterogeneous family of alcohol dehydrogenases, namedshort-chain dehydrogenases/reductases (SDR) [5], and is theonly member that utilizes small alcohols as substrates. All theothers are mostly steroid and prostaglandin dehydrogenases/reductases, of prokaryotic and mammalian origin. Commonethanol-oxidizing enzymes belong to the evolutionary non-related medium-chain dehydrogenases/reductase (MDR) family,reported long ago in mammals, first in horse liver and later inmany other organisms [6]. Therefore, neither the distant MDRmembers nor the closer SDR relatives are good models to
assess the structure±function relationship of the Drosophilaenzyme. A recent breakthrough to this impasse has been theelucidation of both the crystal structure and the reactionmechanism of Drosophila lebanonensis ADH [7,8]. From thesedata, many questions regarding structure±function relationshipscould now be approached, not only in this species, but also inthe paradigmatic D. melanogaster. Substantial differences inADH activity have been reported among Drosophila speciesand also in different allomorphs of the same species. They havebeen related to ethanol tolerance, thus alcohol-rich ecologicalniche exploitation, and finally Drosophila speciation. Naturalpopulation studies showed that the three most frequentD. melanogaster Adh alleles, AdhUF, AdhF and AdhS exhibitdistinct geographic distribution [9], but no molecularexplanation could account for this fact. Few amino-acidsubstitutions are involved in this polymorphism with respectto ADHS: ADHF has only a Lys192Thr replacement, andADHUF has two additional replacements, Asn8Ala andAla45Asp. Another highly studied species was D. simulans,very closely related to D. melanogaster, but with a poorcapacity to exploit ethanol-rich environments [10]. RegardingD. melanogaster ADHS, it differs only in positions 1 (Ala1Ser)and 82 (Lys82Gln). Although in the absence of the tertiarystructure the effects of amino-acid substitutions had beenshown kinetically [11±15], it was not possible to assess howthey could alter the overall conformation or modulate theactivity by providing new chemical interactions. To address allthese questions, a molecular model of the D. melanogasterADHS enzyme based on the tertiary structure of theD. lebanonensis enzyme was constructed. The high sequenceidentity (82.3%) and homology (87.4%) between these two
Correspondence to: R. Ladenstein, Center for Structural Biochemistry,
Karolinska Institutet-NOVUM, Huddinge, Sweden.
Fax: 1 46 8 60 89 290, Tel.: 1 46 8 60 89 222,
E-mail: [email protected]
Abbreviations: ADH, alchol dehydrogenase; DADH, Drosophila alcohol
dehydrogenase; MDR, medium-chain dehydrogenase/reductase SDR,
short-chain dehydrogenase/reductase.
Enzymes: alcohol dehydrogenase (EC 1.1.1.1).
*Note: J. Benach and S. Atrian contributed equally to this study.
(Received 18 February 2000, revised 29 March 2000, accepted
3 April 2000)

ADHs fully justified the construction of a three-dimensionalmodel by molecular modeling. The validity of the proposedstructure was assessed and the ADHF, ADHUF and D. simulansvariants were analyzed. Overall conformation, electrostaticpotentials and coenzyme binding energies were correlated withcatalytic efficiencies.
Other means to ascertain the structure±function relationshipsof individual residues and small protein segments are throughnatural and induced mutants studies. Yet, an alternativeapproach is to obtain mAbs against a particular protein regionand evaluate the effects of binding. Here, we also report theepitope mapping of mAb MMBB8 and show that it recognizesthe peptide encompassing the D. melanogaster ADHS/ADHF
substitution, Thr192Lys, an external disordered loop found inthe apo configuration of ADH. The inhibitory effect ofMMBB8 on the catalytic activity stresses the contribution ofthis region to proper folding of the tertiary complex and allowsus to presume a direct relationship between primary structureand catalytic efficiency.
M A T E R I A L S A N D M E T H O D S
Three-dimensional structure modeling and analysis
Molecular modeling procedures were performed on a SiliconGraphics Octane R10000 workstation. whatif (v4.99) [16] wasused to generate a primary model (BLDPIR), as well as toverify all model structures shown in this work (check).Basic sequence manipulations, alignments, energy minimiza-tion and molecular dynamics routines were performed by theHomology/Discover modules of insight II [17], using thebinary force field library included (cvff.frc 2.3, Biosym).xplor [18] was used in the last steps of positional refinement.Throughout the model building and refinement process, thequality of the model was assessed by a number of criteria,including Ramachandran plots drawn using the procheckprogram [19]. whatif was used to validate the model.promotif [20] helped us to determine secondary structureelements. The grasp utility [21] was used to calculate andrepresent electrostatic surface potentials. Molecular graphicswere created with the programs molscript [22] and ribbons[23].
The template used to build the D. melanogaster ADHS three-dimensional model was the crystal structure of the orthologousapoenzyme D. lebanonensis ADH [7]. When needed, dataabout the position of the coenzyme (NAD1) and the substratewere extracted from the crystal structures of binary and ternaryenzyme complexes [8]. A basic model structure, obtained withwhatif, was submitted to the first rounds of energy mini-mization through discover, disregarding cross terms andMorse-bond stretching potentials. A cut-off of 10 AÊ wasapplied with a switching distance of 1.5 AÊ and an overlapof 0.01 AÊ . A dielectric constant of 1 was used in all mini-mizations. Structures were minimized using conjugate gradientdescent (maximum derivative , 4.2 J. The model was thensubjected to 2000 positional refinement cycles in xplor and aharmonic potential (83 kJ´mol21´AÊ 22) was applied to the mainchain of the model to restrain it to the structure ofD. lebanonensis ADH. Non-crystallographic symmetry wasapplied to the dimer with a restraining potential of1254 kJ´mol21´AÊ 22.
Finally, D. melanogaster ADHF and ADHUF allelomorphs, aswell as D. simulans ADH, were modeled on the ADHS
structure, by the substitution in each position of the corres-
ponding residues. The ADHUF structure was minimized inxplor.
ADH purification and antigenic analysis
Monoclonal antibodies MMBB8, against D. melanogasterADH, and LLBE8, against D. lebanonensis ADH, wereobtained and characterized as described in Fibla et al. [24].The epitope recognized by LLBE8 mapped to the C-terminaltail of the ADH monomer [25]. Pure Drosophila ADH wasprepared according to published protocols [7]. Peptides wereobtained with an automatic synthesizer (Applied Biosystems,Model 430A) using t-BOC amino acids according to themanufacturer's instructions and were kindly provided byL. Enjuanes (CNB-CSIC, Madrid). Peptides carried a cysteinein the first position, not present in the homologous proteinsequence. The amino-acid content of each peptide wasconfirmed by amino-acid analysis.
Dot blotting and immunostaining were performed followingthe procedures reported in Visa et al. [26] using a peroxidase-conjugated MMBB8 [24]. ADH activity was quantified usingpure Drosophila ADH, by measuring the production of NADHat 340 nm. One unit of activity reduces 1 mmol of NAD1 perminute at 25 8C. The standard assays were performed in 0.12 mTris/HCl, buffer pH 8.6. All spectrophotometric measurementswere made on a Pharmacia-LKB Ultrospec III using thekinetics software.
R E S U LT S
Overall structure and quality of the model ofD. melanogaster ADH
Drosophila alcohol dehydrogenase exists as a dimer insolution and in crystals. We have previously reported forD. lebanonensis ADH [7] that the subunit shows the typicalfold of the short-chain dehydrogenase/reductase enzyme cores[5,27] thus exhibiting a three-dimensional structure similar tothat reported for other SDR proteins whose three-dimensionalstructure has been solved to date [28±44]. The subunit ofDADH has an a/b single domain structure with a characteristicRossmann fold dinucleotide-binding motif (bA-aB-bB-aC-bC-aD-bD-aE). It consists of a central b sheet flanked on eachside by three parallel a helices. The motif bD-aE-bE-aF-bF-a1-aG-bG, together with the adjoining loops, is responsible forquaternary association and substrate binding.
The model structure of D. melanogaster ADHS (Fig. 1A) wasbuilt from the crystal structure of D. lebanonensis ADH solvedand refined at 1.9 AÊ resolution [7]. Ramachandran plots of themain chain conformational angles showed that < 89% of theresidues lie within the most favored regions and < 11% inadditionally allowed regions. As in D. lebanonensis ADH, onlyone residue, Lys84, lies in the disallowed region. The rmsdbetween the model and crystal structure was 0.1 AÊ for 508 Caatoms. No severe clashes between side chains were observed inthe model after the last round of model building. Main chainhydrogen bonds are conserved in relation to D. lebanonensisADH as it is also the case for all the secondary structuralelements.
D. melanogaster ADHS vs. D. lebanonensis ADH
Comparison of the three-dimensional structure of D. lebano-nensis and D. melanogaster ADHS clearly showed that most ofthe replacements are clustered, and as many as 67% of the
3614 J. Benach et al. (Eur. J. Biochem. 267) q FEBS 2000
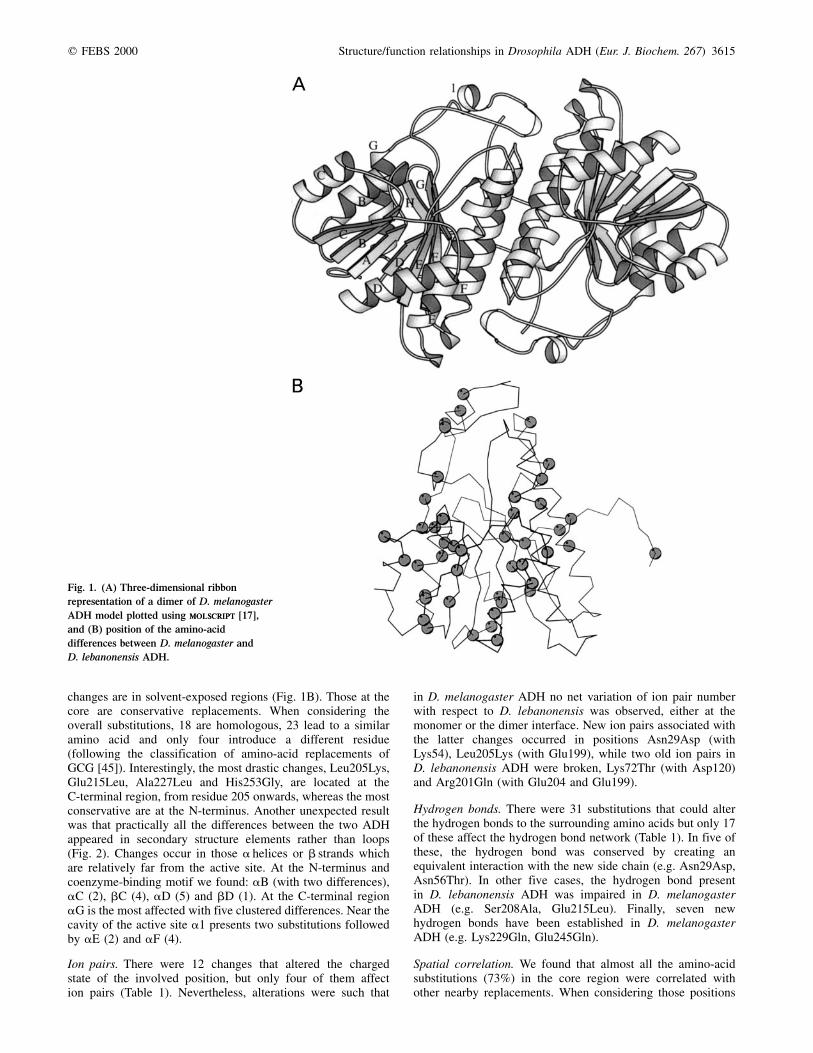
changes are in solvent-exposed regions (Fig. 1B). Those at thecore are conservative replacements. When considering theoverall substitutions, 18 are homologous, 23 lead to a similaramino acid and only four introduce a different residue(following the classification of amino-acid replacements ofGCG [45]). Interestingly, the most drastic changes, Leu205Lys,Glu215Leu, Ala227Leu and His253Gly, are located at theC-terminal region, from residue 205 onwards, whereas the mostconservative are at the N-terminus. Another unexpected resultwas that practically all the differences between the two ADHappeared in secondary structure elements rather than loops(Fig. 2). Changes occur in those a helices or b strands whichare relatively far from the active site. At the N-terminus andcoenzyme-binding motif we found: aB (with two differences),aC (2), bC (4), aD (5) and bD (1). At the C-terminal regionaG is the most affected with five clustered differences. Near thecavity of the active site a1 presents two substitutions followedby aE (2) and aF (4).
Ion pairs. There were 12 changes that altered the chargedstate of the involved position, but only four of them affection pairs (Table 1). Nevertheless, alterations were such that
in D. melanogaster ADH no net variation of ion pair numberwith respect to D. lebanonensis was observed, either at themonomer or the dimer interface. New ion pairs associated withthe latter changes occurred in positions Asn29Asp (withLys54), Leu205Lys (with Glu199), while two old ion pairs inD. lebanonensis ADH were broken, Lys72Thr (with Asp120)and Arg201Gln (with Glu204 and Glu199).
Hydrogen bonds. There were 31 substitutions that could alterthe hydrogen bonds to the surrounding amino acids but only 17of these affect the hydrogen bond network (Table 1). In five ofthese, the hydrogen bond was conserved by creating anequivalent interaction with the new side chain (e.g. Asn29Asp,Asn56Thr). In other five cases, the hydrogen bond presentin D. lebanonensis ADH was impaired in D. melanogasterADH (e.g. Ser208Ala, Glu215Leu). Finally, seven newhydrogen bonds have been established in D. melanogasterADH (e.g. Lys229Gln, Glu245Gln).
Spatial correlation. We found that almost all the amino-acidsubstitutions (73%) in the core region were correlated withother nearby replacements. When considering those positions
Fig. 1. (A) Three-dimensional ribbon
representation of a dimer of D. melanogaster
ADH model plotted using molscriptmolscript [17],
and (B) position of the amino-acid
differences between D. melanogaster and
D. lebanonensis ADH.
q FEBS 2000 Structure/function relationships in Drosophila ADH (Eur. J. Biochem. 267) 3615
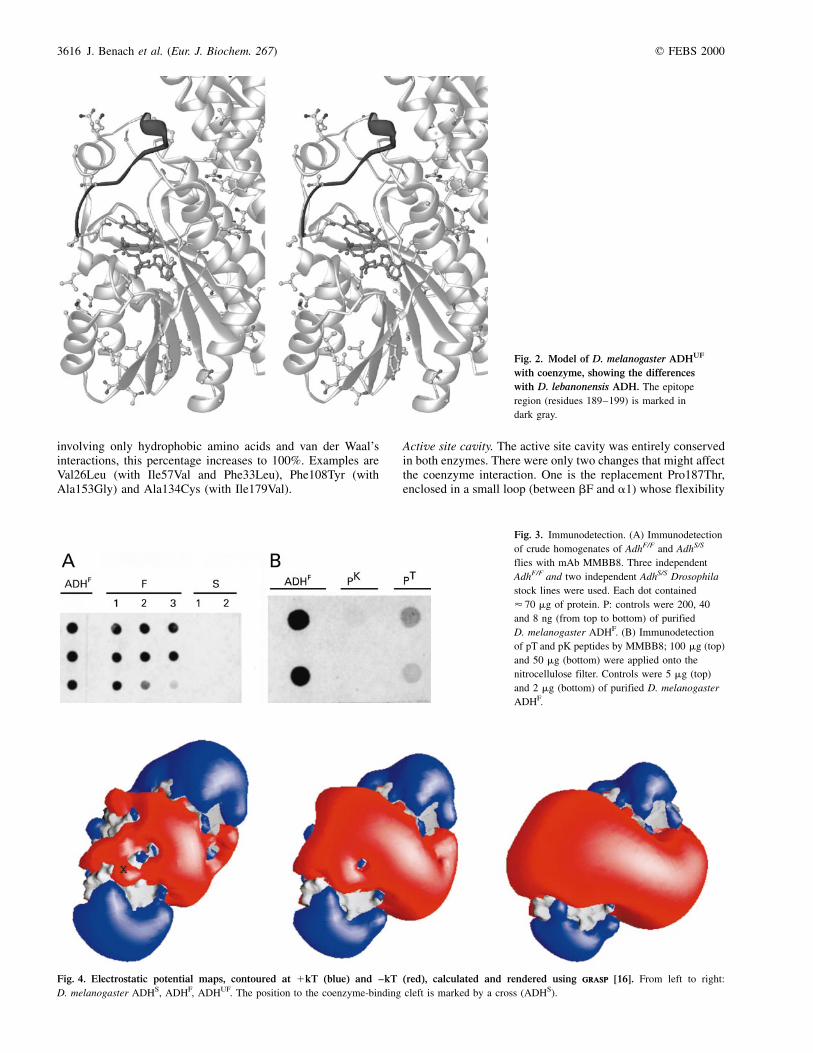
involving only hydrophobic amino acids and van der Waal'sinteractions, this percentage increases to 100%. Examples areVal26Leu (with Ile57Val and Phe33Leu), Phe108Tyr (withAla153Gly) and Ala134Cys (with Ile179Val).
Active site cavity. The active site cavity was entirely conservedin both enzymes. There were only two changes that might affectthe coenzyme interaction. One is the replacement Pro187Thr,enclosed in a small loop (between bF and a1) whose flexibility
Fig. 2. Model of D. melanogaster ADHUF
with coenzyme, showing the differences
with D. lebanonensis ADH. The epitope
region (residues 189±199) is marked in
dark gray.
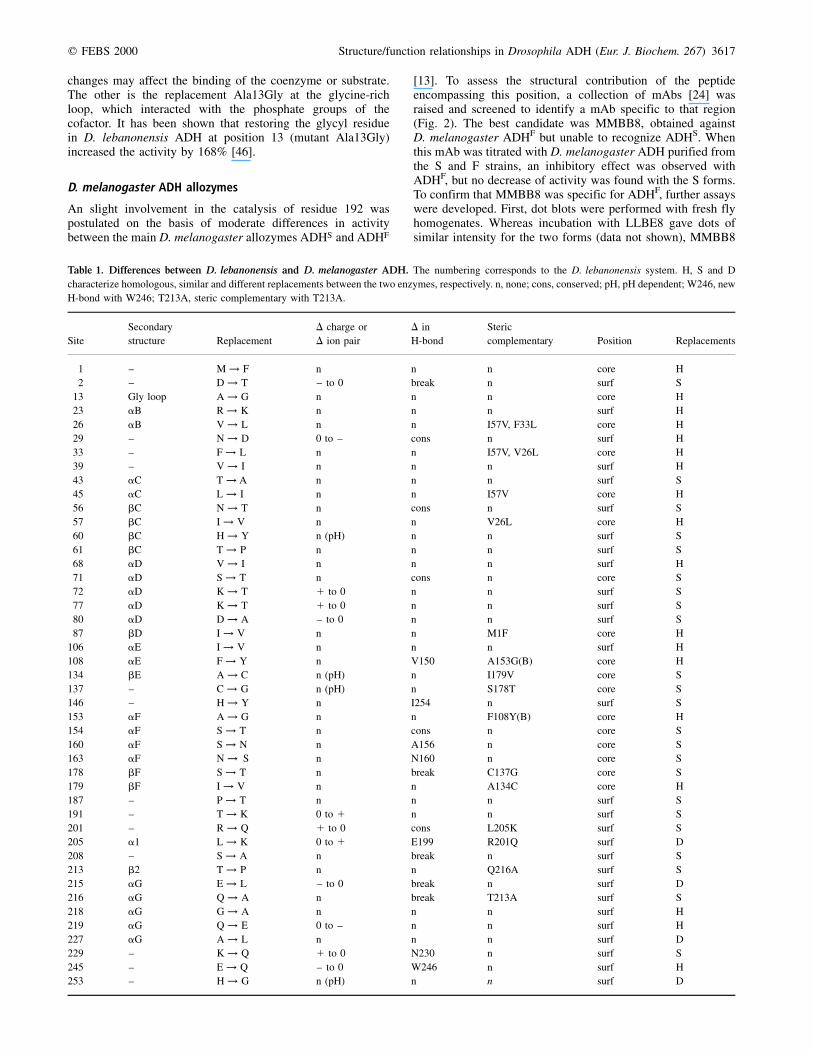
Fig. 3. Immunodetection. (A) Immunodetection
of crude homogenates of AdhF/F and AdhS/S
flies with mAb MMBB8. Three independent
AdhF/F and two independent AdhS/S Drosophila
stock lines were used. Each dot contained
< 70 mg of protein. P: controls were 200, 40
and 8 ng (from top to bottom) of purified
D. melanogaster ADHF. (B) Immunodetection
of pT and pK peptides by MMBB8; 100 mg (top)
and 50 mg (bottom) were applied onto the
nitrocellulose filter. Controls were 5 mg (top)
and 2 mg (bottom) of purified D. melanogaster
ADHF.
Fig. 4. Electrostatic potential maps, contoured at 1kT (blue) and ±kT (red), calculated and rendered using graspgrasp [16]. From left to right:
D. melanogaster ADHS, ADHF, ADHUF. The position to the coenzyme-binding cleft is marked by a cross (ADHS).
3616 J. Benach et al. (Eur. J. Biochem. 267) q FEBS 2000
changes may affect the binding of the coenzyme or substrate.The other is the replacement Ala13Gly at the glycine-richloop, which interacted with the phosphate groups of thecofactor. It has been shown that restoring the glycyl residuein D. lebanonensis ADH at position 13 (mutant Ala13Gly)increased the activity by 168% [46].
D. melanogaster ADH allozymes
An slight involvement in the catalysis of residue 192 waspostulated on the basis of moderate differences in activitybetween the main D. melanogaster allozymes ADHS and ADHF
[13]. To assess the structural contribution of the peptideencompassing this position, a collection of mAbs [24] wasraised and screened to identify a mAb specific to that region(Fig. 2). The best candidate was MMBB8, obtained againstD. melanogaster ADHF but unable to recognize ADHS. Whenthis mAb was titrated with D. melanogaster ADH purified fromthe S and F strains, an inhibitory effect was observed withADHF, but no decrease of activity was found with the S forms.To confirm that MMBB8 was specific for ADHF, further assayswere developed. First, dot blots were performed with fresh flyhomogenates. Whereas incubation with LLBE8 gave dots ofsimilar intensity for the two forms (data not shown), MMBB8
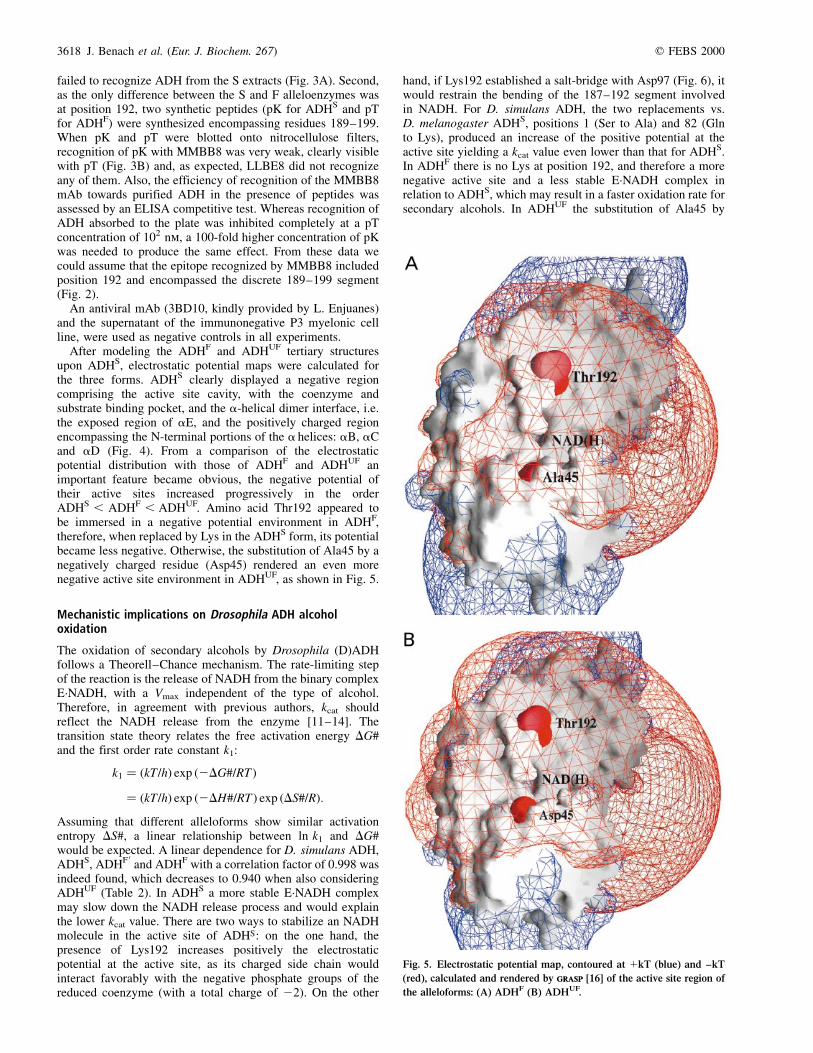
Table 1. Differences between D. lebanonensis and D. melanogaster ADH. The numbering corresponds to the D. lebanonensis system. H, S and D
characterize homologous, similar and different replacements between the two enzymes, respectively. n, none; cons, conserved; pH, pH dependent; W246, new
H-bond with W246; T213A, steric complementary with T213A.
Site
Secondary
structure Replacement
D charge or
D ion pair
D in
H-bond
Steric
complementary Position Replacements
1 ± M ! F n n n core H
2 ± D ! T ± to 0 break n surf S
13 Gly loop A ! G n n n core H
23 aB R ! K n n n surf H
26 aB V ! L n n I57V, F33L core H
29 ± N ! D 0 to ± cons n surf H
33 ± F ! L n n I57V, V26L core H
39 ± V ! I n n n surf H
43 aC T ! A n n n surf S
45 aC L ! I n n I57V core H
56 bC N ! T n cons n surf S
57 bC I ! V n n V26L core H
60 bC H ! Y n (pH) n n surf S
61 bC T ! P n n n surf S
68 aD V ! I n n n surf H
71 aD S ! T n cons n core S
72 aD K ! T 1 to 0 n n surf S
77 aD K ! T 1 to 0 n n surf S
80 aD D ! A ± to 0 n n surf S
87 bD I ! V n n M1F core H
106 aE I ! V n n n surf H
108 aE F ! Y n V150 A153G(B) core H
134 bE A ! C n (pH) n I179V core S
137 ± C ! G n (pH) n S178T core S
146 ± H ! Y n I254 n surf S
153 aF A ! G n n F108Y(B) core H
154 aF S ! T n cons n core S
160 aF S ! N n A156 n core S
163 aF N ! S n N160 n core S
178 bF S ! T n break C137G core S
179 bF I ! V n n A134C core H
187 ± P ! T n n n surf S
191 ± T ! K 0 to 1 n n surf S
201 ± R ! Q 1 to 0 cons L205K surf S
205 a1 L ! K 0 to 1 E199 R201Q surf D
208 ± S ! A n break n surf S
213 b2 T ! P n n Q216A surf S
215 aG E ! L ± to 0 break n surf D
216 aG Q ! A n break T213A surf S
218 aG G ! A n n n surf H
219 aG Q ! E 0 to ± n n surf H
227 aG A ! L n n n surf D
229 ± K ! Q 1 to 0 N230 n surf S
245 ± E ! Q ± to 0 W246 n surf H
253 ± H ! G n (pH) n n surf D
q FEBS 2000 Structure/function relationships in Drosophila ADH (Eur. J. Biochem. 267) 3617
failed to recognize ADH from the S extracts (Fig. 3A). Second,as the only difference between the S and F alleloenzymes wasat position 192, two synthetic peptides (pK for ADHS and pTfor ADHF) were synthesized encompassing residues 189±199.When pK and pT were blotted onto nitrocellulose filters,recognition of pK with MMBB8 was very weak, clearly visiblewith pT (Fig. 3B) and, as expected, LLBE8 did not recognizeany of them. Also, the efficiency of recognition of the MMBB8mAb towards purified ADH in the presence of peptides wasassessed by an ELISA competitive test. Whereas recognition ofADH absorbed to the plate was inhibited completely at a pTconcentration of 102 nm, a 100-fold higher concentration of pKwas needed to produce the same effect. From these data wecould assume that the epitope recognized by MMBB8 includedposition 192 and encompassed the discrete 189±199 segment(Fig. 2).
An antiviral mAb (3BD10, kindly provided by L. Enjuanes)and the supernatant of the immunonegative P3 myelonic cellline, were used as negative controls in all experiments.
After modeling the ADHF and ADHUF tertiary structuresupon ADHS, electrostatic potential maps were calculated forthe three forms. ADHS clearly displayed a negative regioncomprising the active site cavity, with the coenzyme andsubstrate binding pocket, and the a-helical dimer interface, i.e.the exposed region of aE, and the positively charged regionencompassing the N-terminal portions of the a helices: aB, aCand aD (Fig. 4). From a comparison of the electrostaticpotential distribution with those of ADHF and ADHUF animportant feature became obvious, the negative potential oftheir active sites increased progressively in the orderADHS , ADHF , ADHUF. Amino acid Thr192 appeared tobe immersed in a negative potential environment in ADHF,therefore, when replaced by Lys in the ADHS form, its potentialbecame less negative. Otherwise, the substitution of Ala45 by anegatively charged residue (Asp45) rendered an even morenegative active site environment in ADHUF, as shown in Fig. 5.
Mechanistic implications on Drosophila ADH alcoholoxidation
The oxidation of secondary alcohols by Drosophila (D)ADHfollows a Theorell±Chance mechanism. The rate-limiting stepof the reaction is the release of NADH from the binary complexE´NADH, with a Vmax independent of the type of alcohol.Therefore, in agreement with previous authors, kcat shouldreflect the NADH release from the enzyme [11±14]. Thetransition state theory relates the free activation energy DG#and the first order rate constant k1:
k1 � �kT/h� exp �2DG#/RT �� �kT/h� exp �2DH#/RT � exp �DS#/R�:
Assuming that different alleloforms show similar activationentropy DS#, a linear relationship between ln k1 and DG#would be expected. A linear dependence for D. simulans ADH,ADHS, ADHF 0 and ADHF with a correlation factor of 0.998 wasindeed found, which decreases to 0.940 when also consideringADHUF (Table 2). In ADHS a more stable E´NADH complexmay slow down the NADH release process and would explainthe lower kcat value. There are two ways to stabilize an NADHmolecule in the active site of ADHS: on the one hand, thepresence of Lys192 increases positively the electrostaticpotential at the active site, as its charged side chain wouldinteract favorably with the negative phosphate groups of thereduced coenzyme (with a total charge of 22). On the other
hand, if Lys192 established a salt-bridge with Asp97 (Fig. 6), itwould restrain the bending of the 187±192 segment involvedin NADH. For D. simulans ADH, the two replacements vs.D. melanogaster ADHS, positions 1 (Ser to Ala) and 82 (Glnto Lys), produced an increase of the positive potential at theactive site yielding a kcat value even lower than that for ADHS.In ADHF there is no Lys at position 192, and therefore a morenegative active site and a less stable E´NADH complex inrelation to ADHS, which may result in a faster oxidation rate forsecondary alcohols. In ADHUF the substitution of Ala45 by
Fig. 5. Electrostatic potential map, contoured at 1kT (blue) and ±kT
(red), calculated and rendered by graspgrasp [16] of the active site region of
the alleloforms: (A) ADHF (B) ADHUF.
3618 J. Benach et al. (Eur. J. Biochem. 267) q FEBS 2000

aspartate introduced another negative charge near the coenzymeand concomitantly increased the kcat, as in ADHF. Anothersubstitution found between ADHUF and ADHF is Asn8Alawhose location at the beginning of bA could indirectly affectthe binding of the coenzyme. The kcat value of D. lebanonensisADH is between that of ADHS and ADHF, which would be inaccordance with a more negative electrostatic potential in theactive site compared with that of ADHS and a more positivepotential in relation to ADHF. This is not, however, the case.The positively charged residues are placed in a way that rendersthe active site with a more positive character than that of ADHS.This implies that overall conformational differences betweenD. lebanonensis and D. melanogaster ADHS may affect therelease of NADH from the reactive complex. Recently, it hasbeen observed that the catalytic residue that regulates thebinding of NAD1 has a pKa which is 0.5 pH units lower inD. lebanonensis ADH than in D. melanogaster ADHS [47].This could be explained by the more positive characterof the active site of D. lebanonensis ADH comparedwith D. melanogaster ADHS. A more positive electrostaticpotential stabilizes the deprotonated form of the active siteresidue resulting in a lower pKa value.
For oxidation of primary alcohols, Vmax was found to bemuch lower than for secondary alcohols and varied with the
type of alcohol. The rate-limiting step involved at least hydridetransfer and dissociation of NADH from the E´NADH complex[12±14]. From the dependence of kcat on temperature, values ofDH#, DS# and DG# could be obtained [48]. The DH# value forD. simulans ADH turned out to be significantly higher than thatfor the D. melanogaster ADH allozymes. This could beinterpreted in two ways: either the transition state duringhydride transfer is destabilized by the increasingly negativelycharged active sites of ADHS and ADHF or rather, theE´NAD1´alcohol complex becomes stabilized by the negativelycharged active sites of ADHS and ADHF.
D I S C U S S I O N
No allozymic variation in natural populations has beenstudied more intensively than the AdhF/AdhS polymorphismat the Adh locus of D. melanogaster, as well as that of other lessfrequent alleles. After its description in 1964 [54], the ADHgene±enzyme system became the centerpiece of an intenseselectionist±neutralist debate aimed at explaining the main-tenance of enzymic polymorphisms in natural populations. Thestrongest evidence in favor of selection on the Adh locus wasthe account of a global geographical cline, the AdhS alleleincreased towards the Equator, while AdhF predominated athigher latitudes. The molecular bases for this phenomenon weresearched for at the biochemical level. After exhaustivecharacterization of the ADHF and ADHS enzymes threefeatures became evident. First, AdhF/F homozygous fliesshowed an increased amount of enzyme with respect to AdhS
individuals [55]. Second, the ADHF allozyme showed thermo-dynamic and steady-state kinetic differences regarding ADHS,these differences conferring an advantage in catalytic efficiency[13,48,53]. Third, the combination of both factors clearly pro-duced an increase in ethanol tolerance for the AdhF genotype,also supported by the artificial selection of flies grown onalcohol-supplemented media [56]. However, the presence of theAdhS allele at different frequencies in natural populationsremained elusive. Higher thermostability for ADHS wasinvoked to justify its prevalence in hot-climate latitudes [57].However, biochemical studies yielded controversial results: theincreased thermostability shown for pure ADHS preparations[58] was questioned after extensive evidence of lack ofcorrelation between temperature and any kinetic feature [53].
We modeled the D. melanogaster ADHS, ADHF andADHUF structures from the coordinates of the homologousD. lebanonensis enzyme. It is at this high level of resolution,where the effects of minor conformational differences, such aschanges due to a single amino-acid replacement, can be deeplyevaluated. We re-addressed the question of the increased
Table 2. Correlation between the calculated electrostatic DH# and the kcat values.
Enzyme Replacementsa kcat (s21)d DH#elec (kT)b References for kinetic data
ADH-sim Ser1Ala, Gln82Lys 1�.9 5�.2 [49]
ADH-S ± 3�.4±4.0 6�.0 [50±52]
ADH-F 0 Lys192Thr,Ala51Asp 4�.0 6�.2 [50]
ADH-F Lys192Thr 10�.4 7�.7 [13,50,51]
ADH-UF Lys192Thr,Asn8Ala, Ala45Asp 12±18 11�.0 [11,53]
ADH-leb ± c 7�.4 4�.4 [14]
a Replacements in relation to ADH-S. b Electrostatic energy of the NADH molecule in the DADH binary form, calculated using grasp [17]. c Sequence
similarity with ADH-S is 82%. d The catalytic center activities for 2-propanol, representing the dissociation rate of NADH from the enzyme±NADH
complex, extracted from the references mentioned and from Winberg & McKinley-McKee [15].
Fig. 6. Magnified portion of the active site of D. melanogaster ADHUF
with coenzyme in the region around Lys192 and its position in relation
to Asp97.
q FEBS 2000 Structure/function relationships in Drosophila ADH (Eur. J. Biochem. 267) 3619

catalytic efficiency of ADHF and the presumed higherthermostability of ADHS. Moreover, data deduced fromthis structure have allowed us to extend the comparison toother D. melanogaster ADH alleles and, furthermore, to specieswhich split 56 million years ago, such as D. lebanonensis.
D. lebanonensis ADH vs. D. melanogaster ADH
The validation of the D. melanogaster ADHS model was com-pletely satisfactory. Ramachandran plots were in accordancewith those of the template structure (D. lebanonensis), withonly Lys84 in the disallowed region in both cases, and the rmsdwas 0.1 AÊ for 508 Ca atoms, a figure which corresponds to thedimeric active form. Furthermore, no severe internal clasheswere detected after the last round of minimization.
D. melanogaster and D. lebanonensis differ in only 45positions (82.2% identity). It is no surprise that these changesdo no affect the catalytic residues, active site cavity and dimerinterface, as both enzymes share most of basic biochemical andkinetic features. A similar overall conformation relies onconservation of the total number of ion pairs, despite 12replacements altering the charge of a position and thehydrogen-bonding network, only two of the 31 substitutionswhich could alter it generate additional H-bonds in theD. melanogaster protein. Furthermore, in 73% of the sub-stitutions occurring in the core region, the local side-chain pairvolume is preserved, this percentage reaching 100% whenhydrophobic residues are considered.
Surprisingly, the substituted positions were not distributedrandomly or clustered in the connecting loops. Instead, theywere gathered at the exposed faces of the secondary structureelements, in helices and strands relatively distant from theactive site: aB, aC, bC, aD, bD, aG. Only two alterationscould bring about significant activity changes with respectto D. melanogaster: Ala13Gly, affecting one of the threeinvariant Gly residues of the coenzyme dinucleotide bindingregion, and Pro187Thr, presumably modifying the flexibility ofthe active-site closing loop corresponding to positions 186±191of the D. melanogaster ADH amino-acid sequence. It isreasonable to assume that even if these substitutions couldproduce a decrease in enzyme activity, the overall remainingchanges would help to balance this effect and restore thecatalytic efficiency to an intermediate level between ADHF andADHS of D. melanogaster.
D. melanogaster ADH allozymes:structure±function relationship
The absence of drastic differences in activity with secondaryalcohols between ADHS and ADHF forms led us to assume thatposition 192 should not have a direct involvement in thereaction mechanism, later corroborated through kinetic analysis[11±15], site-directed mutagenesis and protein alignments [5].From the structure, it is now possible to envisage the effect ofthe Thr/Lys substitution in the context of the general features ofthe molecule and relate it to activity changes. Epitope mappingof MMBB8 showed that it encompassed the differentialposition between ADHF and ADHS. Recognition and bindingto ADHF interfered with the activity of the enzyme and stressedthe contribution of this loop to the structural changes associatedwith catalysis. However, no significant alterations in the tertiarystructure were observed, either at the Ca main chain or at thesurrounding side chains when both models were compared.However, electrostatic potential determinations for the S, Fand UF allozymes, clearly showed correlation between the
electronegativity of the active site cavity (S , F , UF) andthe kcat values, from lower to higher: ADHS ,ADHF , ADHUF. Increasing the negative electrostatic poten-tial of the cavity should speed up the release of the reducedcoenzyme, which is also negatively charged. Therefore, thesedata provided the structural bases to understand the differencesin activity between allozymes, and agreed with the previousfinding that the rate-limiting step in the oxidation of secondaryalcohols by Drosophila ADH was the dissociation of the binarycomplex E´NADH [14].
In the case of primary alcohols, the rate-limiting stepinvolves the dissociation of the binary complex, as forsecondary alcohols, and the hydride transfer reaction. The kcat
value was much lower than that for secondary alcohols anddepended on the type of alcohol. The kcat with ethanol forADHS was half that of ADHF, following the same trend as forsecondary alcohols. Sequence replacements in the differentallozymes affected the electrostatic character of the active sitemore than its relative conformation. This could indicate that thelysine at position 192 in ADHS not only strengthened enzyme±NADH binding, but also decreased the rate of hydride transferfrom ethanol to the oxidized coenzyme NAD1. A slowerhydride transfer process due to a more positive surroundingcould be caused by a stabilized initial state, representing a morestable negatively charged (a possible candidate could beE´NAD1´EtO2) complex in the active site or by a destabilizedpositive transition state (of unknown nature).
Putative thermostability of the D. melanogaster ADHS
allozyme
From the structural analysis, no data were obtained which couldsupport higher thermostability of ADHS. The substitution ofThr192Lys did not generate any clear additional ionic orhydrogen bond. The neighboring residue with which Lys192could have established a new putative ion pair (Asp97) waslocated 9 AÊ away, clearly too distant (Fig. 6). In fact, nosignificant effect of the temperature on the kinetic constantswas found for all major ADH allozymes [53]. Therefore, themolecular bases of the claimed thermostability of the ADHS
allele [58] have not yet been elucidated. Nevertheless, recentnatural population surveys showed that selection is in factresponsible for the maintenance of the ADHF/ADHS poly-morphism but acting through other linked sites or, epistaticallyon the combination of the F/S site and an insertion/deletionpolymorphism in an intronic region of this gene [59].
General conclusions
The high correlation found between the active site electrostaticpotential differences and the kinetic parameters may allow us toelucidate the function±structure relationships in the mainallozymes of D. melanogaster ADH. From the overalldata, it seems plausible to assume that catalytic efficienciesfor D. melanogaster ADH allozymes, and even for the closelyrelated species D. simulans, are not modulated by individualamino-acid substitutions involved directly in coenzyme orsubstrate interaction or by minor conformational changes.Instead, it is replacements adjusting the electrostatic characterof the active site which specifically alter the magnitude of therate-limiting step of alcohol oxidation.
ADH has recently been associated with both the emergenceof the Drosophila genus, some 65 million years ago [60], andwith intergenic speciation [61]. Now two evolutionary patternsemerge for ADH: discrete amino-acid substitutions that affect
3620 J. Benach et al. (Eur. J. Biochem. 267) q FEBS 2000

the electrostatic potential distribution in the active site, gaverise to biochemically similar new forms, D. melanogasterADHS vs. ADHF (1.8 million years divergence time [62])and D. melanogaster ADHS vs. D. simulans (2.3 millionyears divergence time [63]). Or overall changes, such asthose reported in distant species, as D. lebanonensis andD. melanogaster (56 million years divergence time [64]), wereresponsible for modulation of the enzymatic activity.
Finally, it seems worth widening the Drosophila ADHscenario to other enzyme systems, mainly NAD1/NADP1-dependent dehydrogenases/reductases, to trace possiblesimilarities in their evolutionary patterns. Site-directed muta-genesis could be used to test such hypotheses and to obtain newenzymes with optimized catalytic efficiency.
A C K N O W L E D G E M E N T S
This work was financed by the Swedish Medical Research Council (MFR),
Spanish Ministerio de EducacioÂn y Ciencia grants (PB96-0220 and
UE98-0014) and the EC contract BIO4-CT97-2123. We thank J. Ll. GelpõÂ
for his constant and helpful advice about the insight Software Package.
R E F E R E N C E S
1. Chambers, G.K. (1988) The Drosophila alcohol dehydrogenase gene±
enzyme system. In Advances in Genetics (Caspari, E.W. &
Scandalios, J.G., eds), pp. 39±107. Academic Press, San Diego,
CA, USA.
2. Sullivan, D.T., Atkinson, P.W. & Starmer, W.T. (1990) Molecular
evolution of the alcohol dehydrogenase genes in the genus
Drosophila. In Evolutionary Biology (Hecht, M., Wallace, B. &
Prance, G., eds), pp. 107±147. Plenum Press, London, UK.
3. Lindsley, D.L. & Zimm, G.G. (1992) The Genome of Drosophila
melanogaster, pp. 16±21. Academic Press, San Diego, CA, USA.
4. Heinstra, P.W.H. (1993) Evolutionary genetics of the Drosophila
alcohol dehydrogenase gene±enzyme system. Genetica 92, 1±22.
5. JoÈrnvall, H., Persson, B., Krook, M., Atrian, S., GonzaÁlez-Duarte, R.,
Jeffery, J. & Ghosh, D. (1995) Short-chain dehydrogenase/reductases
(SDR). Biochemistry 34, 6003±6013.
6. JoÈrnvall, H., Danielsson, O., Eklund, H., Hjelmqvist, L., HoÈoÈg, J.O.,
PareÂs, X. & Shafqat, J. (1993) Enzyme and isozyme developments
within the medium-chain alcohol dehydrogenase family. In
Enzymology and Molecular Biology of Carbonyl Metabolism
(Weiner, H., Crabb, D.W., Flynn, T.G., eds), pp. 533±544. Plenum
Press, New York, USA.
7. Benach, J., Atrian, S., GonzaÁlez-Duarte, R. & Ladenstein, R. (1998)
The refined crystal structure of Drosophila lebanonensis alcohol
dehydrogenase at 1.9 AÊ resolution. J. Mol. Biol. 282, 383±399.
8. Benach, J., Atrian, S., GonzaÁlez-Duarte, R. & Ladenstein, R. (1999)
The catalytic reaction and inhibition mechanism of Drosophila
alcohol dehydrogenase. Observation of an enzyme-bound NAD-
ketone adduct at 1.4 AÊ resolution by X-ray crystallography. J. Mol.
Biol. 289, 335±355.
9. Oakeshott, J.G., Gibson, J.B., Anderson, P.R., Knibb, W.R., Anderson,
D.G. & Chambers, G.K. (1982) Alcohol dehydrogenase and glycerol-
3-phosphate dehydrogenase clines in Drosophila melanogaster of
different continents. Evolution 39, 86±96.
10. Bodmer, M. & Ashburner, M. (1984) Conservation and change in the
DNA sequence coding for alcohol dehydrogenase in sibling species
of Drosophila. Nature 309, 425±430.
11. Winberg, J.O., Thatcher, D.R. & McKinley-McKee, J. (1982) Alcohol
dehydrogenase from the fruitfly Drosophila melanogaster substrate
specificity of the alleloenzymes AdhS and AdhUF. Biochim. Biophys.
Acta 704, 7±16.
12. Winberg, J.O., Thatcher, D.R. & McKinley-McKee, J.S. (1982)
Alcohol dehydrogenase from the fruitfly Drosophila melanogaster
inhibition studies of the alleloenzymes AdhS and AdhUF. Biochim.
Biophys. Acta 704, 17±25.
13. Hovik, R., Winberg, J.O. & McKinley-McKee, J.S. (1984) Drosophila
melanogaster alcohol dehydrogenase. Substrate stereospecificity of
the AdhF alleloenzyme. Insect Biochem. 14, 345±351.
14. Winberg, J.O., Hovik, R., McKinley-McKee, J.S., Juan, E. & GonzaÁlez-
Duarte, R. (1986) Biochemical properties of alcohol dehydrogenase
from Drosophila lebanonensis. Biochem. J. 235, 481±490.
15. Winberg, J.O. & McKinley-McKee, J.S. (1992) Kinetic interpretations
of active site topologies and residue exchange in Drosophila alcohol
dehydrogenase. Int. J. Biochem. 24, 169±181.
16. Vriend, G. (1990) WHATIF: a molecular modelling and drug design
program. J. Mol. Graphics 8, 52±56.
17. Biosym/Molecular Simulations Inc. (1997) Insight Version 97.0.
18. BruÈnger, A.T. (1992) XPLOR manual, Version 3.1. Yale University,
New Haven, CT, USA.
19. Laskowski, R.A., MacArthur, M.W., Moss, D.S. & Thornton, J.M.
(1993) procheck: a program to check the stereochemical quality of
protein structures. J. Appl. Crystallogr. 26, 283±291.
20. Hutchinson, E.G. & Thornton, J.M. (1996) promotif: a program to
identify and analyze structural motifs in proteins. Protein Sci. 5,
212±220.
21. Nicholls, A., Sharp, K. & Honig, B. (1991) Protein folding and
association: insights from the interfacial and thermodynamic
properties of hydrocarbons. Proteins 11, 281±296.
22. Kraulis, P.J. (1991) molscript: a program to produce both detailed
and schematic plots of protein structures. J. Appl. Crystallogr. 24,
946±950.
23. Carson, M. (1987) Ribbon models of macromolecules. J. Mol.
Graphics 5, 103±106.
24. Fibla, J., Enjuanes, L. & GonzaÁlez-Duarte, R. (1989) Inter-specific
analysis of Drosophila alcohol dehydrogenase by an immuno-
enzymatic assay using monoclonal antibodies. Biochem. Biophysic.
Res. Commun. 160, 638±646.
25. Albalat, R., Valls, M., Fibla, J., Atrian, S. & GonzaÁlez-Duarte, R.
(1995) Involvement of the C-terminal tail in the activity of
Drosophila alcohol dehydrogenase. Eur. J. Biochem. 233, 498±505.
26. Visa, N., Fibla, J., Santa-Cruz, M.C. & GonzaÁlez-Duarte, R. (1992)
Developmental profile and tissue distribution of Drosophila alcohol
dehydrogenase: an immunochemical analysis with monoclonal
antibodies. J. Histochem. Cytochem. 40, 39±49.
27. Holm, L., Sander, C. & Murzin, A. (1994) Three sisters, different
names. Struct. Biol. 1, 146±147.
28. Ghosh, D., Weeks, C.M., Grochulski, P., Duax, W.L., Erman, M.,
Rimsay, R.L. & Orr, J.C. (1991) Three-dimensional structure of
holo 3a,20b-hydroxysteroid dehydrogenase: a member of a the
short-chain dehydrogenase family. Proc. Natl Acad. Sci. USA 88,
10064±10068.
29. Ghosh, D., Wawrzak, Z., Weeks, C.M., Duax, W.L. & Erman, M.
(1994) The refined three-dimensional structure of 3a,20b-hydro-
xysteroid dehydrogenase and possible roles of the residues conserved
in short-chain dehydrogenases. Structure 2, 629±640.
30. Ghosh, D., Erman, M., Wawrzak, Z., Duax, W.L. & Pangborn, W.
(1994) Mechanism of inhibition of 3a,20b-hydroxysteroid dehydro-
genase by a licorice-derived steroidal inhibitor. Structure 2, 973±980.
31. Ghosh, D., Pletnev, V., Zhu, D.W., Wawrzak, Z., Duax, W.L.,
Pangborn, W., Labrie, F. & Lin, S.X. (1995) Structure of human
estrogenic 17b-hydroxysteroid dehydrogenase at 2.20 AÊ resolution.
Structure 3, 503±513.
32. Bauer, A.J., Rayment, I., Frey, P.A. & Holden, H.M. (1992) The
molecular structure of UDP-galactose 4-epimerase from Escherichia
coli determined at 2.5 AÊ resolution. Proteins 12, 372±381.
33. Varughese, K.I., Skinner, M.M., Whiteley, J.M., Matthews, D.A. &
Xuong, N.H. (1992) Crystal structure of rat liver dihydropteridine
reductase. Proc. Natl Acad. Sci. USA 89, 6080±6084.
34. Su, Y., Varughese, K.I., Xuong, N.H., Bray, T.L., Roche, D.J. &
Whiteley, J.M. (1993) The crystallographic structure of a human
dihydropteridine reductase NADH binary complex expressed in
Escherichia coli by a cDNA constructed from its rat homologue.
J. Biol. Chem. 268, 26836±26841.
35. Rafferty, J.B., Simon, J.W., Baldock, C., Artymiuk, P.J., Baker, P.J.,
q FEBS 2000 Structure/function relationships in Drosophila ADH (Eur. J. Biochem. 267) 3621

Stuitje, A.R., Slabas, A.R. & Rice, D.W. (1995) Common themes in
redox chemistry emerge from the X-ray structure of oilseed rape
(Brassica napus) enoyl acyl carrier protein reductase. Structure 3,
927±938.
36. Andersson, A., Jordan, D., Schneider, G. & Lindqvist, Y. (1996) Crystal
structure of the ternary complex of 1,3,8-trihydroxysteroid naph-
thalene reductase from Magnaporthe grisea with NADPH and an
active-site inhibitor. Structure 4, 1161±1170.
37. Azzi, A., Rehse, P.H., Zhu, D.W., Campbell, R.L., Labrie, F. & Lin,
S.X. (1996) Crystal structure of human estrogenic 17b-hydroxyster-
oid dehydrogenase complexed with 17b-estradiol. Nat. Struct. Biol.
3, 665±668.
38. Breton, R., Housset, D., Mazza, C. & Fontecilla-Camps, J.C. (1996)
The structure of a complex of human 17b-hydroxysteroid dehydro-
genase with estradiol and NADP1 identifies two principal targets for
the design of inhibitors. Structure 4, 905±915.
39. Tanaka, N., Nonaka, T., Nakanishi, M., Deyashiki, Y., Hara, A. &
Mitsui, Y. (1996) Crystal structure of the ternary complex of mouse
lung carbonyl reductase at 1.8 AÊ resolution: the structural origin
of coenzyme specificity in the short-chain dehydrogenase/reductase
family. Structure 4, 33±45.
40. Tanaka, N., Nonaka, T., Tanabe, T., Yoshimoto, T., Tsuru, D. & Mitsui,
Y. (1996) Crystal structures of the binary and ternary complexes
of 7a-hydroxysteroid dehydrogenase from Escherichia coli. Bio-
chemistry 35, 7715±7730.
41. Thoden, J.B., Frey, P.A. & Holden, H.M. (1996) High-resolution
X-ray structure of UDP-galactose 4-epimerase complexed with
UDP-phenol. Protein Sci. 5, 2149±2161.
42. Auerbach, G., Herrmann, A., GuÈtlich, M., Fischer, M., Jacob, U.,
Bacher, A. & Huber, R. (1997) The 1.25 AÊ crystal structure of
sepiapterin reductase reveals its binding mode to pterins and brain
neurotransmitters. EMBO J. 16, 7219±7230.
43. HuÈlsmeyer, M., Hecht, H.J., Niefind, K., Hofer, B., Lindsay, D.E.,
Timmis, K.H. & Schomburg, D. (1998) Crystal structure of cis-
biphenyl-2,3-dihydrodiol-2,3-dehydrogenase from a PCB degrader at
2.0 AÊ resolution. Protein Sci. 7, 1286±1293.
44. Somers, W.S., Stahl, M.L. & Sullivan, F.X. (1998) GDP-fucose
synthetase from Escherichia coli: structure of a unique member of the
short-chain dehydrogenase/reductase family that catalyzes two
distinct reactions at the same active site. Structure 6, 1601±1612.
45. Genetics Computer Group (1994) GCG Program Manual for the
Wisconsin Package, Version 8. Genetics Computer Group, Madison,
WI, USA.
46. Albalat, R., Atrian, S. & GonzaÁlez-Duarte, R. (1994) Drosophila
lebanonensis ADH: analysis of recombinant wild-type enzyme and
site directed mutagenesis. The effect of restoring the consensus
sequence in two positions. FEBS Lett. 341, 171±176.
47. Brendskag, M.K., McKinley-McKee, J.S. & Winberg, J.-O. (1999)
Drosophila lebanonensis alcohol dehydrogenase: pH dependence of
the kinetic coefficients. Biochim. Biophys. Acta 1431, 74±86.
48. Heinstra, P.W., ThoÈrig, G.E.W., Scharloo, W., Drenth, W. & Nolte,
R.J.M. (1988) Kinetics and thermodynamics of ethanol oxidation
catalyzed by genetic variants of the alcohol dehydrogenase from
Drosophila melanogaster and D. simulans. Biochim. Biophys. Acta
967, 224±233.
49. Juan, E. & GonzaÁlez-Duarte, R. (1981) Determination of some
biochemical and structural features of alcohol dehydrogenase from
Drosophila simulans and Drosophila virilis. Biochem. J. 195, 61±69.
50. Retzios, A.D. (1982) Studies on Drosophila Adh. PhD Thesis,
University of Edinburgh, UK.
51. Winberg, J.O., Hovik, R. & McKinley-McKee, J.S. (1985) The alcohol
dehydrogenase alleloenzymes AdhS and AdhF from the fruitfly
Drosophila melanogaster: an enzymatic rate assay to determine the
active-site concentration. Biochem. Genet. 23, 205±216.
52. Winberg, J.O. & McKinley-McKee, J.S. (1988) The Adhs alleloenzyme
of alcohol dehydrogenase from Drosophila melanogaster. Biochem.
J. 255, 589±599.
53. McElfresh, K.C. & McDonald, J.F. (1986) The effect of temperature on
biochemical and molecular properties of Drosophila alcohol
dehydrogenase. Biochem. Genet. 24, 873±889.
54. Johnsen, F.M. & Denniston, C. (1964) Genetic variation of alcohol
dehydrogenase in Drosphilia melanogaster. Nature 204, 906±907.
55. Laurie-Ahlberg, C.C. (1985) Genetic variation affecting the expression
of enzyme-coding genes in Drosophila: an evolutionary perspective.
Isozymes Curr. Top. Biol. Med. Res. 12, 33±88.
56. Heinstra, P.W.H., Scharloo, W. & ThoÈrig, G.E.W. (1987) The
physiological significance of the alcohol dehydrogenase poly-
morphism in larvae of Drosophila. Genetics 117, 75±84.
57. Vigue, C.L. & Johnson, F.M. (1973) Isozyme variability in species
of the genus Drosophila. VI. Frequency±property±environment
relationships of allelic alcohol dehydrogenase in D. melanogaster.
Biochem. Genet. 9, 213±226.
58. Thatcher, D.R. & Seich, R. (1981) The relative conformational stability
of alcohol dehydrogenase allelozymes purified from Drosophila
melanogaster. Biochem. J. 197, 111±117.
59. Kreitman, M. & Berry, A. (1993) Molecular analysis of an allozyme
cline: alcohol dehydrogenase in Drosophila melanogaster of the east
coast of North America. Genetics 134, 869±893.
60. Ashburner, M. (1998) Speculations on the subject of alcohol
dehydrogenase and its properties in Drosophila and other flies.
Bioessays 20, 949±954.
61. Atrian, S., SaÂnchez-Pulido, L., GonzaÁlez-Duarte, R. & Valencia, A.
(1998) Shaping of Drosophila alcohol dehydrogenase through
evolution: relationship with enzyme functionality. J. Mol. Evol. 47,
211±221.
62. Cohn, V.H. & Moore, G.P. (1988) Organization and evolution of
the alcohol dehydrogenase gene in Drosophila. Mol. Biol. Evol.
5, 154±166.
63. Russo, C.A.M., Takezaki, N. & Nei, M. (1995) Molecular phylogeny
and divergence times of drosophilid species. Mol. Biol. Evol. 12,
391±404.
64. Kwiatowski, J., Skarecky, D., Bailey, K. & Ayala, F.J. (1994)
Phylogeny of Drosophila and related genera inferred from the
nucleotide sequence of the Cu, Zn Sod gene. J. Mol. Evol. 38,
443±454.
3622 J. Benach et al. (Eur. J. Biochem. 267) q FEBS 2000




















