
Day and night heat stress trigger different transcriptomic ...

Spo0J regulates the oligomeric state of Soj to triggerits switch from an activator to an inhibitor of DNAreplication initiationmmi_7507 1089..1100
Graham Scholefield,1 Rachel Whiting,2
Jeff Errington1 and Heath Murray1*1Centre for Bacterial Cell Biology, Institute for Cell andMolecular Biosciences, Newcastle University, NewcastleUpon Tyne NE2 4AX, UK.2Sir William Dunn School of Pathology, University ofOxford, Oxford OX1 3RE, UK.
Summary
Control of DNA replication initiation is essential forbacterial cells to co-ordinate the faithful replicationand segregation of their genetic material. The Bacillussubtilis ATPase Soj is a dynamic protein that regu-lates DNA replication initiation by either inhibiting oractivating the DNA replication initiator protein DnaA.Here we report that the key event which switches Sojregulatory activity is a transition in its oligomericstate from a monomer to an ATP-dependenthomodimer capable of DNA binding. We show that theDNA binding activity of the Soj dimer is required bothfor activation of DNA replication initiation and forinteraction with Spo0J. Finally, we demonstrate thatSpo0J inhibits Soj dimerization by stimulating SojATPase activity. The data provide a molecular expla-nation for the dichotomous regulatory activities ofSoj, as well as assigning unique Soj conformations todistinct cellular localization patterns. We discuss howthe regulation of Soj ATPase activity by Spo0J couldbe utilized to control the initiation of DNA replicationduring the cell cycle.
Introduction
Initiation of DNA replication must be co-ordinated with thecell cycle to ensure accurate chromosome segregation. Inbacteria there are often multiple overlapping systems thatorchestrate DNA replication initiation by regulating theactivity of the DNA replication initiator protein DnaA(Katayama et al., 2010). In the model bacterium Bacillus
subtilis the ATPase Soj acts as a molecular switch toregulate the activity of the DNA replication initiator proteinDnaA (Murray and Errington, 2008). Soj is a member ofthe ParA family of ATPases and its gene is transcribedfrom a highly conserved module composed of soj(parA),spo0J(parB) and parS (Gerdes et al., 2000; Livny et al.,2007). parS sites are cis-acting DNA sequence motifslocated proximal to the DNA replication origin that nucle-ate the spreading of Spo0J into flanking regions of DNA tocreate large nucleoprotein complexes (Lin and Gross-man, 1998; Murray et al., 2006; Breier and Grossman,2007).
Structural and biochemical analysis of Soj orthologueshas led to the model that these proteins are ATP-dependent ‘sandwich’ dimers (Leonard et al., 2005;Hester and Lutkenhaus, 2007). In the presence of ADP (orin the absence of nucleotide) Soj proteins are monomeric.ATP binding by Soj facilitates dimerization and is requiredfor Soj to bind DNA. Soj DNA binding activity appears tobe non-specific, highly cooperative and mediated by con-served arginine residues located on one face of the dimer.
Under several conditions, such as in the absence of theregulator Spo0J, when artificially overexpressed, or as amutant allele (SojD40A) that cannot hydrolyse ATP, Soj acti-vates DnaA to stimulate DNA replication initiation (Oguraet al., 2003; Lee and Grossman, 2006; Murray and Err-ington, 2008). Under other conditions, such as whenweakly expressed or when converted to a mutant allele(SojG12V) that cannot bind to DNA, Soj inhibits DnaA torepress DNA replication initiation (Ogura et al., 2003;Murray and Errington, 2008). Although the ability of Soj todifferentially regulate DnaA has been established, themechanism by which Soj switches between its regulatorystates could not been determined because biochemicalcharacterization of the Soj mutants utilized in thesestudies was incomplete.
In this report we have investigated the contributions ofdimerization and DNA binding to the regulatory activitiesof B. subtilis Soj. Our data indicate that DNA replicationinitiation is inhibited by monomeric Soj and activated bydimeric Soj. We find that negative regulation by Soj doesnot require its DNA binding activity, while positive regula-tion by Soj does. We go on to show that the regulatory
Accepted 8 December, 2010. *For correspondence. E-mail [email protected]; Tel. (+44) 191 208 3233; Fax (+44) 191 208 3205.
Molecular Microbiology (2011) 79(4), 1089–1100 � doi:10.1111/j.1365-2958.2010.07507.xFirst published online 16 January 2011
© 2011 Blackwell Publishing Ltd
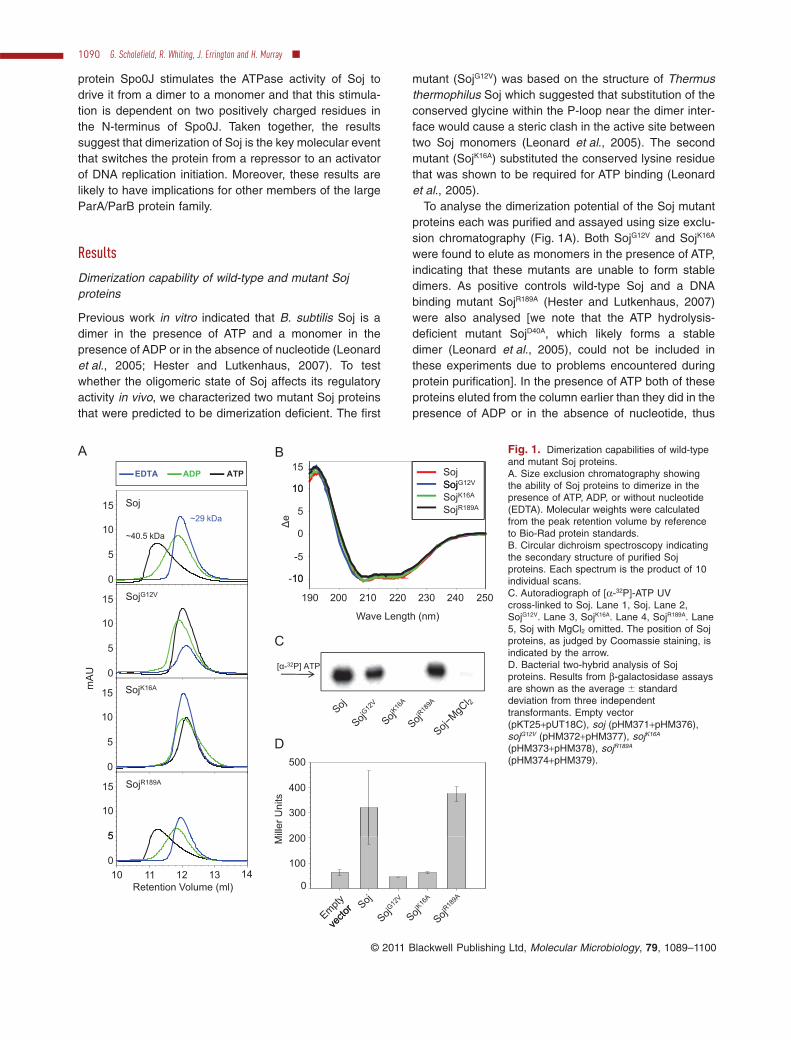
protein Spo0J stimulates the ATPase activity of Soj todrive it from a dimer to a monomer and that this stimula-tion is dependent on two positively charged residues inthe N-terminus of Spo0J. Taken together, the resultssuggest that dimerization of Soj is the key molecular eventthat switches the protein from a repressor to an activatorof DNA replication initiation. Moreover, these results arelikely to have implications for other members of the largeParA/ParB protein family.
Results
Dimerization capability of wild-type and mutant Sojproteins
Previous work in vitro indicated that B. subtilis Soj is adimer in the presence of ATP and a monomer in thepresence of ADP or in the absence of nucleotide (Leonardet al., 2005; Hester and Lutkenhaus, 2007). To testwhether the oligomeric state of Soj affects its regulatoryactivity in vivo, we characterized two mutant Soj proteinsthat were predicted to be dimerization deficient. The first
mutant (SojG12V) was based on the structure of Thermusthermophilus Soj which suggested that substitution of theconserved glycine within the P-loop near the dimer inter-face would cause a steric clash in the active site betweentwo Soj monomers (Leonard et al., 2005). The secondmutant (SojK16A) substituted the conserved lysine residuethat was shown to be required for ATP binding (Leonardet al., 2005).
To analyse the dimerization potential of the Soj mutantproteins each was purified and assayed using size exclu-sion chromatography (Fig. 1A). Both SojG12V and SojK16A
were found to elute as monomers in the presence of ATP,indicating that these mutants are unable to form stabledimers. As positive controls wild-type Soj and a DNAbinding mutant SojR189A (Hester and Lutkenhaus, 2007)were also analysed [we note that the ATP hydrolysis-deficient mutant SojD40A, which likely forms a stabledimer (Leonard et al., 2005), could not be included inthese experiments due to problems encountered duringprotein purification]. In the presence of ATP both of theseproteins eluted from the column earlier than they did in thepresence of ADP or in the absence of nucleotide, thus
Fig. 1. Dimerization capabilities of wild-typeand mutant Soj proteins.A. Size exclusion chromatography showingthe ability of Soj proteins to dimerize in thepresence of ATP, ADP, or without nucleotide(EDTA). Molecular weights were calculatedfrom the peak retention volume by referenceto Bio-Rad protein standards.B. Circular dichroism spectroscopy indicatingthe secondary structure of purified Sojproteins. Each spectrum is the product of 10individual scans.C. Autoradiograph of [a-32P]-ATP UVcross-linked to Soj. Lane 1, Soj. Lane 2,SojG12V. Lane 3, SojK16A. Lane 4, SojR189A. Lane5, Soj with MgCl2 omitted. The position of Sojproteins, as judged by Coomassie staining, isindicated by the arrow.D. Bacterial two-hybrid analysis of Sojproteins. Results from b-galactosidase assaysare shown as the average � standarddeviation from three independenttransformants. Empty vector(pKT25+pUT18C), soj (pHM371+pHM376),sojG12V (pHM372+pHM377), sojK16A
(pHM373+pHM378), sojR189A
(pHM374+pHM379).
1090 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

indicating ATP-dependent dimerization. Far-UV circulardichroism spectroscopy showed that the purified Soj pro-teins had indistinguishable secondary structure composi-tions (Fig. 1B), indicating that the inability of SojG12V andSojK16A to dimerize was not due to protein misfolding.Moreover, UV cross-linking confirmed that SojG12V wasable to bind ATP (Fig. 1C), consistent with the model thatthis mutant is monomeric because the substitution ofglycine for valine introduces a steric clash at the dimerinterface (Leonard et al., 2005).
We found that the Soj dimerization deficient mutantswere unable to stably interact with plasmid DNA using anelectrophoretic mobility shift assay (Fig. S1A), consistentwith previous results demonstrating that the DNA bindingactivity of Soj is ATP-dependent (Leonard et al., 2005;Hester and Lutkenhaus, 2007). The DNA binding activityof Soj appears highly cooperative and it has been foundthat homologous ParA proteins can either form nucleopro-tein filaments (T. thermophilus; Leonard et al., 2005) orassemble into filaments in the absence of DNA (Caulo-bacter crescentus; Ptacin and Shapiro, 2010). We won-dered whether the DNA binding activity of Soj mightinvolve filament formation and therefore we evaluated theability of Soj to form filaments in vitro using 90 degreeangle light scattering (Fig. S1B) and electron microscopy(data not shown). We were unable to detect the assemblyof Soj into higher-order complexes using either approach(with or without DNA), and we note that Soj complexeslarger than dimers were not observed by size exclusionchromatography (data not shown). However, we cannotrule out the possibility that Soj forms filaments underdifferent reaction conditions.
The ability of the Soj proteins to dimerize in vivo wasprobed using an Escherichia coli two-hybrid system thatutilizes a split adenylate cyclase to activate lacZ geneexpression when the two halves of the enzyme arebrought together (Karimova et al., 1998). Wild-type andmutant soj alleles were fused to the adenylate cyclasefragments and the ability to dimerize was assayed using ab-galactosidase assay. An interaction was detected for thewild-type Soj protein and the SojR189A mutant, but not forthe SojG12V and SojK16A mutants (Fig. 1D). We note thatpreviously both the SojG12V and SojK16A mutants have beenshown to interact with DnaA using this two-hybrid assay(Murray and Errington, 2008), suggesting that thesemutants are properly expressed and folded. Takentogether with the in vitro analyses, the results indicate thatboth the SojG12V and SojK16A mutants cannot dimerize andinstead exist as monomers.
Activity of Soj proteins on DNA replication initiation
To analyse the activities of the wild-type and mutant Sojproteins on DNA replication initiation, each allele was
transferred into the chromosome of a Dsoj strain underthe control of a xylose-inducible promoter. Soj proteinswere overexpressed and their effects on DNA replicationinitiation were determined by marker frequency analysis(we hypothesized that high levels of protein expressionwould have two advantages: accentuating the regulatoryactivity of Soj proteins and promoting Soj dimerization).Compared with the uninduced controls, both of the Sojproteins that were capable of dimerization (wild type andSojR189A) showed increased DNA replication initiation,while both of the Soj proteins that were unable to dimer-ize (SojG12V and SojK16A) inhibited DNA replication initia-tion (Fig. 2A). Western blot analysis showed that all ofthe Soj proteins were expressed to a similar level, sig-nifying that the difference in protein activities was notdue to variations in protein concentration (Fig. 2B). Inter-estingly, the SojK16A mutant was not as potent an inhibi-tor of DNA replication initiation as the SojG12V mutant,suggesting that nucleotide binding enhances the activityof the Soj monomer. Additionally, the SojR189A mutantwas not as potent an activator of DNA replication initia-tion as the wild-type protein, indicating that DNA binding
Fig. 2. Effect of wild-type and mutant Soj proteins on DNAreplication initiation.A. The oriC-to-terminus ratio of each Soj protein was determinedusing marker frequency analysis. Cells were grown in CH mediumat 30°C with or without xylose (1%). The average � standarddeviation from three technical replicates is shown. The dashed linerepresents the average ratio for the parent strain.B. Western blot analysis of Soj protein overexpression. Strainswere grown as described above except that xylose was added afterthe cultures had reached the exponential growth phase. Emptyvector (HM359), soj (HM558), sojG12V (HM240), sojK16A (HM625),sojR189A (HM626).
Oligomeric status switches Soj regulatory activity 1091
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

facilitates positive regulation of DNA replication by Soj(see next section below).
To confirm the results of the marker frequency analysis,DNA replication was assayed by observing nucleoid local-ization within single cells. Overexpression of either SojG12V
(Fig. S2C) or SojK16A (Fig. S2D) caused a dramaticdecrease in the number of nucleoids per cell, consistentwith severe inhibition of DNA replication initiation. Con-versely, overexpression of wild-type Soj (Fig. S2B) led tocells containing multiple nucleoids, consistent with activa-tion of DNA replication initiation. Overexpression ofSojR189A (Fig. S2E) did not appear to appreciably affectnucleoid size or distribution, consistent with the modestincrease in DNA replication initiation determined bymarker frequency analysis.
The DNA binding activity of Soj is required foractivation of DnaA
The overexpression analysis described above suggestedthat the DNA binding activity of Soj facilitates maximalactivation of DNA replication initiation. In order to furtherassess the requirement of DNA binding for positive regu-lation by Soj, we investigated both the localization and theregulatory activity of the SojR189A mutant expressed fromits endogenous locus.
Wild-type GFP–Soj localizes at septa and forms foci atthe oriC region of the chromosome (Fig. 3A). In a Dspo0Jmutant GFP–Soj relocalizes to the nucleoid (Fig. 3B). Thelocalization of the GFP–SojR189A mutant was indistinguish-able from GFP–Soj in the presence of Spo0J (Fig. 3C), but
Fig. 3. Soj DNA binding activity is requiredto activate DNA replication initiation.A–D. The localization of GFP–SojR189A in thepresence and absence of Spo0J wasobserved using epifluorescence microscopy.Cells were grown in CH medium at 30°C. (A)gfp-soj (HM4), (B) gfp-soj Dspo0J (HM13), (C)gfp-sojR189A (HM308), (D) gfp-sojR189A Dspo0J(HM310).E. The oriC-to-terminus ratio of SojR189A wasdetermined using marker frequency analysis.Cells were grown in CH medium at 30°C. Theaverage � standard deviation from threetechnical replicates is shown. The dashed linerepresents the average ratio for the parentstrain. Wild type (HM34), Dspo0J (HM42),Dsoj (HM161), Dsoj Dspo0J (HM183), sojR189A
(HM416), sojR189A Dspo0J (HM417).F. The oriC-to-terminus ratio of SojD40A,R189A
was determined using marker frequencyanalysis. Cells were grown in CH medium at30°C. The average � standard deviation fromthree technical replicates is shown. Thedashed line represents the average ratio forthe parent strain. Wild type (HM34), sojD40A
(HM36), sojR189A (HM416), sojD40A,R189A
(HM696).G. The oriC-to-terminus ratio of SojR189A
(HM626) expressed at increasingconcentrations was determined using markerfrequency analysis. Cells were grown in CHmedium at 30°C with increasingconcentrations of xylose (0%, 0.0016%,0.008%, 0.04%, 0.2%, 1%). Theaverage � standard deviation from threetechnical replicates is shown. Below theamount of SojR189A expressed at each xyloseconcentration was determined by Westernblot and compared with the expression ofSojR189A from its native locus (HM416; seehatched box). Western blot analysis of DivIVAwas used as a loading standard.
1092 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

in contrast GFP–SojR189A did not relocalize to the nucleoidin a Dspo0J mutant (Fig. 3D). These results indicate thatGFP–SojR189A maintains the ability to interact with factorsrequired for its localization at septa and at oriC, but that it isunable to bind to the nucleoid, consistent with the modelthat SojR189A is unable to bind to DNA in vivo.
Next, marker frequency analysis was used to determinethe effect of SojR189A on DNA replication initiation in theabsence of Spo0J (Fig. 3E). In a Dspo0J mutant the wild-type Soj protein causes an increase in the ratio of DNA atthe origin of replication compared with DNA at the termi-nus of replication, indicating that Soj activates DNA repli-cation initiation. In contrast, the SojR189A mutant did notactivate DNA replication in a Dspo0J mutant, indicatingthat the DNA binding activity of Soj is required for positiveregulation. Moreover, the SojR189A mutant displayed anori/ter ratio that was below the wild-type ratio, rather thanthe increased ori/ter ratio characteristic of a Dsoj mutant.Although the ~10% decrease in the ori/ter ratio observedin the SojR189A mutant is modest, it has a profound effecton cell cycle regulation and produces a large number ofcells that lack DNA (0.49% for wild type compared with6.8% for SojR189A; Fig. S3A). We suggest that this pheno-type arises due to delayed DNA segregation as a conse-quence of a decreased rate of DNA replication initiation;consequently, as the cell wall elongates it creates a gapbetween the cell pole and the nucleoid where the celldivision machinery can assemble, leading to the produc-tion of ‘stumpy’ cells lacking DNA. Taken together theseresults suggest that the DNA binding activity of Soj is notrequired for it to inhibit DnaA. To substantiate this hypoth-esis we created a sojG12V,R189A double mutant under thecontrol of a xylose-inducible promoter. Overexpression ofthe SojG12V,R189A protein caused the same inhibition of DNAreplication initiation as SojG12V (Fig. S2C and F), verifyingthat the DNA binding activity of Soj is not required fornegative regulation of DNA replication initiation.
Interestingly, we observed that the effect of the SojR189A
mutant on DNA replication initiation was dependent uponthe assay: SojR189A activated DNA replication initiationwhen overexpressed (Fig. 2A) but inhibited DNA replica-tion initiation when expressed from its native locus(Fig. 3E). Although this mutant clearly has the ability todimerize (Fig. 1), its localization and activity suggestedthat it was actually monomeric under the latter condition(Fig. 3). To investigate this possibility we constructed asojD40A,R189A double mutant that was expressed from thenative locus; this aspartic acid is required for ATP hydroly-sis and it has been shown that mutation of this residuelocks T. thermophilus Soj as a dimer (Leonard et al.,2005). If natively expressed SojR189A is dimeric in vivo,then the SojD40A,R189A double mutant should have proper-ties similar to the SojR189A single mutant. We tested theeffect of the double mutant on DNA replication initiation
using marker frequency analysis and on protein localiza-tion using a GFP tag. In contrast to the SojR189A singlemutant, we found that the SojD40A,R189A double mutant dis-played a significantly elevated initiation rate (Fig. 3F) andits localization was clearly different from the GFP–SojR189A
single mutant (Fig. S3B). The SojD40A,R189A double mutantwas expressed to a similar level as the wild type andthe Soj single mutants (Fig. S4A), indicating that theobserved increase in DNA replication was not due tochanges in protein expression. We did find that the GFP–SojD40A,R189A double mutant was consistently underex-pressed compared with other GFP–Soj proteins (Fig.S4B). However, we did not detect any degradation of thisfusion protein, indicating that the observed diffuse local-ization pattern was not due to free GFP. Moreover, wenote that low basal expression of GFP–Soj from thexylose-inducible promoter does not preclude this chimerafrom clearly localizing at cell poles (Murray and Errington,2008; Fig. S1A), suggesting that underexpression cannotaccount for protein mislocalization.
The results using the SojD40A,R189A double mutant areinconsistent with the model that SojR189A is dimeric whenexpressed from its native locus, and they suggested thatthe overexpression of SojR189A might alter the oligomericstatus and activity of the protein. To test this hypothesiswe titrated the level of inducer to incrementally increasethe expression of SojR189A from the xylose-inducible pro-moter and correlated this with the rate of DNA replicationinitiation. Figure 3G shows that at low induction levelsSojR189A decreases the rate of DNA replication initiation.Importantly, the level of protein expression observedunder these conditions was similar to the expression ofSojR189A from its native locus (hatched box in Fig. 3G). Asthe expression level of SojR189A was further increased, thisresulted in a marked upshift in the rate of DNA replicationinitiation. Taken together with the results using theSojD40A,R189A double mutant shown above, the data sug-gests that SojR189A is a monomer when expressed from itsnative locus, but that overexpression drives the proteininto a dimer.
Regulation of Soj through the activation of its ATPaseactivity by Spo0J
Since Soj is a monomer either in the presence of ADP orwithout nucleotide, one way for Soj to switch from a dimerto a monomer could be for it to hydrolyse its bound ATPmolecules. To begin examining this possibility the intrinsicATPase activity of wild-type Soj was determined using amalachite green assay. Figure 4 shows that Soj has a lowlevel of intrinsic ATPase activity that is just above thebackground level.
It has been found that T. thermophilus Spo0J stimulatesthe ATPase activity of T. thermophilus Soj (Leonard et al.,
Oligomeric status switches Soj regulatory activity 1093
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

2005). To determine whether B. subtilis Spo0J could act ina similar manner to regulate Soj, the Spo0J protein waspurified and the ATPase activity of Soj was determined inthe presence of Spo0J. In sharp contrast to the result withSoj alone, Spo0J stimulated the ATPase activity of Sojmore than 50-fold (Fig. 4), much greater than the degreeof stimulation observed for T. thermophilus Soj. TheATPase activities of SojG12V and SojK16A were not stimu-lated by Spo0J, suggesting that Soj must form a dimerto become competent for Spo0J-mediated ATPasestimulation.
We observed that the dramatic stimulation of SojATPase activity by Spo0J was dependent upon DNA in
the reactions (Fig. 4). To further investigate the role thatDNA plays in the Soj:Spo0J interaction we used a surfaceplasmon resonance (SPR) assay. Double-stranded DNAencoding a single parS site was immobilized onto thesurface of a SPR sensor chip and Spo0J was injected ata concentration that rapidly reached equilibrium (Fig. S5).Subsequently, the Spo0J injection was terminated and Sojwas immediately injected over the Spo0J:DNA complexesin the presence and absence of various nucleotides.Figure 5A shows that wild-type Soj only interacted withthe Spo0J:DNA complexes in the presence of ATP, indi-cating that Soj needs to dimerize in order to form acomplex with Spo0J. We then repeated this experimentusing the SojR189A DNA binding mutant. Figure 5B showsthat SojR189A did not significantly interact with Spo0J, eventhough this mutant is able to dimerize in the presence ofATP. This result is consistent with the observation thatSpo0J-dependent foci formation by GFP–SojD40A isdependent upon the DNA binding activity of Soj (Fig. S3B;Murray and Errington, 2008). Taken together, the ATPaseand SPR experiments indicate that Soj must form a dimerand bind to DNA in order to be properly orientated so thatit can efficiently interact with Spo0J:DNA complexes tostimulate its ATPase activity.
Lysines 3 and 7 of Spo0J are required to controlSoj activity
Thermus thermophilus Spo0J, as well as many ortholo-gous plasmid proteins, contain arginine residues withinthe N-terminal tail of the protein that have been shown tobe required for activating their cognate ATPases (Leonardet al., 2005; Barilla et al., 2007; Ah-Seng et al., 2009). B.subtilis Soj does not contain an arginine residue withinthis region; however, it does contain two lysine residues atamino acid positions 3 and 7.
To determine whether these lysines are involved in thestimulation of Soj ATPase activity in vitro, a double alaninesubstitution was constructed and purified. Addition ofSpo0JK3A,K7A to Soj resulted in an approximately ninefolddecrease in ATPase activity compared with wild-typeSpo0J, indicating that these lysine residues play animportant role in regulating Soj (Fig. 4). Interestingly, wefound that these lysine residues could be substituted witharginine residues with almost no loss of ATPase stimula-tion (Fig. 4). This result differs from previous reports forplasmid ParA homologues, in which the analogous argin-ine residues could not be substituted with lysines (Barillaet al., 2007; Ah-Seng et al., 2009).
To further understand the role of the Spo0J N-terminallysine residues, we investigated the ability of Soj to inter-act with the Spo0JK3A,K7A and Spo0JK3R,K7R mutants usingSPR. Consistent with results from the ATPase assays, theSpo0JK3A,K7A mutant was unable to form a complex with
Fig. 4. Efficient stimulation of Soj ATPase activity requires Spo0Jand DNA. Reaction components are described in the lower tableand are associated with symbols shown in the time-courses above.The DNA substrate was a 55-base-pair fragment from the spo0Jgene containing a single parS site. Error bars represent thestandard error of the mean (n = 3). Rate of phosphate releasedwas calculated using a potassium phosphate standard along with alinear regression analysis (coloured lines) for each data set. Acontrol reaction containing ATP alone released 0.8 moles of Pi perhour.
1094 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

Soj (Fig. 5C). In contrast the Spo0JK3R,K7R mutant inter-acted with Soj similarly to wild-type Spo0J (Fig. 5D). Inspite of these differences, both Spo0J mutants bound toDNA in a similar manner to wild-type Spo0J.
To determine whether these lysine residues in Spo0Jare required to regulate Soj in vivo, we replaced theendogenous spo0J gene with both single and doublealanine substitutions and assessed the activity of Sojusing marker frequency analysis and the localization ofGFP–Soj. Marker frequency analysis revealed that eachof the single mutants overinitiated DNA replication, withthe K3A mutation having the strongest effect (Fig. 6A).The degree of over-replication observed in theSpo0JK3A,K7A double mutant appeared to be the additiveeffect of the two single substitutions and was indistin-guishable from that of the Dspo0J null mutant (Fig. 6A),indicating that these lysine residues are critical for theregulation of Soj in vivo. We found that the lysine muta-tions did not affect the ability of Spo0J–GFP to localize as
foci, indicating that these mutations do not interfere withSpo0J dimerization or DNA binding (Fig. S6).
Next the localization of GFP–Soj was examined in thecontext of the Spo0J lysine mutants (Fig. 6B). The trendobserved followed the pattern of the marker frequencyanalysis. While GFP–Soj normally colocalizes with the cellsepta and forms faint foci at oriC, the Spo0JK3A,K7A doublemutant caused GFP–Soj to accumulate into large patchesthat colocalized with the nucleoid, similar to the patternobserved in a Dspo0J null mutant. The single lysinemutants caused GFP–Soj to accumulate as smallerpatches most often located proximal to a cell pole. Wesuspected that these smaller patches represented GFP–Soj binding to DNA. To test this idea we localized the DNAbinding mutant GFP–SojR189A in the context of Spo0JK3A.Figure 6C shows that the fluorescent patches weredependent upon the DNA binding residues of GFP–Soj,indicating that they represent GFP–Soj binding to thenucleoid. Western blot analysis confirmed that all of the
Fig. 5. Interaction of Soj with Spo0J:DNA complexes. Spo0J proteins (500 nM) were injected over DNA substrates for 200 s, followed by aninjection of Soj proteins (1500 nM) with or without nucleotide for 300 s during the Spo0J dissociation phase.A. Injection of wild-type Spo0J followed by wild-type Soj.B. Injection of wild-type Spo0J followed by SojR189A.C. Injection of Spo0JK3A,K7A followed by wild-type Soj.D. Injection of Spo0JK3R,K7R followed by wild-type Soj.
Oligomeric status switches Soj regulatory activity 1095
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

GFP–Soj proteins were expressed to the same level(Fig. S4B), confirming that altered protein localization wasnot a consequence of changes in protein concentration.
Activation of Soj ATPase activity inhibits Sojdimerization
The data presented above suggest a model in whichSpo0J controls Soj localization and activity by altering theoligomeric state of Soj via stimulation of its ATPaseactivity. To test this model we utilized the two-hybrid assayto investigate Soj self-interaction in the presence ofSpo0J. Figure 6D shows that wild-type Spo0J efficientlyinhibits the Soj self-interaction (white colonies), whereasthe Spo0JK3A,K7A mutant does not (blue colonies). Takentogether, the results presented in this work are consistentwith a model in which Spo0J regulates the localization
and regulatory activity of Soj by driving Soj from an ATP-bound dimer to an ADP-bound monomer.
Discussion
Molecular basis for the dichotomous regulatory activitiesof Soj
The B. subtilis ATPase Soj was known to act as either anactivator or an inhibitor of the DNA replication initiatorprotein DnaA under different conditions; however, themolecular basis underlying this switch in regulatory activi-ties had not been established. Here we report that DNAreplication initiation is inhibited by monomeric Soj andactivated by dimeric Soj, thereby indicating that the keyevent which switches Soj regulatory activity in vivo is themonomer-to-dimer transition. We go on to show that spe-
Fig. 6. N-terminal lysine residues of Spo0Jregulate Soj localization and activity.A. The oriC-to-terminus ratio of Spo0Jmutants was determined using markerfrequency analysis. Cells were grown in CHmedium at 30°C. The average � standarddeviation from three technical replicates isshown. The dashed line represents theaverage ratio for the parent strain. Wild type(HM34), Dspo0J (HM42), spo0JK7A (HM329),spo0JK3A (HM337), spo0JK3A,K7A (HM338).B. The localization of GFP–Soj in thepresence of Spo0J N-terminal lysine mutantswas observed using epifluorescencemicroscopy. Cells were grown in CH mediumat 30°C. Patches of GFP–Soj are indicated byyellow asterisks. gfp-soj spo0J+ (HM4), gfp-sojDspo0J (HM13), gfp-soj spo0JK7A (HM391),gfp-soj spo0JK3A (HM392), gfp-soj spo0JK3A,K7A
(HM393). Note that because the end of sojoverlaps the start of spo0J, the K3A mutationeliminates the natural Soj stop codon andresults in the addition of the followingresidues: CRPWSRD.C. The localization of GFP–SojR189A in thepresence of Spo0JK3A was observed usingepifluorescence microscopy. Cells were grownin CH medium at 30°C. Patches of GFP–Sojare indicated by yellow asterisks and polarlocalization is indicated with arrows. gfp-sojspo0JK3A (HM392), gfp-sojR189A spo0JK3A
(HM695).D. Bacterial two-hybrid analysis of Soj in thepresence of Spo0J. Strains were streakedonto nutrient agar plates supplemented withantibiotics and X-gal. All strains contained thelow-copy-number vector pHM371 withcya�-soj, which was paired with the followingplasmids: empty vector (pUT18C), soj(pHM376), soj spo0J (pHM393), sojspo0JK3A,K7A (pHM394).
1096 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

cific amino acids of the regulatory protein Spo0J requiredfor stimulating the ATPase activity of Soj are required forregulating Soj dimerization, thus accounting for themechanism by which Spo0J regulates Soj localization andactivity.
The results suggest that Spo0J plays a key role inflipping the Soj switch to control DNA replication initiation.We hypothesize that during the cell cycle or in response toan undefined cellular cue the regulation of Soj by Spo0J isovercome. One model is that the concentration of Sojrelative to Spo0J increases significantly to shift the equi-librium of Soj protein from a monomer to a dimer. Thiscould occur through increased protein synthesis orthrough an increase in the local concentration of Soj rela-tive to Spo0J (i.e. near the cell pole). Alternatively, it wasrecently shown that Spo0J recruits condensin to the originregion of the chromosome to facilitate DNA segregation(Gruber and Errington, 2009; Sullivan et al., 2009). Wewonder whether the binding of Condensin occludesSpo0J from interacting with Soj, thereby promoting Soj toactivate DNA replication initiation as the Condensin chro-mosome segregation machinery is being recruited. Afurther possibility is that the Spo0J nucleoprotein com-plexes may be displaced during DNA replication by theaction of DNA polymerases. The dispersed Spo0J maynot be able to effectively interact with and/or regulate Soj,thus allowing a window of time before Spo0J reassemblesfor Soj to accumulate as a dimer and activate DnaA. Thiscould help to synchronize the initiation of multiple replica-tion origins since DnaA is sequestered from newly acti-vated origins by YabA-dependent tethering to thereplication machinery (Soufo et al., 2008), thereby tran-siently leaving DnaA bound only to origins that have yet tofire.
Soj DNA binding activity is required to activate DNAreplication initiation and to interact with Spo0J
We have found that the DNA binding activity of Soj isrequired for positive regulation of DNA replication initia-tion, but not for negative regulation. Furthermore, SojDNA binding activity is required for both the Spo0J inter-action and the Spo0J-mediated ATPase stimulation. Weimagine that Soj must be properly orientated by DNA sothat it sits adjacent to Spo0J in order to efficiently interact.As noted previously (Hester and Lutkenhaus, 2007), thelocation of the Soj DNA binding surface on the structure T.thermophilus Soj (Leonard et al., 2004) would allowSpo0J to contact Soj dimers that are located eitherupstream or downstream.
Interestingly, we have found that the SojR189A proteindisplays the localization pattern and regulatory activity ofa monomeric Soj protein, even though this mutant iscapable of dimerization either as a purified protein in vitro
or when overexpressed in vivo. We propose that Sojdimerization is carefully regulated in B. subtilis through, atthe very least, protein expression and protein localizationto ensure proper control of Soj activity. We also suspectthat additional factors are present in vivo that regulate themonomer-to-dimer transition which were not present inour in vitro and two-hybrid experiments (see below).
Regulation of the monomeric Soj/ParA proteins
Soj is a member of a large class of proteins referred to asParA ATPases. These proteins were originally identifiedas stability determinants on low-copy-number plasmidsand have now been identified in the majority of sequencedbacterial genomes (Gerdes et al., 2000; Livny et al.,2007). Although all of the ParA orthologues share homol-ogy, there are also significant differences between them,both at the sequence level and potentially at the functionallevel. Based on structural and biochemical studies ofplasmid-encoded ParA proteins from P1, P7 andpSM19035, one interesting difference appears to be thatthese plasmid ParA proteins do not undergo a monomerto dimer switch, but rather they form stable dimers inde-pendent of the identity of their bound nucleotide (Prattoet al., 2008; Dunham et al., 2009). We note that becauseSoj/ParA proteins from the distantly related organisms B.subtilis (Gram-positive) and T. thermophilus (Gram-negative) appear to share the ATP-dependent dimeriza-tion switch, this mechanism for Soj/ParA regulation waslikely derived from a deeply rooted common ancestor andwe suspect that the majority of chromosomally encodedSoj/ParA proteins will retain this activity.
We speculate that the Soj monomer-to-dimer transitionmay provide the opportunity for regulatory systems toinfluence Soj activity. One candidate for such a mecha-nism would be the Soj polar localization determinant(s)which appears to only interact with monomeric Soj(Murray and Errington, 2008). In the current study weobserved that the patches of GFP–Soj observed in theSpo0J N-terminal lysine mutants were most often locatedadjacent to septa (Fig. 6B) and we have previously shownthat in filamentous cells GFP–Soj preferentially accumu-lates near cell poles (Autret and Errington, 2003). Theseresults suggest that factors localized at the cell pole couldeither spatially or temporarily affect Soj regulation bysequestering monomeric Soj. Although MinD is requiredfor Soj to localize to the cell pole (Murray and Errington,2008), we have not been able to detect a direct interactionbetween Soj and MinD using the bacterial two-hybridassay, nor have we observed a change in the regulation ofDNA replication by Soj in a DminD mutant (data notshown). It will be highly informative to identify the directpolar localization determinant of Soj and to evaluate itsrole regulating Soj dimerization.
Oligomeric status switches Soj regulatory activity 1097
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

Experimental procedures
Strains and plasmids
Construction of strains and plasmids used in this study aredescribed in Supporting information and listed in Tables S1and S2. E. coli strain DH5a (Invitrogen) was used for theconstruction of all plasmids, and strain BL21 (DE3) pLysS(Stratagene) was used to express all proteins. The plasmidpET21-d (Invitrogen) was used as the expression vector forall proteins, and either pUC18 or pBSoriC4 (Krause et al.,1997) was used for biochemical assays.
Media
Nutrient agar (Oxoid) was used for routine selection andmaintenance of both B. subtilis and E. coli strains. For experi-ments in B. subtilis cells were grown in casein hydrolysate(CH) medium, except for the experiment to calculate thefrequency of cells lacking DNAwhere defined minimal mediumwas used [Spizizen minimal medium supplemented with glyc-erol (0.5%), MgSO4 (6 mM), CaCl2 (0.1 mM), MnSO4 (130 mM)and Fe-NH4-citrate (0.01 mg ml-1)]. Supplements were addedas required: 20 mg ml-1 tryptophan, 5 mg ml-1 chlorampheni-col, 2 mg ml-1 kanamycin, 50 mg ml-1 spectinomycin. Forprotein expression in E. coli cells were grown in Luria–Bertani(LB) medium or Nutrient Broth (Oxoid) and supplemented with30 mg ml-1 (for single copy plasmids) or 75 mg ml-1 ampicillin,10 mg ml-1 chloramphenicol, 50 mg ml-1 kanamycin. For thebacterial two-hybrid analysis cells were grown in M9 saltssupplemented with glucose (0.8%), casamino acids (0.4%),MgSO4 (1 mM), CaCl2 (0.1 mM) and thiamine (2 mM).
Microscopy
To visualize cells during exponential growth starter cultureswere grown in CH medium overnight, then diluted 1:100 intofresh medium (with inducer where indicated) and allowed toachieve at least three doublings before observation. Cellswere mounted on ~1.2% agar pads (containing 0.5¥ CH)immobilized within a Gene Frame (ABgene) using a0.13–0.17 mm glass coverslip (VWR). To visualize nucleoidsthe DNA was stained with 2 mg ml-1 4′-6-diamidino-2-phenylindole (DAPI) (Sigma). To visualize individual cells thecell membrane was stained with either 2 mg ml-1 Nile Red(Sigma) or 0.4 mg ml-1 FM5-95 (Molecular Probes). Micros-copy was performed on an inverted epifluorescence micro-scope (Zeiss Axiovert 200M) fitted with a Plan-Neofluarobjective (Zeiss 100¥/1.30 Oil Ph 3). Light was transmittedfrom a 300 Watt xenon arc-lamp through a liquid light guide(Sutter Instruments) and images were collected using a SonyCoolSnap HQ cooled CCD camera (Roper Scientific).All filterswere Modified Magnetron ET Sets from Chroma and detailsare available upon request. Digital images were acquired andanalysed using METAMORPH software (version V.6.2r6).
Marker frequency analysis
Cells were grown in CH medium at 30°C as described formicroscopy. Sodium azide (0.5%; Sigma) was added to expo-nentially growing cells (A600 = 0.2–0.4) to prevent furthergrowth. Chromosomal DNA was isolated using a DNeasy
Blood and Tissue Kit (Qiagen). Power SYBR GreenPCR Master Mix was used for PCR reactions (AppliedBiosystems). Q-PCR was performed in a LightCycler 480Instrument (Roche). By use of crossing points and PCR effi-ciency a relative quantification analysis was performed usingLight-Cycler Software version 4.0 (Roche) to determine theori/ter ratio of each sample. These results were normalized tothe ori/ter ratio of a DNA sample from B. subtilis spores inwhich the ori/ter ratio is 1.
Bacterial two-hybrid assay
Escherichia coli strain BTH101 was transformed using acombination of complimentary plasmids. For quantitativeb-galactosidase assays strains were incubated overnight at30°C in M9 medium containing ampicillin and kanamycin,diluted 1:50 into fresh medium with antibiotics and grown untilthey reached an A600 of 0.4–0.6. Samples were placed on icefor 20 min to stop growth, after which the culture density wasdetermined. Each culture was mixed with Z-buffer, then Bug-Buster (Novagen) was added to permeabilize the cells for20 min at 30°C. b-Galactosidase activity was measured aspreviously described (Miller, 1972; Gourse et al., 1986). Forqualitative b-galactosidase assays single colonies werestreaked onto nutrient agar plates containing ampicillin, kana-mycin and the indicator X-gal (0.008%). Plates were incu-bated at 30°C overnight and imaged using a digital camera.
Protein purification
Purification of Soj-His6 and Spo0J-His6 was performed asdescribed below. The molecular weight, theoretical pI andextinction coefficient for Soj-His6 and Spo0J-His6 were calcu-lated using ProtParam, an online tool present on the ExPASyproteomics sever. All affinity chromatography was performedat 4°C at a flow rate of 0.5 ml min-1 (unless otherwise stated)using an AKTA Purifier FPLC system (GE Healthcare).
Purification of Soj-His6
A BL21 (DES) pLysS strain containing the appropriate soj-his6 plasmid (see Table S2) was inoculated into LB containingampicillin and chloramphenicol and grown overnight at 37°C.The culture was diluted 1:100 into LB containing ampicillinand incubated at 37°C until the A600 reached ~0.6. Cultureswere then supplemented with 1 mM IPTG and shifted to 30°Cfor 3 h to induce protein expression. Cells were collected bycentrifugation at 5000 g (4°C) for 10 min and then resus-pended in Soj purification buffer (30 mM Tris pH 8, 300 mMNaCl, 20 mM MgCl2, 40 mM imidazole and 20% glycerol)such that they were concentrated 25-fold. One completeEDTA-free protease inhibitor tablet (Roche) and lysozyme(32 mg ml-1) were added and cells were incubated on ice for1 h with gentle agitation. Cells were lysed using a Vibra-Cellsonicator (Sonics) on ice for approximately 5 min (powersetting 80; 4 s pulse, 1 s rest). Cell debris was removed bycentrifugation at 31 000 g (4°C) for 45 min. The supernatantwas applied to a 1 ml HisTrap FF column (GE Healthcare),washed with 20 ml of Soj purification buffer and then washedwith 20 ml of Soj buffer supplemented with NaCl (200 mM).Bound protein was eluted with 5 ml of Soj purification buffer
1098 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

supplemented with imidazole (500 mM). The eluate wasapplied to a HiLoad Superdex 75 gel filtration column (GEHealthcare) pre-equilibrated with Soj gel filtration buffer(30 mM Tris pH 7.6, 200 mM potassium glutamate, 1 mMDTT, 1 mM EDTA and 20% glycerol) and run overnight at0.3 ml min-1. Fractions were then analysed by SDS-PAGEand relevant fractions pooled. The protein solution was con-centrated using an Amicon Ultra-15 centrifugal filter spincolumn (Millipore). All Soj mutants were purified in an identi-cal manner.
Purification of Spo0J-His6
A BL21 (DES) pLysS strain containing the appropriate spo0J-his6 plasmid (see Table S2) was inoculated, grown andinduced as described above. Cells were collected by centrifu-gation at 5000 g (4°C) for 10 min and then resuspended inSpo0J purification buffer (50 mM HEPES pH 7.6, 300 mMNaCl, 10 mM imidazole and 20% glycerol). Cells were lysedas described above. The supernatant was applied to a 1 mlHisTrap FF column (GE Healthcare), washed with 20 ml ofSpo0J wash buffer (50 mM HEPES pH 7.6, 300 mM NaCland 40 mM imidazole) and then eluted with a linear gradientof Spo0J wash buffer supplemented with imidazole(500 mM). Fractions were analysed by SDS-PAGE and rel-evant fractions were pooled, diluted to 50 ml with Spo0Jheparin buffer (20 mM HEPES pH 7.6 and 1 mM EDTA) andapplied to a 1 ml HiTrap Heparin column (GE Healthcare).Proteins were eluted using a linear gradient of Spo0J heparinbuffer supplemented with NaCl (1.5 M). Fractions wereanalysed by SDS-PAGE and relevant fractions were pooledand applied to a HiLoad Superdex 75 gel filtration columnpre-equilibrated with Spo0J gel filtration buffer (30 mMHEPES pH 7.6, 300 mM NaCl, 1 mM DTT and 10% glycerol)and run overnight at 0.3 ml min-1. Fractions were then analy-sed by SDS-PAGE and relevant fractions pooled. The proteinsolution was then concentrated using an Amicon Ultra-15centrifugal filter spin column.
Size exclusion chromatography
A Superdex 75 HR 10/30 (GE Healthcare) column was pre-equilibrated with Soj gel filtration buffer (50 mM Tris pH 7.6,150 mM potassium glutamate, 5 mM MgCl2 and 1 mM DTT)supplemented with either 2 mMATP orADP. The target proteinwas pre-incubated with or without nucleotide for 20 min at37°C. One hundred microlitres of the protein solution(2 mg ml-1) was injected onto the column at a flow rate of0.4 ml min-1. The A280 was monitored for the entire columnvolume (24 ml).
Circular dichroism spectroscopy
Fav-UV circular dichroism spectra (190–250 nm) were col-lected using a JASCO 810 (JASCO, UK) in a 0.2 mm path-length cuvette. Prior to analysis Soj-His6 was bufferexchanged into 200 mM Sodium Phosphate (pH 7.0) using aPD-10 column (GE Healthcare) and concentrated using anAmicon Ultra-15 centrifugal filter spin column to 0.5 mg ml-1.A buffer only spectra was collected as a reference and sub-tracted from the sample spectra.
UV cross-linking
[a-32P]-ATP was cross-linked to Soj following exposure to UVlight. Soj (5 mM) was added to cross-linking buffer [50 ml;25 mM HEPES pH 7.6, 50 mM potassium glutamate, 1 mMDTT, 10 mM MgCl2, 0.2 mg ml-1 bovine IgG and 0.5 mM[a-32P]-ATP (400 Ci mmol-1)] in PCR tubes chilled in a metalrack on ice. The reactions were irradiated at 254 nm for45 min in a Spectrolinker (XL-1500 UV Crosslinker, Spectron-ics Corp.). The samples were analysed by SDS-PAGE (Invit-rogen) and protein bands visualized by staining withCoomassie Brilliant Blue. The gel was exposed to a phosphorstorage screen overnight and the 32P-labelled bands visual-ized using a Typhoon imager (GE Healthcare).
ATPase assay
The malachite green assay solution was created by mixing0.0812% (w/v) malachite green, 2.23% (w/v) polyvinyl alcohol,5.72% (w/v) ammonium molybdate in HCl (6 M) and H2O in aratio of 2:1:1:2. The solution was incubated for 2 h with gentleagitation (during this time the solution turned from a muddybrown colour to a golden yellow). Various combinations of Soj,Spo0J and DNA (all at a final concentration of 2 mM) weremixed and diluted into ATPase buffer (50 mM HEPES pH 7.6,100 mM potassium glutamate and 10 mM MgCl2) in a finalvolume of 400 ml. The DNA substrate is a fragment of thespo0J gene containing a parS site and was constructed byannealing complimentary oligonucleotides (5′-CTGAATCAGCAGTTGAATCAGAATGTTCCACGTGAAACAAAGAAAAAAGAACCTG-3′). The protein mixtures were incubated for 5 minat 37°C, followed by addition of ATP (to a final concentration of2 mM).At various time points 50 ml of the reaction solution wasremoved and immediately mixed with 800 ml of malachitegreen assay solution, followed by addition of 100 ml of sodiumcitrate (34%). The colour was allowed to stabilize for 10 minbefore the absorbance was detected at 620 nm. A standardcurve was created using a serial dilution of sodium phosphate(3–500 mM). The A620 was converted into mole Pi produced permole of protein using a phosphate standard.
Surface plasmon resonance
Surface plasmon resonance was performed using a NeutrA-vidin coated (NLC) sensor chip in a Proteon XPR36 system.A 148 bp PCR product carrying a 5′ Biotin-tag and harbouringa parS site in the middle was amplified from the 3′ region ofthe spo0J gene. A PCR product carrying a mutated parS sitewas also created and used as a negative control (Lin andGrossman, 1998). Approximately 500 RU of specific and non-specific DNA substrates were immobilized in separate ligandchannels on the NLC chip in running buffer at 25°C (25 mMHEPES pH 7.6, 200 mM NaCl, 10 mM MgCl2, 1 mM DTT and0.005% Tween-20). Spo0J was exchanged (PD-10 column,GE Healthcare) into running buffer and injected over bothchannels at a flow rate of 50 ml min-1. Soj was exchanged intorunning buffer and pre-incubated with 2 mM of the relevantnucleotide for 20 min at 37°C. To investigate the Spo0J–Sojinteraction, Soj was injected during the Spo0J dissociationphase at a flow rate of 50 ml min-1. The reported data repre-
Oligomeric status switches Soj regulatory activity 1099
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100

sent the response observed using the DNA substrate carryingthe specific parS site following subtraction of the responseobserved using the DNA substrate carrying the mutated parSsite. Affinity analysis was performed using the ProteonManager software.
Acknowledgements
We thank the Lakey Lab (Newcastle University) for advice onSPR and circular dichroism analysis, the Lutkenhaus Lab(Kansas University) for providing the sojR189A allele, DavidAdams for providing purified FtsZ and Ian Selmes for techni-cal assistance. This work was supported by fellowships fromthe European Molecular Biology Organization, the HumanFrontier Science Program and the Royal Society to H.M., andby Grant No. 43/G18654 from the BBSRC to J.E.
References
Ah-Seng, Y., Lopez, F., Pasta, F., Lane, D., and Bouet, J.Y.(2009) Dual role of DNA in regulating ATP hydrolysis by theSopA partition protein. J Biol Chem 284: 30067–30075.
Autret, S., and Errington, J. (2003) A role for division-site-selection protein MinD in regulation of internucleoidjumping of Soj (ParA) protein in Bacillus subtilis. J BiolChem 47: 159–169.
Barilla, D., Carmelo, E., and Hayes, F. (2007) The tail of theParG DNA segregation protein remodels ParF polymersand enhances ATP hydrolysis via an arginine finger-likemotif. Proc Natl Acad Sci USA 104: 1811–1816.
Breier, A.M., and Grossman, A.D. (2007) Whole-genomeanalysis of the chromosome partitioning and sporulationprotein Spo0J (ParB) reveals spreading and origin-distalsites on the Bacillus subtilis chromosome. J Biol Chem 64:703–718.
Dunham, T.D., Xu, W., Funnell, B.E., and Schumacher, M.A.(2009) Structural basis for ADP-mediated transcriptionalregulation by P1 and P7 ParA. EMBO J 28: 1792–1802.
Gerdes, K., Møller-Jensen, J., and Jensen, R.B. (2000)Plasmid and chromosome partitioning: surprises fromphylogeny. J Biol Chem 37: 455–466.
Gourse, R.L., de Boer, H.A., and Nomura, M. (1986) DNAdeterminants of rRNA synthesis in E. coli: growth ratedependent regulation, feedback inhibition, upstream acti-vation, antitermination. Cell 44: 197–205.
Gruber, S., and Errington, J. (2009) Recruitment of condensinto replication origin regions by ParB/SpoOJ promotes chro-mosome segregation in B. subtilis. Cell 137: 685–696.
Hester, C.M., and Lutkenhaus, J. (2007) Soj (ParA) DNAbinding is mediated by conserved arginines and is essen-tial for plasmid segregation. Proc Natl Acad Sci USA 104:20326–20331.
Karimova, G., Pidoux, J., Ullmann, A., and Ladant, D. (1998)A bacterial two-hybrid system based on a reconstitutedsignal transduction pathway. Proc Natl Acad Sci USA 95:5752–5756.
Katayama, T., Ozaki, S., Keyamura, K., and Fujimitsu, K.(2010) Regulation of the replication cycle: conserved anddiverse regulatory systems for DnaA and oriC. Nat RevMicrobiol 8: 163–170.
Krause, M., Ruckert, B., Lurz, R., and Messer, W. (1997)Complexes at the replication origin of Bacillus subtilis withhomologous and heterologous DnaA protein. J Mol Biol274: 365–380.
Lee, P.S., and Grossman, A.D. (2006) The chromosome par-titioning proteins Soj (ParA) and Spo0J (ParB) contribute toaccurate chromosome partitioning, separation of replicatedsister origins, and regulation of replication initiation in Bacil-lus subtilis. Mol Microbiol 60: 853–869.
Leonard, T.A., Butler, P.J., and Lowe, J. (2004) Structuralanalysis of the chromosome segregation protein Spo0Jfrom Thermus thermophilus. J Biol Chem 53: 419–432.
Leonard, T.A., Butler, P.J., and Lowe, J. (2005) Bacterialchromosome segregation: structure and DNA binding ofthe Soj dimer – a conserved biological switch. EMBO J 24:270–282.
Lin, D.C.-H., and Grossman, A.D. (1998) Identification andcharacterization of a bacterial chromosome partitioningsite. Cell 92: 675–685.
Livny, J., Yamaichi, Y., and Waldor, M.K. (2007) Distributionof centromere-like parS sites in bacteria: insights fromcomparative genomics. J Bacteriol 189: 8693–8703.
Miller, J.H. (1972) Experiments in Molecular Genetics. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press.
Murray, H., and Errington, J. (2008) Dynamic control of theDNA replication initiation protein DnaA by Soj/ParA. Cell135: 74–84.
Murray, H., Ferreira, H., and Errington, J. (2006) The bacte-rial chromosome segregation protein Spo0J spreads alongDNA from parS nucleation sites. J Biol Chem 61: 1352–1361.
Ogura, Y., Ogasawara, N., Harry, E.J., and Moriya, S. (2003)Increasing the ratio of Soj to Spo0J promotes replicationinitiation in Bacillus subtilis. J Bacteriol 185: 6316–6324.
Pratto, F., Cicek, A., Weihofen, W.A., Lurz, R., Saenger, W.,and Alonso, J.C. (2008) Streptococcus pyogenespSM19035 requires dynamic assembly of ATP-bound ParAand ParB on parS DNA during plasmid segregation.Nucleic Acids Res 36: 3676–3689.
Ptacin, J.L., and Shapiro, L. (2010) Initiating bacterial mitosis:understanding the mechanism of ParA-mediated chromo-some segregation. Cell Cycle 9: 4033–4034.
Soufo, C.D., Soufo, H.J., Noirot-Gros, M.F., Steindorf, A.,Noirot, P., and Graumann, P.L. (2008) Cell-cycle-dependent spatial sequestration of the DnaA replicationinitiator protein in Bacillus subtilis. Dev Cell 15: 935–941.
Sullivan, N.L., Marquis, K.A., and Rudner, D.Z. (2009)Recruitment of SMC by ParB-parS organizes the originregion and promotes efficient chromosome segregation.Cell 137: 697–707.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
1100 G. Scholefield, R. Whiting, J. Errington and H. Murray �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1089–1100
Copyright © 2022 FDOKUMEN




















