
Structure and Expression of a Membrane Component of the Inhibin Receptor System
8
Structure and Expression of a Membrane Component of the Inhibin Receptor System* HUIRA CHONG, STEPHANIE A. PANGAS, DANIEL J. BERNARD, EILEEN WANG, JOLEE GITCH, WEI CHEN, LAWRENCE B. DRAPER, EDWARD T. COX, AND TERESA K. WOODRUFF Department of Neurobiology and Physiology (H.C., S.A.P., D.J.B., E.W., J.G., L.B.D., T.K.W.), Department of Biochemistry, Molecular Biology, and Cell Biology (W.C.), Northwestern University, Evanston, Illinois 60208-2850; and Genentech, Inc. (E.T.C.), South San Francisco, California 94080-4990 ABSTRACT The purification and cloning of a membrane-anchored proteoglycan with affinity for inhibin A are described. Bovine pituitary membranes were isolated, and membrane-anchored proteins were solubilized and used as an enriched source of inhibin binding protein. The extract was passed over an inhibin A affinity column, and a protein, designated p120, was identified as an inhibin-binding moiety. A partial amino acid sequence was determined for the protein, which matched two human complementary DNAs (cDNAs) in the database. The full- length cDNA predicts a 1336-amino acid glycoprotein. Full-length p120-encoding cDNAs were isolated from human testis RNA and cloned into expression vectors. Two p120 messenger RNA transcripts of 4.6 kb and 2 kb are detected in rat pituitary by RNA blot analysis. Similar analysis of rat testis RNA revealed transcripts of identical molecular mass, albeit at lower abundance. To determine the cellular localization of p120 in pituitary and testis, an antibody directed against the predicted extracellular domain of the protein was gen- erated and used in an immunohistochemical analysis of thin tissue sections. p120 immunostaining is coincident with FSHb immunopo- sitive gonadotrope cells in rat pituitary. p120 staining is intense in the testicular Leydig cells, which bind iodinated inhibin but not iodinated activin. In summary, an inhibin-binding protein has been isolated that is produced in tissues that are targets of inhibin action. (Endo- crinology 141: 2600 –2607, 2000) I N MAMMALS, the reproductive axis is dependent upon endocrine and paracrine acting hormones to regulate follicle maturation and ovulation in females and to maintain tonic sperm production in males. Inhibin is a gonadal- derived protein that is a critical feedback hormone between the gonads and the pituitary (1, 2). Despite the central nature of inhibin to the regulation of the reproductive axis, little is known regarding its molecular mechanism of action, largely because a receptor for this ligand has not been identified and cloned. Here we describe a membrane-associated protein that was isolated based on its affinity for inhibin and that is expressed in inhibin target cells. This novel protein may represent a component of an inhibin signal transduction system. Inhibin is a disulfide-linked, dimeric glycoprotein com- posed of an a-subunit and one of two b-subunits (2). Ho- modimers of the b-subunits form activin, a functional an- tagonist of inhibin. The inhibin b-subunit places that molecule into the larger transforming growth factor-b (TGFb) superfamily (3). Inhibin is distinct from most TGFb- like ligands because its a-subunit is capable of heterodimeric assembly (with activin b-subunits), but it is not able to ho- modimerize with unpaired a-subunits. Because inhibin is a member of the TGFb superfamily and antagonizes activin action, we hypothesized that some aspects of the cellular response pathways activated by this ligand would be similar to those activated by TGFb. Based on the unique structural nature of the inhibin a-subunit and the heterodimeric a-b ligand, we also predicted that novel components of the in- hibin signaling pathway exist. While little is known concerning the mechanism of inhibin action, the activin signaling pathway has been described. The cellular response to activin is transduced through two mem- brane-bound, serine-threonine kinases: a ligand binding re- ceptor subunit (ActRII) and a signal generating protein (ActRI) (4). ActRI subunits are phosphorylated by ligand- bound ActRII. Activated ActRI then phosphorylates cyto- plasmic Smad proteins (5). Three classes of Smad proteins have been identified. These include the receptor Smads, which are phosphorylated by activated type I receptors, the inhibitory Smads, which inhibit receptor-mediated Smad phosphorylation, and the common activator Smad known as Smad4 (5). Inhibin inhibits pituitary FSH synthesis and release and stimulates Leydig and theca cell androgen production (1, 2, 6, 7). One hypothesis regarding inhibin action is that inhibin binds and inactivates the ActRII subunit, thereby antagoniz- ing activin action (8, 9). The association of inhibin with ActRII subunits blocks ActRI subunit phosphorylation and uncou- ples the activin signal transduction pathway (8, 9). Thus, when the inhibin concentration exceeds activin in a target cell, inhibin can act to disengage the activin signal pathway by competitive inhibition of the activin-binding receptor sub- Received November 9, 1999. Address all correspondence and requests for reprints to: Teresa K. Woodruff, Ph.D., Northwestern University, Department of Neurobiol- ogy and Physiology, O.T. Hogan 4 –150, 2153 North Campus Drive, Evanston, Illinois 60208. E-mail: [email protected]. * This study was supported by NIH Grants HD-37096-01 and HD-28048-01 (to T.K.W.); an American Cancer Society Grant, IL Division (96-17) (to T.K.W.), and NIH Grant HD-21921 (to W.C. and Kelly E. Mayo). 0013-7227/00/$03.00/0 Vol. 141, No. 7 Endocrinology Printed in U.S.A. Copyright © 2000 by The Endocrine Society 2600
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Structure and Expression of a Membrane Component of the Inhibin Receptor System

Structure and Expression of a Membrane Component ofthe Inhibin Receptor System*
HUIRA CHONG, STEPHANIE A. PANGAS, DANIEL J. BERNARD, EILEEN WANG,JOLEE GITCH, WEI CHEN, LAWRENCE B. DRAPER, EDWARD T. COX, AND
TERESA K. WOODRUFF
Department of Neurobiology and Physiology (H.C., S.A.P., D.J.B., E.W., J.G., L.B.D., T.K.W.),Department of Biochemistry, Molecular Biology, and Cell Biology (W.C.), Northwestern University,Evanston, Illinois 60208-2850; and Genentech, Inc. (E.T.C.), South San Francisco, California94080-4990
ABSTRACTThe purification and cloning of a membrane-anchored proteoglycan
with affinity for inhibin A are described. Bovine pituitary membraneswere isolated, and membrane-anchored proteins were solubilized andused as an enriched source of inhibin binding protein. The extract waspassed over an inhibin A affinity column, and a protein, designatedp120, was identified as an inhibin-binding moiety. A partial aminoacid sequence was determined for the protein, which matched twohuman complementary DNAs (cDNAs) in the database. The full-length cDNA predicts a 1336-amino acid glycoprotein. Full-lengthp120-encoding cDNAs were isolated from human testis RNA andcloned into expression vectors. Two p120 messenger RNA transcripts
of 4.6 kb and 2 kb are detected in rat pituitary by RNA blot analysis.Similar analysis of rat testis RNA revealed transcripts of identicalmolecular mass, albeit at lower abundance. To determine the cellularlocalization of p120 in pituitary and testis, an antibody directedagainst the predicted extracellular domain of the protein was gen-erated and used in an immunohistochemical analysis of thin tissuesections. p120 immunostaining is coincident with FSHb immunopo-sitive gonadotrope cells in rat pituitary. p120 staining is intense in thetesticular Leydig cells, which bind iodinated inhibin but not iodinatedactivin. In summary, an inhibin-binding protein has been isolatedthat is produced in tissues that are targets of inhibin action. (Endo-crinology 141: 2600–2607, 2000)
IN MAMMALS, the reproductive axis is dependent uponendocrine and paracrine acting hormones to regulate
follicle maturation and ovulation in females and to maintaintonic sperm production in males. Inhibin is a gonadal-derived protein that is a critical feedback hormone betweenthe gonads and the pituitary (1, 2). Despite the central natureof inhibin to the regulation of the reproductive axis, little isknown regarding its molecular mechanism of action, largelybecause a receptor for this ligand has not been identified andcloned. Here we describe a membrane-associated proteinthat was isolated based on its affinity for inhibin and that isexpressed in inhibin target cells. This novel protein mayrepresent a component of an inhibin signal transductionsystem.
Inhibin is a disulfide-linked, dimeric glycoprotein com-posed of an a-subunit and one of two b-subunits (2). Ho-modimers of the b-subunits form activin, a functional an-tagonist of inhibin. The inhibin b-subunit places thatmolecule into the larger transforming growth factor-b(TGFb) superfamily (3). Inhibin is distinct from most TGFb-like ligands because its a-subunit is capable of heterodimericassembly (with activin b-subunits), but it is not able to ho-modimerize with unpaired a-subunits. Because inhibin is a
member of the TGFb superfamily and antagonizes activinaction, we hypothesized that some aspects of the cellularresponse pathways activated by this ligand would be similarto those activated by TGFb. Based on the unique structuralnature of the inhibin a-subunit and the heterodimeric a-bligand, we also predicted that novel components of the in-hibin signaling pathway exist.
While little is known concerning the mechanism of inhibinaction, the activin signaling pathway has been described. Thecellular response to activin is transduced through two mem-brane-bound, serine-threonine kinases: a ligand binding re-ceptor subunit (ActRII) and a signal generating protein(ActRI) (4). ActRI subunits are phosphorylated by ligand-bound ActRII. Activated ActRI then phosphorylates cyto-plasmic Smad proteins (5). Three classes of Smad proteinshave been identified. These include the receptor Smads,which are phosphorylated by activated type I receptors, theinhibitory Smads, which inhibit receptor-mediated Smadphosphorylation, and the common activator Smad known asSmad4 (5).
Inhibin inhibits pituitary FSH synthesis and release andstimulates Leydig and theca cell androgen production (1, 2,6, 7). One hypothesis regarding inhibin action is that inhibinbinds and inactivates the ActRII subunit, thereby antagoniz-ing activin action (8, 9). The association of inhibin with ActRIIsubunits blocks ActRI subunit phosphorylation and uncou-ples the activin signal transduction pathway (8, 9). Thus,when the inhibin concentration exceeds activin in a targetcell, inhibin can act to disengage the activin signal pathwayby competitive inhibition of the activin-binding receptor sub-
Received November 9, 1999.Address all correspondence and requests for reprints to: Teresa K.
Woodruff, Ph.D., Northwestern University, Department of Neurobiol-ogy and Physiology, O.T. Hogan 4–150, 2153 North Campus Drive,Evanston, Illinois 60208. E-mail: [email protected].
* This study was supported by NIH Grants HD-37096-01 andHD-28048-01 (to T.K.W.); an American Cancer Society Grant, IL Division(96-17) (to T.K.W.), and NIH Grant HD-21921 (to W.C. and Kelly E. Mayo).
0013-7227/00/$03.00/0 Vol. 141, No. 7Endocrinology Printed in U.S.A.Copyright © 2000 by The Endocrine Society
2600

unit. Because the affinity of inhibin for ActRII is approxi-mately 10-fold lower than activin for ActRII, this mechanismof action is plausible when inhibin is in excess of the localactivin signal.
In addition to the dominant-negative model of inhibin-mediated activin antagonism, strong evidence exists to sup-port an inhibin-specific receptor or inhibin accessory pro-tein(s) that mediate an inhibin-specific signal. The mostcompelling evidence supporting an independent inhibin re-ceptor is the fact that inhibin is not able to antagonize all ofactivin’s actions. For example, inhibin does not antagonizeactivin-stimulated liver cell apoptosis (10) nor does itblock activin-induced animal cap development (M. Whit-man, personal communication). Further, inhibin and activinboth promote oocyte maturation (11) and Leydig cell P45017a
messenger RNA (mRNA) accumulation (6). These experi-mental observations suggest that a unique inhibin-specificsignal transduction pathway may exist. Alternatively, aninhibin-binding protein may act in concert with activin re-ceptor components to permit the antagonistic as well ascooperative relationship between the two ligands. These twomechanisms are not mutually exclusive, and cells may havethe ability to use inhibin in a number of different ways tomodify an activin signal.
Identifying an inhibin-binding and signal-transducingmolecule will increase knowledge about the normal repro-ductive cycle and will lead to a more complete understand-ing of diseases in which inhibin-signaling cascades may beabnormally regulated. Toward this end, the present manu-script describes efforts to isolate inhibin-binding proteins.The bovine pituitary was used as a starting material to isolateand characterize an inhibin- binding protein based on phys-iological data (inhibin inhibits bovine pituitary FSH secre-tion), direct binding data, and the availability of largeamounts of tissue (12–14). The purification strategy imple-mented in the present study was based on a method devel-oped to isolate inhibin-binding proteins from the gonadaltumors of inhibin a-subunit knockout mice (15). A proteinthat binds inhibin and is expressed in inhibin target tissue,was isolated and cloned, and its characteristics are described.
Materials and MethodsRecombinant ligands
Recombinant human (rh) inhibin A (90B; NU35–45, Genentech, Inc.,South San Francisco, CA) and rh-activin A (NU43–40) were producedin the laboratory. The ligands were formulated in a buffer of 0.15 m NaCland 0.05 m Tris, pH 7.4.
Ligand affinity chromatography and microsequencing
Inhibin A was coupled to two support matrices. Inhibin was coupledto cross-linked agarose (Affi Gel 10) through free amino groups ofinhibin according to the manufacturer’s instructions (Bio-Rad Labora-tories, Inc., Richmond, CA). Inhibin was also coupled to Affi Gel HZthrough carbohydrate groups on the inhibin a-subunit. A blank columnwas prepared by immobilizing Tris to Affi Gel 10; 500 g of bovinepituitaries were suspended in 150 mm NaCl with 1 mm EDTA, 25 mg/mlaprotinin, 0.5 mg/ml leupeptin, and 0.7 mg/ml pepstatin and disruptedin a Waring blender. The pituitary mixture was stirred at 5C for 1 h andthen centrifuged at 10,000 3 g to remove particulate matter. The result-ing supernatant was centrifuged at 105,000 3 g for 3 h. The ultracen-trifugation pellet was resuspended in 85 mm Tris, pH 7.8, 0.1% octyl bglucoside (ObG), 30 mm NaCl, 1 mm EDTA, and 25 mg/ml aprotinin. The
sample was treated with 1 U/mg DNase (Roche Molecular Biochemicals,Indianapolis, IN) overnight to reduce viscosity and improve sedimen-tation of particulate matter during ultracentrifugation. DNase-treatedsamples were sedimented at 105,000 3 g for 3 h, and clear red super-natants were removed and sedimented again at 105,000 3 g for 3 h. Theaffinity columns were washed extensively with 25 mm Tris, pH 7.5, 150mm NaCl, and 0.1% ObG. The membrane proteins were eluted from thecolumns with 100 mm glycine, pH 3, and 0.1% ObG. The eluents wereneutralized with 3 m Tris, pH 8. Addition of excess inhibin A to themembrane-solubilized proteins resulted in inhibition of one band of 120kDa from the Affi Gel 10 column. The protein with molecular mass of120 kDa, which will be referred to as p120, was enriched and purifiedto homogeneity using reverse-phase HPLC (15 mm bead diameter C4resin in a 2-cm column, 0–70% n-propanol/trifluoroacetic acid gradi-ent). The receptor-enriched reverse-phase HPLC fractions were run onreduced 10% SDS-PAGE, transferred to polyvinylidenefluoride (PVDF),and stained with Coomassie blue. The single protein band at 120 kDabelieved to be an inhibin-binding protein was cut out of the transblot andsequenced. This protein was purified only when inhibin was coupledthrough amines and not when the carbohydrate moieties of the inhibina-subunit were used in the coupling chemistry. These data suggest thatthe column purification was specific for binding sites that were blockedwhen the space between the a-subunit (the only subunit that is glyco-sylated) and the inhibin-binding protein was hindered.
p120 complementary DNA (cDNA) cloning
Internal sequence for the isolated protein was obtained from theGenentech microsequencing facility (Genentech, Inc., South San Fran-cisco, CA). Two sequences were identical to the peptide sequence andwere similar to each other [accession numbers: Y10523 (IGDC1) andAF034198 (IGSF1)]. A cDNA was amplified by PCR from human testescDNA (CLONTECH Laboratories, Inc., Palo Alto, CA) giving a full-length Y10523 cDNA. PCR errors, detected by sequence analysis, werecorrected to match Y10523.
Northern blot analysis
RNA was size separated on a formaldehyde-agarose gel and trans-ferred to a nylon filter by capillary action. The filter was hybridizedusing a 32P-labeled rat cDNA probe corresponding to part of the extra-cellular domain of the rat p120 cDNA. After stringent washes, the blotwas exposed to x-ray film for 2 days.
Tissue distribution analysis
In situ ligand binding was performed as described previously (16).Briefly, 12-mm cryocut tissue sections were incubated for 3 h at roomtemperature in blocking buffer: DMEM:F12 (1:1), 20 mm HEPES, 0.05%cytochrome C, 0.3% BSA, 0.01 mg/ml phenylmethylsulfonylfluoride,0.01% bacitracin, 0.4 mg/ml leupeptin. Slides were then incubated atroom temperature overnight in the same buffer containing 40 pm 125I-rhinhibin A, activin A, or in the presence of 40 nm excess homologousligand (to define nonspecific background) or heterologous ligand. Theligands were iodinated using a modified lactoperoxidase method.Briefly, 5 mg of ligand were diluted in 0.4 m sodium acetate, pH 5.6, and0.5 nmol Na125I (0.5 nmol/mCi on calibration date), 0.5 U lactoperox-idase, and 0.25 nmol H2O2 were added sequentially. The ligand wasincubated at ambient temperature with intermittent vortexing for 5 min.The reaction was quenched with 450 ml PBS 1 0.05% Tween 20 1 0.5%BSA (Intergen, Purchase, NY). A 10 ml aliquot of the precolumn fractionwas removed for trichloroacetic acid precipitation. Free iodine wasremoved using Sephadex G-10 column chromatography (PD-10, Phar-macia Biotech, Piscataway NJ). The specific activity of the ligands usedin the in situ ligand-binding studies was approximately 100 mCi/mg. Theslides were subsequently washed in PBS (two times for 1 sec), PBS (twotimes for 10 min), and fixed in 3.7% formalin, 2% glutaraldehyde (10min); rinsed in water (four times for 1 sec)]; and allowed to dry. Dryslides were exposed to x-ray film for 1–14 days and then dipped in NTB-3emulsion (Eastman Kodak Co., Rochester, NY).
Immunohistochemical studies were done using an antibody directedagainst the protein sequence determined for the bovine p120 protein.The bovine and human epitopes are identical; however, 4 of 17 aminoacids (aa) differ between the human and rat epitope. Paraffin-embedded
IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN 2601
testicular sections from adult rats were the gift of Dr. Erwin Goldberg,Northwestern University. The p120 polyclonal primary antibody wasdiluted in PBS (1:500), and the sections were incubated overnight at roomtemperature, washed, incubated with a biotinylated goat-antirabbit sec-ondary antibody (Vector Laboratories, Inc., Burlingame, CA), washed,and incubated in ABC reagent (Vectastain ABC kit, Vector Laboratories,Inc.). The signal was detected using the DAB substrate kit (Vector Lab-oratories, Inc.). Sections were counterstained with hematoxylin. Controlsections were not incubated in the primary antibody and did not havedetectable brown stain (data not shown). The specificity of the antibodywas confirmed by preabsorption of the antibody with recombinant p120(100 mg of protein of a transient transfection extract) or oligopeptide (ata final concentration of 1 mg/ml). Tissue sections were analyzed usinga Eclipse E600 microscope (Nikon, Melville, NY).
ResultsPurification of an inhibin-binding protein
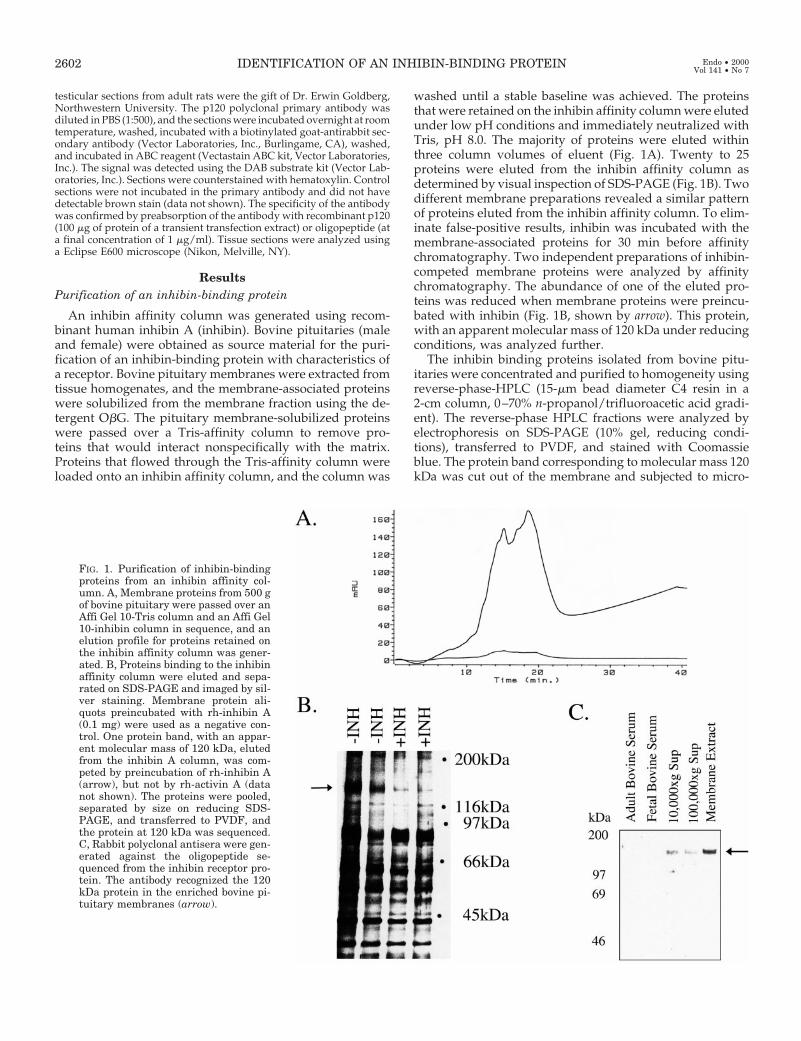
An inhibin affinity column was generated using recom-binant human inhibin A (inhibin). Bovine pituitaries (maleand female) were obtained as source material for the puri-fication of an inhibin-binding protein with characteristics ofa receptor. Bovine pituitary membranes were extracted fromtissue homogenates, and the membrane-associated proteinswere solubilized from the membrane fraction using the de-tergent ObG. The pituitary membrane-solubilized proteinswere passed over a Tris-affinity column to remove pro-teins that would interact nonspecifically with the matrix.Proteins that flowed through the Tris-affinity column wereloaded onto an inhibin affinity column, and the column was
washed until a stable baseline was achieved. The proteinsthat were retained on the inhibin affinity column were elutedunder low pH conditions and immediately neutralized withTris, pH 8.0. The majority of proteins were eluted withinthree column volumes of eluent (Fig. 1A). Twenty to 25proteins were eluted from the inhibin affinity column asdetermined by visual inspection of SDS-PAGE (Fig. 1B). Twodifferent membrane preparations revealed a similar patternof proteins eluted from the inhibin affinity column. To elim-inate false-positive results, inhibin was incubated with themembrane-associated proteins for 30 min before affinitychromatography. Two independent preparations of inhibin-competed membrane proteins were analyzed by affinitychromatography. The abundance of one of the eluted pro-teins was reduced when membrane proteins were preincu-bated with inhibin (Fig. 1B, shown by arrow). This protein,with an apparent molecular mass of 120 kDa under reducingconditions, was analyzed further.
The inhibin binding proteins isolated from bovine pitu-itaries were concentrated and purified to homogeneity usingreverse-phase-HPLC (15-mm bead diameter C4 resin in a2-cm column, 0–70% n-propanol/trifluoroacetic acid gradi-ent). The reverse-phase HPLC fractions were analyzed byelectrophoresis on SDS-PAGE (10% gel, reducing condi-tions), transferred to PVDF, and stained with Coomassieblue. The protein band corresponding to molecular mass 120kDa was cut out of the membrane and subjected to micro-
FIG. 1. Purification of inhibin-bindingproteins from an inhibin affinity col-umn. A, Membrane proteins from 500 gof bovine pituitary were passed over anAffi Gel 10-Tris column and an Affi Gel10-inhibin column in sequence, and anelution profile for proteins retained onthe inhibin affinity column was gener-ated. B, Proteins binding to the inhibinaffinity column were eluted and sepa-rated on SDS-PAGE and imaged by sil-ver staining. Membrane protein ali-quots preincubated with rh-inhibin A(0.1 mg) were used as a negative con-trol. One protein band, with an appar-ent molecular mass of 120 kDa, elutedfrom the inhibin A column, was com-peted by preincubation of rh-inhibin A(arrow), but not by rh-activin A (datanot shown). The proteins were pooled,separated by size on reducing SDS-PAGE, and transferred to PVDF, andthe protein at 120 kDa was sequenced.C, Rabbit polyclonal antisera were gen-erated against the oligopeptide se-quenced from the inhibin receptor pro-tein. The antibody recognized the 120kDa protein in the enriched bovine pi-tuitary membranes (arrow).
2602 IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN Endo • 2000Vol 141 • No 7

sequence analysis. An oligopeptide sequence was deter-mined to be 59-VLIEETEIVMPTPKPEL-39. Using the peptidesequence, polyclonal antisera were raised and purified usingan immobilized protein A/G column (Pierce Chemical Co.,Rockford, IL). The polyclonal antisera detected a single pro-tein band at molecular mass 120 kDa in membrane protein-enriched extracts from bovine pituitaries (Fig. 1C).
cDNA cloning of p120
The oligopeptide sequence derived from microsequenceanalysis was used to search NCBI-GenBank and was foundto match a partial sequence of a human cDNA isolated by twoindependent groups [accession numbers Y10523 andAF034198 (IGSF1)] (Fig. 2A; Refs. 17 and 18). The humanp120 protein has five predicted extracellular N-linked gly-cosylation sites and a large putative extracellular domain(1234 aa in length) followed by a single, hydrophobic, po-tential transmembrane domain and a short cytoplasmic tail(53 aa) that is 19% serine and threonine. The extracellulardomain contains a series of Ig-like domains. Using the Gen-Bank match (Y10523), primers were designed that containedconvenient unique internal restriction sites, and a cDNA wasamplified from human testis cDNA by PCR. The cDNAswere ligated together to form the full length 4.01-kb codingregion encoding a predicted protein of 1336 aa. Subse-quently, partial sequence was obtained for a rat ortholog byscreening a testis cDNA library. Tissue expression and abun-dance of p120 mRNA in male rat pituitary, testis, and liverwere examined (Fig. 2B). Northern blot analysis revealed twomajor transcripts of approximately 2.0 and 4.6 kb, corre-sponding to transcripts of similar sizes detected in humantestis (data not shown).
Tissue distribution of p120
To determine whether p120 colocalizes with FSH-produc-ing gonadotropes, anterior pituitaries from castrate male ratswere examined by immunohistochemistry using the p120polyclonal antibody. Gonadotropes, delineated by FSHb-subunit antibody staining, were abundant in castrate pitu-itaries and p120 immunostaining colocalized with FSHb-subunit immunostaining (Fig. 3, A and B).
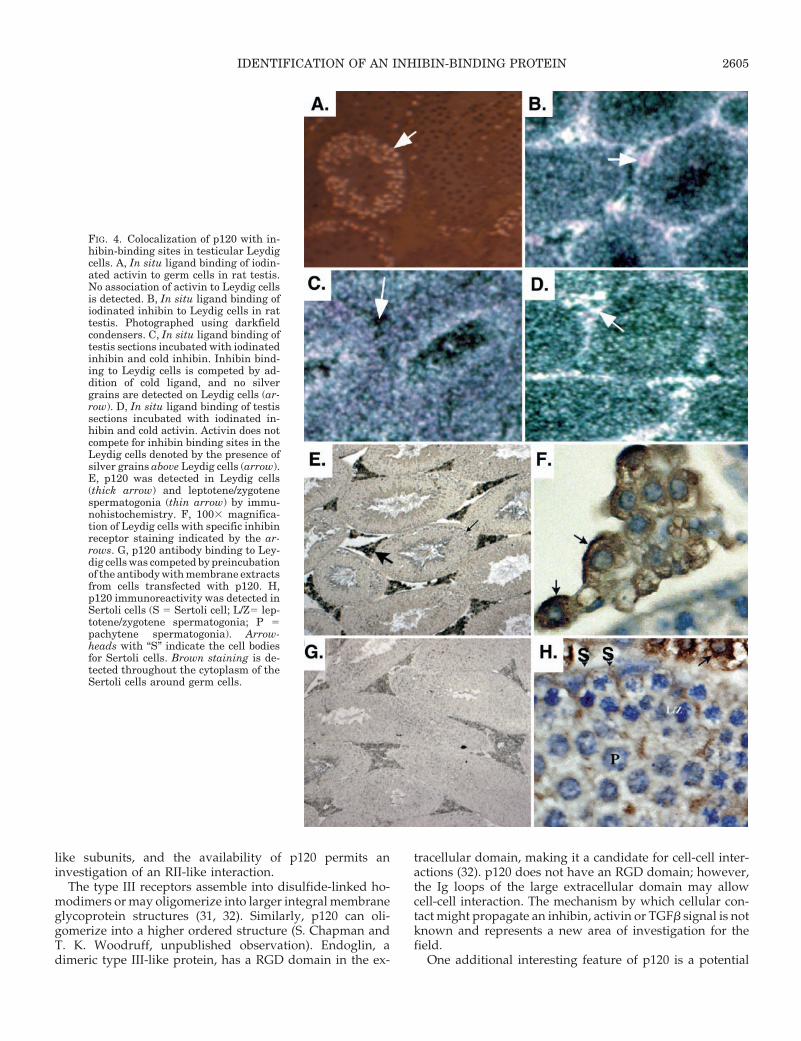
Nonoverlapping inhibin- and activin-binding sites arepresent in the testis (Ref. 19 and Fig. 4, A and B). Activinspecifically associated with testicular germ cells while in-hibin-specific binding sites were detected in Leydig cells (Fig.4, A–D). Low-level inhibin binding was detected in Sertolicells. p120 protein was detected primarily in the Leydig cellby immunohistochemical analysis of thin tissue sections de-rived from adult rat testis (Fig. 4, E and F). The positive signalwas competed by cellular extracts from p120 transfected cells(Fig. 4G) or with the oligopeptide (data not shown), indi-cating that the antibody staining is specific for p120. LongSertoli cell cytoplasmic processes are positive and can bedetected throughout the interior portion of all seminiferoustubules (Fig. 3H). Thus, p120 colocalizes with inhibin-bind-ing sites in the testis.
Discussion
The biological action of inhibin on bovine pituitary cells isto suppress FSH synthesis and secretion (12). Previous bind-ing studies using bovine pituitary membranes demonstratedhigh-affinity binding sites as did initial attempts to showinhibin binding complexes in crude bovine pituitary mem-brane extracts (Ref. 13, and our unpublished observation).Therefore, based on physiological data and direct binding
FIG. 2. Oligopeptide sequence of p120 and expression of p120 mRNA in rat tissues. A, Sequence from NCBI-GenBank match Y10523 spanningexons 9–12 including the linker region and Ig domain 6 is shown. The oligopeptide sequence obtained from microsequencing of the 120-kDainhibin-binding protein is represented by upper case letters and maps to base pairs 1812–1863 in GenBank match accession number Y10523.This region is located within the linker region (red outline) and slightly overlaps Ig domain 6 (blue outline). B, p120 expression was examinedin total RNA (20 mg per lane) extracted from adult male rat testis, pituitary, and liver by Northern blot analysis. Poly A1 RNA (5 mg) purifiedfrom rat testis total RNA was also included in the analysis. The results show a high level of p120 expression in rat pituitary. The mRNA isalso expressed, albeit to a lesser degree, in rat testis. Two major transcripts of approximately 2.0 and 4.6 kb were detected in testes and pituitarytotal and polyA1 RNA. p120 is not expressed at detectable levels in liver-derived mRNA. RNA size standards are indicated at the left.
IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN 2603

data, kilogram quantities of bovine pituitary were used as asource for inhibin receptor purification. Twenty to 25 pro-teins were eluted from an inhibin affinity column. A proteinwith a molecular mass of 120 kDa was selected for furtheranalysis because the interaction of this protein with thecolumn-linked inhibin could be blocked by preincubation ofmembrane protein homogenates with inhibin A. This resultsuggested that the interaction of p120 with inhibin was spe-cific and of high affinity.
p120 was not eluted from the inhibin column when thecoupling chemistry was through the carbohydrate moietiesof the a-subunit. This result suggests that the interactingportion of inhibin for p120 is related to the a-subunit. Thea-subunit has limited sequence similarity to any other TGFbligand, including activin, and is distinct because it is capableof heterodimeric assembly with activin b-subunit but willnot homodimerize (20). Other ligand subunits with distanthomology to the TGFb core ligands include Mullerian-inhibiting substance (whose receptor is similar to the TGFbreceptor structure), growth differentiation factor-9 (GDF-9,whose receptor has not been identified), and glial-derivednerve growth factor (GDNF) (whose receptor includes a gly-cosylphosphatidyl inositol-anchored binding protein thatpresents the ligand to the Ret tyrosine kinase receptor) (21–23). An antibody against the inhibin a-subunit N-terminal1–30 aa bioneutralizes inhibin action in vivo (24). Taken to-gether, these data imply that the a-subunit N-terminal 1–30peptide may serve as a ligand-interacting domain.
Binding assays for inhibin are difficult due to the poorquality of the iodinated ligand. Despite this well documentedimpediment (14), inhibin iodinated using lactoperoxidase iscapable of suppressing pituitary FSH, suggesting that bind-ing studies using modified (iodinated) ligand can bind andact upon receptor proteins. A second challenge in the bindingstudies is that inhibin may interact with numerous surfacereceptors with different relative affinities. The activin type IIor type IIB receptor subunits, follistatin, or a2-macroglobulinall bind inhibin, and the relative abundance of each of theseproteins may affect inhibin binding to its receptor (25–27).None of these potential inhibin-binding proteins were pu-rified from bovine pituitary homogenates; however, p120was efficiently purified. This result suggests that the successof isolating p120 relative to the other molecules is due pri-
marily to the high-affinity interaction of p120 and inhibin A.Thus, in tissues where p120 is expressed, interaction of in-hibin A with p120 is predicted to exceed interactions with theother proteins. Previous binding studies using bovine pitu-itary cells revealed inhibin-specific binding sites with anapparent dissociation constant (Kd) of 1.2 nm (13) while ovinepituitary cells had two inhibin-specific binding sites with Kd
values of 0.28 nm and 3.9 nm (14) consistent with our hy-pothesis. The p120 molecule was purified from bovine pi-tuitary membranes based on its ability to bind inhibin A.Inhibin B may also bind p120, but insufficient purified re-combinant protein is available to determine the relative af-finity of this isoform for p120.
Importantly, nonoverlapping inhibin- and activin-bindingsites are found in the testis. Activin binds germ cells whileinhibin binds Leydig cells, and the predominant site of p120production is the Leydig cell. This cellular localization sug-gests that inhibin and activin action are compartmentalizedby various binding proteins such as p120 and ActRII. Fur-ther, the two binding compartments may permit differentialaction of inhibin and activin within the testis. Additionalstudies will be required to determine whether the inhibin-binding protein has a function and whether that function isrestricted to inhibin alone or whether there is an interactionwith other TGFb ligands.
The p120 Ig-like domains bear structural resemblance toother cytokine receptors (28) and some adhesion molecules(29). p120 bears no sequence or structural homology to thecanonical TGFb-family type II or type I receptor subunits (4,30). It is not surprising that p120 represents a departure fromthe previously described TGFb family of receptor subunits inlight of the unsuccessful attempts to clone an inhibin receptoror inhibin-binding proteins based on homology to these pro-teins (15). p120 appears to be a unique and novel addition tothe family of cell surface receptor molecules that associatewith TGFb-like ligands.
The TGFb or activin type III receptor is also dissimilarfrom other cytokine receptors and bears no sequence orstructural homology to the canonical type II or type I recep-tors (31, 32). The TGFb type III receptor and accessory proteinendoglin are large proteins with no consensus signal trans-duction motif. There is no sequence similarity between thesereceptors and p120; however, p120 may act as a type III-likemolecule in regulating inhibin action. For example, fiveTGFb isoforms are synthesized by various cells, and thepredominant isoforms found in most tissues are TGFb1 andTGFb2 (33). TGFb1 binds directly to type II and activates typeI; however, TGFb2 requires an additional ancillary protein,a single membrane-spanning, nonsignaling protein (type III),that presents TGFb2 to the type II-type I complex (34). Anactivin type III receptor has been identified in endothelialcells and can be assembled with activin A into a type III-typeII-type I complex (35). Recent data in the K562 hematopoieticcell line are suggestive of an ancillary protein requirement forinhibin function (36). Ancillary proteins, such as p120, mayassemble complexes of activin type II and type I receptorsinto a configuration that is not only antagonistic to an in-trinsic activin signal, but also promotes inhibin signal trans-duction. Alternatively, there may exist inhibin-specific RII-
FIG. 3. Colocalization of p120 with FSHb-subunit in pituitary gonad-otropes. A, Immunohistochemistry of castrate male rat pituitary dem-onstrating the increased staining for FSH b-subunit and and coinci-dent staining of p120 (arrow). B, Magnification of the cellularmembrane staining for p120 in the castrate male rat pituitary.
2604 IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN Endo • 2000Vol 141 • No 7
like subunits, and the availability of p120 permits aninvestigation of an RII-like interaction.
The type III receptors assemble into disulfide-linked ho-modimers or may oligomerize into larger integral membraneglycoprotein structures (31, 32). Similarly, p120 can oli-gomerize into a higher ordered structure (S. Chapman andT. K. Woodruff, unpublished observation). Endoglin, adimeric type III-like protein, has a RGD domain in the ex-
tracellular domain, making it a candidate for cell-cell inter-actions (32). p120 does not have an RGD domain; however,the Ig loops of the large extracellular domain may allowcell-cell interaction. The mechanism by which cellular con-tact might propagate an inhibin, activin or TGFb signal is notknown and represents a new area of investigation for thefield.
One additional interesting feature of p120 is a potential
FIG. 4. Colocalization of p120 with in-hibin-binding sites in testicular Leydigcells. A, In situ ligand binding of iodin-ated activin to germ cells in rat testis.No association of activin to Leydig cellsis detected. B, In situ ligand binding ofiodinated inhibin to Leydig cells in rattestis. Photographed using darkfieldcondensers. C, In situ ligand binding oftestis sections incubated with iodinatedinhibin and cold inhibin. Inhibin bind-ing to Leydig cells is competed by ad-dition of cold ligand, and no silvergrains are detected on Leydig cells (ar-row). D, In situ ligand binding of testissections incubated with iodinated in-hibin and cold activin. Activin does notcompete for inhibin binding sites in theLeydig cells denoted by the presence ofsilver grains above Leydig cells (arrow).E, p120 was detected in Leydig cells(thick arrow) and leptotene/zygotenespermatogonia (thin arrow) by immu-nohistochemistry. F, 1003 magnifica-tion of Leydig cells with specific inhibinreceptor staining indicated by the ar-rows. G, p120 antibody binding to Ley-dig cells was competed by preincubationof the antibody with membrane extractsfrom cells transfected with p120. H,p120 immunoreactivity was detected inSertoli cells (S 5 Sertoli cell; L/Z5 lep-totene/zygotene spermatogonia; P 5pachytene spermatogonia). Arrow-heads with “S” indicate the cell bodiesfor Sertoli cells. Brown staining is de-tected throughout the cytoplasm of theSertoli cells around germ cells.
IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN 2605

glycosyl phosphatidylinositol (GPI)-linkage domain. GDNF,neurturin, and artemin are members of a new family offactors within the TGFb superfamily that use accessory re-ceptor proteins for activity (37). Ligand binds a glycosylphosphatidylinositol (GPI)-linked protein called GDNF fam-ily receptor as (GFRa-1 and GFRa-2) (38). These proteinsthen associate with the Ret tyrosine kinase (39). To date, theGDNF-like ligands are the only TGFb superfamily ligandsthat do not use serine-threonine kinase-based receptors andtherefore represent a separate subclass of ligands within thelarger family. Because p120 does not have intrinsic kinaseactivity, it is likely that this ligand-binding protein associateswith other signal-transducing proteins. The interacting pro-teins may include ActRII, ActRI, a type III-like molecule, ora tyrosine-kinase signaling molecule. Each of these possibil-ities will be investigated and should provide new insight intothe mode of inhibin action.
p120 is produced by the pituitary and testis. It was pre-dicted that p120 would be expressed in the pituitary becausethis is the compartment from which the protein was purified.Equally important, if the protein is relevant to FSH regula-tion, then it must be expressed in the gonadotrope cell. p120and FSHb were detected in the same cell, suggesting a co-localization of p120 with gonadotrophs. The testis has themost distinct and nonoverlapping compartmentalization ofinhibin and activin binding of all tissues examined: inhibinbinding is highest in the Leydig cells, and activin binding isin specific germ cell populations (present paper and Ref. 19).Inhibin binding can be specifically competed by inhibin andnot by activin. The Leydig cell was positive for p120 asdetected by the polyclonal p120 antibody. Moreover, previ-ous studies suggested that inhibin binds Sertoli cells, al-though the resolution of in situ ligand binding was not suf-ficient to identify clearly this compartment as inhibinbinding. p120 is positive in the cytoplasm of Sertoli cells,suggesting that the low grain density detected over thesecells represents true inhibin binding. As was discussed ear-lier, this binding may predict activity. Inhibin stimulatesLeydig cell P45017a mRNA accumulation while activin stim-ulates 3b-hydroxysteroid dehydrogenase mRNA (6). As pre-dicted, inhibin can antagonize activin-induced 3b-hydroxy-steroid dehydrogenase mRNA accumulation; however,activin does not antagonize inhibin-stimulated P45017a.Much of the physiology of inhibin has been directed at its rolein pituitary FSH regulation and an apparent antagonism ofactivin function. The identification of p120, together with thedifferential compartmentalization of inhibin- and activin-binding sites and functional actions of the two ligands, sug-gests an additional level of complexity.
p120 had been previously identified as a large molecularweight expressed protein and then as a member of the a celladhesion molecule family. In humans, the gene for the re-ceptor is encoded by the Xq25 chromosome and is 20 kb with19 exons (17, 18). Duplications in this region of the humanchromosome are associated with X-linked pan-hypopituitar-ism or Pettigrew Syndrome, and loss of heterozygosity isrelated to advanced human ovarian carcinoma (40, 41). Boththe gonads and the pituitary respond to inhibin, and thesefunctional abnormalities identified in humans may imply animportant role for the ligand (through p120) in pituitary
development and in ovarian carcinogenesis. In concert withthe later observation, a loss-of-inhibin mouse model has beendeveloped and the animals develop, with 100% penetrance,gonadal tumors (42, 43).
In summary, inhibin plays critical paracrine and endocrineroles in regulating pituitary FSH, and absence of inhibinresults in ovarian and testicular cancer in knockout mice.Until now, none of the downstream signaling proteins in theinhibin signal transduction cascade had been identified. Aninhibin-binding protein (p120) was purified from severalkilograms of bovine pituitaries using affinity chromatogra-phy. An oligopeptide sequence was determined for the iso-lated and purified protein and the cDNA cloned from humantestis RNA. In addition, p120 is abundant in the testicularLeydig cell, a compartment enriched for inhibin-bindingsites and devoid of activin-binding sites. The purification andcloning of an inhibin-binding protein provide a key reagentto our further understanding of the mammalian reproduc-tive axis as well as the origin of reproductive oncogenesis.
Acknowledgments
The authors thank Erwin Goldberg (Northwestern University, Evan-ston, IL) for adult testis sections. The authors also thank Kelly E. Mayo,Stacey Chapman, Jose Y. Santiago, Malcolm Whitman, and Martin Mat-zuk for helpful discussion.
References
1. Woodruff TK, Krummen LA, Lyon R, Stocks DL, Mather JP 1993 Recom-binant human inhibin A and recombinant human activin A regulate pituitaryand ovarian function in the adult female rat. Endocrinology 132:2332–2341
2. Vale W, Rivier C, Hsueh A, Campen C, Meunier H, Bicsak T 1988 Chemicaland biological characterization of the inhibin family of protein hormones.Recent Prog Horm Res 44:1–34
3. Massague J 1990 The transforming growth factor-b family. Annu Rev Cell Biol6:597–641
4. Mathews L 1994 Activin receptors and cellular signaling by the receptor serinekinase family. Endocr Rev 15:310–325
5. Massague J 1996 TGF signaling: receptors, transducers, and MAD proteins.Cell 85:947–950
6. Lejeune H, Chuzel F, Sanchez P, Durand P, Mather JP, Saez JM 1997 Stim-ulating effect of both human recombinant inhibin A and activin A on immatureporcine Leydig cell functions in vitro. Endocrinology 138:4783–4791
7. Hillier S, Yong E, Illingworth P, Baird D, Schwall R, Mason A 1991 Effectof recombinant inhibin on androgen synthesis in cultured human thecal cells.Mol Cell Endocrinol 75:R1–R6
8. Martens JW, de Winter JP, Timmerman MA, McLuskey A., van Schaik RH,Themmen AP, de Jong FH 1997 Inhibin interferes with activin signaling at thelevel of the activin receptor complex in Chinese hamster ovary cells. Endo-crinology 138:2928–2936
9. Xu J, McKeehan K, Matsuzaki K, McKeehan WL 1995 Inhibin antagonizesinhibition of liver cell growth by activin by a dominant-negative mechanism.J Biol Chem 270:6308–6313
10. Schwall R, Robbins K, Jardieu P, Chang L, Lai C, Terrell T 1993 Activininduces cell death in hepatocytes in vivo and in vitro. Hepatology 18:347–356
11. Alak B, Smith G, Woodruff T, Stouffer R, Wolf D 1996 Enhancement ofprimate oocyte maturation and fertilization in vitro by inhibin A and activinA. Fertil Steril 66:646–653
12. Robertson, DM, Foulds LM, Leversha L., Morgan FJ, Hearn MTW, BurgerHG, Wretenhall REH, deKretser DM 1985 Isolation of inhibin from bovinefollicular fluid. Biochem Biophys Res Commun 126:220–226
13. Castillo RJ, Olivieri E, Vega I 1996 Inhibin binding sites in bovine pituitarymembranes. Biol Res 29:183–188
14. Hearn R, Farnworth P, Fitzsimmons K, Robertson D 1999 Identification ofhigh affinity binding sites for inhibin on ovine pituitary cells in culture. En-docrinology 140:6–12
15. Draper LB, Matzuk M, Roberts V, Cox E, Weiss J, Mather J, Woodruff TK1998 Identification of an inhibin receptor in gonadal tumors from inhibina-subunit knockout mice. J Biol Chem 273:398–40
16. Woodruff TK 1998 Cellular localization of mRNA and protein: in situ hy-bridization histochemistry and in situ ligand binding. Methods Cell Biol57:333–351
2606 IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN Endo • 2000Vol 141 • No 7

17. Frattini A, Faranda S, Redolfi E, Allavena P, Vezzoni P 1998 Identification andgenomic organization of a gene coding for a new member of the cell adhesionmolecule family mapping to Xq25. Gene 214:1–6
18. Mazzarella R, Pengue G, Jones J, Jones C, Schlessinger D 1998 Cloning andexpression of an immunoglobulin superfamily gene (IGSF1) in Xq25. Genomics48:157–162
19. Krummen LA, Moore A, Woodruff TK, Covello R, Taylor R, Working P,Mather JP 1994 Localization of inhibin and activin binding sites in the testisduring development by in situ ligand binding. Biol Reprod 50:734–744
20. Wiesmann C, Fuh G, Christinger H, Eigenbrot C, Wells J, deVos A 1997Crystal structure at 1.7A resolution of VEGF in complex with domain 2 of theFlt-1 receptor. Cell 91:695–704
21. Mason AJ, Miall HD, Seeburg PH 1986 Structure of two human ovarianinhibins. Biochem Biophys Res Commun 135:957–964
22. Aaltonen J, Laitinen MP, Vuojolainen K, Jaatinen R, Horelli-Kuitunen N,Seppa L, Louhio H, Tuuri T, Sjoberg J, Butzow R, Hovata O, Dale L, RitvosO 1999 Human growth differentiation factor 9 (GDF-9) and its novel homologGDF-9B are expressed in oocytes during early folliculogenesis. J Clin Endo-crinol Metab 84:2744–2750
23. Mishina Y, Whitworth DJ, Racine C, Behringer RR 1999 High specificity ofMullerian-inhibiting substance signaling in vivo. Endocrinology 140:2084–2088
24. Treanor JJ, Goodman L, de Sauvage F, Stone DM, Poulsen KT, Beck CD,Gray C, Armanini MP, Pollock RA, Hefti F, Phillips HS, Goddard A, MooreMW, Buj-Bello A, Davies AM, Asai N, Takahashi M, Vandlen R, HendersonCE, Rosenthal A 1996 Characterization of a multicomponent receptor forGDNF. Nature 382:80–82
25. Rivier C, Vale W 1989 Immunoneutralization of endogenous inhibin modifieshormone secretion and ovulation rate in the rat. Endocrinology 125:152–157
26. Mathews LS, Vale WW 1991 Expression cloning of an activin receptor, apredicted transmembrane serine kinase. Cell 65:973–982
27. Krummen LA, Woodruff TK, DeGuzman G, Cox ET, Baly DL, Mann E, GargS, Wong WL, Cossum P, Mather JP 1993 Identification and characterizationof binding proteins for inhibin and activin in human serum and follicularfluids. Endocrinology 132:431–443
28. Schneyer AL, Rzucidlo DA, Sluss PM, Crowley Jr WF 1994 Characterizationof unique binding kinetics of follistatin, and activin or inhibin in serum.Endocrinology 135:667–674
29. Harpaz Y, Chothia C 1994 Many of the immunoglobulin superfamily domainsin cell adhesion molecules and surface receptors belong to a new structural setwhich is close to that containing variable domains. J Mol Biol 238:528–539
30. Carcamo J, Weis FM, Ventura F, Wieser R, Wrana JL, Attisano L, Massague
J 1994 Type I receptors specify growth-inhibitory and transcriptional responsesto transforming growth factor b and activin. Mol Cell Biol 14:3810–3821
31. Lopez-Casillas F, Payne HM, Andres JL, Massague J 1994 Betaglycan can actas a dual modulator of TGF-beta access to signaling receptors: mapping ofligand binding and GAG attachment sites. J Cell Biol 124:557–568
32. Barbara NP, Wrana JL, Letarte M 1999 Endoglin is an accessory protein thatinteracts with the signaling receptor complex of multiple members of thetransforming growth factor-b superfamily. J Biol Chem 274:584–594
33. Roberts AB 1998 Molecular and cell biology of TGF-b. Miner Electrolyte Metab24:111–119
34. McCarthy SA, Bicknell R 1994 Activin-A binds to a heterotrimeric receptorcomplex on the vascular endothelial cell surface. Evidence for a type 3 activinreceptor. J Biol Chem 269:3909–3912
35. Durbec P, Marcos-Gutierrez CV, Kilkenny C, Grigoriou M, Wartiowaara K,Suvanto P, Smith D, Ponder B, Costantini F, Saarma M 1996 GDNF signallingthrough the Ret receptor tyrosine kinase. Nature 381:789–793
36. Lebrun JJ, Vale WW 1997 Activin and inhibin have antagonistic effects onligand-dependent heteromerization of the type I and type II activin receptorsand human erythroid differentiation. Mol Cell Biol 17:1682–1691
37. Baloh RH, Tansey MG, Lampe PA, Fahrner TJ, Enomoto H, Simburger KS,Leitner ML, Araki T, Johnson Jr EM, Milbrandt J 1998 Artemin, a novelmember of the GDNF ligand family, supports peripheral and central neuronsand signals through the GFRa3-RET receptor complex. Neuron 21:1291–1302
38. Saarma M, Sariola H 1999 Other neurotrophic factors: glial cell line-derivedneurotrophic factor (GDNF). Microsc Res Tech 45:292–302
39. van Weering DH, Bos JL 1998 Signal transduction by the receptor tyrosinekinase Ret. Recent Results Cancer Res 154:271–281
40. Lagerstrom-Fermer M, Sundvall M, Johnsen E, Warne GL, Forrest SM, ZajacJD, Rickards A, Ravine D, Landegren U, Pettersson U 1997 X-linked recessivepanhypopituitarism associated with a regional duplication in Xq25–q26. Am JHum Genet 60:910–916
41. Choi C, Cho S, Horikawa I, Berchuck A, Wang N, Cedrone E, Jhung SW, LeeJB, Kerr J, Chenevix-Trench G, Kim S, Barrett JC, Koi M 1997 Loss ofheterozygosity at chromosome segment Xq25–26.1 in advanced human ovar-ian carcinomas. Genes Chromosomes Cancer 20:234–242
42. Matzuk MM, Kumar TR, Shou W, Coerver KA, Lau AL, Behringer RR, andFinegold MJ 1996 Transgenic models to study the roles of inhibins and activinsin reproduction, oncogenesis, and development. Recent Prog Horm Res51:123–154
43. Matzuk MM, Finegold MJ, Su JG, Hsueh AJ, Bradley A 1992 a-Inhibin is atumour-suppressor gene with gonadal specificity in mice. Nature 360:313–319
IDENTIFICATION OF AN INHIBIN-BINDING PROTEIN 2607




















