
Reduced olfactory bulb volume and olfactory sensitivity in patients with acute major depression

Journal of Insect Physiology 59 (2013) 542–551
Contents lists available at SciVerse ScienceDirect
Journal of Insect Physiology
journal homepage: www.elsevier .com/ locate/ j insphys
Specificity and sensitivity of plant odor-detecting olfactory sensoryneurons in Ctenarytaina eucalypti (Sternorrhyncha: Psyllidae)
0022-1910/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jinsphys.2013.03.004
⇑ Corresponding author. Tel.: +46 (0) 46 2224997; fax: +46 (0) 46 2224716.E-mail addresses: [email protected] (J.K. Yuvaraj), martin_n.
[email protected] (M.N. Andersson), [email protected] (M.J. Stein-bauer), [email protected] (K. Farnier), [email protected](O. Anderbrant).
Jothi Kumar Yuvaraj a, Martin N. Andersson a, Martin J. Steinbauer b, Kevin Farnier b, Olle Anderbrant a,⇑a Department of Biology, Lund University, Sölvegatan 37, SE-223 62 Lund, Swedenb Department of Zoology, La Trobe University, Melbourne, VIC 3086, Australia
a r t i c l e i n f o a b s t r a c t
Article history:Received 11 December 2012Received in revised form 11 March 2013Accepted 12 March 2013Available online 21 March 2013
Keywords:Ctenarytaina eucalyptiOlfactory sensory neuronSingle sensillum recordingsHeadspace volatilesHost finding
The blue gum psyllid, Ctenarytaina eucalypti (Sternorrhyncha: Psyllidae), is an economic threat to Euca-lyptus subgenus Symphyomyrtus plantations worldwide. To date, no generally applicable control methodis available and the potential for semiochemical-based monitoring or control methods has not yet beeninvestigated. Hence, we conducted the first study on the olfactory sense of C. eucalypti, investigating thespecificity and sensitivity of its olfactory sensory neurons (OSNs) to host plant volatiles using single sen-sillum recordings (SSR). Synthetic compounds were selected from published identifications of Eucalyptusvolatiles and after analysis of headspace collections from Eucalyptus cordata. The antenna of C. eucalypticarries four cavities containing olfactory sensilla (S1–S4). Our recordings revealed that each of these sen-silla houses three OSNs that could be distinguished electrophysiologically based on spike amplitude dif-ferences (A, B, and C neuron with large, intermediate, and small amplitude, respectively). The A neuron insensillum S1 responded primarily to b-caryophyllene and weaker to b-ocimene, whereas the accompany-ing B-neuron responded strongly and very specifically to linalool. Furthermore, the B-neuron in both S2and S3 responded strongly to 1-hexanol, Z3-hexenol, and Z3-hexenyl acetate. OSNs in S4 responded onlyweakly to a few of the synthetic compounds. Response thresholds in strongly responding OSNs to puta-tive key compounds were close to the 1 ng dose on the filter paper and responses exhibited a phasic-tonicprofile irrespective of compound dose. C. eucalypti may use the physiologically active compounds forlong-range host finding. Future laboratory and field experiments will reveal whether plant volatilescan be used in the management and monitoring of C. eucalypti.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The genus Eucalyptus contains some of the most commonly cul-tivated hardwood timber species around the world (Santos et al.,2004). In addition to timber production, Eucalyptus trees are usedfor medicine, ornamental foliage, as landscape trees or wind breaks(Doughty, 2000). In the late 1990s, the commercial usage of Euca-lyptus spp. increased dramatically in range and distribution acrossEurope, North and South America (Hodkinson, 1999). However,over the past three decades Eucalyptus growing as exotics outsideAustralia have suffered increased biotic stress caused by a numberof Australian and endemic herbivorous insects (Gill et al., 1998;Paine et al., 2011). Among these pests is the small (1.5–2.0 mm)blue gum psyllid, Ctenarytaina eucalypti (Maskell) (Sternorrhyncha:Psyllidae), which feeds preferentially on waxy juvenile Eucalyptus
globulus foliage (Burckhardt et al., 1999; Purvis et al., 2002; Hollis,2004). Extralimitally, this psyllid has been reported attacking morethan 25 eucalypts, including Eucalyptus dunnii, Eucalyptus gunnii,Eucalyptus cordata (all subgenus Symphyomyrtus) and evenCorymbia citriodora (Burckhardt et al., 1999; Dahlsten et al.,1998a; Hodkinson, 1999; Purvis et al., 2002; Santana, 2005; San-tana and Burckhardt, 2007). C. eucalypti is native to Australia buthas been introduced into at least 18 countries across the world(Burckhardt, 1998; Burckhardt et al., 1999; Costanzi et al., 2003;Hodkinson, 2007; Olivares, 2000; Purvis et al., 2002; Taylor, 1997).
Female blue gum psyllids can lay 20–100 eggs, normally as agroup in the leaf buds and the axils of young leaves of young plants(Cadahia, 1980; Santana and Burckhardt, 2007). The psyllid canproduce up to four generations per year, depending on climateand plant suitability. The larval instars and the adults feed on plantsap. While feeding they excrete large amounts of honeydew andwaxy secretions that act as a growth medium for sooty molds(Dahlsten et al., 1998b; Santana and Burckhardt, 2007). Accordingto Santana and Burckhardt (2007), damage by C. eucalypti can beinduced directly by sucking the sap, or indirectly by injecting toxicsubstances from the saliva. The direct effects of feeding include

J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551 543
shoot dieback, leaf distortion and leaf discoloration (Elliott and deLittle, 1984; Hodkinson, 1999), reducing the quality of ornamentalfoliage and preventing export of foliage (Dahlsten et al., 1998b).
Different control strategies, such as biological control, culturalcontrol and breeding programs, have been attempted against thisspecies with variable success (Dahlsten et al., 1998a; Collett,2001; Paine et al., 2000). Olfactory-based control measures usingkairomones from host plants or pheromones have not been triedsince background knowledge has been missing. Recently, sex pher-omones were identified for the pear psyllid (Cacopsylla pyricola),the potato psyllid (Bactericera cockerelli), and the Asian citrus psyl-lid, Diaphorina citri (Guédot et al., 2009, 2010; Mann et al., 2012).However, no pheromones have yet been identified for any otherpsyllid. Plants release mixtures of volatile compounds that are pro-duced along different biosynthetic pathways (Pichersky and Ger-shenzon, 2002). These volatile blends contain species-specificinformation that can be used by herbivores to find hosts, oviposi-tion sites, and mates, or to avoid other individuals and food com-petition (Bruce et al., 2005a; Dudareva et al., 2006; Mustaparta,2002; Schoonhoven et al., 1998). Olfactory cues play an importantrole in host selection by psyllids. For instance, the carrot psyllid(Trioza apicalis) (Nehlin et al., 1996), the Asian citrus psyllid (Pattand Sétamou, 2010) and the pear psyllid (Horton et al., 2007,2008) have all shown attraction to either leaves or shoots of theirhost plants.
Volatiles are detected by insect olfactory sensory neurons(OSNs) on the antennae. The antenna of the blue gum psyllid is�0.56 mm long and consists of 10 segments (Hodkinson, 2007).Segments 4, 6, 8, and 9 each contain a sensillum cavity (rhinarium),here shown to house a sensillum with olfactory function similar tothe sensillum organization in T. apicalis (Kristoffersen et al., 2006).Additionally, two bristles are found at the tip of the antennae(Hodkinson, 2007), but it is unknown whether these are olfactoryor are tuned to a different sensory modality. In this study we usedsingle sensillum recordings (SSR) to investigate responses of C. euc-alypti OSNs to potential host-locating cues from Eucalyptus. Thefindings will be used to select compounds for behavioral experi-ments and field tests. Ultimately, we seek to explain psyllid hostspecificity, in particular the role of olfaction in acceptance of noveleucalypts outside Australia. Potentially the findings can be used todevelop olfaction-based pest monitoring and control strategies.
2. Materials and methods
2.1. Insects
Psyllids collected from E. globulus growing at Belfield (Dublin,Ireland) and a plantation of E. globulus near Clonbinane (Victoria,Australia) were shipped to Sweden and thereafter cultured in aclimate chamber. Live psyllids were sent from Melbourne underAustralian Government export permit (WT2012-764) by MNX-GlobalLifeSciences. Psyllids were reared on E. globulus seedlingsunder 20:4 light:dark cycle, 20:15 �C (light:dark) temperature cy-cle and 50–60% RH. Adults of both sexes were used for SSR. Sexeswere separated by the presence of proctiger (#) or terminalia ($).
2.2. Headspace volatile collection and chemical analyses
E. cordata seedlings were used in headspace volatile collection.Volatile collection was performed for 6 h between 12:00 and 18:00under 4 h light and 2 h dark at room temperature. The plants wereenclosed in a 25 � 30 cm plastic oven bag (Toppits, Cofresco, Ger-many) sealed with iron wire. A few slits (ca. 1 cm) were made inthe plastic bag for air circulation. The air around the plants waspumped at a rate of 100 ml/min through a Tenax� (GR 60/80) filter.
The collection was eluted using 250 ll redistilled hexane (purity>98%, LiChrosolv�) and condensed to 30 ll. The samples (2 ll in-jected volume) along with internal standard (methyl stearate10 ng/ll) were analyzed using a Hewlett Packard 6890 GC–MS(gas chromatography–mass spectrometry) equipped with a 7683injector, 5973 MSD detector and splitless injection system. TheGC had a 30 m Innowax capillary column (Agilent 19091N-033MS, 0.25 mm i.d., 0.25 lm film), using helium as carrier gas at6.45 psi (Agilent Technologies, Santa Clara, USA). The oven temper-ature was programmed as follows: 30 �C initial (held for 2 min)and then increased at a rate of 10 �C/min up to 220 �C (held for10 min). The inlet temperature was 225 �C and the interface tem-perature was 280 �C. Identification of compounds was done basedon comparison of mass spectra with those of ADAMS Library andsynthetic reference samples.
2.3. Single sensillum recordings
A live C. eucalypti was placed ventrally in a 100 ll disposableplastic micropipette tip, with its head and antennae protruding.The pipette tip was placed in dental wax on a glass slide and theantennae fixed using double sided sticky tape on a cover slip. Byusing thin copper wire, the position of the antennae was adjustedto a convenient angle. For closer view of the antennae and sensillarcavities, a microscope (Nikon eclipse E600FN: 750�magnification)was used during recordings.
Tungsten microelectrodes were sharpened using saturatedKNO2 solution and 20 V AC (Power Supply Unit MA 4852). The ref-erence electrode was inserted into the head of the insect, normallythrough the eye, and the recording electrode in one of the sensilla,aided with a Piezo micromanipulator (PM 10, Märzhäuser, Wetzlar-Steindorf, Germany). The recording electrode was connected touniversal AC/DC amplifier probe (gain 10�) that, in turn, was con-nected to an IDAC-4 interface board (all from Syntech, Kirchzarten,Germany). Autospike software v3.9 (Syntech) was used to recordthe response. Charcoal-purified and humidified airflow at 1.8 l/min (controlled by airflow meter, Porter Instruments, USA) wascontinuously blown over the antenna via silicone and glass tubing(5.9 mm i.d.). The outlet of the tube was placed approximately25 mm from the antenna.
The odorant test panel comprised 39 synthetic compoundsidentified in the present or previous studies from the headspaceof various C. eucalypti hosts (Table 1). In addition, a crushed E. cordataleaf was also tested to check whether Eucalyptus contains physio-logically active compounds that were lacking in our panel of syn-thetic test odorants. The compounds were diluted in paraffin oil(except for b-ocimene that was diluted in hexane) and applied(10 ll) on 2 � 1 cm filter paper strips (No. 3, Whatman, Maidstone,United Kingdom) inside glass Pasteur pipettes (150 mm soda limeglass, VWR International, Stockholm, Sweden). Response specific-ity of OSNs was characterized using a screening dose of 10 lg com-pound on the filter paper. Compounds were tested in randomorder. Dose–response tests were performed on additional sensilla,using doses from 1 ng to 10 lg (lowest dose tested first). In theserecordings, we focused on the compounds (i.e. linalool, b-caryo-phyllene, 1-hexanol, Z3-hexenol, Z3-hexenyl acetate, and E2-hexe-nal) that elicited the strongest OSN responses at the 10 lgscreening dose, since responses to weakly activating compoundsnormally are absent when the dose is lowered (e.g. Anderssonet al., 2009; Larsson et al., 2001). During stimulation, headspacefrom the stimulus pipettes was introduced into the continuous air-flow, and hence to the insect, at a rate of 0.2 l/min for 0.5 s (con-trolled by stimulus controller CS-02, Syntech). No pulsecompensatory flow was used during puffing. Since repeated puff-ing of stimulus pipettes significantly reduces the airborne stimulusconcentration (Andersson et al., 2012b), pipettes were reloaded

Table 1List of synthetic compounds used in single sensillum recordings with: classification, source, species from which they have been identified, and reference.
Chemicala Chemical source Purity(%)
Chemicalclass
Species Reference
Isoprene Sigma 99 HT E. dunnii, C. citriodora, E. globulus Street et al. (1997) and Zini et al. (2001, 2002)a-Pinene Fluka 99 MT E. dunnii, E. cordata, C. citriodora,
E. globulusThis study, Street et al. (1997), Zini et al. (2001, 2002) and Barataet al. (2000, 2002)
(�)-b-Pinene Fluka >99 MT E. gunnii, E. cordata, C. citriodora,E. globulus
This study, Wirthensohn et al. (2000), Zini et al. (2001) and Barataet al. (2000, 2002)
a-Phellandrene Aldrich >95 MT C. citriodora, E. cordata, E.globulus
This study, Zini et al. (2002) and Barata et al. (2000)
(Z/E)-b-Ocimeneb G. Bergströme 93 MT E. dunnii, C. citriodora, E. globulus Zini et al. (2001, 2002), Barata et al. (2002) and Lopes et al. (2002)c-Terpinene G. Bergströme 95 MT E. dunnii, E. cordata, E. globulus This study, Zini et al. (2002), Yassaa et al. (2000) and Barata et al.
(2000)Camphene Aldrich 95 MT E. saligna, E. globulus Zini et al. (2002), Yassaa et al. (2000) and Barata et al. (2000)(+)-Fenchol G. Bergströme >99 MT–OH E. globulus Yassaa et al. (2000)Limonene Fluka 99 MT E. dunnii, E. cordata, C. citriodora,
E. globulusThis study, Street et al. (1997), Zini et al. (2002) and Barata et al.(2000, 2002)
(Z/E)-Linalool oxidec Dragoco 92 MT E. globulus Barata et al. (2000)Myrcene Fluka 95 MT–OH C. citriodora, E. globulus Street et al. (1997), Zini et al. (2001), Barata et al. (2000, 2002) and
Lopes et al. (2002)Tricyclene Aldrich 99 MT E. globulus Yassaa et al. (2000)D-3-Carene Aldrich 95 MT E. globulus Barata et al. (2000)D-2-Carene G. Bergströme >99 MT E. cordata -P-Cymene Acros >99 MT E. cordata, E. globulus This study, Barata et al. (2000)Sabinene Aldrich 99 MT C. citriodora, E. globulus Zini et al. (2001) and Barata et al. (2000)Terpinolene Fluka 97 MT E. cordata, E. globulus This study, Barata et al. (2000)a-Terpineol Aldrich 90 MT–OH E. globulus Street et al. (1997) and Barata et al. (2000)Terpinen-4-ol Fluka >99 MT–OH E. cordata This studyIsopulegol Sigma >99 MT–OH C. citriodora Zini et al. (2001)Linalool Fluka 97 MT–OH C. citriodora, E. globulus Yassaa et al. (2000), Barata et al. (2000, 2002) and Zini et al.
(2002)Citronellol Sigma 95 MT–OH C. citriodora Zini et al. (2001, 2002)(Z/E)-Thujoned G. Bergströme 96 MT@O E. globulus Barata et al., 2000Piperitone G. Bergströme >99 MT@O E. globulus Barata et al. (2000)1,8-Cineole Aldrich >99 MT–acetal E. dunnii, E. cordata, C. citriodora,
E. globulusThis study, Street et al. (1997), Zini et al. (2001, 2002) and Barataet al. (2000, 2002)
a-Cubebene G. Bergströme 88 ST E. globulus Barata et al. (2000)b-Caryophyllene Fluka 98 ST C. citriodora, E. globulus Zini et al. (2001, 2002)3-Methyl-1-butanol Aldrich 99 C5–OH E. globulus Barata et al. (2000, 2002) and Lopes et al. (2002)1-Hexanol Fluka 99 C6–OH E. globulus Barata et al. (2000)Z2-Hexenol Aldrich 95 C6–OH E. globulus Barata et al. (2000, 2002) and Lopes et al. (2002)E2-Hexenol Acros 96 C6–OH E. globulus Yassaa et al. (2000)Z3-Hexenol Acros 98 C6–OH E. cordata, E. globulus This study, Barata et al. (2000, 2002) and Lopes et al. (2002)Z3-Hexenyl acetate Sigma 99 C6–OAc E. cordata, E. globulus This study, Barata et al. (2000)Ethyl-3-
methylbutanoateAldrich 98 C7–OAc E. globulus Barata et al. (2002)
Citronellyl acetate G. Bergströme 96 C12–OAc C. citriodora Zini et al. (2001)3-Hydroxy-2-butanone Aldrich P97 C4@O E. globulus Barata et al. (2000, 2002) and Lopes et al. (2002)E2-Hexenal Aldrich 98 C6@O E. globulus Barata et al. (2000)6-Methyl-5-hepten-2-
oneSigma 99 C8@O E. cordata This study
(Z) 3,7-Dimethyl-2,6-octadienal
Aldrich 95 C10@O C. citriodora Zini et al. (2001, 2002)
Paraffin oil Merck – – – –
Note: HT: hemiterpene (5C), MT: monoterpene (10C), ST: sesquiterpene (15C),–OH: alcohol, @O: aldehyde and ketone, –OAc: acetate ester.a Chiral compounds were tested as racemic mixtures unless otherwise indicated.b Mixture of Z:E isomers-(30:70%).c Mixture of Z:E isomers-(50:50%).d Mixture of Z:E isomers-(83:17%).e Gift from Prof. Gunnar Bergström.
544 J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551
after a maximum of 10 puffs during screening or after two puffs indose–response tests.
The OSNs in one of the sensilla responded the strongest to twocompounds (b-caryophyllene and linalool activating two differentOSNs) with vastly different evaporation rates from paraffin oil(ca. 30 times lower evaporation of b-caryophyllene; E. Hatano, per-sonal communication). Thus, to provide more accurate sensitivityestimates for these two neurons, GC-coupled SSR was performedby injecting 10 ng of each compound diluted in hexane (for detailssee Kristoffersen et al., 2008). GC–SSR was not used for the otherstrongly activating synthetic compounds, 1-hexanol, Z3-hexenol,and Z3-hexenyl acetate, since the evaporation rate of different
green leaf volatiles (GLVs) from paraffin oil is similar (Anderssonet al., 2012b).
2.4. Data analysis
OSN responses were analyzed offline using Autospike v3.9 (Syn-tech). Net odor responses were calculated by counting the numberof spikes during the first 0.5 s of the response and subtracting thenumber of spikes 0.5 s before the response. The value was doubledto attain a response in spikes/s (Hz). Potential blank responseswere subtracted from all the odor responses in the correspondingOSN. The responses (net�blank) during screening were categorized

J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551 545
based on number of spikes/s, where (0) <10 Hz, (+) = 10–29 Hz,(++) = 30–59 Hz, (+++) = 60–89 Hz, (++++) = 90–119 Hz,(+++++) P 120 Hz. We also analyzed temporal odor response pro-files for strongly responding OSN types. This was done by countingspikes in 200 ms bins, starting at the onset of stimulation and end-ing 8 s after the onset. The average spontaneous activity during 2 spreceding the stimulation was subtracted from these responses toobtain the net responses. GC–SSR responses were quantified bycounting the number of spikes during a 1 s period around the cen-ter of the compound peaks. Net responses were obtained by sub-tracting the spontaneous activity during the 1 s period justbefore the compound peaks.
3. Results
3.1. Headspace volatiles
Twelve volatile compounds were identified from the headspacecollection of E. cordata. Additional compounds were present, butcould not be identified (Supplementary material Fig. S1 andTable S1). The main compound that gives the characteristic odorof Eucalyptus, 1,8-cineole (eucalyptol), was present in the highestquantity followed by a-phellandrene and limonene. a-Pinene andc-terpinene, as well as P-cymene were present in relatively highamounts. Some compounds that existed in low quantities were6-methyl-5-hepten-2-one, the green leaf volatiles Z3-hexenol andZ3-hexenyl acetate, as well as the terpenes, terpinen-4-ol,b-pinene, and terpinolene.
3.2. Antennal morphology and general response characteristics
The antenna of the blue gum psyllid is filiform and contains foursensillar cavities, one on each of segment 4, 6, 8 and 9 (Fig. 1). Welabeled the sensilla in the cavities: S1 (proximal), S2, S3 and S4(distal) according to their location on segments 4–9. Recordingswere obtained from all sensilla. In total, 42 adults (22 males, 20 fe-males) were tested for responses to the synthetic compounds (seeTable 2 and Fig. 3 for n-values for each sensillum in the screeningand dose–response tests, respectively). In each sensillum, threeOSNs were distinguished based on spike amplitudes (labeled A,B, and C neuron with large, intermediate, and small amplitude,respectively; Fig. 2A). However, the C neurons in S1–S4 never re-sponded to any of the tested host volatiles. Inhibitory responses(decrease in spike activity) were elicited only by Z2-hexenol inS2, S3 and S4 (Fig. 2B).
Thirty-three of the 39 tested compounds elicited responses in atleast one OSN type (Table 2). The remaining six compounds(linalool oxide, 3-carene, 2-carene, P-cymene, sabinene, and piperi-tone) were sedentary (Table 2). 1,8-Cineole elicited only minor re-sponses although this volatile dominates the headspace ofEucalyptus and is responsible for the characteristic (at least for hu-mans) Eucalyptus smell. Response to blank stimuli (paraffin oil)was always below 10 Hz.
Males and females appeared to have the same or at least verysimilar classes of OSNs on their antennae. However, the strengthof the responses and the presence of weak (+) responses variedsomewhat between sexes for some of the compounds, especiallyin S2 and S3 (Table 2). The strongest OSN responses (in S1–S3) ran-
Sc PS1 S2 S3 S4
Fig. 1. A schematic ventral view of a Ctenarytaina eucalypti left antenna. S1–S4indicates the olfactory sensilla located on segments 4, 6, 8 and 9. Sc = scape andP = pedicel.
ged between 120 and 160 Hz, and occasionally neurons fired at>190 Hz. These strong responses suggest that the key ligands forthe receptors likely were included in the test panel.
3.3. OSNs in S1 responding primarily to b-caryophyllene and linalool
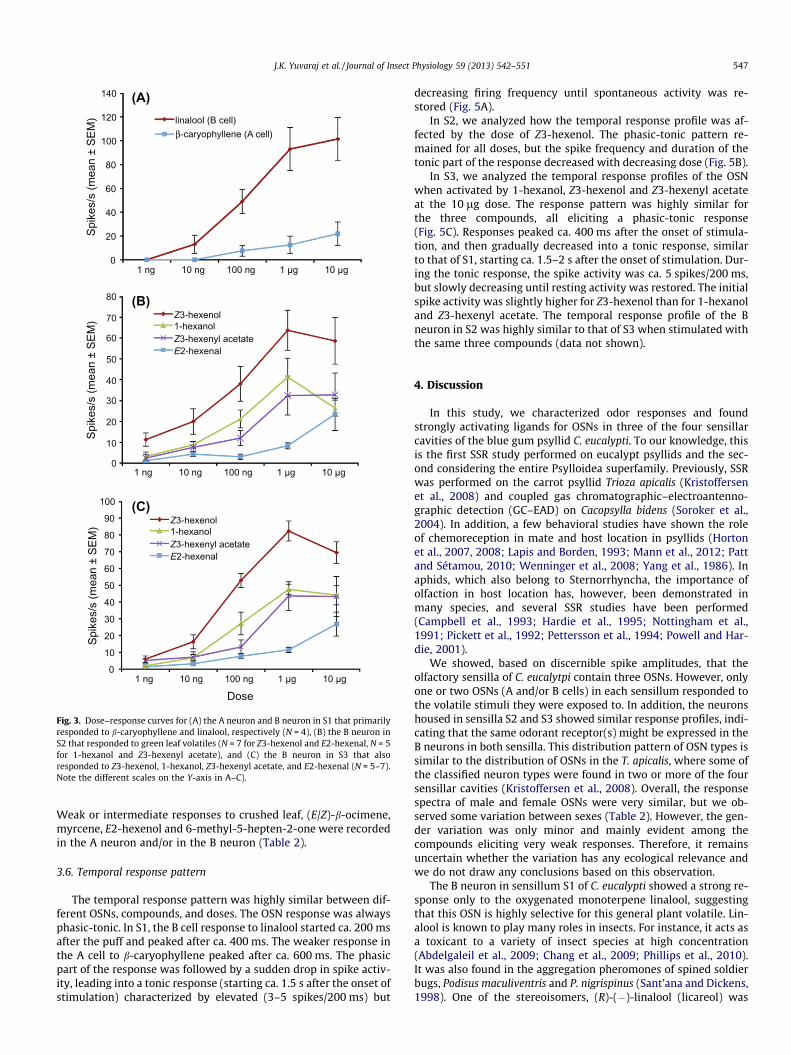
The strongest responses in the study were recorded from the Bneuron in S1 that was highly specific for linalool (Table 2). Some-times the spike activity was elevated for more than a minute afterthe onset of stimulation. In addition, the A neuron in S1 showed anintermediate (++) response to b-caryophyllene (total average formales and females combined: 47 Hz) and a weaker response to(E/Z)-b-ocimene (total average 27 Hz, weaker in males than in fe-males). This OSN also showed a weak (+, male) or intermediate(++, female) response to the crushed E. cordata leaf (Table 2). Bothneurons were subjected to dose–response trials that indicated a re-sponse threshold for linalool below the 10 ng dose on the filter pa-per, whereas the A neuron was 10–100 times less sensitive tob-caryophyllene (Fig. 3A). However, since the evaporation rate ofb-caryophyllene (sesquiterpene) from paraffin oil is ca. 30 timeslower than that of linalool (monoterpene alcohol) (E. Hatano, per-sonal communication), we also delivered the two compounds tothe antenna by means of a GC (10 ng of each compound injected).The GC–SSR results indicated that when the two compounds aredelivered at the same (presumably) airborne concentration, the re-sponses in the two OSNs are more similar. However, the responsein the B cell to linalool (73 spikes/s) was still stronger than theresponse in the A cell to b-caryophyllene (45 spikes/s) (Fig. 4; seeTable 2 for comparison). b-Ocimene, the second most active com-pound in the A cell, was not tested through the GC since a previousstudy showed that the evaporation rates of other monoterpenehydrocarbons were similar to that of linalool (Andersson et al.,2012b). During the B cell response to linalool, the spontaneousactivity of the A cell was simultaneously inhibited.
3.4. OSNs in S2 and S3 responding primarily to green leaf volatiles
The responses of both the A and B cell were similar in S2 and S3(Table 2), especially in the B cell to the three most potent ligands.The B neurons in both sensilla responded most strongly to the GLVs1-hexanol, Z3-hexenol and Z3-hexenyl acetate. The response to theindividual GLVs in the B neuron varied from intermediate to verystrong between contacted sensilla. In the S3 sensillum of threeindividuals there was no response at all to any of the GLVs. Sincethe OSNs within these sensilla might have been damaged orresponsive only to compounds not present in the odor panel, theywere not included in the calculation of average responses (Table 2).In both S2 and S3, some compounds (e.g. 3-methyl-1-butanol, Z2-hexenol, some ketones and acetate esters) that were inactive inmales elicited weak (+) responses in females. The dose–responsecurves of the B neurons in S2 and S3 were highly similar and indi-cated that the response thresholds for Z3-hexenol, 1-hexanol andZ3-hexenyl acetate were near or below the 1 ng dose (Fig. 3B–C).Responses were slightly stronger to Z3-hexenol at all tested doses.The OSNs were less sensitive to E2-hexenal, with a responsethreshold between the 1 and 10 ng dose.
The A neurons in S2 and S3 displayed weaker responses to (E/Z)-b-ocimene and E2-hexenal (for details see Table 2). The other com-pounds in the test panel elicited only very weak responses in thesesensilla. The crushed E. cordata leaf evoked stronger responses inthe B neurons than in the A neurons.
3.5. OSNs in sensillum S4
In contrast to the other sensilla, OSNs within S4 did not respondstrongly to any of the synthetic compounds or to the crushed leaf.

B C
A
(A)
(B)
Fig. 2. Single sensillum recordings from Ctenarytaina eucalypti. (A) Three OSNs (A, B, C) could be distinguished based on differences in the size of the spike amplitude. (B) Aweak excitation followed by inhibition (occasionally seen) in the B neuron of S2 and S3 in response to Z2-hexenol (10 lg).
Table 2Response spectra of OSNs from Ctenarytaina eucalypti to synthetic compounds (10 lg).
Compounds S1 S2 S3 S4
# $ # $ # $ # $
A B A B A B A B A B A B A B A B
E. cordata leaf + 0 ++ 0 ++ +++ + +++ + ++ + ++ ++ + 0 ++Isoprene 0 0 0 0 0 0 0 + 0 0 0 + 0 0 0 0a-Pinene 0 0 0 0 + 0 0 + 0 0 0 + 0 0 0 0(�)-b-Pinene 0 0 0 0 + 0 0 + 0 0 0 + 0 0 0 0a-Phellandrene + 0 0 0 0 0 0 + 0 0 0 + 0 0 0 0(E/Z)-b-Ocimene + 0 ++ 0 ++ + 0 + + 0 0 + + + 0 +c-Terpinene 0 0 0 0 0 0 0 0 0 0 0 + 0 0 0 0Camphene 0 0 0 0 0 0 0 + 0 0 0 + 0 0 0 0(+)-Fenchol + 0 + 0 0 0 0 + 0 0 0 + 0 0 0 0Limonene 0 0 0 0 + + 0 + 0 0 0 + 0 0 0 0(Z/E)-Linalool oxidea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myrcene + 0 + 0 + 0 0 0 0 0 0 + + ++ 0 +Tricyclene 0 0 + 0 0 0 0 + 0 0 0 + 0 0 0 0D-3-Carenea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0D-2-Carenea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0P-Cymenea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Sabinenea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Terpinolenea 0 0 0 0 + 0 0 + 0 0 0 + 0 0 0 0a-Terpineol 0 0 0 + 0 0 0 + 0 0 0 + 0 0 0 0Terpinen-4-ola 0 0 0 + 0 0 0 0 0 0 0 0 0 0 0 0Isopulegol 0 0 + 0 0 0 0 + 0 0 0 + 0 0 0 0Linalool 0 +++++ 0 +++++ + + 0 + 0 0 0 + 0 0 0 0Citronellol 0 0 0 0 0 0 0 + 0 0 0 0 0 0 0 0Thujone 0 0 + 0 0 0 0 + 0 0 0 + + 0 0 0Piperitonea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 01,8-Cineole 0 0 + 0 + + 0 + 0 0 0 + 0 0 0 0a-Cubebenea 0 0 0 + 0 0 0 0 0 0 0 0 0 0 0 0b-Caryophyllene ++ 0 ++ 0 + 0 0 + 0 0 0 + 0 0 0 03-Methyl-1-butanol 0 0 0 0 + 0 0 0 0 0 0 0 0 0 0 01-Hexanola 0 0 0 0 0 +++ 0 +++ 0 +++ 0 +++ 0 0 0 0Z2-Hexenol 0 0 0 0 0 0 0 + 0 0 0 0 0 0 0 0E2-Hexenol 0 0 0 0 + + 0 + 0 0 0 0 0 + 0 0Z3-Hexenolb 0 0 0 0 + ++++ 0 ++ + +++ 0 ++ 0 0 0 0Z3-Hexenyl acetatea 0 0 0 0 0 +++ 0 +++ 0 ++++ 0 +++ 0 0 0 0Ethyl-3-methylbutanoate 0 0 0 0 0 0 0 + 0 0 0 + 0 0 0 0Citronellyl acetate 0 0 0 0 0 0 0 + 0 0 0 + 0 0 0 03-Hydroxy-2-butanone 0 0 0 0 0 0 0 + 0 0 0 + 0 0 0 0E2-Hexenal 0 0 0 + ++ + 0 ++ + 0 0 ++ 0 0 0 06-Methyl-5-hepten-2-onea 0 0 0 0 0 0 0 0 0 0 0 0 0 ++ 0 +(Z) 3,7-Dimethyl-2,6-octadienal 0 0 0 0 0 0 0 + 0 0 0 + + + 0 +Paraffin oil 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Number of OSNs (N-value) 7 7 8 6 6 4 6 4
OSN responses: (0) <10 Hz, (+) 10–29 Hz, (++) 30–59 Hz, (+++) 60–89 Hz, (++++) 90–119 Hz, (+++++) P120 Hz.a For these compounds, N = 3 for male S1–S4, and N = 3, 6, 2, and 5 for female S1, S2, S3, and S4 respectively.b N = 9–11 for Z3-hexenol on S2 (both sexes) and male S3, and N = 5 for female S3. N-values for all other OSN-compound combinations are as stated in the table (bottom
row).
546 J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551
0
10
20
30
40
50
60
70
80
90
100
Spik
es/s
(mea
n ±
SEM
)
Dose
Z3-hexenol
E2-hexenal
1-hexanol Z3-hexenyl acetate
0
20
40
60
80
100
120
140
linalool (B cell) β-caryophyllene (A cell)
1 ng 10 ng 100 ng 1 µg 10 µg
1 ng 10 ng 100 ng 1 µg 10 µg
1 ng 10 ng 100 ng 1 µg 10 µg
Spik
es/s
(mea
n ±
SEM
)Sp
ikes
/s (m
ean
± SE
M)
(A)
(B)
(C)
Z3-hexenol
E2-hexenal
1-hexanol Z3-hexenyl acetate
0
10
20
30
40
50
60
70
80
Fig. 3. Dose–response curves for (A) the A neuron and B neuron in S1 that primarilyresponded to b-caryophyllene and linalool, respectively (N = 4), (B) the B neuron inS2 that responded to green leaf volatiles (N = 7 for Z3-hexenol and E2-hexenal, N = 5for 1-hexanol and Z3-hexenyl acetate), and (C) the B neuron in S3 that alsoresponded to Z3-hexenol, 1-hexanol, Z3-hexenyl acetate, and E2-hexenal (N = 5–7).Note the different scales on the Y-axis in A–C).
J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551 547
Weak or intermediate responses to crushed leaf, (E/Z)-b-ocimene,myrcene, E2-hexenol and 6-methyl-5-hepten-2-one were recordedin the A neuron and/or in the B neuron (Table 2).
3.6. Temporal response pattern
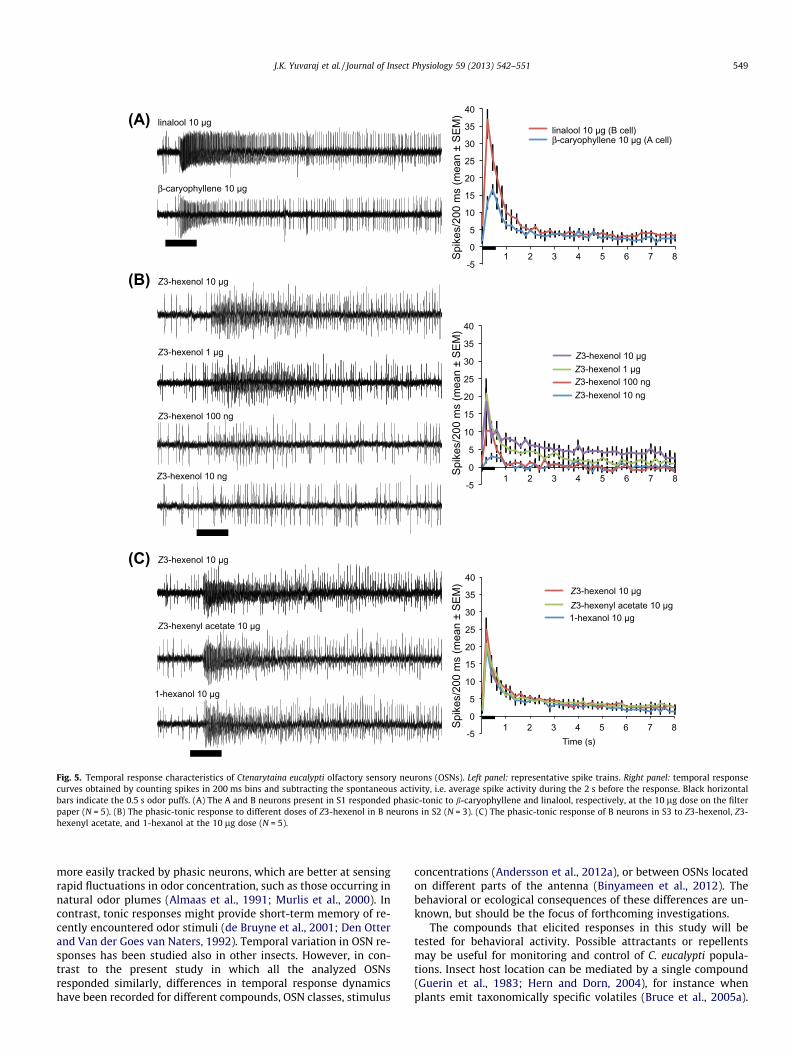
The temporal response pattern was highly similar between dif-ferent OSNs, compounds, and doses. The OSN response was alwaysphasic-tonic. In S1, the B cell response to linalool started ca. 200 msafter the puff and peaked after ca. 400 ms. The weaker response inthe A cell to b-caryophyllene peaked after ca. 600 ms. The phasicpart of the response was followed by a sudden drop in spike activ-ity, leading into a tonic response (starting ca. 1.5 s after the onset ofstimulation) characterized by elevated (3–5 spikes/200 ms) but
decreasing firing frequency until spontaneous activity was re-stored (Fig. 5A).
In S2, we analyzed how the temporal response profile was af-fected by the dose of Z3-hexenol. The phasic-tonic pattern re-mained for all doses, but the spike frequency and duration of thetonic part of the response decreased with decreasing dose (Fig. 5B).
In S3, we analyzed the temporal response profiles of the OSNwhen activated by 1-hexanol, Z3-hexenol and Z3-hexenyl acetateat the 10 lg dose. The response pattern was highly similar forthe three compounds, all eliciting a phasic-tonic response(Fig. 5C). Responses peaked ca. 400 ms after the onset of stimula-tion, and then gradually decreased into a tonic response, similarto that of S1, starting ca. 1.5–2 s after the onset of stimulation. Dur-ing the tonic response, the spike activity was ca. 5 spikes/200 ms,but slowly decreasing until resting activity was restored. The initialspike activity was slightly higher for Z3-hexenol than for 1-hexanoland Z3-hexenyl acetate. The temporal response profile of the Bneuron in S2 was highly similar to that of S3 when stimulated withthe same three compounds (data not shown).
4. Discussion
In this study, we characterized odor responses and foundstrongly activating ligands for OSNs in three of the four sensillarcavities of the blue gum psyllid C. eucalypti. To our knowledge, thisis the first SSR study performed on eucalypt psyllids and the sec-ond considering the entire Psylloidea superfamily. Previously, SSRwas performed on the carrot psyllid Trioza apicalis (Kristoffersenet al., 2008) and coupled gas chromatographic–electroantenno-graphic detection (GC–EAD) on Cacopsylla bidens (Soroker et al.,2004). In addition, a few behavioral studies have shown the roleof chemoreception in mate and host location in psyllids (Hortonet al., 2007, 2008; Lapis and Borden, 1993; Mann et al., 2012; Pattand Sétamou, 2010; Wenninger et al., 2008; Yang et al., 1986). Inaphids, which also belong to Sternorrhyncha, the importance ofolfaction in host location has, however, been demonstrated inmany species, and several SSR studies have been performed(Campbell et al., 1993; Hardie et al., 1995; Nottingham et al.,1991; Pickett et al., 1992; Pettersson et al., 1994; Powell and Har-die, 2001).
We showed, based on discernible spike amplitudes, that theolfactory sensilla of C. eucalytpi contain three OSNs. However, onlyone or two OSNs (A and/or B cells) in each sensillum responded tothe volatile stimuli they were exposed to. In addition, the neuronshoused in sensilla S2 and S3 showed similar response profiles, indi-cating that the same odorant receptor(s) might be expressed in theB neurons in both sensilla. This distribution pattern of OSN types issimilar to the distribution of OSNs in the T. apicalis, where some ofthe classified neuron types were found in two or more of the foursensillar cavities (Kristoffersen et al., 2008). Overall, the responsespectra of male and female OSNs were very similar, but we ob-served some variation between sexes (Table 2). However, the gen-der variation was only minor and mainly evident among thecompounds eliciting very weak responses. Therefore, it remainsuncertain whether the variation has any ecological relevance andwe do not draw any conclusions based on this observation.
The B neuron in sensillum S1 of C. eucalypti showed a strong re-sponse only to the oxygenated monoterpene linalool, suggestingthat this OSN is highly selective for this general plant volatile. Lin-alool is known to play many roles in insects. For instance, it acts asa toxicant to a variety of insect species at high concentration(Abdelgaleil et al., 2009; Chang et al., 2009; Phillips et al., 2010).It was also found in the aggregation pheromones of spined soldierbugs, Podisus maculiventris and P. nigrispinus (Sant’ana and Dickens,1998). One of the stereoisomers, (R)-(�)-linalool (licareol) was

10.85 10.90
1603312454
04.0103.0152.01
linaloolβ-caryophyllene
GCmin
SSR
10.35 10.95 11.010.20
s/sekips 54s/sekips 37
RT
Fig. 4. GC–SSR traces showing responses to b-caryophyllene (right, A neuron, 45 spikes/s) and linalool (left, B neuron, 75 spikes/s) in the olfactory sensory neurons (OSNs)inside S1. 10 ng of each compound was injected into the GC. Net OSN responses were calculated during 1 s (as indicated in graph) after subtracting spontaneous activitybefore the elution of the compounds. Numbers under GC peaks represent GC area units. RT = retention time. Note the inhibition of the A cell during the excitatory response tolinalool in the B cell.
548 J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551
reported as a short-range kairomone and synergist in both Tetropi-um fuscum and Anoplophora glabripennis (long horned beetles:Coleoptera) (Silk et al., 2010) whereas the other stereoisomer (S)-(+)-linalool (coriandrol) was reported as an attractive pheromonecomponent in Colletes cunicularius (a solitary bee: Hymenoptera)(Borg-Karlson et al., 2003). Conversely, linalool interferes with hostfinding by repelling economically important aphids such as Aphisfabae (black bean aphid), Cavariella aegopodii (carrot aphid), Myzuspersicae (green peach aphid) and Acyrthosiphon pisum (pea aphid)(Bruce et al., 2005b; Chapman et al., 1981), which suggests thepresence of linalool-detecting OSNs in these species too. The Asiancitrus psyllid and the carrot psyllid responded behaviorally andphysiologically, respectively, to linalool as well as to other mono-and sesquiterpenes, such as (E)-b-ocimene, b-caryophyllene,terpinolene and terpinen-4-ol (Kristoffersen et al., 2008; Patt andSétamou, 2010). Since OSNs of C. eucalypti also responded to linal-ool, (E)-b-ocimene and b-caryophyllene, it is possible that terpene-detecting OSNs are a general characteristic of psyllid olfaction.Furthermore, our SSR and GC–SSR from the OSNs inside S1 demon-strate that differences in compound release rates, in this case linal-ool and b-caryophyllene, from Pasteur pipette stimulus cartridgescan have large effects on the strength of the OSN responses. Thiswas previously demonstrated in various other insect species(Andersson et al., 2012b). Furthermore, the GC–SSR also revealedthat the A neuron in S1 was inhibited while the B neuron simulta-neously responded to linalool, suggesting that lateral interactionsoccur between OSNs in the periphery, as previously found in otherspecies (Andersson et al., 2010; Su et al., 2012).
In this study, C. eucalypti showed strong responses to GLV alco-hols and acetates (1-hexanol, Z3-hexenol and Z3-hexenyl acetate)(Table 2). GLVs are produced in large amounts by angiosperm trees(Zhang and Schlyter, 2004), and most studied insect species haveone or several classes of OSNs that are tuned to these compounds(e.g. Andersson et al., 2009; Bengtsson et al., 2009; Binyameenet al., 2012), sometimes very specifically (Andersson et al.,2012a; Hansson et al., 1999; Larsson et al., 2001). GLV compoundssuch as E2-hexenal, 1-hexanol, Z3-hexenol and Z3-hexenyl acetateare electrophysiologically active in many aphid species (Hardieet al., 1995; Visser and Piron, 1995; Visser et al., 1996). GLVs canact as attractants to some insect species (Visser and Avé, 1978),and as repellents to others, indicating the presence of unsuitable
hosts (Unsicker et al., 2009; Zhang and Schlyter, 2004). It remainsto be determined whether GLVs may attract or repel C. eucalypti.The weak response of the OSNs in S4 and the absence of responsesin all C neurons suggest that these OSNs may detect plant com-pounds not tested in this study or possible pheromone compounds,if they exist for C. eucalypti.
Insect host location often relies on a tiny proportion of the com-pounds present in the host plant headspace. Thus, the headspacecontains a surplus of compounds that are not used in host location(Birkett et al., 2004; Nojima et al., 2003; Tasin et al., 2007). Thiswas clearly demonstrated in the present study, in which C. eucalyp-ti was non-responsive or responded only weakly to many of thecompounds in the odor panel in spite of the high screening dose(10 lg). In addition, the neurons in S1–S3 were highly confinedin their response specificity, responding strongly to only one (S1)or three (S2 and S3) compounds. High specificity of plant odor-responding OSNs (see Mustaparta, 2002 for a discussion about spe-cific and broadly tuned OSNs) is commonly found in insects and isconsidered integral to their capacity to identify host from non-hostplant species (Andersson et al., 2009, 2012a; Hansson et al., 1999;Stensmyr et al., 2001; Wibe and Mustaparta, 1996).
When comparing the organization of sensilla in C. eucalytpi tothat of T. apicalis (Kristoffersen et al., 2008), there is no differencein the number of sensilla and in their position on the differentantennal filaments. However, the OSN response profiles varied be-tween the two species. Trioza apicalis responded to terpinene-4-olin sensilla S1 and S4, and to terpinolene in S1 (Kristoffersen et al.,2008). Ctenarytaina eucalytpi did not respond strongly to either ter-pinene-4-ol or terpinolene. On the other hand, OSNs in sensilla S2and S3 of both species responded to chemically related GLVs, i.e. T.apicalis responded to Z3-hexenal and C. eucalytpi responded pri-marily to the corresponding alcohol, Z3-hexenol. Thus, both psyl-lids have GLV-detecting OSNs in sensilla S2 and S3.Unfortunately, only Z3-hexenal was tested for T. apicalis, so wedo not know whether the GLVs that elicited responses in C. euca-lypti also are active on T. apicalis.
The OSNs in S1–S3 showed highly similar phasic-tonic responsedynamics to different compounds and different concentrations(Fig. 4). The importance of the temporal structure of OSN responsesis poorly understood, but might be associated with the ability of aninsect to follow pulsed stimuli. Pulsed stimuli are thought to be
Z3-hexenol 10 µg
Z3-hexenol 1 µg
Z3-hexenol 100 ng
Z3-hexenol 10 ng
Z3-hexenol 10 µg
1-hexanol 10 µg
Z3-hexenyl acetate 10 µg
(A)
(B)
(C)
linalool 10 µg
β-caryophyllene 10 µg
Spik
es/2
00 m
s (m
ean
± SE
M)
-5
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8
-5
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8 Time (s)
-5
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8
Z3-hexenol 10 µg
Z3-hexenol 100 ng Z3-hexenol 1 µg
Z3-hexenol 10 ng
linalool 10 µg (B cell)β-caryophyllene 10 µg (A cell)
1-hexanol 10 µg
Z3-hexenol 10 µg Z3-hexenyl acetate 10 µg
Spik
es/2
00 m
s (m
ean
± SE
M)
Spik
es/2
00 m
s (m
ean
± SE
M)
Fig. 5. Temporal response characteristics of Ctenarytaina eucalypti olfactory sensory neurons (OSNs). Left panel: representative spike trains. Right panel: temporal responsecurves obtained by counting spikes in 200 ms bins and subtracting the spontaneous activity, i.e. average spike activity during the 2 s before the response. Black horizontalbars indicate the 0.5 s odor puffs. (A) The A and B neurons present in S1 responded phasic-tonic to b-caryophyllene and linalool, respectively, at the 10 lg dose on the filterpaper (N = 5). (B) The phasic-tonic response to different doses of Z3-hexenol in B neurons in S2 (N = 3). (C) The phasic-tonic response of B neurons in S3 to Z3-hexenol, Z3-hexenyl acetate, and 1-hexanol at the 10 lg dose (N = 5).
J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551 549
more easily tracked by phasic neurons, which are better at sensingrapid fluctuations in odor concentration, such as those occurring innatural odor plumes (Almaas et al., 1991; Murlis et al., 2000). Incontrast, tonic responses might provide short-term memory of re-cently encountered odor stimuli (de Bruyne et al., 2001; Den Otterand Van der Goes van Naters, 1992). Temporal variation in OSN re-sponses has been studied also in other insects. However, in con-trast to the present study in which all the analyzed OSNsresponded similarly, differences in temporal response dynamicshave been recorded for different compounds, OSN classes, stimulus
concentrations (Andersson et al., 2012a), or between OSNs locatedon different parts of the antenna (Binyameen et al., 2012). Thebehavioral or ecological consequences of these differences are un-known, but should be the focus of forthcoming investigations.
The compounds that elicited responses in this study will betested for behavioral activity. Possible attractants or repellentsmay be useful for monitoring and control of C. eucalypti popula-tions. Insect host location can be mediated by a single compound(Guerin et al., 1983; Hern and Dorn, 2004), for instance whenplants emit taxonomically specific volatiles (Bruce et al., 2005a).

550 J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551
However, in this study we found strong responses to commonplant volatiles, and no response to 1,8-cineole (eucalyptol) thatdominates the eucalypt headspace. Moreover, since many eucalyp-ts, especially those within the same subgenus, share so many ter-pene compounds (discussed in Paine et al., 2011), host locationby C. eucalypti may be influenced by specific volatile mixturesand/or by other cues (e.g. visual stimuli) that characterize the pre-ferred species and leaf type (juvenile or adult). The relative influ-ences of these different cues and mechanisms are the focus ofon-going behavioral studies.
In conclusion, the current study characterized the specificity ofplant odor-detecting OSNs in the blue gum psyllid. Even thoughmore studies need to be conducted to identify the functions ofthe active compounds identified here, the knowledge gained fromthis study contributes to our understanding of the chemical ecol-ogy of psyllids.
Acknowledgements
M.J.S. thanks Bronwyn Meredith (Wildlife Trade Assessments,Department of Sustainability, Environment, Water, Populationand Communities, Canberra) for assistance obtaining insect exportpermits and the Australian Research Council for Future Fellowshipfunding (FT100100199). K.F. was funded by an Australian Post-graduate Award from the Australian Government with top-upfunding provided by La Trobe University. The authors would liketo thank Jan-Robert Baars (School of Biology & Environment Sci-ence, University College Dublin, Ireland) for also collecting andshipping psyllids for this study.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.jinsphys.2013.03.004.
References
Abdelgaleil, S.A.M., Mohamed, M.I.E., Badawy, M.E.I., El-arami, S.A.A., 2009.Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) andTribolium castaneum (Herbst) and their inhibitory effects onacetylcholinesterase activity. Journal of Chemical Ecology 35, 518–525.
Almaas, T.J., Christensen, T.A., Mustaparta, H., 1991. Encoding of different featuresof an olfactory stimulus by the sex pheromone receptors in Heliothis zea. Journalof Comparative Physiology A 169, 249–258.
Andersson, M.N., Larsson, M.C., Schlyter, F., 2009. Specificity and redundancy in theolfactory system of the bark beetle Ips typographus: single-cell responses toecologically relevant odors. Journal of Insect Physiology 55, 556–567.
Andersson, M.N., Larsson, M.C., Blazenec, M., Jakuš, R., Zhang, Q.-H., Schlyter, F.,2010. Peripheral modulation of pheromone response by inhibitory hostcompound in a beetle. Journal of Experimental Biology 213, 3332–3339.
Andersson, M.N., Larsson, M.C., Svensson, G.P., Birgersson, G., Rundlöf, M., Lundin,O., Lankinen, Å., Anderbrant, O., 2012a. Characterization of olfactory sensoryneurons in the white clover seed weevil, Apion fulvipes (Coleoptera: Apionidae).Journal of Insect Physiology 58, 1325–1333.
Andersson, M.N., Schlyter, F., Hill, S.R., Dekker, T., 2012b. What reaches theantenna? How to calibrate odor flux and ligand–receptor affinities. ChemicalSenses 37, 403–420.
Barata, E.N., Pickett, J.A., Wadhams, J., Woodcock, C.M., Mustaparta, H., 2000.Identification of host and nonhost semiochemicals of eucalyptus woodborerPhoracantha semipunctata by gas chromatography–electroantennography.Journal of Chemical Ecology 26, 1877–1895.
Barata, E.N., Mustaparta, H., Pickett, J.A., Wadhams, L.J., Araujo, J., 2002. Encoding ofhost and non-host plant odours by receptor neurons in the eucalyptuswoodborer, Phoracantha semipunctata (Coleoptera: Cerambycidae). Journal ofComparative Physiology 188, 121–133.
Binyameen, M., Anderson, P., Ignell, R., Seada, M.A., Hansson, B.S., Schlyter, F., 2012.Spatial organization of antennal olfactory sensory neurons in the femaleSpodoptera littoralis moth: differences in sensitivity and temporalcharacteristics. Chemical Senses 37, 613–629.
Bengtsson, J.M., Wolde-Hawariat, Y., Khbaish, H., Negash, M., Jembere, B., Seyoum,E., Hansson, B.S., Larsson, M.C., Hillbur, Y., 2009. Field attractants for Pachnodainterrupta selected by means of GC–EAD and single sensillum screening. Journalof Chemical Ecology 35, 1063–1076.
Birkett, M.A., Bruce, T.J.A., Martin, J.L., Smart, L.E., Oakley, J., Wadhams, L.J., 2004.Responses of female orange wheat blossom midge, Sitodiplosis mosellana, towheat panicle volatiles. Journal of Chemical Ecology 30, 1319–1328.
Borg-Karlson, A.-K., Tengö, J., Valterová, I., Unelius, C.R., Taghizadeh, T., Tolasch, T.,Francke, W., 2003. (s)-(+)-Linalool, a mate attractant pheromone component inthe bee Colletes cunicularius. Journal of Chemical Ecology 29, 1–14.
Bruce, T.J.A., Wadhams, L.J., Woodcock, C.M., 2005a. Insect host location: a volatilesituation. Trends in Plant Science 10, 269–274.
Bruce, T.J.A., Birkett, M.A., Blande, J., Hooper, A.M., Martin, J.L., Khambay, B., Prosser,I., Smart, L.E., Wadhams, L.J., 2005b. Response of economically important aphidsto components of Hemizygia petiolata essential oil. Pest Management Science61, 1115–1121.
Burckhardt, D., 1998. Ctenarytaina eucalypti (Maskell) (Hemiptera, Psylloidea) neufür Mitteleuropa mit Bemerkungen zur Blattflohfauna von Eucalyptus.Mitteilungen der Entomologischen Gesellschaft Basel 48, 59–67.
Burckhardt, D., Santana, D.L.Q., Terra, A.L., Andrade, F.M., Penteado, S.R.C., Iede, E.T.,Morey, C.S., 1999. Psyllid pests (Hemiptera: Psylloidea) in South Americaneucalypt plantations. Bulletin de la Société Entomologique Suisse 72, 1–10.
Cadahia, D., 1980. Proximidad de dos nuevos enemigos de los Eucalyptus en Espan á.Bol Serv Plagas 6, 165–192.
Campbell, C.A.M., Pettersson, J., Pickett, J.A., Wadhams, L.J., Woodcock, C.M., 1993.Spring migration of damson-hop aphid, Phorodon humuli (Homoptera:Aphididae), and summer host plant-derived semiochemicals released onfeeding. Journal of Chemical Ecology 19, 1569–1576.
Chang, C., Cho, I., Li, Q.X., 2009. Insecticidal activity of basil oil, trans-anethole,estragole, and linalool to adult fruit flies of Ceratitis capitata, Bactrocera dorsalis,and Bactrocera cucurbitae. Journal of Economic Entomology 102, 203–209.
Chapman, R.F., Bernays, E.A., Simpson, S.J., 1981. Attraction and repulsion of theaphid, Cavariella aegopodii, by plant odors. Journal of Chemical Ecology 7, 881–888.
Collett, N., 2001. Biology and control of psyllids, and the possible causes fordefoliation of Eucalyptus camaldulensis Dehnh., river red gum in south-easternAustralia – a review. Australian Forestry 64, 88–95.
Costanzi, M., Costanzi, J.C., Cocqempot, C., 2003. Un nouveau psylle sur lesEucalyptus de la Riviera Ligure et de la Côte d’Azur. Premières observations deCtenarytaina spatulata Taylor dans le Bassin Méditerranéen occidental. Phytoma– La Défense des Végétaux 566, 48–51.
Dahlsten, D.L., Hansen, E.P., Zuparko, R.L., Norgaard, R.B., 1998a. Biological control ofthe blue gum psyllid proves economically beneficial. California Agriculture 52,35–40.
Dahlsten, D.L., Rowney, D.L., Copper, W.A., Tassan, R.L., Chaney, W.E., Robb, K.L.,Tjosvold, S., Bianchi, M., Lane, P., 1998b. Parasitoid wasp controls blue gumpsyllid. California Agriculture 52, 31–34.
de Bruyne, M., Foster, K., Carlson, J.R., 2001. Odor coding in the Drosophila antenna.Neuron 30, 537–552.
Den Otter, C.J., Van der Goes van Naters, W.M., 1992. Single cell recordings fromtsetse (Glossina m. morsitans) antennae reveal olfactory, mechano- and coldreceptors. Physiological Entomology 17, 33–42.
Doughty, R.W., 2000. The Eucalyptus: A Natural and Commercial History of the GumTree. The Johns Hopkins University Press, Baltimore, MD, p. 237.
Dudareva, N., Negre, F., Nagegowda, D.A., Orlova, I., 2006. Plant volatiles: recentadvances and future perspectives. Critical Reviews in Plant Sciences 25, 417–440.
Elliott, H.J., de Little, D.W., 1984. Insect Pests of Trees and Timber in Australia.Forestry Commission, Hobart, p. 90.
Gill, R.J., Oswalt, S.M., Veling, S.M., 1998. Recently introduced pests of Eucalyptus.California Plant Pest and Disease Report 17, 21–24.
Guédot, C., Millar, J.G., Horton, D.R., Landolt, P.J., 2009. Identification of a sexattractant pheromone for male winterform pear psylla, Cacopsylla pyricola.Journal of Chemical Ecology 35, 1437–1447.
Guédot, C., Horton, D.R., Landolt, P.J., 2010. Sex attraction in Bactericera cockerelli(Hemiptera: Triozidae). Environmental Entomology 39, 1302–1308.
Guerin, P.M., Stadler, E., Buser, H.R., 1983. Identification of host plant attractants forthe carrot fly, Psila rosae. Journal of Chemical Ecology 9, 843–861.
Hansson, B.S., Larsson, M.C., Leal, W.S., 1999. Green leaf volatile-detecting olfactoryreceptor neurones display very high sensitivity and specificity in a scarabbeetle. Physiological Entomology 24, 121–126.
Hardie, J., Visser, J.H., Piron, P.G.M., 1995. Peripheral odour perception by adultaphid forms with the same genotype but different host plant preference. Journalof Insect Physiology 41, 91–97.
Hern, A., Dorn, S., 2004. A female-specific attractant for the codling moth, Cydiapomonella, from apple fruit volatiles. Naturwissenschaften 91, 77–80.
Hodkinson, I.D., 1999. Biocontrol of eucalyptus psyllid Ctenarytaina eucalypti by theAustralian parasitoid Psyllaephagus pilosus: a review of current programmes andtheir success. Biocontrol News and Information 20, 129–134.
Hodkinson, D.I., 2007. A new introduced species of Ctenarytaina (Hemiptera,Psylloidea) damaging cultivated Eucalyptus parvula (=parvifolia) in Europe.Deutsche Entomologische Zeitschrift 54, 27–33.
Hollis, D., 2004. Australian Psylloidea: Jumping Plantlice and Lerp Insects.Australian Biological Resources Study, Canberra, p. 216.
Horton, D.R., Guédot, C., Landolt, P.J., 2007. Diapause status of females affectsattraction of male pear psylla, Cacopsylla pyricola, to volatiles from female-infested pear shoots. Entomologia Experimentalis et Applicata 123, 185–192.
Horton, D.R., Guédot, C., Landolt, P.J., 2008. Attraction of male summerform pearpsylla to volatiles from female pear psylla: effects of female age, mating statusand presence of the host. Canadian Entomologist 140, 184–191.

J.K. Yuvaraj et al. / Journal of Insect Physiology 59 (2013) 542–551 551
Kristoffersen, L., Hallberg, E., Wallén, R., Anderbrant, O., 2006. Sparse sensillar arrayon Trioza apicalis (Homoptera, Triozidae) antennae—an adaptation to highstimulus levels? Arthropod Structure & Development 35, 85–92.
Kristoffersen, L., Larsson, M.C., Anderbrant, O., 2008. Functional characteristics of atiny but specialized olfactory system: olfactory receptor neurons of carrotpsyllids (Homoptera: Triozidae). Chemical Senses 33, 759–769.
Lapis, E.B., Borden, J.H., 1993. Olfactory discrimination by Heteropsylla cubana(Homoptera: Psyllidae) between susceptible and resistant species of Leucana(Legunimosae). Journal of Chemical Ecology 19, 83–90.
Larsson, M.C., Leal, W.S., Hansson, B.S., 2001. Olfactory receptor neurons detectingplant odours and male volatiles in Anomala cuprea beetles (Coleoptera:Scarabaeidae). Journal of Insect Physiology 47, 1065–1076.
Lopes, O., Barata, E.N., Mustaparta, H., Araújo, J., 2002. Fine structure of antennalsensilla basiconica and their detection of plant volatiles in the eucalyptuswoodborer, Phoracantha semipunctata Fabricius (Coleoptera: Cerambycidae).Arthropod Structure & Development 31, 1–13.
Mann, R.S., Rouseff, R.L., Smoot, J., Rao, N., Meyer, W.L., Lapointe, S.L., Robbins, P.S.,Cha, D., Linn, C.E., Webster, F.X., Tiwari, S., Stelinski, L.L., 2012. Chemical andbehavioral analysis of the cuticular hydrocarbons from Asian citrus psyllid,Diaphorina citri. Insect Science. http://dx.doi.org/10.1111/j.1744-7917.2012.01541.x.
Murlis, J., Willis, M.A., Cardé, R.T., 2000. Spatial and temporal structures ofpheromone plumes in fields and forests. Physiological Entomology 25, 211–222.
Mustaparta, H., 2002. Encoding of plant odour information in insects: peripheraland central mechanisms. Entomologia Experimentalis et Applicata 104, 1–13.
Nehlin, G., Valterová, I., Borg-Karlson, A.-K., 1996. Monoterpenes released fromApiaceae and the egg-laying preferences of the carrot psyllid, Trioza apicalis.Entomologia Experimentalis et Applicata 80, 83–86.
Nojima, S., Linn, C., Morris, B., Zhang, A.J., Roelofs, W., 2003. Identification of hostfruit volatiles from hawthorn (Crataegus spp.) attractive to hawthorn-originRhagoletis pomonella flies. Journal of Chemical Ecology 29, 321–336.
Nottingham, S.F., Hardie, J., Dawson, G.W., Hick, A.J., Pickett, J.A., Wadhams, L.J.,Woodcock, C.M., 1991. Behavioral and electrophysiological responses of aphidsto host and nonhost plant volatiles. Journal of Chemical Ecology 17, 1231–1242.
Olivares, T.S., 2000. Ctenarytaina eucalypti (Maskell 1890): El psilido del eucalipto enChile (Hemiptera: Sternorryncha: Psylloidea: Spondyliaspininae). Gayana(Concepción) 64, 239–241.
Paine, T.D., Dahlsten, D.L., Millar, J.G., Hoddle, M., Hanks, L.M., 2000. UC scientistsapply IPM techniques to new eucalyptus pests. California Agriculture 54, 8–13.
Paine, T.D., Steinbauer, M.J., Lawson, S.A., 2011. Native and exotic pests ofEucalyptus: a worldwide perspective. Annual Review of Entomology 56, 181–201.
Patt, J.M., Sétamou, M., 2010. Responses of the Asian citrus psyllid to volatilesemitted by the flushing shoots of its rutaceous host plants. Journal of ChemicalEcology 39, 618–624.
Pettersson, J., Pickett, J.A., Pye, B.J., Quiroz, A., Smart, L.E., Wadhams, L.J., Woodcock,C.M., 1994. Winter host component reduces colonization by bird-cherry-oataphid, Rhopalosiphum padi (L.) (Homoptera, Aphididae), and other aphids incereal fields. Journal of Chemical Ecology 20, 2565–2574.
Phillips, A.K., Appel, A.G., Sims, S.R., 2010. Topical toxicity of essential oils to theGerman cockroach (Dictyoptera: Blattellidae). Journal of Economic Entomology103, 448–459.
Pichersky, E., Gershenzon, J., 2002. The formation and function of plant volatiles:perfumes for pollinator attraction and defense. Current Opinion in Plant Biology5, 237–243.
Pickett, J.A., Wadhams, L.J., Woodcock, C.M., Hardie, J., 1992. The chemical ecologyof aphids. Annual Review of Entomology 37, 67–90.
Powell, G., Hardie, J., 2001. The chemical ecology of aphid host alternation: how doreturn migrants find the primary host plant? Applied Entomology and Zoology36, 259–267.
Purvis, G., Chauzat, M.P., Segonds-Pichon, A., Dunne, R., 2002. Life history andphenology of the eucalyptus psyllid, Ctenarytaina eucalypti in Ireland. Annals ofApplied Biology 141, 283–292.
Sant’ana, J., Dickens, J.C., 1998. Comparative electrophysiological studies of olfactionin predaceous bugs Podisus maculiventris and P. nigrispinus. Journal of ChemicalEcology 24, 965–984.
Santana, D.L.Q., 2005. Psilídeos em Eucaliptos no Brasil. Embrapa Florestas. CircularTécnica, 109, pp. 1–14.
Santana, D.L.Q., Burckhardt, D., 2007. Introduced Eucalyptus psyllids in Brazil.Journal of Forest Research 12, 337–344.
Santos, P.E.T.D., Geraldi, I.O., Garcia, J.N., 2004. Estimates of genetic parameters ofwood traits for sawn timber production in Eucalyptus grandis. Genetics andMolecular Biology 27, 567–573.
Schoonhoven, L.M., Jermy, T., Van loon, J.J.A., 1998. Insect–Plant Biology: FromPhysiology to Evolution. Chapman and Hall, London, ISBN 0-412-58700-9.
Silk, P.J., Lemay, M.A., LeClair, G., Sweeney, J., MaGee, D., 2010. Behavioral andelectrophysiological responses of Tetropium fuscum (Coleoptera: Cerambycidae)to pheromone and spruce volatiles. Environmental Entomology 39, 1997–2005.
Soroker, V., Talebaev, S., Harari, A.R., Wesley, S.D., 2004. The role of chemical cues inhost and mate location in the pear psylla Cacopsylla bidens (Homoptera:Psyllidae). Journal of Insect Behavior 17, 613–626.
Stensmyr, M.C., Larsson, M.C., Bice, S., Hansson, B.S., 2001. Detection of fruit- andflower-emitted volatiles by olfactory receptor neurons in the polyphagous fruitchafer Pachnoda marginata (Coleoptera: Cetoniinae). Journal of ComparativePhysiology A 187, 509–519.
Street, R.A., Hewitt, C.N., Mennicken, S., 1997. Isoprene and monoterpene emissionsfrom a Eucalyptus plantation in Portugal. Journal of Geophysical Research 102,15875–15887.
Tasin, M., Bäckman, A.-C., Coracini, M., Casado, D., Ioriatti, C., Witzgall, P., 2007.Synergism and redundancy in a plant volatile blend attracting grapevine mothfemales. Phytochemistry 68, 203–209.
Su, C.-Y., Menuz, K., Reisert, J., Carlson, J.R., 2012. Non-synaptic inhibition betweengrouped neurons in an olfactory circuit. Nature 492, 66–71.
Taylor, K.L., 1997. A new Australian species of Ctenarytaina Ferris and Klyver(Hemiptera: Psyllidae: Spondyliaspidinae) established in three other countries.Australian Journal of Entomology 36, 113–115.
Unsicker, S.B., Kunert, G., Gershenzon, J., 2009. Protective perfumes: the role ofvegetative volatiles in plant defense against herbivores. Current Opinion inPlant Biology 12, 479–485.
Visser, J.H., Avé, D.A., 1978. General green leaf volatiles in the olfactory orientationof the Colorado beetle, Leptinotarsa decemlineata. Entomologia Experimentaliset Applicata 24, 538–549.
Visser, J.H., Piron, P.G.M., 1995. Olfactory antennal responses to plant volatiles inapterous virginoparae of the vetch aphid Megoura viciae. EntomologiaExperimentalis et Applicata 77, 37–46.
Visser, J.H., Piron, P.G.M., Hardie, J., 1996. The aphids’ peripheral perception of plantvolatiles. Entomologia Experimentalis et Applicata 80, 35–38.
Wenninger, E.J., Stelinski, L.L., Hall, D.G., 2008. Behavioral evidence for a femaleproduced sex attractant in Diaphorina citri. Entomologia Experimentalis etApplicata 128, 450–459.
Wibe, A., Mustaparta, H., 1996. Encoding of plant odours by receptor neurons in thepine weevil (Hylobius abietis) studied by linked gas chromatography–electrophysiology. Journal of Comparative Physiology A 179, 331–344.
Wirthensohn, M.G., Sedgley, M., Jones, G.P., 2000. Epicuticular wax of juvenileEucalyptus leaves and headspace analysis of leaf volatiles. Journal of EssentialOil Research 12, 401–411.
Yang, M.M., Yang, C.T., Chao, J.T., 1986. Reproductive isolation and taxonomy of twoTaiwanese Paurocephala species (Homoptera: Psylloidea). Monograph of TaiwanMuseum 6, 176–203.
Yassaa, N., Meklati, B.Y., Cecinato, A., 2000. Evaluation of monoterpenic biogenicvolatile organic compounds in ambient air around Eucalyptus globulus, Pinushalepensis and Cedrus atlantica trees growing in Algiers city area by chiral andachiral capillary gas chromatography. Atmospheric Environment 34, 2809–2816.
Zhang, Q.H., Schlyter, F., 2004. Olfactory recognition and behavioral avoidance ofangiosperm nonhost volatiles by conifer inhabiting bark beetles. Agriculturaland Forest Entomology 6, 1–19.
Zini, C.A., Augusto, F., Christensen, E., Smith, B.P., Caramão, E.B., Pawliszyn, J., 2001.Monitoring biogenic volatile compounds emitted by Eucalyptus citriodora usingSPME. Analytical Chemistry 73, 4729–4735.
Zini, C.A., Augusto, F., Christensen, E., Caramão, E.B., Pawliszyn, J., 2002. SPMEapplied to the study of volatile organic compounds emitted by three Species ofEucalyptus in situ. Journal of Agriculture and Food Chemistry 50, 7199–7205.
Copyright © 2022 FDOKUMEN




















