
Surface modifications of cellulose nanocrystals for biobased ...

©20
15N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature structural & molecular biology advance online publication �
b r i e F com m u n i c at i o n s
Structural insights into the role of rRNA modifications in protein synthesis and ribosome assemblyYury S Polikanov1,2,4, Sergey V Melnikov1,4, Dieter Söll1,3 & Thomas A Steitz1–3
We report crystal structures of the Thermus thermophilus ribosome at 2.3- to 2.5-Å resolution, which have enabled modeling of rRNA modifications. The structures reveal contacts of modified nucleotides with mRNA and tRNAs or protein pY, and contacts within the ribosome interior stabilizing the functional fold of rRNA. Our work provides a resource to explore the roles of rRNA modifications and yields a more comprehensive atomic model of a bacterial ribosome.
Current models suggest that the high fidelity of protein synthesis by the ribosome requires numerous modifications of rRNA1–3. In bacteria, there are three major types of rRNA modifications: conversion of uridines to pseudouridines, methylation of the 2′-hydroxyl groups of ribose and methylation of nucleotide bases1. Modified nucleotides represent only a small fraction of all rRNA resi-dues (23 out of ~4,400 total in T. thermophilus ribosomes4) and cluster mainly near the ribosome
functional centers. Numerous studies have shown that loss of rRNA modifications results in alteration of the structures of active sites5–7; this in turn causes slower rates and lower accuracy of translation8–10 as well as impaired responses to metabolites and antibiotics11–13. Despite the functional importance of rRNA modifications, the reso-lution of previous structural studies of the 70S ribosomes has been insufficient to unambiguously model modified nucleotides, which were instead modeled as regular RNA nucleotides.
Here, we set out to apply our recent advances in crystal treatment to reach diffraction resolution suitable for visualization of modified nucleotides within the 70S ribosome (Online Methods). As a result, we report two crystal structures of the T. thermophilus (Tth) 70S ribosome reflecting two functional states: hibernating ribosomes, in complex with the hibernation factor pY, and translating ribosomes, in complex with mRNA and tRNAs, determined at 2.3-Å and 2.5-Å resolution, respectively (I/σI = 1; Online Methods). The resolution at which I/σI = 2 is 2.5 Å and 2.7 Å for each structure, respectively. At this resolution, we could directly visualize methylation of ribose and nucleotide bases in the unbiased electron density maps (Figs. 1 and 2). Pseudouridines cannot be distinguished from uridines, and therefore their modeling was guided by biochemical data4. In one particular case, however, our electron density map confirmed the presence of pseudouridine (Ψ2605), a result consistent with hydrogen-bonding
1Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, Connecticut, USA. 2Howard Hughes Medical Institute at Yale University, New Haven, Connecticut, USA. 3Department of Chemistry, Yale University, New Haven, Connecticut, USA. 4These authors contributed equally to this work. Correspondence should be addressed to T.A.S. ([email protected]).
Received 5 December 2014; accepted 17 February 2015; published online 16 March 2015; doi:10.1038/nsmb.2992
4Fe-4S
Iron-sulfur cluster
i
ms-Asp88
β-methylthioaspartate
h
m6A1519 2
N6-dimethyladenosineN2-dimethylguanosine
m7G527
7-methylguanosine
m2G1207
N2-methylguanosine
m4Cm1402c
m3U1498
N3-methyluridine
b
4Fe-4S
m6G15182
m6G15192
m3U1498
m5C1407
m5C1404
m4Cm1402 m5C1400
m2G1207
m7G527
ψ516
m5C967m2G9662
Head
Body
Protein modifications16S-rRNA modifications
30S
ms-Asp88
a
Cys31
Cys12
Cys9
Cys26
ge fd
m4Cm1402
N4,O2 ′-dimethylcytidine
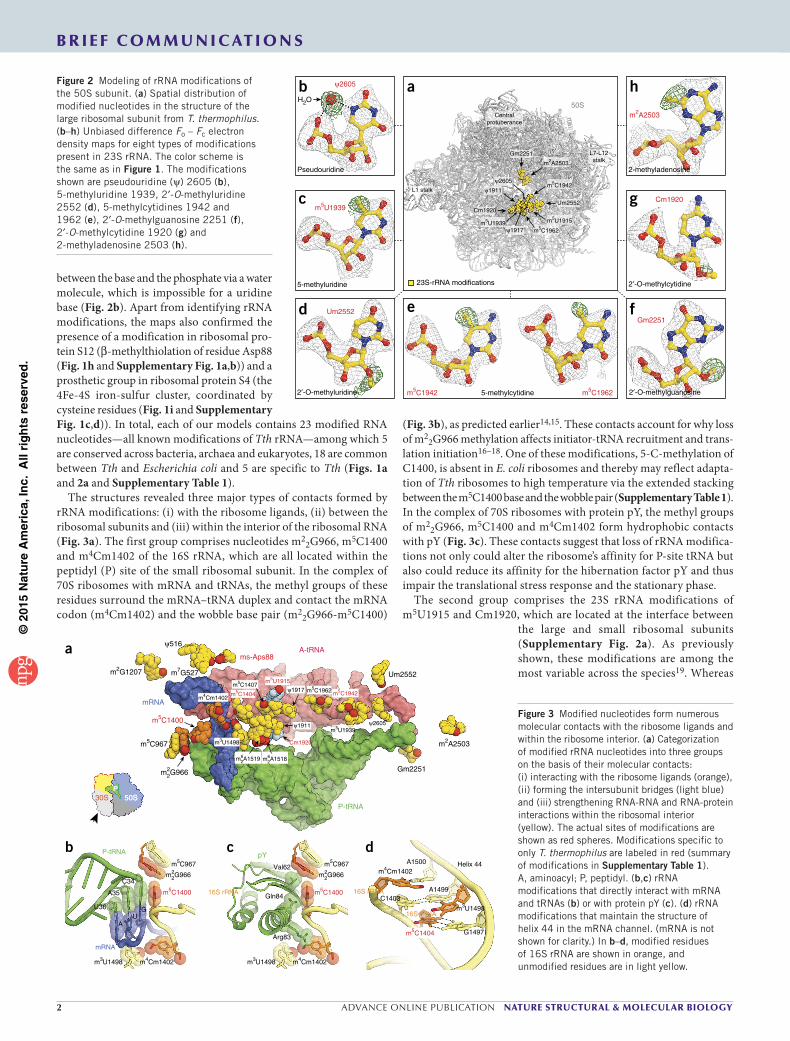
m22G966
Figure 1 Electron density maps allow comprehensive modeling of rRNA and ribosomal-protein modifications of the 30S subunit. (a) Spatial distribution of modified nucleotides in the structure of the small ribosomal subunit from T. thermophilus. Here and throughout, rRNA nucleotides are named according to the E. coli numbering system. (b–i) Unbiased difference Fo – Fc electron density maps for eight types of modifications present in 16S rRNA and ribosomal proteins S12 and S4. Gray mesh, Fo – Fc map after refinement, with modified nucleotides or amino acid residues omitted (contoured at 2.7–3.0σ). Green mesh, Fo – Fc electron density map after refinement with unmodified nucleotides (contoured at 1.5–2.5σ). The modifications shown are N3-methyluridine 1498 (b), N 4,O 2′-dimethylcytidine 1402 (c), N 2-methylguanosine 1207 (d), N2-dimethylguanosine 966 (e), N 7-methylguanosine 527 (f), N 6-dimethyladenosine 1519 (g), β-methylthioaspartate of protein S12 (h) and 4Fe-4S iron-sulfur cluster of protein S4 (i).
©20
15N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
2 advance online publication nature structural & molecular biology
b r i e F com m u n i c at i o n s
between the base and the phosphate via a water molecule, which is impossible for a uridine base (Fig. 2b). Apart from identifying rRNA modifications, the maps also confirmed the presence of a modification in ribosomal pro-tein S12 (β-methylthiolation of residue Asp88 (Fig. 1h and Supplementary Fig. 1a,b)) and a prosthetic group in ribosomal protein S4 (the 4Fe-4S iron-sulfur cluster, coordinated by cysteine residues (Fig. 1i and Supplementary Fig. 1c,d)). In total, each of our models contains 23 modified RNA nucleotides—all known modifications of Tth rRNA—among which 5 are conserved across bacteria, archaea and eukaryotes, 18 are common between Tth and Escherichia coli and 5 are specific to Tth (Figs. 1a and 2a and Supplementary Table 1).
The structures revealed three major types of contacts formed by rRNA modifications: (i) with the ribosome ligands, (ii) between the ribosomal subunits and (iii) within the interior of the ribosomal RNA (Fig. 3a). The first group comprises nucleotides m2
2G966, m5C1400 and m4Cm1402 of the 16S rRNA, which are all located within the peptidyl (P) site of the small ribosomal subunit. In the complex of 70S ribosomes with mRNA and tRNAs, the methyl groups of these residues surround the mRNA–tRNA duplex and contact the mRNA codon (m4Cm1402) and the wobble base pair (m2
2G966-m5C1400)
(Fig. 3b), as predicted earlier14,15. These contacts account for why loss of m2
2G966 methylation affects initiator-tRNA recruitment and trans-lation initiation16–18. One of these modifications, 5-C-methylation of C1400, is absent in E. coli ribosomes and thereby may reflect adapta-tion of Tth ribosomes to high temperature via the extended stacking between the m5C1400 base and the wobble pair (Supplementary Table 1). In the complex of 70S ribosomes with protein pY, the methyl groups of m2
2G966, m5C1400 and m4Cm1402 form hydrophobic contacts with pY (Fig. 3c). These contacts suggest that loss of rRNA modifica-tions not only could alter the ribosome’s affinity for P-site tRNA but also could reduce its affinity for the hibernation factor pY and thus impair the translational stress response and the stationary phase.
The second group comprises the 23S rRNA modifications of m5U1915 and Cm1920, which are located at the interface between
the large and small ribosomal subunits (Supplementary Fig. 2a). As previously shown, these modifications are among the most variable across the species19. Whereas
m2A2503
2-methyladenosine
h
Cm1920
2′-O-methylcytidine
g
Gm2251
2′-O-methylguanosine
f
m5C1942 5-methylcytidine m5C1962
eUm2552
2′-O-methyluridine
d
m5U1939c
H2O
ψ2605
Pseudouridine
b
ψ2605
Um2552
m2A2503
Gm2251
m5C1962
m5C1942
m5U1939
Cm1920
ψ1917
m5U1915
ψ1911
Centralprotuberance
L7-L12stalk
L1 stalk
a50S
23S-rRNA modifications5-methyluridine
Figure 2 Modeling of rRNA modifications of the 50S subunit. (a) Spatial distribution of modified nucleotides in the structure of the large ribosomal subunit from T. thermophilus. (b–h) Unbiased difference Fo – Fc electron density maps for eight types of modifications present in 23S rRNA. The color scheme is the same as in Figure 1. The modifications shown are pseudouridine (ψ) 2605 (b), 5-methyluridine 1939, 2′-O-methyluridine 2552 (d), 5-methylcytidines 1942 and 1962 (e), 2′-O-methylguanosine 2251 (f), 2′-O-methylcytidine 1920 (g) and 2-methyladenosine 2503 (h).
ms-Aps88
m6A15182m6A15192
m2G9662
m3U1498
m5C1407
m5C1404m4Cm1402
m5C1400
m2G1207
m5C967
m7G527
ψ516
ψ2605
Um2552
m2A2503
Gm2251
m5C1962 m5C1942
m5U1939
Cm1920
ψ1917
m5U1915
ψ1911
P-tRNA
A-tRNA
mRNA
30S 50S50SPP
a
Gln84
Arg83
Val62
pY
m3U1498 m4Cm1402
m5C1400
cm5C967
m2G9662
16S rRNA
U36
A35
C34
P-tRNA
GU
A
mRNA
m3U1498 m4Cm1402
m5C1400
m5C967
m2G9662
bA1500
A1499
m3U1498
G1497m5C1404
C1403
m4Cm1402Helix 44
16S rRNA
d
16S rRNA
Figure 3 Modified nucleotides form numerous molecular contacts with the ribosome ligands and within the ribosome interior. (a) Categorization of modified rRNA nucleotides into three groups on the basis of their molecular contacts: (i) interacting with the ribosome ligands (orange), (ii) forming the intersubunit bridges (light blue) and (iii) strengthening RNA-RNA and RNA-protein interactions within the ribosomal interior (yellow). The actual sites of modifications are shown as red spheres. Modifications specific to only T. thermophilus are labeled in red (summary of modifications in Supplementary Table 1). A, aminoacyl; P, peptidyl. (b,c) rRNA modifications that directly interact with mRNA and tRNAs (b) or with protein pY (c). (d) rRNA modifications that maintain the structure of helix 44 in the mRNA channel. (mRNA is not shown for clarity.) In b–d, modified residues of 16S rRNA are shown in orange, and unmodified residues are in light yellow.

©20
15N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature structural & molecular biology advance online publication 3
b r i e F com m u n i c at i o n s
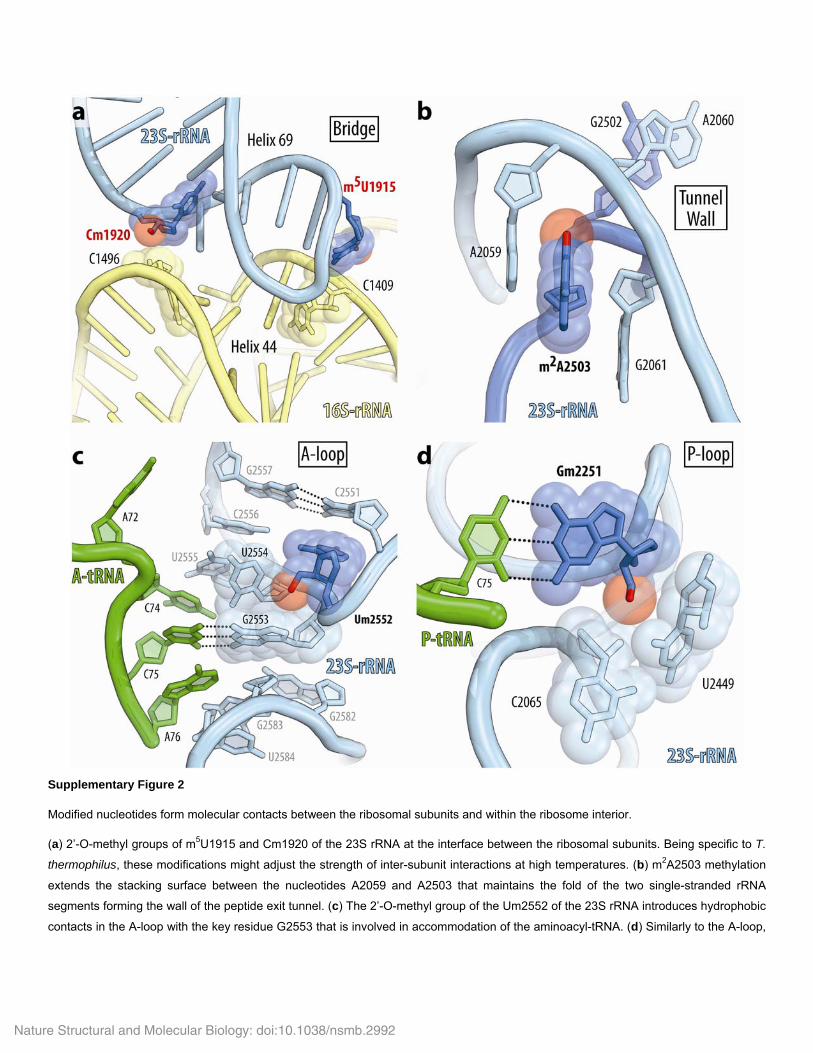
U1915 is 5-C-methylated in Tth, it is 3-N-methylated and pseudouri-dylated in E. coli and unmethylated but pseudouridylated in archaea (Haloarcula marismortui) and eukaryotes (Saccharomyces cerevisiae and Homo sapiens)19. The second residue, C1920, is 2′-O-methylated in Tth, unmethylated in E. coli and pseudouridylated in eukaryotic ribosomes. In the classical state of the ribosome in both of our complexes, the 2′-O-methyl group forms hydrophobic interactions between the large and the small subunits (Supplementary Fig. 2a). Therefore, these two modifications might adjust the strength of the intersubunit contacts to meet the requirements of translation at different temperatures or compensate for the interspecies variability in the intersubunit bridges.
The third and the most abundant group comprises the remaining nucleotides that are buried inside and reside within the single-stranded rRNA segments or within highly distorted helices, in which rRNA adopts energetically unfavorable folds. Our structures revealed that, except for pseudouridines, most of these modifications form or extend the hydrophobic interactions that further stabilize rRNA structure. Perhaps the most unusual example of structural motifs involving rRNA modifications is our observation of a cluster of modified nucleotides in the mRNA channel (Fig. 3d). In the middle part of this cluster, nucleotides C1403, m3U1498 and A1499 form a base triple, whose structure is supported by 3-N-methylation of U1498 and by stacking with the flanking bases of m4Cm1402 and m5C1404. These inter-actions occur at the interface of the mRNA channel and thus might be important for proper accommodation of the mRNA. As predicted by previous studies, similar motifs are found in other functional sites of the ribosome, such as the peptide-exit tunnel or the tRNA-binding sites on the large ribosomal subunit3. In the peptide-exit tunnel, the 2-C-methyl group of A2503 (m2A2503) extends the stacking between A2059 and A2503. This stacking supports the fold of the two single-stranded rRNA segments of the peptidyl transferase loop, which form the wall of the peptide-exit tunnel (Supplementary Fig. 2b). In the A loop of the 23S rRNA, the 2′-O-methyl group of Um2552 intercalates between the bases of G2553 and U2554. Apparently, these contacts maintain the active conformation of the G2553 base, which is directly involved in accommodation of the aminoacyl-tRNA (Supplementary Fig. 2c). Similarly, the P loop of the 23S rRNA carries 2′-O-methylated Gm2251, which is the key residue required for accommodation of the P-site tRNA via Watson-Crick base-pairing with the 3′ end of tRNA. The 2′-O-methyl group of Gm2251 forms additional hydrophobic con-tacts with the sugar of C2065 and the base of U2449 (Supplementary Fig. 2d). Because the m2A2503, Gm2251 and Um2552 modifications are universally conserved across kingdoms of life, similar or identical interactions are expected to occur in the ribosomes from other species, including humans. Overall, Tth rRNA carries 18 rRNA modifications, which are buried or oriented toward the interior of rRNA helices and loops, where they form additional hydrophobic contacts with neighboring nucleotides and, in a few cases, with ribosomal proteins (Supplementary Table 1). Therefore, the major role of modified nucleotides within the ribosome is possibly related to the maintenance of distorted rRNA folds at the functional interfaces of the ribosome.
In summary, we have visualized previously unseen modifications of ribosomal RNA. This allowed us to create what are, to our knowledge,
the most comprehensive models of bacterial ribosome and provide a resource to explore unique properties of modified nucleotides in the context of translating or hibernating ribosomes. Because RNA modifications are present in most noncoding RNAs and mRNAs, our structures pave the way to understanding the general principles of how nucleotide modifications control RNA structure and function.
MeThOdsMethods and any associated references are available in the online version of the paper.
Accession codes. Coordinates and structure factors have been depos-ited in the Protein Data Bank under accession codes 4Y4O for the Tth 70S ribosome in complex with protein pY and 4Y4P for the Tth 70S ribosome in complex with mRNA and tRNA ligands.
Note: Any Supplementary Information and Source Data files are available in the online version of the paper.
AcknowleDgMenTSWe thank J. Lin, C.A. Innis and N.V. Sumbatyan for bringing this project to our attention; R.L. Grodzicki for preparation of the unmodified tRNAs and for critical reading of the manuscript; and members of the T.A.S. and D.S. laboratories for discussions. We also thank the staff at the Advanced Photon Source (beamline 24ID), supported by award GM103403 from the US National Center for Research Resources at the US National Institutes of Health, the staff at the US National Synchrotron Light Source (beamline X25) for help during data collection and the staff at the Richards Center at Yale University for computational support. This work was supported by US National Institutes of Health grants GM022778 (T.A.S.) and GM022854 (D.S.).
AUTHoR conTRIBUTIonSY.S.P. and S.V.M. devised experiments; Y.S.P. crystallized ribosomal complexes and performed crystallographic data collection and processing, S.V.M. and Y.S.P. analyzed the data, and S.V.M., Y.S.P., D.S. and T.A.S. wrote the manuscript.
coMPeTIng FInAncIAl InTeReSTSThe authors declare no competing financial interests.
Reprints and permissions information is available online at http://www.nature.com/reprints/index.html.
1. Decatur, W.A. & Fournier, M.J. Trends Biochem. Sci. 27, 344–351 (2002).2. Chow, C.S., Lamichhane, T.N. & Mahto, S.K. ACS Chem. Biol. 2, 610–619 (2007).3. Sergiev, P.V. et al. in Ribosomes: Structure, Function, and Dynamics (eds. Rodnina,
M.V. et al.) 97–110 (Springer, 2011).4. Mengel-Jørgensen, J. et al. J. Biol. Chem. 281, 22108–22117 (2006).5. Demirci, H. et al. RNA 16, 2319–2324 (2010).6. Desaulniers, J.P. et al. Org. Biomol. Chem. 6, 3892–3895 (2008).7. Sumita, M., Jiang, J., SantaLucia, J. Jr. & Chow, C.S. Biopolymers 97, 94–106 (2012).8. Liang, X.H., Liu, Q. & Fournier, M.J. Mol. Cell 28, 965–977 (2007).9. Baudin-Baillieu, A. et al. Nucleic Acids Res. 37, 7665–7677 (2009).10. Baxter-Roshek, J.L., Petrov, A.N. & Dinman, J.D. PLoS ONE 2, e174 (2007).11. Helser, T.L., Davies, J.E. & Dahlberg, J.E. Nat. New Biol. 233, 12–14 (1971).12. Doi, Y. & Arakawa, Y. Clin. Infect. Dis. 45, 88–94 (2007).13. Prokhorova, I.V. et al. Sci. Rep. 3, 3236 (2013).14. Prince, J.B., Taylor, B.H., Thurlow, D.L., Ofengand, J. & Zimmermann, R.A.
Proc. Natl. Acad. Sci. USA 79, 5450–5454 (1982).15. Kimura, S. & Suzuki, T. Nucleic Acids Res. 38, 1341–1352 (2010).16. Burakovsky, D.E. et al. Nucleic Acids Res. 40, 7885–7895 (2012).17. Jemiolo, D.K., Taurence, J.S. & Giese, S. Nucleic Acids Res. 19, 4259–4265 (1991).18. Arora, S. et al. Nucleic Acids Res. 41, 4963–4975 (2013).19. Piekna-Przybylska, D., Decatur, W.A. & Fournier, M.J. Nucleic Acids Res. 36,
D178–D183 (2008).

©20
15N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature structural & molecular biology doi:10.1038/nsmb.2992
20. Polikanov, Y.S., Steitz, T.A. & Innis, C.A. Nat. Struct. Mol. Biol. 21, 787–793 (2014).
21. Polikanov, Y.S., Blaha, G.M. & Steitz, T.A. Science 336, 915–918 (2012).22. Kabsch, W. Acta Crystallogr. D Biol. Crystallogr. 66, 125–132 (2010).23. Emsley, P., Lohkamp, B., Scott, W.G. & Cowtan, K. Acta Crystallogr. D Biol.
Crystallogr. 66, 486–501 (2010).24. Adams, P.D. et al. Acta Crystallogr. D Biol. Crystallogr. 66, 213–221 (2010).
ONLINe MeThOdsPurification of T. thermophilus ribosomes and preparation of tRNAs. 70S ribosomes from T. thermophilus and unmodified tRNAi
Met and tRNAPhe from E. coli were purified as previously described20. Synthetic mRNA with the sequence 5′-GGC AAG GAG GUA AAA AUG UUC UAA-3′ was obtained from Integrated DNA Technologies.
Crystallization and data collection. The ribosome complex with protein pY was formed essentially as described previously21. The ribosome complex with mRNA and tRNAs was formed by programming 5 µM Tth 70S ribosomes with 10 µM mRNA and 20 µM P- and A-site tRNA substrates. All Tth 70S ribosome complexes were formed in buffer containing 5 mM HEPES-KOH, pH 7.6, 50 mM KCl, 10 mM NH4Cl, and 10 mM Mg(CH3COO)2 and were then crystal-lized in buffer containing 100 mM Tris-HCl, pH 7.6, 2.9% (w/v) PEG 20000, 7–12% (v/v) MPD, 100–200 mM arginine and 0.5 mM β-mercaptoethanol. Crystals were grown by the vapor-diffusion method in sitting drops at 19 °C and were stabilized as previously described20. Diffraction data were collected at beamline 24ID-C at the Advanced Photon Source. A complete data set for each ribosome complex was collected at 0.979-Å wavelength at 100 K from multiple regions of the same crystal with 0.2° oscillations.
Crystallographic data processing and model building. The raw data were integrated and processed with the XDS package22. All crystals belonged to the primitive orthorhombic space group P212121 with approximate unit-cell dimen-sions of 210 Å × 450 Å × 620 Å and contained two copies of the 70S ribosome per asymmetric unit. The initial molecular-replacement solutions were refined by
rigid-body refinement and subsequent positional and individual B-factor refine-ment. Determination of locations of rRNA modifications within the rRNA was facilitated by the use of the Fournier laboratory’s database of ribosomal RNA modifications19 and biochemical data for T. thermophilus4. The electron density maps allowed location of the previously unknown position of methyl groups in the nucleotides C1915 (m5C1915) and C1940 (m5C1940). All modified nucleo-tides were also modeled in both tRNAi
Met and tRNAPhe. The final models of the 70S ribosome in complex with either pY or mRNA and tRNAs were generated by multiple rounds of model building in COOT23 and subsequent refinement in PHENIX24. The improved quality of the electron density maps allowed us to build a more accurate model of the 70S ribosome with more residues of the ribosomal proteins falling into the favored and allowed regions of the Ramachandran plot: 95.79% favored, 3.73% allowed and 0.47% disallowed for the 70S–pY complex, and 94.80% favored, 4.29% allowed and 0.91% disallowed for the 70S–mRNA–tRNAs complex. The statistics of data collection and refinement for each complex are compiled in Supplementary Table 2. In both structures, all rRNA nucleotides were renumbered according to the E. coli numbering system.

Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992
Supplementary Figure 1
Post-translational modification of ribosomal protein S12 and prosthetic group of protein S4.
(a, b) β-methyl-thiolation of Asp88 (ms-Asp88) in ribosomal protein S12 is a bacteria-specific and essential modification with unknown
role in protein synthesis (Kowalak, J.A. & Walsh, K.A. Beta-methylthio-aspartic acid: identification of a novel posttranslational
modification in ribosomal protein S12 from Escherichia coli. Protein Sci. 5, 1625-1632, 1996). We show that this post-translational
modification establishes an additional RNA-protein contact, mediated by the N7-methyl group of the modified nucleotide m7G527. This
interaction, observed in a close proximity to the decoding center, suggests that the essential function of msD88 might be related to the

Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992
assembly of the decoding center and that the effect of msD88 on the ribosome structure may depends on methylation of m7G527. (a)
The 30S subunit is shown in light yellow with protein S12 highlighted in light blue. (b) Close-up view of panel (a). (c, d) The iron-sulfur
cluster coordinated by protein S4 has been recently identified in a crystallographic study of T. thermophilus ribosomes (for details see
Online Methods section in (Polikanov, Y.S., Steitz, T.A. & Innis, C.A. A proton wire to couple aminoacyl-tRNA accommodation and
peptide-bond formation on the ribosome. Nat. Struct. Mol. Biol. 21, 787-793, 2014)). It is located in the vicinity of the mRNA entrance
tunnel on the cytoplasmic side of the small ribosomal subunit. The covalent association of the 4Fe-4S cluster with protein S4 is
mediated by four cysteine residues (Cys9, Cys12, Cys26, and Cys31) in the N-terminal globular domain. Presence of the iron-sulfur
cluster within T. thermophilus ribosomes is consistent with metabolism of these bacteria that require high concentration of iron for
optimal growth. It also explains why samples of pure ribosome have a yellow/red color. Importantly, it has been reported previously that
protein S4 can associate with RNA polymerase in vivo (Torres, M., Condon, C., Balada, J.M., Squires, C. & Squires, C.L. Ribosomal
protein S4 is a transcription factor with properties remarkably similar to NusA, a protein involved in both non-ribosomal and ribosomal
RNA antitermination. Embo J. 20, 3811-3820, 2001). Although the function of this prosthetic group has not been examined, it is
appealing to propose that protein S4 can mediate the ribosome interaction with RNA-polymerase during the coupled transcription-
translation, where iron-sulfur cluster stabilizes the complex at high temperatures. Location of protein S4 on the 30S subunit is viewed
from the cytoplasm with the 50S subunit removed, as indicated by the inset. (c) The 30S subunit is shown in light yellow with protein S4
highlighted in green. The mRNA is shown in blue, and arrows are pointing to its entrance and exit channels. (d) Close-up view of panel
(c).
Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992
Supplementary Figure 2
Modified nucleotides form molecular contacts between the ribosomal subunits and within the ribosome interior.
(a) 2’-O-methyl groups of m5U1915 and Cm1920 of the 23S rRNA at the interface between the ribosomal subunits. Being specific to T.
thermophilus, these modifications might adjust the strength of inter-subunit interactions at high temperatures. (b) m2A2503 methylation
extends the stacking surface between the nucleotides A2059 and A2503 that maintains the fold of the two single-stranded rRNA
segments forming the wall of the peptide exit tunnel. (c) The 2’-O-methyl group of the Um2552 of the 23S rRNA introduces hydrophobic
contacts in the A-loop with the key residue G2553 that is involved in accommodation of the aminoacyl-tRNA. (d) Similarly to the A-loop,

Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992
the 2’-O-methyl group of Gm2251 fills the cavity between the sugar of C2065 and the base of U2449 in the P-loop of the 23S rRNA.
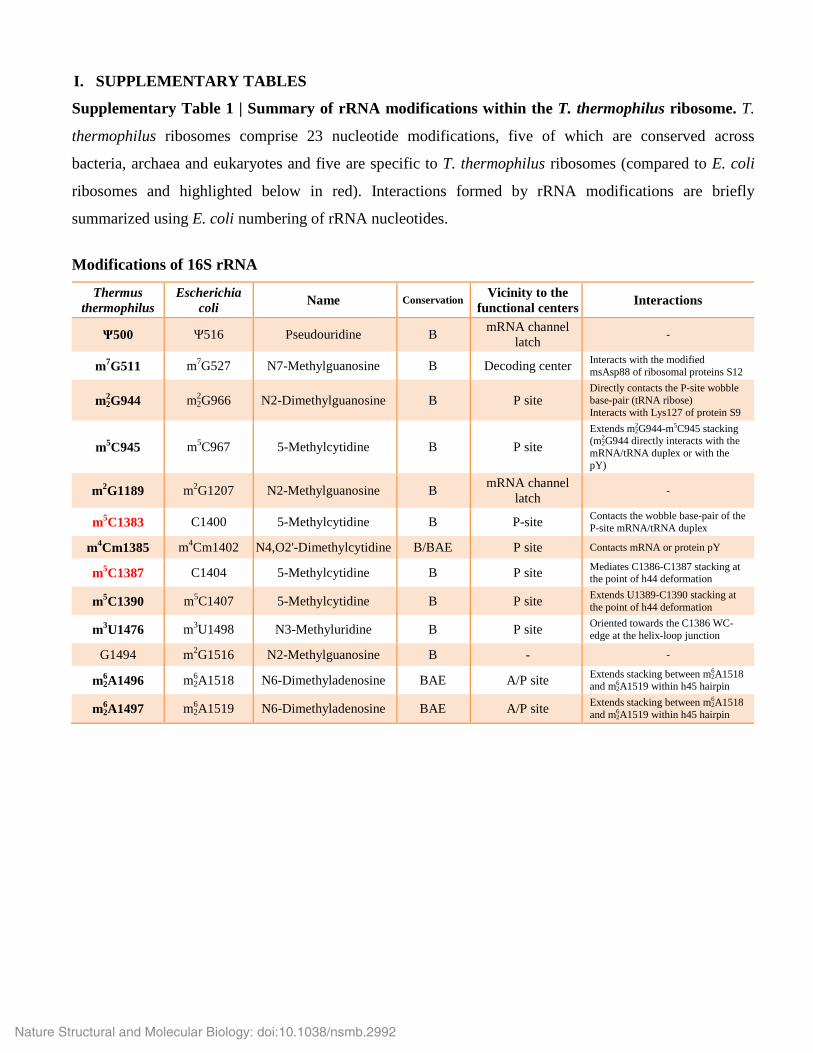
I. SUPPLEMENTARY TABLES
Supplementary Table 1 | Summary of rRNA modifications within the T. thermophilus ribosome. T.
thermophilus ribosomes comprise 23 nucleotide modifications, five of which are conserved across
bacteria, archaea and eukaryotes and five are specific to T. thermophilus ribosomes (compared to E. coli
ribosomes and highlighted below in red). Interactions formed by rRNA modifications are briefly
summarized using E. coli numbering of rRNA nucleotides.
Modifications of 16S rRNA
Thermus thermophilus
Escherichia coli Name Conservation Vicinity to the
functional centers Interactions
Ψ500 Ψ516 Pseudouridine B mRNA channel latch -
m7G511 m7G527 N7-Methylguanosine B Decoding center Interacts with the modified msAsp88 of ribosomal proteins S12
m22 G944 m2
2 G966 N2-Dimethylguanosine B P site Directly contacts the P-site wobble base-pair (tRNA ribose) Interacts with Lys127 of protein S9
m5C945 m5C967 5-Methylcytidine B P site Extends m2
2 G944-m5C945 stacking (m2
2 G944 directly interacts with the mRNA/tRNA duplex or with the pY)
m2G1189 m2G1207 N2-Methylguanosine B mRNA channel latch -
m5C1383 C1400 5-Methylcytidine B P-site Contacts the wobble base-pair of the P-site mRNA/tRNA duplex
m4Cm1385 m4Cm1402 N4,O2'-Dimethylcytidine B/BAE P site Contacts mRNA or protein pY
m5C1387 C1404 5-Methylcytidine B P site Mediates C1386-C1387 stacking at the point of h44 deformation
m5C1390 m5C1407 5-Methylcytidine B P site Extends U1389-C1390 stacking at the point of h44 deformation
m3U1476 m3U1498 N3-Methyluridine B P site Oriented towards the C1386 WC-edge at the helix-loop junction
G1494 m2G1516 N2-Methylguanosine B - -
m62 A1496 m6
2 A1518 N6-Dimethyladenosine BAE A/P site Extends stacking between m62 A1518
and m62 A1519 within h45 hairpin
m62 A1497 m6
2 A1519 N6-Dimethyladenosine BAE A/P site Extends stacking between m62 A1518
and m62 A1519 within h45 hairpin
Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992

Modifications of 23S rRNA
Thermus thermophilus
Escherichia coli Name Conservation Vicinity to the
functional centers Interactions
G791 m1G745 N1-Methylguanosine B - - A792 Ψ746 Pseudouridine B - - U793 m5U747 5-Methyluridine B - - C999 Ψ955 Pseudouridine B - -
A1663 m6A1618 N6-Methyladenosine B - - G1865 m2G1835 N2-Methylguanosine B - - Ψ1932 Ψ1911 Pseudouridine B Helix 69 -
m5U1936 m3Ψ1915 5-Methyluridine B Helix 69 Extends U1915-A1916 stacking and faces h44 of the small subunit
Ψ1938 Ψ1917 Pseudouridine B Helix 69 -
Cm1941 C1920 2'-O-Methylcytidine B Helix 69 Directly contacts h44 of the small ribosomal subunit
m5U1960 m5U1939 5-Methyluridine B - Extends A1937-U1939 stacking at the helix-bulge border
m5C1963 C1942 5-Methylcytidine B A/P site Extends C1941-C1942-U1943 stacking within the H70-H71 helical junction
m5C1983 m5C1962 5-Methylcytidine B A/P site Extends G1935-C1962 stacking within the H70-H71 helical junction
A2051 m6A2030 N6-Methyladenosine B - -
G2090 m7G2069 N7-Methylguanosine B - -
Gm2262 Gm2251 2'-O-Methylguanosine BAE P site
Intercalates between the ribose of C2065 and the base of U2449 (G2251 is a key residue which base-pairs with the CCA-end of the P-site tRNA)
G2456 Gm2445 N2-Methylguanosine B - -
U2460 D2449 5,6-Dihydrouridine B - -
U2468 Ψ2457 Pseudouridine B - -
C2509 Cm2498 2'-O-Methylcytidine B - -
m2A2514 m2A2503 2-Methyladenosine BAE Peptide tunnel Extends A2059-A2503 stacking between two segments of single-stranded rRNA
U2515 Ψ2504 Pseudouridine B - -
Um2563 Um2552 2'-O-Methyluridine BAE A site Intercalates between the bases of U2554 and G2553 (G2553 is a key residue which base-pairs with the CCA-end of the A-site tRNA)
U2591 Ψ2580 Pseudouridine B - -
U2615 Ψ2604 Pseudouridine B - -
Ψ2616 Ψ2605 Pseudouridine B - Coordinates water molecule between the base and the phosphate using pseudouridine-specific H-bonds
Nature Structural and Molecular Biology: doi:10.1038/nsmb.2992
Copyright © 2022 FDOKUMEN




















