
Functional organization of the main olfactory bulb
15
Functional Organization of the Main Olfactory Bulb JOHN W. SCOTT, DAVID P. WELLIS, MARCIA J. RIGGOTT, AND NATHALIE BUONVISO Department of Anatomy and Cell Biology, Emory University School of Medicine, Atlanta, Georgia 30322 (J. W.S.); Department of Neurosurgery, Tufts-New England Mea ical Center, Boston, Massachusetts 021 11 (D.P. W.); Department of Neurobiology, Duke University, Durham, North Carolina 27710 (M.J.R.); Laboratoire de Physiologie Neurosensorielle, UCB Lyon I-CNRS L'RA 180, 69622 Villerbanne Ceder, France (N.B.) KEY WORDS Sensory processing, Olfactory coding, Olfaction, Odor stimulation ABSTRACT Completcb understanding of the role of the mammalian main olfactory bulb in sensory processing has remained elusive despite many detailed studies on its anatomy and phys- iology. Several lines of recent evidence viewed in the context of earlier knowledge have provided new insights into the bullsar mechanisms of olfactory coding. The output cells of the olfactory bulb receive a localized olfactory nerve input and interneuronal input via dendrodendritic synapses on distinct sets of dendrites. The spatial arrangement of granule cell contacts on output cell basal dendrites suggests that lat,eral inhibitory interactions may occur between neighboring output cells. The input from olfactory receptor cell axons to the bulb also has spatial order, but does not repre- sent a precise map of the ceceptor surface. Recent studies with antibodies and lectins suggest that different groups of axons from chemically similar receptor cells collect into certain glomeruli, even if the axons originate from cells that are not contiguous in the mucosa. Electrophysiological studies have begun to explore the participation of spatially organized circuits in olfactory processing. The degree to which neighboring output cells respond similarly to odor stimulation, for example, de- pends on the distance between the cells, with those further apart showing complementary re- sponses. Also, a single output cell can show 2 or more different temporal response patterns when different odors are presented. Intracellular recordings indicate that these responses are shaped by IPSPs. Electrical stimulation during such recordings shows that some mitral cells are excited by nerve inputs close to their glomerular tufts, while they are inhibited by nerve inputs to other parts of the bulb. Finally, recordings from granule and periglomerular cells indicate their potential in mediating components of output cell odor responses. These considerations suggest that the olfactory bulb performs a spatially based analysis on the information coming from the receptor cells. While the spatial organization of the olfactory bulb is probably not faithfully represented in the projec- tions to the olfactory cortex, bulbocortical projections are not random. The fact that spatial factors exist at each of these levels in the olfactory system must be considered in developing models of Central olfactory processing. 0 1993 Wiley-Liss, Inc INTRODUCTION The study of olfaction has not yet provided a convinc- ing full description of the role of the main olfactory bulb in the process of odor discrimination. However, a number of features give strong 3ues about the role of this intriguing structure in procnessing olfactory infor- mation. We will review some of these features, discuss some new data, and attempt to outline some unresolved issues standing in the way of a systematic view of mammalian main olfactory bulh function. This will be a selective review and will not deal with some of the issues covered in other articles in this series. We will first review the arrangement of sensory input to the glomeruli and speculate upon its functional conse- quences. After describing some morphological features of the output cells and the interneurons that can act upon those cells, we will discuss some of the physiolog- ical properties of these circuits and suggest some con- sequences of these properties fcjr the further analysis by more central parts of the system. ORGANIZATION OF THE MAIN OLFACTORY BULB SENSORY AFFERENTS The mammalian olfactory bulb contains both output neurons and interneurons arranged in a concentric laminar fashion (Fig. 1). The output neurons, the mi- tral and tufted cells, are located within the bulb in the mitral cell layer and external plexiform layer, respec- tively. The main groups of interneurons exist at 2 lev- els within the bulb: Granule cells are located deep to the mitral and tufted cells, whereas periglomerular neurons lie more superficially, in the glomerular layer. Dendrodendritic synapses are formed between mitral: tufted cell basal dendrites in the external plexiform layer and between mitralitufted apical dendrites and periglomerular cells within the glomeruli. It is within Received January 24, 1991; accepted in revised form July 8, 1991. Address reprint requests to John W. Scott, Department of Anatomy and Cell Biology, Emory University School of Medicine, Atlanta, GA 30322. 0 1993 WILEY-LISS. INC.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Functional organization of the main olfactory bulb

Functional Organization of the Main Olfactory Bulb JOHN W. SCOTT, DAVID P. WELLIS, MARCIA J. RIGGOTT, AND NATHALIE BUONVISO Department of Anatomy and Cell Biology, Emory University School of Medicine, Atlanta, Georgia 30322 (J. W.S.); Department of Neurosurgery, Tufts-New England Mea ical Center, Boston, Massachusetts 021 11 (D.P. W.); Department of Neurobiology, Duke University, Durham, North Carolina 27710 (M.J.R.); Laboratoire de Physiologie Neurosensorielle, UCB Lyon I-CNRS L'RA 180, 69622 Villerbanne Ceder, France (N.B.)
KEY WORDS Sensory processing, Olfactory coding, Olfaction, Odor stimulation
ABSTRACT Completcb understanding of the role of the mammalian main olfactory bulb in sensory processing has remained elusive despite many detailed studies on its anatomy and phys- iology. Several lines of recent evidence viewed in the context of earlier knowledge have provided new insights into the bullsar mechanisms of olfactory coding. The output cells of the olfactory bulb receive a localized olfactory nerve input and interneuronal input via dendrodendritic synapses on distinct sets of dendrites. The spatial arrangement of granule cell contacts on output cell basal dendrites suggests that lat,eral inhibitory interactions may occur between neighboring output cells. The input from olfactory receptor cell axons to the bulb also has spatial order, but does not repre- sent a precise map of the ceceptor surface. Recent studies with antibodies and lectins suggest that different groups of axons from chemically similar receptor cells collect into certain glomeruli, even if the axons originate from cells that are not contiguous in the mucosa. Electrophysiological studies have begun to explore the participation of spatially organized circuits in olfactory processing. The degree to which neighboring output cells respond similarly t o odor stimulation, for example, de- pends on the distance between the cells, with those further apart showing complementary re- sponses. Also, a single output cell can show 2 or more different temporal response patterns when different odors are presented. Intracellular recordings indicate that these responses are shaped by IPSPs. Electrical stimulation during such recordings shows that some mitral cells are excited by nerve inputs close to their glomerular tufts, while they are inhibited by nerve inputs to other parts of the bulb. Finally, recordings from granule and periglomerular cells indicate their potential in mediating components of output cell odor responses. These considerations suggest that the olfactory bulb performs a spatially based analysis on the information coming from the receptor cells. While the spatial organization of the olfactory bulb is probably not faithfully represented in the projec- tions to the olfactory cortex, bulbocortical projections are not random. The fact that spatial factors exist at each of these levels in the olfactory system must be considered in developing models of Central olfactory processing. 0 1993 Wiley-Liss, Inc
INTRODUCTION
The study of olfaction has not yet provided a convinc- ing full description of the role of the main olfactory bulb in the process of odor discrimination. However, a number of features give strong 3ues about the role of this intriguing structure in procnessing olfactory infor- mation. We will review some of these features, discuss some new data, and attempt to outline some unresolved issues standing in the way of a systematic view of mammalian main olfactory bulh function. This will be a selective review and will not deal with some of the issues covered in other articles in this series. We will first review the arrangement of sensory input to the glomeruli and speculate upon its functional conse- quences. After describing some morphological features of the output cells and the interneurons that can act upon those cells, we will discuss some of the physiolog- ical properties of these circuits and suggest some con- sequences of these properties fcjr the further analysis by more central parts of the system.
ORGANIZATION OF THE MAIN OLFACTORY BULB SENSORY AFFERENTS
The mammalian olfactory bulb contains both output neurons and interneurons arranged in a concentric laminar fashion (Fig. 1). The output neurons, the mi- tral and tufted cells, are located within the bulb in the mitral cell layer and external plexiform layer, respec- tively. The main groups of interneurons exist a t 2 lev- els within the bulb: Granule cells are located deep to the mitral and tufted cells, whereas periglomerular neurons lie more superficially, in the glomerular layer. Dendrodendritic synapses are formed between mitral: tufted cell basal dendrites in the external plexiform layer and between mitralitufted apical dendrites and periglomerular cells within the glomeruli. It is within
Received January 24, 1991; accepted in revised form July 8 , 1991. Address reprint requests to John W. Scott, Department of Anatomy and Cell
Biology, Emory University School of Medicine, Atlanta, GA 30322.
0 1993 WILEY-LISS. INC.

143 MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION
ONL ---------------\ 7 - ------
47 EPL
Fig. 1. Schematic representation of the layeringin the mammalian olfactory bulb. The principal cells exist in 2 main types, the mitral (M) and tufted (TI cells. These cells have apical dendrites that ascend to have dense branching in the circular glomeruli (GL). The olfactory nerve axons pass over the surface of the bulb, forming the olfactory nerve layer (ONL), from which they descend into the glomeruli to make synaptic contact. One major group of interneurons, the periglomerular cells (PG), makes reciprocal dendrodendritic contacts with the mitrali tufted cell apical dendrites in the glomeruli. The best studied inter- neurons in the bulb are the granule cells (GI, with somata in the granule cell layer (GCL) below the mitral cell layer (MCL). These cells make reciprocal dendrodendritic synapses with the basal dendrites of mitralitufted cells within the external plexiform layer (EPL).
these specialized regions of neuropil that the second order cells (mitralkufted cells and periglomerular cells) also receive synaptic input from the olfactory re- ceptor neurons. There are a variety of short axon cells, whose physiology has not been studied and will not be considered here. Further detail about the anatomical organization of the olfactory bulb can be found in sev- eral reviews (Harrison and Scott, 1986; Mori, 1987b; Shepherd, 1972; Shepherd and Greer, 1990).
The receptor cells of the olfactory epithelium project axons to the olfactory bulb in a spatially organized fashion (Astic et al., 1987; Clancy et al., 1985; Free- man, 1974c; Jastreboff et al., 1984; Land, 1973; Le Gros Clark, 1951), but the fact that the projection does not have a point-to-point topography has been open to sev- eral possible interpretations. One possibility is that the mechanisms generating the topography are simply not precise and that central mechanisms must act to cor- rect this lack of precision. Kauer (1987) has discussed another possibility, that there is good topography from some classes of receptor cells and diffuse projections from other types of receptor cells. These 2 groups of axons might end on different cell types within the glomeruli and have different functional consequences. Another notion, originally suggested by Le Gros Clark (1951), is that receptor cells with similar response spec- t ra might converge onto the same glomerulus or group of glomeruli. An interesting variant of this notion is the suggestion by observations in some insects that there is convergence of s o n s from receptors of dissim- ilar function onto particular glomeruli (Boeckh et al., 1990; Rodrigues, 1988). Other insect examples, partic- ularly in the pheromone system, show examples of con- vergence of similar receptor inputs to specific glomer- ular regions (Hansson et al., 1992).
Le Gros Clark (1951) suggested that there are “cat- egories of receptors” whose axons are rearranged in the olfactory nerve layer so that they enter “glomeruli of corresponding categories” without complete restriction by their place of origin. He further suggested that these inputs might remain separated as far as the mitral cell projections to the piriform cortex. A variant of this pro- posal is that the rearrangement of s o n s as they pass onto the surface of the olfactory bulb places inputs from the nasal recesses together and those from the promi- nences of the turbinal bones together. This type of ar- rangement was indicated by Adrian’s (1950) idea of regional excitation of the mucosa due to differential absorption of odorant molecules into the mucus and is supported by recent observations with retrograde trac- ers (Astic et al., 1987; Clancy et al., 1985).
The first data to support Le Gros Clark’s idea of func- tional segregation of glomeruli came from electrophys- iological studies with DC shifts in recordings from the glomeruli (Leveteau and MacLeod, 1966). More recent studies with 2-deoxyglucose autoradiography (Cooper- smith and Leon, 1984; Jourdan et al., 1980; Stewart et al., 1979) have shown that some glomeruli respond to different odor stimuli than do neighboring glomeruli. Mori et al. (1992) have shown that mitral cells in the dorsomedial region of the rabbit olfactory bulb respond differentially to fatty acids. In high resolution 2-deoxy- glucose studies, there was a uniform intensity of reac- tion throughout each glomerulus (Lancet et al., 1982). Combination of cfos and 2-deoxyglucose labeling has shown activation of the neurons forming the borders of the activated glomeruli (Guthrie et al., 1993). These observations have led to the use of the term functional unit to describe the discrete activation of certain glom- eruli or glomerular groups by specific odors.
These 2-deoxyglucose observations did not determine the mechanism by which adjacent glomeruli differ in their odor responses or how each glomerulus responds as a unit. These properties might arise by convergence of receptor cells with similar properties onto a single glomerulus. An alternate possibility is that many dif- ferent types of afferents converge into some individual glomeruli. This has been suggested for some insects (Boeckh et al., 1990; Rodrigues, 1988). In the latter case the unity of response would come from some in- teraction within the glomerulus, either of a synaptic nature or from some nonsynaptic process, such as an elevated external potassium concentration caused by the glial wrapping of the glomeruli (Freeman, 1974b; Khayari et al., 1988; Pinching and Powell, 1971). Func- tionally, the convergence of unlike receptor inputs would provide a mechanism by which new, nonlinear combinations of inputs from receptors could be made, adding to the complexity of molecular properties that could be coded at the glomerular level.
In order to ascertain the mechanisms that determine the response properties of the glomeruli, i t will be nec- essary to measure the odor responses of receptor axon populations that innervate particular groups of glo- meruli. These physiological experiments cannot be eas- ily done in the mammal. Nevertheless, there are sev- eral anatomical markers, both antibodies and lectins, that label groups of receptors, their axons, and glom-

144 J.W. SCOTT ET AL.
eruli. If one makes the reasonable assumption that these markers correlate either with the response prop- erties of the receptors or, more likely, with the mech- anisms that guided axons to their targets during de- velopment, then the labeling pattern of receptors and glomeruli may give clues to the range of sensory infor- mation that is transmitted t o a glomerulus.
The only main olfactory bulb glomeruli that can be identified morphologically across individuals lie in the posterior region. In the rat , thess are the modified glo- merular complex (Greer et a l , 1982), the atypical glomeruli (Zheng et al., 1987), and the necklace glo- meruli (Shinoda et al., 1989). The modified glomerular complex appears to receive affepents only from a sub- population of receptor cells along the septa1 wall and nearby regions (Jastreboff et al., 1984). The atypical glomeruli are a set of specialized glomeruli that stain for acetylcholinesterase from centrifugal sources and also receive a special set of olfactory afferents that are electron lucent and contain dense core vesicles unlike most olfactory afferents, which Ese electron opaque and contain clear vesicles (Zheng arid Jourdan, 1988). The necklace glomeruli, like the atypical glomeruli, circle the caudal portion of the bulb and label with an anti- body to a human placental antigen. The same antibody stains receptor cells scattered widely through the ol- factory epithelium (Shinoda et al., 19891, leading to the conclusion that these glomeruli receive a wide spatial convergence. It is not yet established whether the neck- lace and atypical glomeruli are identical.
Data also exist on the distribution of receptors pro- jecting to ordinary glomeruli of the main olfactory bulb. Some data appear to favor a topographic organi- zation of the afferents. For both the rabbit (Fujita et al., 1985) and the rat (Schwob and Gottleib, 19861, anti- bodies have been found that evenly label a population of glomeruli in one region of the bulb and label a cor- responding set of spatially localized olfactory nerve ax- ons andlor receptor cells. These are quite spatially de- fined and suggest that some glomerular innervation might be strictly topographic. For example, in the case of the rat, the labeled receptor; lie in the recesses be- tween the turbinal bones (Schvrob and Gottleib, 1986). Other data with antibodies and lectins suggest that there are additional nontopographic factors in the pro- jection. Mori (1987a) reported that another antibody, which labeled receptors scattered throughout the epi- thelium, labeled glomeruli in :in uneven pattern. The labeled glomeruli were often found interspersed among unlabeled glomeruli, and the label varied in intensity among the labeled glomeruli. This appears to be an example in which a subpopulation of receptors has no localization in the mucosa and for which there may be a local cue leading the receptor axons to bypass certain glomeruli in order to innervate others.
Lectins have also been found to mark olfactory re- ceptors and/or olfactory glomeruli. Among the lectins that have shown differential labeling patterns in the adult rat are soybean agglulhin (Key and Giorgi, 19861, Uler europaeus 1 (UEAL, Barber, 1989; Riggott and Scott, 1989), and Lotus retragonolobus (Stewart and Touloukian, 1990). We surveyed a number of plant lectins, including those listed, and found the fucose
binding lectin UEAl to produce the strongest and most reliable labeling. Figure 2 illustrates a micrograph of typical labeling with this lectin, showing glomeruli la- beled with a gradation of densities and showing a re- gion where glomeruli of different labeling densities are intermixed. The labeling of receptor cells in the epithe- lium seems to be correlated with their position, so that heavily labeled receptor cells are found in the recesses between pairs of turbinal bones (Fig. 1B). There is a generalized topography of the labeling in the olfactory bulb, with the ventral and lateral parts of the bulb showing the most intense label. The great variation in intensity of UEAl labeling among adjacent glomeruli supports some of the antibody staining, mentioned above, indicating the possibility that local cues might override topography in determining some aspects of the projection pattern.
Le Gros Clark (1951) reported that there is a sub- stantial rearrangement of the olfactory receptor axons as they enter onto the surface of the olfactory bulb. This rearrangement might explain the local variations in projection implied by the labeling studies. While both Freeman (1974) and Wellis and Scott (1991) have demonstrated that there is a good topographic organi- zation of the axons in the olfactory nerve layer of the lateral surface of the bulb that is useful for electrophys- iological studies, this organization has some variation. Small fascicles of axons commonly deviate 1 or 2 glo- merular widths at their point of termination and sev- eral fascicles may enter a glomerulus from different directions. This behavior is consistent with a pattern in which factors other than topography or random error determine the axon population entering a glomerulus.
In general, the literature on the olfactory nerve pro- jection seems consistent with the presence of both topo- graphic and nontopographic principles of organization. These 2 modes of organization in the projection produce considerable complexity in the afferent pattern and re- quire sophisticated analysis by the central nervous sys- tem.
Recently Buck and Axel (1991) identified a large multigene family that appears to encode olfactory re- ceptor proteins. Their recent work suggests that cells expressing particular genes may be localized within the mucosa.
HOW DOES THE OLFACTORY BULB PROCESS THE INFORMATION FROM THE
OLFACTORY NERVE? At least 2 opposing viewpoints could be taken about
central processing of olfaction. By one view, rather sim- ilar to that expressed by Le Gros Clark (19511, the classes of olfactory experience are rather similar to the classes of receptor cells (or of binding sites on receptor cells). The influential suggestions of Amoore et al. (1964) are an example of this idea. In this view, the purpose of central processing is the separation of infor- mation that may be mixed because of the potential oc- currence of multiple molecular receptors on the same receptor cell or because of poor precision in the mu- cosobulbar projection. An analogy would be the mech- anisms suggested for sharpening of spatial localization
MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION
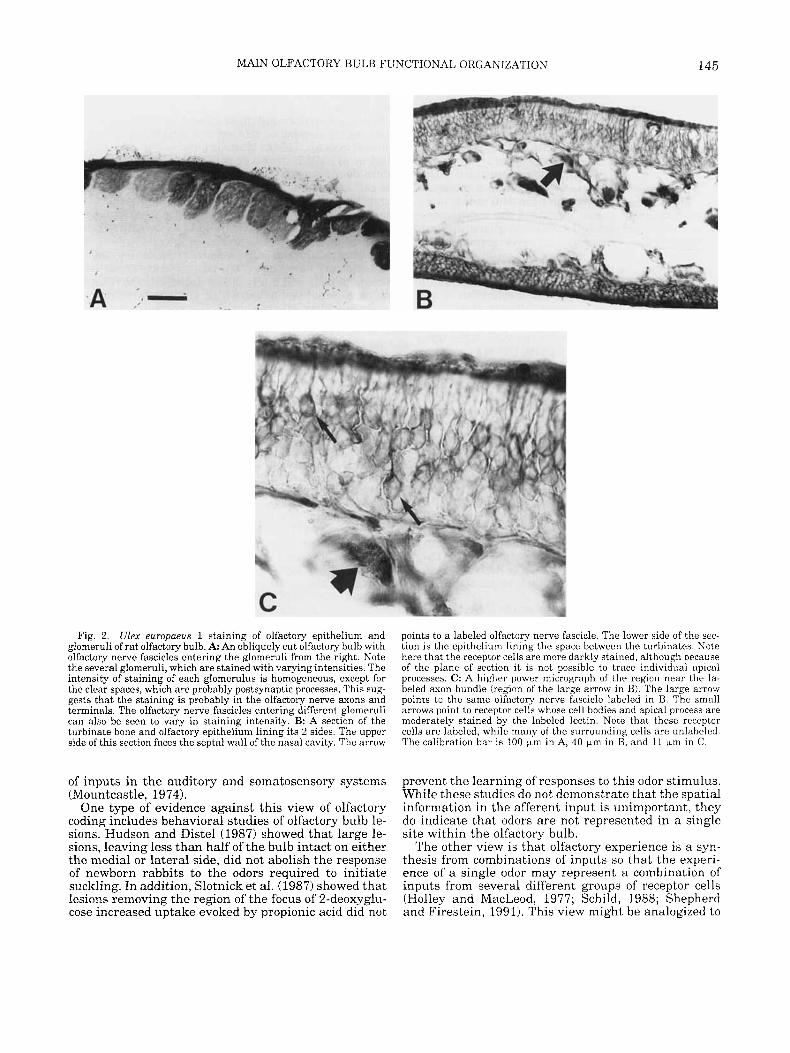
Fig. 2. Ulex europuew 1 staining of olfactory epithelium and glomeruli of rat olfactory bulb. A: An obliquely cut olfactory bulb with olfactory nerve fascicles entering the glomeruli from the right. Note the several glomeruli, which are stained with varying intensities. The intensity of staining of each glomerulus is homogeneous, except for the clear spaces, which are probably postsynaptic processes. This sug- gests that the staining is probably in the olfactory nerve s o n s and terminals. The olfactory nerve fascicles entering different glomeruli can also be seen to vary in staining intensity. B: A section of the turbinate bone and olfactory epithelium lining its 2 sides. The upper side of this section faces the septa1 wall of the nasal cavity. The arrow
of inputs in the auditory and somatosensory systems (Mountcastle, 1974).
One type of evidence against this view of olfactory coding includes behavioral studies of olfactory bulb le- sions. Hudson and Distel (1987) showed that large le- sions, leaving less than half of the bulb intact on either the medial or lateral side, did not abolish the response of newborn rabbits to the odors required to initiate suckling. In addition, Slotnick et al. (1987) showed that lesions removing the region of the focus of 2-deoxyglu- cose increased uptake evoked by propionic acid did not
145
points to a labeled olfactory nerve fascicle. The lower side of the sec- tion is the epithelium lining the space between the turbinates. Note here that the receptor cells are more darkly stained, although because of the plane of section it is not possible to trace individual apical processes. C: A higher power micrograph of the region near the la- beled axon bundle (region of the large arrow in B). The large arrow points to the same olfactory nerve fascicle labeled in B. The small arrows point to receptor cells whose cell bodies and apical process are moderately stained by the labeled lectin. Note that these receptor cells are labeled, while many of the surrounding cells are unlabeled. The calibration bar is 100 km in A, 40 km in B, and 11 K r n in C.
prevent the learning of responses to this odor stimulus. While these studies do not demonstrate that the spatial information in the afferent input is unimportant, they do indicate that odors are not represented in a single site within the olfactory bulb.
The other view is that olfactory experience is a syn- thesis from combinations of inputs so that the experi- ence of a single odor may represent a combination of inputs from several different groups of receptor cells (Holley and MacLeod, 1977; Schild, 1988; Shepherd and Firestein, 1991). This view might be analogized to

146 J.W. SCOTT ET AL.
visual cortex cell responses to lines. These responses are put together from the responses of ganglion cells to concentric, antagonistic receptive fields (Hubel and Wiesel, 1962), which are in turn put together from the simpler responses of receptor cell!; (Kuffler, 1953). This second view changes the role of the olfactory bulb from a stage involved in refining away unwanted noise, through a process of lateral inhibition, to a process of combining afferent information from different sources. It is noteworthy in this context that the hypothesized convergence of receptor cell axons into glomeruli could serve either function; that is, the convergence of axons from cells with like responsiveiiess could serve as a mechanism for purification of input. On the other hand, convergence of unlike receptor cell axons into single glomeruli would provide one mechanism of com- bination of inputs from receptor cells sensitive to dif- ferent properties of the same odor molecule. SPECIFIC MORPHOLOGICAL FEATURES OF
OLFACTORY BULB CIRCUITRY The morphology of the principal cells of the mamma-
lian olfactory bulb, the mitral and tufted cells, is a good illustration of the ideas about olfactory bulb function. Two of the most striking aspect 3 of the morphology of these cells are their glomeruIar dendritic arboriza- tions, through which they receive olfactory nerve syn- apses, and their very long basal dendrites, which have excitatory output on granule cells and receive inhibi- tory feedback from those cells. These basal dendrites extend as far as a millimeter for rat mitral cells (Orona et al., 1984), a significant fraction of the circumference of the olfactory bulb. These 2 features of the mitrali tufted cells emphasize the discrete nature of the input at the glomerulus as well as the potential for continu- ous interaction with many interneurons along the basal dendrites. This section will briefly describe some of these features with a view to their significance in olfactory processing.
The glomeruli are sites of considerable convergence of olfactory nerve axons onto t?e apical dendrites; in rabbits this convergence is thought to average about 1,000 receptor axons per mi t rd cell (Allison, 1953). The glomeruli are surrounded by glial sheaths (Pinching and Powell, 1971) and by groups of small neurons. Among these small neiirons are a type known as periglomerular cells, which are defined as cells send- ing spiny dendrites into the glomeruli, where they form dendrodendritic synaptic conthcts with mitralitufted cell apical dendrites and in some cases also receive di- rect olfactory nerve contacts 1 Pinching and Powell, 1971; Schneider and Macrides, 1978; White, 1972). The periglomerular cells contain gamma amino-butyric acid (GABA) and/or dopamine (Baker, 1988; Halasz et al., 1981; Mugnaini et al., 19841. They have often been suggested to have inhibitory action on the basis of their GABA content and the presence of flattened synaptic vesicles after aldehyde fixation (Mori, 1987b; Shep- herd, 1972). We will make further comments on the function of these cells in a la tw section. The periglo- merular region also contains a population of tufted cells, many of which lack the long basal dendrites of deeper tufted cells. Some of these tufted cells appear to
give rise to long association connections within the bulb (Schoenfeld et al., 1985).
Most mammalian mitral cells and tufted cells send their apical dendrites into only a single glomerulus (Buonviso et al., 1991a; Macrides and Schneider, 1982; Mori et al., 1983; Orona et al., 1984). The mitral cell apical dendrites can vary in their angle with respect to the cell layers, but there is a strong tendency for these dendrites to be parallel, so that most of the mitral cells innervating a single glomerulus lie very close together (Buonviso et al., 1991a). This might be expected to pro- duce similar responses in neighboring cells.
In contrast to the very localized excitatory input from olfactory nerve axons onto the primary dendrites, the mitral cells and many of the tufted cells have very extensive basal dendrites, which enter into reciprocal dendrodendritic synapses with the specialized spines (or gemmules) of granule cells (Price and Powell, 1970; Rall et al., 1966; Shepherd, 1972). The mitral cell side of this dendrodendritic synapse is excitatory, probably through the release of the excitatory amino acid gluta- mate (Jacobson, 1986; Trombley and Westbrook, 1990). The granule cell input onto mitral cell dendrites is in- hibitory and is mediated at least in part by GABA (Nicoll, 1971; Jahr and Nicoll, 1982). The anatomical data on the lengths of the mitral cell basal dendrites suggest that a mitral cell can receive inhibitory inputs from sources that are up to 1 mm away from the cell body. The spatial convergence of these inputs through granule cell action onto mitralitufted basal dendrites is probably amplified by a number of connections involv- ing axon collaterals and other interneurons, such as short axon cells. Many authors have suggested that this convergent inhibitory input serves to transform the incoming signal in some way. We shall review these suggestions and the data that bear on this issue.
The mitral and tufted cells exist in several subtypes that vary in the lamination of their basal dendrites within the external plexiform layer. Three sublaminae of the rat external plexiform layer have been described on the basis of these patterns of lamination and on the basis of density of cytochrome oxidase staining (Orona et al., 1984; Mouradian and Scott, 1988). Mitral cells extend basal dendrites within either the deep or inter- mediate sublaminae. The several subtypes of tufted cells extend basal dendrites within the intermediate or superficial sublaminae, usually the same sublamina containing the cell body. Certain external tufted cells of the hamster do not have basal dendrites (Macrides and Schneider, 1982). The details of these subtypes have been reviewed more extensively by Scott and Har- rison (1987).
The mitralhufted cell types of the rabbit are similar, although a displaced mitral cell has been described that may correspond to the internal tufted cell of the rat and hamster. Some hamster tufted cell axons prob- ably do not leave the bulb but contribute to intrabulbar association connections (Schoenfeld et al., 1985). Most rat tufted cells having basal dendrites project axons out of the bulb, where they reach at least as far as the anterior olfactory nucleus (Scott et al., 1985). Mitral cells as a group have longer axons than tufted cells, and those mitral axons project back to the piriform

MAIN OLFACTORY BULB FUNCTIONAL ORGAIVIZATION 147
cortex in several species (Davis et al., 1978; Haberly and Price, 1977; Skeen and Hall, 1977). This differen- tial projection has been useful in providing an electro- physiological distinction between mitral and tufted cells (Scott, 1981; Schneider and Scott, 1983).
Granule cells of the mammalian bulb exist in at least 2 populations, although the exact details of these pop- ulations may vary with species. One population of granule cells has been found with maximal spine den- sity in the upper 2 sublayers of the external plexiform layer, while other granule cells have maximal spine density in the 2 lower sublayers of the external plexi- form layer. In the rat these 2 groups of granule cells were reported to correlate with the position of granule cell somata (Orona et al., 1983). Reports in other spe- cies have noted subpopulations of granule cells with similarly restricted spine distributions, but they dif- fered in finding other types that spanned all layers and in also failing to see a correlation with the position of the soma (Greer, 1987; Mori et al., 1983; Wahle et al., 1990). It is as yet unclear whether the inputs to the basal processes of these granule cells are in any way laminated, although it is clear in both rat and hamster that there are laminated afferents from the forebrain that terminate in the granule cell layer (Davis et al., 1978; Luskin and Price, 1983).
THE PHYSIOLOGICAL ACTION OF OLFACTORY BULB CIRCUITS
The morphological studies reviewed above showed that the olfactory bulb contains a complicated circuitry that should be capable of comparing inputs over spa- tially separated glomeruli as one step in recognizing a complex odor from a combination of molecular proper- ties. The function of this circuitry is poorly understood, in spite of a substantial literature on the electrophys- iology of the mammalian olfactory bulb during odor stimulation, because of the difficulty in specifying the distribution of receptor cells activated by an individual odor and in specifying the precise distribution of mi- tralitufted cells that will be directly activated by those receptor cells. For this and other reasons, it has been useful to activate these circuits by electrical stimula- tion of the olfactory nerve or the lateral olfactory tract to study certain aspects of the system. In addition there are several recent papers that have approached the is- sue of spatial factors in odor processing by recording at several sites in the bulb (Buonviso and Chaput, 1990; Meredith, 1986; Wilson and Leon, 1987) or have pro- vided important information about the synaptic pro- cesses evoked by odor stimuli (Hamilton and Kauer, 1989; Wellis et al., 1989; Wellis and Scott, 1990).
Organization of the Mitral Cell Connections With Glomeruli
Each glomerulus in the rat is connected with an av- erage of 25 mitral cells and 50 tufted cells (Meisami and Safari, 1981). Since the glomerulus probably re- sponds as a unit, it is useful to know whether this uni- tary behavior is conserved in the response of the mi- tral/tufted cells that innervate that glomerulus. A recent experiment comparing odor responses in pairs of mitral cells, simultaneous recorded with small ( < 40
Fm) and large (150-200 pm) electrode separations, showed more similar response patterns for the close recordings than the distant ones (Buonviso and Chaput, 1990). The close pairs of recordings are consis- tent with the cells being connected with the same glo- merulus, while the distant recordings were substan- tially further apart than the average size of a rat glomerulus (approximately 130 pm). Anatomical stud- ies with small injections of horseradish peroxidase to label populations of mitral cells have shown that the majority of cells that are as close together as this 130 Fm distance project into the same or adjacent glomer- uli (Buonviso et al., 1991a). One might expect that strong lateral inhibitory interaction between mitral cells making contacts with the same granule cell pop- ulation could produce a situation where the most strongly activated cell entering the glomerulus would inhibit all its close neighbors. This type of behavior is seen in some network models (Ambros-Ingerson et al., 1990; Shamma, 1989). Such dominance by one cell in a local activated region may be prevented by a strong self-inhibition of the mitral cells (Shepherd, 1981). An- other mechanism that could locally suppress lateral inhibition could be the arrangement of synapses on mi- tral cell basal dendrites. The greatest density of con- nections from granule cells appears to be at interme- diate distances between the soma and the tips of the basal dendrites (Mori, 1987b), so that the strongest lat- eral inhibition may not be on immediate neighbors.
Lateral Inhibitory Interactions Inhibitory postsynaptic potentials (IPSPs) are
evoked in mitralitufted cells during odor stimulation, and these inhibitory mechanisms can act over substan- tial distances in the bulb to provide for interactions between inputs several hundred micrometers apart. In- tracellular recordings of mitralitufted cells during odor responses were first reported from salamander olfac- tory bulb (Hamilton and Kauer, 1989). These record- ings demonstrated hyperpolarizations underlying pauses in spiking, providing strong evidence for the role of inhibition in shaping the responses of these cells. Recordings from rat mitralitufted cells have re- peated this observation (Wellis, 1990) and added the observation that many of these cells show increased membrane conductance during the hyperpolarizations, demonstrating that most periods of hyperpolarization do not result from disfacilitation. Figure 3 and Table 1 illustrate these results.
Recordings from multiple sites in the same bulb have been used to test the expectation that lateral inhibition would produce differences between the odor responses of mitral cells separated by distances that are close enough to involve interactions with the same popula- tion of granule cells (Meredith, 1986; Wilson and Leon, 1987). Both of these papers indicate that cells recorded at distances of 300-500 pm tend to show complemen- tary patterns of response to the same odor, as would be expected from a strong lateral inhibitory influence. These distances are within the range of the long basal dendrites of mitral cells and are large relative to the size of a glomerulus, implying that the olfactory nerve inputs are different. Intracellular recordings of re-
A -06 mV
..... ......I 4
...
i -75mV AA0.2 i 1 i
.... ..... ..... 4
...... ...... 4
Spontaneous Aver age D 130 (4 snif fs)
....................
F Current ( n N
1 5 - 1.0 -0.5 I
/'
I I I
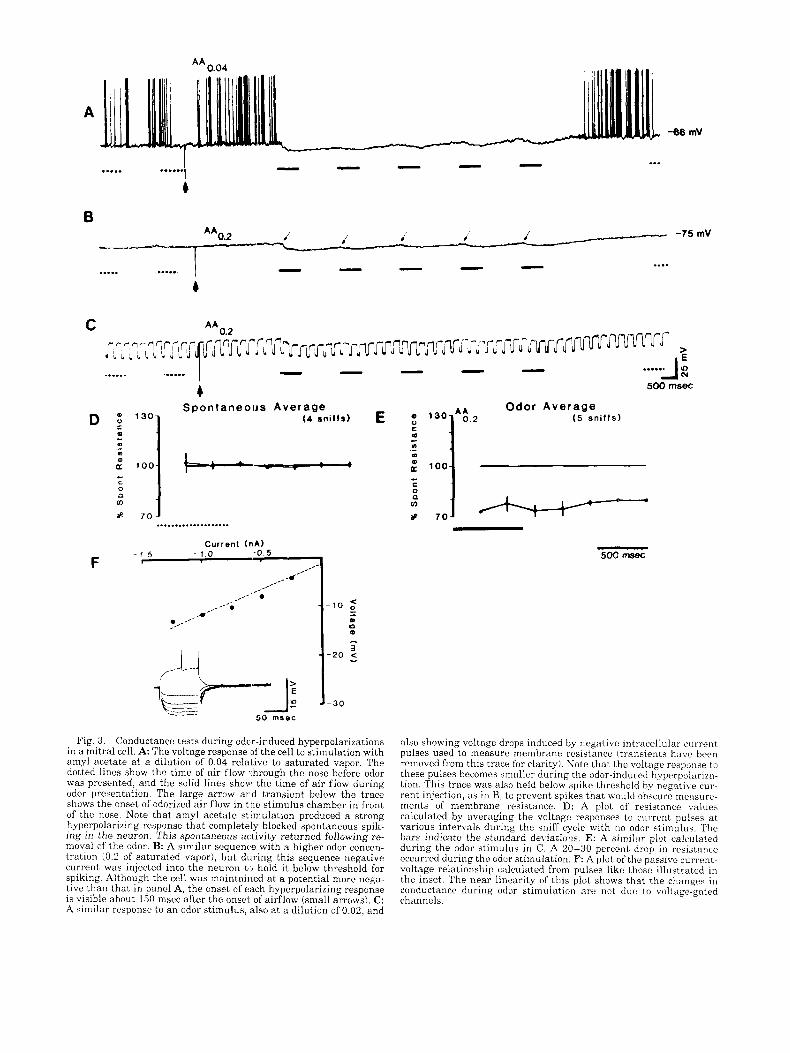
Fig. 3 . Conductance tests during odcr-induced hyperpolarizations in a mitral cell. A: The voltage response If the cell to stimulation with amyl acetate at a dilution of 0.04 relative to saturated vapor. The dotted lines show the time of a i r flow 1,hrough the nose before odor was presented, and the solid lines show the time of a i r flow during odor presentation. The large arrow a r d transient below the trace shows the onset of odorized air flow in the stimulus chamber in front of the nose. Note that amyl acetate stimulation produced a strong hyperpolarizing response tha t completely blocked spontaneous spik- ing in the neuron. This spontaneous activity returned following re- moval of the odor. B: A similar sequenc'e with a higher odor concen- tration (0.2 of saturated vapor), but during this sequence negative current was injected into the neuron t J hold i t below threshold for spiking. Although the cell was maintained at a potential more nega- tive than that in panel A, the onset of each hyperpolarizing response is visible about 150 msec after the onsel. of airflow (small arrows). C: A similar response t o a n odor stimulus, also at a dilution of 0.02, and
.....A 2 500 rnsec
Odor Average ( 5 sn i f fs )
; 100
- 500 ms8c
also showing voltage drops induced by negative intracellular current pulses used to measure membrane resistance (transients h a w been removed from this trace for clarity!. Note that the voltage response t o these pulses becomes smaller during the odor-induced hyperpolariza- tion. This trace was also held below spike threshold by negative cur- rent injection, as in B, to prevent spikes that would obscure me;isurr- ments of membrane resistance. D: A plot of resistance values calculated by averaging the voltage responses t o current pulses a t various intervals during the sniff cycle with no odor stimulus. The bars indicate the standard deviations. E: A similar plot calculated during the odor stimulus in C. A 20-30 percent drop in resistance occurred during the odor stimulation. F: A plot of the passive current- voltage relationship calculated from pulses like those illustrated in the inset. The near linearity of this plot shows that the changes in conductance during odor stimulation are not due to voltage-gated channels.

149 MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION
TABLE 1. Summary of membrane impedance changes during odor-induced potentials' ~~
Resistance Em R M n change h V ) IMR) Eodor
Odor-induced potential Depolarization
Decreased resistance 9 31.1 2 11.5% -70.8 t 10 24.8 2 10.2 17.6 f 8.3 5.1 -+ 2.7
Increased resistance 9 19.1 c 10.2% -71.3 f 9.6 20.0 2 10 24.2 f 12.9 4.9 2 2.9
No change in resistance 3 Less than 10% -93.3 t 11.5 30.1 2 0.9 27.0 f 0.9 12.3 -r 2.7
Range 16-48%
Range 11-4270
Hyperpolarization Decreased resistance 7 28.5 f 11.6% -66.9 ? 7.3 17.7 lr 8.8 12.4 I 7.3 -6.9 2 2.9
Range 18-50%
Range 20-29% Increased resistance 2 24.5 c 6.4 -68.0 2 4.2 30.6 36.7 -4.7 f 1.1
No change in resistance 1 Less than 10% - 75 27.7 25.7 - 1.8 ~~
'n = number of tests (different neurons or odors showing different response patterns for the same odor); R = resistance; Em = resting membrane potential, R,% = resting membrane resistance; Rod,,, = resistance during the odor response; E,,,, = change in membrane potential during the odor response All measurement; are mean 2 standard deviation.
sponses to electrical stimulation of the olfactory nerve layer show that sites on the nerve layer as far apart as 1,000 pm often produce IPSPs in the same mitral or tufted cell (Fig. 41, sometimes even in the absence of an action potential (Fig. 5). This indicates that the hyper- polarization was synaptically produced and not simply a result of intrinsic membrane properties. The excita- tory effects of such nerve stimulation are generally more localized, especially in mitral cells (Schneider and Scott, 1983; Wellis and Scott, 1990). Recordings of field potential responses to electrical stimulation of the nerve layer in cat led to similar conclusions. In that case the average region of inhibition in mitralitufted cells was calculated to extend 569 p.m beyond the av- erage regions of excitation in those cells (Freeman, 1974a).
Interneuron Actions During Odor Responses The interneurons of the mammalian olfactory bulb
have been well described morphologically but have been difficult to study physiologically because of their small size. The oscillatory behavior of the olfactory bulb EEG during odor responses has long been attrib- uted to currents in the granule cells (Freeman, 1975; Scott and Aaron, 1977; Shepherd, 1972). Because the granule cell interactions with mitral cell basal den- drites occur at a reciprocal synapse in an isolated gem- mule, it has been speculated that the granule cells might not require an action potential for feedback in- hibition of mitral cells (Shepherd, 19811, even though these cells fire action potentials to electrical stimula- tion (Mori, 1987). Recent intracellular recordings from granule cells show that many of them do spike during odor responses, while others failed to spike in spite of substantial EPSPs (Wellis and Scott, 1990). Marking of active granule cells by cfos expression shows restric- tion to a region near activated glomeruli (Guthrie et al., 1993).
The significance of spikes in granule cells can be appreciated by considering the inhibitory effect pro- duced by a single granule cell following a single mitral cell spike. Modeling of granule cells, based on the as- sumption of passive membrane properties, indicates
that the depolarization should be substantially greater in the gemmule contacted by the activated mitral cell (Shepherd, 1981). Therefore more transmitter should be released at that gemmule and the inhibitory effect should be greater on the activated mitral cell (feedback inhibition) than on the neighboring mitral cells (lat- eral inhibition). A spike in that granule cell would, in contrast, lead to more widespread inhibitory effects on the total population of cells contacted by that granule cell. These effects have been further discussed in the recent report by Woolf et al. (1991). The presence of 2 types of granule cell response, either resulting from intrinsic differences between superficial and deep cells and/or resulting from differences in control by other neurons, could have substantial effects on the role of the granule cells in lateral inhibition.
The periglomerular cells have also been difficult to identify and observe. Extracellular recordings with paired pulse stimulation have suggested that these cells have inhibitory effects, probably both within a single glomerulus via the dendrodendritic synapse and between glomeruli via their axons (Getchell and Shep- herd, 1975; Shepherd, 1971). Modeling and poststimu- lus time histogram analysis in the cat and rabbit have suggested that some population of cells in the peri- glomerular region is mutually excitatory (Freeman, 1975). While these 2 results appear contradictory, it is not entirely certain that they involved the same group of cells.
While recent recordings from neonatal rabbit olfac- tory slices have suggested that periglomerular cells do not spike (Bufler et al., 1992a,b), intracellular record- ings and marks have been made from intact adult rats of cells tentatively identified as periglomerular cells (Wellis and Scott, 1990). These recordings suggest that periglomerular cells receive a very wide spatial conver- gence of excitatory and inhibitory inputs. This conver- gence is greater than the dispersion seen in the pri- mary olfactory nerve fibers on the part of the olfactory nerve layer stimulated (Wellis and Scott, 1991) and suggests some excitatory interconnection between cells of the periglomerular region. Extracellular recordings from animals inspiring clean air showed a population

150 J.W. SCOTT ET AL.
DORSAL
NE3
NE4 I)
!I1 i NE5 I)
N7- - NE6 I)
VENTRAL
Fig. 4. A biocytin-filled superficial tufted cell and its responses to olfactory nerve stimulation. The intracellular records shown on the left are the responses to nerve stimulati,m between the points (arrows labeled NE3-NE6) indicated to the left of the glomerular layer illus- trated on the right. This cell had a resting potential of -70 mV. The small spike (about 50 mV) is probably d i e to the recording site, which was apparently in a basal dendrite. The second and third spikes (when
present) were smaller, apparently due to shunting by synaptic con- ductances. Note that both excitatory and inhibitory post synaptic po- tentials were evoked by nerve stimuli applied dorsally or ventrally to the apical dendrites. The field of excitatory and inhibitory responses to this cell was asymmetrical (as were the basal dendrites), suggest- ing that the basal dendritic field was important in modulating t h c excitability of the cell.
of superficial cells that had relatively simple excitatory responses in phase with the inspiration (Onoda and Mori, 1980), as distinguished from the more compli- cated temporal patterns of deeper cells. These superfi- cia1 cells are likely to have been periglomerular cells. Similar odor responses were s e m in a small number of intracellularly recorded cells 0'" the periglomerular re- gion (Wellis and Scott, 1990).
WHAT COMPUTATIONS ARE MADE IN THE OLFACTORY BULB CIRCUITRY?
The output cells are not easily activated by focal elec- trical stimuli applied to the olfactory nerve layer. It has been repeatedly observed that (electrical stimula- tion of the olfactory nerve evokes very few spikes from mitralitufted cells because a powerful recurrent inhi-

MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION 151
A
4
Fig. 5. A mitral cell, showing potent odor responses and for which the response to nerve stimulation was confined to a single spike evoked by only 1 of the 7 olfactory nerve electrode sites. A The re- sponse to the effective nerve electrode site (N5) and the 2 adjacent stimulating sites. The stimulus intensity was maximal for our sys- tem, 1 mAmp for 0.1 msec. Note that both adjacent electrodes evoke large IPSPs, even though they do not produce a spike. This is evidence of lateral inhibition. Electrode 6 evoked an early brief hyperpolariza- tion (arrow) that preceded the spike response. B: Responses of this cell to 3 odors. The response pattern to amyl acetate (AA) shows a brief
bition quickly hyperpolarizes the cells below their fir- ing threshold (Shepherd, 1963). Subsequent work has demonstrated that mitral cells are less responsive to single pulse stimulation of olfactory nerve than are tufted cells (Schneider and Scott, 1983). In this study, about 40 percent of mitral cells fail to respond to focal stimulation and cannot be driven to fire by manipulat- ing the stimulus intensity or stimulus rate.
This result has been supported by further studies in this laboratory (Wellis and Scott, 1990; Wellis et al., 1989). Recent evidence shows that many of these mi- tral cells, unresponsive to olfactory nerve stimulation, are the mitral cells, with basal dendrites confined to the deepest portion of the mitral cell layer (Ezeh and Scott, 1992). Nevertheless, even the mitral cells with more superficial basal dendrites (cf. Orona et al., 1984, for a description of these types) show only one or at most 2 spikes to a n electrical stimulus. Some mitral cells that fail entirely to respond to electrical stimula- tion respond vigorously to odor stimulation (Wellis et al., 1989). Figure 5 shows a n example of a mitral cell with a very poor olfactory nerve response and a strong odor response. It is possible that strong odor responses by mitral cells depend upon the release of tonic inhibi- tion by either granule cells or periglomerular cells.
hyperpolarization soon after the onset of air flow, followed by a de- polarization with spiking and a return to baseline after air flow ceased. A strong hyperpolarization followed depolarization in the re- sponse to ethyl butyrate (EB). Limonene (LIM), in contrast, produced a prolonged hyperpolarization, most evident during the first sniff. This response habituated but is still evident during subsequent sniffs. High rates of spiking occurred at times corresponding to the hypolar- ization in the AA and EB responses. The large arrows below the odor traces indicate the start of odor flow in the system, and the bars under the traces indicate the time of inspiration through the rat's nose.
This disinhibition could occur because the odor acti- vated several patches of mitralitufted cells at an appro- priate distance apart. The effect of disinhibition would occur because the activity of these cells would inhibit intervening mitralitufted cells and disfacilitate inter- neurons driven by those neurons. This postulated dis- inhibition has not been demonstrated. Our observa- tions of changes in resistance during odor responses have shown some cases where membrane conductance was decreased during depolarization, as would be ex- pected if tonic inhibitory synapses were turned off (Table 1). On the other hand, we were able to find only 1 granule cell (out of 16 that responded to odor) with spontaneous activity that was decreased during odor stimulation. Therefore, we have yet to conclusively confirm the hypothesis that mammalian mitralitufted cells produce their maximum response only to a specific spatial pattern of input.
Observations from amphibians raise some cautions about the existence of these spatially based lateral in- teractions involving mitral cells. Kauer and Moulton (1974) were able to verify that the same stimulus odor presented in different parts of the salamander olfactory mucosa could evoke different sequences of spikes from a single olfactory bulb neuron, but the responses t o

152 J.W. SCOTT ET AL.
local stimulation often had essentially the same form as the response to stimulation 0 ” the total mucosa. In neither this study nor a more recent study using local- ized electrical stimulation of the frog mucosa (Jiang and Holley, 1992) was it possible to convert a mitral cell from a predominately inhibitory to a predomi- nately excitatory response by moving the stimulus. It is possible that these failures t 3 observe antagonistic responses from stimulation of different sites on the olfactory mucosa may reflect the rearrangement of axons as they enter the olfactory bulb. (We have by- passed this problem by directly stimulating the olfac- tory nerve layer of the bulb.) In addition, direct com- parisons between mammals and amphibians are difficult because the olfactory bulb structure is quite different (for example, amphib (an mitral cells have multiple apical dendritic processes that enter several glomeruli (Herrick, 1931). Certainly, analogous exper- iments with localized stimulatim of the mammalian mucosa would be useful for testing the question of how the optimal inputs to mitralitufted cells are orga- nized.
Inhibitory processing within 1 he olfactory bulb also produces different temporal sequences of spikes evoked in response to different odors. It had been observed previously that a single olfactclry bulb neuron could show a t least 2 patterns of spiki ig depending upon the odor stimulus (Hamilton and Kauer, 1989; Kauer, 1974; Macrides and Chorover, 1972). Harrison and Scott (1986) had cautioned the necessity to control for intensity dependence of the spike pattern, since some mitral and tufted cells show dramatic changes in spike pattern with changes in odor s.,imulus concentration. Wellis et al. (1989) showed by comparison of several odors over a series of concentration steps that some mitralitufted cells show diverging patterns for pairs of odors at all suprathreshold odors tested. These pattern differences were not specific to the odor (i.e., one cell might show excitation to amyl acetate and inhibition to ethyl butyrate, while a second cell could show the op- posite pattern), and not all cdls showed the same groupings of odor differences. These differential pat- terns of mitralitufted cell response are likely to be dependent upon the distribution of olfactory nerve af- ferents influencing neighboring interneuron popula- tions as well as the afferents reaching the glomerulus for the recorded neurons. I t is not demonstrated that these temporal patterns are used in coding, although there are indications that the 1,emporal patterns con- tain more information than a simple spike count (Har- rison and Scott, 1986; Scott, 1990). Similar observa- tions have been made in the visual system (Cattaneo et al., 1981; Richmond and Optican, 1987). Certainly some recordings in alert animals show temporally pat- terned odor responses (Chapd and Holley, 1980; Chaput and Panhuber, 19821, indicating that temporal pattern effects seen in anesthelized preparations may also have significance in unanesthetized animals. Cer- tain observations have suggestcd a synchronization of activity of the olfactory bulb ,md hippocampus with sniffing that could serve as a time base for analysis of temporally coded informal ion (Macrides et al., 1982).
GENERAL CONCLUSIONS The Significance of Spatial Organization in
Olfactory Processing
One issue that has surfaced in anatomical and phys- iological studies of the olfactory bulb is the extent to which the projection pattern of receptor cell axons is spatially organized and whether that ;spatial organiza- tion has meaning for olfactory sensory analysis. There are accumulating data, reviewed above, that the pri- mary projection may have topographic and nontopo- graphic components. The issue of strict topography in the primary projection is separate from the issue of whether the spatial interactions within the olfactory bulb are significant for coding. The fact that the pri- mary axons in a substantial portion of the olfactory nerve layer are spatially organized, the systematic dif- ferences in excitability of the morphologically different groups of mitralitufted cells, the physiological data in- dicating spatial restrictions on interactions between cells when activated by odors, and t h e anatomical ar- rangements indicating that granule cells mediate lat- eral inhibitory effects all suggest that there is an im- portant spatially organized, local interaction between bulb neurons. Recent modeling efforts attempt t o ac- count for these lateral interactions in the bulb (White et al., 1992) and, in some cases, have tried to take the different properties of mitral and tufted cells into ac- count (Ant6n et al., 1991).
The nontopographic arrangement of many of the af- ferents from the receptor sheet to the glomeruli creates a problem for the study of the mechanisms in the bulb involved in odor sensory information processing. If a completely precise topographic arrangement of these primary axons existed, it would be possible to specify the afferents reaching a mitral cell simply by knowing its position within the bulb, as one could for a cell in the lateral geniculate. Then, knowing the odor spectra of the afferent receptor cells, one could describe the changes in the receptive fields of olfactory bulb and cortex cells relative to the receptive fields of receptor cells. This specification seems very unlikely for olfac- tion, partly because of the discontinuous nature of the projection. Glomeruli appear to receive convergent and divergent projections.
Since the difficulties in specifying the projection onto the glomeruli result not exclusively from technical de- ficiencies, but also from significant discontinuities in the projection (Astic et al., 19871, whi2t approaches can be followed in studying processing in the olfactory bulb? One important method used already by Wilson and Leon (1987) and by Buonviso and Chaput (1990) is a search for spatial order in the way that bulbar neu- rons respond to odors, using that order to probe the synaptic relationships that must exist between the tested cells. Each of these investigators had specific hypotheses about the extent of common excitatory in- puts to mitral cells or the extent of mutual inhibitory influences. Further progress with this type of compar- ison will require more specific measurements of the response pattern (cf. Duchamp-Viret et al., 1990; Har- rison and Scott, 1986; Scott, 1990) as well as more in-

MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION 153
formation about the spatial extent of the various levels of contact between cell types.
OLFACTORY PROCESSING IN THE CORTEX AND ITS RELATION TO PROCESSING
IN THE OLFACTORY BULB We have attempted to develop the idea that the ol-
factory bulb is an important intermediate stage of sen- sory processing that looks for contrasts in the activity arriving in a set of glomeruli. Although the bulb has a well-defined anatomy, its function, and perhaps its structure, seems to be modifiable by experience (Coo- persmith and Leon, 1984; Freeman, 1979). Those who have attempted to make comparisons between re- sponses at various levels of the system have generally concluded that there is improvement in the ability to discriminate between odors at each stage (Duchamp and Sicard, 1984; Giachetti and MacLeod, 1973; Tan- abe et al., 1975). This conclusion does not necessarily imply a “purification” of responses to simple odor re- ceptors, a view rejected above, since the discrimination seen electrophysiologically at any intermediate level need not be identical with the final subjective odor clas- sification.
Interpretation of the role of the olfactory bulb in sen- sory processing also requires consideration of what the more central regions do with the information that leaves the bulb. Recent efforts at modeling the behav- ior of the piriform cortex have been built on the as- sumption that the output from the bulb is coded in an ensemble fashion. This assumption is based on 2 points in the literature, most clearly articulated by Freeman (1975) and by Haberly (1985). These are (1) failure to observe evidence of strict topography in the bulbocor- tical projection and (2) failure to see evidence of specific feature detectors in the olfactory bulb that would indi- cate the type of information processing done in the bulb.
Haberly pointed out that these features, combined with the high degree of interconnectivity between cells of the piriform cortex (Haberly and Bower, 1984) and with the relative lack of effect of restricted cortical de- nervation on olfactory discrimination (Slotnick and Berman, 1980), suggest that processing occurs over a widely distributed region of the olfactory cortex. Re- cent models propose that the piriform cortex works by connectionist principles in which the strength of syn- aptic connections between neurons is modified by ac- tivity (Ambros-Ingerson et al., 1990; Haberly and Bower, 1989). These models will surely hasten the un- derstanding of sensory processing in olfaction. Never- theless, the assumptions about the nature of sensory processing in the olfactory bulb and the spatial order in the bulbocortical projection should be examined with care.
As Price (1987) has pointed out, failure to observe a point-to-point topography in the bulbocortical connec- tions does not imply absence of spatial order in this projection, It is recognized that there is a good topo- graphic organization between the olfactory bulb and the pars externa of the anterior olfactory nucleus (Schoenfeld and Macrides, 1984; Scott et al., 1985). Many efforts to observe a similar topography in the
posterior part of the piriform cortex by simple lesion or transport experiments have had negative results. An attempt t o test spatial order by mapping the position of labeled mitralitufted cells after retrograde transport from cortical injections could account for about 25% of the variance in the projection (Scott et al., 1980). The presence of extensive collaterals on many mitral cell axons (Luskin and Price, 1982; Ojima et al., 1984; Scott, 1981) probably has contributed to the impression of diffuse projection, but there have not been investi- gations of whether neighboring cells or cells of similar type have similar collateralization patterns,
Recent observations on the bulbocortical projection (Buonviso, 1991b) indicate that neighboring mitral cells probably project to closely related terminal re- gions in the piriform cortex. This result raises the pos- sibility that, even without a strict topography, the spa- tially organized analysis conducted in the bulb could be maintained to some degree in the cortical projection. Experiments with 2-deoxyglucose mapping indicate that this organization does not produce a separation of responses to particular odors in identifiable parts of the piriform cortex (Astic and Cattarelli, 1982; Sharp et al., 19771, a further indication against the notion ad- vanced by Le Gros Clark (1951) that there might be isolation of the inputs from functional receptors a t each stage of central processing.
This review has outlined evidence supporting the ex- istence of substantial processing within the circuits of the olfactory bulb that act upon a spatially organized input. The responses of mitralitufted cells are signifi- cantly altered as compared to the responses of receptor cells. The existence of neurons showing different re- sponse patterns to different odors is one example. The strong temporal patterning that is seen in olfactory bulb mitralitufted cells, and the changes in temporal patterns of response to different odors, suggest a tem- poral mechanism in coding.
It has been pointed out that stimulation of the olfac- tory tract evokes a sequence of activation of the piri- form cortex in which the primary and association in- puts have a temporal separation that depends upon the position in the cortex. This spatiotemporal pattern of electrical activity might indicate a system tuned to pro- cess temporally coded information from the olfactory bulb (Ketchum and Haberly, 1991). On the other hand, behavioral studies using direct electrical stimulation through an array of electrodes in the olfactory bulb as a training cue have argued against a temporal code. Rats could learn a response to a stimulus train pre- sented during either inspiration or expiration, but they could not learn a response based on the timing of that stimulus train relative t o the inspiration cycle (Monod et al., 1989). This experiment suggests that the tempo- ral patterning in bulb responses is not used to code information in the cortex but, instead, reflects the mechanism controlling the spatial distribution of neu- ral activity.
These observations imply that the processing that occurs at the piriform cortex and other central regions must act on an input that is significantly modified as compared to the output of the receptor sheet. While it is important that there be better understanding of the

154 J.W. SCOTT ET AL.
actions of the olfactory bulb circuits so that these can be included in the cortical models, it is also important to consider the possible actions of the cortex in assess- ing the meaning of observations on the bulb circuits.
ACKNOWLEDGMENTS The work from our laboratory described in this re-
view was supported by NIH grant DC00113. The au- thors gratefully acknowledge the assistance of Brad- ford Priddy in preparation of the figures.
REFERENCES Adrian, E.D. (1950) Sensory discrimination with some recent evi-
dence from the olfactory organ. Br. Ned. Bull., 6:330-333. Allison, A.C. (1953) The morphology o ’ the olfactory system i n the
vertebrates. Biol. Rev., 28:195-244. Ambros-Ingerson, J., Granger, R., and Lynch, G. (1990) Simulation of
paleocortex performs hierarchal clustering. Science, 247:1344- 1348.
Amoore, J .E . , Johnston, J.W., and Rubin, M. (1964) The stereochem- ical theory of odor. Sci. Am. 2120:42- 49.
Antbn, P.S., Lynch, G., and Granger, € I . (1991) Computation of fre- quency-to-spatial transform by olfactory bulb glomeruli. Biol. Cy- bernetics, 65:407-414.
Astic, L., and Cattarelli, M. (1982) Metabolic mapping of functional activity in the rat olfactory system after a bilateral transection of the lateral olfactory tract. Brain Res
Astic, L,, Saucier, D., and Holley, A. ( ships between olfactory receptor cells and glomerular foci in the ra t olfactory bulb. Brain Res., 424:144-152.
Baker, 13. (1988) Neurotransmitter p1a:;ticity in the juxtaglomerular cells of the olfactory bulb. In: Molecu a r Neurobiology of the Olfac- tory System. F.L. Margolis and T.V. Getchell, eds. Plenum Press, New York, pp. 185-216.
Barber, P.C. (1989) Ulex europeus a g g h tinin binds exclusively to pri- mary olfactory axons in the ra t nervous system. Neuroscience, 30: 1-9.
Boeckh, J., Distler, P., Ernst, K.D., Hiisl, M., and Malun, D. (1990) Olfactory bulb and antenna1 lobe. In: Chemosensory Information Processing. D. Schild, ed. NATO AS1 Series. Springer-Verlag, Ber- lin, pp. 201-228.
Buck, L., and Axel, R. (1991) A novel rnultigene family may encode odorant receptors: A molecular basis for odor recognition. Cell, 65: 175-187.
Bufler, J., Zufall, F., Franke, C., and Hatt, H. (1992a) Patch-clamp recordings of spiking and nonspiking: interneurons from rabbit ol- factory bulb slices: GAEA- and other transmitter receptors. J. Comp. Physiol. A, 170:153-159.
Bufler, J., Zufall, F., and Hat t , H. (199213) Patch-clamp recordings of spiking and nonspiking interneurons from rabbit olfactory bulb slices: Membrane properties and ionic currents. J. Comp. Physiol.
Buonviso, N., and Chaput, M.A. (1990) .3esponse similarity to odors in olfactory bulb output cells presumed to be connected to the same glomerulus: Electrophysiological stu i y using simultaneous single- unit recordings. J. Neurophysiol., 63:447-454.
Buonviso, N., Chaput, M.A., and Scott, J.W. (19913) Mitral cell-to- glomerulus connectivity: a n HRP study of the orientation of mitral cell apical dendrites. J. Comp. Neurol., 30757-64.
Buonviso, N., Revial, M.F., and Jourdaa, F. (1991b) The projections of mitral cells from small regions of the olfactory bulb: an anterograde tracing study using PHA-L (Phaseolus uuZgaris leucoagglutinin). Eur. J. Neurosci., 1991:493-501.
Cattaneo, A., Maffei, L., and Morrone, C. (1981) Patterns in the dis- charge of simple and complex visual cortical cells. Proc. R. SOC. Lond. €3, 212:279-297.
Chaput, M., and Holley, A. (1980) Single unit responses of olfactory bulb neurones t o odour presentation in awake rabbits. J. Physiol. (Paris,, ’76:551-558.
Chnput, M.A., and I’anhuber, H. (1982) Effects of long duration odor exposure on the unit activity of o l fxtory bulb cells in awake rab- bits. Brain Res., 250:41-52.
Clancy, A.W., Schoenfeld, T.A., and Macrides, F. (1985) Topographic organization of peripheral input to the hamster main olfactory bulb. Chem. Senses, 10:399.
A, 170:145-152.
Coopersmith, R., and Leon, M. (1984) Enhanced neural response to familiar olfactory cues. Science, 225:849-851..
Davis, B.J., Macrides, F., Youngs, W.M., Schneider, S.P., and Rosene, D.L. (1978) Efferents and centrifugal afferents of the main and accessory olfactory bulbs in the hamster. Brain Res. Bull., 3:59-72.
Duchamp, A, , and Sicard, G . (1984) Odour discrimination by olfactory bulb neurons: Statistical analysis of electrophysiological responses and comparison with odour discrimination by receptor cells. Chem. Senses, 9:l-14.
Duchamp-Viret, P., Duchamp, A, , and Vigourou.x, M. (1990) Temporal aspects of information processing in the first two stages of the frog olfactory system: Influence of stimulus intensity. Chem. Senses, 15:349-365.
Ezeh, P.I., and Scott, J.W. (1992) Morphology and electrophysiology of ra t olfactory bulb mitral and tufted cells. Chem. Senses, in press.
Freeman, W.J. (1974a) Average transmission distance from mitral- tufted to granule cells in olfactory bulb. Electroenceph. Clin. Neu- rophysiol., 36:609-618.
Freeman, W.J. (1974b) Relation of glomerular neuronal activity to glomerular transmission attenuation. Brain Kes., 65:91-107.
Freeman, W.J. (1974~) Topographic organization of primary olfactory nerve in cat and rabbit a s shown by evoked potentials. Electroen- ceph. Clin. Neurophysiol., 36:33-45.
Freeman, W.J. (1975) Mass Action in the Nervous System. Academic Press, New York.
Freeman, W.J. (1979) EEG analysis gives model of neuronal tem- plate-matching mechanism for sensory search with olfactory bulb. Biol. Cybernetics, 35:221-234.
Fujita, S.C., Mori, K.R., Imamura, D., and Obata, K. (1985) Sub- classes of olfactory receptor cells and their segregated central pro- jections demonstrated by a monoclonaY antibody. Brain Res., 326: 192-196.
Getchell, T.V., and Shepherd, G.M. (1975) Short-axon cells in the olfactory bulb: Dendrodendritic aynaptic interactions. J. Physiol. (Lond.), 251523-548.
Giachetti, I., and MacLeod, P. (1973) Superiorite du pouvoir discrim- inateur des cellules mitrales compare a celui des recepteur olfactifs. 3. Physiol. (Paris), 66:399-407.
Greer, C.A. (1987) Golgi analyses of dendritic Organization among denervated olfactory bulb granule cells. J. Comp. Neurol., 257:442- 451.
Greer, C.A., W.B. Stewart, ‘Teicher, M.B., and Shepherd, G.M. (1982) Functional development of the olfactory bulb and a unique glomor- ular complex in the neonatal rat. J. Neurosci., 2:1744-1759.
Guthrie, K.M., Anderson, A.J., Leon, M., and Gall, C. (1993) Odor induced increases in cfos niRNA expression reveal an anatomical unit for odor processing in olfactory bulb. Proc. Natl. Acad. Sci. USA, in press.
Haberly, L.B. (1985) Neuronal circuitry in olfactory cortex: anatomy and functional implications. Chein. Senses, 10:219-238.
Haberly, L.B., and Bower, J .M. (1984) Analysis of association fiber system in piriform cortex with intracellular recording and staining techniques. J. Neurophysiol., 51:90-112.
Haberly, L.B., and Bower, J.M. (1989) Olfactory cortex: model circuit for study of associative memory? Trends Neurosci., 12:258-264.
Haberly, L., and J.L. Price (1977) The ‘axonal projection patterns of the mitral and tufted cells of the olfactory bulb in the rat . Brain Res., 129:152-157.
Halasz, N. , Johansson, O., Hokfelt, T., Ljungdahl, and Goldstein, M. (1981) Immunohistochemical identification of two types of dopa- mine neurons in the r a t olfactory bulb as seen by serial sectioning. J. Neurocytol., 10:251-259.
Hamilton, K.A., and Kauer, J.S. (1989) Patterns of intracellular po- tentials in salamander mitralitufted cells in response to odor stirn- ulation. J. Neurophysiol., 62509-625.
Harrison, T.A., and Scott, J.W. (1986) Olfactory bulb responses to odor stimulation: Analysis of response pattern and intensity relation- ships. J. Neurophysiol., 56:1571-1589.
Hansson, B.S., Ljungberg, H., Hallberg, E., .and Lofstedt, C. (19921 Functional specialization of olfactory glomeruli in a moth. Science, 256:1313-1315.
Herrick, C.J. (1931) The amphibian forebrain. J. Comp. Neurol., 53: 55-69.
Holley, A., and MacLeod, P. (1977) Transduction et codage des infor- mations olfactives chez les vertebres. J. Physiol. (Paris), 73:725- 828.
Hubel, D.H., and Wiesel, T.N. (1962) Receptive fields, binocular in-

teraction and functional architecture in the cat's visual cortex. J. Physiol. (Lond.), 160:106-154.
Hudson, R., and Distel, H. (1987) Regional autonomy in the periph- eral processing of odor signals in newborn rabbits. Brain Res., 421: 85-94.
Jacobson, I., Butcher, S., and Hamburger, A. (1986) An analysis of the effects of excitatory amino acid receptor antagonists on evoked field potentials in the olfactory bulb. Neuroscience, 19267-273.
Jahr, C.E., and Nicoll, R.A. (1982) An intracellular analysis of den- drodendritic inhibition in the turtle in uitro olfactory bulb. J. Phys- iol. (Lond.), 326:213-234.
Jastreboff, P.J., Pedersen, P.E., Greer, C.A., Stewart, W.B., Kauer, J.S., Benson, T.E., and Shepherd, G.M. (1984) Specific olfactory receptor populations projecting to identified glomeruli in the rat olfactory bulb. Proc. Natl. Acad. Sci. USA, 81:5250-5254.
Jiang, T., and Holley, A. (1992) Some properties of receptive fields of olfactory mitralitufted cells in the frog. J. Neurophysiol., 68:726- 722 . --.
Jourdan, F., Duveau, A., Astic, L., and Holley, A. (1980) Spatial dis- tribution of I4C 2-deoxyglucose uptake in the olfactory bulbs of rats stimulated with two different odours. Brain Res., 188:139-154.
Kauer, J.S. (1974) Response patterns of amphibian olfactory bulb neurones to odour stimulation. J. Physiol. (Lond.), 2433395-715.
Kauer, J.S. (1987) Coding in the olfactory system. In: Neurobiology of Taste and Smell. T.E. Finger and W.L. Silver eds., John Wiley, New York.
Kauer, J.S., and Moulton, D.G. (1974) Responses of olfactory bulb neurones to odour stimulation of small nasal areas in the sala- mander. J. Physiol. (Lond.), 243:717-737.
Ketchum, K.L., and L.B. Haberly (1991) Fast oscillations and disper- sive propagation in olfactory cortex and other cortical areas: a func- tional hypothesis. In: Olfaction as a Model System for Computa- tional Neuroscience. J . Davis and H. Eichenbaum, eds. MIT Press, Cambridge.
Key, B., and Giorgi, P.P. (1986) Soybean agglutinin binding to the olfactory systems of rat and mouse. Neurosci. Lett., 69:131-136.
Khayari, A., Math, F., and Davrainville, J.-L. (1988) Evidence for an absence of K f spread in the glomerular layer of the rat olfactory bulb. Neurosci. Lett., 93:56-60.
Kuffler, S.W. (1953) Discharge patterns and functional organization of mammalian retina. J . Neurophysiol., 16:37-68.
Lancet, D., Greer, D.A., Kauer, J.S., and Shepherd, G.M. (1982) Map- ping of odor related neuronal activity in the olfactory bulb by high- resolution 2-deoxyglucose autoradiography. Proc. Natl. Acad. Sci.
Land, L.J. (1973) Localized projection of olfactory nerves to rabbit olfactory bulb. Brain Res., 63:153-166.
Le Gros Clark, W.E. (1951) The projection of the olfactory epithelium on the olfactory bulb in the rabbit. J . Neurol. Neurosurg. Psychiat.,
Leveteau, J., and MacLeod, P. (1966) La discrimination des odeurs par les glomBrules olfactifs du lapin (etude Blectrophysiologique) J. Physiol. (Paris), 58:717-729.
Luskin, M.B., and Price, J.L. (1982) The distribution of axon collat- erals from the olfactory bulb and the nucleus of the horizontal limb of the diagonal band to the olfactory cortex, demonstrated by double retrograde labeling techniques. J. Comp. Neurol., 209:249-263.
Luskin, M.B., and Price, J.L. (1983) The topographic organization of association fibers of the olfactory system in the rat, including cen- trifugal fibers to the olfactory bulb. J. Comp. Neurol., 216:264-291.
Macrides, F., and Chorover, S.L. (1972) Olfactory bulb activity corre- lated with inhalation cycles and odor quality. Science, 175:84-87.
Macrides, F., and Schneider, S.P. (1982) Laminar organization of mi- tral and tufted cells in the main olfactory bulb of the adult hamster. J . Comp. Neurol., 208:419-430.
Macrides, F., Eichenbaum, H.B., and Forbes, W.B. (1982) Temporal relationship between sniffing and the limbic 0 rhythm during odor discrimination reversal learning. J. Neurosci., 21705-1717.
Meisami, E., and Safari, L. (1981) A quantitative study of the effects of early unilateral olfactory deprivation on the number and distri- bution of mitral and tufted cells and of glomeruli in the rat olfactory bulb. Brain Res., 221:81-107.
Meredith, M. (1986) Patterned response to odor in mammalian olfac- tory bulb: The influence of intensity. J . Neurophysiol., 56:572-597.
Monod, B., Mouly, A.M., Vigouroux, M., and Holley, A. (1989) An investigation of some temporal aspects of olfactory coding with the model of multi-site electrical stimulation of the olfactory bulb in the
USA, 79~670-674.
14:1-10.
Mori, K. (1987a) Monoclonal antibodies (2C5 and 4C9) against lacto- series carbohydrates identify subsets of olfactory and vomeronasal receptor cells and their axons in the rabbit. Brain Res., 408:215- 221.
Mori, K. (1987bj Membrane and synaptic properties of identified neu- rons in the olfactory bulb. Prog. Neurobiol., 29975-320.
Mori, K., Kishi, K., and Ojima, H. (1983) Distribution of dendrites of mitral, tufted, and granule cells in the rabbit olfactory bulb. J. Comp. Neurol,, 2 19: 339 -355.
Mori, K., Mataga, N., and Imamura, K. (1992) Differential specifici- ties of single mitral cells in rabbit olfactory bulb for a homologous series of fatty acid odor molecules. J. Neurophysiol., 67:786-789.
Mountcastle, V.B. (1974) Medical Physiology, Vol. 1, 13th ed. C.V. Mosby, St. Louis, pp. 307-411.
Mouradian, L.E., and Scott, J.W. (1988) Cytochrome oxidase staining marks dendritic zones of the rat olfactory bulb external plexiform layer. J. Comp. Neurol., 271:507-518.
Mugnaini, E., Oertel, W.H., and Wouterlood, F.F. (1984) Immunocy- tochemical localization of GABA neurons and dopamine neurons in the rat main and accessory olfactory bulbs. Neurosci. Lett., 47:221- 226.
Nicoll, R.A. (1971) Pharmacological evidence for GABA as the trans- mitter in granule cell inhibition in the olfactory bulb. Brain Res., 35:137-149.
Ojima, H., Mori, K., and Kishi, K. (1984) The trajectory of mitral cell axons in the rabbit olfactory cortex revealed by intracellular HRP injection. J. Comp. Neurol., 230:77-87.
Onoda, N., and Mori, K. (1980) Depth distribution of temporal firing patterns in olfactory bulb related to air-intake cycles. J. Neuro- physiol., 4429-39.
Orona, E., Rainer, E.C., and Scott, J.W. (1984) Dendritic and axonal organization of mitral and tufted cells in the rat olfactory bulb. J. Comp. Neurol., 217227-237.
Orona, E., Scott, J.W., and Rainer, E.C. (1983) Different granule cell populations innervate superficial and deep regions of the external plexiform layer in rat olfactory bulb. J. Comp. Neurol., 217227- 237.
Pinching, A.J., and Powell, T.P.S. (1971) The neuron types of the glomerular layer of the olfactory bulb. J. Cell. Sci., 9:305-345.
Price, J .L. (1987) The central olfactory and accessory olfactory sys- tems. In: Neurobiology of Taste and Smell. T.E. Finger and W.L. Silver. eds. John Wilev and Sons. New York. DD. 179-203.
Price, J.L., and Powell, '?.P.S. (1970) The synaptdlogy of the granule cells of the olfactory bulb. J. Cell Sci., 7:125-155.
Rall, W., Shepherd, G.M., Reese, T.S., and Brightman, M.W. (1966) Dendro-dendritic synaptic pathway for inhibition in the olfactory bulb. Exp. Neural.; 14:44-56.
Richmond, B.J., and Optican, L.M. (1987) Temporal encoding of two- dimensional patterns by single units in primate inferior temporal cortex. 11. Quantification of response waveform. J. Neurophysiol., 57: 147-161.
Riggott, M.J., and Scott, J.W. (1989) Specific lectin labeling of rat olfactory bulb glomeruli. Chem. Senses, 14:740-741.
Rodrigues, V. (1988) Spatial coding of olfactory information in the antenna1 lobe of Drosophila melanogaster. Brain Res., 453: 299-307.
Schild, D. (1988) Principles of odor coding and a neural network for odor discrimination. Biophys. J., 54: 1001-1011.
Schneider, S.P., and Macrides, F. (1978) Laminar distributions of in- terneurons in the main olfactory bulb of the adult hamster. Brain Res. Bull., 3:73-82.
Schneider, S.P., and Scott, J.W. (1983) Orthodromic response proper- ties of rat olfactory bulb mitral and tufted cells correlate with their projection patterns. J. Neurophysiol., 50:358-378.
Schoenfeld, T.A., and Macrides, F. (1984) Topographic organization of connections between the main olfactory bulb and pars externa of the anterior olfactory nucleus in the hamster. J. Comp. Neurol., 227:121-135.
Schoenfeld, T.A., Marchand, J.E., and Macrides, F. (1985) Topo- graphic organization of tufted cell axonal projections in the hamster main olfactory bulb: An intrabulbar association system. J. Comp. Neurol., 235:503 -5 18.
Schwob, J .E. , and Gottleib, D.I. (1986) The primary olfactory projec- tion has two chemically distinct zones. J. Neurosci., 6:3393-3404.
Scott, J.W. (1981) Electrophysiological identification of mitral and tufted cells and distribution of their axons in the olfactory system of the rat. J. Neurouhvsiol.. 46:918-931.
MAIN OLFACTORY BULB FUNCTIONAL ORGANIZATION 155
' " I
rat. Behav. Brain. Res., 33:51-63. Scott, J.W. (1990) Extracting information from spike trains of olfac-

156 J.W. SCOTT ET AL.
tory bulb neurons. In: Chemosensory Information Processing. D. tion of propionic acid vapor by rats with lesions of olfactory bulb Schild, ed. NATO AS1 Series. Springer-Verlag, Berlin, pp, 277-290. areas associated with high 2-DG uptake. Brain Res., 417:343-346.
Scott, J.W., and Aaron, D.D. (1977) Averaged induced waves in the Stewart, W.B., and Touloukian, C.E. (1990) Lectins label subpopula- olfactory bulb of the rat during odor stimulation. Exp. Neurol., 55: tions of olfactory receptor cells. Neurosci. Abjjtr., 16:103. 654-665. Stewart, W.B., Kauer, J.S., and Shepherd, G.M. (1979) Functional
Scott, J.W., and Harrison, T.A. (1987) The olfactory bulb: anatomy organization of ra t olfactory bulb a n a b e d by the 2-deoxyglucose and physiology. In: Neurobiology of Taste and Smell. T.E. Finger method. J. Comp. Neurol., 185:715-734. and W.L. Silver, eds. John Wiley and Sons, New York, pp. 151-178. Switzer, R.C., and Johnson, J.I. (1977) Absence of mitral cells in
Scott, J.w., McBride, R,L,, and Schneidc.r, s,p. (1980) The organiza- monolayer in monotremes. Variations in vertebrate olfactory bulbs. tion of projections from the olfactory b d b to the piriform cortex and Acta Anat., 99:36-42. olfactory tubercle in the rat . J . Comp. Neurol., 194:519-534. Tanabe, T., Iino, M., and Takagi, S.F. (1975) Discrimination of odors
Scott, J.W., Rainer, E.C., Pemberton, J.L., Orena, E., and Mouradian, in olfactory bulb, pyriform-amygdaloid areas, and orbitofrontal cor- L.E. (1985) Pattern of rat olfactory bulb mitral and tufted cell con- tex of the nections to the anterior olfactory bulb pars externa. J. Comp. Neu- RombleY, p.&,, and Westbrook, G.L. (1990) Excitatory synaptic rol., 242:415-424. transmission in cultures of rat olfactory bulb. J. Neurophysiol., 64:
Shamma, S. (1989) Spatial and temporal processing in central audi- 598-606. tory networks. In: Methods in Neuronal Modeling. C. Koch and 1. Wahle, P., Sanides-Kohlrausch, c., Meyer, G., and Lubke, J. (1990) Segev, eds. MIT Press, Cambridge, MA, pp. 247-290. Substance P- and opioid-immunoreactive structures in olfactory
Sharp, F.R., Kauer, J.S., and Shepherd, G.M. (1977) Laminar analysis centers the cat: pattern and postnatal J. of 2-deoxyglucose uptake in olfactory bulb and olfactory cortex of 'Omp. Neurol., 302:349-369.
Wellis, D.P. (1990) Intracellular Studies of Olfactory Bulb Interneu- rons. Doctoral dissertation, Emory University, Atlanta, GA. rabbit and rat. J. Neurophysiol., 40:8(10-813.
volleys in the rabbit. J . Physiol. (Lontl.), 168:89-100. fied rat olfactory bulb interneurons to electrical and odor stimula- tion. J . Neurophysiol., 64:932-947.
Wellis, D.P,, and Scott, J,W, (1991) Localizedl denervation demon- strates the innervation pattern of olfactory bulb glomeruli and sec-
Wellis, D.P., Scott, J.W., and Harrison, T.A. (1989) Discrimination among odorants by single neurons of the rat olfactory bulb. J. Neu- rophysiol,, 61:1161-1177,
m i t e , E.L. (1972) Synaptic organization in the olfactory glomerulus of the mouse. Brain Res., 37:69-80.
m i t e , J., Hamilton, K.A., Neff, S.R., and Kauer, J.S. (1992) Emer- gent properties of odor information coding in a representational model of the salamander olfactory bulb. J. Neurosci., 12:1772-1780.
Wilson, D.A., and Leon, M. (1987) Evidence of lateral synaptic inter- actions in olfactory bulb output cell responses to odors, Brain Res., 417:175-180.
Woolf, T.B., Shepherd, G.M., and Greer, C.A. (1991) Local information processing in dendritic trees: Subsets of spines in granule cells of the mammalian olfactory bulb. J . Neurosci., 11:1837-1854.
Zheng, L.M., and Jourdan, F. (1988) Atypical olfactory glomeruli con- tain original olfactory axon terminals: an ultrastructural horserad- ish peroxidase study in the rat. Neuroscience, 362:367-378.
Zheng, L.M., Ravel, N., and Jourdan, F. (1987) Topography of centrif- ugal acetyl-cholinesterase-positive fibres in the olfactory bulb of the rat: Evidence for original projections in atypical glomeruli. Neuro- science, 23:1083-1093.
J. NeuroPhYsiol.~ 38:1284-1296.
G.M' (1963) Responses Of mitral to nerve Wellis, D,P,, and Scott, J,W, (1990) Intracellular responses of identi- Shepherd, G,M, (1971) Physiological evidence for dendrodendritic
synaptic interactions in the rabbit's olfactory glomerulus. Brain Res., 32:212-217.
Shepherd, G.M. (1972) Synaptic organization of the mammalian 01- factory bulb. Physiol. Rev., 52:864-917.
Shepherd, G.M. (1981) Synaptic and inlpulse loci in olfactory bulb dendritic circuits. In: Neurones Withwt Impulses. A. Roberts and B.M.H. Bush, eds. Cambridge University Press, Cambridge, U.K., pp. 255-267.
Shepherd, G.M., and Firestein, s. (1991) Toward a Pharmacolo&3' of odor receptors and the processing of odor images. J. Steroid Bio- chem. Molec. Biol., 39:583-592.
Shepherd, G.M., and Greer, C.A. (1990) Olfactory bulb. In: The SYn- aptic Organization of the Brain. G.M. ,Shepherd, ed. Oxford Univer- sity Press, New York, pp. 133-169.
Shinoda, K., Shiotana, y., and Osawa, y . (1989) Necklace olfactory glomeruli form unique components of :he rat Primary olfactory SYS- tem. J. Comp. Neurol., 284:362-373.
Skeen, L.C., and Hall, W.C. (1977) Eflwent Projections of the main and the accessory olfactory bulb in the tree shrew (TupaZa g l i ) J. Comp. Neurol., 172:l-36.
Slotnick, B.M., and Berman, E.J. (1983) Transection of the lateral olfactory tract does not produce anosinia. Brain Res. Bull., 5:141- 145.
Slotnick, B.M., Graham, S., Laing, D.G., and Bell, G.A. (1987) Detec-
order cells, J, Camp, Neural,, 304:544-554.




















