
Structural Basis for the Carbohydrate Recognition of the Sclerotium rolfsii Lectin
17
Structural Basis for the Carbohydrate Recognition of the Sclerotium rolfsii Lectin Demetres D. Leonidas 1 ⁎, Bale M. Swamy 2 , George N. Hatzopoulos 1 Sathisha J. Gonchigar 2 , Vishwanath B. Chachadi 2 Shashikala R. Inamdar 2 , Spyros E. Zographos 1 and Nikos G. Oikonomakos 1 1 Institute of Organic & Pharmaceutical Chemistry, The National Hellenic Research Foundation, 48 Vas. Constantinou Avenue, 11635 Athens, Greece 2 Department of Biochemistry, Karnatak University, Dharwad 580003, India The crystal structure of a novel fungal lectin from Sclerotium rolfsii (SRL) in its free form and in complex with N-acetyl-D-galactosamine (GalNAc) and N-acetyl- D -glucosamine (GlcNAc) has been determined at 1.1 Å, 2.0 Å, and 1.7 Å resolution, respectively. The protein structure is composed of two β-sheets, which consist of four and six β-strands, connected by two α-helices. Sequence and structural comparisons reveal that SRL is the third member of a newly identified family of fungal lectins, which includes lectins from Agaricus bisporus and Xerocomus chrysenteron that share a high degree of structural similarity and carbohydrate specificity. The data for the free SRL are the highest resolution data for any protein of this family. The crystal structures of the SRL in complex with two carbohydrates, GalNAc and GlcNAc, which differ only in the configuration of a single epimeric hydroxyl group, provide the structural basis for its carbohydrate specificity. SRL has two distinct carbohydrate-binding sites, a primary and a secondary. GalNAc binds at the primary site, whereas GlcNAc binds only at the secondary site. Thus, SRL has the ability to recognize and probably bind at the same time two different carbohydrate structures. Structural comparison to Agaricus bisporus lectin–carbohydrate complexes reveals that the primary site is also able to bind the Thomsen-Friedenreich antigen (Galβ1→3 GalNAc-α- glycan structures) whereas the secondary site cannot. The fea- tures of the molecular recognition at the two sites are described in detail. © 2007 Published by Elsevier Ltd. *Corresponding author Keywords: Sclerotium rolfsii; lectin; structure; carbohydrate; X-ray crystallography Introduction Cell recognition and adhesion is an important event in many biological processes including cell differen- tiation and development. Amongst various types of molecules that act as mediators of cellular interac- tions, lectins have a unique and pivotal role. 1 Lectins are widely distributed in nature, and they are defined as mono- or multivalent carbohydrate-binding, (glyco)proteins, different from immunoglobulins that have no enzymic activity towards their carbohy- drate ligand. 2 Since lectins display unique glycan specificity, they have been described as tools to read sugar-encoded messages expressed in glycans attached to soluble and integral glycoconjugates. 3 The recognition mechanism and the fine specificity of lectins towards complex carbohydrate moieties of cell Present address: G. N. Hatzopoulos, EMBL Hamburg Outstation, Notkestrasse 85, Building 25A, D-22603 Hamburg, Germany. Abbreviations used: SRL, Sclerotium rolfsii lectin; ABL, Agaricus bisporus lectin; XCL, Xerocomus chrysenteron lectin; AAL, Aleuria aurantia lectin; Fve, Flammulina velutipes lectin; CGL2, Coprineus cinerea lectin; ACG, Agrocybe cylindracea lectin; LSL, Laetiporus sulphureus lectin; PVL, Psathyrella velutina lectin; GalNAc, N-acetyl-D-galactosamine; GlcNAc, N-acetyl-D-glucosamine; MPD, 2-methyl-2,4-pentanediol; r.m.s.d., root-mean-square deviation. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2007.02.092 J. Mol. Biol. (2007) 368, 1145–1161 0022-2836/$ - see front matter © 2007 Published by Elsevier Ltd.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Structural Basis for the Carbohydrate Recognition of the Sclerotium rolfsii Lectin

doi:10.1016/j.jmb.2007.02.092 J. Mol. Biol. (2007) 368, 1145–1161
Structural Basis for the Carbohydrate Recognition ofthe Sclerotium rolfsii Lectin
Demetres D. Leonidas1⁎, Bale M. Swamy2, George N. Hatzopoulos1
Sathisha J. Gonchigar2, Vishwanath B. Chachadi2
Shashikala R. Inamdar2, Spyros E. Zographos1
and Nikos G. Oikonomakos1
1Institute of Organic &Pharmaceutical Chemistry,The National Hellenic ResearchFoundation, 48 Vas.Constantinou Avenue,11635 Athens, Greece2Department of Biochemistry,Karnatak University,Dharwad 580003, India
Present address: G. N. HatzopoulOutstation, Notkestrasse 85, BuildinHamburg, Germany.Abbreviations used: SRL, Sclerotiu
Agaricus bisporus lectin; XCL, Xerocolectin; AAL, Aleuria aurantia lectin; Fvelutipes lectin; CGL2, Coprineus cineAgrocybe cylindracea lectin; LSL, Laetlectin; PVL, Psathyrella velutina lectinN-acetyl-D-galactosamine; GlcNAc,N-acetyl-D-glucosamine; MPD, 2-mer.m.s.d., root-mean-square deviationE-mail address of the correspondi
0022-2836/$ - see front matter © 2007 P
The crystal structure of a novel fungal lectin from Sclerotium rolfsii (SRL) inits free form and in complex with N-acetyl-D-galactosamine (GalNAc) andN-acetyl- D -glucosamine (GlcNAc) has been determined at 1.1 Å, 2.0 Å,and 1.7 Å resolution, respectively. The protein structure is composed of twoβ-sheets, which consist of four and sixβ-strands, connected by twoα-helices.Sequence and structural comparisons reveal that SRL is the third member ofa newly identified family of fungal lectins, which includes lectins fromAgaricus bisporus and Xerocomus chrysenteron that share a high degree ofstructural similarity and carbohydrate specificity. The data for the free SRLare the highest resolution data for any protein of this family. The crystalstructures of the SRL in complex with two carbohydrates, GalNAc andGlcNAc, which differ only in the configuration of a single epimeric hydroxylgroup, provide the structural basis for its carbohydrate specificity. SRL hastwo distinct carbohydrate-binding sites, a primary and a secondary.GalNAc binds at the primary site, whereas GlcNAc binds only at thesecondary site. Thus, SRL has the ability to recognize and probably bind atthe same time two different carbohydrate structures. Structural comparisonto Agaricus bisporus lectin–carbohydrate complexes reveals that the primarysite is also able to bind the Thomsen-Friedenreich antigen (Galβ1→3GalNAc-α- glycan structures) whereas the secondary site cannot. The fea-tures of the molecular recognition at the two sites are described in detail.
© 2007 Published by Elsevier Ltd.
Keywords: Sclerotium rolfsii; lectin; structure; carbohydrate;X-ray crystallography
*Corresponding authoros, EMBL Hamburgg 25A, D-22603
m rolfsii lectin; ABL,mus chrysenteronve, Flammulinarea lectin; ACG,iporus sulphureus; GalNAc,
thyl-2,4-pentanediol;.ng author: [email protected]
ublished by Elsevier Ltd.
Introduction
Cell recognition and adhesion is an important eventin many biological processes including cell differen-tiation and development. Amongst various types ofmolecules that act as mediators of cellular interac-tions, lectins have a unique and pivotal role.1 Lectinsare widely distributed in nature, and they are definedas mono- or multivalent carbohydrate-binding,(glyco)proteins, different from immunoglobulinsthat have no enzymic activity towards their carbohy-drate ligand.2 Since lectins display unique glycanspecificity, they have been described as tools to readsugar-encoded messages expressed in glycansattached to soluble and integral glycoconjugates.3
The recognition mechanism and the fine specificity oflectins towards complex carbohydratemoieties of cell

†http://plab.ku.dk/tcbh/Lectins15/Sawmy/paper.htm
1146 Crystal Structure of the Sclerotium rolfsii Lectin
surfaces has been the focus of current research in-terests in glycobiology. For example galectins, afamily of vertebrate lectins, with high affinity for N-acetyllactosamine that regulate a number of biologicalprocesses including inflammatory responses, infec-tious processes and tumour growth and invasion,have been the target for the discovery of anti-galectincompounds as potential anti-cancer drugs.4 Anothergoal is to exploit this fine glycan specificity and uselectins as biosensors that target specific cells for thedevelopment of new prognostic and diagnostic toolsfor cancer.5 Lectins also have the potential to mediatedrug targeting and delivering.6
Different roles and functions have been ascribed tolectins. The principal function of animal lectins is toact as recognition molecules within the immune sys-tem,7 while plant lectins function as storage proteinsand they have been implicated in defence mechan-isms against invading pathogens and predators.2
Most of the bacterial and viral lectins are involved inhost recognition, cell surface adhesion, biofilm for-mation and toxicity. Unlike plant, animal and bac-terial lectins functional role(s) for fungal lectins hasyet to be established. However, many suggestionshave been put forward based on their location,including mediation of host–parasite interactions,storage, mycoparasitism2 and morphogenesis, anddevelopment of fungi.8 Despite reports on theirdiverse biological functions and knowledge of mole-cular characterization, limited information is avail-able on structural aspects of fungal lectins comparedto plant and animal lectins. Thus, although thestructural basis of the carbohydrate recognitionmode for animal, plant, and to some extent forbacterial and viral, lectins has been elucidated,9 thatof fungal lectins is still in its infancy. Ever since theelucidation of the crystal structure of first fungallectin, Aleuria aurantia (AAL) in 2003,10,11 three-dimensional structures of seven fungal lectins havebeen determined by X-ray crystallography to date.This sudden surge of interest is because of theirunique protein fold different, which is from anypreviously described lectin fold, and their novelsugar-binding properties.12
To date, eight mushroom lectin crystal structureshave been described. A. aurantia fucose-binding lec-tin (AAL) was found to be a six-bladed β-propeller,different from any previously known lectin fold.10,11
Lectin purified from Flammulina velutipes (Fve),showing specificity for complex cell-surface carbo-hydrates with immunomodularory activity,13 dis-plays structural similarity to human fibronectin.14
Whereas lectin from Xerocomus chrysenteron (XCL)15
and Agaricus bisporus (ABL)16 both resemble actino-porins, a family of pore-forming toxins. Both XCLand ABL recognize N-acetyl galactosamine and ga-lactose, while ABL also recognizes Thomsen–Frie-denreich (TF) antigen, a disaccharide (Galβ1-3 N-acetyl-D-galactosamine (GalNAc)) over-expressedon cell-surface glycoproteins in a variety of humancarcinomas.17,18 Remarkably, ABL exhibits reversi-ble antiproliferative activity on a variety of epithelialcells.19 XCL also shows antiproliferative activity on
different cell lines and interestingly exhibits potentinsecticidal activity.20,21
Lectins from Coprineus cinerea (CGL2)22 and Agro-cybe cylindracea (ACG)23 share a common carbohy-drate recognition domain with galectins, a largefamily of β-galactoside-binding animal lectins witha highly conserved carbohydrate recognition domain(CRD). CGL2 has unique sugar specificity and bindsstrongly to the blood group A tetrasaccharideGalNAcα1-3 [Fucα1-2]Galβ1-4Glc;24 on the otherhand, ACG recognizes sialoconjugates.25 Recently,the crystal structures of Laetiporus sulphureus lectin(LSL)26 and Psathyrella velutina lectin (PVL)27 havebeen reported. LSL has a distinct N-terminal lectindomain with a β-trefoil scaffold and a C-terminalpore-formingmodule, similar to those found in toxinssuch as ricin,28 abrin29 and Clostridium botulinum.30
PVL, unlike other known fungal lectins ,binds to Ca2+
with a characteristic seven-bladedβ-propeller foldingseen in integrins,31 apart form its sugar-binding site.Sclerotium rolfsii, a soil-borne plant pathogenic
fungus capable of forming fruiting bodies referred toas sclerotial bodies, secretes a cell wall-associated,developmental-stage specific lectin (SRL)†.32,33 Re-cent findings indicate that SRL is expressed on themycelia at the time of sclerotial body formation andfacilitates the aggregation of the mycelium to formsclerotial bodies by interacting with its endogenousglycosyl ceramide receptor(s), which have specificcarbohydrate moieties.33 SRL is a monomer underacidic conditions (pH 4.3) with a molecular mass of17 kDa and all experimental evidence so far suggestthat it forms a dimer at neutral or basic pH.32
Matrix-assisted laser desorption/ionization tandemtime-of-flight (MALDI TOF/TOF) mass spectro-metry experiments at pH 3.0, 4.5, 6.8, 7.8 and 9.0have shown that SRL is dimeric at neutral or basicpH and monomeric at acidic pH (B.M.S. et al.,unpublished results). SRL displays a clear specifi-city for Galβ1→3GalNAc-α- (TF, Thomsen-Frieden-reich antigen) and GalNAc-α- (Tn antigen) glycanstructures.34The TF antigen, is a carbohydrate antigen specifi-
cally over-expressed in human tumour cells.18 In re-cent years, TF antigen-binding lectins have attractedconsiderable interest, since they have potentialapplications in cancer diagnostics and therapy.Thus, it has been shown that ABL, and jacalin(a lectin from Artocarpus integrifolia seeds) inhibitthe proliferation of HT29 colonic cancer cells,whereas peanut (Arachis hypogaea) lectin (PNA) andAmaranthus caudatus lectin (ACL), stimulate theirproliferation.35 Although these lectins share specifi-city for the TF antigen, they exhibit subtle differencesin their finer specificities towards modifications ofthis glycan structure and hence they bring aboutmarkedly different effects in cancerous epithelia. It isof interest therefore to understand the molecularbasis of the TF-recognition specificity by such lectins

1147Crystal Structure of the Sclerotium rolfsii Lectin
to elucidate their mode of carbohydrate recognitionand engineer novel binding activities.Here, we report the crystal structures of SRL in free
form at near-atomic resolution (1.1 Å) and in complexwith N-acetyl-D-glucosamine (GlcNAc) and GalNAcat 1.7 Å and 2.0 Å resolution. Both the crystalstructure and the amino acid sequence of SRL revealthat SRL is similar to fungal lectins XCL15 and ABL,16
with which it shares similar carbohydrate specificityfor TF antigen-based glycan structures. The SRLcomplex structures reveal that SRL has two distinctcarbohydrate-binding sites, a primary and a second-ary, and elicits the structural determinants of themolecular recognition for carbohydrate binding.
Results and Discussion
Description of the overall structure
There are two SRL molecules in the crystal-lographic asymmetric unit and all residues arewell defined within the electron density map. Adifference map calculated using phases from therefined SRL model showed unambiguously addi-tional electron density for an acetate group cova-lently attached to the first residue in both monomers(molecule A and molecule B) of the non-crystal-lographic dimer in agreement with attempted fail-ure to sequence the protein by chemical methods,which indicated that the N terminus was blocked(B.M.S. et al., unpublished results).SRL is a compact molecule with approximate
dimensions of 38 Å×36 Å×23 Å. (Figure 1(a)). Thetwo monomers in the asymmetric unit have nearlyidentical structures with r.m.s. deviations of 0.21 Å(Cα atoms), 0.20Å (main-chain) and 0.57Å (all atoms).In the native structure, the non-crystallographicdimer contains 462 bound water molecules, in ad-dition to a tris(hydroxymethyl)aminomethane, twoacetate, and three 2-methyl-2,4-pentanediol (MPD)molecules from the crystallization medium. The N-terminal residue (Thr) is acetylated while six residuesin molecule A and nine residues in molecule B of theSRL dimer display alternative conformations for theirside-chains.SRL is a typical α+β protein consisting of two β
sheets (F and S). Four antiparallel β-strands (F1–F4)compose the F β-sheet, while the S β-sheet consistsof six strands (S1–S6). The topology of the S β-sheetis mixed with strands S1, S2, S4 and S6 antiparallelto S3 and S5 (Figure 1(b)). The packing of the twoβ-sheets creates a cavity inside the moleculealigned mostly by hydrophobic residues. Two α-helices (H1, H2) connect strands F4 and S4, orientedalmost perpendicular to each other (Figure 1(a)).These α-helices pack against the F β-sheet mainlywith polar interactions, while the outer surface ofthe S β-sheet is mostly negatively charged (Figure2). The SRL molecule has a total charge of −3.0,with the C-terminal part of the molecule chargedand the N-terminal part uncharged or neutral(Figure 2). The β-strands and α-helices are con-
nected with very short loops (the longest is fourresidues), resulting in a very close packing of thewhole molecule. The SRL molecule contains asingle cysteine residue at position 76. It is buriedin the middle of strand F4 in both protein moleculesof the dimer in the crystal, and the thiol group isnot involved in any crystal packing contact.
Quaternary structure
The two SRL monomers are related by a non-crystallographic 2-fold symmetry (spherical polarangles, ω=135.6°, φ=−45.5°, and χ=179.4°). Thetwo molecules pack antiparallel with their β-sheetsinclined ∼45°, while the α-helices are located at thesame side of the dimer (Figure 1(a)). Thus, thecavities created by the two β sheets in each SRLmolecule are not aligned, and each one is blocked bythe β-strand F1 of the adjacent molecule. The otherend of each cavity is blocked by β-strand S4. Thedimer interface involves the association of residuesfrom F1, S2, F2, and H1, and from the loopsconnecting F1 to S2, and F4 to H1 (Figure 1(a)).These are held together by 17 intermolecular hydro-gen bonding interactions between five residues frommolecule A and eight residues from molecule B, andan extensive network of van der Waals interactions(Table 1). In addition, seven water molecules at thedimer interface mediate hydrogen bond interactionsbetween the two molecules.The solvent accessibilities of the SRL monomers
are 6907 Å2 and 6990 Å2 for molecules A and B,respectively. Upon dimer association, these surfacesreduce to 6019 Å2 and 6110 Å2, indicating that areasof 888 Å2 and 880 Å2 become inaccessible to solventin molecules A and B, respectively. This representsapproximately 13% of the surface of each moleculethat becomes buried. The greatest contributioncomes from the non-polar residues, which contri-bute 569 Å2 (64%) and 574 Å2 (65%) of the surface,which becomes inaccessible in SRL molecules A andB, respectively. The total buried surface area in thecomplex is 1768 Å2. The shape correlation statisticSc,36 which is used to quantify the shape comple-mentarity of interfaces and gives an idea of the“goodness of fit” between two surfaces, is 0.741 forthe association of SRL molecules A and B.
The binding of N-acetylgalactosamine in thecrystal
GalNAc is one of the most potent saccharideligands of SRL.34 In attempts to identify the car-bohydrate recognition site of SRL, various soakingexperiments were performed with different sugarligands based on the SRL carbohydrate specificity.34
Soaking SRL crystals with the TF antigen (Galβ1-3GalNAcα1-Thr), the most potent sugar ligand forSRL,34 caused total destruction of the crystals within30 s, evenwith carbohydrate concentrations as low as500 μM. The same phenomenon, but to a muchsmaller extent, was observed while soaking SRLcrystalswithGalNAc.However,wewere able to form

Figure 1. (a) A presentation ofthe SRL dimer. Strands F1–F4 andS1–S6 and helices H1 and H2 arelabelled. (b) Topology diagram ofthe SRL structure. Strands and he-lices are represented as arrows andcylinders, respectively. The diagramwas drawn with TopDraw.73
1148 Crystal Structure of the Sclerotium rolfsii Lectin
the SRL-GalNAc complex in the crystal by soaking asingle SRL crystal with a solution containing 30 mMGalNAc in the crystallization media for 10 min atroom temperature. After this short soaking time, theSRL crystal was cooled immediately to 100 K and X-ray diffraction data were collected. However, thecrystal of the complex diffracted to significantly lowerresolution (1.96 Å) than the crystal of the free protein(1.1 Å) using synchrotron radiation. Attempts tocrystallize SRL in the presence of GalNAc or the TF-antigen were unsuccessful. Furthermore, soakingcrystals of the SRL-GlcNAc complex with solutionscontaining GalNAc also caused severe deteriorationof the crystals and even after 5 min of soaking the X-ray diffraction pattern was totally abolished. X-raydata from SRL crystals soaked with 50 mM galactosefor several days or grown in the presence of 10 mMgalactose did not show any binding.Two distinct binding sites, approximately 21 Å
apart (Figure 3), have been found for GalNAc in
SRL. The primary, which has GalNAc bound in bothSRL molecules of the non-crystallographic dimer,and the secondary, which is occupied by GalNAconly in molecule B of the SRL dimer in the crystal. Inthe complex structure, the carbohydrate moiety iswell defined within the electron density map in thethree binding sites of the SRL dimer (Figure 3,insets). Structural comparison between the free andthe complexed SRL structures indicates that theprotein does not undergo any significant conforma-tional change upon carbohydrate binding. The r.m.s.deviation between the free and the liganded SRLstructures is 0.19 Å (main chain atoms) and 0.27 Å(all atoms). The primary binding site is formed as ashallow groove created by the loops connectingstrands F1 to S1, S3 to F2, F3 to F4, and the end of theH2 helix (Figure 3). The binding of GalNAc to theprimary binding site is almost identical in both SRLmolecules of the asymmetric unit. Two MPD mol-ecules that participate in crystal packing contacts
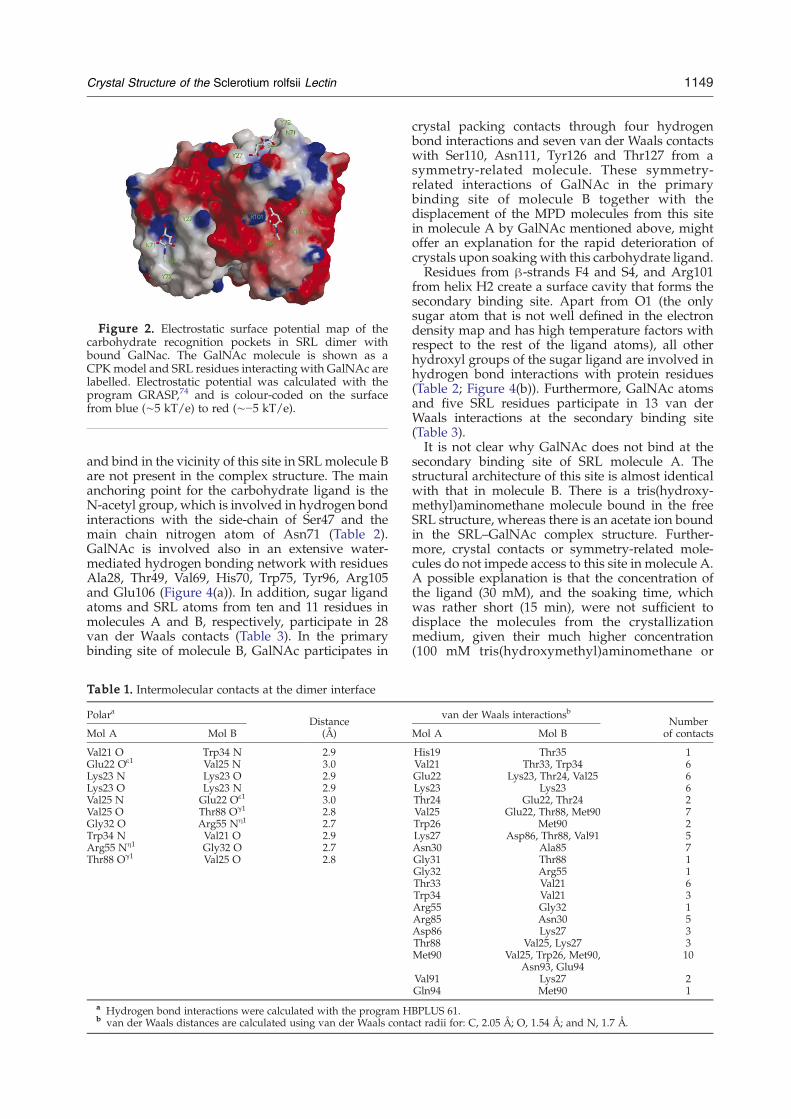
Figure 2. Electrostatic surface potential map of thecarbohydrate recognition pockets in SRL dimer withbound GalNac. The GalNAc molecule is shown as aCPKmodel and SRL residues interacting with GalNAc arelabelled. Electrostatic potential was calculated with theprogram GRASP,74 and is colour-coded on the surfacefrom blue (∼5 kT/e) to red (∼−5 kT/e).
1149Crystal Structure of the Sclerotium rolfsii Lectin
and bind in the vicinity of this site in SRL molecule Bare not present in the complex structure. The mainanchoring point for the carbohydrate ligand is theN-acetyl group, which is involved in hydrogen bondinteractions with the side-chain of Ser47 and themain chain nitrogen atom of Asn71 (Table 2).GalNAc is involved also in an extensive water-mediated hydrogen bonding network with residuesAla28, Thr49, Val69, His70, Trp75, Tyr96, Arg105and Glu106 (Figure 4(a)). In addition, sugar ligandatoms and SRL atoms from ten and 11 residues inmolecules A and B, respectively, participate in 28van der Waals contacts (Table 3). In the primarybinding site of molecule B, GalNAc participates in
Table 1. Intermolecular contacts at the dimer interface
PolaraDistance
(Å)Mol A Mol B
Val21 O Trp34 N 2.9Glu22 Oε1 Val25 N 3.0Lys23 N Lys23 O 2.9Lys23 O Lys23 N 2.9Val25 N Glu22 Oε1 3.0Val25 O Thr88 Oγ1 2.8Gly32 O Arg55 Nη1 2.7Trp34 N Val21 O 2.9Arg55 Nη1 Gly32 O 2.7Thr88 Oγ1 Val25 O 2.8
a Hydrogen bond interactions were calculated with the program Hb van der Waals distances are calculated using van der Waals conta
crystal packing contacts through four hydrogenbond interactions and seven van der Waals contactswith Ser110, Asn111, Tyr126 and Thr127 from asymmetry-related molecule. These symmetry-related interactions of GalNAc in the primarybinding site of molecule B together with thedisplacement of the MPD molecules from this sitein molecule A by GalNAc mentioned above, mightoffer an explanation for the rapid deterioration ofcrystals upon soaking with this carbohydrate ligand.Residues from β-strands F4 and S4, and Arg101
from helix H2 create a surface cavity that forms thesecondary binding site. Apart from O1 (the onlysugar atom that is not well defined in the electrondensity map and has high temperature factors withrespect to the rest of the ligand atoms), all otherhydroxyl groups of the sugar ligand are involved inhydrogen bond interactions with protein residues(Table 2; Figure 4(b)). Furthermore, GalNAc atomsand five SRL residues participate in 13 van derWaals interactions at the secondary binding site(Table 3).It is not clear why GalNAc does not bind at the
secondary binding site of SRL molecule A. Thestructural architecture of this site is almost identicalwith that in molecule B. There is a tris(hydroxy-methyl)aminomethane molecule bound in the freeSRL structure, whereas there is an acetate ion boundin the SRL–GalNAc complex structure. Further-more, crystal contacts or symmetry-related mole-cules do not impede access to this site in molecule A.A possible explanation is that the concentration ofthe ligand (30 mM), and the soaking time, whichwas rather short (15 min), were not sufficient todisplace the molecules from the crystallizationmedium, given their much higher concentration(100 mM tris(hydroxymethyl)aminomethane or
van der Waals interactionsbNumber
of contactsMol A Mol B
His19 Thr35 1Val21 Thr33, Trp34 6Glu22 Lys23, Thr24, Val25 6Lys23 Lys23 6Thr24 Glu22, Thr24 2Val25 Glu22, Thr88, Met90 7Trp26 Met90 2Lys27 Asp86, Thr88, Val91 5Asn30 Ala85 7Gly31 Thr88 1Gly32 Arg55 1Thr33 Val21 6Trp34 Val21 3Arg55 Gly32 1Arg85 Asn30 5Asp86 Lys27 3Thr88 Val25, Lys27 3Met90 Val25, Trp26, Met90,
Asn93, Glu9410
Val91 Lys27 2Gln94 Met90 1
BPLUS 61.ct radii for: C, 2.05 Å; O, 1.54 Å; and N, 1.7 Å.

Figure 3. A presentation of SRL in complex with N-acetyl-galactosamine (white in the primary and grey in thesecondary binding sites, respectively). Inset: plots of the σA 2|Fo| – |Fc| electron density map, contoured at the 1.0 σlevel, for theN-acetylgalactosamine molecule at the primary and the secondary SRL carbohydrate-binding sites before theincorporation of the sugar ligand models.
1150 Crystal Structure of the Sclerotium rolfsii Lectin
200 mM acetate) from this site and achieve binding.In the secondary site of SRL molecule B, GalNAcparticipates in three crystal packing contacts: (i) awater-mediated hydrogen bond interaction of thehydroxyl O1 group to the main chain nitrogen atomof Lys115 from a symmetry-related molecule (this
Table 2. Potential hydrogen bonds of GalNAc with SRL
GalNAcatom
Primary binding siteSecondarybinding site
Distance (Å) Distance (Å)
SRL atom Mol A Mol B SRL atom Mol B
O1 Water 2.8 Water 3.3O3 Water 3.1 Tyr112 Oη 2.5O3 Water 2.9 2.3 Ile78 O 2.7O4 Gly48 O 2.7 2.7 Asp77 Oδ1 2.8O4 Water 2.9 2.7 Asp77 Oδ2 2.7O4 Ile78 O 2.9O5 Arg101 Nη1 3.1O6 Water 2.7 2.9 Asp77 Oδ2 3.0O6 Arg101 Nη1 2.8O7 Water 3.1 Thr80 Oγ1 3.0O7 Ser47 Oγ 2.8 2.9 Thr80 N 2.8O7 Asn71 N 2.8 2.8O7 Gly48 N 3.3
Hydrogen bond interactions were calculated with the programHBPLUS.61 Atom names are shown in Figure 3.
interaction might be an artefact of the model, sinceatom O1 is modelled in the GalNAc molecule due tothe lack of electron density for this atom); (ii) awater-mediated hydrogen bond from the N1 atomto the N1 atom of the GalNAc molecule in thesecondary binding site of a symmetry-relatedmolecule; and (iii) a van der Waals interaction toresidue Val114 from a symmetry-related molecule.These contacts, albeit weak, might contribute furtherto ligand binding to this site in molecule B, sincethey are not possible in the secondary binding site ofSRL molecule A. Modelling the binding of GalNActo the secondary site of SRL molecule A by super-imposition of the structure of the SRL molecule B –GalNAc complex onto the structure of SRL moleculeA has shown that binding of GalNAc to this site inmolecule A is feasible. However, we cannot rule outthe possibility that binding of GalNAc at this site isan artefact imposed by crystal packing forces.On forming the complex with the protein, GalNAc
becomes buried in both binding sites. The solvent-accessible surface area of the freeGalNAcmolecule is372 Å2. This area, upon binding, shrinks to 133 Å2 atthe primary binding site and 103 Å2 at the secondarybinding site, indicating that surface areas of 239 Å2
(64%) and 269 Å2 (72%) become buried in the proteincomplex at the two sites, respectively. The greatest
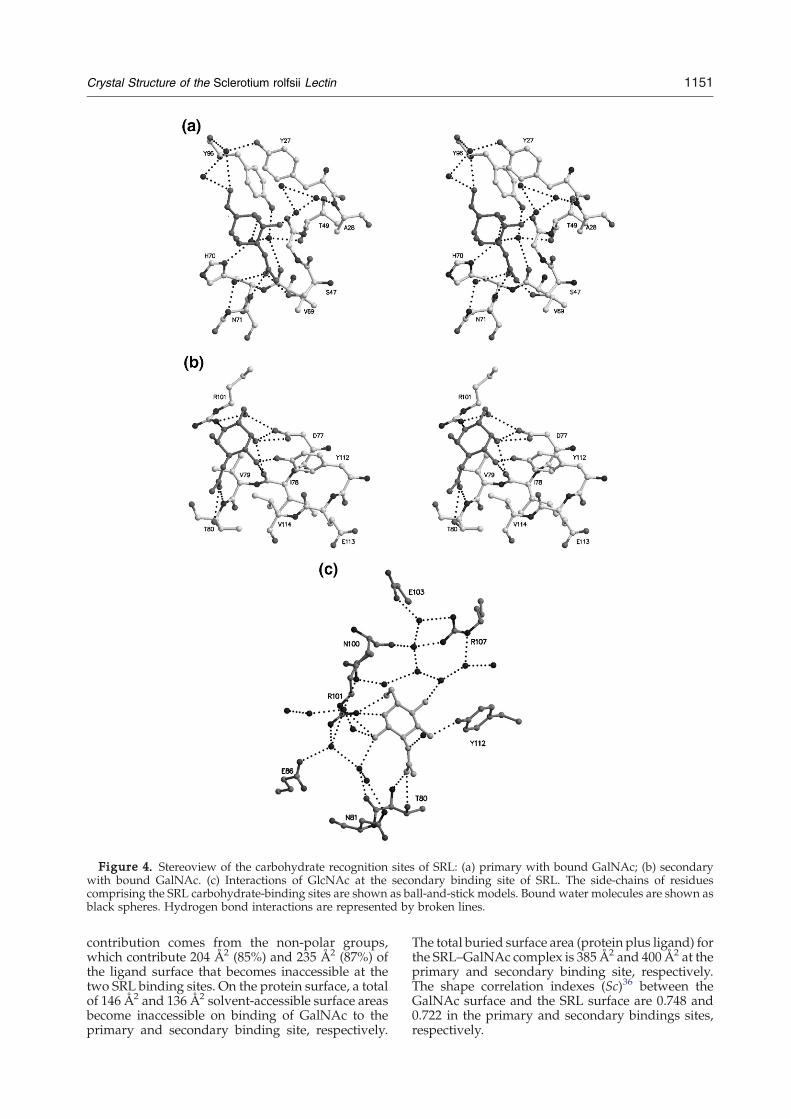
Figure 4. Stereoview of the carbohydrate recognition sites of SRL: (a) primary with bound GalNAc; (b) secondarywith bound GalNAc. (c) Interactions of GlcNAc at the secondary binding site of SRL. The side-chains of residuescomprising the SRL carbohydrate-binding sites are shown as ball-and-stick models. Bound water molecules are shown asblack spheres. Hydrogen bond interactions are represented by broken lines.
1151Crystal Structure of the Sclerotium rolfsii Lectin
contribution comes from the non-polar groups,which contribute 204 Å2 (85%) and 235 Å2 (87%) ofthe ligand surface that becomes inaccessible at thetwo SRL binding sites. On the protein surface, a totalof 146 Å2 and 136 Å2 solvent-accessible surface areasbecome inaccessible on binding of GalNAc to theprimary and secondary binding site, respectively.
The total buried surface area (protein plus ligand) forthe SRL–GalNAc complex is 385 Å2 and 400 Å2 at theprimary and secondary binding site, respectively.The shape correlation indexes (Sc)36 between theGalNAc surface and the SRL surface are 0.748 and0.722 in the primary and secondary bindings sites,respectively.

Table 3. van der Waals interactions between GalNAc and SRL in the crystal
GalNAcatoms
Primary site Secondary site
SRL residues (mol A) SRL residues (mol B) SRL residues (mol B)
C1 His70, Cε1; Asn71, Oδ1 His70, Cε1
C2 Gly48, Cα Gly48, Cα
C3 Gly48, Cα Gly48, Cα Tyr112, Oη
C4 Tyr27, Cε2, Cδ2 Tyr27, Cδ2 Asp77, Cγ, Oδ1
C6 Tyr96, Cδ1; Arg105, Cζ, Nη1 Arg105, Nη1, Nη2 Arg101, Nη1
C7 Ser47, Oγ, C, O; Gly48, Cα; Ser47, Oγ, C Thr80, Oγ1, Val114, Cγ2
Asn71, Cα, Oδ1 Gly48, Cα
C8 Ser47, Cβ, C, O; Thr127, O Ser47, Cβ, Oγ; Tyr72, Cζ, Oη, Cε2 Thr80, Oγ1; Val114, Cγ2
O3 Gly48, Cα Gly48, Cα Tyr112, Cζ, Cε2
O4 Tyr27, Cδ2; Gly48, C Gly48, Cα, C Asp77, Cγ
O5 His70 Cε1
O6 Asp14, Cβ Ty27 Cε2 Arg101, Cδ
O7 Ser47, Oγ, C; Gly48, Cα; Asn71, Cα Ser47, Oγ; Gly48, Cα; Asn71, Cα Thr80, Cα
Total Nine residues (28 contacts) Seven residues (21 contacts) Five residues (13 contacts)
van der Waals distances are calculated using van der Waals contact radii for: C 2.05 Å; O, 1.54 Å; and N, 1.7 Å.
1152 Crystal Structure of the Sclerotium rolfsii Lectin
The binding of N-acetylglucosamine in thecrystal
GlcNAc is a weak ligand for SRL.34 In contrast tothe formation of the SRL–GalNAc complex, theSRL–GlcNAc complex was formed without anydifficulties by soaking a single SRL crystal in100 mM GlcNAc in the crystallization media forthree days. Although the structure presented here isbased on a soaking experiment, data from SRL co-crystallized with 10 mMGlcNAc were also availableat 2.0 Å resolution. Structural analysis showed thatthe carbohydrate is bound in exactly the same wayas in the soaked crystal.GlcNAc has been found bound in SRL only in the
secondary carbohydrate-binding site of both the SRLmolecules of the non-crystallographic dimer. Allatoms of the ligand are well defined within theelectron density map, which has shown that GlcNAcbinds to this site with two alternative conformationswith respect to atomO1, α and β (Figure 5) that haveequal occupancies in molecule A and 0.7 and 0.3 inmolecule B. This finding is consistent with theambiguity in the location of atom O1 of GalNAcobserved in the ligand upon binding at the second-ary binding site of SRL. The primary binding site inthe complex is occupied bywater molecules in mole-
cule A and twoMPDmolecules inmolecule B. One ofthese MPDmolecules aligns well with location of theN-acetyl moiety of GalNAc in the SRL–GalNAccomplex structure. Structural comparison betweenthe free and the SRL–GlcNAc complex structureindicates that the protein does not undergo anysignificant conformational change upon carbohy-drate binding. The r.m.s. deviation between the freeand the liganded SRL structures is 0.12 Å (main chainatoms) and 0.27 Å (all atoms).The binding of GlcNAcis almost identical in both molecules of the non-crystallographic dimer and thus we will discuss thebinding of this ligand only in molecule A. With theexception of hydroxyl group O1, all other hydroxylgroups of the sugar ligand are involved in hydrogenbond interactions with protein residues (Table 4;Figure 4(c)), a fact that offers an explanation for thealternative conformations of this group. In bothalternative conformations of GlcNAc the hydroxylgroup O1 is involved in hydrogen bond interactionsonly with water molecules. The rest of the hydroxylgroups of GlcNAc are involved in hydrogen bondinteractions with residues Asp77, Ile78, Thr80,Arg101, and Tyr112 (Table 4, Figure 4(c)).With the exception of hydroxyl group O4 (the epi-
meric hydroxyl group), the binding mode GlcNAc issimilar to the binding of GalNAc at this site in
Figure 5. Stereoview of σA2|Fo|–|Fc| electron densitymap, contoured at the 1.0 σ level,for the N-acetylglucosamine mole-cule at the secondary SRL carbohy-drate-binding sites before theincorporation of the carbohydrateligand model.
Table 5. van der Waals interactions between GlcNAc andSRL in the crystal
GalNAcatoms
SRL residues
Molecule A Molecule B
C3 Ile78, O; Tyr112, Oη Ile78, O; Tyr112, Oη
C4 Asp77, Cγ, Oδ1, Oδ2; Ile78, O Asp77, Cγ, Oδ1, Oδ2
C6 Asn100, Cγ, C; Arg101, Cα, Nη1 Arg101, Cα, Nη1
C7 Thr80, Oγ1;Val114, Cγ2 Thr80, Cγ2;Val114, Cγ2
C8 Thr80, Oγ1;Val114, Cγ2 Thr80, Cγ2;Val114, Cγ2
O3 Tyr112, Cζ, Cε2 Tyr112, Cζ, Cε2
O4 Asp77, Cγ Asp77, Cγ
O6 Arg101, Cγ, Cδ Arg101, Cγ, Cδ
O7 Thr80, Cβ Val79, Cα, C; Thr80, Cγ2
Total Seven residues(20 contacts)
Seven residues(19 contacts)
van der Waals distances are calculated using van der Waalscontact radii for: C, 2.05 Å; O, 1.54 Å; and N, 1.7 Å.
Table 4. Potential hydrogen bonds of GlcNAc with SRL
Distance (Å)
GlcNAc atom SRL atomMolecule A(conf. A/B)
Molecule B(conf. A/B)
O1 Water 2.8/3.0 2.8/2.5O1 Water 2.8/3.3 2.6/3.3O1 Water 2.8/−O3 Tyr112 Oη 2.6/2.8 2.7/2.7O3 Ile78 O 2.7/2.6 2.7/2.6O4 Asp77 Oδ1 2.6/2.6 2.7/2.7O4 Tyr112 Oη 3.3/3.2 −/3.2O4 Water 2.6/2.7O5 Arg101 Nη1 2.9/3.0 2.9/3.1O6 Arg101 Nη1 2.9/2.8 3.0/2.9O6 Asp77 Oδ2 2.7/2.7 2.7/2.8O7 Thr80 N 2.9/2.7 2.7/2.7O7 Thr 80 O −/3.3O7 Thr80 Oγ1 2.9/3.0O7 Water −/3.3 −/2.7N2 Water 2.9/2.9 3.2/3.2
Hydrogen bond interactions were calculated with the programHBPLUS.61 Atom names are the same as in GalNAc and areshown in Figure 3.
1153Crystal Structure of the Sclerotium rolfsii Lectin
molecule B of the SRL–GalNAc complex. In addition,in SRL molecule B, GlcNAc like GalNAc, participatesin three crystal packing contacts through: (i) a water-mediated hydrogen bond interaction of the hydroxylO1 group to the main chain nitrogen atom of Lys115from a symmetry-relatedmolecule (observed only forconformation α); (ii) a water-mediated hydrogenbond from theN1 atom to theN1 atom of the GlcNAcmolecule in the secondary binding site of a symmetry-related molecule; and (iii) a van der Waals interactionto residue Val114 from a symmetry-related molecule.Furthermore, GlcNAc atoms and seven SRL residuesparticipate in 19 (moleculeA) and 20 (molecule B) vander Waals interactions at the secondary binding site(Table 5).On forming the complex with the protein, GlcNAc
becomes buried. The solvent-accessible surface areaof the free GlcNAcmolecule is 369 Å2. Upon binding,this area shrinks to 88 Å2, indicating that a surfacearea of 281 Å2 (76%) becomes buried in the proteincomplex. The greatest contribution comes from thenon-polar groups, which contribute 200 Å2 (71%) ofthe ligand surface that becomes inaccessible at theSRL secondary binding site. On the protein surface, atotal of 142 Å2 of solvent-accessible surface areabecomes inaccessible on binding GalNAc. The totalburied surface area (protein plus ligand) for the SRL–GlcNAc complex is 423 Å2. The shape correlationindex (Sc)36 between the GlcNAc surface and theSRL surface is 0.747 and 0.769 in molecules A and Bof the non-crystallographic dimer, respectively.
Comparison to other fungal lectins
A sequence-based homologue search usingFASTA37 against the UNIPROT protein database38revealed that SRL is homologous (37–64% aminoacid identity) to fungal lectins ABL,39 XCL,20 Paxillusinvolutus (PIL),40 Pleurotus cornucopiae (cornucopiamushroom; isolectins PCL and PCL-2),41 Arthrobo-
trys oligospora (nematode-trapping fungus; AOL),42
Podospora anserina43 (PAL) and Neurospora crassa(NCL)44 (Figure 6).A Dali45 three-dimensional structural similarity
search against all proteins in the Protein Data Bank46revealed that the SRL structure is most similar toXCL.20 The Z-score (strength of structural similarityin standard deviations) was 26.5, three times higherthan the second best, with a positional r.m.s.d. of0.9 Å for 138 equivalent Cα atoms. XCL has potentinsecticidal activity,20 and blocks the proliferation ofseveral cell lines by disrupting the cell–substrateadhesion.21 XCL is similar to ABL, which alsoexpresses selective cytotoxicity against malignantepithelial cell lines by blocking the sequence-depen-dent nuclear protein import.47,48 SRL shares withABL and XCL similar carbohydrate specificities forN-acetyl galactosamine and the TF-antigen.20,34,49
The r.m.s. distances between the structures of SRLand XCL are 0.66 Å (Cα atoms), 0.66 Å (main chain)and 0.91 Å (all atoms) for 138 structurally equivalentresidues. The structures of SRL and ABL have r.m.s.deviations of 0.53 Å (Cα atoms), 0.51 Å (main chain)and 0.92 Å (all atoms) for 141 equivalent residues.The marginally larger difference found with ABLwhen considering all atoms might be related todifferences in the sequence and the fact that adifferent number of amino acids were considered.The corresponding r.m.s. deviation between thestructures of XCL and ABL is 0.80 Å, 0.78 Å, and1.02 Å for 138 equivalent residues. Differencesbetween the three structures are focused mainly inthe loop regions, with the most notable in the loopconnecting strands F2 and F3 (Figure 7). Also in thestructure of XCL, the loop between the two α-heliceshas a conformation different from the one it adoptsin the structures of SRL and ABL (Figure 7).The structures of ABL complexes with four mono-
saccharides and seven disaccharides have shownthat ABL also has distinct primary and secondarycarbohydrate binding sites.16 Comparison of thecarbohydrate-binding pockets in the three fungallectins, SRL, ABL and XCL shows that both theprimary and the secondary binding sites are con-

Figure 6. Structure-based sequence alignment of SRL, ABL (PDB ID 1y2t),16 and XCL (1xi0)15 as performed by Dali.45
The sequences of fungal lectins from Paxillus involutus (PIL),40 Pleurotus cornucopiae (isolectins PCL and PCL-2),41
Arthrobotrys oligospora (AOL),42 Podospora anserina43 (PAL) and Neurospora crassa (NCL)44 aligned by CLUSTALW75 to theSRL sequence are shown. Every tenth residue in the SRL sequence is numbered. In SRL, residues involved in carbohydrateligand recognition are marked by open (primary) and filled (secondary) triangles. Residues identical between lectins areshaded. Secondary structural elements (β-strands F1–F4, S1–S6, α-helices H1 and H2) as determined by DSSP areshown.76
1154 Crystal Structure of the Sclerotium rolfsii Lectin
served in terms of amino acid sequence (Figure 6),and structural architecture. The only differences arein XCL, where there is phenylalanine instead oftyrosine and valine instead of isoleucine at positions27 and 78 (SRL numbering), respectively, which areconservative substitutions. In the primary bindingsite, GalNAc occupies similar positions in thestructures of the SRL–GalNAc and the ABL–GalNAc complexes, and it forms similar interactionswith each protein. Furthermore, superposition of theABL–TF antigen complex structure (PDB ID 1y2x)16
onto the SRL–GalNAc complex (Figure 8(a)) revealsthat the GalNAc moiety of the TF glycan structuresuperimposes very well with the GalNAc molecule
of the SRL complex. Inspection of the superimposedstructures suggests that the rest of the TF antigenmolecule can be accommodated in the SRL primarybinding site without any steric conflicts (Figure 8(a))by displacing four water molecules. All SRLresidues in this site adopt conformations very closeto their structural equivalents in ABL, favouringpotential hydrogen bond interactions with thehydroxyl groups O2, O3, and O4 of the galactosemoiety of the TF-antigen similar to those observed inthe ABL–TF-antigen complex.16 The serine moiety inSRL, as in the ABL complex, could form van derWaals interaction with His72 and Arg107 but notwith Tyr72 (the structural equivalent of Tyr74 in
Figure 7. Stereo view of thesuperposition of the Cα traces ofthe SRL (grey), ABL (red) and XCL(green) monomers.

Figure 8. Structural comparison of (a) the SRL–GalNAc complex (white) and the ABL–TF antigen complex (grey) atthe primary carbohydrate-binding site (an MPD molecule from the free SRL structure is also shown), and (b) the SRL–GlcNAc complex (grey) and the ABL–GlcNAc complex (white) at the secondary carbohydrate-binding site. (c) Amodel ofthe TF-antigen molecule in the SRL secondary carbohydrate-binding site.
1155Crystal Structure of the Sclerotium rolfsii Lectin
ABL) (Figure 8(a)). However, a small shift in theconformation of the phenyl ring might allow non-polar interactions of Tyr72 with the serine moiety ofthe TF-antigen similar to those observed for Tyr74 inthe ABL complex structure.16 The shape comple-mentarity between the surfaces of the TF-antigenand SRL in the primary binding site is 0.792.In the primary binding site of SRL molecule B of
the non-crystallographic dimer, crystal packing con-tacts impede binding of TF-antigen. Superposition ofthe ABL–TF-antigen complex structure onto the SRL–GalNAc complex shows that the serine moiety of TF-antigen clashes with residues from a symmetry-
related molecule in this site in SRL. This offers anexplanation for the rapid destruction of SRL crystalsupon soaking with solutions containing TF-antigen.ABL, like SRL, has a secondary carbohydrate-
binding site. However, ABL binds GlcNAc and notGalNAc at this site. The architecture of this site isconserved between the two lectins both in sequenceand structural terms with the exception of Asn100 ofSRL, which is Ile in ABL. Superposition of the SRL–GlcNAc complex structure onto the ABL–GlcNAcstructure reveals that the sugar binds similarly inboth lectins (Figure 8(b)), forming similar hydrogenbond and van der Waals interactions with SRL and

1156 Crystal Structure of the Sclerotium rolfsii Lectin
ABL residues. The secondary site is conserved alsoin XCL, with the exception again of residue Asn100(Gly104 in XCL), and the loop between residues 96and 101, which adopts a different conformationfrom the structurally equivalent loop in XCL (resi-dues 100-105) in SRL. Superposition of the GalNAcmoiety of TF-antigen onto the GalNAc molecule inthe SRL secondary binding site shows that this sitecannot accommodate the TF-antigen without anysignificant conformational changes, since the galac-tose moiety clashes with the side-chain of Tyr112and other residues from strand S4 (Figure 8(c)).Although the SRL, XCL and ABL crystals belong
to different space groups, the crystallographicasymmetric unit consists of two protein molecules.The dimer association for all three lectins is verysimilar and the r.m.s. deviations between the SRLand XCL dimers is 0.71 Å (Cα atoms), 0.68 Å (mainchain) and 0.98 Å (all atoms) for 276 equivalentresidues while between SRL and ABL is 0.61 Å (Cα
atoms), 0.59 Å (main-chain) and 0.95 Å (all atoms)for 282 equivalent residues. The r.m.s. deviationsbetween the non-crystallographic dimers of ABLand XCL is 0.90 Å (Cα atoms), 0.87 Å (main chain)and 1.09 Å (all atoms) for 278 equivalent residues.However, XCL and ABL are both tetramers insolution,15,16 while all experimental evidence so farsuggests that SRL is a dimer;32 hence, XCL and ABLhave four primary carbohydrate-binding sites incontrast to SRL that has two. Although the asym-metric unit of the XCL consists of two proteinmolecules, it has been proposed that its distinctbiological tetrameric assembly could be derived byapplying the crystallographic 2-fold axis.15 A tetra-meric form of ABL was found by co-crystallizingABL with GlcNAc (space group P43212).16 Interest-ingly, applying the crystallographic symmetryoperations to the SRL dimer of the asymmetricunit provides a tetrameric form, which superim-poses very well on the ABL and XCL tetramers withr.m.s. deviations of 0.74 Å and 0.76 Å (Cα atoms) for564 and 554 equivalent residues, respectively. Thecorresponding value between the ABL and XCLtetramer is 1.10 Å (Cα atoms) for 556 equivalentresidues. This indicates that SRL may have theability to form tetramers, in agreement withprevious studies showing that the purified lectin insolution (both at acidic and neutral pH) aggregatesand loses activity after prolonged storage at 4 °C.32
SRL does not display any significant sequence orstructural similarity with the other fungal lectinswith known structure (AAL, CGL2, Fve) or to anyother plant or animal lectin. Furthermore, SRL doesnot show any significant structural homology to anyTF-antigen-binding lectins like the Vicia villosaisolectin (VVLB4),50 PNA,51 jacalin,52 or ACL.53
The binding mode of TF-antigen in fungal lectinsseems to be quite different from the binding mode inother TF-specific plant lectins. Comparative struc-tural analysis of the SRL primary binding siteagainst the binding sites of other TF-antigen-bindingplant lectins PNA (PDB ID 2tep),51 jacalin (1m26),52ACL (1jlx)53 and VVLB4 (1n47)50 has revealed that
the structural architecture of the SRL site is quitedifferent. In peanut lectin, the galactose moiety ofthe TF-antigen is the anchoring point for thebinding, while the GalNAc part extends towardsthe solvent. In jacalin, the GalNAc part of the TF-antigen is the anchoring point for the ligandbinding; however, the sugar-binding mode is totallydifferent from that in the SRL primary binding site.In VVLB4 the galactopyranoside ring of the Tnresidue is stacked between the aromatic side-chainof a tyrosine residue and the aliphatic side-chain of aleucine residue,50 a binding mode different fromthat of SRL where GalNAc binds primarily withpolar interactions. Thus, the structural determinantsof TF-antigen specificity in SRL, jacalin, PNA, ACL,and VVLB4 are entirely different.Since, the three fungal lectins (SRL, ABL, XCL)
share structural topology, glycan specificity, twoconserved carbohydrate-binding sites and a com-mon species origin, they can form a new lectinsubfamily. On the basis of sequence comparisons(Figure 6), some more lectins of fungal origin seemto belong to this protein family.
Conclusions
The crystal structure of SRL has been determinedat near-atomic resolution. The primary, secondaryand tertiary structure of SRL has classified thisprotein as a new member of the recently identifiedfamily of TF-antigen-specific fungal lectins, and thedata for the free SRL are the highest resolution datafor a protein of this family. The structures of the SRLin complex with GalNAc and GlcNAc have revealedthe existence, the location and the structuralarchitecture of two distinct carbohydrate-bindingsites, designated primary and secondary. Our studyon the free and the carbohydrate-bound forms ofSRL provides a structural basis for carbohydraterecognition. The N-acetyl group of the carbohydrateligands is involved in both the primary and thesecondary binding sites in a network of polar andnon-polar interactions with the protein, providing astructural explanation for the higher specificity ofSRL for N-acetyl capping saccharides.34
Although we have observed binding of GalNAc tothe secondary binding site of SRL, which is sup-ported by an extensive network of interactions (bothpolar and non-polar) between the protein and theligand, and this was achieved by soaking SRLcrystals with a solution of a relatively low concentra-tion of GalNAc, in a short time (15 min), we cannotrule out the possibility that binding of GalNAc at thissite is an artefact of crystal packing, since itparticipates in crystal packing contacts and we didnot observe it in both molecules of the asymmetricunit, as we found for GlcNAc. Screening for newcrystallization conditions of SRL in the presence ofGalNAc is now in progress and may provide a wayto investigate further the specificity of this site.SRL shows a clear preference for GalNAc over
GlcNAc,34 and it seems able to differentiate struc-

1157Crystal Structure of the Sclerotium rolfsii Lectin
turally between the two sugar ligands. The primarybinding site recognizes GalNAc but not GlcNAc,whereas the secondary binding site recognizesGlcNAc. The structural basis of the molecularrecognition lies in the interactions of the epimerichydroxyl group (O4), which is the structuraldifference between the two carbohydrate molecules,and the hydroxyl group O6. In the primary bindingsite the hydroxyl group O4 of GalNAc forms (i) ahydrogen bond with the carbonyl oxygen of Gly48and (ii) a water-mediated hydrogen bond with theside chain of His70. The hydroxyl group O6, whichpoints to the opposite direction of O4 in the galactosemoiety, forms a hydrogen bond to a water molecule.In the secondary site the hydroxyl group O4 ofGlcNAc forms a hydrogen bond with the side chainatoms of As77. The hydroxyl groupO6,which pointsto the same direction with O4 in the glucosyl moietyofGlcNac, forms hydrogen bondswith the side chainatoms of Asp77 and Arg101.The endogenous receptors of SRL have been iden-
tified as glycosphingolipids that resemble glycosyli-nositolphosphoryl ceramides.33 These glycolipidsare quite long and might have parts that bind inboth the carbohydrate-binding sites of SRL, whichare 21 Å apart. The existence of two distinct carbo-hydrate-binding sites has been reported for anotherfungal lectin, the Psathyrella velutina mushroomlectin (PVL) where heparin/pectin binding occursat a site different from the N-acetylglucosamine/N-acetylneuraminic acid-specific PVL site.54 Thus, PVLis capable in binding, with high affinity, bothsialoglycans and GlcNAc-exposed glycans.55 Recentevidence established that ABL has dual distinctsugar specificity for Gal-exposed O-linked glycansand GlcNAc-exposed N-linked glycans.56 Therefore,the simultaneous binding of different carbohydrateligands in SRL needs to be investigated. Further-more, we are currently investigating the binding ofmodified Tantigen structures and different Tantigenconformations.Our results may offer an explanation for the
complex carbohydrate specificity displayed by PVLand other lectins. However, further investigation isrequired to establish the physiological role of thetwo carbohydrate-binding sites in the fungal lectins.This may assist in the elucidation of the carbohy-drate recognition mode of tumour-associated glyco-proteins by other similar fungal lectins.
‡http://wolf.bms.umist.ac.uk/naccess/
Materials and Methods
Purification of SRL from the fungus S. rolfsii andcrystallization conditions have been described.57,58 Thecrystal structure of SRLwas determined at 1.1 Å resolutionby the multiple isomorphous replacement (MIR) methodusing HoCl3 and Kr for derivatization. The holmiumderivative was prepared by soaking one SRL crystal for1 h in the crystallization buffer containing 100 mM HoCl3.The Kr derivative was prepared using a gas pressurizationapparatus at the EMBL-Hamburg outstation. GalNAc,GlcNAc and other chemicals were purchased fromSIGMA-Aldrich (Hellas) while the TF antigen (Galβ1-
3GalNAcα1-Thr) was provided by Dr O. Blixt through theConsortium for Functional Glycomics.59
Structure determination
Data for the holmium derivative to 1.65 Å resolutionwere collected using the Synchrotron Radiation Source atDaresbury (U.K.) on station PX9.6 (λ=0.87 Å) at 100 K.Diffraction data for the native, and for the Kr derivative to1.1 Å and 1.75 Å resolution, respectively, were collected at100 K using synchrotron radiation at the EMBL-Hamburgon station BW7B (λ=0.8441 Å and λ=0.8631 Å for thenative and the Kr derivative, respectively). Data for theSRL–GalNAc complex were collected to 1.96 Å resolutionat the EMBL-Hamburg on station X11 (λ=0.8128 Å) at100 K, while data for the SRL–GlcNAc complex werecollected at 100 K using the Synchrotron Radiation Sourceat Daresbury (U.K.) on station PX14.1 (λ=1.488 Å) to 1.7 Åresolution. Raw data images were indexed, integrated,corrected for Lorentz and polarization, and scaled usingthe HKL program suite.60 Intensities were truncated toamplitudes by the TRUNCATE program.61 Harker sec-tions of the isomorphous difference Patterson map fromthe Ho derivative calculated using the programs availablein the PHASES package62 showed two peaks correspond-ing to a single Ho atom per monomer in the asymmetricsite. Cross Fourier difference maps revealed the position ofone Kr atom in the asymmetric unit. Scaling of native andderivative data, heavy-atom refinement, MIR phasescalculations to 2.0 Å resolution were performed usingprograms within the CCP463 suite of programs andMLPHARE.63 Phase extension, and solvent flatteningwith the program DM64 generated new phases for thenative data to 1.1 Å resolution.An automatic construction of a poly-alanine model with
ARP/wARP produced a model of two monomers, each141 residues long.65 The quality of the high-resolutionelectron density σAmaps allowed the identification of theamino-acid sequence. However, difficulties arose in distin-guishing between residues Asp-Asn and Glu–Gln. Thesesequence ambiguities were resolved by tandem massspectrometry (the results will be published elsewhere)that also confirmed the initial X-ray derived sequence.Several cycles of refinement with CNS66 without any
non-crystallographic symmetry restraints improved themodel and the electron density maps allowed theincorporation of one tris(hydroxymethyl)aminomethane,three acetate, three MPD and 462 water molecules fromthe crystallization medium to the model. Finally, ten cyclesof anisotropic B-factor refinement were performed withthe program REFMAC.67 For the SRL–GalNAc and theSRL–GlcNAc complexes the structure of the free SRL wasused as a starting model and refinement was carried outwith REFMAC.67 The ligand molecules were incorporatedinto the model at the final stages of the refinement. Theprogram PROCHECKwas used to assess the quality of thefinal structure.68 Analysis of the Ramachandran (φ-ψ) plotshowed that all residues lie in the allowed regions. Detailsof the data collection, phasing and refinement statistics arepresented in Table 6. Accessible molecular surfaces werecalculated with the program NACCESS‡.
Protein Data Bank accession codes
The atomic coordinates of the free SRL, the SRL–GalNAc,and the SRL–GlcNAc complexes have been deposited in the
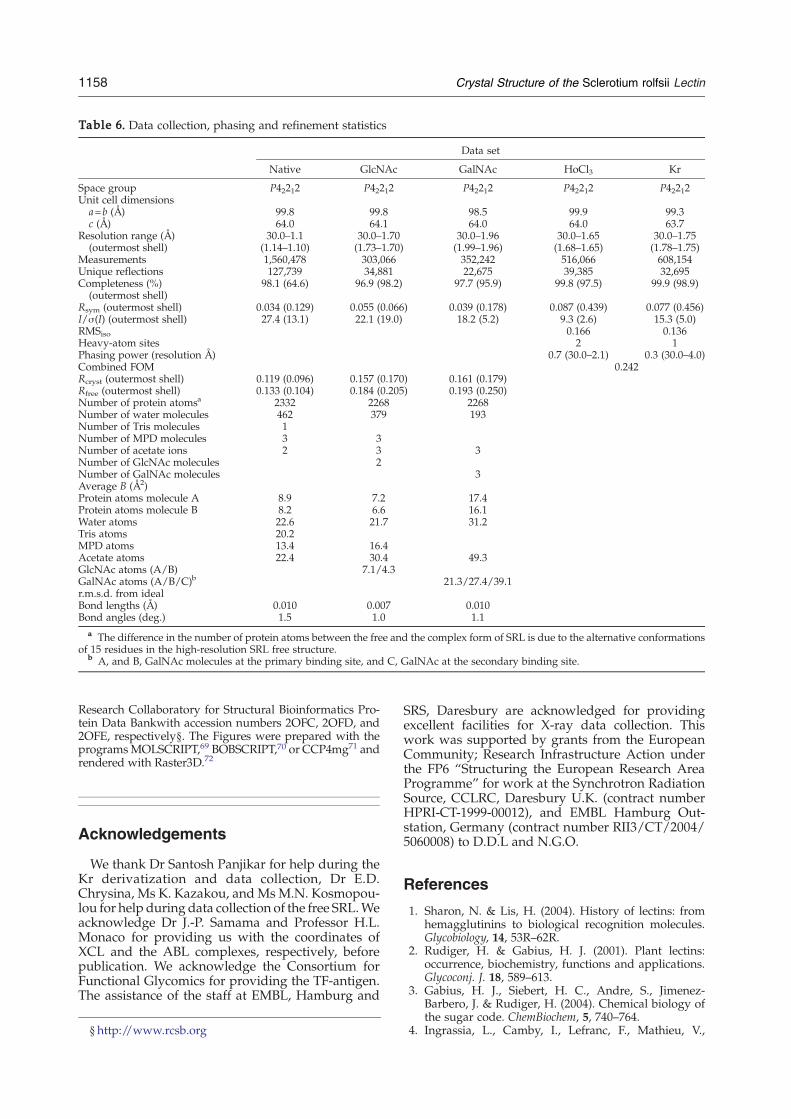
Table 6. Data collection, phasing and refinement statistics
Data set
Native GlcNAc GalNAc HoCl3 Kr
Space group P42212 P42212 P42212 P42212 P42212Unit cell dimensions
a=b (Å) 99.8 99.8 98.5 99.9 99.3c (Å) 64.0 64.1 64.0 64.0 63.7
Resolution range (Å)(outermost shell)
30.0–1.1(1.14–1.10)
30.0–1.70(1.73–1.70)
30.0–1.96(1.99–1.96)
30.0–1.65(1.68–1.65)
30.0–1.75(1.78–1.75)
Measurements 1,560,478 303,066 352,242 516,066 608,154Unique reflections 127,739 34,881 22,675 39,385 32,695Completeness (%)
(outermost shell)98.1 (64.6) 96.9 (98.2) 97.7 (95.9) 99.8 (97.5) 99.9 (98.9)
Rsym (outermost shell) 0.034 (0.129) 0.055 (0.066) 0.039 (0.178) 0.087 (0.439) 0.077 (0.456)I/σ(I) (outermost shell) 27.4 (13.1) 22.1 (19.0) 18.2 (5.2) 9.3 (2.6) 15.3 (5.0)RMSiso 0.166 0.136Heavy-atom sites 2 1Phasing power (resolution Å) 0.7 (30.0–2.1) 0.3 (30.0–4.0)Combined FOM 0.242Rcryst (outermost shell) 0.119 (0.096) 0.157 (0.170) 0.161 (0.179)Rfree (outermost shell) 0.133 (0.104) 0.184 (0.205) 0.193 (0.250)Number of protein atomsa 2332 2268 2268Number of water molecules 462 379 193Number of Tris molecules 1Number of MPD molecules 3 3Number of acetate ions 2 3 3Number of GlcNAc molecules 2Number of GalNAc molecules 3Average B (Å2)Protein atoms molecule A 8.9 7.2 17.4Protein atoms molecule B 8.2 6.6 16.1Water atoms 22.6 21.7 31.2Tris atoms 20.2MPD atoms 13.4 16.4Acetate atoms 22.4 30.4 49.3GlcNAc atoms (A/B) 7.1/4.3GalNAc atoms (A/B/C)b 21.3/27.4/39.1r.m.s.d. from idealBond lengths (Å) 0.010 0.007 0.010Bond angles (deg.) 1.5 1.0 1.1
a The difference in the number of protein atoms between the free and the complex form of SRL is due to the alternative conformationsof 15 residues in the high-resolution SRL free structure.
b A, and B, GalNAc molecules at the primary binding site, and C, GalNAc at the secondary binding site.
1158 Crystal Structure of the Sclerotium rolfsii Lectin
Research Collaboratory for Structural Bioinformatics Pro-tein Data Bankwith accession numbers 2OFC, 2OFD, and2OFE, respectively§. The Figures were prepared with theprogramsMOLSCRIPT,69 BOBSCRIPT,70 or CCP4mg71 andrendered with Raster3D.72
Acknowledgements
We thank Dr Santosh Panjikar for help during theKr derivatization and data collection, Dr E.D.Chrysina, Ms K. Kazakou, and MsM.N. Kosmopou-lou for help during data collection of the free SRL.Weacknowledge Dr J.-P. Samama and Professor H.L.Monaco for providing us with the coordinates ofXCL and the ABL complexes, respectively, beforepublication. We acknowledge the Consortium forFunctional Glycomics for providing the TF-antigen.The assistance of the staff at EMBL, Hamburg and
§http://www.rcsb.org
SRS, Daresbury are acknowledged for providingexcellent facilities for X-ray data collection. Thiswork was supported by grants from the EuropeanCommunity; Research Infrastructure Action underthe FP6 “Structuring the European Research AreaProgramme” for work at the Synchrotron RadiationSource, CCLRC, Daresbury U.K. (contract numberHPRI-CT-1999-00012), and EMBL Hamburg Out-station, Germany (contract number RII3/CT/2004/5060008) to D.D.L and N.G.O.
References
1. Sharon, N. & Lis, H. (2004). History of lectins: fromhemagglutinins to biological recognition molecules.Glycobiology, 14, 53R–62R.
2. Rudiger, H. & Gabius, H. J. (2001). Plant lectins:occurrence, biochemistry, functions and applications.Glycoconj. J. 18, 589–613.
3. Gabius, H. J., Siebert, H. C., Andre, S., Jimenez-Barbero, J. & Rudiger, H. (2004). Chemical biology ofthe sugar code. ChemBiochem, 5, 740–764.
4. Ingrassia, L., Camby, I., Lefranc, F., Mathieu, V.,

1159Crystal Structure of the Sclerotium rolfsii Lectin
Nshimyumukiza, P., Darro, F. & Kiss, R. (2006). Anti-Galectin compounds as potential anti-cancer drugs.Curr. Med. Chem. 13, 3513–3527.
5. Nangia-Makker, P., Conklin, J., Hogan, V. & Raz, A.(2002). Carbohydrate-binding proteins in cancer, andtheir ligands as therapeutic agents. Trends Mol. Med. 8,187–192.
6. Bies, C., Lehr, C. M. & Woodley, J. F. (2004). Lectin-mediated drug targeting: history and applications.Advan. Drug Deliv. Rev. 56, 425–435.
7. Kilpatrick, D. C. (2002). Animal lectins: a historicalintroduction and overview. Biochim. Biophys. Acta,1572, 187–197.
8. Cooper, D. N., Boulianne, R. P., Charlton, S., Farrell,E. M., Sucher, A. & Lu, B. C. (1997). Fungal galectins,sequence and specificity of two isolectins from Coprinuscinereus. J. Biol. Chem. 272, 1514–1521.
9. Loris, R. (2002). Principles of structures of animal andplant lectins. Biochim. Biophys. Acta, 1572, 198–208.
10. Wimmerova, M., Mitchell, E., Sanchez, J. F., Gautier,C. & Imberty, A. (2003). Crystal structure of fungallectin: six-bladed beta-propeller fold and novel fucoserecognition mode for Aleuria aurantia lectin. J. Biol.Chem. 278, 27059–27067.
11. Fujihashi, M., Peapus, D. H., Kamiya, N., Nagata, Y.& Miki, K. (2003). Crystal structure of fucose-specificlectin from Aleuria aurantia binding ligands at threeof its five sugar recognition sites. Biochemistry, 42,11093–11099.
12. Imberty, A., Mitchell, E. P. & Wimmerova, M. (2005).Structural basis of high-affinity glycan recognition bybacterial and fungal lectins. Curr. Opin. Struct. Biol. 15,524–534.
13. Ko, J. L., Hsu, C. I., Lin, R. H., Kao, C. L. & Lin, J. Y.(1995). A new fungal immunomodulatory protein,FIP-fve isolated from the edible mushroom, Flammu-lina velutipes and its complete amino acid sequence.Eur. J. Biochem. 228, 244–249.
14. Paaventhan, P., Joseph, J. S., Seow, S. V., Vaday, S.,Robinson, H., Chua, K. Y. & Kolatkar, P. R. (2003). A1.7Å structure of Fve, a member of the new fungalimmunomodulatory protein family. J. Mol. Biol. 332,461–470.
15. Birck, C., Damian, L., Marty-Detraves, C., Lougarre,A., Schulze-Briese, C., Koehl, P. et al. (2004). A newlectin family with structure similarity to actinoporinsrevealed by the crystal structure of Xerocomus chry-senteron lectin XCL. J. Mol. Biol. 344, 1409–1420.
16. Carrizo, M. E., Capaldi, S., Perduca, M., Irazoqui, F. J.,Nores, G. A. &Monaco, H. L. (2005). The antineoplasticlectin of the common edible mushroom (Agaricus bis-porus) has two binding sites, each specific for a differentconfiguration at a single epimeric hydroxyl. J. Biol.Chem. 280, 10614–10623.
17. Springer, G. F. (1997). Immunoreactive T and Tnepitopes in cancer diagnosis, prognosis, and immu-notherapy. J. Mol. Med. 75, 594–602.
18. Hakomori, S. (2001). Tumor-associated carbohydrateantigens defining tumor malignancy: basis for devel-opment of anti-cancer vaccines.Advan. Expt. Med. Biol.491, 369–402.
19. Yu, L., Fernig, D. G., Smith, J. A., Milton, J. D. &Rhodes, J. M. (1993). Reversible inhibition of prolif-eration of epithelial cell lines by Agaricus bisporus(edible mushroom) lectin. Cancer Res. 53, 4627–4632.
20. Trigueros, V., Lougarre, A., Ali-Ahmed, D., Rahbe, Y.,Guillot, J., Chavant, L. et al. (2003). Xerocomuschrysenteron lectin: identification of a new pesticidalprotein. Biochim. Biophys. Acta, 1621, 292–298.
21. Marty-Detraves, C., Francis, F., Baricault, L., Fournier,D. & Paquereau, L. (2004). Inhibitory action of a newlectin from Xerocomus chrysenteron on cell-substrateadhesion. Mol. Cell Biochem. 258, 49–55.
22. Walser, P. J., Haebel, P. W., Kunzler, M., Sargent, D.,Kues, U., Aebi, M. & Ban, N. (2004). Structure andfunctional analysis of the fungal galectin CGL2.Structure (Camb), 12, 689–702.
23. Yagi, F., Miyamoto, M., Abe, T., Minami, Y., Tadera, K.& Goldstein, I. J. (1997). Purification and carbohy-drate-binding specificity of Agrocybe cylindracea lectin.Glycoconj. J. 14, 281–288.
24. Walser, P. J., Haebel, P. W., Kunzler, M., Sargent, D.,Kues, U., Aebi, M. & Ban, N. (2004). Structure andfunctional analysis of the fungal galectin CGL2.Structure, 12, 689–702.
25. Ban, M., Yoon, H.-J., Demirkan, E., Utsumi, S.,Mikami, B. & Yagi, F. (2005). Structural basis of afungal galectin from Agrocybe cylindracea for recogniz-ing sialoconjugate. 351, 695–706.
26. Mancheno, J. M., Tateno, H., Goldstein, I. J.,Martinez-Ripoll, M. & Hermoso, J. A. (2005). Struc-tural analysis of the Laetiporus sulphureus hemolyticpore-forming lectin in complex with sugars. J. Biol.Chem. 280, 17251–17259.
27. Cioci, G., Mitchell, E. P., Chazalet, V., Debray, H.,Oscarson, S., Lahmann, M. et al. (2006). Beta-propellercrystal structure of Psathyrella velutina lectin: anintegrin-like fungal protein interacting with mono-saccharides and calcium. J. Mol. Biol. 357, 1575–1591.
28. Rutenber, E., Ready, M. & Robertus, J. D. (1987). Struc-ture and evolution of ricin B chain.Nature, 326, 624–626.
29. Tahirov, T. H., Lu, T. H., Liaw, Y. C., Chen, Y. L. &Lin, J. Y. (1995). Crystal structure of abrin-a at 2.14 Å.J. Mol. Biol. 250, 354–367.
30. Inoue, K., Sobhany, M., Transue, T. R., Oguma, K.,Pedersen, L. C. & Negishi, M. (2003). Structuralanalysis by X-ray crystallography and calorimetry ofa haemagglutinin component (HA1) of the progenitortoxin from Clostridium botulinum. Microbiology, 149,3361–3370.
31. Springer, T. A. (1997). Folding of the N-terminal,ligand-binding region of integrin alpha-subunits intoa beta-propeller domain. Proc. Natl Acad. Sci. USA, 94,65–72.
32. Swamy, B. M., Hedge, G. V., Naik, R. S. & Inamdar, S.R. (2001). T-antigen binding lectin from the phyto-pathogenic fungus Sclerotium rolfsii. Lect. Biol. Biochem.Clin. Biochem. 15, [Available online at http://plab.ku.dk/tcbh/Lectins15/Sawmy/paper.htm].
33. Swamy, B. M., Bhat, A. G., Hegde, G. V., Naik, R. S.,Kulkarni, S. & Inamdar, S. R. (2004). Immunolocaliza-tion and functional role of Sclerotium rolfsii lectin indevelopment of fungus by interaction with its endog-enous receptor. Glycobiology, 14, 951–957.
34. Wu, A. M., Wu, J. H., Tsai, M.-S., Hedge, G. V.,Inamdar, S. R., Swamy, B. M. & Herp, A. (2001).Carbohydrate specificity of a lectin isolated from thefungus Sclerotium rolfsii. Life Sci. 69, 2039–2050.
35. Milton, J. D., Fernig, D. G. & Rhodes, J. M. (2001). Useof a biosensor to determine the binding kinetics of fivelectins for galactosyl-N-acetylgalactosamine. Glyco-conj. J. 18, 565–569.
36. Lawrence, M. C. & Colman, P. M. (1993). Shapecomplementarity at protein/protein interfaces. J. Mol.Biol. 234, 946–950.
37. Pearson, W. R. & Lipman, D. J. (1988). Improved toolsfor biological sequence comparison. Proc. Natl Acad.Sci. USA, 85, 2444–2448.

1160 Crystal Structure of the Sclerotium rolfsii Lectin
38. Apweiler, R., Bairoch, A., Wu, C. H., Barker, W. C.,Boeckmann, B., Ferro, S. et al. (2004). UniProt: theUniversal Protein knowledgebase. Nucl. Acids Res. 32,D115–D119.
39. Crenshaw, R. W., Harper, S. N., Moyer, M. & Privalle,L. S. (1995). Isolation and characterization of a cDNAclone encoding a lectin gene from Agaricus bisporus.Plant Physiol. 107, 1465–1466.
40. Johansson, T., Le Quere, A., Ahren, D., Soderstrom, B.,Erlandsson, R., Lundeberg, J. et al. (2004). Transcrip-tional responses ofPaxillus involutus andBetula penduladuring formation of ectomycorrhizal root tissue. Mol.Plant Microbe Interact. 17, 202–215.
41. Iijima, N., Yoshino, H., Ten, L. C., Ando, A., Watanabe,K. & Nagata, Y. (2002). Two genes encoding fruit bodylectins of Pleurotus cornucopiae: sequence similaritywith the lectin of a nematode-trapping fungus. Biosci.Biotechnol. Biochem. 66, 2083–2089.
42. Rosen, S., Kata, M., Persson, Y., Lipniunas, P. H.,Wikstrom, M., Van Den Hondel, M. J. et al. (1996).Molecular characterization of a saline-soluble lectinfrom a parasitic fungus. Extensive sequence simila-rities between fungal lectins. Eur. J. Biochem. 238,822–829.
43. Silar, P., Barreau, C., Debuchy, R., Kicka, S., Turcq, B.,Sainsard-Chanet, A. et al. (2003). Characterization ofthe genomic organization of the region bordering thecentromere of chromosome V of Podospora anserinaby direct sequencing. Fungal Genet. Biol. 39, 250–263.
44. Galagan, J. E., Calvo, S. E., Borkovich, K. A., Selker,E. U., Read, N. D., Jaffe, D. et al. (2003). The genomesequence of the filamentous fungus Neurospora crassa.Nature, 422, 859–868.
45. Holm, L. & Sander, C. (1993). Protein structure com-parison by alignment of distance matrices. J. Mol. Biol.233, 123–138.
46. Berman, H. M., Westbrook, J., Feng, Z., Gilliland, G.,Bhat, T. N., Weissig, H. et al. (2000). The Protein DataBank. Nucl. Acids Res. 28, 235–242.
47. Yu, L. G., Fernig, D. G., White, M. R., Spiller, D. G.,Appleton, P., Evans, R. C. et al. (1999). Edible mush-room (Agaricus bisporus) lectin, which reversibly in-hibits epithelial cell proliferation, blocks nuclearlocalization sequence-dependent nuclear protein im-port. J. Biol. Chem. 274, 4890–4899.
48. Yu, L. G., Andrews, N., Weldon, M., Gerasimenko,O. V., Campbell, B. J., Singh, R. et al. (2002). An N-terminal truncated form of Orp150 is a cytoplasmicligand for the anti-proliferative mushroom Agaricusbisporus lectin and is required for nuclear localizationsequence-dependent nuclear protein import. J. Biol.Chem. 277, 24538–24545.
49. Rosen, S., Bergstrom, J., Karlsson, K. A. & Tunlid, A.(1996). A multispecific saline-soluble lectin from theparasitic fungus Arthrobotrys oligospora. Similarities inthe binding specificities compared with a lectin fromthe mushroom agaricus bisporus. Eur. J. Biochem. 238,830–837.
50. Babino, A., Tello, D., Rojas, A., Bay, S., Osinaga, E. &Alzari, P. M. (2003). The crystal structure of a plantlectin in complex with the Tn antigen. FEBS Letters,536, 106–110.
51. Ravishankar, R., Ravindran,M., Suguna, K., Surolia, A.& Vijayan, M. (1997). Crystal structure of the peanutlectin - T-antigen complex. Carbohydrate specificiftygenerated by water bridges. Curr. Sci. 72, 855–861.
52. Jeyaprakash, A. A., Geetha Rani, P., BanuprakashReddy, G., Banumathi, S., Betzel, C., Sekar, K. et al.(2002). Crystal structure of the jacalin-T-antigen
complex and a comparative study of lectin-T-antigencomplexes. J. Mol. Biol. 321, 637–645.
53. Transue, T. R., Smith, A. K., Mo, H., Goldstein, I. J. &Saper, M. A. (1997). Structure of benzyl T-antigendisaccharide bound to Amaranthus caudatus agglu-tinin. Nature Struct. Biol. 4, 779–783.
54. Ueda, H., Saitoh, T., Kojima, K. & Ogawa, H. (1999).Multi-specificity of a Psathyrella velutina mushroomlectin: heparin/pectin binding occurs at a site differentfrom the N-acetylglucosamine/N-acetylneuraminicacid-specific site. J. Biochem. 126, 530–537.
55. Ueda, H., Matsumoto, H., Takahashi, N. & Ogawa,H. (2002). Psathyrella velutina mushroom lectin exhi-bits high affinity toward sialoglycoproteins posses-sing terminal N-acetylneuraminic acid alpha 2,3-linked to penultimate galactose residues of trisialylN-glycans. Comparison with other sialic acid-specificlectins. J. Biol. Chem. 277, 24916–24925.
56. Nakamura-Tsuruta, S., Kominami, J., Kuno, A. &Hirabayashi, J. (2006). Evidence that Agaricus bis-porus agglutinin (ABA) has dual sugar-binding speci-ficity. Biochem. Biophys. Res. Commun. 347, 215–220.
57. Swamy, B. M., Hedge, G. V., Naik, R. S. & Inamdar, S.R. (2000). T-antigen binding lectin from the phyto-pathogenic fungus Sclerotium rolfsii. Lect. Biol. Biochem.Clin. Biochem. 14, 45–55.
58. Leonidas, D. D., Swamy, B. M., Bhat, A. G., Inamdar,S. R., Kosmopoulou, M. N., Chrysina, E. D. &Oikonomakos, N. G. (2003). Crystallization and pre-liminary X-ray crystallographic analysis of Sclerotiumrolfsii lectin. Acta Crystallog. sect. D, 59, 363–365.
59. Blixt, O., Allin, K., Pereira, L., Datta, A. & Paulson, J. C.(2002). Efficient chemoenzymatic synthesis of O-linked sialyl oligosaccharides. J. Am. Chem. Soc. 124,5739–5746.
60. Otwinowski, Z. & Minor, W. (1997). Processing ofX-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
61. French, S. & Wilson, K. S. (1978). On the treatment ofthe negative intensity observations. Acta Crystallog.sect. A, 34, 517–525.
62. Furey, W. & Swaminathan, S. (1997). PHASES-95: aprogram package for the processing and analysis ofdiffraction data from macromolecules. Methods Enzy-mol. 277, 540.
63. Collaborative Computing Project Number 4. (1994).The CCP4 suite: programs for protein crystallography.Acta Crystallog. sect. D, 50, 760–763.
64. Cowtan, K. (1994). Density modification package.Joint CCP4 and ESF-EACBM Newsletter on ProteinCrystallography, 31, 34–38.
65. Perrakis, A., Morris, R. & Lamzin, V. S. (1999). Au-tomated protein model building combined withiterative structure refinement. Nature, Struct. Biol. 6,458–463.
66. Brünger, A. T., Adams, P. D., Clore, G. M., DeLano,W. L., Gros, P., GrosseKunstleve, R. W. et al. (1998).Crystallography and NMR system: a new softwaresuite for macromolecular structure determination.Acta Crystallogr. D, 54, 905–921.
67. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. (1997).Refinement of macromolecular structures by themaximum-likelihood method. Acta Crystallog. sect. D,53, 240–255.
68. Laskowski, R. A., MacArthur, M. W., Moss, D. S. &Thornton, J. M. (1993). PROCHECK - a program tocheck the stereochemical quality of protein structures.J. Appl. Crystallog. 26, 283–291.
69. Kraulis, P. J. (1991). MOLSCRIPT - a program to

1161Crystal Structure of the Sclerotium rolfsii Lectin
produce both detailed & schematic plots of proteinstructures. J. Appl. Crystallog. 24, 946–950.
70. Esnouf, R. M. (1997). An extensively modified versionof Molscript that includes greatly enhanced coloringcapabilities. J. Mol. Graph. Model. 15, 132–134.
71. Potterton, L., McNicholas, S., Krissinel, E., Gruber, J.,Cowtan, K., Emsley, P. et al. (2004). Developments inthe CCP4 molecular-graphics project. Acta Crystallog.sect. D, 60, 2288–2294.
72. Merritt, E. A. & Bacon, D. J. (1997). Raster3D:photorealistic molecular graphics. Macromol. Crystal-log. B, 277, 505–524.
73. Bond, C. S. (2003). TopDraw: a sketchpad for proteinstructure topology cartoons. Bioinformatics, 19, 311–312.
74. Nicholls, A. & Honig, B. (1991). A rapid finite dif-ference algorithm, utilising successive over relaxationto solve the Poisson-Boltzmann equation. J. Comput.Chem. 12, 435–445.
75. Thompson, J. D., Higgins, D. G. & Gibson, T. J.(1994). CLUSTAL W: improving the sensitivity ofprogressive multiple sequence alignment throughsequence weighting, position-specific gap penaltiesand weight matrix choice. Nucl. Acids Res. 22,4673–4680.
76. Kabsch, W. & Sanders, C. (1983). Dictionary of theprotein secondary structure: pattern recognition ofhydrogen bonded and geometrical features. Biopoly-mers, 22, 2577–2637.
Edited by R. Huber
(Received 5 January 2007; received in revised form 26 February 2007; accepted 27 February 2007)Available online 7 March 2007




















