
Multiple interaction sites of galnon trigger its biological effects
12
Multiple interaction sites of galnon trigger its biological effects Anders Flore ´n a , Ulla Sollenberg a , Linda Lundstro ¨m a , Matjaz ˇ Zorko b , Jure Stojan b , Metka Budihna c , Mark Wheatley d , Negin P. Martin e , Kalle Kilk f , Andrey Mazarati g , Tamas Bartfai h , Maria Lindgren a ,U ¨ lo Langel a, * a Department of Neurochemistry, Stockholm University, S. Arrheniusv 21 A, SE-106 91 Stockholm, Sweden b Institute of Biochemistry, Medical Faculty, University of Ljubljana, Vrazov trg 2, 1000 Ljubljana, Slovenia c Institute of Pharmacology and Experimental Toxicology, Medical Faculty, University of Ljubljana, Korytkova 2, 1000 Ljubljana, Slovenia d School of Biosciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK e The Howard Hughes Medical Institute, Duke University Medical Center, Box 3821, Durham, NC, 27710, USA f Centre of Molecular and Clinical Medicine, Tartu University, Ravila 19, 51014 Tartu, Estonia g Department of Neurology, University of California, Los Angeles, West Los Angeles Veteran Administration Medical Center, 90073, Los Angeles, CA, USA h Department of Neuropharmacology, The Scripps Research Institute, La Jolla, CA 92037, USA Received 23 May 2005; accepted 24 September 2005 Abstract Galnon was first reported as a low molecular weight non-peptide agonist at galanin receptors [Saar et al. (2002) Proc. Natl. Acad. Sci. USA 99, 7136–7141]. Following its systemic administration, this synthetic ligand affected a range of important physiological processes including appetite, seizures and pain. Physiological activity of galnon could not be explained solely by the activation of the three known galanin receptors, GalR1, GalR2 and GalR3. Consequently, it was possible that galnon generates its manifold effects by interacting with other signaling pathway components, in addition to via GalR1-3. In this report, we establish that galnon: (i) can penetrate across the plasma membrane of cells, (ii) can activate intracellular G-proteins directly independent of receptor acti- vation thereby triggering downstream signaling, (iii) demonstrates selectivity for different G-proteins, and (iiii) is a ligand to other G-protein coupled receptors (GPCRs) in addition to via GalR1-3. We conclude that galnon has multiple sites of interaction within the GPCR signaling cascade which mediate its physiological effects. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Galanin; Galnon; G-protein; Receptor; Signaling 0143-4179/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.npep.2005.09.005 Abbreviations: AR, adrenoreceptor; AVP, [Arg 8 ]vasopressin; CHO, Chinese hamster ovary; D1(2)R, dopamine receptors, type 1 (2); DMSO, dimethylsulfoxide; GalR, galanin receptor; GPCR, G-protein coupled receptors; HKR, Hepes–Krebs–Ringer solution; HPLC, high performance liquid chromatography; 5-HT, 5-hydroxytryptamine; IC 50 , concentration giving half-maximal inhibition; LC–MSD-Trap-XCT, liquid chromato- graph–mass spectrometer-ion trap instrument; LDH, lactate dehydrogenase; M1, (2,5) mAChR muscarinic acetylcholine receptor, type 1 (2, 5); mAChR, muscarinic acetylcholine receptor, not specified; MALDI-TOF MS, matrix assisted laser desorption/ionization-time-of-flight mass spec- trometer; MC3 (4), melanocortin receptor, type 3 (4); OTR, oxytocin receptor; PBS, phosphate buffered saline; PTX, pertussis toxin; PPS, perforant path stimulation; RP, reverse phase; SSSE, self-sustaining status epilepticus; TFA, trifluoroacetic acid; V 1a R, V 1a vasopressin receptor; V 1b R, V 1b vasopressin receptor; V 2 R, V 2 vasopressin receptor * Corresponding author. Tel.: +46 816 1793; fax: +46 816 1371. E-mail address: [email protected] (U ¨ . Langel). www.elsevier.com/locate/npep Neuropeptides xxx (2005) xxx–xxx Neuropeptides ARTICLE IN PRESS
Transcript of Multiple interaction sites of galnon trigger its biological effects

ARTICLE IN PRESS
www.elsevier.com/locate/npep
Neuropeptides xxx (2005) xxx–xxx
Neuropeptides
Multiple interaction sites of galnon trigger its biological effects
Anders Floren a, Ulla Sollenberg a, Linda Lundstrom a, Matjaz Zorko b, Jure Stojan b,Metka Budihna c, Mark Wheatley d, Negin P. Martin e, Kalle Kilk f, Andrey Mazarati g,
Tamas Bartfai h, Maria Lindgren a, Ulo Langel a,*
a Department of Neurochemistry, Stockholm University, S. Arrheniusv 21 A, SE-106 91 Stockholm, Swedenb Institute of Biochemistry, Medical Faculty, University of Ljubljana, Vrazov trg 2, 1000 Ljubljana, Slovenia
c Institute of Pharmacology and Experimental Toxicology, Medical Faculty, University of Ljubljana, Korytkova 2, 1000 Ljubljana, Sloveniad School of Biosciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK
e The Howard Hughes Medical Institute, Duke University Medical Center, Box 3821, Durham, NC, 27710, USAf Centre of Molecular and Clinical Medicine, Tartu University, Ravila 19, 51014 Tartu, Estonia
g Department of Neurology, University of California, Los Angeles, West Los Angeles Veteran Administration Medical Center,
90073, Los Angeles, CA, USAh Department of Neuropharmacology, The Scripps Research Institute, La Jolla, CA 92037, USA
Received 23 May 2005; accepted 24 September 2005
Abstract
Galnon was first reported as a low molecular weight non-peptide agonist at galanin receptors [Saar et al. (2002) Proc. Natl. Acad.
Sci. USA 99, 7136–7141]. Following its systemic administration, this synthetic ligand affected a range of important physiologicalprocesses including appetite, seizures and pain. Physiological activity of galnon could not be explained solely by the activationof the three known galanin receptors, GalR1, GalR2 and GalR3. Consequently, it was possible that galnon generates its manifoldeffects by interacting with other signaling pathway components, in addition to via GalR1-3. In this report, we establish that galnon:(i) can penetrate across the plasma membrane of cells, (ii) can activate intracellular G-proteins directly independent of receptor acti-vation thereby triggering downstream signaling, (iii) demonstrates selectivity for different G-proteins, and (iiii) is a ligand to otherG-protein coupled receptors (GPCRs) in addition to via GalR1-3. We conclude that galnon has multiple sites of interaction withinthe GPCR signaling cascade which mediate its physiological effects.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Galanin; Galnon; G-protein; Receptor; Signaling
0143-4179/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.npep.2005.09.005
Abbreviations: AR, adrenoreceptor; AVP, [Arg8]vasopressin; CHO, Chinese hamster ovary; D1(2)R, dopamine receptors, type 1 (2); DMSO,dimethylsulfoxide; GalR, galanin receptor; GPCR, G-protein coupled receptors; HKR, Hepes–Krebs–Ringer solution; HPLC, high performanceliquid chromatography; 5-HT, 5-hydroxytryptamine; IC50, concentration giving half-maximal inhibition; LC–MSD-Trap-XCT, liquid chromato-graph–mass spectrometer-ion trap instrument; LDH, lactate dehydrogenase; M1, (2,5) mAChR muscarinic acetylcholine receptor, type 1 (2, 5);mAChR, muscarinic acetylcholine receptor, not specified; MALDI-TOF MS, matrix assisted laser desorption/ionization-time-of-flight mass spec-trometer; MC3 (4), melanocortin receptor, type 3 (4); OTR, oxytocin receptor; PBS, phosphate buffered saline; PTX, pertussis toxin; PPS, perforantpath stimulation; RP, reverse phase; SSSE, self-sustaining status epilepticus; TFA, trifluoroacetic acid; V1aR, V1a vasopressin receptor; V1bR, V1b
vasopressin receptor; V2R, V2 vasopressin receptor* Corresponding author. Tel.: +46 816 1793; fax: +46 816 1371.E-mail address: [email protected] (U. Langel).

2 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
1. Introduction
G-protein coupled receptors (GPCR) comprise one ofthe most important pharmaceutical targets, with 60% ofall clinically prescribed drugs being GPCR ligands.Alternative therapeutic agents which bypass GPCR,but rather directly target downstream intracellular sig-naling cascades have not been developed to-date.
Galanin receptors (GalRs) are members of the rho-dopsin-like family A GPCR superfamily of proteins.Three subtypes of GalRs have been identified, termedGalR1, GalR2 and GalR3. In an attempt to generatea novel ligand for these receptors, a combinatoriallibrary was constructed based on a tripeptide whichincorporated the major pharmacophore of the naturalagonist galanin, namely Trp2, Asn5 and Tyr9. Screeningof this library identified galnon, 7-[(9-fluorenylmethoxy-carbonyl) cyclohexylalanyllysyl] amino-4-methylcouma-rin, a low molecular weight substance (MW of 677 Da)capable of displacing [125I]-porcine galanin binding tothe GalR1 with an affinity of 3–12 lM (Bartfai et al.,2004; Saar et al., 2002). Galnon was subsequently dem-onstrated to be an agonist at this receptor as it inhibitedadenylyl cyclase in rat hippocampal membranes (Saaret al., 2002), which are known to be rich in GalR1.
Administration of galnon generates a wide range ofphysiological effects. For example, galnon has beenreported to act as an anti-convulsant in vivo (Saaret al., 2002), to decrease symptoms of opiate withdrawal(Zachariou et al., 2003), to regulate appetite, therebyreducing food intake (Abramov et al., 2004), to inducelong-term potentiation in dentate gyrus of C57BL/6mice (Badie-Mahdavi et al., 2005), to increase insulinrelease in rat pancreatic islets (Quynh et al., 2005), toproduce an antidepressant effect in rats (Lu et al.,2005a), and to alleviate nociceptive responses to partialsciatic nerve injury (Wu et al., 2003); summarized in(Sollenberg et al., 2005). While some of the effects of gal-non have been shown to involve GalR, others cannot beexplained solely by this mechanism. Hence, additionalpathways to interaction of galnon with galanin receptorsmust be involved to explain the observed results. Forexample, intraperitoneally applied galnon prolongs heatwithdrawal latency in sciatic nerve injured rats in a dose-dependent manner. However, galnon does not affectmechanical or cold hypersensitivity, even though thiseffect was observed previously with intrathecally appliedgalanin (Wu et al., 2003). Furthermore, while intracere-broventricular injection of galanin stimulates food con-sumption (Kyrkouli et al., 1990), galnon causes apronounced, dose-dependent, reduction of food intakein rats and mice following both intraperitoneal andintracerebroventricular administration (Abramovet al., 2004). In vivo anticonvulsant activity of galnonwas comparable to that of galanin despite the differencesin the binding affinity between the two compounds (Saar
et al., 2002). Taken together, these data suggest that gal-non must have other modes of action in addition to thedirect activation of GalRs.
It has been reported previously that a small numberof compounds, such as neuropeptide Y and substanceP, and several basic secretagogues, can activate G-pro-teins directly in a receptor-independent manner, albeitat micromolar concentrations (Ferry et al., 2002; Ogawaet al., 1999; Raimondi et al., 2002). In this study, weestablish that galnon can cross the plasma membraneof cells to activate G-proteins directly and selectively.In addition, we demonstrate that galnon is also a ligandat GPCRs other than GalR1-3. The ability of a smallmolecule to affect G-proteins directly, rather than viaa GPCR, may provide a novel approach for pharmaceu-tical intervention of intracellular signaling.
2. Materials and methods
2.1. Chemicals and reagents
Lipofectamine and Fugene were purchased from LifeTechnologies/Invitrogen (Carlsbad, CA) and Roche(Basel, Switzerland), respectively. SKF38393 was pur-chased from Research Biochemicals International(Natick, MA). [125I]-(�) iodocyanopindolol, [N-Methyl-3H]- SCH23390, [125I]-galanin, [Phe3-3,4,5-3H]AVP, [N-methyl-3H] scopolamine and [Tyr-2,6- 3H]oxytocin wereall from Perkin–Elmer Life Sciences (Boston, MA). C18ZipTips were obtained from Millipore (Billerica, MA).Amino acids were from Novabiochem (Darmstadt, Ger-many). Cellmedia and reagents were purchased fromLife Technologies/Invitrogen (Carlsbad, CA) orSigma–Aldrich (St. Louis, MO). AVP and OT were pur-chased from Bachem Ltd (St. Helens, UK). Soluene 350(Packard, USA), HiSafe3TM (Wallac, UK). Carboxyflu-oroscein was purchased from Molecular Probes (Leiden,The Netherlands) and lactate dehydrogenase (LDH)leakage assay Cytotox-one, from Promega (SDSbio-sciences, Sweden). Additional chemicals and reagentswere all from SigmaAldrich (St. Louis, MO).
2.2. Cell cultures and transfection
HEK293 and chinese hamster ovary, CHO, cells weremaintained in Minimal Essential Medium and F-12Medium, respectively. COS-7 and HEK 293T were cul-tured in Dulbecco�s Modified Eagle�s Medium. Rinm5Fcells were grown in RPMI-1640 medium with 2 mM L-glutamine. Bowes human melanoma cells (AmericanType Culture Collection CRL-9607) were grown inEagle�s Minimal Essential Medium with Glutamax with1% non-essential amino acids and 1% sodium pyruvate.Sf9 cells were maintained in Grace�s insect medium. Allmedia were supplemented with 10% fetal bovine serum,

A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx 3
ARTICLE IN PRESS
100 units/ml penicillin and 100 lg/ml streptomycin.Cells were kept in 5% CO2 atmosphere at 37 �C, exceptSf9 cells that were kept at 28 �C.
Transient transfections of COS-7 and CHO cells werecarried out with Lipofectamine reagent, while HEK-293cells were transiently transfected with Fugene. Prepara-tion of CHO and HEK293 cells was as follows. Fortyeight hours following transfection, cells were lysed usinghypotonic lysis solution of 50 mM Tris–HCl, 4 mMEDTA, pH 7.4. HEK 293T cells were seeded at a densityof approximately 5 · 105 cells/100 mm dish and trans-fected after 48 h using a DNA-calcium phosphate co-precipitation protocol with 10 lg DNA/dish ofpcDNA3 expressing the respective receptor (Table 1).After incubation of 16 h, the medium was replaced withgrowth medium and cells incubated for a further 48 hbefore harvesting. A cell membrane preparation ofHEK 293T cells was prepared as described previously(Wesley et al., 2002). Sf9 cells were cotransfected withrecombinant Baculoviruses vectors carrying differentalpha subunits of heterotrimeric G-proteins (Gas,Gai1, Gao, Ga11) together with b1c2 subunits, asdescribed (Bavec et al., 2003). Briefly, approximately6 · 106 cells/75 cm2 flask were infected with high titerrecombinant Baculovirus stock solution. After 60 minof incubation at 28 �C the virus stock was removed,the fresh medium was added, cells were grown for threedays at 28 �C. Rat heart and hippocampus membranesprepared as described previously (Wheatley et al.,1997) were used as the source of M2 and M1 mAChRs,respectively.
2.3. Synthesis
Synthesis of the peptides was carried out on a model431A peptide synthesizer (Applied Biosystems, FosterCity, CA) using t-Boc or Fmoc strategies of solid-phasepeptide synthesis as described earlier (Langel et al.,1992). Galnon was synthesised following the schemedescribed by Saar et al. (2002). The purity of the prod-
Table 1G-protein coupled receptors used in displacement studies with galnon
Receptor IC50 (lM) Radioligand
GalR1 3.0 ± 0.2 [125I]-porcinb1-AR 3.6. ± 0.3 [125I]-(�)Iodb2-AR 16 ± 2 [125I]-(�)IodD1 R 18 ± 3 [N-Methyl-3
V1aR (rat) 5.9 ± 0.8 [3H]AVP #V1aR (human) 3.9 ± 1.0 [3H]AVP #V1bR > 10 [3H]AVP #V2R > 10 [3H]AVP #OTR 5.4 ± 1.5 [3H]oxytocimAChR 8 ± 0.5 [3H]QNB #M1mAChR No binding [3H]NMS #M2mAChR No binding [3H]NMS #
The IC50 values gathered, the receptors, and competing radioligand used.
ucts was analyzed by HPLC on an analytical Nucleosil120–3 C18 RP-HPLC column (0.4 · 10 cm). The molec-ular masses of the peptides were confirmed by MALDI-TOF Mass Spectrometer analysis (Applied Biosystems,Foster City, CA).
2.4. Cellular uptake of galnon
2.4.1. Quantitative detection of intracellular galnon by
fluorometry
Human Bowes melanoma and CHO cells were usedfor the uptake studies. Stock solutions of tested drugswere prepared from 1 mM to 10 and 25 lM (in phos-phate buffered saline, PBS, buffer, pH 7.4). Two hun-dred thousand cells/well were seeded one day beforeexperiments onto a 12-well plate and washed (2 · 1 mlPBS), then exposed to the drug in PBS at 37 �C for30 min at 300 rpm (on Thermo mixer, Eppendorf).
The drug treated cells were washed thoroughly withice cold PBS and then lysed in 0.1% Triton X-100 inHKR at 4 �C for 10 min. The cell lysates were trans-ferred to a black 96-well plate for fluorescence readoutat 492/520 nm (carboxy fluorescein) and 325/415 nm(galnon�s substituted coumarin as determined by spectraanalysis). The samples were compared to the fluores-cence of the added amount of drug and the total proteinamount (nmol/mg) as determined by protein determina-tion kit (Bio-Rad).
In parallel with the translocation studies, the mem-brane disturbance was determined by lactate dehydroge-nase (LDH) leakage assay. Samples were collected fromthe supernatant of drug solution on top of the cells at30 min. The LDH analysis was carried out accordingto the manufacturer�s description. Data are given as per-centage of 0.1% Triton X-100 treated cells LDH release.
2.4.2. Qualitative mass-spectrometry
Bowes human melanoma cells and CHO cells wereseeded in 6-well plates (100000 cells/well). When sub-confluent (48 h), the cells were washed twice with
(agonist #, antagonist ##) Source
e galanin # Bowes melanoma cellsocyanopindolol ## CHO cellsocyanopindolol ## CHO cellsH]SCH23390 ## HEK 293 cells
HEK 293T cellsHEK 293T cellsHEK 293T cellsHEK 293T cells
n # HEK 293T cells# Rat cortex# Rat hippocampus# Rat heart

4 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
Hepes–Krebbs–Ringer solution (HKR). One milliliter oftest substance (10 lM in HKR) was added per well andthe plate was incubated under 300 rpm rotation for60 min at 37 �C. After incubation the cells were washedwith HKR, followed by treatment with trypsin/EDTA(10 mg/ml trypsin, 2 mg/ml EDTA) for 2 min at roomtemperature. The trypsin solution was carefully aspi-rated and the cells were washed three additional times.Subsequently, the cells were lysed in 0.1% HCl, 200 ll/well, for 10 min on ice. The lysate was purified withC18 ZipTips according to the manufacturer�s instruc-tions. Mass spectrometry was performed with a-cya-nohydroxycinnamic acid type matrix (10 mg/ml in 50%AcN in 0.1% TFA/H2O), the data were collected fromat least three different locations on the spot andaccumulated. Analysis of the mass spectrometry datawas performed on a Voyager-DE STR (Applied Biosys-tems, Foster City, CA) and processed on the freewareMoverZ program (http://65.219.84.5/moverzDL.html).
2.4.3. Quantitative mass-spectrometryRinm5F cells were cultivated in 24-well plates to 50%
confluence and treated with 10 lM galnon in HKR.After 1 h incubation at 37 �C the extracellular fractionwas collected and saved for further analysis. Cells werewashed three times with HKR and thereafter lysed in0.1 M HCl by four freeze–thaw cycles. For reference,galnon was mixed with un-treated cell lysates or extra-cellular fractions to obtain 10, 5 and 1 lM final concen-trations in respective matrix. Samples were purified byC18 ZipTips (Millipore, Billeriva, MA, USA) accordingto the manufacturer�s protocol. Quantification was per-formed on an Agilent 1100 LC–MSD-Trap-XCT. Thegradient water–acetonitrile (0.1 % HCOOH) was usedfor separation. Iontrap mass detector was set to registeronly masses 678 ± 0.5 Da. Quantitative values wereobtained by comparison of peak heights in mass spectraof unknown samples and known concentrations.
2.5. GTPcS binding assay
Frozen pieces of rat brain and porcine arterial tissuewere mechanically pulverized, after which the mem-branes were obtained according to the protocol ofMcKenzie (McKenzie, 1992) with minor modifications(Bavec et al., 1999). The membranes were stored frozenat the protein concentrations of 1.5 mg/ml (brain) and0.6 mg/ml (porcine coronary artery), respectively, asdetermined by the method of Lowry et al. (Lowryet al., 1951). The modified protocol of McKenzie wasused also for the preparation of membranes from cells;protein concentrations of these membrane preparationswere between 0.2 and 0.4 mg/ml.
The rate of GTPcS binding to G-proteins from mem-branes was followed as described by McKenzie
(McKenzie, 1992). Briefly, the membranes (final proteinconcentration in the assay mixture was 0.05 mg/ml)were incubated for 3 min in the absence or presence ofgalnon in different concentrations with 0.5 nM[35S]GTPcS at 13 �C in TE-buffer (10 mM Tris–HCl + 0.1 mM EDTA), pH 7.5. The unbound[35S]GTPcS was removed by rapid filtration of the reac-tion mixture through Millipore GF/C glass-fibber filtersunder vacuum. The remaining radioactivity in filters wasdetermined in the LKB 1214 Rackbeta liquid scintilla-tion counter. Galnon was dissolved in 100% DMSOand the final DMSO concentration in all experimentalmixtures was kept fixed at 5%.
2.6. Receptor binding, interference by galnon
Receptor binding assays were used to quantify theability of galnon to displace radioactively labelledligands on cells/tissue expressing various G-protein cou-pled receptors, according to Table 1. The assay for bind-ing to galanin receptors on Bowes melanoma cells ispreviously described in detail by Land et al. (Landet al., 1991). For binding studies of b1 and b2 adrenergicand the D1 dopamine receptors 20 lg of cell lysate fromthe supernatant was aliquoted into sample tubes, receiv-ing one of the following solutions: (1) radioligand alone,(2) radioligand and antagonist, (3) radioligand plus var-ious concentrations of galnon or (4) radioligand, com-peting antagonist, and galnon. Antagonists for b1 andb2 adrenergic receptors; [125I]-(�)iodocyanopindolol(radioligand), and alprenolol (competing antagonist)were present at 166 pM and 1 lM concentrations,respectively. D1 dopamine receptor antagonist,[N-methyl-3H]-SCH23390, was used at 15 nM concen-tration and was competed off using 10 lM of the D1specific agonist SKF23383.
Binding assays of the vasopressin receptors, oxytocinreceptors and muscarinic acetylcholine receptors (rathippocampus and heart) were essentially as describedpreviously (Hawtin et al., 2001) comprising membranes(50–300 lg protein) diluted in binding buffer (20 mMHepes, 1 mM EGTA, 10 mM Mg(CH3COO2)2, 1 mg/ml BSA; pH 7.4), radioligand and competing ligand atthe concentration indicated in a final volume of 0.5 ml.Non-specific binding was defined by a saturating con-centration (1 lM) of unlabeled [Arg8]-vasopressin(AVP), oxytocin or atropine for the vasopressin recep-tors, oxytocin receptors and muscarinic acetylcholinereceptors, respectively. After incubation for 90 min at30 �C to establish equilibrium, bound and free ligandswere separated by centrifugation at 12,000g for10 min. Membrane pellets were washed twice with waterand solubilized with 50 ll of Soluene 350 prior to addi-tion of 1 ml HiSafe3TM liquid scintillation cocktail forcounting. Binding data were analysed by non-linearregression to fit theoretical Langmuir binding isotherms

A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx 5
ARTICLE IN PRESS
to the experimental data using the GraphPad Prism pro-gram (GraphPad Software Inc., San Diego, USA).
2.7. 3D visualization of galnon binding to G-proteins;
docking
To explain the effects of galnon on Gi or Gs, and tounderstand the putative interaction between galnonand G-proteins, we performed 3D modeling of galnonwith the a-subunits. We based the modeling on twoassumptions; in the first approach we modeled galnonmanually on the backbone of the only lysine-containingloop on the Gi-b subunit and adenylate cyclase, respec-tively, which interact with the corresponding a-subunit.In our other approach, we initially built and optimized a3D structure of galnon quantum mechanically, takinginto account transconfiguration of the three amidebonds. Based on kinetic data and structural similarity,we identified a guanine-binding cavity on the Ga as apotentially attractive surface for the substituted couma-rine moiety of galnon. Additionally, the entrance intothis cavity serves as a portion of buried surface, whichmight accept the remaining two hydrophobic radicals.After docking galnon in this way, molecular dynamicsimulations were performed. For the protein models,we used the crystal structures of a subunits Gi (1AS0)and Gs (1AZS) (Raw et al., 1997), respectively. The pro-tein molecule was put in a cube of water molecules(10120), subjected to 150 relaxation steps (50 steps ofsteepest descent optimization, 50 steps of optimizationby adopted basis Newton–Raphson method, 50 stepsof descent lattice optimization) and followed by 100 psconstant pressure and temperature (CPT) dynamic sim-ulation (300 K, 1 bar, time step of 1 fs) invoking theEWALD summation for calculating the electrostaticinteractions, with the aim to check the stability of theproposed complex. For optimization and dynamics cal-culations, we used CHARMM molecular simulationprogram (Brooks et al., 1983) and for docking and qual-ity checks, we used WHATIF molecular modeling anddrug design program (Vriend, 1990). The remainingthree galnon residues were modeled in the conformationsuperimposed to the backbone of each particular loop,accordingly.
2.8. Effect of galnon on seizures
The experiments were performed on 10–12 week-oldmale Wistar rats (Harlan, Indianapolis, IN). The studieshad been approved by West Los Angeles Animal Careand Use Committee and conducted according to theNIH guidelines. Under isoflurane anesthesia, the ani-mals were stereotaxically implanted with the bipolarelectrode–cannula guide (this and other electrodes,cannulas from PlasticsOne, Roanoke, VA) into the leftdentate gyrus (4.16 mm ventral, 2.5 mm lateral from
Bregma, 3.5 mm down from brain surface) and a guidecannula into the symmetrical side of the right dentategyrus. Bipolar stimulating electrode was placed intothe left angular bundle of perforant path (4.5 mm leftfrom lambda, 0.5 mm rostral from lambdoid fissure).After one-week recovery, self-sustaining status epilepti-cus (SSSE) was induced as described earlier (Mazaratiet al., 1998). Briefly, the animals were subjected to30 min of perforant path stimulation using Grass stimu-lator model 8800, for 30 min with the following param-eters: 10 s 20 Hz trains of 1 ms 30 V pulses deliveredevery min, together with the continuous 2 Hz stimula-tion using the same parameters. Electrographic activitywas acquired and analyzed offline using Harmonie Soft-ware (Stellate Systems, Montreal, Quebec), configuredfor automatic detection and saving of seizures (ampli-tude threshold 2.5, detection threshold 0, minimal fre-quency 3 Hz, maximal coefficient of variation 60). Toquantify seizures during SSSE, cumulative seizure time,i.e., the time between the first and the last seizure lessinterictal activity, was calculated. PTX (List laborato-ries, Campbell, CA) was dissolved in sterile saline, andinjected in the amount of 2.5 lg/hippocampus in a vol-ume of 2.5 ll using Hamilton microsyringe connectedto the injection cannula, with the latter placed into thelumen of guide cannula. Galnon was injected as a 5%DMSO solution in a dose of 10 nmole/side in a volumeof 2 ll. The following groups were used; Group 1. Con-trol, received vehicle and PPS; Group 2. PTX followedby DMSO (control for galnon), followed by PPS; Group3. Galnon, followed by saline (control for PTX) fol-lowed by PPS; Group 4. PTX followed by galnon fol-lowed by PPS; Group 5. Galnon, followed by PTXfollowed by PPS. Each group consisted of four animals.The time between PTX and other treatments was60 min; the time between galnon and other treatmentswas 15 min. Data were analyzed by One-way ANOVAfollowed by posthoc Student–Newman–Keuls test formultiple comparisons, using Sigma Stat software (SPSS,Chicago, IL, USA); p values less than 0.05 wereaccepted as statistically significant.
2.9. Effect of galnon on porcine coronary artery
Dissected porcine hearts (260–390 g) were trans-ported in ice-cold Krebs–Henseleit solution (119 mMNaCl, 23.8 mM NaHCO3, 3 mM KCl, 1.14 mMNaH2PO4, 1.63 mM CaCl2, and 16.5 mM glucose).The left anterior descending coronary artery and greatcardiac vein were isolated. Blood vessel contractionmeasurement was performed as already described in(Japelj et al., 1999). In brief, 5 mm wide rings of theblood vessels were cut and mounted into 10 ml tissuechambers, filled with Krebs–Henseleit solution (37 �C,pH 7.4), which was oxygenated with a mixture of 95%O2 and 5% CO2. Initial tension of 50 mN was applied

20
25
30
35
40
45a
mo
l/m
g
6 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
and after equilibration 30 mM KCl was added to obtainstable isometric contraction. The presence of endothe-lium was verified by application of substance P
(0.1 lM). After washing out of KCl and substance P
and after equilibration of the system was restored,72 lM galnon was added into the tissue chamber andblood vessel contraction was recorded.
m/z675.0 715.0
Inte
nsity
40
0 1
7
36
700.98679.00
II
III
.00 1
7
36
.0
galnon 10
µM
galnon 25
µM
carb
oxyflu
orosc
ein 5µ
M
penet
ratin
5µM
0
5
10
15
b
nI
Fig. 1. Intracellular localization of galnon. (a) Quantitative cellularuptake determined by spectrofluorometry. Galnon�s fluorescence wasdetermined at 325em and 415ex nm in PBS. The diagram shows theamount of galnon, nmol/mg protein, detected intracellularly after30 min of incubation in two different cell lines. Grey bars representCHO cells and white bars represent Bowes melanoma cells. (b)Translocation of galnon (MW 678 Da) into Bowes human melanomacells, analyzed by mass spectrometry. Experiments were conducted inparallel on CHO cells with comparable results (data not shown). Thelabeled peaks correspond to galnon (MW + H+) and its Na salt,respectively. I spectra from purified cell lysate from untreated cells, IIspectra from purified cell lysate from galnon treated cells, III spectrafrom purified extracellular media from galnon treated cells. Forexperimental details see Section 2.
3. Results
3.1. Galnon accesses the intracellular compartment of
cells
For galnon to directly activate downstream compo-nents of a GPCR signaling cascade, it must first gaindirect access to the latter. Therefore, a pre-requisite tosuch a mechanism of action is that galnon must be ableto penetrate the plasma membrane. Possible cellularuptake of galnon was studied by two methods, fluores-cence spectrometry and mass spectrometry.
3.1.1. Detection of intracellular galnon by fluorometry
The intracellular localization of galnon was demon-strated directly by utilizing the natural fluorescence ofthe coumarinyl moiety within the galnon structure, theinternalised galnon could be determined in cell lysates,and correlated to a reference standard of known concen-trations of galnon. The uptake of galnon was deter-mined at 10 and 25 lM, and was found to be dosedependent (Fig. 1(a)). The result did not differ signifi-cantly with a trypsin treatment of the cells prior to lysis(data not shown), eliminating the possibility that galnonmay be trapped in extracellular protein structures. Thegalnon translocation was dose-dependent with 24.0and 40.9 nmol/mg protein for 10 and 25 lM in Bowescells, respectively. The translocation was generally lowerin CHO cells; 10 and 19.8 nmol/mg protein for 10 and25 lM galnon, respectively. In addition, as control sub-stances, carboxy fluorescein and the well characterizedcell penetrating peptide penetratin (Derossi et al.,1998) were used (Fig. 1(a)). Carboxy fluorescein onlyentered the cells in very low amounts, below 1 nmol/mg protein, while penetratin entered 5.4 nmol/mg inBowes cells and 1.4 nmol/mg in CHO cells. The differ-ence of drug translocating into the two different cell linesmay reflect the difference in cell size. LDH release indi-cates cell membrane disturbance. At 10 lM galnon,there was no significant release of LDH, but at 25 lM,15–20% of LDH was released.
3.1.2. Qualitative mass spectrometry
To further confirm the intracellular location of gal-non, Bowes human melanoma and CHO cells wereexposed to it (10 lM) for 60 min at 37 �C. Intracellulardistribution of galnon was then investigated by using
MALDI-TOF mass spectrometry to analyze cell lysateas described in Section 2. Fig. 1(b) (I and III) shows thatthe presence of galnon (678 Da) in the extracellular med-ium which could be identified by two characteristicpeaks, m/z = 678.00 (MH+) and 700.98 (MNa+). These�signature peaks� are absent in the lysate of control cellsbut are present in the lysate of cells which were exposedto galnon (Fig. 1(b), I and II). Although results fromthis analysis are strictly qualitative, they, nevertheless,
-7 -6 -5 -460
80
100
120
140
160
180
//
//
log[galnon, M]
rate
of
[35S
]GT
PγS
bind
ing,
%of
bas
al
80
100
120
140
160
180
//
rate
of
[35S
]GT
PγS
bind
ing,
%of
bas
al
a
b
A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx 7
ARTICLE IN PRESS
establish that galnon effectively penetrates the plasmamembrane. As a control, the cell-penetrating peptidepVEC (Elmquist et al., 2001) showed the correspondinguptake pattern, similar to that obtained with galnon(data not shown). Only data on Bowes cells are shownfor clarity, the spectra of uptake in CHO cells showthe corresponding pattern.
3.1.3. Quantitative mass-spectrometry
The amount of galnon translocated was confirmed inanother cell line Rinm5F, and with a LC–MS setup. Theresults indicate that 26 ± 5% of total galnon is found inthe intracellular fraction at given experimental condi-tions (data not shown), which is comparable to the datafrom the quantitative fluorometry at 10 lM galnonexposure. Since the peak intensity in mass-spectra highlydepends on total ion composition, i.e., other substancespresent in sample, intra- and extracellular concentra-tions of galnon were estimated separately.
-7 -6 -5 -460 //
log[galnon, M]
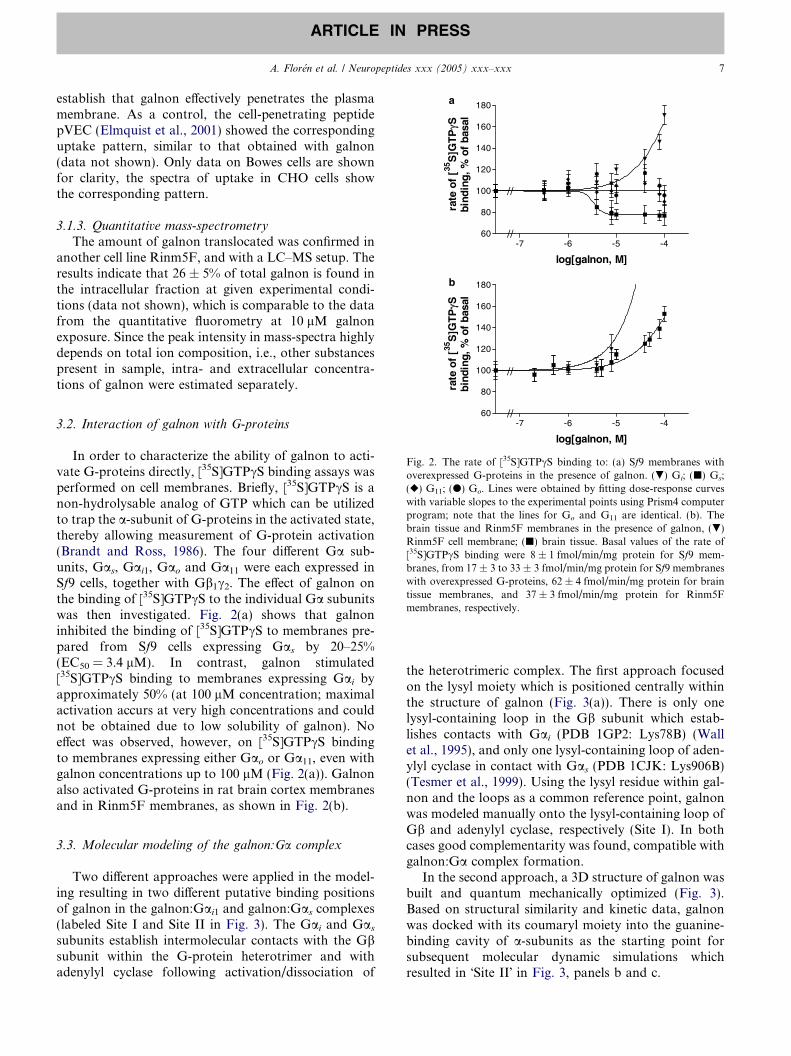
Fig. 2. The rate of [35S]GTPcS binding to: (a) Sf9 membranes withoverexpressed G-proteins in the presence of galnon. (.) Gi; (j) Gs;(r) G11; (d) Go. Lines were obtained by fitting dose-response curveswith variable slopes to the experimental points using Prism4 computerprogram; note that the lines for Go and G11 are identical. (b). Thebrain tissue and Rinm5F membranes in the presence of galnon, (.)Rinm5F cell membrane; (j) brain tissue. Basal values of the rate of[35S]GTPcS binding were 8 ± 1 fmol/min/mg protein for Sf9 mem-branes, from 17 ± 3 to 33 ± 3 fmol/min/mg protein for Sf9 membraneswith overexpressed G-proteins, 62 ± 4 fmol/min/mg protein for braintissue membranes, and 37 ± 3 fmol/min/mg protein for Rinm5Fmembranes, respectively.
3.2. Interaction of galnon with G-proteins
In order to characterize the ability of galnon to acti-vate G-proteins directly, [35S]GTPcS binding assays wasperformed on cell membranes. Briefly, [35S]GTPcS is anon-hydrolysable analog of GTP which can be utilizedto trap the a-subunit of G-proteins in the activated state,thereby allowing measurement of G-protein activation(Brandt and Ross, 1986). The four different Ga sub-units, Gas, Gai1, Gao and Ga11 were each expressed inSf9 cells, together with Gb1c2. The effect of galnon onthe binding of [35S]GTPcS to the individual Ga subunitswas then investigated. Fig. 2(a) shows that galnoninhibited the binding of [35S]GTPcS to membranes pre-pared from Sf9 cells expressing Gas by 20–25%(EC50 = 3.4 lM). In contrast, galnon stimulated[35S]GTPcS binding to membranes expressing Gai byapproximately 50% (at 100 lM concentration; maximalactivation accurs at very high concentrations and couldnot be obtained due to low solubility of galnon). Noeffect was observed, however, on [35S]GTPcS bindingto membranes expressing either Gao or Ga11, even withgalnon concentrations up to 100 lM (Fig. 2(a)). Galnonalso activated G-proteins in rat brain cortex membranesand in Rinm5F membranes, as shown in Fig. 2(b).
3.3. Molecular modeling of the galnon:Ga complex
Two different approaches were applied in the model-ing resulting in two different putative binding positionsof galnon in the galnon:Gai1 and galnon:Gas complexes(labeled Site I and Site II in Fig. 3). The Gai and Gassubunits establish intermolecular contacts with the Gbsubunit within the G-protein heterotrimer and withadenylyl cyclase following activation/dissociation of
the heterotrimeric complex. The first approach focusedon the lysyl moiety which is positioned centrally withinthe structure of galnon (Fig. 3(a)). There is only onelysyl-containing loop in the Gb subunit which estab-lishes contacts with Gai (PDB 1GP2: Lys78B) (Wallet al., 1995), and only one lysyl-containing loop of aden-ylyl cyclase in contact with Gas (PDB 1CJK: Lys906B)(Tesmer et al., 1999). Using the lysyl residue within gal-non and the loops as a common reference point, galnonwas modeled manually onto the lysyl-containing loop ofGb and adenylyl cyclase, respectively (Site I). In bothcases good complementarity was found, compatible withgalnon:Ga complex formation.
In the second approach, a 3D structure of galnon wasbuilt and quantum mechanically optimized (Fig. 3).Based on structural similarity and kinetic data, galnonwas docked with its coumaryl moiety into the guanine-binding cavity of a-subunits as the starting point forsubsequent molecular dynamic simulations whichresulted in �Site II� in Fig. 3, panels b and c.

Fig. 3. Structure of galnon (a) and two putative positions of galnon incomplex with a subunits of Gi (b) and Gs (c), respectively. (a) 3Dstructure of galnon, by CHARMM molecular simulation program. (b)The lysyl moiety of galnon is superimposed on the only lysinecontaining loop of Gb in contact with Gai (b; Site I) and galnon�scoumarin moiety is superimposed on guanine base of GTP in thenucleotide binding site of Gai (b; Site II). (c) The lysyl moiety ofgalnon is superimposed on the only lysine containing loop of adenylatecyclase in contact with Gas (c; Site I) and galnon�s coumarin moiety issuperimposed on guanine base of GTP in the nucleotide binding site ofGas (c; Site II). For details see text. In panels (b) and (c) galnon isyellow and GTP is red.
Fig. 4. Anticonvulsant effects of galnon are Gi/o protein dependent.Self-sustaining status epilepticus (SSSE) was induced in rats by 30 minperforant path stimulation. Seizure activity was quantified by calcu-lating the total time spent in electrographic seizures (cumulative seizuretime). The table under the x-axis depicts the sequence of treatments. *,p < 0.02 vs control (Bartfai et al., 2004); #, p < 0.05 vs Group 2; �,p < 0.05 vs Group 3; +, p < 0.05 vs Group 4 (One-way ANOVA withStudent–Newman–Keuls post hoc test).
8 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
3.4. Effect of galnon on GPCR ligand binding
The effect of increasing galnon concentrations onradioligand binding to a range of GPCRs was investi-gated, Table 1. Galnon displaced binding to bothbiogenic amine receptors (b1-, and b2-adrenergic recep-tors, D1 dopamine receptor) and to peptide hormonereceptors (V1a vasopressin receptor and the oxytocinreceptor) in the low micromolar range. This effect of gal-non was observed when either agonist or antagonist wasused as radio-tracer in the ligand binding assay.
Although galnon displaced the binding of radioligandto several GPCRs, galnon binding was neverthelessselective, as the compound did not bind to all of theGPCRs investigated. For example, although galnonbound to the V1a vasopressin receptor, it did not bindto the V2 vasopressin receptor (Table 1).
3.5. Effect of galnon on seizures in pertussis toxin
(PTX)-treated rats
We have previously established the anticonvulsanteffects of galnon. In addition, we have shown that
pertussis toxin (PTX), which prevents activation of Gi/
o-protein, augments the severity of seizures during self-sustaining status epilepticus (SSSE) in the rat whenadministered bilaterally into the dentate gyrus (2.5 lg/side, but not 0.1 or 0.5 lg/side) (Mazarati and Lu,2005). Consequently, we utilized this system to examinethe regulation of G-protein signaling by galnon.
Galnon (10 nmole/side) administered bilaterally intothe hippocampus in the absence of PTX significantlyshortened seizure duration (cumulative seizure time51 ± 5.5 min vs. 316 ± 11 min in control). However, inPTX-pretreated animals galnon completely failed toinhibit seizures (seizure time 621 ± 28 min comparedto 668 ± 37 min after PTX treatment only) (Fig. 4).When the sequence of the treatments was reversed, i.e.,the animals had first been treated with galnon and theninjected with PTX, galnon significantly shortened cumu-lative seizure time (435 ± 19 min) as compared to PTX-treated animals, although seizures still lasted longer thaneither controls or galnon-only treated rats (Fig. 4).
3.6. Effect of galnon on porcine coronary artery
Relaxation of blood vessels is a response caused byseveral second messengers originating from stimulationof GPCRs of the endothelium; we used a model systemto confirm the ability of galnon to activate these cas-cades. Dissected porcine coronary artery was contractedwith KCl, after which galnon was applied. A very slightrelaxation was observed initially, followed after 20–

Fig. 5. Effect of galnon on porcine coronary artery. Arrows indicatethe following procedures: (1) application of KCl, (2) washing out, (3)application of 1% DMSO, (4) application of galnon.
A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx 9
ARTICLE IN PRESS
25 min by a pronounced relaxation (Fig. 5). Further-more, this relaxation could not be reversed by additionalKCl treatment, thereby indicating that the effect of gal-non was mediated by an intracellular, rather than anextracellular, site of action. Galnon showed no effecton relaxed arteries.
4. Discussion
The first systemically active galanin receptor agonist,galnon, has been shown to generate several importantbiological effects which cannot be explained by assumingthat galnon only exhibits a single mode of action,namely activation of GalR signaling. In this study, weshow that galnon can stimulate G-proteins directly, inaddition to eliciting effects via activation of GalR1-3and several other GPCRs.
A pre-requisite to direct activation of G-proteinswould be the ability of galnon to translocate acrossthe plasma membrane of cells in order to gain accessto the G-proteins. Unfortunately, 7-amino-4-methyl-coumarin is not the best fluorophore for microscopy,as the weak signal is hampered by photo-bleachingand auto-fluorescence. However, we utilized a combina-tion of mass spectrometry and fluorescence spectrome-try to establish the cellular uptake of galnon intoseveral mammalian cell lines, which occurs after only ashort incubation at 37 �C. This is in accordance withother small, lipophilic compounds that readily traversethe lipid bilayer. The observation that galnon-inducedblood vessel relaxation showed a lag-period of 10–15 min which is consistent with our suggestion that someGalR-independent effects of galnon are generated onlyafter intracellular accumulation of the compound. Thislag-period may reflect the time required for galnon to
translocate into the cytoplasm of cells and to achieve asufficient intracellular concentration to mediate itseffects. Moreover, the lack of effect of washing-outextracellular galnon, together with the inability of extra-cellular KCl to reverse galnon�s action, strongly indi-cates an intracellular site of action for galnon-inducedblood vessel relaxation.
Multiple GPCRs are expressed by blood vessels, andgalnon could cause its effects by interacting with thesesince it has been reported recently that galnon recog-nizes with low affinity several 7TM receptors such as5-HT1A and -1B, a1 adrenergic, D2, M5, ghrelin,MC3, MC4 and Y1 NPY receptors (Lu et al., 2005b).However, it is unlikely that the galnon-induced bloodvessel relaxation in the present study was mediated byactivation of GPCRs (such as b-adrenergic receptors,see Table 1) due to the difference in kinetics as shownin the case of isoproterenol-induced relaxation of bloodvessels (Chruscinski et al., 2001).
One could argue that the DMSO present in mostexperiments as a vehicle could enhance the intracellulardelivery of galnon since, due to low solubility of galnon,the complete removal of DMSO from these experimentsis not feasible. Moreover, as most in vivo experimentsare obtained with 1% DMSO in the vehicle, it wouldbe unwise to exclude this parameter from the uptake-verification. The presence of 1% DMSO does not affectthe viability of Rinm5F cells and the membrane integrityand it does not influence the internalization of cell-pene-trating peptides, nor does it induce the internalization ofmembrane impermeable agents such as FITC-labeledavidin (data not shown), suggesting that the registereduptake of galnon is not induced by the presence ofDMSO.
Having established that galnon accumulated in theintracellular compartment of cells to access G-proteins,we then investigated whether direct galnon:G-proteininteraction actually occurred. In order to do this, wegenerated recombinant Sf9 cell membranes expressingdifferent individual G-proteins. Subsequent [35S]GTPcSbinding experiments demonstrated that galnon doesindeed interact with G-proteins, resulting in the inhibi-tion of Gas and the activation of Gai. Furthermore, gal-non also stimulated a dose-dependent increase in[35S]GTPcS binding in brain tissue. The concentrationrange in which galnon affected G-proteins (viewed inFig. 2) was consistent with the concentration range inwhich it induced the relaxation of coronary artery(Fig. 5).
Molecular modeling of galnon:Ga complexes wasused to provide insight into a feasible mechanism bywhich galnon could regulate G-protein activation. Onthe basis of the high flexibility of the Ga observed bythe molecular dynamic simulations, we first hypothe-sized allosteric cooperativity between the suggestedbinding site of galnon (Site I in Fig. 3(b) and (c)) and

Fig. 6. Possible interaction scheme of galnon with GPCRs and Ga-proteins. A ‘‘two-state’’ model showing GPCR on the cell membranerecognizing galnon (a), inside the cell (b) galnon interacts directly withGa.
10 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
the GTP/GDP binding site. We assumed that two differ-ent conformations of Ga could be adopted when incomplex with galnon: a relaxed GTP preferring confor-mation in Gi, and a tenser GDP preferring one in Gs.Under resting conditions, G-protein heterotrimers arein equilibrium with their dissociated forms and galnoninterferes by shifting equilibrium towards free Ga. Con-sequently, GTP turnover substantially increases in Gi,but only moderately decreases in Gs. One could inferthat, in comparison to Gi, more than 10 times higheraffinity of galnon towards Gs (estimated from Fig. 2),provides energy for the stabilization towards GDP-likeconformation, thus lowering the rate of GTPcSassociation.
Another explanation considers G-proteins as notori-ously inefficient enzymes that, in most cases, encountertheir substrates non-productively. Galnon might inter-fere with this process in two ways: it can compete withits coumarinyl or Fmoc moiety for the guanine bindingpocket (Site II in Fig. 3(b) and (c)), or it can bind to themouth of this pocket and sterically hinder the entranceor exit of the nucleotide. We have tested this possibilityby the kinetic re-examination of the data previouslypublished (Ferguson et al., 1986; Sternweis and Robi-shaw, 1984). While the explanation of galnon inhibitionwas possible by its slow binding, we could only explainthe increased initial rates of GTPcS binding, as observedin the case of Gi, by the modified allosteric mechanismaccording to Monod et al. (Monod et al., 1965).Namely, we assumed the specific affinities of galnonand GTPcS for two different conformations of the Ga.Once with GTPcS in the pocket, galnon can only bindto the mouth of the active site according to rapid equi-librium, thus hindering the exit of the substrate ana-logue, or it may enter as a slow binding ligand intothe GDP/GTP-binding pocket of the free Ga and in thisway lock it in GDP-binding conformation. The shift ofequilibrium between two conformations, resulting inopposite effects on Gi and Gs, now depends only onthe relative affinities in each event.
In seizure models, the effects of galnon were com-pletely prevented by PTX, indicating complete depen-dence of anticonvulsant effects of galnon on Gi/o
proteins or on Gi/o coupled receptors interacting withgalnon. However, the inefficiency of galnon in PTX-pre-treated animals could result from higher severity ofSSSE, which could become resistant to therapeutic inter-ventions. To address this concern, we reversed thesequence of treatments, and injected galnon prior toPTX, to allow galnon interaction with G-protein. Underthese conditions, galnon exhibited significant anticon-vulsant activity. The fact that in this scenario galnondid not completely inhibit seizures, as it did in PTX-freeanimals, not even returned seizure severity to there con-trol level, is probably due to the fact that G-proteinscoupled to other GPCR were still inhibited by PTX, par-
ticularly those coupled to receptors for other hippocam-pal neuropeptides with anticonvulsant properties (e.g.,somatostatin, or Neuropeptide Y). Alternatively, theeffects of galnon on G-protein at the applied concentra-tion simply did not match PTX-induced inhibition of Gi/
o. However, examination of this possibility was compli-cated because PTX increased seizure severity in a dose-independent manner (lower doses of 0.2 and 1 lg didnot affect seizures at all), and also because the anticon-vulsant effects of galnon were saturated at the applieddose (Saar et al., 2002).
When galnon displaces radioligand from a GPCR, itexhibits a consistent low apparent affinity. We proposetwo theoretical models of why galnon is able to displacea native ligand from its receptor. In the first model, gal-non is taken up by the cell and interacts with G-proteinsin a receptor independent manner. The interaction ofgalnon and G-proteins leads to a subsequent allostericconformational change of the receptor and dissociationof the original ligand. This scenario is in accordancewith almost the same value of EC50 obtained with alltested receptors, which also roughly matches the concen-trations of galnon that are operating in blood vessel con-traction and direct G-protein activation. However, notall obtained data are consistent with this scenario, sincegalnon affected the binding of some antagonists to thereceptors, see Table 1, and also failed to affect allGPCRs investigated. For example, galnon decreasedbinding to b-AR but did not affect the binding of AVPto the V2R even though both of these receptors coupleto Gs, see Table 1. The second alternative is simply thatgalnon has an affinity for GPCRs and hence competeswith the ligand for its binding site, see Fig. 6. All resultspresented in this study were found at relatively highdoses/concentrations of galnon, why we consider directG-protein activation an alternative pathway alongsidereceptor binding.
As GPCR/G-protein signaling is of such great impactin almost all physiological events and the target of themajority of today�s drugs, the possibility to syntheticallymanufacture small molecules that directly affect G-pro-teins can be a new area for drug targeting. In conclusion,we present evidence for galnon being a low MW sub-stance with multiple important biological actions, recog-nizing several G-protein coupled receptors and directly

A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx 11
ARTICLE IN PRESS
activating G-proteins in a receptor independent manner,additionally to its galanin–receptor interactions. Thislatter novel pathway for synthetic low MW moleculesis similar to the direct G-protein coupled receptor acti-vation, previously proposed for small cationic peptideslike mastoparan.
Acknowledgments
The authors are grateful to Dr. Ursel Soomets forfruitful discussions and Dr. Tonu Pussa for access toLC–MS.This work was supported by grants from theSwedish Research Council (VR-Med), Research GrantNS043409 from NIH (Mazarati et al.) and Estonian Sci-ence Foundation Grant No. 6503 (Kilk, K.).
References
Abramov, U., Floren, A., Echevarria, D.J., Brewer, A., Manuzon, H.,Robinson, J.K., Bartfai, T., Vasar, E., Langel, U., 2004. Regula-tion of feeding by galnon. Neuropeptides 38, 55–61.
Badie-Mahdavi, H., Behrens, M., Rebek, J., Bartfai, T., 2005. Effect ofgalnon on induction of long-term potentiation in dentate gyrus ofC57BL/6 mice. Neuropeptides 39, 249–251.
Bartfai, T., Lu, X., Badie-Mahdavi, H., Barr, A.M., Mazarati, A.,Hua, X.Y., Yaksh, T., Haberhauer, G., Ceide, S.C., Trembleau,L., Somogyi, L., Krock, L., Rebek Jr., J., 2004. Galmic, anonpeptide galanin receptor agonist, affects behaviors in seizure,pain, and forced-swim tests. Proc. Natl. Acad. Sci. USA 101,10470–10475.
Bavec, A., Jureus, A., Cigic, B., Langel, U., Zorko, M., 1999.Peptitergent PD1 affects the GTPase activity of rat brain corticalmembranes. Peptides 20, 177–184.
Bavec, A., Hallbrink, M., Langel, U., Zorko, M., 2003. Different roleof intracellular loops of glucagon-like peptide-1 receptor in G-protein coupling. Regul. Pept. 111, 137–144.
Brandt, D.R., Ross, E.M., 1986. Catecholamine-stimulated GTPasecycle. Multiple sites of regulation by b-adrenergic receptor andMg2+ studied in reconstituted receptor-Gs vesicles. J. Biol. Chem.261, 1656–1664.
Brooks, B.R., Bruccoleri, R.E., Olafson, B.D., States, D.J., Swamina-than, S., Karplus, M., 1983. J. Comp. Chem. 4, 187–217.
Chruscinski, A., Brede, M.E., Meinel, L., Lohse, M.J., Kobilka, B.K.,Hein, L., 2001. Differential distribution of b-adrenergic receptorsubtypes in blood vessels of knockout mice lacking b(1)- or b(2)-adrenergic receptors. Mol. Pharmacol. 60, 955–962.
Derossi, D., Chassaing, G., Prochiantz, A., 1998. Trojan peptides: thepenetratin system for intracellular delivery. Trends Cell Biol. 8, 84–87.
Elmquist, A., Lindgren, M., Bartfai, T., Langel, U., 2001. VE-cadherin-derived cellpenetrating peptide, pVEC, with carrier func-tions. Exp. Cell Res. 269, 237–244.
Ferguson, K.M., Higashijima, T., Smigel, M.D., Gilman, A.G., 1986.The influence of bound GDP on the kinetics of guanine nucleotidebinding to G proteins. J. Biol. Chem. 261, 7393–7399.
Ferry, X., Brehin, S., Kamel, R., Landry, Y., 2002. G-protein-dependent activation of mast cell by peptides and basic secreta-gogues. Peptides 23, 1507–1515.
Hawtin, S.R., Tobin, A.B., Patel, S., Wheatley, M., 2001. Palmitoy-lation of the vasopressin V1a receptor reveals different conforma-tional requirements for signaling, agonist-induced receptor
phosphorylation, and sequestration. J. Biol. Chem. 276, 38139–38146.
Japelj, I., Budihna, M.V., Stanovnik, L., Krisch, I., 1999. Stereo-selective and endothelium-independent action of nicardipine onthe isolated porcine coronary artery. Eur. J. Pharmacol. 369, 43–47.
Kyrkouli, S.E., Stanley, B.G., Seirafi, R.D., Leibowitz, S.F., 1990.Stimulation of feeding by galanin: anatomical localization andbehavioral specificity of this peptide�s effects in the brain. Peptides11, 995–1001.
Land, T., Langel, U., Low, M., Berthold, M., Unden, A., Bartfai, T.,1991. Linear and cyclic N-terminal galanin fragments and analogsas ligands at the hypothalamic galanin receptor. Int. J. Pept.Protein Res. 38, 267–272.
Langel, U., Land, T., Bartfai, T., 1992. Design of chimeric peptideligands to galanin receptors and substance P receptors. Int. J. Pept.Protein Res. 39, 516–522.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951.Protein measurement with the Folin phenol reagent. J. Biol. Chem.193, 265–275.
Lu, X., Barr, A.M., Kinney, J.W., Sanna, P., Conti, B., Behrens,M.M., Bartfai, T., 2005a. A role for galanin in antidepressantactions with a focus on the dorsal raphe nucleus. Proc. Natl. Acad.Sci. USA 102, 874–879.
Lu, X., Lundstrom, L., Langel, U., Bartfai, T., 2005b. Galaninreceptor ligands. Neuropeptides 39, 143–146.
Mazarati, A., Lu, X., 2005. Regulation of limbic status epilepticus byhippocampal galanin type 1 and type 2 receptors. Neuropeptides39, 277–280.
Mazarati, A.M., Liu, H., Soomets, U., Sankar, R., Shin, D.,Katsumori, H., Langel, U., Wasterlain, C.G., 1998. Galaninmodulation of seizures and seizure modulation of hippocampalgalanin in animal models of status epilepticus. J. Neurosci. 18,10070–10077.
McKenzie, F.R., 1992. In: Milligan, E. (Ed.), Signal Transduction.Oxford University Press, Oxford, NY, Tokyo, pp. 33–39, 46, and52.
Monod, J., Wyman, J., Changeux, J.P., 1965. On the nature ofallosteric transitions: a plausible model. J. Mol. Biol. 12, 88–118.
Ogawa, K., Nabe, T., Yamamura, H., Kohno, S., 1999. Nanomolarconcentrations of neuropeptides induce histamine release fromperitoneal mast cells of a substrain of Wistar rats. Eur. J.Pharmacol. 374, 285–291.
Quynh, N.T., Islam, S.M., Floren, A., Bartfai, T., Langel, U.,Ostenson, C.G., 2005. Effects of galnon, a non-peptide galanin-receptor agonist, on insulin release from rat pancreatic islets.Biochem. Biophys. Res. Commun. 328, 213–220.
Raimondi, L., Banchelli, G., Matucci, R., Stillitano, F., Pirisino, R.,2002. The direct stimulation of Gi proteins by neuropeptide Y(NPY) in the rat left ventricle. Biochem. Pharmacol. 63, 2063–2068.
Raw, A.S., Coleman, D.E., Gilman, A.G., Sprang, S.R., 1997.Structural and biochemical characterization of the GTPcS-,GDP.Pi-, and GDP-bound forms of a GTPase-deficientGly42! Val mutant of Gialpha1. Biochemistry 36, 15660–15669.
Saar, K., Mazarati, A., Mahlapuu, R., Halnemo, G., Soomets, U.,Kilk, K., Hellberg, S., Pooga, M., Tolf, B.R., Shi, T.S., Hokfelt, T.,Wasterlain, C., Bartfai, T., Langel, U., 2002. Anticonvulsantactivity of a nonpeptide galanin receptor agonist. Proc. Natl. Acad.Sci. USA 99, 7136–7141.
Sollenberg, U., Bartfai, T., Langel, U., 2005. Galnon – a low-molecular weight ligand of the galanin receptors. Neuropeptides39, 161–163.
Sternweis, P.C., Robishaw, J.D., 1984. Isolation of two proteins withhigh affinity for guanine nucleotides from membranes of bovinebrain. J. Biol. Chem. 259, 13806–13813.

12 A. Floren et al. / Neuropeptides xxx (2005) xxx–xxx
ARTICLE IN PRESS
Tesmer, J.J.G., Sunahara, R.K., Johnson, R.A., Gosselin, G., Gilman,A.G., Sprang, S.R., 1999. Two metal ion catalysis in adenylylcyclase revealed by its complex with ATP analogs, Mg2+, Mn2+,and Zn2+. Science 285, 756–760.
Wall, M.A., Coleman, D.F., Lee, E., Iniguez-Lluhi, J.A., Posner,B.A., Gilman, A.G., Sprang, S.R., 1995. The structure of theG-protein heterotrimer Gi alpha1beta1gamma2. Cell 83, 1047–1058.
Wesley, V.J., Hawtin, S.R., Howard, H.C., Wheatley, M., 2002.Agonist-specific, high affinity binding epitopes contributed by theN-terminus of the oxytocin receptor are provided by a singlearginyl. Biochemistry 41, 5086–5092.
Wheatley, M., Howl, J., Yarwood, N.J., Hawtin, S.R., Davies, A.R.,Matthews, G., Parslow, R.A., 1997. Structure and function of
neurohypophysial hormone receptors. Biochem. Soc. Trans. 25,1046–1051.
Vriend, G., 1990. WHAT IF: a molecular modeling and drug designprogram. J. Mol. Graph. 8, 52–56, 29.
Wu, W.P., Hao, J.X., Lundstrom, L., Wiesenfeld-Hallin, Z., Langel,U., Bartfai, T., Xu, X.-J., 2003. Systemic galnon, a low-molecular weight galanin receptor agonist, reduces heathyperalgesia in rats with nerve injury. Eur. J. Pharmacol. 482,133–137.
Zachariou, V., Brunzell, D.H., Hawes, J., Stedman, D.B., Bartfai, T.,Steiner, R., Wynick, D., Langel, U., Picciotto, M.R., 2003. Theneuropeptide galanin modulates behavioral and neurochemicalsigns of opiate withdrawal. Proc. Natl. Acad. Sci. USA 100, 9028–9033.



















