
From Banned to Celebrated, yet Relegated and Segregated: The Struggles of the Female Footballer
Upload
independentCategory
view
0download
0
Segregated pathways to the vomeronasal amygdala:differential projections from the anterior and posteriordivisions of the accessory olfactory bulb
Alicia Mohedano-Moriano,1,* Palma Pro-Sistiaga,1,* Isabel Ubeda-Banon,1 Carlos Crespo,2 Ricardo Insausti1 andAlino Martinez-Marcos1
1Laboratorio de Neuroanatomıa Humana, Departamento de Ciencias Medicas, Facultad de Medicina, Centro Regional deInvestigaciones Biomedicas, Universidad de Castilla-La Mancha, 02006 Albacete, Spain2Departamento de Biologıa Celular, Facultad de Ciencias Biologicas, Universidad de Valencia, 46100 Burjassot, Spain
Keywords: G-protein-coupled receptor, pheromone, rat, vomeronasal receptor
Abstract
Apically and basally located receptor neurons in the vomeronasal sensory epithelium express Gi2a- and Goa-proteins, V1R and V2Rvomeronasal receptors, project to the anterior and posterior accessory olfactory bulb and respond to different stimuli, respectively.The extent to which secondary projections from the two portions of the accessory olfactory bulb are convergent in the vomeronasalamygdala is controversial. This issue is addressed by using anterograde and retrograde tract-tracing methods in rats includingelectron microscopy. Injections of dextran-amines, Fluoro Gold, cholera toxin-B subunit and Fast Blue were delivered to the anteriorand posterior accessory olfactory bulb, bed nucleus of the stria terminalis, dorsal anterior amygdala and bed nucleus of the accessoryolfactory tract ⁄ anteroventral medial amygdaloid nucleus. We have demonstrated that, apart from common vomeronasal-recipientareas, only the anterior accessory olfactory bulb projects to the bed nucleus of the stria terminalis, medial division, posteromedialpart, and only the posterior accessory olfactory bulb projects to the dorsal anterior amygdala and deep cell layers of the bed nucleusof the accessory olfactory tract and the anteroventral medial amygdaloid nucleus. These results provide evidence that, excludingareas of convergence, the V1R and V2R vomeronasal pathways project to specific areas of the amygdala. These two vomeronasalsubsystems are therefore anatomically and functionally separated in the telencephalon.
Introduction
Parallel to the olfactory system, most mammals possess a vomeronasalsystem that includes a vomeronasal organ, an accessory olfactory bulb(AOB) and some vomeronasal-recipient amygdaloid structures (Halp-ern & Martinez-Marcos, 2003). The olfactory system is able to sense alarge number of odorants (Malnic et al., 1999), including some volatilesubstances that arise from pheromones (Lin et al., 2004); whereas thevomeronasal system detects mainly (Luo et al., 2003), but notexclusively, pheromones (Restrepo et al., 2004; Baxi et al., 2006).
In many mammals, the vomeronasal system is anatomically andfunctionally dichotomous (Halpern et al., 1998a,b). Two classes ofvomeronasal receptor neurons, apically and basally placed in thesensory epithelium, express Gi2a- and Goa-proteins (Berghard & Buck,1996; Jia & Halpern, 1996), V1R and V2R vomeronasal receptors(Dulac & Axel, 1995; Herrada & Dulac, 1997; Matsunami & Buck,1997; Ryba & Tirindelli, 1997), respond to different stimuli (Del Puntaet al., 2002a) and project to specific glomeruli in the anterior andposterior AOB (Belluscio et al., 1999; Rodriguez et al., 1999),respectively. The two portions of the AOB are also functionallydifferent (Sugai et al., 1997). The multiple apical dendrites of mitral-tufted cells tend to restrict their distribution to glomeruli innervated by
axons of sensory neurons expressing the same vomeronasal receptorwithin one of the divisions of the AOB (Jia & Halpern, 1997; DelPunta et al., 2002b; Wagner et al., 2006). How this segregatedinformation is transferred to the vomeronasal amygdala is a matter ofcontroversy.In opossums, the posterior AOB receives differential centrifugal
inputs from the posteromedial cortical amygdaloid nucleus (PMCo;Martinez-Marcos & Halpern, 1999a). Also in opossums, differentialprojections from the posterior AOB to deep cell layers of the medialamygdaloid complex (Me) have been reported (Martinez-Marcos &Halpern, 1999b). Anterograde (von Campenhausen & Mori, 2000) andretrograde (Salazar & Brennan, 2001) tracing studies, however, havenot revealed divergent projections from the two portions of the AOBin mice.In the present report we show, for the first time, vomeronasal-
recipient structures in the telencephalon of rats receiving specificinputs from the anterior and posterior divisions of the AOB.Accordingly, apart from a common vomeronasal pathway, twoseparate pathways can be traced from V1R and V2R receptors togiven areas in the vomeronasal amygdala.
Materials and methods
Thirty-six adult male and female Sprague–Dawley rats from theUniversity Hospital of Albacete were used in the present study. Animalswere maintained under veterinary supervision in a 12 h light : dark
Correspondence: Dr A. Martinez-Marcos, as above.E-mail: [email protected]
*A.M.-M. and P.P.-S. have equally contributed to this work.
Received 24 July 2006, revised 6 February 2007, accepted 8 February 2007
European Journal of Neuroscience, Vol. 25, pp. 2065–2080, 2007 doi:10.1111/j.1460-9568.2007.05472.x
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
E u r o p e a n J o u r n a l o f N e u r o s c i e n c e

cycle at 21 ± 2 �C. Experimental procedures were carried out accordingto the guidelines of the European Community on welfare of researchanimals (directive 86 ⁄ 609 ⁄ EEC) and the local Ethical Committee ofAnimal Research of the University of Castilla La Mancha.Animals were intraperitoneally injected with a combined dose of
ketamine hydrochloride (Ketolar�, Parke-Davis, Madrid, Spain,1.5 mL ⁄ kg, 75 mg ⁄ kg) and xylazine (Xilagesic�, Calier, Barcelona,Spain, 0.5 mL ⁄ kg, 10 mg ⁄ kg). Eye-drops (Lacryvisc�, Alcon, Bar-celona, Spain) were applied to prevent eye ulceration during surgery.The animals were placed into a Kopf stereotaxic apparatus and theskull trepanned at the intended injection site (Paxinos & Watson,2005). Ionophoretic injections of dextran-amines conjugated to biotin(BDA), tetramethylrhodamine (RDA) or fluorescein (FDA; 10 000MW, lysine fixable, Molecular Probes, Eugene, OR, USA), Fluoro-Gold� (FG) (methanesulphonate hydroxystilbamidine, Biotium, CA,USA) and cholera toxin B-subunit (CTB; List Biological Laboratories,CA, USA) were performed. Tracers were delivered from micropipettes(30–50-lm diameter tips) by means of positive current pulses (7 ⁄ 7 s,2–7 lA, 10–20 min). Fast-Blue (FB, Polysciences, Warrington, PA,USA) was injected using a Hamilton syringe (Table 1). A singlesubcutaneous dose of an analgesic (Buprenorphine, Buprex;0.17 mL ⁄ kg, 0.05 mg ⁄ kg, Schering-Plough, Madrid, Spain) wasadministered after surgery.Five days later, animals were anaesthetized (as above) and perfused
with saline solution followed by 4% paraformaldehyde in phosphatebuffer. The brains were postfixed for 4 h and cryoprotected overnightwith 30% buffered sucrose. Parasagittal (olfactory bulbs) or frontalsections (50 lm) were obtained with a freezing microtome. For BDAdetection, endogenous peroxidase activity was quenched by 1% H2O2
(in 0.05 m Tris buffer saline, pH 7.6, for 30 min). Sections wereincubated for 2 h in avidin–biotin complex (ABC elite kit, Vector,Burlingame, CA, USA; diluted in 0.05 m Tris buffer, pH 7.6) andvisualized with 0.02% 3,3¢-diaminobenzidine (Sigma, St. Louis, MO,USA) diluted in 0.05 m Tris buffer (pH 8.0) with 0.1% ammoniumnickel sulphate and 0.01% H2O2. Sections were mounted, dried, somesections stained with thionin (Panreac, Barcelona, Spain) andcoverslipped with DPX (BDH Laboratory, Poole, England). Injectionsof CTB were revealed by means of immunocytochemistry (goat anti-CTB antibody 1 : 4000, List Biological Laboratories; donkey anti-
goat antibody 1 : 400 and peroxidase anti-peroxidase 1 : 1000,Lutherville, MA, USA).Six animals injected with BDA (three AOBr, three AOBc) were
processed for electron microscopy. These animals were perfused with4% paraformaldehyde and 0.5% glutaraldehyde in 0.1 m phosphatebuffer. After perfusion, the brains were removed and 60-lm-thicktransverse sections were cut on a vibratome (VT 1000S, Leica,Nussloch, Germany). Sections were collected in cold 0.1 m phosphatebuffer (4 �C) and treated with 1% NaBH4 in phosphate buffer for30 min and thoroughly rinsed in the same buffer. Free-floatingsections were cryoprotected overnight by immersion in 25% sucroseand 10% glycerol in 0.05 m phosphate buffer, freeze-thawed threetimes in liquid nitrogen to enhance antibody penetration and processedfor BDA detection as previously described. Afterwards, sections werewashed in phosphate buffer and then lipids were postfixed with 1%osmium tetroxide (EMS) for 60 min at room temperature, stained with2.5% uranyl acetate (Sigma) in maleate buffer for 90 min at 4 �C,dehydrated through increasing graded ethanol series, cleared inpropylene oxide and flat-embedded in Durcupan (Fluka). Afteranalysis of the sections under the light microscope, some fieldscontaining stained fibres in dorsal anterior amygdala (AAd), antero-ventral medial amygdaloid nucleus (MeAV), bed nucleus of theaccessory olfactory tract (BAOT), bed nucleus of the stria terminalis,medial division, posteromedial part (BSTMPM) and st werere-embedded, and 60-nm-thick ultrathin sections were cut with anultramicrotome. Ultrathin sections were serially collected on single-slot Formvar-coated nickel grids, stained using lead citrate andanalysed under the electron microscope.Immunofluorescence for Goa-proteins was performed using anti-
Goa-protein (1 : 2000, Medical and Biological Laboratories, Woburn,MA, USA) and Alexa� 350 or 488 (1 : 200, Molecular Probes). Toobserve fluorescent tracers, sections were mounted and coverslippedwith 30% glycerin in 0.4 m potassium bicarbonate. Images weremerged using Adobe Photoshop 8.0.1 software, and figure panelsarranged and lettered using Canvas 7.0 (Deneba).
Results
Thirty-six animals received one, two or three injections in the samehemisphere or different hemispheres, totalling 60 injections (Table 1).First, anterograde tracer experiments using BDA are described;second, anterograde tracing experiments using RDA and retrogradetracer experiments are used to confirm projections; third, double-labelling tracer experiments allowed us comparisons of projectionsfrom the two portions of the AOB in the same sections.
Injections of BDA in the AOB
A total of 10 ionophoretic injections of BDA were aimed at the AOB(three total, four rostral, three caudal). Injections affecting bothportions of the AOB gave rise to anterograde labelling in basaltelencephalic structures similar to that described in previous reports(Winans & Scalia, 1970; Scalia & Winans, 1975; Price, 1987;Martinez-Marcos & Halpern, 1999b; von Campenhausen & Mori,2000). Counterstained sections help in the cytoarchitectonic identifi-cation of telencephalic structures. Rostrally, fibres were seen in layer Iof the nucleus of the lateral olfactory tract and anterior corticalamygdaloid nucleus (ACo), some of which turned dorsally to reachthe dorsal and ventral anterior amygdalae (AAd and AAv, respec-tively), and eventually enter the loop-shaped stria terminalis (st)ventrally (asterisks in Fig. 1A, not visible at this magnification). These
Table 1. Animals used including tracers delivered and injection sites
Cases and verified injection sites
AOBt AOBr AOBc AAd BSTMPM MeAV ⁄ BAOT
BDA4705 BDA4805 RDA4705 CTB8705 FG8205 FDA7705FDA4705 RDA4805 BDA5605 FB9305 FG9105 FDA8405BDA6005 BDA5205 FDA5605 CTB9905 FG2406 FDA8705RDA6005 BDA5505 BDA6605 RDA1306 RDA5506 FDA9305FDA6005 FDA6305 RDA6605 FG1306 FDA5506 FG9305BDA6505 RDA6505 FDA6605 FG1406 RDA5806 FB9405RDA8205 BDA6805 BDA9105 FG5506 – –FDA9105 RDA6805 RDA2406 FG5806 – –– RDA9605 FDA9605 – – –– RDA10205 FDA10205 – – –– RDA6106 FDA6106 – – –– RDA7306 FDA7306 – – –– RDA7606 FDA7606 – – –– BDA8906 BDA9006 – – –– BDA9106 BDA9206 – – –– BDA9306 BDA9406 – – –
Numbers correspond to animals and letters to tracers delivered. For abbrevia-tions, see list.
2066 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

fibres continued caudally within the st, then dorsally (asterisk inFig. 1B) and rostrally to reach the medial aspect of the supracapsularbed nucleus of the st (BSTS; Alheid et al., 1998; Shammah-Lagnadoet al., 2000) (asterisk in Fig. 1C), and finally more rostrally to end inthe BSTMPM (asterisk in Fig. 1D). Labelled fibres within the stfollowed the dorsal component, as described by de Olmos (de Olmos& Ingram, 1972; de Olmos, 1972), and the medial division of theextended amygdala (Alheid et al., 1998; Shammah-Lagnado et al.,2000; de Olmos et al., 2004). Ventral fibres continued caudally toreach superficial layer I of the BAOT and anteroventral andanterodorsal medial amygdaloid nuclei (MeAV and MeAD, respect-ively; Fig 1E and F). Note that fibres in the BAOT and MeAV reacheddeep cell layers (asterisks in Fig. 1E and F, not visible at thismagnification). More caudally, fibres reached the posteroventral andposterodorsal medial amygdaloid nuclei (MePV and MePD; Fig. 1G).At this level, fibres arising from the MePD were seen entering the st(asterisk in Fig. 1G). Finally, fibres reached the full extent of thePMCo (Fig. 1G and H).
Injections into the anterior AOB (Figs 2A and 3A) gave rise toanterograde labelling similar to that described for total injectionsincluding the BAOT, Me and PMCo (Figs 2C, E and G, and 3E, G andI). As above, labelled fibres entered the loop-shaped st ventrally withinthe amygdala and were seen along its entire course, running firstcaudal (arrows in Figs 2C, 3C, and 4A and B). More caudally, thesefibres joined a contingent of labelled fibres arising from the MePD(Fig. 4C), running then together dorsally (Fig. 4D), then rostrally inthe BSTS (arrows in Figs 2G and E, and 4E) to end rostrally in theBSTMPM (arrow in Fig. 2C) where they formed a dense terminal field(Fig. 4F). Electron microscope studies reveal that labelled fibres alongthe st corresponded to fibres of passage both at caudal (Fig. 6A) anddorsal (BSTS, Fig. 6B) levels. High-power micrographs of the st in theBSTS, however, suggest the presence of synaptic boutons (arrowheadsin Fig. 4E) that could not be confirmed with electron microscopy.Therefore, it cannot be confirmed whether these axons synapse toneurons of the BSTS (Alheid et al., 1998; Shammah-Lagnado et al.,2000). High-power micrographs revealed synaptic boutons in theBSTMPM (arrowhead in Fig. 4F) that were confirmed using electronmicroscopy (Fig. 6C). The course of labelled fibres within the stperfectly matches the description of the st and medial division of theextended amygdala by de Olmos and colleagues (de Olmos et al.,2004).
Ionophoretic injections of BDA in the posterior AOB (Figs 2B and3B) gave rise to a pattern of anterograde labelling similar to thatdescribed for total injections in the AOB (Figs 2D, F and H, and 3D,F, H and J). Compared with rostral injections, labelling was similar inthe st at the level of the AAv (Figs 2D, and 5A and B). Labelling,however, was much reduced at levels of the MePD (Fig. 5C), morecaudally when fibres turned dorsally (Fig. 5D), in BSTS (Figs 2F and5E) and BSTMPM (Figs 2D and 5F). Labelled fibres were observed inthe AAd (arrow in Figs 2D and 3D), which exhibited synaptic boutons(Figs 5G and 6D) and layers II and III of the BAOT and MeAV(arrows in Figs 2F and H, and 3F and H) also showing synapticcontacts (Figs 5H, I and J, and 6E and F).
Injections of fluorescent dextrans in the AOB and retrogradeexperiments
A total of 14 injections of fluorescent dextrans involved both (3FDA,2RDA), rostral (1FDA, 3RDA) or caudal (2FDA, 3RDA) divisions ofthe AOB. Injections of fluorescent dextrans had two advantagescompared with BDA injections. Injections of RDA and FDA were
more restricted and they could be combined with immunofluorescentantibodies against G-proteins to delineate the anterior–posteriorsubdivisions of the AOB. Retrograde experiments included eightinjections in the AAd (2CTB, 1FB, 1RDA, 4FG), six in the BSTMPM(3FG, 2RDA, 1FDA) and six in the MeAV ⁄ BAOT (4FDA, 1FG,1FB).Injections of RDA restricted to the anterior AOB (immunofluores-
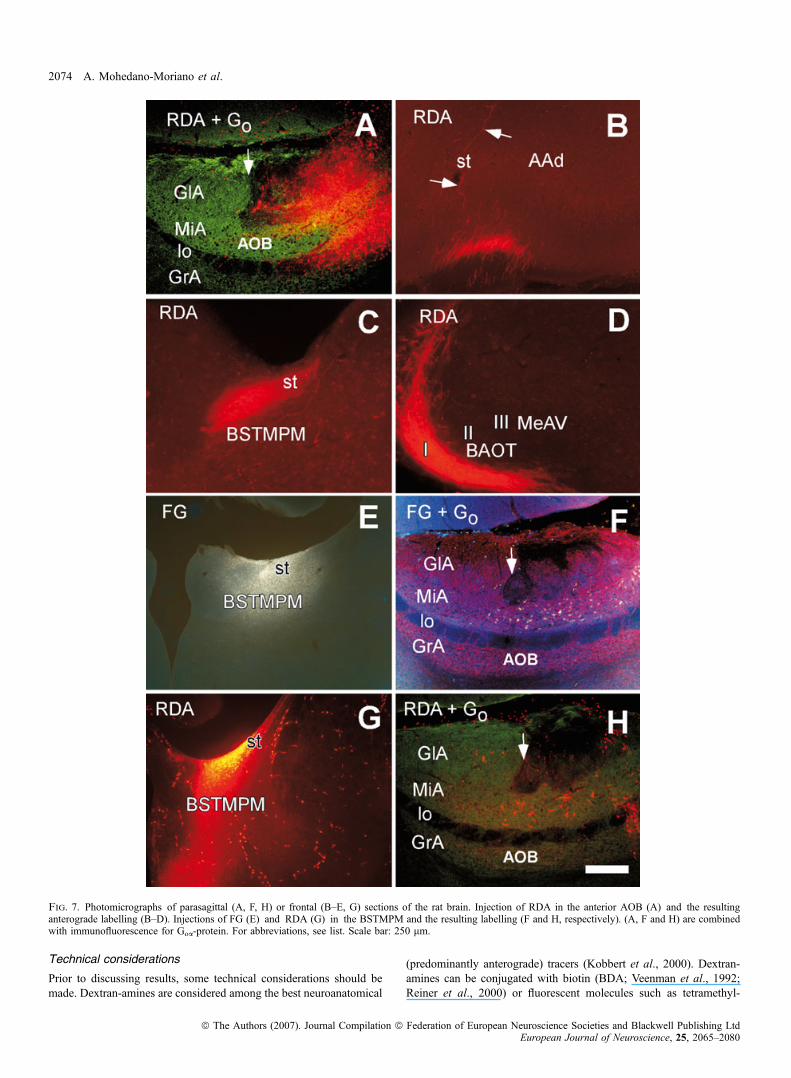
cence for Goa-protein showed the anterior ⁄ posterior AOB border,arrow in Fig. 7A) resulted in a similar pattern of labelling to thoseinjections of BDA in the rostral AOB including fibres in the st (arrowsin Fig. 7B), BSTMPM (Fig. 7C) and layer I of the BAOT, Me(Fig. 7D) and PMCo (not shown).A total of six injections of the retrograde tracers (3FG, 1FDA,
2RDA) were aimed at the BSTMPM. Fluorescent dextrans (FDA andRDA, even 10 000 MW) can function as retrograde tracers undercertain conditions (Nance & Burns, 1990; Schmued et al., 1990;Richmond et al., 1994; Kaneko et al., 1996; Martinez-Marcos &Halpern, 1999a; Kobbert et al., 2000; Reiner et al., 2000). Injectionsof FG (Fig. 7E) or RDA (Fig. 7G) in the st near the BSTMPM gaverise to retrogradely labelled cells that were virtually restricted to theanterior AOB (Fig. 7F and H, respectively; Goa-protein immuno-fluorescence indicated the border between AOB divisions, arrows inFig. 7F and H).Injections of RDA restricted to the posterior AOB (immunofluo-
rescence for Goa-protein indicated the rostro-caudal AOB border,arrow in Fig. 8A) gave rise to anterograde labelling in the AAd(Fig. 8B, compare with Fig. 7B) and layers II and III of the BAOT andMeAV (Fig. 8C, compare with Fig. 7D). In contrast to injections ofBDA in the posterior AOB, injections of RDA restricted to theposterior AOB (as confirmed using immunofluorescence for Goa-protein) did not give rise to anterograde labelling in the BSTMPM (notshown).Three animals received simultaneous injections of FG in the
BSTMPM (Fig. 8D), combined with injections of RDA involving bothanterior and posterior or the posterior division of the AOB (notshown). These injections produced double-labelling of RDA and FGin the AAd and MeAV. RDA-labelled axons arising from the posteriorAOB appeared to make synaptic contacts with neurons in the AAd(arrow in Fig. 8E), the cell bodies of which were retrogradely labelledafter injections into the BSTMPM (Fig. 8D). Similarly, cells located inlayers II and III of the MeAV (projecting to the BSTMPM) appeared toreceive inputs from the posterior AOB in close proximity to their cellbodies (arrows in Fig. 8F) and different from those occurring in layer Ibetween axons of mitral ⁄ tufted cells and superficially directeddendrites of amygdaloid neurons.Injections of retrograde tracers in the AAd and layers II and III of
the BAOT ⁄ MeAV were used as confirmatory experiments. Injectionsof CTB in the AAd, not affecting the st (Fig. 8G), gave rise toretrogradely labelled cells exclusively in the posterior AOB (Fig. 8H).Similar experiments in layers II and III of the BAOT ⁄ MeAV alsoproduced retrograde labelling in the posterior, but not anterior, AOB(not shown).
Simultaneous injections of RDA and FDA in the anterior andposterior AOB
In order to compare, in the same sections, the projections arising fromthe two divisions of the AOB, five animals were injected with RDAand FDA in the anterior and posterior AOB, respectively (Table 1).Injections of RDA in the anterior AOB (Fig. 9A) yielded a pattern
of labelling similar to that described above, including labelled fibres inthe BSTMPM (Fig. 9C, E, G and I). Injections of FDA in the posterior
Segregated pathways to the vomeronasal amygdala 2067
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
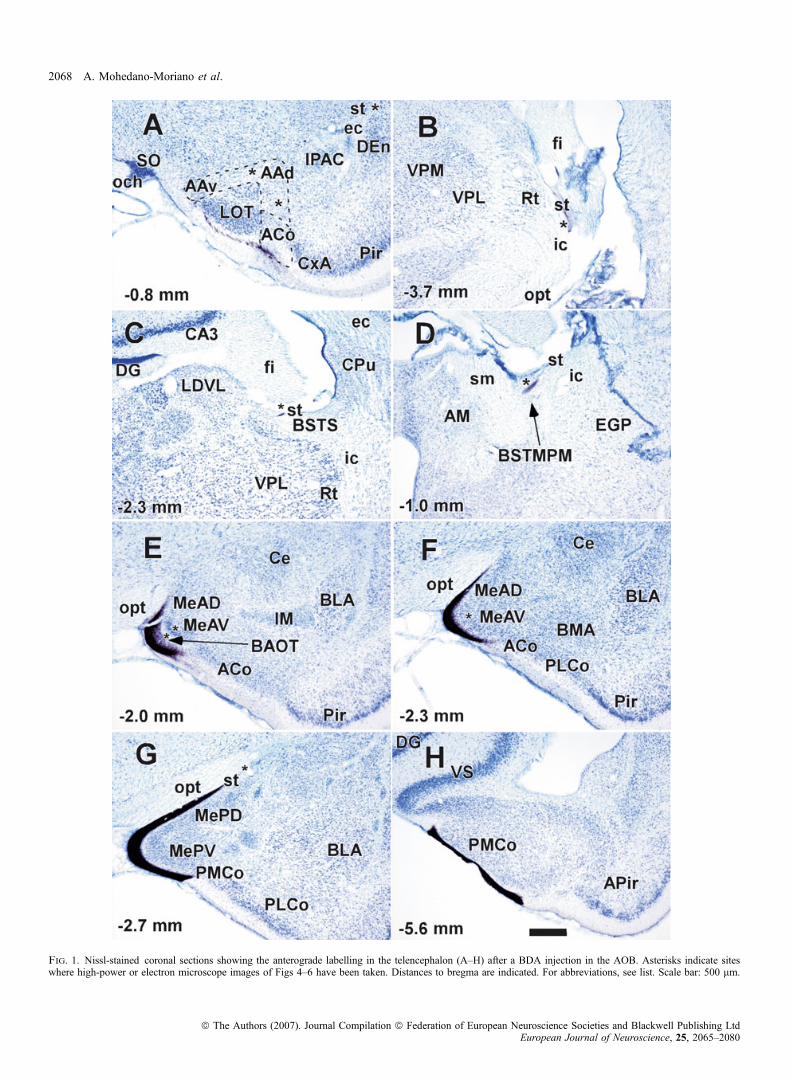
Fig. 1. Nissl-stained coronal sections showing the anterograde labelling in the telencephalon (A–H) after a BDA injection in the AOB. Asterisks indicate siteswhere high-power or electron microscope images of Figs 4–6 have been taken. Distances to bregma are indicated. For abbreviations, see list. Scale bar: 500 lm.
2068 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
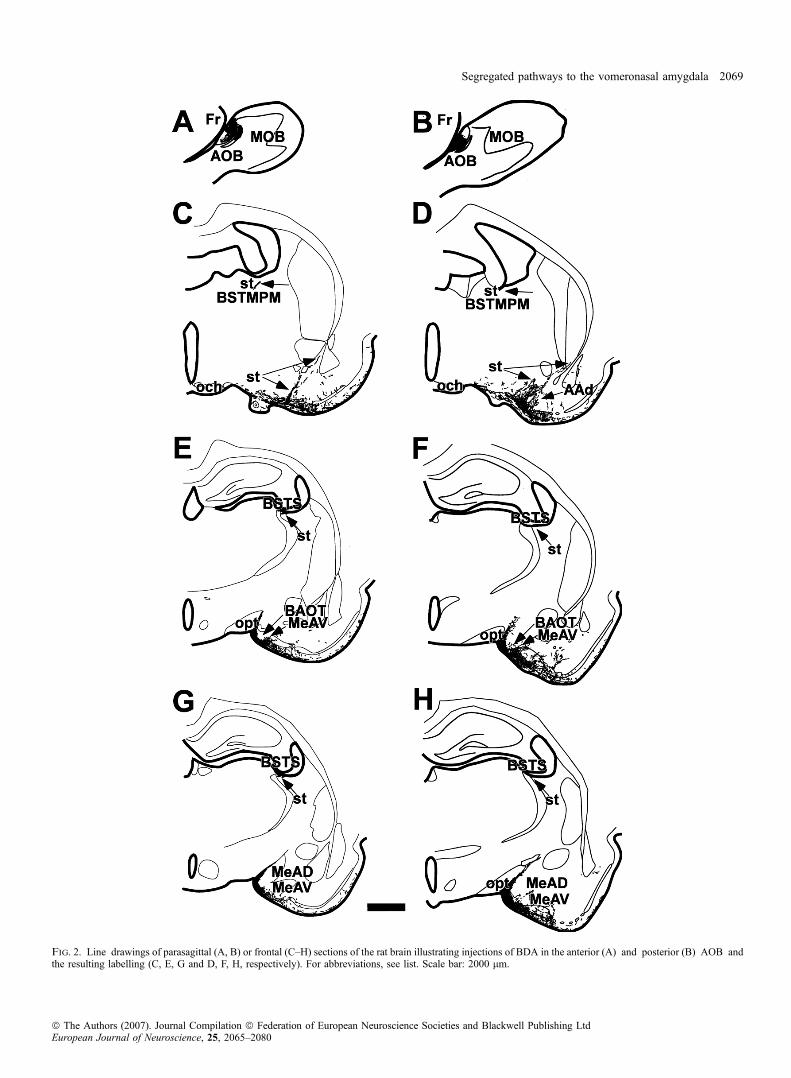
Fig. 2. Line drawings of parasagittal (A, B) or frontal (C–H) sections of the rat brain illustrating injections of BDA in the anterior (A) and posterior (B) AOB andthe resulting labelling (C, E, G and D, F, H, respectively). For abbreviations, see list. Scale bar: 2000 lm.
Segregated pathways to the vomeronasal amygdala 2069
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
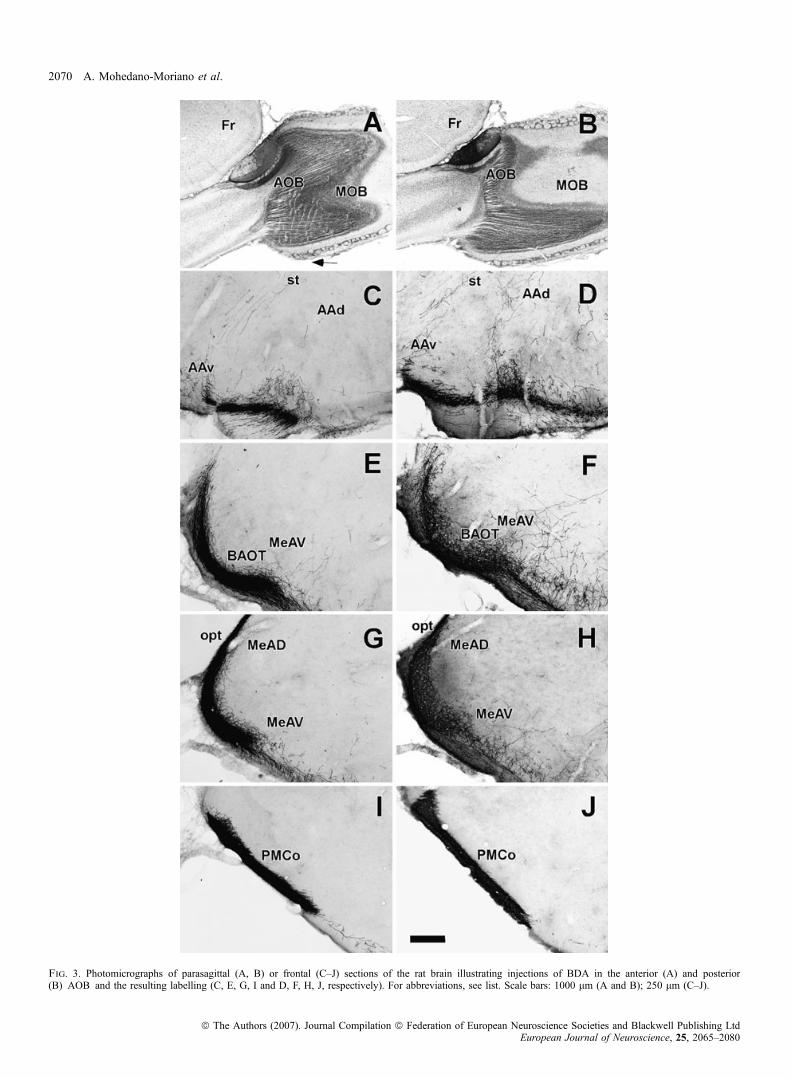
Fig. 3. Photomicrographs of parasagittal (A, B) or frontal (C–J) sections of the rat brain illustrating injections of BDA in the anterior (A) and posterior(B) AOB and the resulting labelling (C, E, G, I and D, F, H, J, respectively). For abbreviations, see list. Scale bars: 1000 lm (A and B); 250 lm (C–J).
2070 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

AOB (Fig. 9B) also resulted in a pattern of labelling similar to thatdescribed above for caudal injections (Fig. 9D, F, H and J), includingthe absence of labelling in the BSTMPM (Fig. 9D) and the presence oflabelled fibres in the AAd (Fig. 9F) and layers II and III of theBAOT ⁄ MeAV (Fig. 9H).
Discussion
In the present report, the efferent projections from the anterior andposterior divisions of the AOB were investigated. Apart from common
vomeronasal-recipient structures where both projections converge(layer I of the BAOT, Me and PMCo), differential projections havebeen identified. A projection from the anterior AOB to the BSTMPMis described in rats using anterograde tracers and confirmed usingretrograde tracers. The posterior AOB does not appear to project to theBSTMPM. The posterior AOB projects to the AAd and deep layers ofthe BAOT ⁄ MeAv, a projection not shared by the anterior AOB(Fig. 10). These results constitute the first demonstration of differentialprojections from both divisions of the AOB to specific areas of thevomeronasal amygdala.
Fig. 4. High-power photographs illustrating the resulting labelling at different levels of the st (A–F) after a BDA injection in the rostral AOB. Locations of picturesA and B approximately correspond to asterisks in Fig. 1A and C, to asterisks in Fig. 1G and D, to asterisks in Fig. 1B and E, to asterisks in Fig. 1C and F, and toasterisks in Fig. 1D. For abbreviations, see list. Scale bar: 25 lm.
Segregated pathways to the vomeronasal amygdala 2071
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
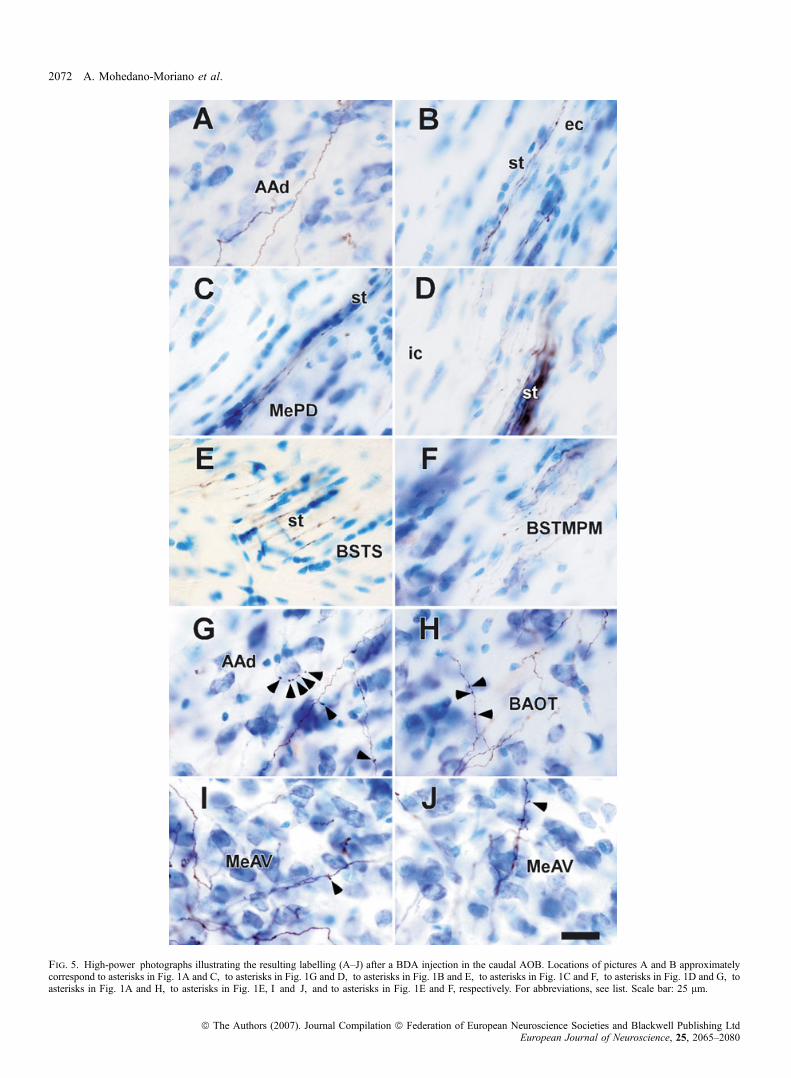
Fig. 5. High-power photographs illustrating the resulting labelling (A–J) after a BDA injection in the caudal AOB. Locations of pictures A and B approximatelycorrespond to asterisks in Fig. 1A and C, to asterisks in Fig. 1G and D, to asterisks in Fig. 1B and E, to asterisks in Fig. 1C and F, to asterisks in Fig. 1D and G, toasterisks in Fig. 1A and H, to asterisks in Fig. 1E, I and J, and to asterisks in Fig. 1E and F, respectively. For abbreviations, see list. Scale bar: 25 lm.
2072 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

Fig. 6. Electron microscope photographs illustrating the resulting labelling after BDA injections in the rostral (A–C) and caudal (D–F) AOB. BDA-stained fibres ofpassage (arrows) in the caudal st (A, location corresponds to asterisk in Fig. 1B) and BSTS (B, location corresponds to asterisk of Fig. 1C). Boutons from BDA-containing axons (asterisks) made asymmetrical synaptic contacts (arrows) upon dendritic profiles of the BSTMPM (C, location corresponds to asterisk in Fig. 1D),AAd (D, location corresponds to asterisk in Fig. 1A), BAOT (E, location corresponds to asterisk in Fig. 1E) and MeAV (F, location corresponds to asterisk inFig. 1F). Note that the postsynaptic element shown in (D) receives an additional synapse (open arrow) from an unidentified axon. For abbreviations, see list. Scalebars: 1 lm (A and B); 500 nm (C and D).
Segregated pathways to the vomeronasal amygdala 2073
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
Technical considerations
Prior to discussing results, some technical considerations should bemade. Dextran-amines are considered among the best neuroanatomical
(predominantly anterograde) tracers (Kobbert et al., 2000). Dextran-amines can be conjugated with biotin (BDA; Veenman et al., 1992;Reiner et al., 2000) or fluorescent molecules such as tetramethyl-
Fig. 7. Photomicrographs of parasagittal (A, F, H) or frontal (B–E, G) sections of the rat brain. Injection of RDA in the anterior AOB (A) and the resultinganterograde labelling (B–D). Injections of FG (E) and RDA (G) in the BSTMPM and the resulting labelling (F and H, respectively). (A, F and H) are combinedwith immunofluorescence for Goa-protein. For abbreviations, see list. Scale bar: 250 lm.
2074 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

rhodamine (RDA; Nance & Burns, 1990; Schmued et al., 1990) orfluorescein (FDA; Nance & Burns, 1990). In addition, BDA tolerateselectron microscope fixation and processing (Kobbert et al., 2000;
Reiner et al., 2000). Like other anterograde tracers, apart from theproblem of optimizing the survival time or concentration to achieveoptimal injection sites, the main problem with dextran-amines is the
Fig. 8. Photomicrographs of parasagittal (A, H) or frontal (B–G) sections of the rat brain. Injection of RDA in the posterior AOB (A) and the resulting anterogradelabelling (B and C). Injection of FG (D) in the BSTMPM and injection of RDA in the AOB (not shown) and the resulting double labelling (E and F). Injection ofCTB in the AAd (G) and the resulting labelling (H). (A) is combined with Goa-protein. For abbreviations, see list. Scale bars: 250 lm (A, B, D and H); 125 lm(C and F); 25 lm (E); 1562 lm (G).
Segregated pathways to the vomeronasal amygdala 2075
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
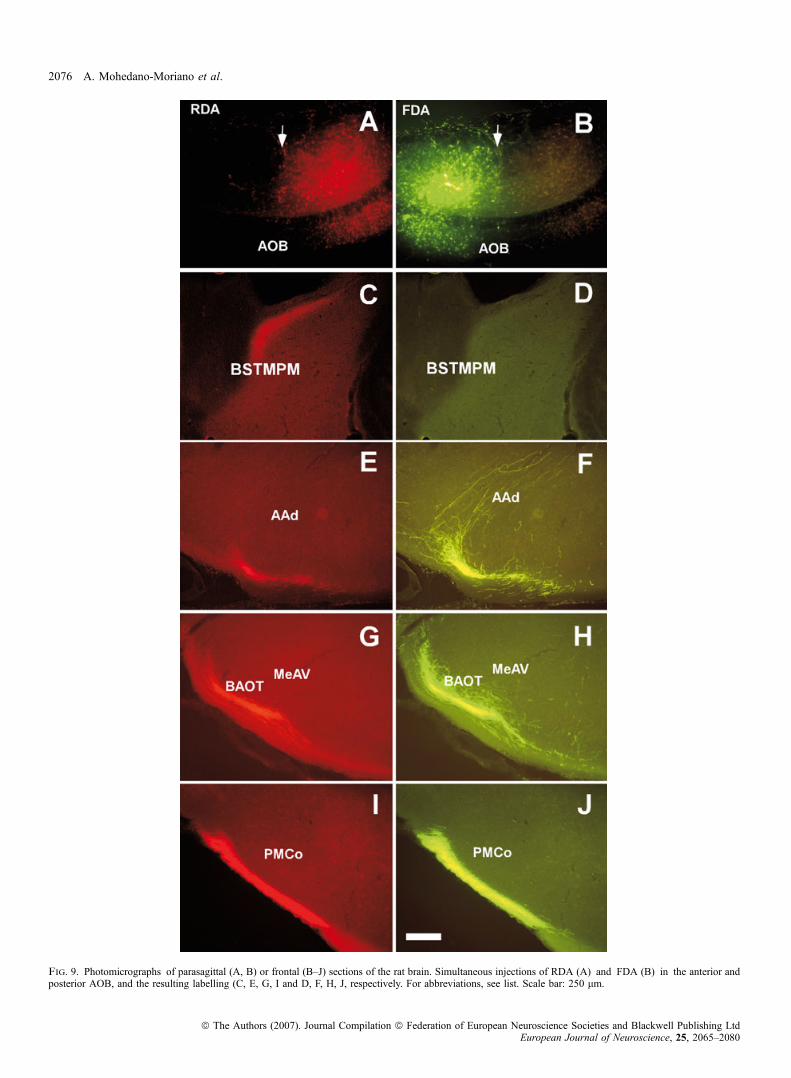
Fig. 9. Photomicrographs of parasagittal (A, B) or frontal (B–J) sections of the rat brain. Simultaneous injections of RDA (A) and FDA (B) in the anterior andposterior AOB, and the resulting labelling (C, E, G, I and D, F, H, J, respectively. For abbreviations, see list. Scale bar: 250 lm.
2076 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
potential for labelling fibres of passage. Regarding the first problem,because the AOB is quite a small structure (between 1500 and2000 microns from rostral to caudal, i.e. about 750 and 1000 micronsfor subdivision), we had to adjust the concentration of tracers andinjection parameters (see Materials and methods) to get injection sitesfrom 250 to 500 microns and avoid the problem of contaminationbetween subdivisions of the AOB. In the second technical problem,the tracer is being taken up by axons damaged by the injection or evenby intact axons (Nance & Burns, 1990; Reiner et al., 2000). In ourexperiments, for instance, this possibility could arise when injecting inthe posterior AOB, as tracer could be taken up by axons of mitral cellsof the anterior AOB travelling through the posterior AOB to theamygdala. This potential problem has also been taken into account byusing retrograde tracing to confirm projections.
Regarding retrograde tracing, typical retrograde tracers such as FB,FG and CTB (Kobbert et al., 2000) have been used. Given ourdifficulty of getting small injection sites with FB or FG or enoughretrograde labelling with CTB (Fig. 8G), RDA has also been used(Schmued et al., 1990) to confirm some projections. When compared,however, RDA or FDA are less effective retrograde tracers than FB orFG (Richmond et al., 1994), although RDA can be reliably used as aretrograde tracer (Schmued et al., 1990) and its retrograde effective-ness is improved by using acidic vehicles for dissolving the tracer(Kaneko et al., 1996).
Anatomical remarks
In the past, the efferent projections of the AOB were usuallyinvestigated assuming that the AOB was a unitary entity that belongedto a chemosensory system (the vomeronasal or accessory olfactorysystem) parallel to the olfactory system (Winans & Scalia, 1970;Raisman, 1972; Broadwell, 1975; Scalia & Winans, 1975; Devor,1976; Skeen & Hall, 1977; Davis et al., 1978; Shammah-Lagnado &Negrao, 1981; Lehman & Winans, 1982). This anatomical distinctionbetween olfactory and vomeronasal systems was predicted to subservedifferent functions, the so-called ‘dual olfactory hypothesis’ (Halpern,1987).
Early reported vomeronasal-recipient structures include the anterioramygdala, BSTMPM, BAOT, Me and PMCo. The anterior amygdalahas been anatomically considered as the rostral pole of the amygdala.Functionally, it belongs to the medial division of the extendedamygdala. Hodologically, it receives inputs from the MOB and it hasbeen considered as part of the olfactory amygdala, although somerelationship with the vomeronasal amygdala has been proposed aswell (de Olmos et al., 2004). Only recently, the ventral and dorsalcomponents of the anterior amygdala (AAv and AAd) have beendescribed receiving inputs from the AOB (Martinez-Marcos &Halpern, 1999b). In fact, our unpublished observations indicate thatthe AAv receives both olfactory and vomeronasal inputs, whereas theAAd receives single inputs from the posterior AOB (see below). TheBSTMPM, according to its connections, has been involved inreproductive behaviour (Dong & Swanson, 2004; Wood & Swann,2005). The BAOT has been described as a sexually dimorphicstructure (Collado et al., 1990) related to maternal behaviour (DelCerro et al., 1991; Komisaruk et al., 2000). The Me is thought to playa key role in male sexual behaviour. The anterior and posteriordivisions of the Me may contribute to a different extent tochemosensory and hormonal components of such behaviour (Gomez& Newman, 1992; Coolen & Wood, 1998). Further, the Me has alsobeen divided into dorsal (MePD) and ventral (MeAD, MeAV, MePV)divisions devoted to reproductive and agonistic behaviours (Canteraset al., 1995). Finally, the PMCo, as most of the vomeronasal system, isalso sexually dimorphic (Vinader-Caerols et al., 1998), and it has beendemonstrated that species-specific chemosignals provoke a delayedexcitation in this amygdaloid nucleus (Mucignat-Caretta et al., 2006).Until recently, however, vomeronasal-recipient structures have notbeen studied from a dual perspective.During the last decade, the molecular (Shinohara et al., 1992; Dulac
& Axel, 1995; Halpern et al., 1995; Berghard & Buck, 1996; Jia &Halpern, 1996; Herrada & Dulac, 1997; Matsunami & Buck, 1997;Ryba & Tirindelli, 1997), anatomical (Belluscio et al., 1999;Rodriguez et al., 1999; Del Punta et al., 2002b) and functional(Leinders-Zufall et al., 2000; Del Punta et al., 2002a) heterogeneity inthe vomeronasal system from the vomeronasal epithelium to the AOBhas been established (Halpern et al., 1998a,b; Halpern & Martinez-Marcos, 2003). Attempts have been made to investigate the efferentprojections of the anterior and posterior AOB from this dualperspective.In opossums, differential centrifugal afferents from the PMCo to the
posterior granule cell layer of the AOB have been reported (Martinez-Marcos & Halpern, 1999a). Regarding efferent connections inopossums, a projection from the posterior AOB to deep cell layersof different nuclei of the Me was described (Martinez-Marcos &Halpern, 1999b). This situation contrasts with data in mice whereefferent projections from the anterior and posterior AOB convergealong the vomeronasal-recipient structures including the Me and theBSTMPM (von Campenhausen & Mori, 2000). Also in mice,retrograde tracing experiments in different vomeronasal amygdaloidnuclei did not reveal differential projections from the anterior andposterior AOB (Salazar & Brennan, 2001). The current view of theprojections from the two divisions of the AOB of rodents is that thereis no evidence for segregated pathways in the vomeronasal amygdala(Keverne, 1999; Mori et al., 2000). However, the present data in ratsindicate that, apart from the convergent projections in vomeronasal-recipient areas such as layer I of the Me and BAOT, there aredifferential projections arising from the two divisions of the AOB.The rostral AOB alone projects to the BSTMPM. The projection
from the AOB to the BSTMPM has been clearly recognized in rabbits(Scalia & Winans, 1975), but not observed in opossums (Scalia &
Fig. 10. Schematic diagram of frontal sections of the basal telencephalonsummarizing the main results. Black corresponds to convergent projectionsfrom both divisions of the AOB; dark grey corresponds to projections arisingfrom the rostral AOB; and light grey corresponds to projections arising fromthe caudal AOB. For abbreviations, see list.
Segregated pathways to the vomeronasal amygdala 2077
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

Winans, 1975; Martinez-Marcos & Halpern, 1999b), and appeared tobe minor in rats (Scalia & Winans, 1975), which is contrary to thepresent results. In mice, the projection appears to be robust and arisesfrom both divisions of the AOB (von Campenhausen & Mori, 2000).Studies of the afferent connections of the BSTMPM are not conclusiveregarding differential projections from the AOB (Weller & Smith,1982; Dong et al., 2001). Therefore, the projection from the AOB tothe BSTMPM appears to show interspecific variability, which must beconsidered when describing the projections of the AOB.According to our results, the AAd receives a differential projection
from the posterior AOB. The anterior amygdala is a poorlycharacterized area that has been classified as an olfactory as well asa vomeronasal structure in rats (de Olmos et al., 2004). This area canbe divided into ventral and dorsal components. The ventral anterioramygdala has been described receiving olfactory inputs in rats (deOlmos et al., 2004) and vomeronasal inputs in opossums (Martinez-Marcos & Halpern, 1999b), whereas the dorsal component (AAd) hasbeen reported receiving vomeronasal inputs in opossums (Martinez-Marcos & Halpern, 1999b) that arise specifically from the posteriorAOB in rats (present results).Finally, our results indicate that the deep cell layers (II and III) of
the BAOT and MeAV receive inputs from the posterior AOB, but notthe anterior AOB. In mice, studies using sensitive tracers such asdextrans (Kobbert et al., 2000; Reiner et al., 2000) reveal that both theBAOT and the Me receive inputs from the two divisions of the AOBthat reach not only layer I, but also deep cell layers (von Campen-hausen & Mori, 2000). In opossums, the BAOT is not recognizable(Scalia & Winans, 1975), and layers II and III of the anterodorsal,anteroventral and posteroventral nuclei of the Me receive differentialprojections from the posterior AOB (Martinez-Marcos & Halpern,1999b). Therefore, the projections from the AOB to the deep cell layerof the Me and related structures also appears to show interspecificdifferences that need to be taken into account.
Functional considerations
The present results provide, for the first time, anatomical data linkingvomeronasal stimuli sensed by V1R vomeronasal neurons to theBSTMPM. Similarly, stimuli sensed by V2R vomeronasal neuronsreach the AAd and layers II and III of the BAOT and MeAV. Forinstance, it is known that BSTMPM receives inputs from the AOB(von Campenhausen & Mori, 2000; Dong et al., 2001). According tothe present results, chemosensory inputs to gonadotropin-releasinghormone (GnRH) neurons from the BSTMPM (Boehm et al., 2005)may originate exclusively in V1R-expressing receptor neurons.Interestingly, GnRH neurons are essential for reproduction, and theBST has been described as critical for copulatory behaviour (Kollack-Walker & Newman, 1997; Newman, 1999), particularly erection inmale rats (Liu et al., 1997). Boehm and colleagues (Boehm et al.,2005) also demonstrate that GnRH neurons are heavily connected tothe ACo, traditionally described as an olfactory structure (Shipleyet al., 2004), but according to our results (Fig. 1A) also receivesvomeronasal inputs (unpublished results). On the other hand, differ-ential inputs from the posterior AOB to the AAd indicate that this areais receiving, exclusively, vomeronasal information processed throughV2R receptors. The AAd is a non-cortical structure only recentlyconsidered as vomeronasal-recipient (Martinez-Marcos & Halpern,1999b). Stimuli sensed specifically by V2R vomeronasal receptors, forinstance sex-specific peptides from exocrine glands (Kimoto et al.,2005), could be processed in areas such as the AAd. Finally, in theBAOT and MeAV, a number of neurons with dendrites directed to
layer I may receive inputs from the anterior and posterior AOB,whereas others neurons may be receiving specific inputs from theposterior AOB, i.e. from V2R-expressing neurons. Furthermore,dendrites of Me neurons change their morphology under hormonalinfluence (Gomez & Newman, 1991). Therefore, the possibility arisesthat neurons normally receiving two inputs in layer I receive only oneinput from V2R receptor neurons if the dendritic tree is retracted underhormonal influence. It would be very interesting to check thishypothesis with methods similar to that used in the olfactory cortexwith combinatorial mixes of odours (Zou & Buck, 2006), but usingstimuli specific for one or two types of vomeronasal receptors.Collectively, these data open a new dual perspective to the study of thevomeronasal system in the basal telencephalon.
Acknowledgements
The authors thank Dr Mimi Halpern for critical review of the manuscript,Mr Ichiro Ikuta for help with English grammar, members of the LNH fortechnical and scientific support, and Marıa Picazo and Dr Garcıa Olmo for theirhelp with animals. This work has been supported by grants GC04007 andC03 ⁄ 06 to R.I., and grant PAC-05-007 to A.M.M.
Abbreviations
AAd, dorsal anterior amygdala; AAv, ventral anterior amygdala; ACo, anteriorcortical amygdaloid nucleus; AM, anteromedial thalamic nucleus; AOB (t, r, c),accessory olfactory bulb (total, rostral and caudal, respectively); Apir,amygdalopiriform transition area; BAOT, bed nucleus of the accessoryolfactory tract; BDA, biotinylated-labelled dextran-amine; BLA, basolateralamygdaloid nucleus, anterior part; BMA, basomedial amygdaloid nucleus,anterior part; BST, bed nucleus of the stria terminalis; BSTMPM, bed nucleusof the stria terminalis, medial division, posteromedial part; BSTS, supracap-sular bed nucleus of the stria terminalis; CA3, field CA3 hippocampus;Ce, central amygdaloid nucleus; CPu, caudate putamen; CTB, cholera toxin-Bsubunit; CxA., cortex-amygdala transition zone; Den, dorsal endopiriformnucleus; DG, dentate gyrus; ec, external capsule; EGP, external globus pallidus;FB, Fast-Blue; FDA, fluorescein-labelled dextran-amine; FG, Fluoro-Gold;Fr, frontal cortex; GlA, glomerular layer of the AOB; GnRH, gonadotropin-releasing hormone; Goa, Goa-protein; GrA, granule cell layer of the AOB; fi,fimbria of hippocampus; ic, internal capsule; IM, intercalated amygdaloidnucleus; IPAC, interstitial nucleus of the posterior limb of the anteriorcommissure; LDVL, laterodorsal thalamic nucleus, ventrolateral part; lo, lateralolfactory tract; LOT, nucleus of the lateral olfactory tract; Me, medialamygdaloid complex; MeAD, anterodorsal medial amygdaloid nucleus; MeAV,anteroventral medial amygdaloid nucleus; MePD, posterodorsal medial amy-gdaloid nucleus; MePV, posteroventral medial amygdaloid nucleus; MiA,mitral cell layer of the AOB; MOB, main olfactory bulb; och, optic chiasm; opt,optic tract; Pir, piriform cortex; PLCo, posterolateral cortical amygdaloidnucleus; PMCo, posteromedial cortical amygdaloid nucleus; RDA, tetrameth-ylrhodamine-labelled dextran-amine; RF, rhinal fissure; Rt, reticular thalamicnucleus; sm, stria medullaris; SO, supraoptic nucleus; st, stria terminalis; VPL,ventral posterolateral thalamic nucleus; VPM, ventral posteromedial thalamicnucleus; VS, ventral subiculum; I, II, III, cortical cell layers.
References
Alheid, G.F., Beltramino, C.A., de Olmos, J.S., Forbes, M.S., Swanson, D.J. &Heimer, L. (1998) The neuronal organization of the supracapsular part of thestria terminalis in the rat: the dorsal component of the extended amygdala.Neuroscience, 84, 967–996.
Baxi, K.N., Dorries, K.M. & Eisthen, H.L. (2006) Is the vomeronasal systemreally specialized for detecting pheromones? Trends Neurosci., 29, 1–7.
Belluscio, L., Koentges, G., Axel, R. & Dulac, C. (1999) A map of pheromonereceptor activation in the mammalian brain. Cell, 97, 209–220.
Berghard, A. & Buck, L.B. (1996) Sensory transduction in vomeronasalneurons: evidence for G alpha o, G alpha i2, and adenylyl cyclase II as majorcomponents of a pheromone signaling cascade. J. Neurosci., 16, 909–918.
Boehm, U., Zou, Z. & Buck, L.B. (2005) Feedback loops link odor andpheromone signaling with reproduction. Cell, 123, 683–695.
2078 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

Broadwell, R.D. (1975) Olfactory relationships of the telencephalon anddiencephalon in the rabbit. I. An autoradiographic study of the efferentconnections of the main and accessory olfactory bulbs. J. Comp. Neurol.,163, 329–345.
von Campenhausen, H. & Mori, K. (2000) Convergence of segregatedpheromonal pathways from the accessory olfactory bulb to the cortex in themouse. Eur. J. Neurosci., 12, 33–46.
Canteras, N.S., Simerly, R.B. & Swanson, L.W. (1995) Organization ofprojections from the medial nucleus of the amygdala: a PHAL study in therat. J. Comp. Neurol., 360, 213–245.
Collado, P., Guillamon, A., Valencia, A. & Segovia, S. (1990) Sexualdimorphism in the bed nucleus of the accessory olfactory tract in the rat.Brain Res. Dev. Brain Res., 56, 263–268.
Coolen, L.M. & Wood, R.I. (1998) Bidirectional connections of the medialamygdaloid nucleus in the Syrian hamster brain: simultaneous anterogradeand retrograde tract tracing. J. Comp. Neurol., 399, 189–209.
Davis, B.J., Macrides, F., Youngs, W.M., Schneider, S.P. & Rosene, D.L.(1978) Efferents and centrifugal afferents of the main and accessory olfactorybulbs in the hamster. Brain Res. Bull., 3, 59–72.
Del Cerro, M.C., Izquierdo, M.A., Collado, P., Segovia, S. & Guillamon, A.(1991) Bilateral lesions of the bed nucleus of the accessory olfactory tractfacilitate maternal behavior in virgin female rats. Physiol. Behav., 50, 67–71.
Del Punta, K., Leinders-Zufall, T., Rodriguez, I., Jukam, D., Wysocki, C.J.,Ogawa, S., Zufall, F. & Mombaerts, P. (2002a) Deficient pheromoneresponses in mice lacking a cluster of vomeronasal receptor genes. Nature,419, 70–74.
Del Punta, K., Puche, A., Adams, N.C., Rodriguez, I. &Mombaerts, P. (2002b) Adivergent pattern of sensory axonal projections is rendered convergent bysecond-order neurons in the accessory olfactory bulb.Neuron, 35, 1057–1066.
Devor, M. (1976) Fiber trajectories of olfactory bulb efferents in the hamster.J. Comp. Neurol., 166, 31–47.
Dong, H.W., Petrovich, G.D. & Swanson, L.W. (2001) Topography ofprojections from amygdala to bed nuclei of the stria terminalis. Brain Res.Brain Res. Rev., 38, 192–246.
Dong, H.W. & Swanson, L.W. (2004) Projections from bed nuclei of the striaterminalis, posterior division: implications for cerebral hemisphere regulationof defensive and reproductive behaviors. J. Comp. Neurol., 471, 396–433.
Dulac, C. & Axel, R. (1995) A novel family of genes encoding putativepheromone receptors in mammals. Cell, 83, 195–206.
Gomez, D.M. & Newman, S.W. (1991) Medial nucleus of the amygdala in theadult Syrian hamster: a quantitative Golgi analysis of gonadal hormonalregulation of neuronal morphology. Anat. Rec., 231, 498–509.
Gomez, D.M. & Newman, S.W. (1992) Differential projections of the anteriorand posterior regions of the medial amygdaloid nucleus in the Syrianhamster. J. Comp. Neurol., 317, 195–218.
Halpern, M. (1987) The organization and function of the vomeronasal system.Annu. Rev. Neurosci., 10, 325–362.
Halpern, M., Jia, C. & Shapiro, L.S. (1998a) Segregated pathways in thevomeronasal system. Microsc. Res. Techn., 41, 519–529.
Halpern, M. & Martinez-Marcos, A. (2003) Structure and function of thevomeronasal system: an update. Prog. Neurobiol., 70, 245–318.
Halpern, M., Shapiro, L.S. & Jia, C. (1995) Differential localization of Gproteins in the opossum vomeronasal system. Brain Res., 677, 157–161.
Halpern, M., Shapiro, L.S. & Jia, C. (1998b) Heterogeneity in the accessoryolfactory system. Chem. Senses, 23, 477–481.
Herrada, G. & Dulac, C. (1997) A novel family of putative pheromonereceptors in mammals with a topographically organized and sexuallydimorphic distribution. Cell, 90, 763–773.
Jia, C. & Halpern, M. (1996) Subclasses of vomeronasal receptor neurons:differential expressionofGproteins (Gi alpha2 andG (o alpha)) and segregatedprojections to the accessory olfactory bulb. Brain Res., 719, 117–128.
Jia, C. & Halpern, M. (1997) Segregated populations of mitral ⁄ tufted cells inthe accessory olfactory bulb. Neuroreport, 8, 1887–1890.
Kaneko, T., Saeki, K., Lee, T. & Mizuno, N. (1996) Improved retrogradeaxonal transport and subsequent visualization of tetramethylrhodamine(TMR)-dextran amine by means of an acidic injection vehicle and antibodiesagainst TMR. J. Neurosci. Meth., 65, 157–165.
Keverne, E.B. (1999) The vomeronasal organ. Science, 286, 716–720.Kimoto, H., Haga, S., Sato, K. & Touhara, K. (2005) Sex-specific peptides
from exocrine glands stimulate mouse vomeronasal sensory neurons. Nature,437, 898–901.
Kobbert, C., Apps, R., Bechmann, I., Lanciego, J.L., Mey, J. & Thanos, S.(2000) Current concepts in neuroanatomical tracing. Prog. Neurobiol., 62,327–351.
Kollack-Walker, S. & Newman, S.W. (1997) Mating-induced expression ofc-fos in the male Syrian hamster brain: role of experience, pheromones, andejaculations. J. Neurobiol., 32, 481–501.
Komisaruk, B.R., Rosenblatt, J.S., Barona, M.L., Chinapen, S., Nissanov, J.,O’Bannon, R.T., Johnson, B.M. & Del Cerro, M.C. (2000) Combined c-fosand 14C-2-deoxyglucose method to differentiate site-specific excitation fromdisinhibition: analysis of maternal behavior in the rat. Brain Res., 859, 262–272.
Lehman, M.N. & Winans, S.S. (1982) Vomeronasal and olfactory pathways tothe amygdala controlling male hamster sexual behavior: autoradiographicand behavioral analyses. Brain Res., 240, 27–41.
Leinders-Zufall, T., Lane, A.P., Puche, A.C., Ma, W., Novotny, M.V., Shipley,M.T. & Zufall, F. (2000) Ultrasensitive pheromone detection by mammalianvomeronasal neurons. Nature, 405, 792–796.
Lin, W., Arellano, J., Slotnick, B. & Restrepo, D. (2004) Odors detected bymice deficient in cyclic nucleotide-gated channel subunit A2 stimulate themain olfactory system. J. Neurosci., 24, 3703–3710.
Liu, Y.C., Salamone, J.D. & Sachs, B.D. (1997) Lesions in medial preoptic areaand bed nucleus of stria terminalis: differential effects on copulatorybehavior and noncontact erection in male rats. J. Neurosci., 17, 5245–5253.
Luo, M., Fee, M.S. & Katz, L.C. (2003) Encoding pheromonal signals in theaccessory olfactory bulb of behaving mice. Science, 299, 1196–1201.
Malnic, B., Hirono, J., Sato, T. & Buck, L.B. (1999) Combinatorial receptorcodes for odors. Cell, 96, 713–723.
Martinez-Marcos, A. & Halpern, M. (1999a) Differential centrifugal afferentsto the anterior and posterior accessory olfactory bulb. Neuroreport, 10,2011–2015.
Martinez-Marcos, A. & Halpern, M. (1999b) Differential projections from theanterior and posterior divisions of the accessory olfactory bulb to the medialamygdala in the opossum, Monodelphis domestica. Eur. J. Neurosci., 11,3789–3799.
Matsunami, H. & Buck, L.B. (1997) A multigene family encoding a diversearray of putative pheromone receptors in mammals. Cell, 90, 775–784.
Mori, K., von Campenhause, H. & Yoshihara, Y. (2000) Zonal organization ofthe mammalian main and accessory olfactory systems. Philos. Trans. R. Soc.Lond. B Biol. Sci., 355, 1801–1812.
Mucignat-Caretta, C., Colivicchi, M.A., Fattori, M., Ballini, C., Bianchi, L.,Gabai, G., Cavaggioni, A. & Della, C.L. (2006) Species-specificchemosignals evoke delayed excitation of the vomeronasal amygdala infreely-moving female rats. J. Neurochem., 99, 881–891.
Nance, D.M. & Burns, J. (1990) Fluorescent dextrans as sensitive anterogradeneuroanatomical tracers: applications and pitfalls. Brain Res. Bull., 25, 139–145.
Newman, S.W. (1999) The medial extended amygdala in male reproductivebehavior. A node in the mammalian social behavior network. Ann. N.Y.Acad. Sci., 877, 242–257.
de Olmos, J.S. (1972) The amygdaloid projection field in the rat as studied withthe cupric silver method. In Eleftheriou, B. (Ed.), The Neurobiology of theAmygdala. Plenum Press, New York, pp. 145–204.
de Olmos, J.S., Beltramino, C.A. & Alheid, G. (2004) Amygdala and extendedamygdala of the rat: a cytoarchitectonical, fibroarchitectonical, andchemoarchitectonical survey. In Paxinos, G. (Ed.), The Rat Nervous System.Elsevier, San Diego, pp. 509–603.
de Olmos, J.S. & Ingram, W.R. (1972) The projection field of the striaterminalis in the rat brain. An experimental study. J. Comp. Neurol., 146,303–334.
Paxinos, G. & Watson, C. (2005) The Rat Brain in Stereotaxic Coordinates.Elsevier, San Diego.
Price, J.L. (1987) The central olfactory and accessory olfactory systems. InFinger, T.T. & Silver, W.L. (Eds), Neurobiology of Taste and Smell. Wiley,New York, pp. 179–204.
Raisman, G. (1972) An experimental study of the projection of the amygdala tothe accessory olfactory bulb and its relationship to the concept of a dualolfactory system. Exp. Brain Res., 14, 395–408.
Reiner, A., Veenman, C.L., Medina, L., Jiao, Y., Del Mar., N. & Honig, M.G.(2000) Pathway tracing using biotinylated dextran amines. J. Neurosci.Meth., 103, 23–37.
Restrepo, D., Arellano, J., Oliva, A.M., Schaefer, M.L. & Lin, W. (2004)Emerging views on the distinct but related roles of the main and accessoryolfactory systems in responsiveness to chemosensory signals in mice. Horm.Behav., 46, 247–256.
Richmond, F.J., Gladdy, R., Creasy, J.L., Kitamura, S., Smits, E. &Thomson, D.B. (1994) Efficacy of seven retrograde tracers, compared inmultiple-labelling studies of feline motoneurones. J. Neurosci. Meth., 53,35–46.
Segregated pathways to the vomeronasal amygdala 2079
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080

Rodriguez, I., Feinstein, P. & Mombaerts, P. (1999) Variable patterns of axonalprojections of sensory neurons in the mouse vomeronasal system. Cell, 97,199–208.
Ryba, N.J. & Tirindelli, R. (1997) A new multigene family of putativepheromone receptors. Neuron, 19, 371–379.
Salazar, I. & Brennan, P.A. (2001) Retrograde labelling of mitral ⁄ tufted cells inthe mouse accessory olfactory bulb following local injections of thelipophilic tracer DiI into the vomeronasal amygdala. Brain Res., 896, 198–203.
Scalia, F. & Winans, S.S. (1975) The differential projections of the olfactorybulb and accessory olfactory bulb in mammals. J. Comp. Neurol., 161, 31–55.
Schmued, L., Kyriakidis, K. & Heimer, L. (1990) In vivo anterograde andretrograde axonal transport of the fluorescent rhodamine-dextran-amine,Fluoro-Ruby, within the CNS. Brain Res., 526, 127–134.
Shammah-Lagnado, S.J., Beltramino, C.A., McDonald, A.J., Miselis, R.R.,Yang, M., de Olmos, J., Heimer, L. & Alheid, G.F. (2000) Supracapsular bednucleus of the stria terminalis contains central and medial extended amygdalaelements: evidence from anterograde and retrograde tracing experiments inthe rat. J. Comp. Neurol., 422, 533–555.
Shammah-Lagnado, S.J. & Negrao, N. (1981) Efferent connections of theolfactory bulb in the opossum (Didelphis marsupialis aurita): a Fink-Heimerstudy. J. Comp. Neurol., 201, 51–63.
Shinohara, H., Asano, T. & Kato, K. (1992) Differential localization ofG-proteins Gi and Go in the accessory olfactory bulb of the rat. J. Neurosci.,12, 1275–1279.
Shipley, M.T., Ennis, M. & Puche, A.C. (2004) Olfactory system. In Paxinos,G. (Ed.), The Rat Nervous System. Elsevier, San Diego, pp. 923–964.
Skeen,L.C.&Hall,W.C. (1977)Efferentprojectionsof themainand theaccessoryolfactory bulb in the tree shrew (Tupaia glis). J. Comp. Neurol., 172, 1–35.
Sugai, T., Sugitani, M. & Onoda, N. (1997) Subdivisions of the guinea-pigaccessory olfactory bulb revealed by the combined method with immuno-histochemistry, electrophysiological, and optical recordings. Neuroscience,79, 871–885.
Veenman, C.L., Reiner, A. & Honig, M.G. (1992) Biotinylated dextran amineas an anterograde tracer for single- and double-labeling studies. J. Neurosci.Meth., 41, 239–254.
Vinader-Caerols, C., Collado, P., Segovia, S. & Guillamon, A. (1998) Sexdifferences in the posteromedial cortical nucleus of the amygdala in the rat.Neuroreport, 9, 2653–2656.
Wagner, S., Gresser, A.L., Torello, A.T. & Dulac, C. (2006) A multireceptorgenetic approach uncovers an ordered integration of VNO sensory inputs inthe accessory olfactory bulb. Neuron, 50, 697–709.
Weller, K.L. & Smith, D.A. (1982) Afferent connections to the bed nucleus ofthe stria terminalis. Brain Res., 232, 255–270.
Winans, S.S. & Scalia, F. (1970) Amygdaloid nucleus: new afferent input fromthe vomeronasal organ. Science, 170, 330–332.
Wood, R.I. & Swann, J.M. (2005) The bed nucleus of the stria terminalis in theSyrian hamster: subnuclei and connections of the posterior division.Neuroscience, 135, 155–179.
Zou, Z. & Buck, L.B. (2006) Combinatorial effects of odorant mixes inolfactory cortex. Science, 311, 1477–1481.
2080 A. Mohedano-Moriano et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2065–2080
Copyright © 2022 FDOKUMEN




















