
Spécificité de Zymoseptoria tritici en Tunisie
169
INSTITUT NATIONAL AGRONOMIQUE DE TUNISIE THESE DE DOCTORAT EN SCIENCES AGRONOMIQUES Spécialité : Science de la Production Végétale Thème : Préparée et présentée publiquement le 10 Mars 2020 par : Rim Bel Hadj Chedli Ep Tayari Devant le Jury : Pr. Faycal Ben Jeddi (INAT) : Président de Jury Dr. Hanène Chaabane (INAT) : Directrice de thèse Pr. Walid Hamada (INAT) : Rappoteur Dr. Naceur Djbali (CBBC) : Rapporteur Pr. Sonia Hamza (INAT) : Examinatrice Dr. Sarrah Ben M’barek (CRRGC) : Invitée Ministère de l’Enseignement Supérieur et de la Recherche Scientifique ***** Université de Carthage Spécificité de Zymoseptoria tritici en Tunisie Ministère de l’Agriculture, des Ressources Hydrauliques et de la Pêche ***** Institution de la Recherche et de l’Enseignement Supérieur Agricoles REPUBLIQUE TUNISIENNE
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Spécificité de Zymoseptoria tritici en Tunisie

INSTITUT NATIONAL AGRONOMIQUE DE TUNISIE
THESE DE DOCTORAT EN SCIENCES AGRONOMIQUES
Spécialité : Science de la Production Végétale
Thème :
Préparée et présentée publiquement le 10 Mars 2020 par :
Rim Bel Hadj Chedli Ep Tayari
Devant le Jury :
Pr. Faycal Ben Jeddi (INAT) : Président de Jury
Dr. Hanène Chaabane (INAT) : Directrice de thèse
Pr. Walid Hamada (INAT) : Rappoteur
Dr. Naceur Djbali (CBBC) : Rapporteur
Pr. Sonia Hamza (INAT) : Examinatrice
Dr. Sarrah Ben M’barek (CRRGC) : Invitée
Ministère de l’Enseignement
Supérieur et de la Recherche
Scientifique
*****
Université de Carthage
Spécificité de Zymoseptoria tritici en
Tunisie
Ministère de l’Agriculture, des Ressources
Hydrauliques et de la Pêche
*****
Institution de la Recherche et de
l’Enseignement Supérieur Agricoles
REPUBLIQUE TUNISIENNE


DEDICACES
Je tiens tout d’abord à remercier Dieu, qui m’a donné la force
et la patience d’accomplir ce modeste travail
Je dédie cette thèse
A mes chers parents, Jamel et Rekaya
A mon frère Mohamed et à ma sœur Raja
A mon cher mari Wassim
A mon cher fils Iskander
A toute ma famille
Merci pour votre amour, patience, encouragements et soutien
qui ont toujours illuminé mon chemin …
J’ai de la chance de vous avoir dans ma vie….
Que Dieu vous protège
RIM

Remerciements
Ce travail de thèse a été réalisé au sein du Laboratoire de génétique et d’amélioration des
plantes de l’Institut National Agronomique de Tunisie (INAT) en collaboration avec la plateforme
septoriose « Tunisia STB-Precision Phenotyping Platform (TunSPP) » dirigée par le Centre
International du Maïs et du Blé ‘CIMMYT’.
Au terme de ce travail, je tiens à remercier ma directrice de thèse Dr. Hanène Chaabane ;
Maître de conférences à l’INAT de m'avoir accueillie dans son équipe malgré ses occupations
professionnelles. Je tiens à rendre hommage à la pertinence de ses conseils et ses remarques
constructives au cours de la rédaction de ce mémoire.
Je remercie très sincèrement Pr. Salah Rezgui, mon ancien directeur de thèse retraité pour
m’avoir encadrée durant toutes ces années de thèse et pour ses conseils prodigués au cours de la
réalisation de ce travail.
Mes profonds remerciements et ma vive gratitude s’adressent aussi au Pr. Amor Yahyaoui,
Consultant et Coordinateur de Septoria Phenotyping Platform, pour son implication personnelle, son
soutien matériel et moral, ses orientations qui ont ponctué ces années de thèse et tout le temps qu'il
m'a consacré malgré ses nombreuses occupations. Je souhaiterais ici le témoigner ma sincère
reconnaissance pour tous les conseils et les remarques objectives qu’il m’a apporté.
Je remercie chaleureusement Dr. Sarrah Ben M’barek Maitre-assistant à CRRGC (Béjà), pour
sa patience, son aide, ses encouragements et ses conseils précieux. Qu’elle trouve ici l’expression de
toute ma reconnaissance, de ma profonde admiration et de ma respectueuse considération.
Ma grande reconnaissance s’adresse aussi au Pr. Gerrit Kema du ‘Wageningen University and
Research Center’, et à Dr. Lamia Aouini de ‘Purdue University’ pour leur contribution dans les
analyses moléculaires. Qu’ils trouvent ici mes chaleureux remerciements.
J’adresse mes plus sincères remerciements également à Dr. Bochra Amina Bahri de l’université
de Georgia-INAT, Dr. Zakaria Kehel de l’ICARDA-Maroc et Amir Souissi doctorant à l’INAT pour
leur aide dans les analyses statistiques et bio-informatiques. Je souhaiterais leur témoigner ma grande
gratitude.
Une pensée bien particulière à Dr. Sameh Boukef, Maitre-assistant à L’Institut Supérieur
Agronomique de Chott Meriem, qui m’a beaucoup encadrée, m’a appris les techniques d’isolements
du pathogène et m’a aimablement guidée vers les premiers pas de la recherche. Qu’elle trouve ici ma
grande reconnaissance.
Je tiens également à adresser mes vifs remerciements aussi à Dr Abdennour Sbei du CRRGC
(Béjà), Dr Fatiha Ben Tata de INRA Maroc et Dr. Abdelkader Benbelkacem de INRA Algérie d’avoir
assuré les semences de blé utilisées dans cette étude.
Je remercie tout particulièrement Pr. Faycel Ben Jeddi d’avoir accepté de juger et présider cette
thèse.

Je tiens à remercier également mes deux rapporteurs, Pr. Walid Hamada et Dr. Naceur Djbali
pour le temps qu’ils ont consacré à ce travail, et pour l’intérêt qu’ils ont bien voulu porter au
manuscrit.
Je remercie très sincèrement Pr. Sonia Hamza pour l’effort qu’elle a fourni pour examiner ce
travail.
Je n’oublierai pas d’adresser mes remerciements à tous les membres de la plateforme
septoriose en Tunisie, Maroua Laribi, Lamia Ben Naceur, Hajer Kouki, et Radhouan… pour leur
aide et leur soutien.
Mes remerciements seraient bien incomplets si ma grande famille n’y était pas associée. Merci
donc à mes chers parents Jamel et Rekaya, ma sœur Raja et mon frère Mohamed, mon petit garçon
Iskander, aux Familles Bel Hadj Chedli, Tayari et Ben Taher pour m’avoir soutenue et avoir partagé
ce parcours avec moi. Cette thèse leur est dédiée en reconnaissance des efforts et des sacrifices
consentis durant toutes ces années.
Mes remerciements les plus particulières s’adressent à mon cher mari Wassim pour ses
consolations pendant mes moments de détresse, son soutien sans faille, sa grande indulgence, sa
compréhension et surtout sa contribution dans la réussite de ce travail. Qu’il trouve ici ma plus grande
reconnaissance.
Je ne saurais terminer cette liste de remerciements sans évoquer les efforts de mon cher oncle
Mouhamed Taher pour son dévouement et sa disponibilité dans la réalisation de l’enquête et les essais
de plein champ à El Haouaria.
Enfin, à tous ceux qui ont contribué de près ou de loin à la réalisation de ce modeste travail de
recherche et dont le nom m’échappe à cet instant et que je regretterais de n’avoir pas cités, tous mes
remerciements.
RIM

Résumé
La septoriose du blé (Septoria tritici blotch (STB)) causée par Zymoseptoria tritici (Z. tritici)
reste la maladie la plus dommageable des cultures du blé dur en Tunisie. Etant donné que cette maladie
fut observée annuellement sur blé dur, peu de travaux sont disponibles sur la distribution et
l’occurrence de ce pathogène chez le blé tendre en Tunisie.
Ainsi, l’enquête menée au Nord et au Nord-Ouest de la Tunisie a révelé des moyennes
d’incidence et de sévérité les plus élevées sur blé dur, à Jendouba, Bizerte, Béjà et au Kef. D’autre part,
sur blé tendre, les moyennes d’incidences et de sévérités les plus importantes ont été signalées à El
Haouaria particulièrement sur une ancienne variété de blé tendre désignée localement par ‘Farina Arbi’.
Des incidences importantes ont été également notées sur triticale dans certaines régions.
Ensuite, l’évaluation de la résistance relative à STB de 89 variétés de blé dur et tendre a indiqué
que la majorité des variétés de blé dur Tunisien, Marocain et Algérien sont classées comme sensibles
et hautement sensibles dans les deux sites (Béjà et Cap Bon) avec des moyennes du rAUDPC allant de
0,5 à 0,8 et des valeurs de N et PC variant entre 30 et 65%, et 40 et 75% respectivement. Toutefois, le
blé tendre Marocain était sensible dans les régions du Cap Bon alors que le blé tendre Tunisien a été
classé comme résistant à totalement résistant à l’exception de la ‘Farina Arbi’, sensible au Cap Bon et
résistante à Béjà.
Le génotypage moyennant 12 marqueurs microsatellites (SSR) a englobé une collection de 184
isolats mono-pycnidiaux de Z. tritici issues du blé tendre (El Haouaria, Béjà et Jendouba) et 65 isolats
échantillonnés de blé tendre, blé dur et triticale cultivés dans le même champ à El Haouaria. Au niveau
régional, la région d’El Haouaria présente les indices de Nei’s (0,42), de Shannon (0,84) et le nombre
d’allèles privés (36) les plus importants. En outre, une différenciation modérée entre les populations
(Fst=0,16) et un important flux de gène (Nm = 1,85) ont été ainsi signalés. De point de vue espèces, la
diversité génétique de Nei’s (0,52), la diversité génétique non biaisée (0,58) et richesse allélique (4,43)
sont égales.
Par conséquent, cette étude a démontré d’une part que la diversité et la structure des populations
de Z. tritici semblent ne pas être affectées par l’espèce hôte à l’échelle de la parcelle, et elle a dévoilé
d’autre part une absence de structure des populations de Z. tritici en Tunisie.
Mots-clés: Blé tendre, blé dur, triticale, El Haouaria, diversité génétique, structure des populationst, Z.
tritici.

Abstract
Septoria tritici blotch (STB) caused by Zymoseptoria tritici (Z. tritici) has become an inherent
disease of durum wheat in Tunisia. Although Septoria was observed on durum wheat annually, up to
now not much is known on the occurence of STB on bread wheat. In this study, the STB survey
conducted in North and Northwestern Tunisia revealed that important incidence and severity were
recorded on durum wheat at Jendouba, Bizerte, Béjà, and Kef while STB was nearly absent in these
regions on bread wheat. However, the highest mean incidence and severity were recorded at El
Haouaria region mainly on bread wheat landrace of unknown origin called ‘Farina Arbi’. STB was
noted also on Triticale during the survey.
In addition, the performance of 89 wheat varieties from Morocco, Algeria and Tunisia were
screened in Tunisia for their relative resistance to STB at Cap Bon and Béjà regions. Results indicated
that the majority of Tunisian, Algerian and Moroccan durum wheat varieties were susceptible to STB
at both locations ranging from susceptible to highly susceptible where the average of the relative Area
inder disease progresse curve (rAUDPC) was ranged from 0.5 to 0.8 and the Necrosis (N) and the
Pycnidial coverage (CP) values were ranged from, 30 to 65 % and 40 to 75% respectively. The
Moroccan bread wheat varieties were susceptible in Cap Bon area, while the commercial Tunisian
bread wheat were resistant at both locations with the exception of “Farina Arbi” which was susceptible
at El Haouaria and resistant at Béjà.
Genotyping using 12 polymorphic microsatellite (SSR) was carried out using a set of 184 single-
pycnidial isolates sampled from infected bread wheat fields at EL Haouaria, Béjà and Bizerte, and 65
isolates from infected wheat species: durum wheat, bread wheat and triticale from the same field at
Cap Bon Area. At regional level, the highest Nei’s index value (0.42), Shannon Index (0.84), and the
private alleles number (36) were observed at El Haouaria region. Moderate population differentiation
(Fst=0.16) and a high gene flow (Nm =1.85) were then observed between the sampled fields. However,
at species level, an equal Nei’s gene diversity (0.52), unbiased gene diversity (0.58) and allele richness
(4.43) within Z. tritici-durum and bread wheat populations were observed. Therefore, weak population
differentiation (0.13) between species population explained by high levels of gene flow (3.26) were
then observed. The high degree of diversity could be due to and active sexual reproduction. Overall,
results of this study revealed an absence of relationship between genetic diversity and structure and
wheat-host species at regional and field level in Tunisia.
Key-words: Bread wheat, durum wheat, triticale, El Haouaria, genetic diversity, genetic structure, Z.
tritici

ملخص
لدى القمح السبتوري من أهم األمراض شيوعا ع المتسبب في مرض التبق ' Zymoseptoria tritici (Z. tritici)يعتبر الفطر'
الرغم من تسجيل أعراض هذا المرض على القمح الصلب سنويا في تونس إال وعلى. والتريتيكالأقل لدى القمح اللين الصلب وبصفة
.............................. . أن انتشاره على أصناف القمح اللين يضل مجهوال.
2017-2016و 2016-2015الدراسة االستقصائية لمائة وستة وعشرون حقل خالل الفصول الفالحية أولى كشفتة في مرحل
مرتفعة في كل من: باجة، جندوبة، بنزرت والكاف وخطورةعن حساسية كامل أصناف القمح الصلب للتبقع السبتوري مع معدالت حد ة
ناطق باستثناء صنف واحد 'فارينة عربي' الذي أثبت حساسية عالية في منطقة الهوارية مقابل مقاومة أنواع القمح اللين للمرض بهذه الم
............................................... .أين يتم إنتاجه. كما أثبتت هذه الدراسة أيضا مدى حساسية الترتيكال لهذا المرض في عدة مناطق
أنه يمكن والمغرب والجزائرمن تونس واللينلى عدة أصناف من القمح الصلب في مرحلة ثانية أثبتت هذه الدراسة المنجزة ع
(rAUDPC) لتطورالمرضالرسم البياني تحت المساحة بمعدالت والهواريةتصنيفها من حساس إلى حساس جدا في منطقتي باجة
و بين %75و %40متراوحة على التوالي بين (N) النخرية والمنطقة(PC) بالبيكنديا المساحة المغطات ومعدالت 0,8إلى 0,5من
الصنف المغربي حساس مقارنة بالصنف التونسي الذي أبرز مقاومته التجارب أن أما بخصوص القمح اللين أظهرت .%65و 30%
على صنف 'فرينة عربي'. وخاصةالسبتوري في جل المناطق ما عدى منطقة الهوارية للتبق عبالنسبة
عزلة لدى 65عزلة من هذا فطر لدى القمح اللين من عدة مناطق و 184ٲثبت هذا البحث من ناحية أخرى مدى التنوع الجيني ل
Nei (0,42 ،)لتريتيكال من نفس الحقل. على مستوى الجهات، سجلت ٲعلى معدالت مؤشر عدة أنواع من القمح الصلب واللين واا
( ومؤشر 1,85( في الهوارية باإلضافة إلى معدل جيني كبير )36) Allèles privés(، ٲعلى ٲعداد 0,84) Shannonٲعلى مؤشر
سجلت معدالت متساوية من مؤشر ني ،مستوى األصناف(. أما على 0,16اختالف متوسط للتركيز السكاني بين مختلف الحقول )
....... (. 3,26( و ٲيضا معدل جيني كبير )4,43) Allèle(، ثراء 0,58) Shannon(، مؤشر 0,52)
وجود أخرى عدمجهة ومنبصفة عامة، أثبتت الدراسة من جهة عدم وجود عالقة بين التنوع الجيني وأصناف القمح المعتمدة
في تونس. سكانية للفطرتركيبة
Zymoseptoria tritici جينية، ةالجيني، تركيبالصلب، القمح اللين، الهوارية، التنوع ح: القمالمفاتيحالكلمات

Table des matières
RÉSUMÉ
ABSTRACT
ملخص
INTRODUCTION GÉNÉRALE 1
1. Importance du secteur céréalier dans le monde .................................................................................. 4
3. La septoriose du blé : Importance dans le Monde, au nord de l’Afrique et en Tunisie ...................... 7
4. Généralités sur la septoriose ............................................................................................................... 9
4.1. Classification et plantes hôtes .................................................................................................. 9
4.2. Symptômatologie ................................................................................................................... 10
4.3. Cycle biologique de Zymoseptoria tritici .............................................................................. 10
4.4. Hétérothallisme et reproduction sexuée................................................................................. 13
4.5. Intéraction entre Zymoseptoria tritici-blé et spécificité ......................................................... 13
4.6. Structure des populations de Zymoseptoria tritici ................................................................. 15
4.7. Diversité génétique des populations ...................................................................................... 16
4.8. Les forces évolutives affectant la diversité génétique des populations ................................. 16
5. Influence des facteurs climatiques sur le développement de la maladie .......................................... 17
CHAPITRE 2. OCCURRENCE OF SEPTORIA TRITICI BLOTCH (ZYMOSEPTORIA
TRITICI) DISEASE ON DURUM WHEAT, TRITICALE, AND BREAD WHEAT IN
NORTHERN TUNISIA 20
1. Introduction ...................................................................................................................................... 21
2. Materials and Methods ..................................................................................................................... 22
2.1. Study area description .......................................................................................................................... 22 2.2. Climatic conditions of the surveyed regions ........................................................................................ 23 2.3. Cereal crops .......................................................................................................................................... 24 2.4. Septoria leaf blotch disease assessment ............................................................................................... 25 2.5. Data analysis ........................................................................................................................................ 26
3. Results .............................................................................................................................................. 26
3.1. Incidence of wheat Septoria tritici blotch ............................................................................................. 26 3.2. Incidence of Septoria tritici blotch on commercial wheat varieties...................................................... 31
4. Discussion ........................................................................................................................................ 33
5. Conclusion ........................................................................................................................................ 34
CHAPITRE 3. GENETIC DIFFERENTIATION BETWEEN ‘ZYMOSEPTORIA TRITICI’
POPULATIONS SAMPLED FROM BREAD WHEAT IN TUNISIA REVEALED BY SSR
MARKERS……………………………………………………………………………………………36
1. Introduction ...................................................................................................................................... 36

2. Materials and Methods ..................................................................................................................... 38
2.1. Fungal sampling and isolation .............................................................................................................. 38 2.2. DNA extraction and mating type’s determination ................................................................................ 39
2.3. Microsatellites analysis ........................................................................................................................ 41 2.4. Data analysis .......................................................................................................................... 42
3. Results .............................................................................................................................................. 42
3.1. Genetic diversity within and among sampled populations ................................................................... 42 3.2. Genetic differentiation between populations ........................................................................................ 44 3.3. Relationship between geographic populations and genetic structure ................................................... 45 3.4. Occurrence of sexual reproduction ....................................................................................................... 47
4. Discussion ........................................................................................................................................ 48
5. Conclusion ........................................................................................................................................ 51
CHAPITRE 4. SCREENING FOR RESISTANCE OF TUNISIAN, MOROCCAN AND
ALGERIAN WHEAT VARIETIES TO ZYMOSEPTORIA TRITICI IN NORTHERN
TUNISIA………………………………………………………………………………………………53
1. Introduction ...................................................................................................................................... 53
2. Materials and Methods ..................................................................................................................... 55
2.1. Description of the study areas and experimental design ...................................................................... 55 2.2. Plant materials ...................................................................................................................................... 56 2.3. Evaluation of disease severity and area under disease progress curve ................................................. 59 2.4. Statistical analysis ................................................................................................................................ 59
3. Results .............................................................................................................................................. 60
3.1. Meteorological conditions during the crop cycle ................................................................................. 60 3.2. Genotype by region interaction ............................................................................................................ 60 3.3. Varieties response to STB .................................................................................................................... 62 3.4. Varieties classification ......................................................................................................................... 63 3.5. Significant correlation between quantitative traits ............................................................................... 67
4. Discussion ........................................................................................................................................ 68
5. Conclusion ........................................................................................................................................ 70
CHAPITRE 5. EFFECT OF HOST-WHEAT SPECIES ON GENETIC DIFFERENTIATION
OF ‘ZYMOSEPTORIA TRITICI’ AT SINGLE FIELD IN NORTHERN
TUNISIA………………………………………………………………………………………………72
1. Introduction ...................................................................................................................................... 72
2. Materiel and Methods ....................................................................................................................... 74
2.1. Wheat varieties and Z. tritici sampling................................................................................................. 74 2.2. Mating types determination .................................................................................................................. 75 2.3. Microsatellite analysis .......................................................................................................................... 76 2.4. Data analysis ........................................................................................................................................ 76
3. Results .............................................................................................................................................. 77
3.1. Distribution of mating-type alleles at single field ................................................................................ 77 3.2. Genetic variability in core chromosome according to host species ...................................................... 77 1.1. Genetic diversity and differentiation between varieties populations .................................................... 78 1.2. Lack of genetic structure within total population ................................................................................. 80
2. Discussion ........................................................................................................................................ 82

3. Conclusion ........................................................................................................................................ 85
DISCUSSION GÉNÉRALE ET CONCLUSION…………………………………………………..86
RÉFÉRENCES BIBLIOGRAPHIQUES……………………………………………………………93

Liste des Figures
CHAPITRE 1
Figure1. Etages bioclimatiques en Tunisie (Anonyme 1) .............................................................................. 7
Figure 2. Symptômes typiques de la septoriose du blé. (A) Les pycnides de la phase asexuée de Z.tritici (B)
(photo adapté par suffert et al.2016), (B) Taches nécrotiques avec de petits points brun foncés à noirs. (Photo
adapté par Gigot, 2013) ................................................................................................................................. 10
Figure3. Cycle biologique de Zymoseptoria tritici…………………………………………………………………12
Figure4. Schéma représentatif de la dynamique des ascospores (d'inoculum primaire) et celui des
pycnidiospores (inoculum secondaire) et leurs rôles dans l’induction d’une épidémie (Suffert et al.2016). 12
CHAPITRE 2
Figure 1. Map of Tunisia showing the location of survey areas across different climatic regions during 2016
and 2017 cropping seasons. Sub-humid: Cap Bon North (A), Bizerte (B) and Béjà (C). Semi-arid region of
Northern Tunisia: Cap Bon South (D), Manouba (E), Zaghouan (F), Jendouba (G), and Le Kef (H). ......... 23
Figure 2. Incidence of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops (bread
wheat, durum wheat and triticale). ................................................................................................................ 29
Figure 3. Severity of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops (bread wheat,
durum wheat and triticale). ........................................................................................................................... 30
Figure 4. Incidence of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops species
(bread wheat, durum wheat and triticale). ..................................................................................................... 30
Figure 5. Severity of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops (bread wheat,
durum wheat and triticale) ............................................................................................................................ 31
Figure 6. Incidence of STB on durum wheat, bread wheat and triticale varieties. ....................................... 32
Figure 7. Severity of STB on durum wheat, bread wheat, and triticale varieties. ........................................ 32
CHAPITRE 3
Figure 1. Schematic map of the sampling locations in Northern Tunisia. A: El Haouaria region, B:Bizete
(Ichkeul), C: Goubellat; D: Oued Zarga. ...................................................................................................... 40
Figure 2. Population structure of the 162 ‘Zymoseptoria tritici’ isolates sampled from four locations using
Structure software version 2.3.4 with K=4. .................................................................................................. 46
Figure. 3. Principal coordinates analysis (PCoA). Individuals from the same region are marked using the
same symbol. The first and second principal coordinates account for 15.07 % and 46.20 % of the variation,
respectively. .................................................................................................................................................. 46
Figure 4. Dendrogram showing the genetic clustering of the 162 Zymospetoria. Tritici isolates sampled from
bread wheat across three geographic locations in Northern Tunisia. The tree was constructed using the
weighted neighbor-joining method implemented in DARwin 6 software. Isolates from the same field were
indicated with the same color. Fields 1,2,3,4,5 and 6 are located at Ca Bon Area, fields 7 and 8 are located at
Béjà while field 9 belongs to Bizerte region. ................................................................................................ 47

CHAPITRE 4
Figure1. Map of Tunisia showing the location of study area (Béjà and Cap Bon area) during 2017-2018
cropping season. The STB experiments were set in an augmented design at both locations. ....................... 56
Figure 2. Variation of PC and N for all varieties in Cap Bon region during the two cropping seasons. TDM:
Tunisian durum wheat; TBW: Tunisian bread wheat; ADM: Algerian durum wheat; MBW: Moroccan durum
wheat; MDW: Moroccan durum wheat. ........................................................................................................ 62
Figure 3. Variation of PC and N across varieties in Béjà station. TDM: Tunisian durum wheat; TBW:
Tunisian bread wheat; ADM: Algerian durum wheat; MBW: Moroccan durum wheat; MDW: Moroccan
durum wheat.................................................................................................................................................. 63
Figure 4. PCA showing the major correlated variability of varieties as shown by axes 1 and 2. The first
Dimension1 accounted for 98% of the total variability expressed by quantitative traitswhile the second
component (Dimension2) accounts only 1.64% of the total variation. ACP revealed 6 clusters at Béjà region:
Cluster 1: Very resistant; Cluster 2: resistant; Cluster3: moderately resistant; Cluster4: moderately
susceptible; Cluster5: susceptible; Cluster 6: very susceptible. Details about genotypes of each group are
shown in table 3. ........................................................................................................................................... 65
Figure 5. PCA showing the major correlated variability of varieties as shown by axes 1 and 2 accounting
98% and 1.64% respectively of the total variability expressed by quantitative traits. ACP revealed 6 clusters
at Cap Bon region: Cluster 1: Very resistant; Cluster 2: resistant; Cluster3: moderately resistant; Cluster4:
moderately susceptible; Cluster5: susceptible; Cluster 6: very susceptible. Details about genotypes of each
group are shown in table 4. ........................................................................................................................... 67
Figure 6. Dimensional relationships among the measured parameters of STB infection showing a significant
correlation between AUDPC, rAUDPC, N and PC as revealed by principal component analyses over two
years. ............................................................................................................................................................ 68
CHAPITRE 5
Figure 1. Summary of different measured index: Genetic diversity (h); Shannon's Information Index (I), and
Unbiased genetic diversity (uh) across three wheat species. DW: durum wheat, BW: bread wheat and TRIT:
triticale. ......................................................................................................................................................... 81
Figure 2. PCoA analysis of 65 Z. tritici isolates sampled from three wheat species cropped in single field at
Cap Bon region during 2016-2017 cropping season. .................................................................................... 82
Figure 3. Lack of population structure as revealed by Structure software with K=2, K=3 and K=4 within a
total of 65 Zymoseptoria tritici population sampled from single field in Tunisia. ........................................ 83
Figure 4. Genetic clustering and relationships between 65 Z. tritici isolates sampled from bread wheat, durum
wheat and triticale from 22 wheat genotypes cultivated in Northern Tunisia. The tree was constructed using
the weighted neighbor-joining method implemented in DARwin 6 software. Isolates from bread wheat,
durum wheat and triticale were colored, black, red and blue respectively. Bootstrap values are indicated in
blue and references-isolates we designed with the green color. .................................................................... 84

Liste des Tableaux
CHAPTER 2
Table 1. Climatic conditions of inspected regions during the survey period. .................................................. 23
Table 2. Geographical coordinates of inspected regions during the survey period ......................................... 24
Table 3. Survey designated Classes for Septoria tritici blotch (STB) prevalence, severity and incidence. ..... 25
Table 4. Triticum species and wheat varieties identified during the survey during the two cropping seasons.27
Table 5. Prevalence of Septoria tritici blotch in inspected areas during 2015-2016. ....................................... 28
Table 6. Prevalence of Septoria tritici blotch in inspected areas during 2016-2017 …………………………28
CHAPITRE 3
Table 1. Origin, number and geographical coordinates of Zymoseptoria tritici isolates used in this study .... 40
Table 2. Mating type’s specific primers and microsatellites markers used in this study ................................. 41
Table 3.Genetic diversity based on twelve microsatellite markers related to Zymoseptoria tritici
populations/sub-populations collected from three regions: Cap Bon Area, Bizerte and Béjà (Goubellat and
Oued Zarga) in Northern Tunisia during 2015-2016. ...................................................................................... 44
Table 4. Analysis of molecular variance (AMOVA) of the bread-wheat Zymoseptoria tritici population ..... 45
Table 5. Distribution of Z.tritici mating types across different regions. .......................................................... 48
CHAPTER 4
Table1. Wheat varieties screened for resistance to Septoria tritici blotch disease during 2016-2017 and 2017-
2018 cropping seasons. .................................................................................................................................... 57
Table 2. ANOVA analysis for Pycnidial coverage (PC), Necrotic area (N) and the relative area under disease
progress curve rAUDPC for 89 wheat varieties at Béjà and Cap Bon regions ................................................ 61
Table 3. Means and ranges of AUDPC and rAUDPC for all identified clusters at Béjà region ...................... 64
Table 4. Means and ranges of AUDPC and rAUDPC for all identified clusters at Cap Bon region ............... 65
CHAPTER 5
Table 1. Summary of information about ‘Zymoseptoria tritici’ isolates investigated in this study. ................ 75
Table 2. Distribution of Z. tritici mating types within wheat species from single field at Cap Bon region. ... 77
Table 4. Genetic diversity of ‘Zymoseptoria tritici’ population, genetic differentiation between population and
gene flow. ........................................................................................................................................................ 79
Table 5. Analysis of Molecular Variance (AMOVA) for 65 Zymoseptoria tritici isolates ............................. 79

Liste des Abréviations
ADN :Acide désoxyribonucléique
AMOVA : Analysis of Molecular Variance
AFLP : amplified fragment length polymorphisms
BW: bread wheat
CIMMYT : Centre d’amélioration du blé et du maïs
DW: Durum wheat
°C : Degré Celsius
ddl : Degré de liberté
dNTP : Désoxyribonuléotide triphosphate
FST : differentiation entre les populations
H: diversité génétique
I : l’index de Shannon
ISEPTON : International Septoria Observation Nursery
Max : Maximum
Mat1-1: Mating type 1-1
Mat1-2: Mating type 1-2
Min : Minimum
Mm : Millimètre
Mn : flux de gène
PCR: Polymerase Chain Reaction
PDA Potato Dextrose Agar
P : polymorphisme
RFLP : amplified fragment length polymorphisms
STB : septoria tritici blotch
SNP Single Nucleotide Polymorphism
SSR : simple sequence repeats
Trit: triticale
UV: ultrat violet
X2 : Chi-carré

Introduction générale
1
Introduction Générale
Les céréales occupent aujourd’hui une place prépondérante aussi bien dans la production
agricole qu’agroalimentaire, à l’échelle nationale et internationale (Rastoin et Benabderrazek,
2014). Néanmoins, la production céréalière en Tunisie est marquée par une forte irrégularité
conditionnée par les aléas climatiques. La stabilité des productions céréalières constitue
actuellement une priorité de la stratégie tunisienne d’amélioration des céréales (El Felah et
Gharbi, 2014). Ainsi, l’optimisation des rendements nécessite la maîtrise d’un maximum de
facteurs de nature abiotiques (tels que la sécheresse et la salinité) et biotiques (comme les
insectes, les nématodes et les maladies foliaires causées par les virus et les champignons
phytopathogènes). La septoriose du blé est considérée parmis les maladies les plus importantes
en Tunisie qui semble entraver les productions céréalières depuis des années (Gharbi et al.,
2002).
L’émergence de la maladie des taches septorienne (STB), causée par Zymoseptoria tritici
(Z. tritici), a commencé avec la domestication du blé dans le croissant fertile à partir des
ancêtres, Z. ardabiliae et Z. pseudotritici qui ont été isolés des volontaires Elymus repens,
Dactylis glomerata et Lolium perenne en Iran (Ponomarenko et al., 2011 ; Stukenbrock et al.,
2011). Par la suite, ce pathogène a été largement distribué en causant une grande menace des
cultures du blé tendre et du blé dur dans le monde surtout dans les zones humides (Nord de la
France, Allemagne, Royaume-Uni…) (Linde et al., 2002 ; Zhan et al., 2003 ; Kabbage et al.,
2008 ; Singh et al., 2016). Toutefois, en Europe, STB est considérée comme la maladie la plus
importante économiquement causant des pertes annuelles allant jusqu’à 1 milliard d’euro
(Torriani et al., 2015). Les pertes de rendement sont estimées à 50% dans les champs non traités,
et de 5 à 10% dans les champs traités ou avec des variétés résistantes (Fones and Gurr, 2015).
Cette maladie a suscité beaucoup d’attention après la grande épidémie enregistrée durant
les années 1968-1969 en Afrique du Nord, particulièrement après l’introduction du blé semi-
nain ou Mexicain (Ammar et al., 2011) qui a conduit considérablement à d’énormes attaques
de Z. tritici au Maroc, en Algérie et en Tunisie (Yahyaoui et al., 2002).
En Tunisie des pertes de rendements allant de 30 à 50% ont été enregistrées sur blé dur
durant les années pluvieuses (Ben Mohamed et al., 2000), en particulier lorsque les pluies de
printemps persistent après l’émergence de la feuille drapeau (Suffert et al., 2013). En
particulier, le nord du pays, qui compte les emblavures les plus importantes de blé dur et fournit
à peu près 85% de la production nationale, connaît les attaques les plus sévères surtout dans les

Introduction générale
2
régions de Mateur, Bizerte, Béjà qui ont été signalées comme un « Hot Spot » pour la septoriose
(Gharbi et al., 2002 ; Fakhfekh et al., 2011 ; El Felah et Gharbi, 2014).
Ce pathogène était omniprésent dans les climats tempérés favorables pour les cultures de
blé tendre, dans les climats arides et semi-arides notamment dans le bassin méditerranéen et en
Afrique du Nord, mais également dans les climats chauds du Nord de l’Amérique (Linde et al.,
2002 ; Zhan et al., 2003 ; Singh et al., 2016). Par ailleurs, une grande spécificité de Z. tritici
aux divers écosystèmes agricoles a été notée (Eyal et al., 1985). D’une part, certains chercheurs
ont suggéré que cette spécificité est due à la grande plasticité de son génome illustré par le
grand nombre de chromosomes accessoires qui peuvent être perdus ou gagnés durant le cycle
d’infection (Stukenbrock et al., 2011 ; Gautier et al., 2014). D’autres part, une certaine
spécificité de Z. tritici envers le genre Triticum a été signalée grâce à la relation gène pour gène
et trois catégories d’isolats ont été ainsi rapportées : des isolats collectés de blé tendre qui ne
peuvent infecter que le blé tendre, des isolats isolés de blé dur qui ne s’attaquent qu’au blé dur
et ceux qui peuvent infester les deux espèces (Kema et al., 1996 a et b).
L’étude de l’effet de la spécificité du pathogène envers l’hôte sur la variation génétique
des populations paraît d’une grande importance. Dans le cas de Z. tritici, la structure génétique
des populations peut être altérée par des cycles réguliers de la reproduction sexuée (McDonald,
2016). Plusieurs marqueurs moléculaires à savoir : fragment length polymorphisms (RFLP) ;
amplified fragment length polymorphisms (AFLP) ; et simple sequence repeats (SSR), ont été
utilisés pour étudier et caractériser la génétique des populations de Z. tritici dans plusieurs pays
(Chen and McDonald, 1996 ; Linde et al., 2002 ; Zhan et al., 2003). Une grande diversité
génétique chez Z. tritici ainsi qu’un taux de migration très important chez des isolats de blé dur
ont été révélés en Tunisie (Boukef, 2012 ; Berrais et al., 2013 ; Nouari et al., 2016). Jusqu’à
présent, nous n’avons pas relevé de travaux qui ont étudié la diversité génétique de Z. tritici
isolée à partir du blé tendre en Tunisie.
C’est dans ce cadre que s’intègre ce travail de recherche qui constitue une première étude
de la distribution de la septoriose chez le blé tendre et le blé dur en Tunisie, de la spécificité de
ce pathogène en Tunisie, et de l’investigation de la diversité génétique d’une population de Z.
tritici isolée à partir des variétés Tunisiennes et étrangères de blé tendre, blé dur et triticale
cultivées en plein champ.

Introduction générale
3
Ce manuscrit est organisé en cinq chapitres :
Le premier chapitre consiste en une synthèse bibliographique qui relate l’importance de
la culture du blé, des connaissances générales sur la septoriose et l’importance de l’étude des
populations du pathogène pour développer une stratégie de lutte intégrée contre cette maladie.
Le deuxième chapitre se base sur une enquête effectuée dans les principales régions
céréalières du Nord du pays pour étudier la prévalence et la distribution géographique de la
septoriose chez le blé tendre et le blé dur en Tunisie ainsi que la spécificité de Z. tritici envers
la plante l’hôte.
Le troisième chapitre comporte une étude de la diversité génétique d’une population
Tunisienne de Z. tritici isolée à partir du blé tendre dans différents étages bioclimatiques en
Tunisie en adoptant une approche basée sur l’analyse du polymorphisme de séquences
microsatellites SSR.
Le quatrième chapitre s’intéresse à une étude épidémiologique de la septoriose dans des
régions céréalières considérées comme un hot-spot sur blé dur. Etant donné que ce pathogène
s’attaque plus au blé dur qu’au blé tendre en Tunisie et inversement au Maroc, deux hypothèses
seront vérifiées : S’agit-il de la susceptibilité des variétés marocaines de blé tendre et de la
tolérance des variétés Tunisiennes à ce pathogène ou bien plutôt de la spécificité du pathogène
envers l’espèce ‘Tritcum durum’ et ‘Triticum aestivum’ ?
Le cinquième chapitre présente une investigation de la diversité génétique d’une
collection de 65 isolats de Z. tritici isolée à partir des variétés Marocaines, Algériennes, et
Tunisiennes de blé tendre, blé dur et triticale cultivées en Tunisie suite à une infection naturelle.
Les questions suivantes ont été abordées : La structure des populations de Z. tritici change-t-
elle en fonction de l’espèce hôte et de la variété de la même espèce ? Quels sont les facteurs qui
peuvent expliquer des différenciations entre ces populations ?

Chapitre 1. Synthèse bibliographique

Chapitre 1. Synthèse Bibliographique
4
1. Importance du secteur céréalier dans le monde
Les céréales sont les principales sources de la nutrition humaine et animale dans le monde.
Le maïs, le blé et le riz sont les trois principales céréales cultivées et ils constituent 89% de la
production mondiale (Rastoin et Benabderrazek, 2014). En 2015, la Chine était le premier
producteur mondial de céréales (21% du total), devant les États-Unis (16%), l’Union
Européenne (11%) et l’Inde (10%). La Chine, le Japon, le Mexique et l’Egypte sont les
principaux pays importateurs de céréales tandis que les Etats Unis, l’Union Européenne,
l’Argentine et l’Australie sont considérés les premiers exportateurs mondiaux (Graphagri,
2016).
Le blé occupe la première place au niveau mondial en termes de production et la
deuxième, après le riz, comme source de nourriture pour les populations humaines. La
production mondiale de blé a atteint un record en 2015 et elle est estimée à 735 000 Tonnes,
dont l’UE produit 21% et la France 6%. L’Amérique du Nord est aussi un producteur et un
exportateur majeur, les États-Unis et le Canada récoltant à eux deux 11% de la production
mondiale du blé (Graphagri, 2016).
2. Importance économique et répartition géographique des céréales en Tunisie
Le blé dur (Triticum turgidum L. ssp. durum) constitue avec l’olivier l’une des cultures
les plus anciennes de la Tunisie. Les agriculteurs tunisiens cultivent traditionnellement le blé
dur pour leurs besoins en semoule et ils le consomment sous plusieurs formes dont
essentiellement le couscous, les pâtes alimentaires, le pain et le borghol (Ben Salem et al.,
1995). La mise en place d’un programme d’amélioration variétale des céréales en Tunisie a
commencé il y a plus d’un siècle et les archives du programme des céréales de l’INRAT
attestent du nombre impressionnant de lignées introduites, collectées localement et créées à
travers les programmes de croisements annuels des blés et de l’orge (Gharbi et al., 2013).
Ainsi, le démarrage des croisements a eu lieu dès 1923 (Bœuf, 1936) et les sélectionneurs
ont commencé par des collectes du germoplasme local, suivies de sélections des meilleures
lignées tout en gardant leurs noms d’origine tels que Biskri, Mahmoudi, Sebeï, Hamira, et
Roussia, etc… (El Felah et Gharbi, 2014).
L’introduction du ‘blé semi-dwarf’ ou ‘blé Mexican’ a commencé vers la fin de l’année
1970 (Ammar et al., 2011). Amal 72 et Magherbi 72 étaient les premières variétés introduites,

Chapitre 1. Synthèse Bibliographique
5
suivies par Ben Bechir et INRAT 69. Les rendements des nouvelles variétés de céréales étaient
trois à quatre fois plus élevés que ceux des anciennes variétés (Gharbi et al., 2011).
L’introduction de la variété Karim en 1980 a créé une véritable percée avec les plus hauts
rendements et depuis elle est restée la plus populaire chez les agriculteurs tunisiens. De plus,
les variétés Razzak (1987), Khiar (1992), Nasr (2004) et Maali (2007) ont-elles aussi réalisé de
véritables succès (Ammar et al., 2011)
Pour le blé tendre (Triticum aestivum L.), exogène à la Tunisie, il n’a pris de l’importance
qu’avec la colonisation française. Cependant, des efforts de recherche sur le blé tendre ont
entraîné l'extension des superficies cultivées par l’introduction du « Florence Aurore » par les
colons depuis 1930 (Lasram, 2013). Ce dernier a eu beaucoup de succès grâce à sa rentabilité
et sa résistance aux maladies. Toutefois, au début des années 1950, le blé tendre était produit
en Tunisie, à 80% à partir de Florence-Aurore et à 20% de blés boulangers (EAP ou Guelma et
Etoile de Choisy) (Saade 1996 ; Ammar et al., 2011).
Avec l’avènement des blés semi-nains, ‘Sonora 63, Inia 66 et Tobari 66’ étaient les
premières variétés Mexicaines qui ont été introduites, et elles ont été abandonnées après des
années d’expérimentations. Au début des années 1980, le deuxième groupe de blé semi-nain a
été introduit (Soltane 72, Dougga 74, Carthage 74, Tanit 80) grâce à leurs bonnes performances.
Cettes dernières n’étaient pas appréciées par les agriculteurs à cause de leur sensibilité envers
la rouille et la septoriose (Ammar et al., 2011). Entre temps, la variété ‘Salammbô 80’ est
devenue la plus populaire en occupant 70% des superficies réservées au blé tendre, cette
dernière a réalisé le plus haut rendement. Un peu plus tard, la variété ‘Byrsa 87’ est apparue
avec de bonnes performances mais n’a pas pris une grande importance à cause de sa sensibilité
envers la rouille jaune et au mildiou. En 1996, avec de bons rendements, la variété ‘Utique 96’
vient de s’installer et devient en compétition avec ‘Salammbô 80’ (Ammar et al., 2011). Durant
la même période, plusieurs variétés à savoir ‘Tebica 96, Vaga 92’, ont été introduites mais elles
n’ont pas trouvé autant d’intérêt. Jusqu'à présent, ‘Haïdra et Tahent’ constituent les variétés de
blé tendre les plus récentes, elles ont été sélectionnées pour leurs caractéristiques
agronomiques : haut rendement, résistance à la sécheresse, tolérance à la rouille et à la
septoriose (Saade et al., 1996).
En Tunisie, le secteur demeure l’un des plus importants de la production agricole. Les
emblavures réservées aux céréales (blé dur, blé tendre, orge et triticale) sont en moyenne de 1,5
million d’hectares répartis en 700 000 hectares de blé dur, 500 000 hectares d’orge, 300 000
hectares de blé tendre et 20 000 hectares de triticale (Anonyme 1, 2015). Ainsi, 58% des
emblavures sont localisées au Nord et au Nord-Ouest du pays, avec 54% des emblavures qui

Chapitre 1. Synthèse Bibliographique
6
sont réservées au blé dur contre 36% pour le blé tendre alors que l’orge ne représente quant à
elle, que 10% des superficies emblavées (Anonyme 2, 2016).
Des rendements de 60 q/ha sont obtenus par les agriculteurs performants dans les zones
favorables du Nord de la Tunisie telles que BéjàBéjà, Jendouba et leLe Kef (El Faleh et al.,
2014). Les variétés Karim, Utique et Rihane dominent respectivement les emblavures de blé
dur, de blé tendre et d’orge, (El Faleh et al., 2014). Néanmoins, les productions nationales n’ont
pas permis la satisfaction des besoins domestiques et la Tunisie n’arrive pas à atteindre son
autosuffisance en céréales et a annuellement recours à l’importation de cette denrée.
Toutefois, le blé dur, qui représente 70% de la production des céréales, satisfait en
moyenne, 72% de la demande nationale alors que le blé tendre ne couvre que 20% des besoins
nationaux et les importations de blé tendre et de blé dur tournent autour de 79 et 27%
respectivement (Bachta, 2011). Les emblavures et les rendements sont largement dépendants
des conditions climatiques, notamment la pluviométrie. La culture du blé est principalement
conduite en pluvial au Nord du pays (Ammar et al., 2011). Par conséquent, on peut distinguer
4 étages bioclimatiques là où le blé est cultivé (figure 1) :
- Etage bioclimatique Humide : avec une pluviométrie dépassant 500 mm, c’est l’étage
bioclimatique le plus favorable pour la culture de blé, répartis sur quelques régions du
gouvernorat de Bizerte et de Jendouba.
- Etage bioclimatique Sub-humide : étage avec une pluviométrie allant de 400 à 500 mm
sur toute l’année, répartis entre les gouvernorats de Bizerte, La Manouba, Nabeul (El Haouaria)
et Béjà.
- Etage bioclimatique Semi-aride : dans cette zone la pluviométrie annuelle varie entre
250 et 400 mm. Cet étage s’étend sur quelques régions du gouvernorat de Jendouba, Le Kef,
La Manouba, Ariana, Ben Arous, Nabeul (Menzel Temim), Zaghouan et Siliana.
- Etage bioclimatique Aride : Centre et sud, zones avec des précipitations très limitées (<
200mm) influençant négativement la production céréalière dans les régions appartenant à cet
étage.

Chapitre 1. Synthèse Bibliographique
7
Figure 1. Etages bioclimatiques adaptés à la culture de blé en Tunisie (Anonyme 3, 2004)
La production céréalière en Tunisie est fortement liée aux stress abiotiques (salinité,
sécheresse) et aux stress biotiques (adventices, ravageurs et surtout les maladies). En effet,
Slama et al. (2005) ont signalé que la culture de blé est soumise souvent à des sécheresses très
fréquentes, entraînant des pertes considérables de rendement qui varient de 10 à 80% selon les
années. Des pertes de rendements importantes ont été également attribuées à l’effet néfaste
causé par les maladies foliaires notamment la septoriose qui entrave les cultures céréalières en
Tunisie (Ben Mohamed et al., 2000 ; Gharbi et al., 2000)
3. La septoriose du blé : Importance dans le Monde, au nord de l’Afrique et en
Tunisie
Ce pathogène est présent partout dans le monde, là où le blé est cultivé (Eyal, 1987). Il
est considéré comme le pathogène le plus destructif dans les pays du bassin méditerranéen,
l’Afrique et l’Amérique du sud. L’importance économique de la septoriose foliaire découle des
pertes de rendement importantes qu’elle occasionne sur le blé, surtout quand les trois dernières

Chapitre 1. Synthèse Bibliographique
8
feuilles sont sévèrement touchées. Des attaques sévères ont été enregistrées en Argentine
(Pampas) (Cordo et al., 2017), l’état de l’Oregon et l’ensemble des USA (Estep et al., 2015),
Canterbury (Drummond et al., 2016) et la Nouvelle Zélande (Stewart et al., 2014). En Australie,
des épidémies importantes ont été enregistrées chez des variétés précoces et les dégâts ont
atteint 50% dans les régions à pluviométrie élevée (Bathgate et Loughman, 1994). En Urugay
et au Brésil, les pertes de rendement étaient de l’ordre de 60 et 50% respectivement (Van
Beuningen et Kohli, 1990). Dans les pays européens, les pertes de rendements peuvent atteindre
30 à 40% (Morais et al., 2015) et plus qu’un milliard d’euro par an sont consacrés aux
fongicides pour lutter contre cette maladie (Kettles et kanyuka, 2016).
En Afrique du Nord, la première épidémie de septoriose s’est produite avec l’introduction
des variétés naines à maturité précoce et à haut rendement durant la campagnecampagne
agricole 1968/1969, qui a particulièremet connu des températures douces et une humidité très
élevée, des conditions très favorables à l’installation de la septoriose. Ainsi, tous les pays du
Maghreb ont connu durant cette période une épidémie spectaculaire et toutes les variétés
commerciales ont été détruites par ce pathogène (Mamluk et al., 1995 ; Mazouz et al., 1995).
En Algérie, Lounis-lalou (2005) a déclaré que toutes les variétés de blé tendre et de blé
dur ont montré des niveaux de sensibilité vis-à-vis de la septoriose. Dans ce sujet, Ayad et al.
(2014) ajoutent qu’en Algérie ce pathogène s’est montré très préjudiciable au cours des
dernières décennies, pouvant réduire les rendements de plus de 60%.
D’autre part, au Maroc Zahri (2008) a rapporté que ce pathogène s’attaque beaucoup plus
au blé tendre qu’au blé dur et que les pertes de rendement sont estimées entre 28 et 30% pour
la variété de blé tendre ‘Nesma’ et peuvent atteindre 35% (Mazouz et al., 1995). Dans le même
sujet, Jilbene (1996) a signalé des pertes dues à Z. tritici de l’ordre de 80% pour certaines
variétés de blé tendre sensibles, notamment pendant les années pluvieuses.
En Tunisie, en conditions favorables et pendant les années pluvieuses, ce pathogène se
place en tête du complexe parasitaire inféodées à la culture de blé dur (Harrabi et Cherif, 1990).
Les pertes de rendements chez le blé dur occasionnées par ce pathogène varient entre 10 et
15%, pouvant atteindre 60% quand les conditions environnementales sont favorables au
développement de la maladie. L’incidence de la maladie croît avec l’augmentation des
superficies irriguées, l’utilisation intensive de la fertilisation azotée et surtout avec les
conditions climatiques favorables caractérisant les régions céréalières du Nord et du Nord-
Ouest qui sont considérées comme étant un Hot-spot pour la septoriose (Rezgui et al., 2008 ;
Fakhfakh et al., 2011 ; Berrais et al., 2013).

Chapitre 1. Synthèse Bibliographique
9
De point de vue sensibilité variétale, Gharbi et al. (2011) ont rapporté que les variétés de
blé dur inscrites avant 2003 à savoir Karim (1980) Razzak (1987) et Khiar (19992), ont monté
une sensibilité très élevée à la septoriose. Par contre, les variétés Nasr (2004), Maali (2007) et
Salim (2010) ont montré des niveaux de résistance partielle à cette maladie. De plus, Gharbi et
Deghaies (1997) et Gharbi et al. (2000) suggèrent que l’incidence de cette maladie était
beaucoup plus importante sur le blé dur que sur le blé tendre.
Contrairement à la situation en Tunisie, Zahri et al. (2008), ont rapporté qu’au Maroc, les
attaques étaient plus fréquentes et plus sévères chez le blé tendre que chez le blé dur. Dans ce
contexte, Eyal et al. (1985), Kema et al. (1995) et Sayoud (1995) ont suggéré une certaine
spécialisation du pathogène sur le blé dur. En Tunisie, et jusqu’à présent pratiquement peu ou
pas de travaux ont été menés sur la septoriose chez le blé tendre en Tunisie et pas de données
exactes de rendements sont ainsi disponibles pour cette culture.
4. Généralités sur la septoriose
La spetoriose causée par ‘Zymosepotia tritici’ est présente chez le blé dur pendant tout
le cycle de la plante, dès l’apparition des premières feuilles jusqu’à la formation de la feuille
drapeau. Ce champignon est apparu comme étant un agent pathogène du blé coïncidant avec la
domestication de cette plante cultivée et sa spéciation a eu lieu avec une adaptation forte à
l'agro-écosystème, car le blé a été soumis à une forte pression de sélection pour augmenter le
rendement (Stukenbrock et al., 2010). Cette pression de sélection a été accompagnée par des
changements dans la diversité génétique des espèces végétales. La co-évolution à long terme
avec le blé a donné lieu à cet agent pathogène très spécialisé du blé et qui est difficile à contrôler
(McDonald, 2014).
4.1.Classification et plantes hôtes
La septoriose du blé, ‘Septoria tritici leaf blotch (STB)’ causée par Zymoseptoria tritici
(Roberge ex Desm.) Quaedvl. & Crous anamorph, et Mycosphaerella graminicola (Fuckel)
J.SchorÖt 1894 teleomorphe, est un champignon ascomycete de la classe des Dothideomycètes
qui s’attaque principalement au blé dur (Triticum turgidum L. subsp. durum (Desf.)), blé tendre
(Triticum aestivum L. subsp. aestivum) et triticale (×Triticosecale spp.).
Observée non seulement sur blé mais aussi sur plusieurs poacées (avoine, triticale, seigle),
cette maladie attaque principalement la partie foliaire (Nasraoui, 2008). De nombreux
adventices courants des cultures de blé, de seigle ou de triticale, tels que Brachypodium spp.,
Bromus spp., Dactylis spp., Festuca spp., peuvent jouer le rôle de réservoirs pour le champignon

Chapitre 1. Synthèse Bibliographique
10
pathogène en maintenant un inoculum à proximité immédiate des céréales cultivées (Gigot,
2013).
4.2.Symptômatologie
Les symptômes engendrés par ce pathogène se manifestent sur blé par des taches brunes,
irrégulières plus ou moins allongées souvent entourées par une bordure chlorotique mince.
Cette dernière se dessèche et devient de plus en plus clair blanchâtre, portant des fructifications
asexuées (pycnides) sous forme de petits points brun foncé à noir qui se forment alors au niveau
des nécroses foliaires, comme le montre la figure 2 (Nasraoui, 2008). Chaque pycnide est
capable de produire des milliers de pycnidiospores qui possèdent une forme allongée
légèrement arquée, d'une longueur comprise entre 20 et 98 μm (Suffert et al., 2013).
Figure 2. Symptômes typiques de la septoriose du blé. (A) Les pycnides de la phase asexuée de Z. tritici (B)
(suffert et al., 2016), (B) Taches nécrotiques avec de petits points brun foncé à noir (Gigot, 2013)
4.3.Cycle biologique de Zymoseptoria tritici
Zymoseptoria tritici est caractérisé par un cycle de vie hémibiotrophe qui commence par
une phase biotrophe par la colonisation du tissu vivant avant de devenir nécrotrophe et tuer les
cellules vivantes. Le stade biotrophe commence dès la pénétration du champignon à travers les
stomates des tissus foliaires vivants (feuilles vertes). L’infection se déclenche dans la cavité
stomatale et le cycle d’infection se déroule en trois phases : 1- l’entrée du champignon, 2- la
A B

Chapitre 1. Synthèse Bibliographique
11
colonisation des tissus de la plante et 3- la formation des pycnides ou ‘fruiting bodies’
(Steinberg, 2015) (figure 3).
Le premier jour de l’infection est connu par l’absence totale de symptômes (O’Driscoll
et al., 2014 ; Sánchez-Vallet et al., 2015). Dix jours après, le champignon entre en phase de
nécrotrophie et commence à causer des chloroses, des nécroses et des pycnides. En effet, cette
phase est étroitement couplée avec la croissance rapide et le début de la reproduction asexuée.
Cependant, le déclencheur moléculaire et environnemental de ce changement entre les deux
phases reste inconnu. Cepandant, Sanchez Vallet et al. (2015) ont rapporté que le stress
environnemental tel que la lumière, la disponibilité de l'eau et les fluctuations de la température
peuvent également jouer un rôle. A ce sujet, Brunner et al. (2013) et Gohari et al. (2015) ont
suggéré aussi que Z. tritici utilise probablement plusieurs mécanismes pour induire une nécrose,
y compris la production de protéines effectrices et que plusieurs gènes codant pour des protéines
riches en cystéine sont fortement exprimés au cours de la phase nécrotrophe. Cela peut indiquer
le rôle majeur des autres acteurs moléculaires tels que les métabolites secondaires, et les
enzymes dégradant les parois dans l’induction de la nécrose. Toutefois, trente jours après
l’infection, le pathogène entre en stade saprophyte (Sanchez Vallet et al., 2015).
Pendant la saison culturale, la maladie se propage d’une plante à une autre (progression
horizontale) et entre les feuilles (progression verticale) sur de courtes distances par dispersion
pluviale des pycnidiospores (figure 4). Il est désormais acquis que ces dernières, transportées
par la pluie sur de courtes distances provoquent l’infection secondaire alors que l’infection
primaire est assurée par les ascospores, potentiellement transportées sur de longues distances
par le vent. Les ascospores se forment sur les débris d’une culture de blé et contaminent le blé
de la saison suivante, en cas de monoculture (Suffert et Sache, 2011).

Chapitre 1. Synthèse Bibliographique
12
Figure 3. Cycle biologique de Zymoseptoria tritici (adapté par O’Driscoll et al., 2014)
Figure 4. Schéma représentatif de la dynamique des ascospores (d'inoculum primaire) et des pycnidiospores
(inoculum secondaire) ainis que leurs rôles dans l’induction d’une épidémie (Suffert et al., 2016).

Chapitre 1. Synthèse Bibliographique
13
4.4. Hétérothallisme et reproduction sexuée
La phase sexuée de Z. tritici a été observée pour la première fois par Sanderson (1972)
en Nouvelle-Zélande. Ce pathogène a été décrit par la suite par plusieurs chercheurs dans
plusieurs pays à travers le monde, au Chili, aux Etats-Unis, aux Pays-Bas, en France, en Algérie
et en Tunisie (Madariaga, 1986 ; Garcia et Marshall, 1992 ; Kema et al., 1996 ; Halama 1996 ;
Ayad et al., 2014 ; Ben Hassine et Hamada, 2014). Pour les espèces hétérothalliques, cette
phase sexuée commence par une étape de reconnaissance entre les types sexuels et entraine la
fusion temporaire de deux souches fongiques portant les mating types compatibles, suivie par
une méiose et un échange d’information génétique entre les individus (Zhan et al., 2004).
Dans le cas de Z. tritici, la reproduction sexuée est possible uniquement lorsque les deux
idiomorphes sont disponibles au même temps, au niveau de la même feuille et à la même
localité géographique (Waalwijk et al., 2002). L’identification, le clonage et le séquençage des
idiomorphes de Z. tritici ont été réalisés à partir de deux isolats de référence IPO323 et
IPO94269 (Waalwijk et al., 2002). Ces idiomorphes, lors du stade sexué de Z. tritici, donnent
naissance aux pseudothèces qui sont produits sous certaines conditions durant l’année (Hunter
et al., 1999). En effet, la distribution géographique des types sexuels parait fortement liée à
l’évolution et la biologie des populations des champignons hétérothalliques. Toutefois, la
reproduction sexuée possède un grand effet sur la diversité génétique des populations par
l’introduction de nouveaux allèles venant d’autres populations (Zhan et al., 1998 ; Kebbage et
al., 2008). Dans des études précédentes, il a été rapporté que la forme sexuée Mycrosphaerella
graminicola joue un rôle très important dans la diversité génétique des populations tout en
influençant le développement épidémiologique de la maladie à travers les saisons de croissance
du blé (Zhan et al., 1998 ; Zhan et al., 2004).
4.5.Intéraction entre Zymoseptoria tritici-blé et spécificité
L'histoire de Z. tritici peut être reliée au Croissant Fertile ~ il y a 11.000 ans (McDonald
et Mundt, 2016). Ainsi un hot-spot et une diversité génétique ont été découverts dans le Moyen
Orient à travers une population de 1673 isolats collectés à partir de plusieurs pays à travers le
monde (Zhan et al., 2003). L'existence d'une spécificité de l’hôte chez Z. tritici a été décrite
depuis plus de 25 ans (Eyal et al., 1973 ; Ware, 2006). La spécificité de l'hôte décrit le degré
d'adaptation d'un parasite à une espèce hôte spécifique. Pour les champignons phytopathogènes,
cette spécificité diffère beaucoup. Par exemple, les champignons nécrotrophes comme Botrytis

Chapitre 1. Synthèse Bibliographique
14
cinerea, l'agent causal de la pourriture grise ont une large gamme d'hôtes. Contrairement, Z.
tritici a une gamme d'hôtes très étroite qui est limitée au blé tendre (Triticum aestivum) et au
blé dur (T. durum), mais il infecte parfois d'autres espèces de graminées, comme l'orge et le
triticale ainsi que certains adventices (Stukenbrock et al., 2011 ; Ponomarenko et al., 2011). En
effet, il a été rapporté par Eyal et al. (1999), que ce champignon peut hiverner sur plusieurs
hôtes alternatifs comme « Agropyron spp., Agrostis spp., Brachypodium spp., Bromus spp.,
Dactylis spp., Festuca spp., Hordeum spp. ».
Seifbarghi et al. (2009) ont démontré que des souches de Z. tritici qui ont été isolées à
partir de T. aestivum étaient capables de causer des symptômes chez les espèces « T. aestivum,
T. durum, T. dicoccum et T. compactum ». Par contre, Kema et al. (2002) ont montré qu’en
général les isolats de blé tendre sont avirulents sur les cultivars de blé dur et les isolats de blé
dur sont avirulents sur des cultivars de blé tendre et il a attribué ceci à l’existence d’une relation
gène pour gène entre le pathogène et la plante. Dans ce contexte, Eyal et al. (1999) et Kema et
al. (1996) ont mentionné trois classes d’isolats : des souches isolées de blé tendre qui n’infectent
que T. aestivum, des souches isolées de blé dur qui infectent exclusivement T. durum et des
souches de blé dur et de blé tendre qui sont capables d’infecter les deux espèces.
Des études récentes ont prouvé une interaction complexe entre le blé et Z. tritici pendant
le processus d'infection (Kellner et al., 2014 ; Rudd et al., 2015). Ces mêmes chercheurs ont
montré que le champignon bénéficie probablement des carbohydrates produits au début de la
phase nécrotrophe. Cette étape est caractérisée également par une forte surexpression des gènes
de défense des plantes, qui coïncide avec l'activation des gènes codant pour les métabolismes
secondaires et les petites protéines sécrétées. Ainsi, il a été démontré que ces petites protéines
effectrices sécrétées jouent un rôle majeur dans la surexpression des réactions de défense dans
l’interaction plante-pathogène (Mirzadi et al., 2015 ; Poppe et al., 2015 ; Rudd et al., 2015).
En 2007, des travaux ont été menés par Goodwin sur l’interaction plante-pathogène et ont
abouti à la découverte des sources de résistance des cultures à Zymoseptoria tritici. Le blé
possède essentiellement deux types de résistance à la septoriose. Cette résistance peut être de
nature qualitative ou spécifique, et quantitative. La résistance qualitative, spécifique ou
verticale est pratiquement totale, monogénique et gouvernée par une relation gène-pour-gène
(Brown et al., 2015 ; Chartrain et al., 2005). Cette dernière est indépendante du stade de
croissance de la plante. Par contre la résistance quantitative ou horizontale peut être dépendante
du stade phénologique, elle n’est pas totale et elle est polygénique (contrôlée par plusieurs
gènes) et dans plusieurs cas elle est efficace contre Z. tritici (Brown et al., 2015).

Chapitre 1. Synthèse Bibliographique
15
Goodwin a montré l’existance de 12 gènes de résistance, dits Stb, en 2007. En tenant
compte de la mise à jour de l’inventaire par Cuthbert (2011) auquel se rajoutent les travaux de
Ghaffary et al. (2011 et 2012), 21 gènes Stb et 167 QTLs ont été identifiés (Hartmann, 2017).
Certains gènes Stb ont été tout à fait durables, tandis que d'autres ont échoué en raison du
changement génétique rapide de la population d'agents pathogènes. Par exemple, Stb1 est resté
efficace dans l'Indiana depuis plus de 25 ans, tandis que Stb 4 était efficace en Californie
pendant 14 ans puis il a échoué, et il a duré seulement un ou deux ans dans l'Oregon. Souvent,
les cultivars de blé rapportés comme résistants dans une région se sont revelés sensibles dans
une autre (Ponomarenko et al., 2011 ; Brown et al., 2015). Cela parait être dû en premier lieu à
la grande diversité génétique du pathogène, qui peut être affectée par les cultivars, la pertinence
de l'environnement et l'importance relative du stade sexuel du pathogène. En Tunisie, Ferjaoui
et ses collaborateurs (2015) ont identifiées la première source de résistance à la septoriose
identifié chez le blé dur a travers la résistance de l’accession ‘Agili39’ à l’isolat TunBz1 aux
stades plantule et adulte.
4.6. Structure des populations de Zymoseptoria tritici
L’identification de la structure génétique de la population de l'agent pathogène est utile
pour le développement des stratégies de gestion de la maladie. En outre, elle reflète l’histoire
évolutive du pathogène ainsi que son potentiel d’adaptation (McDonald, 1997). La structure
génétique se réfère à la quantité et la distribution de la variation génétique au sein et entre les
populations de pathogènes (McDonald et Mundt, 2016). La génétique des populations permet
de comprendre les processus évolutifs impliqués dans la création et le maintien de la variation
génétique au sein et entre des populations, en analysant les fréquences alléliques. En outre deux
types de diversité génétique contribuent à la structure génétique : la diversité génique et la
diversité génotypique.
La diversité génique est estimée grace au nombre et des fréquences d’allèles au niveau des loci
individuels dans une population. Elle augmente au fur et à mesure que le nombre d’allèles
augmente et les fréquences relatives de ces allèles deviennent égales. Cette diversité génique
peut être affectée par la taille et l’âge de la population ainsi par le flux de gène. La diversité
génotypique se réfère au nombre et à la fréquence des génotypes multi-locus ou d’individus
génétiquement distincts (génotypes) au sein d’une population donnée (McDonald et Linde,
2002).

Chapitre 1. Synthèse Bibliographique
16
4.7.Diversité génétique des populations
Plusieurs études antérieures ont été conduites sur l’analyse de la diversité génétique des
populations de Z. tritici, à tous les niveaux : pays, région, champ, plante et feuille (McDonald
et Martinez, 1990 ; Linde et al., 2002 ; Abrinbana et al., 2010 ; Gurung et al., 2011 ; El
Chartouni et al., 2012 ; Boukef et al., 2013 ; Nouari et al., 2016 ; Siah et al., 2018).
La caractérisation des populations à l’aide des premières générations de marqueurs
moléculaires a permis de mieux comprendre la structure génétique des populations de ce
pathogène. Ainsi, une faible différenciation génétique a été observée chez les populations de Z.
tritici sur les plans régional et mondial. En plus, une diversité génétique très élevée et un flux
de gène très important ont été ainsi signalés entre plusieurs populations à travers le monde
(Schnieder et al., 2001 ; Linde et al., 2002 ; Zhan et al., 2003 ; Drabesova et al., 2013). Le cas
contraire a été signalé en Iran où Abrinbana et al. (2011) ont démontré une faible diversité
génétique, et un flux de gène très bas dans cinq populations de Z. tritici. Des études précédentes
ont indiqué que la reproduction asexuée n’avait aucun effet sur la variabilité génétique des
populations du Z. tritici alors que la reproduction sexuée y joue un rôle important en favorisant
les flux de gènes et l'apparition rapide de résistance aux fongicides (Zhan et al., 1998;
Abrinbana et al., 2010 ; Boukef et al., 2012). De plus, les populations fongiques ayant une
variation génétique élevée sont plus capables de développer une résistance aux fongicides que
les populations ayant une faible variation génétique (Drabešová et al., 2013).
En Tunisie, une grande diversité génétique des populations de Z. tritici isolées à partir du
blé dur a été révelée à travers plusieurs études (Berraies et al., 2013; Boukef, 2012, Nouari et
al., 2016). Toutefois, étant donné que ce pathogène s’attaque plus au blé dur en Tunisie, la
diversité génétique des populations de Z. tritici isolées à partir du blé tendre reste inonnue.
4.8. Les forces évolutives affectant la diversité génétique des populations
Il a été rapporté que la diversité génétique des populations peut être affectée
principalement par cinq forces évolutives à savoir : la mutation, la taille de la population, le
flux de gène, la reproduction sexuée et la sélection (McDonald, 2014).
La mutation constitue la source la plus importante dans les variations génétiques en
conduisant directement à des changements dans la séquence d'ADN et créant ainsi de nouveaux
allèles dans les populations. Cette dernière pourrait également créer des souches avec une
pathogénicité accrue (McDonald et al., 2016). Une importance particulière a été attribuée aux
flux de gène dans la dispersion du pathogène. En effet, c’est au cours de ce processus que

Chapitre 1. Synthèse Bibliographique
17
s’effectue l’échange de gènes entre des populations séparées géographiquement. Toutefois, le
flux de gènes élevé limite la différenciation entre les populations, par l’homogénéisation des
fréquences des allèles, et augmente la diversité génétique dans la population (McDonald et al.,
1997). Cette dernière peut être aussi affectée par la reproduction sexuée tout en favorisant
l’apparition rapide de nouvelles recombinaisons d’allèles virulents qui contribuent eux-mêmes
par la suite à l’apparition de nouveaux gènes de résistances aux fongicides (Zhan et al., 1998).
La sélection est en outre une force évolutive non négligeable car c’est elle qui contribue aux
changements des fréquences alléliques qui se traduit par la perte des allèles au fil du temps
(McDonald et Linde, 2002).
5. Influence des facteurs climatiques sur le développement de la maladie
Les maladies des plantes cultivées peuvent occasionner des pertes considérables lorsque
les conditions climatiques sont favorables à leur développement. L'identification des facteurs
climatiques qui favorisent ou défavorisent le développement des agents pathogènes est
nécessaire à la compréhension de l'épidémiologie et l’évolution des maladies des plantes. De
plus, il est reconnu que c’est le climat qui gouverne en grande partie les changements au niveau
du pathogène, de l’épidémie et aussi de la plante hôte (Rapilly, 1991). Le développement d'une
maladie foliaire résulte de l'interaction dynamique entre la plante hôte, l'agent pathogène et
l'environnement (Suffert et al. 2015).
Dans le cas de la septoriose, la maladie se développe à une température modérée et à une
humidité relative élevée (Gouache et al., 2013). Dans ce contexte, Cordo et al. (2017)
rapportent que, la radiation, la température, l’intensité des pluies et l’humidité relative affectent
significativement la dispersion des ascospores et des pycnidiospores du pathogène. Cependant,
le processus de l’infection peut avoir lieu idéalement au cours des journées nuageuses et
pluvieuses et le progrès et l’intensité de l’épidémie sont déterminés par le nombre de cycles
asexués accomplis par le pathogène (généralement 4 à 6) qui sont fortement gouvernés par les
scénarios de la pluviométrie et de la température (Suffert et al., 2015).
Toutefois, l’humidité est déterminante pour toutes les étapes de l’infection : la
germination, la pénétration et le développement du mycelium dans le tissu de la plante hôte. En
effet, pour germer, les spores ont besoin d’eau sur les feuilles, ainsi qu’une période humide
assez longue pour pouvoir pénétrer dans l’hôte (Rapilly, 1991).
D’autre part, la température peut avoir plusieurs effets sur le changement du
comportement parasitaire du champignon. En effet, les températures basses affectent la

Chapitre 1. Synthèse Bibliographique
18
germination des pycnidiospores et la croissance mycelienne (Eyal et al., 1987). La germination
des spores exige une température optimale entre 5 et 35°C et une humidité relative saturée
constante de 3 à 100% sur le feuillage (Kema et al., 1996a). L’infection peut être retardée dans
le champ si la température est au dessous de 7°C durant 2 nuits consécutives. Cordo et al. (2017)
et Eyal et al. (1987) rapportent que la libération des pycnidiospores est fortement corrélée avec
la radiation et qu’un optimum de 8.000 à 12.000 lux est nécessaire pour la croissance du
mycelium.
Selon Suffert et al. (2015), le vent intervient directement sur la dissémination, la
dispersion et le transport à longue distance des ascospores pour initier une nouvelle épidémie.
Il agit aussi sur les flux de chaleur et de vapeur d’eau qui s’établissent au sein d’un couvert
végétal. En effet, par ses vitesses et ses changements brutaux de direction que le vent joue un
rôle dans la libération des spores (Rapilly, 1991).
Certains groupes de parasites nécrotrophes sont dépendants de l’eau libre pour la
libération de l’inoculum primaire et même l’inoculum secondaire (grâce à la libération de la
gelée). Toutefois, l’humidité saturante et l’eau libre sont indispensables pour la contamination,
par contre une humidité relative élevée est suffisante pour l’incubation, la latence et l’extension
des surfaces sporulantes. En effet, selon Rapilly (1991), à peu près 25% de l’intensité de la
septoriose causée par Septoria nodurum sur les deux derniers limbes du blé est expliquée par la
persistance de la rosée. Toutefois, la vapeur d’eau peut accroitre l’adhésion des spores sur les
feuilles tout en augmentant la pression de l’inoculum. En outre, Cordo et al. (2017), ont signalé
que plus les pluies sont intenses plus la dispersion des spores par splash est rapide vers les
surfaces foliaires saines.
D’autre part, la pluie et la rosée interviennent aussi par la dilution de la concentration de
l’inoculum primaire, parfois par la dissolution des substances sporales du pathogène et même
l’élimination de ces derniers. Selon Morais (2015), la vitesse de développement d'une épidémie
est déterminée par le nombre de cycles de multiplication asexuée qui dépend des conditions de
température et du nombre d'épisodes pluvieux.
6. La résistance variétale comme alternative de lutte contre la septoriose
Face aux limites de la lutte chimique via des molécules unisites, la méthode de contrôle
la plus importante, économique et respectueuse de l'environnement est d’utiliser des cultivars
résistants. L’utilisation de variétés résistantes constitue une composante essentielle pour la
majorité des programmes de sélection et un moyen de contrôle efficace (Fones et Gurr, 2015).

Chapitre 1. Synthèse Bibliographique
19
De leur part, Ben Mbarek et al. (2019) ont souligné l’importance du recours au mélange variétal
à raison de 25% (pour les variétés résistantes) qui a améliore significativement la réduction de
la septoriose dans des essais de plein champ à Béjà.
C’est dans ce cadre que s’intègre ce travail de thèse qui s’est basé sur une étude de la
distribution géographique de Z. tritici en Tunisie chez le genre Triticum particulièrement
l’espèce T. aestivum dans les différentes régions céréalières en une première étape. Ensuite, il
s’est orienté vers l’investigation de la sensibilité/résistance d’un ensemble de 89 génotypes de
blé de différentes origines Tunisienne, Marocaine et Algérienne, cultivés en Tunisie.
Finalement, ce travail s’est interessé à l’étude de la diversité et la structure génétique d’une
collection de Z. tritici et de la différentiation génétique entre les populations à plusieurs
niveaux : région, parcelle, espèce et cultivar.

Chapitre2. Occurrence of Septoria tritici blotch (Zymoseptoria
tritici) disease on durum wheat, triticale, and bread wheat in
Northern Tunisia
Rim Bel Hadj Chedli1, Sarrah Ben M’Barek2, Amor Yahyaoui3, Zakaria Kehel4, and Salah
Rezgui1
1 National Agronomic Institute of Tunisia (INAT), 43 Avenue Charles Nicolle, 1002 Tunis, Tunisia. 2 Regional Field Crop Research Center of Beja (CRRGC) BP 350, 9000 Beja, Tunisia. 3 Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), km 45 Carretera México-Veracruz El Batán,
Texcoco, Estado de México, México. 4 International Center for Agricultural Research in the Dry Areas (ICARDA), Rue Hafiane Cherkaoui, Agdal Rabat
Po Box 6299 PC: 10112, Morocco
Bel Hadj Chedli et al. Occurrence of Septoria tritici blotch (Zymoseptoria tritici ) disease on durum
wheat, triticale, and bread wheat in Northern Tunisia. Chil. j. agric. res. [online]. 2018, vol.78, n.4,
pp.559-568. ISSN 0718-5839. http://dx.doi.org/10.4067/S0718-58392018000400559.

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
20
Objectifs et démarche
La sévérité de l’attaque par Z. tritici dans les champs de blé dur (Triticum turgidum L.
subsp. durum (Desf.)) est influencée par la sensibilité élevée des variétés améliorées ‘Karim’,
‘Khiar’ et ‘Razzak’. Les infections de ce champignon phytopathogène se sont intensifiées dans
les zones humides notamment à Bizerte et à Béjà et les pertes de rendements ont dépassé 60%
chez les variétés sensibles comme ‘Karim’ (Ben Mouhamed et al., 2000 ; El Faleh et al., 2014)
Bien que la septoriose fût observée annuellement sur blé dur, le blé tendre ‘Triticum
aestivum’ semble être beaucoup plus résistant à cette maladie en Tunisie. Cette hypothèse a été
supportée par plusieurs chercheurs à savoir Dhgaies et al. (1996) et Gharbi et al. (2000), qui
ont signalé que le blé tendre en Tunisie a montré depuis toujours un haut niveau de résistance
à ce pathogène. Cependant, jusqu’à présent, aucune information n’a été publiée concernant
l’importance de cette maladie et sa répartition géographique en Tunisie chez le blé tendre.
Ainsi, les principaux objectifs de ce chapitre sont :
✓ L’étude de la prévalence et la distribution géographique de Z. tritici chez le genre
« Triticum » dans différentes régions céréalières au Nord de la Tunisie,
✓ La réévaluation de l'importance relative de cette maladie qui s’attaque
principalement au blé dur en Tunisie
✓ L’identification des microclimats favorables à ce champignon pathogène pour
développer des stratégies de lutte spécifiques pour chaque situation.
Pour cela, une enquête a été menée dans les principales régions céréalières du nord de la
Tunisie (Béjà, Bizerte, Manouba, Jendouba, Cap Bon, Zaghouan et Le Kef) où 57 champs ont
été prospectés durant la saison agricole 2015-16, et 69 champs ont été inspectés en 2016-17. La
prévalence de la maladie a été mesurée au sein de chaque région alors que l’incidence et la
sévérité ont été déterminées au niveau de chaque champ en se basant sur l’échelle de notation
« Saari-Prescott ». En plus, un protocole de classification adapté par ‘CIMMYT-septoria
phenotyping platform’ a été aussi appliqué pour identifier les différentes régions à risque.

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
21
Abstract
Wheat (Triticum turgidum L. subsp. durum (Desf.) is the most important cereal crop in
Tunisia, nonetheless production is highly affected by drought and diseases mainly Septoria
tritici blotch (STB) caused by ‘Zymoseptoria tritici’.The main objective of this work was to
study prevalence and geographical distribution of this pathogen on triticale, durum wheat and
particularly on bread wheat in different cereal growing regions of North and Northwestern
Tunisia to confirm its presence/absence on bread wheat. For this study, 126 wheat fields were
surveyed in North and Northwestern Tunisia during 2015-2016 and 2016-2017 cropping
seasons. STB on durum wheat was present in the majority of inspected durum wheat fields,
where high mean incidence (60%) and severity (40%) were recorded at Jendouba, Bizerte, Béjà,
and Kef. The survey data revealed low risk on bread wheat with an incidence of 23% and 29%
at Bizerte and Béjà, respectively. However high incidence of 84% and 52% was recorded at
Cap Bon in 2016 and 2017, respectively and mainly at El Haouaria where STB severity was
relatively high on bread wheat landrace of unknown origin but called by local farmers as ‘Farina
Arbi’. High (100%) and moderate (33%) incidence was recorded on Triticale at Bizerte and
Jendouba respectively during the two-cropping season. The survey data revealed low risk on
bread wheat except at El Haouaria where STB severity was relatively high on a bread wheat
landrace; while it was considered as high risk at all durum wheat fields in Béjà, Bizerte,
Jendouba, Zaghouan and Kef regions, such distinct occurrence could lead to clarify host
specificity in Z. tritici.
Key words: Farina Arbi, survey, Triticum, Tunisia, wheat, Zymoseptoria.
1. Introduction
The cereal sector is of major economic importance in Tunisia. It provides major staple
food commodities for most Tunisian households. Cereals are cultivated on almost one third of
agricultural land (1.5 million hectares) (Tunisian Ministry of Agriculture, 2015), 58% are
located in the Northern and Western regions where durum wheat (Triticum turgidum L. subsp.
durum (Desf.) van Slageren) represents 54%, against 36% for bread wheat (Triticum aestivum
L. subsp. aestivum) and 10% for barley (FAO, 2017). Average production is around 1.05
million tons, which represent approximately 80% of the country needs (Gharbi et al., 2000).
However, cereal production in Tunisia faces many challenges of which drought is the most
limiting abiotic stress in semi arid zones (Slama et al., 2005), while biotic stress, mainly leaf

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
22
rust and Septoria tritici blotch (STB), cause important yield losses particularly on durum wheat
in sub-humid regions (Ben Mohamed et al., 2000; Gharbi et al., 2000) of North and
Northwestern Tunisia.
STB caused by the ascomycete fungus Zymoseptoria tritici (Roberge ex Desm.) Quaedvl.
& Crous became more important in Tunisia since the introduction of early maturing, semi
dwarf, high yielding varieties. It has become an inherent disease of durum wheat, and thus a
significant challenge for breeders to release varieties which combine good resistance and higher
yields (Ammar et al., 2011). In contrast to durum wheat, bread wheat varieties grown in Tunisia
are almost indemn of Septoria. High incidence of STB on durum compared to bread wheat in
Tunisia suggests either an adaptation of Z. tritici isolates to durum rather than bread wheat
(Yahyaoui et al., 2000) or high levels of resistance in bread wheat. The observed levels of
resistance amongst cultivated bread wheat varies from year to year, most likeley based on the
environmental conditions and the dynamics of STB populations.
Although Septoria was observed on durum wheat annually, up to now not much is known
on the occurence of STB on bread wheat. Hence, the main objective of this paper was to study
the prevalence and geographical distribution of Z. tritici on Triticum species and particularly
on T. aestivum in different cereal growing regions of North and Northwestern Tunisia to
eventually confirm its presence/absence on bread wheat.
2. Materials and Methods
2.1.Study area description
Surveys were conducted during two cropping seasons (2015-2016; 2016-2017) at seven
major wheat-growing areas in North and Northwestern Tunisia (Figure 1). Fifty-seven fields
were surveyed in Cap Bon North (El Haouaria), Cap Bon South, Bizerte, Manouba, Béjà, and
Jendouba during 2016 and sixty-nine fields were surveyed in Cap Bon regions, Bizerte,
Manouba, Béjà, Jendouba, Zaghouan, and Kef during 2017 (Figure1). Certain varieties were
more predominant than others rendering therefore inter region comparison rather difficult to
make.
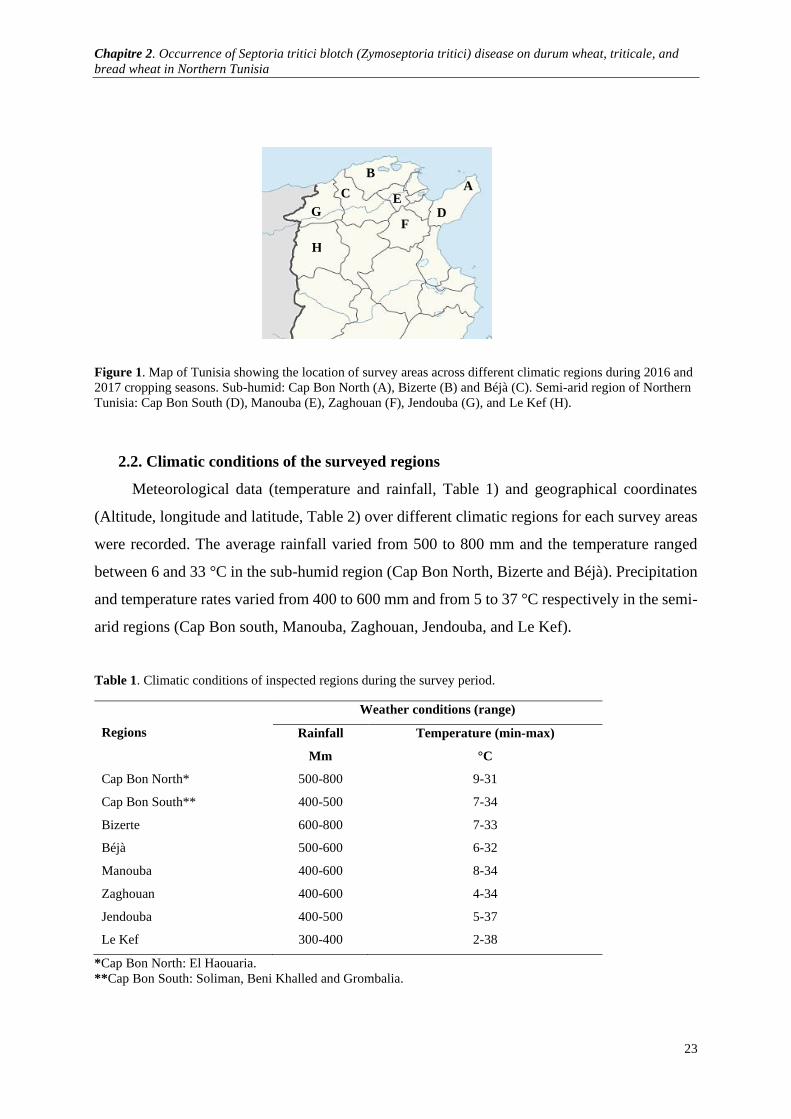
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
23
Figure 1. Map of Tunisia showing the location of survey areas across different climatic regions during 2016 and
2017 cropping seasons. Sub-humid: Cap Bon North (A), Bizerte (B) and Béjà (C). Semi-arid region of Northern
Tunisia: Cap Bon South (D), Manouba (E), Zaghouan (F), Jendouba (G), and Le Kef (H).
2.2. Climatic conditions of the surveyed regions
Meteorological data (temperature and rainfall, Table 1) and geographical coordinates
(Altitude, longitude and latitude, Table 2) over different climatic regions for each survey areas
were recorded. The average rainfall varied from 500 to 800 mm and the temperature ranged
between 6 and 33 °C in the sub-humid region (Cap Bon North, Bizerte and Béjà). Precipitation
and temperature rates varied from 400 to 600 mm and from 5 to 37 °C respectively in the semi-
arid regions (Cap Bon south, Manouba, Zaghouan, Jendouba, and Le Kef).
Table 1. Climatic conditions of inspected regions during the survey period.
Regions
Weather conditions (range)
Rainfall Temperature (min-max)
Mm °C
Cap Bon North* 500-800 9-31
Cap Bon South** 400-500 7-34
Bizerte 600-800 7-33
Béjà 500-600 6-32
Manouba 400-600 8-34
Zaghouan 400-600 4-34
Jendouba 400-500 5-37
Le Kef 300-400 2-38
*Cap Bon North: El Haouaria.
**Cap Bon South: Soliman, Beni Khalled and Grombalia.
A B
D G E
H
C
F
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
24
Table 2. Geographical coordinates of inspected regions during the survey period
Region Altitude
(range)
Longitude (N)
(range)
Latitude (E)
(range)
Bizerte 16-354 09°03’1 09°69’83 32°50’56 37°14’12
Cap bon North* 11-876 10°02’73 10°10’95 36°52’41 36°67’82
Cap bon South** 24- 446 10°45’27 10°49’10 36°47’47 36°92’68
Manouba 58-244 09°59’1 09°91’41 36°30’70 36°85’04
Zaghouan 81-320 09°45’43 10°4’42 36°21’18 36°30’53
Béjà 18-290 09°09’01 10°78’62 36°22’26 36°68’31
Jendouba 21-341 08°32’35 08°42’45 36°28’4 36°32’48
Kef 100-327 08’39°12 08°48’50 36°22’26 36°11’44
*Cap Bon North : El Haouaria.
**Cap Bon South : Soliman, Beni Khalled and Grombalia.
2.3. Cereal crops
The survey covered the major cereal growing areas in North and Northwestern Tunisia
where commercial durum wheat varieties (‘Karim’, ‘Razzak’, ‘Maali’) occupy over 60% of the
area compared to the introduced varieties (‘Saragola’, ‘Carioca’, ‘Sculpture’, ‘Soudaine’) that
cover so far less than 10%. Commercial bread wheat varieties (‘Salambo’, ‘Utique’, ‘Haidra’)
and introduced varieties (‘Zanzibar’) occupy no more than 20% of the area; while triticale
(‘Bienvenue’ and others) covers about 1-3%; the rest of the area is covered by barley and oats.
A unique situation in Cap Bon region where a landrace bread wheat (‘Farina Arbi’) occupies
over 60% of the area, the rest is covered by commercial barley, durum and bread wheat
varieties. ‘Farina arbi’, a tall low yielding bread wheat (landrace) of unknown origin is
cultivated annually for over a century, according to local farmers, in the same region and
exclusively used for pastry known as “Kaak”. The seed is maintained by local farmers and not
commercialized. Bread wheat landrace (‘Farina arbi’) and Septoria tritici blotch (STB)
differentials, (comprised within CIMMYT’s ISEPTON) were phenotyped at experimental
station of Bou Salem (Tunisia) under artificial inoculation with Zymoseptoria tritici populations
sampled from durum wheat. Inoculation was performed at tillering stage using bulk isolates at
a rate of 106 spores/ml, according to Ferjaoui et al. (2015) with slight modifications.

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
25
2.4. Septoria leaf blotch disease assessment
Field surveys were conducted during flowering stage of bread and durum wheat where
each field was visited once. STB prevalence was assessed within and between regions based on
number of fields surveyed and the presence/absence of Septoria at each location. The incidence
was reported on this study based on Saari-Prescott modified “0-9” Cobb-scale (Saari and
Prescott, 1975). In this survey, we designed five field classes (Table 3) to assess STB within
each region where prevalence, severity and incidence were the main criteria. The relative
importance of STB was based on its prevalence at each location where incidence and severity
were assessed and averaged at each surveyed field. In this study, prevalence indicates how wide
spread is STB, whereas incidence conveys information on the risk of the disease within a
severity range. In the survey protocol of the CIMMYT septoria phenotyping platform five
classes (Class I-V) were adopted. Class I: Low prevalence (%), severity (0-9), and low
incidence indicating insignificant risk; Class II: low prevalence and incidence indicating low
risk. Class III: moderate prevalence and incidence indicating moderate risk to be monitored.
Class IV: fields where STB was wide spread and apparent high severity observed at flag leaf,
indicating high risk of the disease. Class V: includes fields heavily infested by STB; situation
where the disease is obvious at each field surveyed and where the severity is at its most, i.e.
severe symptom on flag leaf and spikes, this is a situation where STB is a high risk. Relevant
agronomic data such as, variety name, sowing date, fertilization, crop density and spatial pattern
and previous crops were recorded. Altitude, longitude and latitude were also recorded using
Global Positioning System (GPS).
Table 3. Survey designated Classes for Septoria tritici blotch (STB) prevalence, severity and incidence.
Class Prevalence1 (%) Severity2
(Cobb-Scale: 0-9)
Incidence3
H Severity
I 0-10 0-3 0-2 Insignificant
II 10-20 3-5 2-3 Low
III 20-40 5-6 3-5 Moderate
IV 40-60 6-8 5-7 High
V 60-100 8-9 7-9 Severe
1(Number of infected fields (STB present)/total number of fields surveyed) × 100. 2H: Level of infection plant level. 3Percentage of STB within each class level at surveyed field.

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
26
2.5. Data analysis
Linear mixed model was used to analyze the disease data (incidence and severity)
collected during the survey using ASReml-R software (Gilmour et al., 2002). The years,
regions, species, varieties together with their interactions were assumed to be fixed.
3. Results
3.1. Incidence of wheat Septoria tritici blotch
Survey results showed that STB incidence on bread wheat was very limited across
surveyed areas in Northern Tunisia Triticum species and wheat varieties identified during the
survey during the two cropping seasons are presented in Table 4. STB prevalence during the
two cropping seasons (2015-2016 and 2016-2017) was insignificant to low on commercial
bread wheat varieties in the majority of the surveyed regions (Tables 5 and 6). It was ranked as
class I or II except at El Haouaria (region A, Figure 1) where prevalence and incidence were
relatively high (Tables 5 and 6; Figures 2 and 4) putting it as class IV-V level (Table 3).
Insignificant prevalence levels were recorded at Zaghouan, Bizerte, and Béjà (Tables 5 and 6).
Even though relatively high incidence was recorded at Bizerte, the severity was still low;
hence it is of low to moderate risk (class II-III). The high incidence observed at Béjà was only
at 1 out of 2 fields surveyed, hence it is not quite representative and we considered it low. The
mean disease incidence and severity on bread wheat landrace reached the maximum levels in
2016 with 84% and 52% respectively at El Haouaria (Figures 2 and 3). Same trend was
observed in 2017, where over 77% and 32% disease incidence and severity respectively were
recorded in the same region (Figures 4 and 5).
Low STB incidence on bread wheat were recorded in 2016 on bread wheat fields at
Bizerte 23% and Béjà 29%, and in 2017 at Bizerte 17% and Zaghouan 5% (Figures 2 and 4).
The severity percentages in these regions did not exceed 5% during the two cropping seasons
(Figures 3 and 5). These data showed that STB level was at class I and II ratings except at El
Haouaria, where it was rated class IV and V.
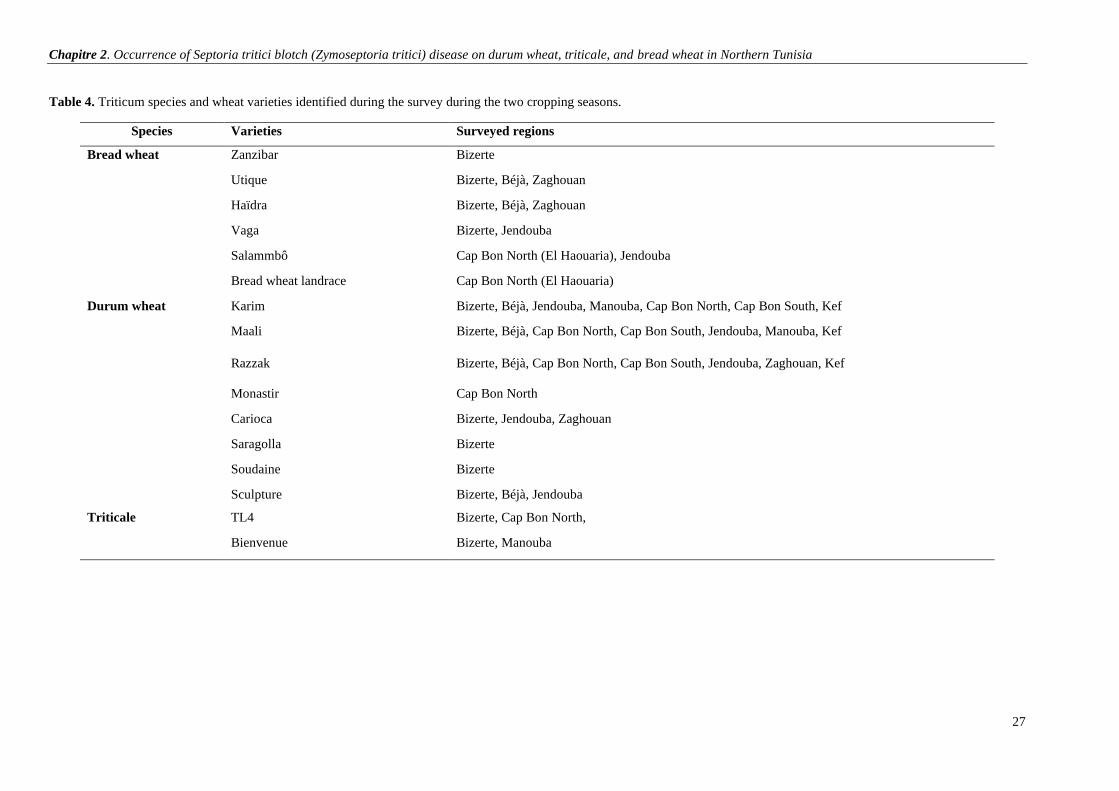
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and bread wheat in Northern Tunisia
27
Table 4. Triticum species and wheat varieties identified during the survey during the two cropping seasons.
Species Varieties Surveyed regions
Bread wheat Zanzibar Bizerte
Utique Bizerte, Béjà, Zaghouan
Haïdra Bizerte, Béjà, Zaghouan
Vaga Bizerte, Jendouba
Salammbô Cap Bon North (El Haouaria), Jendouba
Bread wheat landrace Cap Bon North (El Haouaria)
Durum wheat Karim Bizerte, Béjà, Jendouba, Manouba, Cap Bon North, Cap Bon South, Kef
Maali Bizerte, Béjà, Cap Bon North, Cap Bon South, Jendouba, Manouba, Kef
Razzak Bizerte, Béjà, Cap Bon North, Cap Bon South, Jendouba, Zaghouan, Kef
Monastir Cap Bon North
Carioca Bizerte, Jendouba, Zaghouan
Saragolla Bizerte
Soudaine Bizerte
Sculpture Bizerte, Béjà, Jendouba
Triticale TL4 Bizerte, Cap Bon North,
Bienvenue Bizerte, Manouba
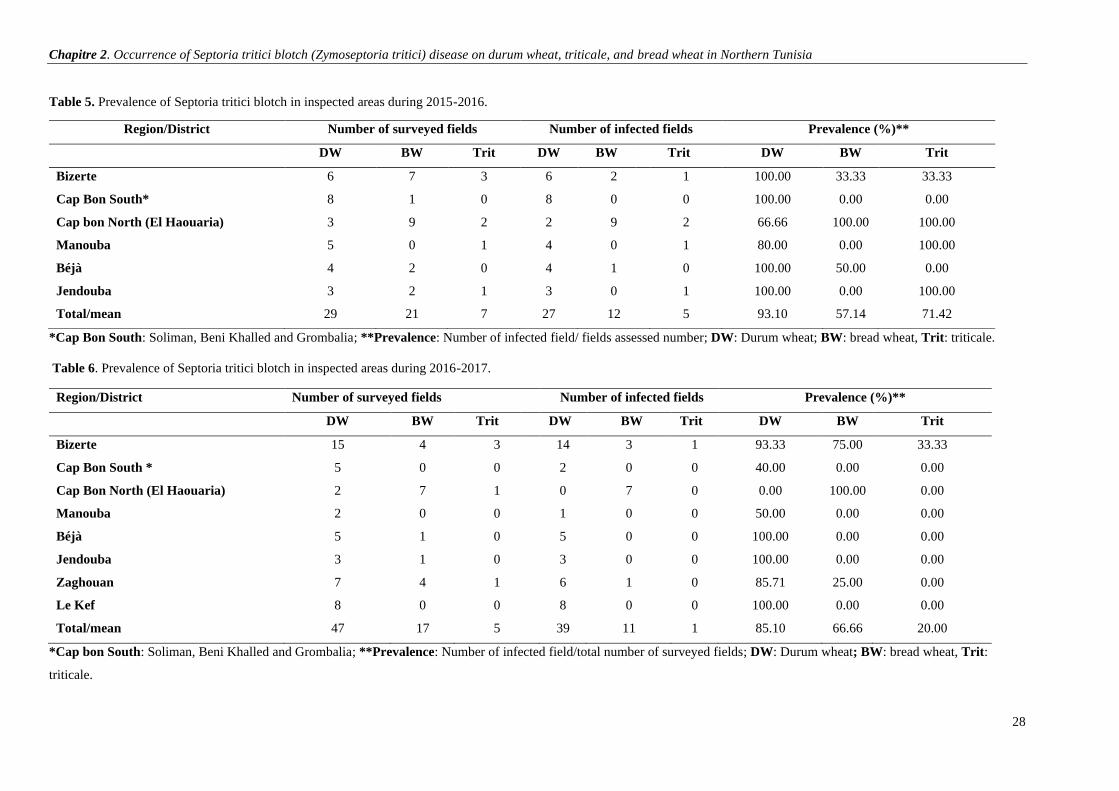
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and bread wheat in Northern Tunisia
28
Table 5. Prevalence of Septoria tritici blotch in inspected areas during 2015-2016.
*Cap Bon South: Soliman, Beni Khalled and Grombalia; **Prevalence: Number of infected field/ fields assessed number; DW: Durum wheat; BW: bread wheat, Trit: triticale.
Table 6. Prevalence of Septoria tritici blotch in inspected areas during 2016-2017.
Region/District Number of surveyed fields Number of infected fields Prevalence (%)**
DW BW Trit DW BW Trit DW BW Trit
Bizerte 15 4 3 14 3 1 93.33 75.00 33.33
Cap Bon South * 5 0 0 2 0 0 40.00 0.00 0.00
Cap Bon North (El Haouaria) 2 7 1 0 7 0 0.00 100.00 0.00
Manouba 2 0 0 1 0 0 50.00 0.00 0.00
Béjà 5 1 0 5 0 0 100.00 0.00 0.00
Jendouba 3 1 0 3 0 0 100.00 0.00 0.00
Zaghouan 7 4 1 6 1 0 85.71 25.00 0.00
Le Kef 8 0 0 8 0 0 100.00 0.00 0.00
Total/mean 47 17 5 39 11 1 85.10 66.66 20.00
*Cap bon South: Soliman, Beni Khalled and Grombalia; **Prevalence: Number of infected field/total number of surveyed fields; DW: Durum wheat; BW: bread wheat, Trit:
triticale.
Region/District Number of surveyed fields Number of infected fields Prevalence (%)**
DW BW Trit DW BW Trit DW BW Trit
Bizerte 6 7 3 6 2 1 100.00 33.33 33.33
Cap Bon South* 8 1 0 8 0 0 100.00 0.00 0.00
Cap bon North (El Haouaria) 3 9 2 2 9 2 66.66 100.00 100.00
Manouba 5 0 1 4 0 1 80.00 0.00 100.00
Béjà 4 2 0 4 1 0 100.00 50.00 0.00
Jendouba 3 2 1 3 0 1 100.00 0.00 100.00
Total/mean 29 21 7 27 12 5 93.10 57.14 71.42
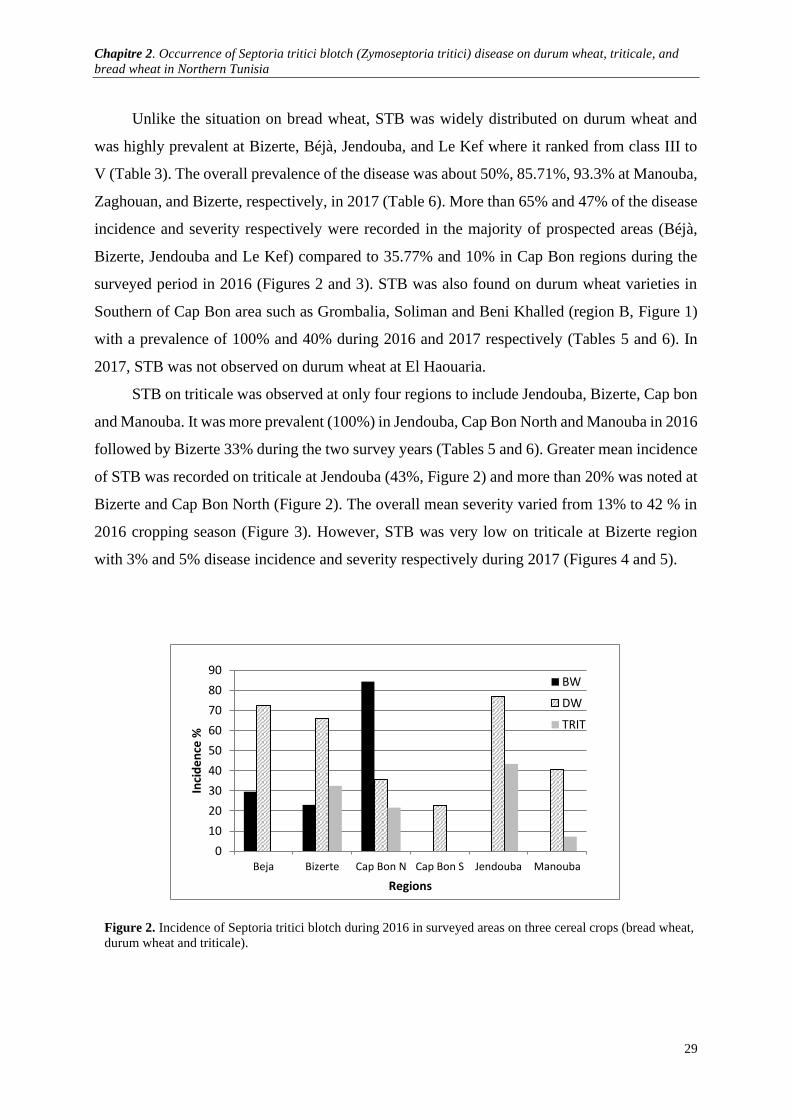
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
29
Unlike the situation on bread wheat, STB was widely distributed on durum wheat and
was highly prevalent at Bizerte, Béjà, Jendouba, and Le Kef where it ranked from class III to
V (Table 3). The overall prevalence of the disease was about 50%, 85.71%, 93.3% at Manouba,
Zaghouan, and Bizerte, respectively, in 2017 (Table 6). More than 65% and 47% of the disease
incidence and severity respectively were recorded in the majority of prospected areas (Béjà,
Bizerte, Jendouba and Le Kef) compared to 35.77% and 10% in Cap Bon regions during the
surveyed period in 2016 (Figures 2 and 3). STB was also found on durum wheat varieties in
Southern of Cap Bon area such as Grombalia, Soliman and Beni Khalled (region B, Figure 1)
with a prevalence of 100% and 40% during 2016 and 2017 respectively (Tables 5 and 6). In
2017, STB was not observed on durum wheat at El Haouaria.
STB on triticale was observed at only four regions to include Jendouba, Bizerte, Cap bon
and Manouba. It was more prevalent (100%) in Jendouba, Cap Bon North and Manouba in 2016
followed by Bizerte 33% during the two survey years (Tables 5 and 6). Greater mean incidence
of STB was recorded on triticale at Jendouba (43%, Figure 2) and more than 20% was noted at
Bizerte and Cap Bon North (Figure 2). The overall mean severity varied from 13% to 42 % in
2016 cropping season (Figure 3). However, STB was very low on triticale at Bizerte region
with 3% and 5% disease incidence and severity respectively during 2017 (Figures 4 and 5).
Figure 2. Incidence of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops (bread wheat,
durum wheat and triticale).
0
10
20
30
40
50
60
70
80
90
Beja Bizerte Cap Bon N Cap Bon S Jendouba Manouba
Inci
de
nce
%
Regions
BW
DW
TRIT
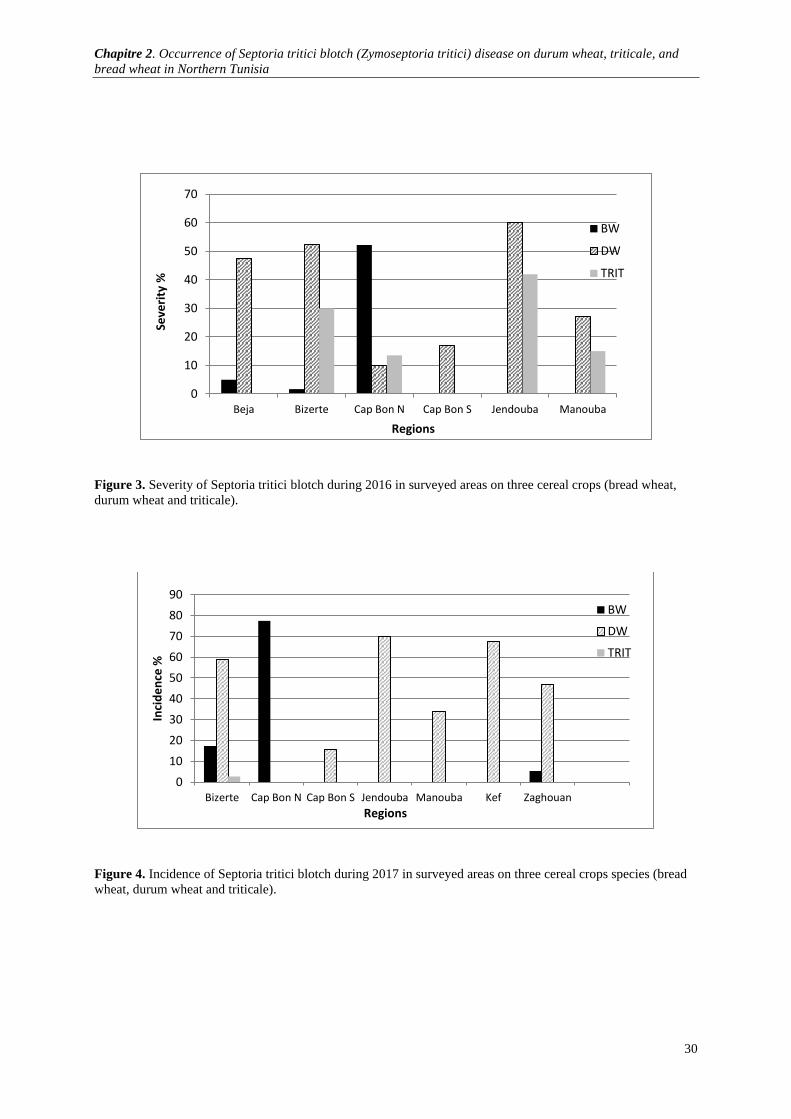
Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
30
Figure 3. Severity of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops (bread wheat,
durum wheat and triticale).
Figure 4. Incidence of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops species (bread
wheat, durum wheat and triticale).
0
10
20
30
40
50
60
70
Beja Bizerte Cap Bon N Cap Bon S Jendouba Manouba
Seve
rity
%
Regions
BW
DW
TRIT
0
10
20
30
40
50
60
70
80
90
Bizerte Cap Bon N Cap Bon S Jendouba Manouba Kef Zaghouan
Inci
de
nce
%
Regions
BW
DW
TRIT

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
31
Figure 5. Severity of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops (bread wheat,
durum wheat and triticale)
3.2.Incidence of Septoria tritici blotch on commercial wheat varieties
Even though the variety distribution between years and surveyed areas varied, the
general trends show that most durum wheat varieties were highly susceptible to STB at different
levels (Figures 6 and 7). The disease incidence reached 100% on the commercial durum wheat
varieties ‘Saragolla’, followed by ‘Soudaine’ (90%), ‘Carioca’ (80%) and ‘Sculpture’ (60%).
High incidence was also recorded on the lead commercial durum wheat ‘Razzak’ (75%),
‘Maali’ (60%), and ‘Karim’ (45%) (Figure 6). The lower incidence of the local cultivars was
showed by the low STB levels at Cap Bon region, particularly that of ‘Karim’ that could have
been affected by late planting. Despite the high STB disease pressure on durum wheat across
the surveyed areas, it was nearly absent at El Haouaria (Cap Bon North) where mainly bread
wheat was cultivated. In 2016 high STB incidence (90%) and severity (70%) were observed
mainly on the bread wheat landrace ‘Farina arbi’ at El Haouaria (Figure 6). Mean incidence and
severity of 30 and 25%, respectively, were recorded on the bread wheat ‘Salammbô’. Lower
rates (<10%) were recorded on other commercial bread wheat varieties such as ‘Zanzibar’,
‘Utique’ and ‘Haïdra’, which were below 10%.When tested at experimental station in Northern
Tunisia, ‘Farina arbi’ and the other bread wheat varieties showed no infection of STB despite
high levels of infection on most if not all commercial durum wheat varieties. In addition, low
levels of susceptibility to STB were recorded on triticale varieties where incidence and severity
ranged from 0% to 30%. Out of three triticale varieties, the disease was totally absent on
‘Bienvenue’ (Figures 6 and 7).
0
10
20
30
40
50
60
70
80
Beja Bizerte Cap Bon N Cap Bon S Jendouba Manouba Kef Zaghouan
Seve
rity
%
Regions
BW
DW
TRIT

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
32
Figure 6. Incidence of STB on durum wheat, bread wheat and triticale varieties.
Figure 7. Severity of STB on durum wheat, bread wheat and triticale varieties.

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
34
Gerbreslassie, 2015; Tekele et al., 2015; ÜNAL et al., 2017) showed that the impact and
distribution of diseases varied due to the continuous release and extensive cultivation of
susceptible varieties. Thus, the magnitude of virulence and disease incidence are variable and
closely related to the frequency of the variety used in a particular area as well as the proportion
of durum wheat area as compared to that of bread wheat (Yahyaoui et al., 2000).
Testing ‘Farina arbi’ land race for its resistance/susceptibility to Z. tritici at other
Northern regions where durum wheat is mostly cultivated showed no STB infection. This
unique bread wheat landrace, completely susceptible at El Haouaria (North Eastern Tunisia)
and completely resistant at Béjà Northwestern Tunisia, could be that we are definitely dealing
with two distinct Z. tritici populations and could give more highlight on STB specificity. Further
studies will be conducted to characterize the STB populations from El Haouaria that are mostly
specific to the bread wheat landrace ‘Farina arbi’ and have no effect on other bread and durum
wheat varieties. Such phenomenon has not been observed before and could lead to further
understanding of STB host specificity.
5. Conclusion
The survey data revealed low risk of Z. tritici on bread wheat except at Cap Bon region
especially at El Haouaria where Septoria tritici blotch severity was relatively high on the old
bread wheat landrace, while rare occurrence at other sites was observed on some commercial
bread wheat varieties. High incidence and severity were observed on triticale across the
surveyed fields. Although Tunisia is primarily a durum-wheat producing country with Z. tritici
being mostly prevalent on durum wheat; bread wheat is of great economic importance, even
though it occupies small areas. The occurrence of STB on the landrace could lead to
development of Septoria population that could become of major importance on bread wheat as
is the case in Morocco and other regions. The presence of an STB population at one site and
infecting a single cultivar will be further investigated and will possibly lead to better
understanding of Z. tritici population dynamics that could become an important tool in
screening for disease resistance.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’
populations sampled from bread wheat in Tunisia revealed by
SSR markers
Bel Hadj Chedli Rim1,2, Aouini Lamia3, Ben M’Barek Sarrah4,2, Bochra Amina Bahri5,6,
Verstappen Els7, Kema Gerrit H.J.7, Rezgui Salah1, Yahyaoui Amor8,2, Chaabane Hanène6
1Laboratory of genetics and plant breeding, National Agronomic Institute of Tunisia (INAT), 43 Avenue Charles
Nicolle, 1002 Tunis, Tunisia. 2 CRP Wheat Septoria Precision Phenotyping Platform, Tunisia. 3 Department of Agronomy, Purdue University, 915 West State Street, West Lafayette, IN 47907 USA. 4 Regional Field Crops Research Center of Beja (CRRGC) BP 350, 9000 Beja, Tunisia. 5 Department of Plant Pathology, College of Agricultural and Environmental Sciences, University of Georgia, 228
Turfgrass Research and EducationCenter, 1109 Experiment Street, Griffin, GA 30223 USA. 6Laboratory of Bioagressors and Integrated protection in agriculture, National Agronomic Institute of Tunisia
(INAT), 43 Avenue Charles Nicolle, 1002 Tunis, Tunisia. 7 WageningenUniversity and Research Center, Wageningen, The Netherlands. 8 Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), km 45 Carretera México-Veracruz El Batán,
Texcoco.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
35
Objectifs et démarche
Plusieurs marqueurs moléculaires ont été utilisés pour analyser la diversité génétique des
populations de Z. tritici issues de nombreux pays à travers le monde. Toutefois, l’observation
récente d’une grande incidence de Z. tritici sur blé tendre dans la région du Cap Bon comme
nous l’avons souligné dans le chapitre 2, montre l’importance et l’urgence d’entamer une
caractérisation moléculaire de la diversité génétique des populations de ce pathogène dans cette
région.
C’est dans ce cadre que s’insère l’objectif principal de ce chapitre qui permet d’étudier la
diversité et la structure génétique d’une population de 184 isolats de Z. tritici isolés à partir des
feuilles de blé tendre infestées naturellement dans trois régions céréalières du Nord de la Tunisie
(Cap Bon, Béjà et Bizerte) moyennant 12 marqueurs microsatellites.
De plus, une approche bio-informatique a été appliquée pour déterminer la diversité
génétique (H) (Nei, 1973), l’indice de Shannon (I), le polymorphisme (P), le flux de gène (Nm),
la différentiation génétique entre les populations (Fst) et la structure génétique des populations
de Z. tritici à deux échelles : champ et région. Les proportions de la distribution de deux types
sexules Mat1-1 et Mat1-2 dans tous les champs ont été aussi calculées dans cette étude.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
36
Abstract
Septoria tritici blotch (STB) is the primary biotic stress for durum wheat production in
Tunisia and has not been common on bread wheat. During the 2015-2016 growing season,
strong STB disease severity was observed in the littoral Tunisian area on an old bread wheat
landrace known as “Farina Arbi” particularly at ‘El Haouaria’ region. A total of 184 single-
pycnidial fungal isolates were sampled from nine naturally infected bread wheat fields in three
main wheat growing area in Tunisia (Cap Bon area, Bizerte and Béjà). The collected isolates
were fingerprinted using 12 polymorphic microsatellite (SSR) markers in order to assess the
genetic diversity and population structure of Zymoseptoria tritici (Z. tritici) at regional and field
scale levels. A high genetic diversity was observed within the collected Z. tritici population,
with the highest Nei’s index value (0.42), Shannon Index (0.84), and important private alleles
number (36) at El Haouaria region. However, moderate population differentiation (Fst=0.16)
and a high gene flow (Nm =1.85) were observed between the nine sampled fields across the
surveyed regions. A lack of genetic structure was observed at both regional and field levels.
The high degree of diversity was likely due to an active sexual recombination in the investigated
areas, as revealed by equal proportions of the two mating types that was also assessed during
this study.
Key words: Bread wheat, genetic diversity, population structure, Zymoseptoria tritici.
1. Introduction
The cereal sector is one of the pillars of the Tunisian agriculture in terms of its output and
cultivated area with nearly a total of 1.5 million of hectares (Ben Hamouda et al., 2016). It is
characterized by a predominance of durum wheat (Triticum turgidum L. subsp. durum (Desf.)
van Slageren) that has been cultivated in Tunisia since the Roman era while the bread wheat
(Triticum aestivum L. subsp. aestivum) was introduced relatively recently after the French
colonization (El Felah et al., 2015). Since then, bread wheat was commonly cultivated in
mixture with durum wheat landraces (Ben Hammouda et al., 2016). Tunisian farmers and rural
communities used the Arabic word “Gameh” to designate durum wheat against the non-Arabic
word “Farina” to indicate bread wheat (Ammar et al.,2011). The gain in wheat production has
often been hampered by low yields caused by drastic losses due to abiotic and biotic constraints.
Septoria tritici blotch (STB) caused by the hemibiotrophic fungus Zymoseptoria tritici (Z.
tritici) (Desm.) Quaedvlieg and Crous (formerly Mycosphaerella graminicola [Fuckel] J.
Schröt. in Cohn), is an important disease in North Africa and particularly in Tunisia where an

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
37
early widespread epidemic occurred between 1968-1969 (Saari and Wilcoxcon, 1974; Brown
et al., 2015). Epidemics have become recurrent since the 1990’s. In Tunisia, STB mainly
infects durum wheat (Triticum turgidum L. ssp. durum (Desf.), genome BBAA), particularly in
the North Western region to include the main durum wheat producing districts of Bizerte, Béjà
and Jendouba (Fakhfakh et al., 2011). Nevertheless, Z. tritici also infects the hexaploid wheat
(Triticum aestivum, L., genome, AABBDD) exclusively in the North Eastern region (Bel Hadj
Chedli et al. 2018).
Z. tritici has a heterothallic bipolar mating system with two mating type alleles, mat1-1
and mat1-2 (Waalwijk et al. 2002) and undergoes sexual and asexual sporulation. Sexual
reproduction requires contact between the two separate mating types (Kema et al., 1996),
resulting in fruiting bodies (pseudothecia) that contain ascospores (Eyal et al., 1987). These
latter are released from wheat debris, dispersed by wind and hence constitute the primary
inoculum (Ponomarkenko et al., 2011; Suffert et al., 2011). An evident and frequent sexual
reproduction has been observed in most Z. tritici populations worldwide as even frequencies of
both mating types idiomorphs were observed (Waalwijk et al. 2002; Siah et al. 2010). On the
other hand, through asexual reproduction, pycnidiospores that are formed in the pycnidia, are
locally dispersed by rain splash and are considered as secondary source of inoculum,
contributing thus to the disease progression during the cropping season (Steinberg, 2015).
Since sexual reproduction of this fungus may occur year-round, it plays a crucial role in
its epidemiology and has a major impact on its population structure (Zhan et al., 2003;
McDonald, 2015). It maintains the diversity of the pathogen populations that are more likely to
adapt to selection pressures such as those exerted by resistant hosts or fungicides treatments
compared to populations with less genetic variability (McDonald et al., 1997; Zhan and
McDonald; 2004).
Genetic structure is the study of genetic variation within and among populations and
reflects thus its evolutionary history and its potential to evolve (Banke and McDonald, 2005).
Many factors contribute to genetic change (i.e., evolution) within populations. These factors
include mutation, mating systems/recombination, gene flow or migration, population size, and
selection (McDonald et al., 1997). In nature, all of these forces interact to determine the course
of evolution of a pathogen and to generate the genetic structure of its populations. According
to McDonald and Linde (2002), pathogens that generate the greatest risk of breaking down
resistance genes have a mixed reproduction system, a high potential for gene flow, large
effective population sizes, and high mutation rates compared to pathogens with strict asexual
reproduction that have low potential for gene flow, small effective population sizes, and low

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
38
mutation rates. Therefore, information on the distribution of genetic variation is important and
could be used to deploy more efficient control strategies against Z. tritici (McDonald and Mundt
2016).
In many countries, population genetic studies using RFLP, AFLP and SSR markers,
showed that Z. tritici-populations are characterized by high genetic diversities and low genetic
differentiation between populations (Schnieder et al., 2001; Linde et al., 2002; Zhan et al.,
2003; Banke and McDonald, 2005). Significant rates of population structure were also reported
in many studies, and important genetic differentiation was noted within and among different Z.
tritici populations sampled at different levels and from many geographical locations: Czech
republic, Iran, California, Indiana, Kansas, North Dakota, France (Linde et al. 2002; Abrinbana
et al. 2010; El Chartouni et al., 2011; Gurung et al., 2011; Drabesova et al., 2013).
In Tunisia, the genetic structure of Z. tritici has been extensively studied mainly using
SSR markers which revealed a strong genetic diversity (Berraies et al., 2013; Boukef, 2012,
Nouari et al., 2016). Although, these studies were concentrated only on populations sampled
from durum wheat. Studying the genetic diversity of the Tunisian bread wheat Z. tritici
population may lead to a better understanding of the epidemiological and evolutionary driving
forces in Z. tritici in Tunisia, particularly at El Haouaria region where STB was recently
intensified on bread wheat ‘Farina Arbi’. In this study, we present the first description of a
Tunisian population of Z. tritici collected from bread wheat which was sampled from three
provinces throughout Northern Tunisia (Béjà, Bizerte and Cap Bon Area).
This study aims to evaluate the molecular polymorphism and the genetic diversity of Z.
tritici at regional and field scales with a focus on the Cap-Bon area region where previously,
high STB severity was reported on an old bread wheat landrace and to estimate the gene flow
among the studied locations and finally to investigate the population structure.
2. Materials and Methods
2.1. Fungal sampling and isolation
Leaf samples were collected from nine bread wheat fields located at three regions in
Northern Tunisia: Cap Bon area (El Haouaria), Bizerte (Ichkeul) and Béjà (Goubellat and Oued
Zarga) where STB infections were reported during 2015-2016 cropping season (Bel Hadj
Chedli et al., 2018) (Figure1). Out of the nine assessed fields, four fields were located at El
Haouaria region where the bread wheat landrace ‘Farina Arbi’ was grown as a monoculture
over several years and two fields where cropped with the variety ‘Salammbô’. Three sampled

Chapitre 2. Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and
bread wheat in Northern Tunisia
33
4. Discussion
The response of durum wheat, bread wheat, and triticale to Z. tritici varied according to
the crop species. During the surveyed period, Z. tritici was more prevalent on durum wheat at
the majority of surveyed areas except Cap Bon North (El Haouaria) and confirms the high to
moderate risk of STB at Northern and Cap Bon regions of Tunisia, respectively.
This result supports conclusions of previous reports and confirms that Septoria diseases
hot spots are prevalent in the sub humid and semi-arid areas at the beginning of winter season
(Fakhfakh et al., 2011). The new commercial durum wheat ‘Sculpture’, ‘Saragolla’, ‘Carioca’
and ‘Soudaine’ were susceptible to Septoria as they were mainly grown at Septoria hot spots
where monoculture of durum wheat particularly the susceptible ‘Karim’ and relatively high
rainfall contributed to the development of high infection levels. In particular, high incidence
and severity were recorded on ‘Karim’ and ‘Razzak’, which confirmed previous findings
conducted by Ltifi and Sakkouhi (2008), and Ben Mohamed et al. (2000). In contrast, ‘Maali’,
which was previously characterized by a good level of resistance in Béjà (Gharbi et al., 2011),
was susceptible to STB in the majority of surveyed areas in this study, which could be explained
by a slow decline of host resistance (Kema et al., 2018). The survey data also revealed that
triticale was also susceptible to STB across the majority of surveyed areas posing therefore a
serious threat to this crop.
On the other hand, the data revealed that STB was very low in the majority of inspected
regions on the commercial bread wheat cultivars such as ‘Haïdra’, ‘Vaga’, ‘Utique’, and
‘Zanzibar’, which could explained by the relative resistance of these varieties to Septoria (Ben
Hamouda et al., 2016) while it was higher in ‘Salammbô’ (Saade, 1996). ‘Salammbô’ was
released in 1980, period that has known a substantial expansion in bread wheat acreage
particularly for varieties with high yield and good level of diseases resistance. It seems that this
variety has undergone a slow decline of host resistance over time that is commonly observed in
this pathosystem, particularly for bread wheat in Europe (Kema et al., 2018).
Surprisingly from this study, STB on bread wheat poses a great risk only at one region,
El Haouaria, where it was rated class IV and V and mainly only on the old bread wheat landrace
‘Farina arbi’. The important incidence of Septoria observed on this variety reveals a specific
presence of Z. tritici population that only develops on this old bread wheat landrace with little
or no apparent effect on other bread wheat varieties. This could be mainly associated with the
wheat-based mono-cropping system and monoculture of this landrace over several decades
facilitating thereby the adaptation of the pathogen to this specific variety (Holloway, 2014;
McDonald and Mundt, 2016). Similar research reviews on wheat diseases surveys (Teferi and

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
39
fields located at Béjà and Bizerte were cropped with the cultivars ‘Utique’ and ‘Zanzibar’,
respectively. Sampling details and GPS coordinates were shown in Table 1.
Hierarchical sampling was carried out according to McDonald et al. (1999) where
infected leaves were obtained from six to nine locations approximately 10 m apart within
individual fields. From each location, five leaves were randomly collected from different plants
and only one isolate was selected from each leaf for genotyping. Isolates from each region are
considered as separate populations and isolates sampled from each field are referred as sub-
populations. Mono-pycnidial isolates were obtained as described by Siah et al. (2010) resulting
in a total of 184 Z. tritici isolates (Table 1) that were subsequently grown on PDA medium
(potato dextrose agar, 39 g L−1) and stored at (-80°C) for further analyses.
2.2. DNA extraction and mating type’s determination
Fungal DNA was extracted using the Sbeadex® mini plant kit (LGC genomics) extraction
performed on a King Fisher KF96 system according to the manufacturer's instructions. Mating
type of each isolate was identified using a multiplex Polymerase Chain Reaction (PCR)
amplification of partial two mating type loci as described by Waalwijk et al. (2002) (Table 2).
PCR for mating type amplifications were performed using a mixture of 2.5µL (6µM) of each
primer, 5µL (600 µM) of dNTPs, 5 µLof 10X reaction buffer, 0.25µL(5U/ul) of Taq polymerase
(Ampli Taq Gold, Roche, Someville MA) and 1 µL (10 ng to 20 ng/µL) of genomic DNA. The
total PCR reaction volume was adjusted with nuclease free water to 50 µL per PCR reaction.
DNA samples were amplified using an MJ Research PT-100 Thermocycle (Biorad, Hercules,
CA) adjusted at thermal cycling conditions that started with an initial denaturation at 94°C for
2 min, followed by 39 cycles at 94°C for 1 min for denaturation, and at 68°C for 30 min for
annealing, and a final extension at 72°C for 1 min. DNA samples were subsequently incubated
at 72°C for 10 min for a final elongation and stored at 4°C for further processing. Amplification
products were stained with GelRed, separated in 1% agarose gel and electrophoresis at 100 V
for 45 min and visualized under UV light.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in Tunisia revealed by SSR markers
40
Table 1. Origin, number and geographical coordinates of Zymoseptoria tritici isolates used in this study
Regions Year Location Host Cultivar Field
number
Isolates
number
Altitude
range
Longitude (N) (range) Latitude (E)
(range)
Bizerte 2015 Ichkeul Bread wheat Zanzibar 1 8 16-354 09°03’1 09°69’83 32°50’56 37°14’12
Béjà 2015 Goubellat Bread wheat Utique 1 3 11-876 09°09’01 10°78’62 36°47’47 36°92’68
Oued Zarga Bread wheat Unknown 1 13
Cap Bon 2015 El Haouaria Bread wheat Salammbô 2 18 18-290 10°02’73 10°49’10 36°22’26 36°68’31
El Haouaria Bread wheat Farina Arbi 4 120
Total 9 162
Figure 1. Schematic map of the sampling locations in Northern Tunisia. A: El Haouaria region, B: Bizerte
(Ichkeul), C: Goubellat; D: Oued Zarga.
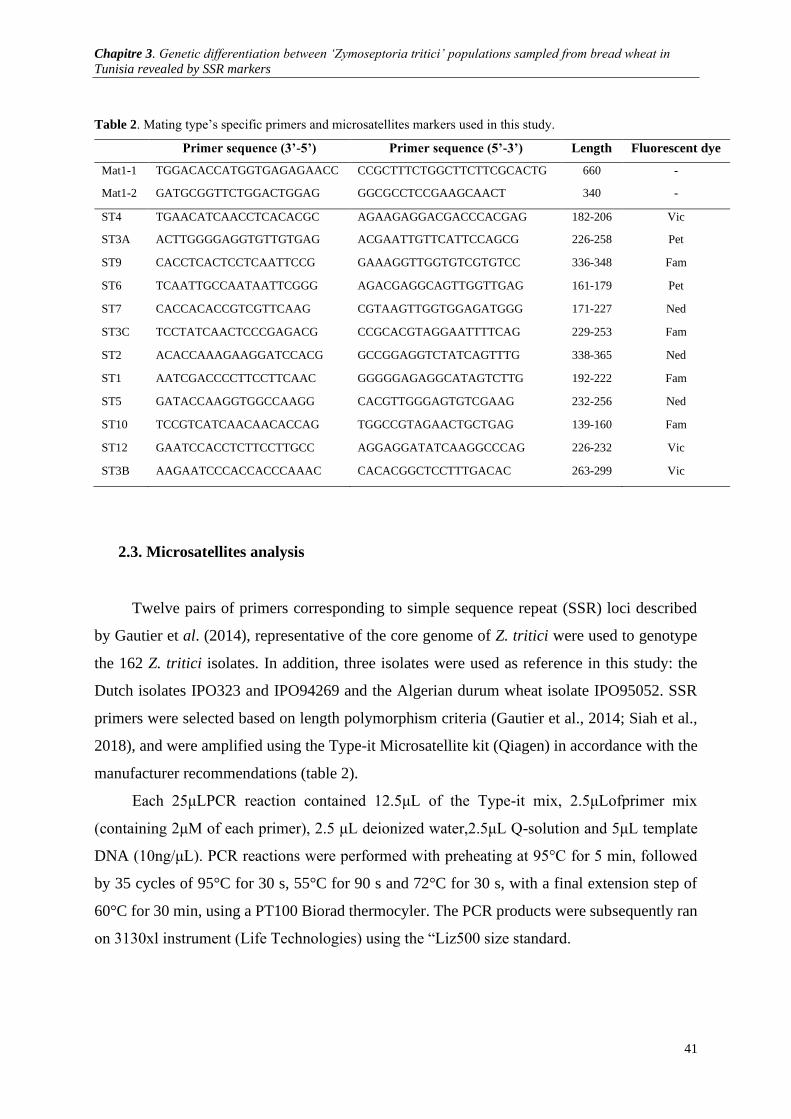
Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
41
Table 2. Mating type’s specific primers and microsatellites markers used in this study.
Primer sequence (3’-5’) Primer sequence (5’-3’) Length Fluorescent dye
Mat1-1 TGGACACCATGGTGAGAGAACC
T
CCGCTTTCTGGCTTCTTCGCACTG 660 -
Mat1-2 GATGCGGTTCTGGACTGGAG GGCGCCTCCGAAGCAACT 340 -
ST4 TGAACATCAACCTCACACGC AGAAGAGGACGACCCACGAG 182-206 Vic
ST3A ACTTGGGGAGGTGTTGTGAG ACGAATTGTTCATTCCAGCG 226-258 Pet
ST9 CACCTCACTCCTCAATTCCG GAAAGGTTGGTGTCGTGTCC 336-348 Fam
ST6 TCAATTGCCAATAATTCGGG AGACGAGGCAGTTGGTTGAG 161-179 Pet
ST7 CACCACACCGTCGTTCAAG CGTAAGTTGGTGGAGATGGG 171-227 Ned
ST3C TCCTATCAACTCCCGAGACG CCGCACGTAGGAATTTTCAG 229-253 Fam
ST2 ACACCAAAGAAGGATCCACG GCCGGAGGTCTATCAGTTTG 338-365 Ned
ST1 AATCGACCCCTTCCTTCAAC GGGGGAGAGGCATAGTCTTG 192-222 Fam
ST5 GATACCAAGGTGGCCAAGG CACGTTGGGAGTGTCGAAG 232-256 Ned
ST10 TCCGTCATCAACAACACCAG TGGCCGTAGAACTGCTGAG 139-160 Fam
ST12 GAATCCACCTCTTCCTTGCC AGGAGGATATCAAGGCCCAG 226-232 Vic
ST3B AAGAATCCCACCACCCAAAC CACACGGCTCCTTTGACAC 263-299 Vic
2.3. Microsatellites analysis
Twelve pairs of primers corresponding to simple sequence repeat (SSR) loci described
by Gautier et al. (2014), representative of the core genome of Z. tritici were used to genotype
the 162 Z. tritici isolates. In addition, three isolates were used as reference in this study: the
Dutch isolates IPO323 and IPO94269 and the Algerian durum wheat isolate IPO95052. SSR
primers were selected based on length polymorphism criteria (Gautier et al., 2014; Siah et al.,
2018), and were amplified using the Type-it Microsatellite kit (Qiagen) in accordance with the
manufacturer recommendations (table 2).
Each 25μLPCR reaction contained 12.5μL of the Type-it mix, 2.5μLofprimer mix
(containing 2μM of each primer), 2.5 μL deionized water,2.5μL Q-solution and 5μL template
DNA (10ng/μL). PCR reactions were performed with preheating at 95°C for 5 min, followed
by 35 cycles of 95°C for 30 s, 55°C for 90 s and 72°C for 30 s, with a final extension step of
60°C for 30 min, using a PT100 Biorad thermocyler. The PCR products were subsequently ran
on 3130xl instrument (Life Technologies) using the “Liz500 size standard.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
42
2.4. Data analysis
Molecular data analyses were carried out using the GenALEx software version 6.5
(Peakall and Smouse, 2012). The Genetic diversity was assessed by investigating the number
of private alleles (alleles found only in a single location), the Nei’s genetic diversity (H) (Nei
1973) and the Shannon’s information Index (I). The relationship between the Nei’s genetic
diversity (GD) and the geographic distance (GGD) was investigated using Mantel test
implemented in GenALEx 6.5 in order to examine the level of genetic isolation by geographical
distances. Gene flow (Nm) within and among population and the percentage of polymorphic
loci present at frequencies>1% were also determined.
The magnitude of the genetic differentiation according to each locus was assessed using
the Nei’s Fst fixation index (Nei, 1973) implemented in GenALEX 6.5 in order to assess the
degree of genetic differentiation between populations and to identify the correlation between
alleles within populations relative to the entire population. Genetic variation within and among
populations was further partitioned by analysis of molecular variance (AMOVA). The
relationship between individuals was calculated using principal coordinate analysis (PCoA) to
detect genetic divergence among subpopulations (Sun et al. 2013).
Population structure was inferred using the Structure 2.3.4 program (Pritchard et al. 2000)
which uses Bayesian algorithm to estimate the number of populations. The burn-in period was
followed by a run phase of 100000 iterations, with the number of clusters K ranging from 1 to
10, and 10 replicates for each value of K. A dendrogram estimating genetic clustering of the
studied Z. tritici population was subsequently produced using the weighted neighbor-joining
method based on the dissimilarity matrix (500 bootstraps), as implemented in the DARwin6
software (Perrier and Jacquemoud-Collet, 2006).
For mating type’s determination, among 162 Z. tritici isolates, only 141 were further
assessed for mating type analysis. the null hypothesis of a 1:1 ratio of two mating types within
each collection at different scales among and within regions, was evaluated using a χ2-test at
the significance level of P=0.05 (Waalwijk et al. 2002).
3. Results
3.1. Genetic diversity within and among sampled populations
In this study, 162 Z. tritici isolates collected from nine bread-wheat fields were analyzed
using 12 microsatellites markers in order to assess the genetic diversity and structure of a Z.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
43
tritici population collected from different region and field scales in Northern Tunisia. Isolates
that did not amplify for more than 6 loci (50%) were removed from the data set and only a total
of 162 isolates were then conserved. A multilocus analysis identified 128 distinct genotypes
(MLG) among 162 Z. tritici isolates (Table 3). The most important MLG number (116) were
identified at El Haouaria region and a total of 4 multilocus were shared between El Haouaria
fields while only one multilocus was shared between Goubellat, Oued Zarga and Bizerte
populations (Table 3).
Results revealed that El Haouaria population possessed the highest number of private
alleles (36), whereas Bizerte isolates presented only one private allele with a frequency of
0.33% (Annexe 1, Table 2,). No private alleles were observed within Oued Zarga population
(Table 3).
Of the 12 loci scored, 100% were polymorphic at El Haouaria population and 75% were
polymorphic at Bizerte and Goubellat populations while 0% polymorphism was observed on
Oued Zarga population (Béjà region) (Table 3). Allele diversity analyses revealed high levels
of genetic diversity among the studied populations. Within the region scale, the most important
Nei's gene diversity (0.4) and Shannon's index (0.8) were recorded at El Haouaria region
whereas there were similar at Goubellat and Bizerte regions with (0.5) and (0.3) respectively
(Table 3).
At the field scale, the Nei’s gene diversity and Shannon's indexes were approximately
similar at El Haouaria region and averaged of 0.7 and 0.4 values respectively with very slight
variations among the sub-collections (field populations).
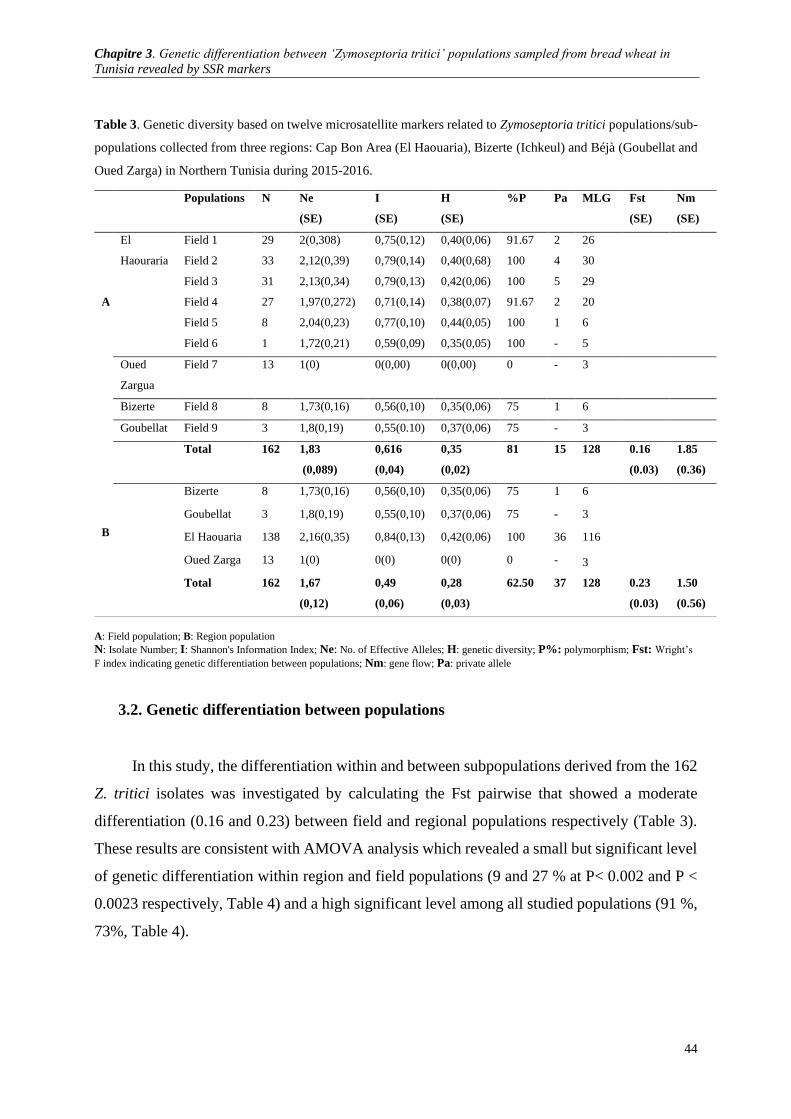
Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
44
Table 3. Genetic diversity based on twelve microsatellite markers related to Zymoseptoria tritici populations/sub-
populations collected from three regions: Cap Bon Area (El Haouaria), Bizerte (Ichkeul) and Béjà (Goubellat and
Oued Zarga) in Northern Tunisia during 2015-2016.
A: Field population; B: Region population
N: Isolate Number; I: Shannon's Information Index; Ne: No. of Effective Alleles; H: genetic diversity; P%: polymorphism; Fst: Wright’s
F index indicating genetic differentiation between populations; Nm: gene flow; Pa: private allele
3.2. Genetic differentiation between populations
In this study, the differentiation within and between subpopulations derived from the 162
Z. tritici isolates was investigated by calculating the Fst pairwise that showed a moderate
differentiation (0.16 and 0.23) between field and regional populations respectively (Table 3).
These results are consistent with AMOVA analysis which revealed a small but significant level
of genetic differentiation within region and field populations (9 and 27 % at P< 0.002 and P <
0.0023 respectively, Table 4) and a high significant level among all studied populations (91 %,
73%, Table 4).
Populations N Ne
(SE)
I
(SE)
H
(SE)
%P Pa MLG Fst
(SE)
Nm
(SE)
A
El
Haouraria
Field 1 29 2(0,308) 0,75(0,12) 0,40(0,06) 91.67 2 26
Field 2 33 2,12(0,39) 0,79(0,14) 0,40(0,68) 100 4 30
Field 3 31 2,13(0,34) 0,79(0,13) 0,42(0,06) 100 5 29
Field 4 27 1,97(0,272) 0,71(0,14) 0,38(0,07) 91.67 2 20
Field 5 8 2,04(0,23) 0,77(0,10) 0,44(0,05) 100 1 6
Field 6 1 1,72(0,21) 0,59(0,09) 0,35(0,05) 100 - 5
Oued
Zargua
Field 7 13 1(0) 0(0,00) 0(0,00) 0 - 3
Bizerte Field 8 8 1,73(0,16) 0,56(0,10) 0,35(0,06) 75 1 6
Goubellat Field 9 3 1,8(0,19) 0,55(0.10) 0,37(0,06) 75 - 3
Total 162 1,83
(0,089)
0,616
(0,04)
0,35
(0,02)
81 15 128 0.16
(0.03)
1.85
(0.36)
B
Bizerte 8 1,73(0,16) 0,56(0,10) 0,35(0,06) 75 1 6
Goubellat 3 1,8(0,19) 0,55(0,10) 0,37(0,06) 75 - 3
El Haouaria 138 2,16(0,35) 0,84(0,13) 0,42(0,06) 100 36 116
Oued Zarga 13 1(0) 0(0) 0(0) 0 - 3
Total 162 1,67
(0,12)
0,49
(0,06)
0,28
(0,03)
62.50 37 128 0.23
(0.03)
1.50
(0.56)
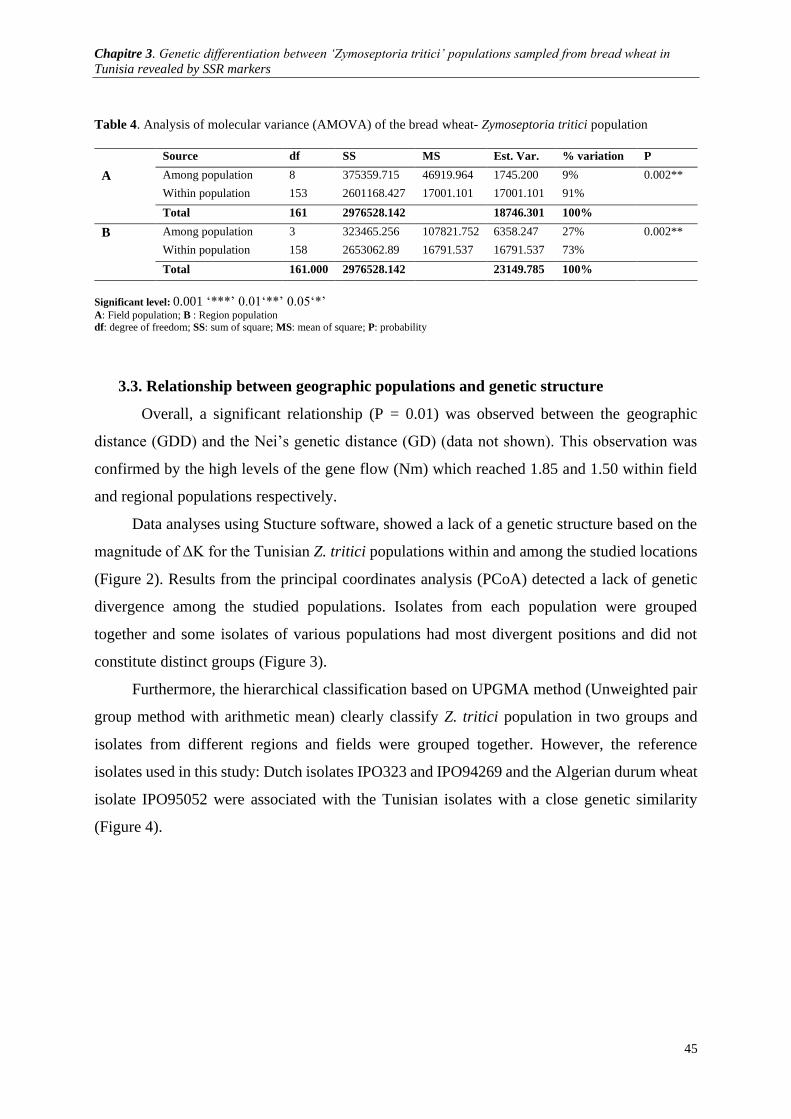
Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
45
Table 4. Analysis of molecular variance (AMOVA) of the bread wheat- Zymoseptoria tritici population
Significant level: 0.001 ‘***’ 0.01‘**’ 0.05‘*’ A: Field population; B : Region population df: degree of freedom; SS: sum of square; MS: mean of square; P: probability
3.3. Relationship between geographic populations and genetic structure
Overall, a significant relationship (P = 0.01) was observed between the geographic
distance (GDD) and the Nei’s genetic distance (GD) (data not shown). This observation was
confirmed by the high levels of the gene flow (Nm) which reached 1.85 and 1.50 within field
and regional populations respectively.
Data analyses using Stucture software, showed a lack of a genetic structure based on the
magnitude of ΔK for the Tunisian Z. tritici populations within and among the studied locations
(Figure 2). Results from the principal coordinates analysis (PCoA) detected a lack of genetic
divergence among the studied populations. Isolates from each population were grouped
together and some isolates of various populations had most divergent positions and did not
constitute distinct groups (Figure 3).
Furthermore, the hierarchical classification based on UPGMA method (Unweighted pair
group method with arithmetic mean) clearly classify Z. tritici population in two groups and
isolates from different regions and fields were grouped together. However, the reference
isolates used in this study: Dutch isolates IPO323 and IPO94269 and the Algerian durum wheat
isolate IPO95052 were associated with the Tunisian isolates with a close genetic similarity
(Figure 4).
Source df SS MS Est. Var. % variation P
A Among population 8 375359.715 46919.964 1745.200 9% 0.002**
Within population 153 2601168.427 17001.101 17001.101 91%
Total 161 2976528.142 18746.301 100%
B Among population 3 323465.256 107821.752 6358.247 27% 0.002**
Within population 158 2653062.89 16791.537 16791.537 73%
Total 161.000 2976528.142 23149.785 100%
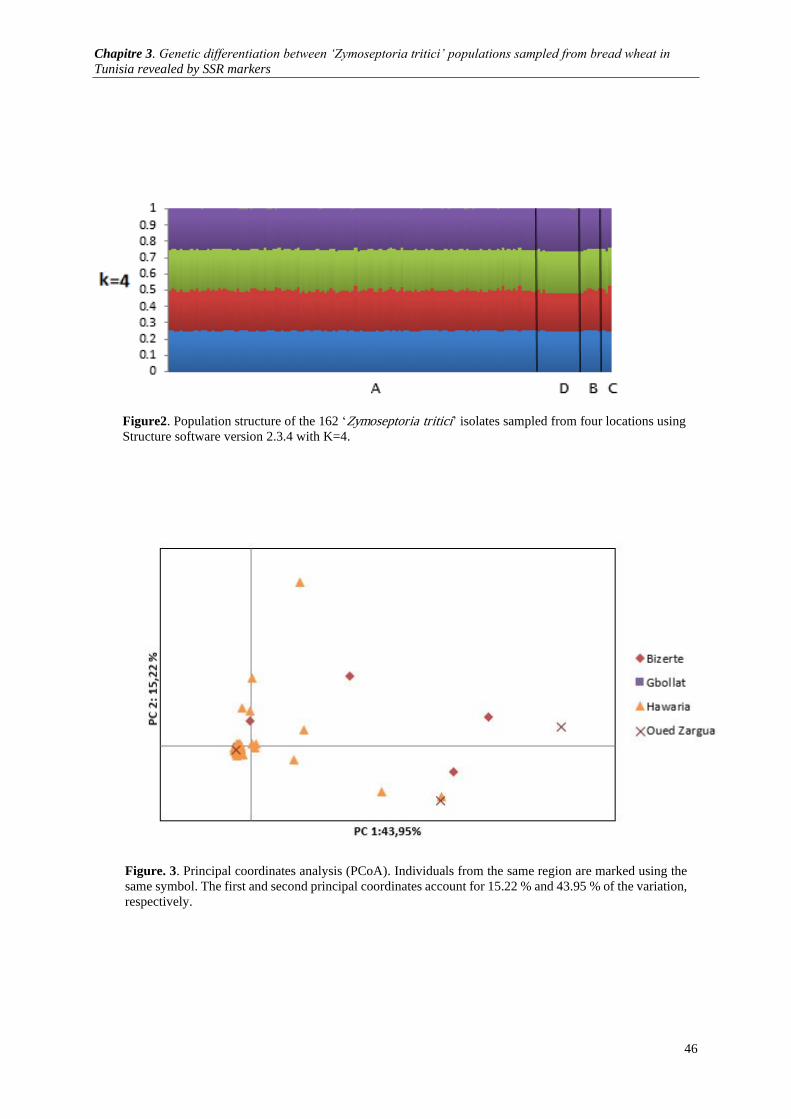
Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
46
Figure2. Population structure of the 162 ‘Zymoseptoria tritici’ isolates sampled from four locations using
Structure software version 2.3.4 with K=4.
Figure. 3. Principal coordinates analysis (PCoA). Individuals from the same region are marked using the
same symbol. The first and second principal coordinates account for 15.22 % and 43.95 % of the variation,
respectively.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
47
3.4. Occurrence of sexual reproduction
The results of this study indicate that the two mating types coexisted in fungal populations
sampled from all geographical scales including Cap Bon and Bizerte but also in different fields
within the same region (Table 5).
While the absolute ratio of the two mating types varied among sampling units (fields, and
locations), none of the differences differed significantly from the null hypothesis that the same
frequency of mating types existed among all sampling units (Table 5) except at Béjà region,
where MAT1-2 was exclusively predominant across 13 Z. tritici isolates and a significant
deviation (P<0.05) from the expected 1:1 ratio was noted.
Fig 4. Dendrogram showing the genetic clustering of the 162 Zymospetoria. Tritici isolates sampled from bread
wheat across three geographic locations in Northern Tunisia. The tree was constructed using the weighted neighbor-
joining method implemented in DARwin 6 software. Isolates from the same field were indicated with the same
color. Fields 1,2,3,4,5 and 6 are located at Ca Bon Area, fields 7 and 8 are located at Beja while field 9 belongs to
Bizerte region.
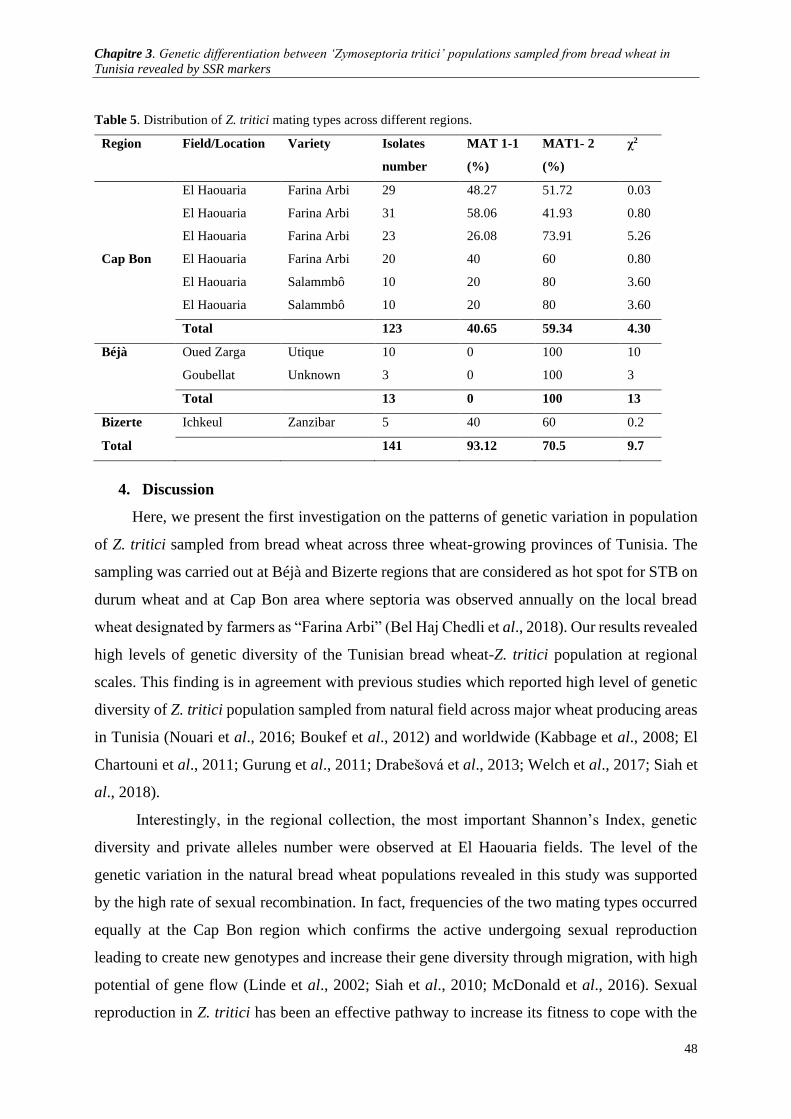
Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
48
Table 5. Distribution of Z. tritici mating types across different regions.
Region Field/Location Variety Isolates
number
MAT 1-1
(%)
MAT1- 2
(%)
χ2
Cap Bon
El Haouaria Farina Arbi 29 48.27 51.72 0.03
El Haouaria Farina Arbi 31 58.06 41.93 0.80
El Haouaria Farina Arbi 23 26.08 73.91 5.26
El Haouaria Farina Arbi 20 40 60 0.80
El Haouaria Salammbô 10 20 80 3.60
El Haouaria Salammbô 10 20 80 3.60
Total 123 40.65 59.34 4.30
Béjà Oued Zarga Utique 10 0 100 10
Goubellat Unknown 3 0 100 3
Total 13 0 100 13
Bizerte Ichkeul Zanzibar 5 40 60 0.2
Total 141 93.12 70.5 9.7
4. Discussion
Here, we present the first investigation on the patterns of genetic variation in population
of Z. tritici sampled from bread wheat across three wheat-growing provinces of Tunisia. The
sampling was carried out at Béjà and Bizerte regions that are considered as hot spot for STB on
durum wheat and at Cap Bon area where septoria was observed annually on the local bread
wheat designated by farmers as “Farina Arbi” (Bel Haj Chedli et al., 2018). Our results revealed
high levels of genetic diversity of the Tunisian bread wheat-Z. tritici population at regional
scales. This finding is in agreement with previous studies which reported high level of genetic
diversity of Z. tritici population sampled from natural field across major wheat producing areas
in Tunisia (Nouari et al., 2016; Boukef et al., 2012) and worldwide (Kabbage et al., 2008; El
Chartouni et al., 2011; Gurung et al., 2011; Drabešová et al., 2013; Welch et al., 2017; Siah et
al., 2018).
Interestingly, in the regional collection, the most important Shannon’s Index, genetic
diversity and private alleles number were observed at El Haouaria fields. The level of the
genetic variation in the natural bread wheat populations revealed in this study was supported
by the high rate of sexual recombination. In fact, frequencies of the two mating types occurred
equally at the Cap Bon region which confirms the active undergoing sexual reproduction
leading to create new genotypes and increase their gene diversity through migration, with high
potential of gene flow (Linde et al., 2002; Siah et al., 2010; McDonald et al., 2016). Sexual
reproduction in Z. tritici has been an effective pathway to increase its fitness to cope with the

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
49
constant changing environment and to overcome host resistance (Kema et al., 2018; McDonald
and Linde, 2002) and this may have resulted in the pathogen diversification in this region (Zhan
et al., 2003; Siah et al., 2013; Siah et al., 2018).
On the other hand, the predominance of Mat1-2 at Oued Zarga could not be attributed to
the lack or to the low rate of sexual recombination which was considered as the only
evolutionary process that affect gametic equilibrium (Zhan et McDonald, 2004; Abrinbana et
al., 2010) but probably to the biased and an insufficient sampling.
A high polymorphism (91 % and 100%) was recorded at six sampled fields at El Haouaria
region where annual incidence of the disease occurred on the old landrace ‘Farina Arbi’
exclusively cultivated in this area. In this context, McDonald and Mundt (2016) and Dalvant et
al. (2018) reported that fungal populations having higher degrees of polymorphism naturally
possess a greater number of genes to exchange which could consequently influence diversity in
pathogenicity.
In contrast, the fungal populations sampled from Bizerte and Goubellat appeared to
possess lower genetic diversity compared to El Haouaria collection. Oued Zarga Z. tritici
population seems to be not polymorphic and lower levels of genetic diversity was recorded at
this region. Moreover, monitoring of STB during recent years in Tunisia demonstrated that this
pathogen is more adapted to durum wheat in these regions where it was mostly cultivated
(Fakhfakh et al., 2011). In Tunisia, STB was found at a very lower severity in few bread wheat
fields in regions such as Bizerte, Goubellat and Oued Zarga (Bel Hadj Chedli et al., 2018).
Consequently, our sampled populations for these regions (Bizerte, Goubellat and Oued Zarga)
may be considered as small which led to biased results because of the limited sample sizes (El
Chatrouni et al., 2012). Hence, the lower genetic diversity of Z. tritici in these regions suggests
that sexual reproduction might be less active than in other regions.
AMOVA analysis revealed that 73% of genetic variation could be attributed to
differences within regional populations and 27% to differences among individuals within
population. Similar results were observed with Razavi and Hughes (2004) findings which
indicated that 88% and 12% of the genetic diversity in 90 isolates were related to intra-
population and inter-population diversity respectively. In contrast, Abrinbana et al. (2010)
reported that the most important genetic variation was attributed among Z. tritici populations
sampled from different wheat provinces in Iran.
Nonetheless, moderate differentiation between regional subpopulations was noted by Fst
index which could be reflected by the high values of gene flow (Nm).The important (Nm) index
as revealed in the present study which suggests the occurrence of a strong gene flow between

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
50
the studied locations would further explain the higher intrapopulation diversity compared to
interpopulation diversity (Dalvant et al., 2018). Even so, Siah et al. (2018) reported a none
significant genetic differentiation at the field, plant and leaf layer scales in the French Z. tritici
population whereas a significant genetic differentiation was observed among Z. tritici
populations and higher degree of homogenization over many countries: Syria, Iran, Algeria,
United Kingdom, Switzerland, Germany, France, Argentina, Australia, Iran, and Czech
Republic (Zhan et al., 2003; Jurgens et al., 2006; El Chartouni et al., 2011; Gurung et al., 2011;
Drabešová et al., 2013). Therefore, many researchers reported that genetic differentiation
depends on the geographical locations studied and the markers used (Linde et al., 2002;
Abrinbana et al., 2010).
However, in this finding, the sampling sites were located in regions where different wheat
genotypes were cultivated. The geographic barriers and the differences in the used bread wheat
varieties may lead to conductive conditions for an important gene flow between Z. tritici
populations across the studied regions (Abrinbana et al., 2010; Zhan and McDonald, 2004).
Data analyses using Mantel test confirms this conclusion and revealed a significant correlation
between the genetic and the geographic distance indicating a great relationship isolation-
distance within the global population.
In this study, no population structure was observed at regional and field scales. This
finding is in agreement with the previous study of Boukef et al. (2012), revealing a lack of a
genetic structure of the Tunisian durum wheat derived Z. tritici populations according to nuclear
microsatellite markers genotyping. The lack of structure has also been reported in the
mitochondrial DNA, with high levels of gene flow and an active sexual reproduction among
durum wheat Tunisian Z. tritici population (Naouari et al., 2016). Even so, several earlier
studies that reported a lack of population structure for Z. tritici at regional and global scales,
attributed the lack of a geographic structure to the spore dispersal that probably occurs over a
wide geographic area and thus to long distance gene flow (McDonald et al. 1999; Linde et
al.2002; Zhan et al.2003). Another possible explanation could be related to the sampled area,
the population size or to the type of marker used (SSR) that cannot accurately capture the small
genomic variation that would occur at a single nucleotide level (Vali et al., 2008; Siah et al.,
2018). However, using the high Single nucleotide polymorphisms (SNP), Gibriel (2019)
investigated a large-scale structural variation in accessory and core chromosomes in the Middle
East-bread and durum wheat Z. tritici population. In this context, Drabešová et al. (2012)
reported that different genetic markers are known to produce different levels of genetic structure
for the same samples.

Chapitre 3. Genetic differentiation between ‘Zymoseptoria tritici’ populations sampled from bread wheat in
Tunisia revealed by SSR markers
51
The Principal coordinates analysis (PCoA) confirmed the lack of genetic variation among
the studied populations where the majority of Z. tritici isolates of each population were grouped
together. However, few exceptions were observed with some isolates of various populations
which were divergent and constituted distinct groups. Bayesian and unweighted neighbor-
joining results are consistent with the lack of the genetic structure where no differentiated
clusters in the bread-wheat Z. tritici populations sampled from different geographical locations
were observed. Furthermore, the dendrogram showed that isolates from different regions and
fields tended to cluster together with the Dutch bread wheat isolates (IPO323, IPO94269) and
with the Algerian durum wheat isolate IPO95052 and were associated with a close genetic
similarity. This observation suggests that sources of inoculum originating from genetically
diverse Z. tritici local population spores conserved a limited familial structure (Berraies et al.,
2013).
In this study, the same genotypes were shared among different regional populations
studied which could be explained by a great migration between different populations or perhaps
by the presence of some individuals that had the same multilocus genotype by chance (Gurung
et al., 2011). However, additional sampling from many bread and durum wheat growing area
in Tunisia is needed to confirm this result and to determine the causes. To our knowledge, this
work is the first study assessing fungal genetic diversity on Z. tritici population sampled from
bread wheat in Tunisia at regional and field scales. This research could provide new information
to further explore the epidemiology and management strategies of Septoria tritici blotch disease
in Northern Tunisia.
5. Conclusion
This study revealed high genetic variation and a clear evidence for recombination within
and among geographic populations of Z. tritici on bread-wheat fields in Tunisia consistent with
sexual reproduction of this organism in nature. Moderate differentiation between populations
leading to an important gene flow and a lack of structuration were further demonstrated. To
further examine the genetic diversity of Z. tritici in Tunisia, larger population size with large
number of isolates and greater number of microsatellite markers should be considered to study
the genetic structure of Z. tritici on both bread and durum wheat. These investigations could be
carried out not only at the field level but also on populations coming from different years and
at different time periods during the cropping season.

Chapter 4. Screening for resistance of Tunisian, Moroccan and
Algerian wheat varieties to Zymoseptoria tritici in Northern
Tunisia
Rim Bel Hadj Chedli1,4, Sarrah Ben M’Barek2,4, Amir Suissi1, Amor Yahyaoui3,4, Salah Rezgui1 and
Hanène Chaabane5
1Laboratory of genetics and plant breeding, National Agronomic Institute of Tunisia (INAT), 43 Avenue Charles Nicolle, 1002
Tunis, University of Carthage, Tunisia. 2Regional Field Crops Research Center of Beja (CRRGC) BP 350, 9000 Beja, Tunisia. 3 Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), km 45 Carretera México-Veracruz El Batán, Texcoco. 4 CRP Wheat Septoria Precision Phenotyping Platform, Tunisia 5Laboratory of Bioagressors and Integrated protection in agriculture, National Agronomic Institute of Tunisia (INAT), 43
Avenue Charles Nicolle, 1002 Tunis, University of Carthage,Tunisia.
Hanène Chaabane : ORCID : 0000-0002-0958-3818

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
52
Objectifs et démarche
Le choix d’une variété résistante permet d’abaisser la pression parasitaire et donc la
nuisibilité. Les variétés commerciales Tunisiennes de blé tendre ont montré une certaine
résistance à la septoriose. Au contraire, au Maroc une situation inverse existait et une résistance
des variétés de blé dur contre une sensibilité du blé tendre a été observée. En Algérie, toutes
les variétés de blé tendre et de blé dur ont montré une sensibilité à cette maladie. Face à cette
situation, il est raisonnable d'examiner les conditions associées à l'expansion rapide de Z. tritici
et aux variations de la sévérité d'attaque d’un génotype a un autre en Afrique du Nord.
Cependant, la recherche des sources de résistance a été faite sur 89 variétés de blé tendre
et de blé dur d’origine Algérienne, Marocaine et Tunisienne et une série différentielle constituée
de 49 ISEPTONS durant les deux campagnes agricoles 2016-2017 et 2017-2018 dans le cadre
d’essais multi-locaux. Les essais ont été menés sous l’infection naturelle dans la région du Cap
bon (El Haouaria et Menzel Temim) là où une grande incidence de septoriose a été observée
sur le blé tendre et dans la station expérimentale Oued Béjà qui est connue comme un hot spot
pour le blé dur.
D’une part, l’évolution de la maladie a été effectuée en évaluant la sévérité (pourcentage
de surface foliaire attaquée) et la hauteur de l’attaque selon l’échelle de Saari et Prescott (00-
99) décrite par Eyal et al. (1987), et aussi en mesurant la couverture pycnidiale (PC) et la surface
nécrosée (N). D’autre part, la progression de la maladie au cours du cycle a été estimée par la
mesure de l’AUDPC (area under disease progress curve) et le rAUDPC (relative area under
disease progress curve). En plus, l’identification de differentes classes de variétés (allant de la
classe immune jusqu’à hautement sensible) a eu lieu en utilisant la méthode statistique “K-
means” classification.

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
53
Abstract
In this study, we investigated the performance of 89 wheat varieties from Morocco, Algeria and
Tunisia, screened in Tunisia for their relative resistance to STB. Field experiments were carried
out in an Augmented design, during 2016-17 and 2017-18 cropping seasons at two locations in
Northern Tunisia: Béjà (Oued Béjà station) and Cap Bon regions (Menzel Temim and El
Haouaria). All trials were conducted under natural infection. Visual disease assessments were
quantified using the percentage of leaf area covered by pycnidia (PC), necrotic area (N), area
under disease progress curve of each variety (AUDPC) and the relative area under disease
progress curve (rAUDPC). Results indicated that the majority of Tunisian, Algerian and
Moroccan durum wheat populations were susceptible to STB at both locations ranging from
susceptible to highly susceptible, with the rAUDPC, N and PC ranging from 0.5 to 0.8, 30 to
65 % and 40 to 75% respectively. On the other hand, the Moroccan bread wheat varieties were
susceptible in Cap Bon area, while the Tunisian bread wheat varieties were resistant; with levels
varying from immune to resistant classes, where rAUDPC value was 0.2, and PC and N not
exceeding 0.2 and 10% respectively, with the exception of local bread wheat known as “Farina
Arbi”.
Key words: wheat varieties, Zymoseptoria tritici, resistance, susceptibility
1. Introduction
North Africa has been the cradle of wheat production for centuries and was the bread
basket for the Romain Empire (Bachta, 2011). Nowadays, the Maghreb zone of North Africa is
still the major durum wheat producer which is the basis for their traditionnal dishes such as
couscous and pasta (Rastoin and Benabdrazik, 2014). Tunisia is among the countries with high
cereal consumption and the average per capita consumption reached 259 kg (Rastoin and Ben
Abderrazik, 2014; Hanson et al., 2016). Durum wheat (Triticum turgidum L. subsp. durum
(Desf.)) is commonly cultivated in Tunisia probably since the Roman era, while cultivation of
bread wheat (Triticum aestivum L. subsp. aestivum) was introduced by French colonists in the
early 1900s (El Falleh et al., 2015). Hence, the cultivation of bread wheat in Tunisia started
with the selection of the cultivar Florence-Aurore that covered up to 80% of the Tunisian bread
wheat acreage until 1952 then dropped to about 50% by 1959 (Ammar et al., 2011; Ben
Hamouda, 2016).

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
54
After the introduction of the semi-dwarf high yielding bread wheats varities selected from
CIMMYT nurseries known as « Mexican wheat », the bread wheat varieties such as Baroota
52, Sonora 63, Inia 66, Tobari 66, Ariana 66 and Mahon 73, selected from introduced
populations were released and covered large areas due to their high-yield performances (Saade,
1996). These varieties were soon replaced by many wheats varieties such as ‘Soltane 72’, but
also rapidly abandoned due to their succeptibility to many diseases namely yellow rust and
septoria. During the late 1980’s and early 1990’s, ‘Salammbô 80’ was the most grown bread
wheat variety and covered 70% of the bread wheat acreage followed by ‘Byrsa 87’. These two
varieties became the most popular because of their high-yield potentiel and resistance to rust
and septoria (Saade 1996). More recent release such as Utique (1996), Haidra (2004), and
Tahent (2009) had good performances and acceptable diseases resistance mainly to powdery
mildew, yellow rust, leaf rust, and septoria (Ammar et al.,2011).
Across the Magreb countries, the most prevalent foliar disease is Septoria tritici blotch
(STB) caused by the ascomycete fungus Zymoseptoria tritici (Roberge ex Desm.): Quaedvl.
and Crous anamorph and Mycosphaerella graminicola (Fuckel) J. Schröt. 1894 teleomorph.
This disease can reach epidemic levels on early planted cereals particularly when rainfall occurs
in late winter and/or spring which is common in the Mediterranean-type environments
(Chartrain et al., 2005).
In Tunisia, STB causes major yield losses on durum wheat (Ben Mohamed et al. 2000)
where most commercial durum wheat varities are highly susceptible to this pathogen while
bread wheat has been resistant to STB (Gharbi and Deghaies 1997; Gharbi et al., 2000). Under
favorable growing conditions STB disease could reduce yield by 40% (Gharbi et al., 2011;
Berraies et al., 2014). The prevailing strains seem to have developed a unique virulence towards
durum wheat (Yahyaoui et al., 2000). The opposite situation exists in Morocco, where bread
wheat is the major crop affected by STB (Mazzouz et al., 1995; Zahri et al., 2014). It has been
previously reported that isolates of Z. tritici exhibit both cultivar specificity (ability to infect
only some cultivars of either durum or bread-wheat) and host species specificity (ability to only
infect one or the other wheat (Kema et al.,1996; Kema and Van Silfhout, 1997). This contrast
between Morocco and Tunisia may explain further that specificity of Z. tritici isolates exists in
wheat (Yahyaoui et al., 2000). However, in Algeria, the STB becomes the major threat to both
durum and bread wheat on coastal and sub-littoral regions of the country (Benbelkacem et al.,
2011; Ayad et al., 2014).

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
55
Recent research provided better insight on the epidemiology of Z. tritici in the Maghreb
countries. Siah et al. (2015) reported a high level of genetic diversity within the Moroccan Z.
tritici population. The occurrence of the teleomorph stage of Z. tritici has been confirmed in
Algeria and Tunisia (Meamiche-Neddaf et al., 2017; Ben Hassine and Hamada, 2014) where
the two mating types were found at equal frequencies (Boukef et al., 2012; Ayad et al., 2014).
Thus, knowledge of the host-pathogen relationship and understanding the basis of host-
specificity and resistance in the Mediterranean area are essential for successful genetic control
of STB both on durum and bread wheat in the Maghreb region.
The objectives of this study were: (i) to evaluate bread and durum wheat varieties from
Tunisia, Algeria and Morocco for STB resistance under natural infection in Tunisia, (ii) to
evaluate the phenotypic differences in resistance of wheat varieties to STB using the area under
disease progress curve (AUDPC), the relative AUDPC (rAUDPC), pycnidial coverage (PC),
necrosis (N) in order to compare STB development between wheat species and finally (iii) to
investigate the relationships between quantitative traits.
2. Materials and Methods
2.1. Description of the study areas and experimental design
Field experiments were conducted during 2016-2017 and 2017-2018 wheat-growing
seasons, in two regions under different sub-humid zones where STB epidemics regularly occur
(Figure 1). The first experiment was carried out at the CRP Wheat Septoria Precision
Phenotyping Platform-experimental station of the CRRGC at Oued-Béjà (36°44’05’’N,
9°13’’35’’E, governorate of Béjà, northwest of Tunisia) located in the sub-humid bioclimatic
zone where the average annual rainfall ranges from 500 to 850 mm and a daily mean
temperature varies between 10 and 28°C. This area is particularly known to be a hot spot for
STB especially on durum wheat. The second experimental was carried out at a farmer field
located at Cap Bon Area (36°47’47’’N, 11°0’8’’E, governorate of Nabeul, northwest of
Tunisia) with precipitation and temperature rates varying from 400 to 600 mm and from 6 to
33 °C, respectively. This region has been recently identified as a hot spot for STB especially on
bread wheat (Bel Hadj Chedli et al., 2018).

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
56
The experiments were set up in an augmented design. During 2016-17, the trials were
sown on November 17 and 18 at Béjà and Cap Bon respectively while during the 2017-18, these
fields were sown on November 6 and 11 at Cap Bon and Béjà respectively. Each variety was
sown in paired rows of 2m with 4g of seeds per row. Standard experimental station plot was
applied (herbicide, hand weeding and fertilizer application) to ensure adequate crop
development.
2.2. Plant materials
A set of 89 wheat varieties including bread wheat from CIMMYT’s International Septoria
Observation Nursery (ISEPTON) (Annexes 2); bread and durum wheat varieties from Tunisia,
Morocco and Algeria (Table 1), were screened for their relative resistance to STB. Two
susceptible checks were used in this study: the bread wheat landrace identified as susceptible
in Cap Bon region during a survey conducted in 2015-16 cropping season (Bel Hadj Chedli et
al., 2018) and the most susceptible durum wheat ‘Karim’.
Beja governorate
36°44’05’’N, 9°13’35’’E
Nabeul governorate (Cap Bon)
36°47’47’’N, 11°0’8’’E
Figure1. Map of Tunisia showing the location of study area (Beja and Cap Bon area) during 2017-2018
cropping season. The STB experiments were set in an augmented design at both locations.
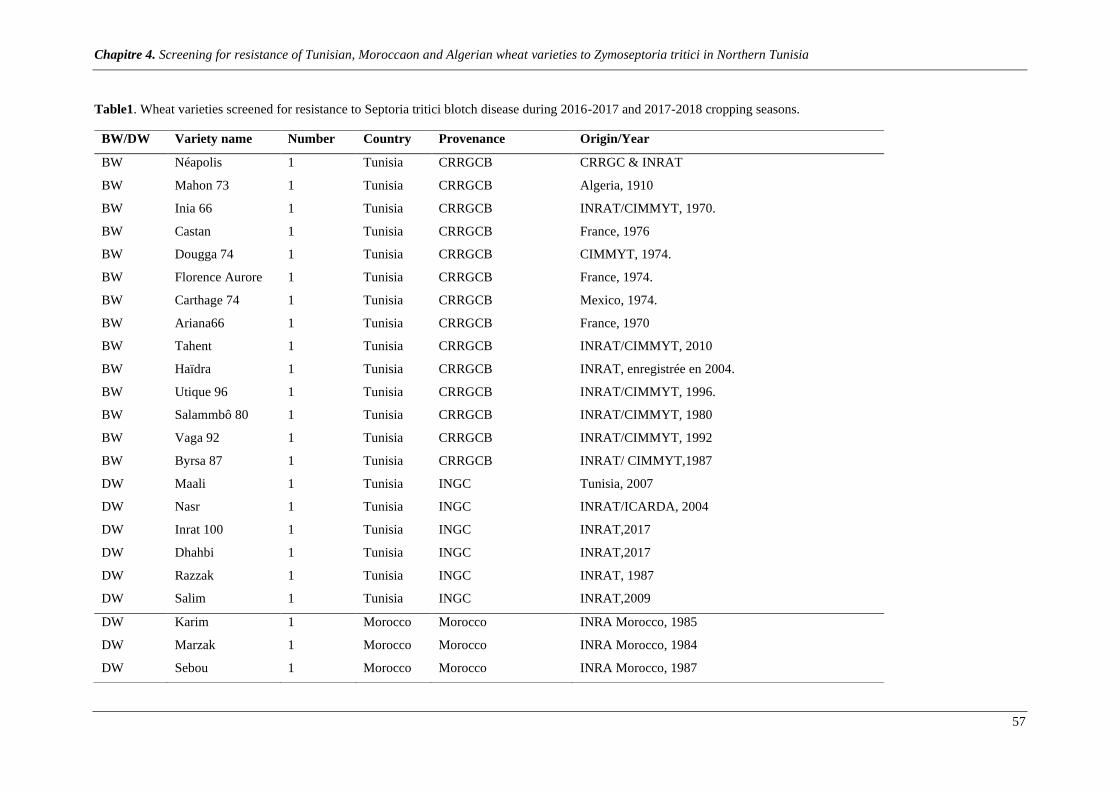
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria tritici in Northern Tunisia
57
Table1. Wheat varieties screened for resistance to Septoria tritici blotch disease during 2016-2017 and 2017-2018 cropping seasons.
BW/DW Variety name Number Country Provenance Origin/Year
BW Néapolis 1 Tunisia CRRGCB CRRGC & INRAT
BW Mahon 73 1 Tunisia CRRGCB Algeria, 1910
BW Inia 66 1 Tunisia CRRGCB INRAT/CIMMYT, 1970.
BW Castan 1 Tunisia CRRGCB France, 1976
BW Dougga 74 1 Tunisia CRRGCB CIMMYT, 1974.
BW Florence Aurore 1 Tunisia CRRGCB France, 1974.
BW Carthage 74 1 Tunisia CRRGCB Mexico, 1974.
BW Ariana66 1 Tunisia CRRGCB France, 1970
BW Tahent 1 Tunisia CRRGCB INRAT/CIMMYT, 2010
BW Haïdra 1 Tunisia CRRGCB INRAT, enregistrée en 2004.
BW Utique 96 1 Tunisia CRRGCB INRAT/CIMMYT, 1996.
BW Salammbô 80 1 Tunisia CRRGCB INRAT/CIMMYT, 1980
BW Vaga 92 1 Tunisia CRRGCB INRAT/CIMMYT, 1992
BW Byrsa 87 1 Tunisia CRRGCB INRAT/ CIMMYT,1987
DW Maali 1 Tunisia INGC Tunisia, 2007
DW Nasr 1 Tunisia INGC INRAT/ICARDA, 2004
DW Inrat 100 1 Tunisia INGC INRAT,2017
DW Dhahbi 1 Tunisia INGC INRAT,2017
DW Razzak 1 Tunisia INGC INRAT, 1987
DW Salim 1 Tunisia INGC INRAT,2009
DW Karim 1 Morocco Morocco INRA Morocco, 1985
DW Marzak 1 Morocco Morocco INRA Morocco, 1984
DW Sebou 1 Morocco Morocco INRA Morocco, 1987
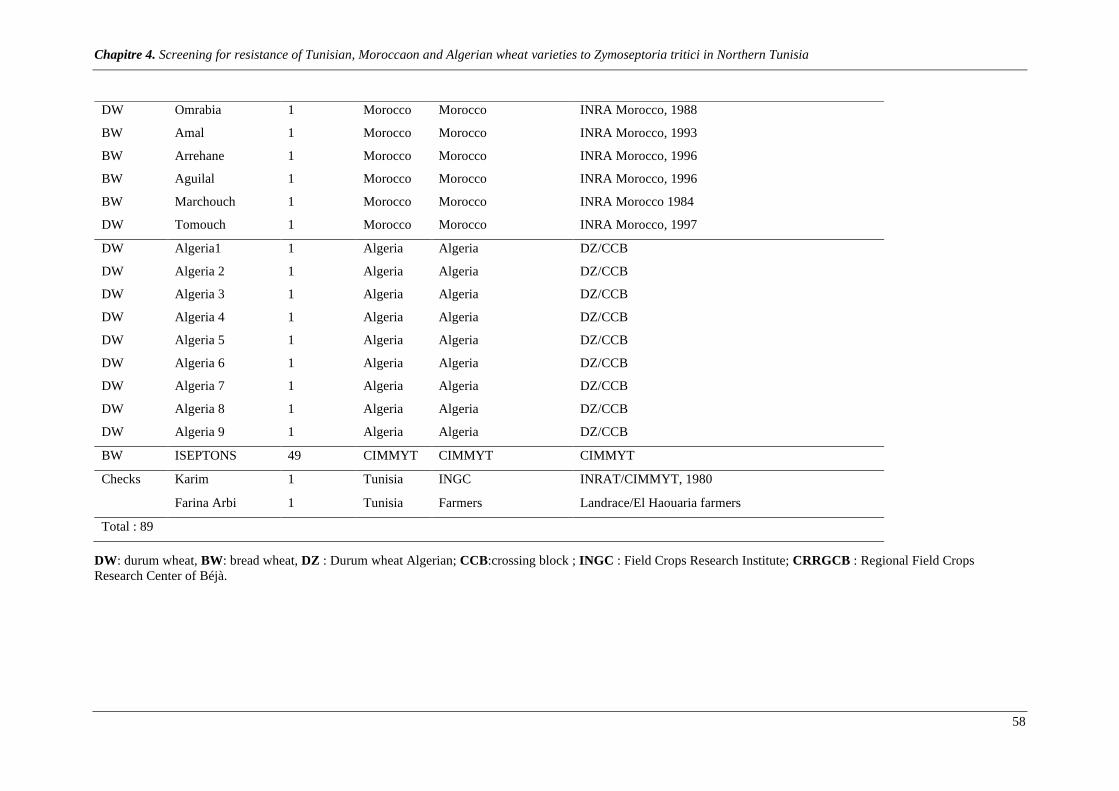
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria tritici in Northern Tunisia
58
DW: durum wheat, BW: bread wheat, DZ : Durum wheat Algerian; CCB:crossing block ; INGC : Field Crops Research Institute; CRRGCB : Regional Field Crops
Research Center of Béjà.
DW Omrabia 1 Morocco Morocco INRA Morocco, 1988
BW Amal 1 Morocco Morocco INRA Morocco, 1993
BW Arrehane 1 Morocco Morocco INRA Morocco, 1996
BW Aguilal 1 Morocco Morocco INRA Morocco, 1996
BW Marchouch 1 Morocco Morocco INRA Morocco 1984
DW Tomouch 1 Morocco Morocco INRA Morocco, 1997
DW Algeria1 1 Algeria Algeria DZ/CCB
DW Algeria 2 1 Algeria Algeria DZ/CCB
DW Algeria 3 1 Algeria Algeria DZ/CCB
DW Algeria 4 1 Algeria Algeria DZ/CCB
DW Algeria 5 1 Algeria Algeria DZ/CCB
DW Algeria 6 1 Algeria Algeria DZ/CCB
DW Algeria 7 1 Algeria Algeria DZ/CCB
DW Algeria 8 1 Algeria Algeria DZ/CCB
DW Algeria 9 1 Algeria Algeria DZ/CCB
BW ISEPTONS 49 CIMMYT CIMMYT CIMMYT
Checks
Karim 1 Tunisia INGC INRAT/CIMMYT, 1980
Farina Arbi 1 Tunisia Farmers Landrace/El Haouaria farmers
Total : 89

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
59
2.3. Evaluation of disease severity and area under disease progress curve
Visual disease assessment based on the leaf area covered with pycnidia (PC) and the
necrotic area (N) were estimated as a percentage of the uppermost-infected leaves either on
Flag leaf or Flag leaf-1 at the end of the growth stage, i.e GS70. In addition, the symptoms and
lesion development over the assessment period were summarized by the area under disease
progress curve (AUDPC) that allows the identification of different classes of resistance. Disease
severity was scored for each plot using the double-digit scale (Saari and Prescott, 1975). The
first digit (D1) indicates disease progress on the infected plants, and the second digit (D2) refers
to severity of infection. Three consecutive evaluations were made at 10 days interval, at GS51,
GS59 and GS65 respectively according to Zadok’s scales (Zadoks et al., 1974). The AUDPC
and the relative area under disease progress curve (rAUDPC) were subsequently calculated
according to Simko and Piepho (2012) formula:
𝐴𝑈𝐷𝑃𝐶 = ∑𝑦𝑖 + 𝑦𝑖+1
2
𝑛−1
𝑖=1
× (𝑡𝑖+1 − 𝑡𝑖)
Where:
Yi: STB severity at time ti,
t(i+1)-ti = time interval (days) between two disease scores,
n = number of times when STB was recorded.
𝑟𝐴𝑈𝐷𝑃𝐶 =𝐴𝑈𝐷𝑃𝐶 (𝑔é𝑛𝑜𝑡𝑦𝑝𝑒)
𝐴𝑈𝐷𝑃𝐶 (𝐾𝑎𝑟𝑖𝑚)
Where: Karim is the most susceptible variety used as a check.
2.4. Statistical analysis
All the observations in the experimental field and dependent variables were subjected to
analysis of variance (ANOVA) using ‘aov’ function from R package ‘daewr’ (Lawson 2016)
implemented in R software v3.4.2 (R Core Team 2017) and least-Squares Means using R
package ‘lsmeans’ (V. Lenth 2016). Distances between wheat varieties using hierarchical
clustering method and correlations coefficients between characters were calculated for all traits
analyzed in the study. A weighted clustering algorithm K-mean (K =6) (Duda and Hart, 1973)
was performed using JMP®11.0 in order to group the different varieties in classes.
Subsequently, Principal component analysis (PCA) was performed using the JMP®11.0

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
60
statistical software (SAS Institute Inc., Cary, NC, USA) with component analysis procedure
(SAS Institute, 2014).
3. Results
3.1. Meteorological conditions during the crop cycle
Meteorological data (temperature and rainfall) over different climatic regions and during
the two cropping seasons were recorded. The annual average rainfall varied from 500 to 800
mm and the temperature ranged between 6 and 33°C in Cap Bon and Béjà regions. The variation
of temperature and rainfall from November to May during the two cropping seasons of 2017
and 2018 is shown in (Annexe 3).
Although epidemics of STB are associated with favorable weather conditions (frequent
rains and moderate temperature) that are encountered in these two regions, the different
responses of wheat varieties across regions were rather associated to the specialization of the
pathogen to the one or the other wheat species.
3.2. Genotype by region interaction
Results of this study revealed a good STB development at the two locations. Therefore, a
significant effect (P<0.0001 and P<0.01, Table 2) of variety and region for pycnidial coverage
(PC), necrotic area (N) and area under the disease progress curves (AUDPC) was observed.
The variety-by-region interaction term in the ANOVA analysis was significant at P0.05, for N,
PC and the rAUDPC indicating different responses of wheat varieties to Z. tritici across regions.
Therefore, any significant effect was observed with year as source of variation and also with
the interaction variety-region-year.
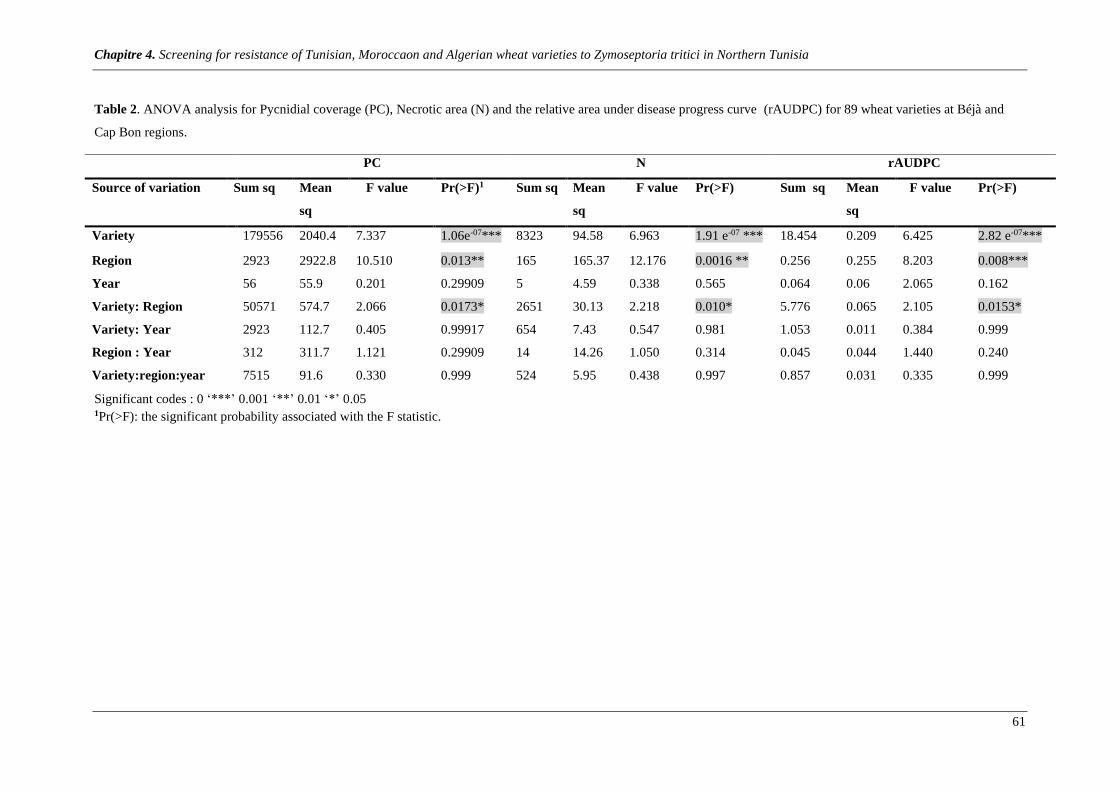
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria tritici in Northern Tunisia
61
Table 2. ANOVA analysis for Pycnidial coverage (PC), Necrotic area (N) and the relative area under disease progress curve (rAUDPC) for 89 wheat varieties at Béjà and
Cap Bon regions.
Significant codes : 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 1Pr(>F): the significant probability associated with the F statistic.
PC N rAUDPC
Source of variation Sum sq Mean
sq
F value Pr(>F)1 Sum sq Mean
sq
F value Pr(>F) Sum sq Mean
sq
F value Pr(>F)
Variety 179556 2040.4 7.337 1.06e-07*** 8323 94.58 6.963 1.91 e-07 *** 18.454 0.209 6.425 2.82 e-07***
Region 2923 2922.8 10.510 0.013** 165 165.37 12.176 0.0016 ** 0.256 0.255 8.203 0.008***
Year 56 55.9 0.201 0.29909 5 4.59 0.338 0.565 0.064 0.06 2.065 0.162
Variety: Region 50571 574.7 2.066 0.0173* 2651 30.13 2.218 0.010* 5.776 0.065 2.105 0.0153*
Variety: Year 2923 112.7 0.405 0.99917 654 7.43 0.547 0.981 1.053 0.011 0.384 0.999
Region : Year 312 311.7 1.121 0.29909 14 14.26 1.050 0.314 0.045 0.044 1.440 0.240
Variety:region:year 7515 91.6 0.330 0.999 524 5.95 0.438 0.997 0.857 0.031 0.335 0.999
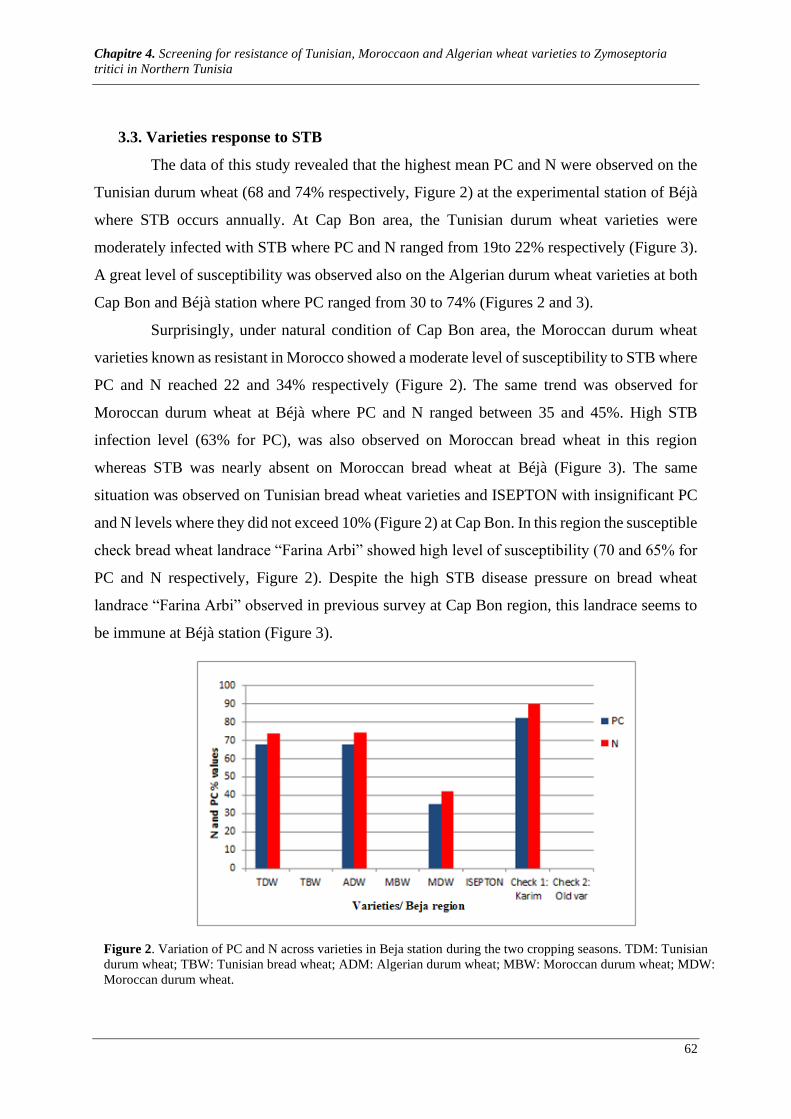
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
62
3.3. Varieties response to STB
The data of this study revealed that the highest mean PC and N were observed on the
Tunisian durum wheat (68 and 74% respectively, Figure 2) at the experimental station of Béjà
where STB occurs annually. At Cap Bon area, the Tunisian durum wheat varieties were
moderately infected with STB where PC and N ranged from 19to 22% respectively (Figure 3).
A great level of susceptibility was observed also on the Algerian durum wheat varieties at both
Cap Bon and Béjà station where PC ranged from 30 to 74% (Figures 2 and 3).
Surprisingly, under natural condition of Cap Bon area, the Moroccan durum wheat
varieties known as resistant in Morocco showed a moderate level of susceptibility to STB where
PC and N reached 22 and 34% respectively (Figure 2). The same trend was observed for
Moroccan durum wheat at Béjà where PC and N ranged between 35 and 45%. High STB
infection level (63% for PC), was also observed on Moroccan bread wheat in this region
whereas STB was nearly absent on Moroccan bread wheat at Béjà (Figure 3). The same
situation was observed on Tunisian bread wheat varieties and ISEPTON with insignificant PC
and N levels where they did not exceed 10% (Figure 2) at Cap Bon. In this region the susceptible
check bread wheat landrace “Farina Arbi” showed high level of susceptibility (70 and 65% for
PC and N respectively, Figure 2). Despite the high STB disease pressure on bread wheat
landrace “Farina Arbi” observed in previous survey at Cap Bon region, this landrace seems to
be immune at Béjà station (Figure 3).
Figure 2. Variation of PC and N across varieties in Beja station during the two cropping seasons. TDM: Tunisian
durum wheat; TBW: Tunisian bread wheat; ADM: Algerian durum wheat; MBW: Moroccan durum wheat; MDW:
Moroccan durum wheat.
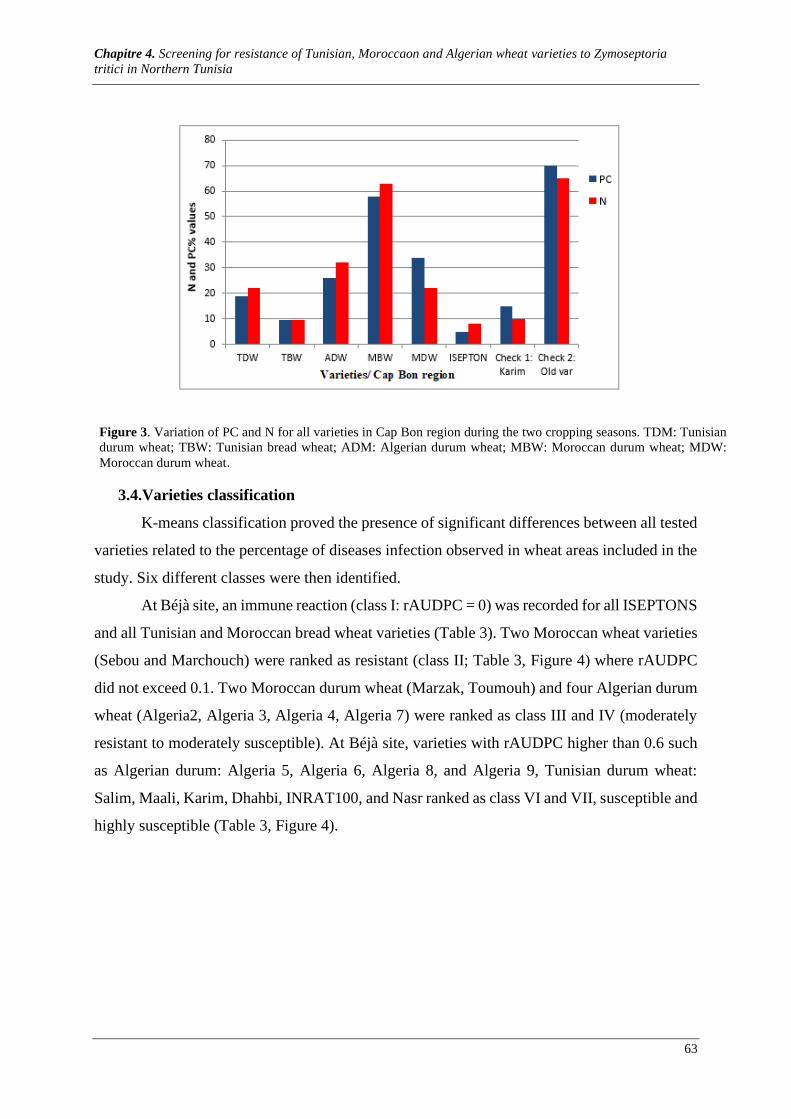
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria
tritici in Northern Tunisia
63
3.4.Varieties classification
K-means classification proved the presence of significant differences between all tested
varieties related to the percentage of diseases infection observed in wheat areas included in the
study. Six different classes were then identified.
At Béjà site, an immune reaction (class I: rAUDPC = 0) was recorded for all ISEPTONS
and all Tunisian and Moroccan bread wheat varieties (Table 3). Two Moroccan wheat varieties
(Sebou and Marchouch) were ranked as resistant (class II; Table 3, Figure 4) where rAUDPC
did not exceed 0.1. Two Moroccan durum wheat (Marzak, Toumouh) and four Algerian durum
wheat (Algeria2, Algeria 3, Algeria 4, Algeria 7) were ranked as class III and IV (moderately
resistant to moderately susceptible). At Béjà site, varieties with rAUDPC higher than 0.6 such
as Algerian durum: Algeria 5, Algeria 6, Algeria 8, and Algeria 9, Tunisian durum wheat:
Salim, Maali, Karim, Dhahbi, INRAT100, and Nasr ranked as class VI and VII, susceptible and
highly susceptible (Table 3, Figure 4).
Figure 3. Variation of PC and N for all varieties in Cap Bon region during the two cropping seasons. TDM: Tunisian
durum wheat; TBW: Tunisian bread wheat; ADM: Algerian durum wheat; MBW: Moroccan durum wheat; MDW:
Moroccan durum wheat.
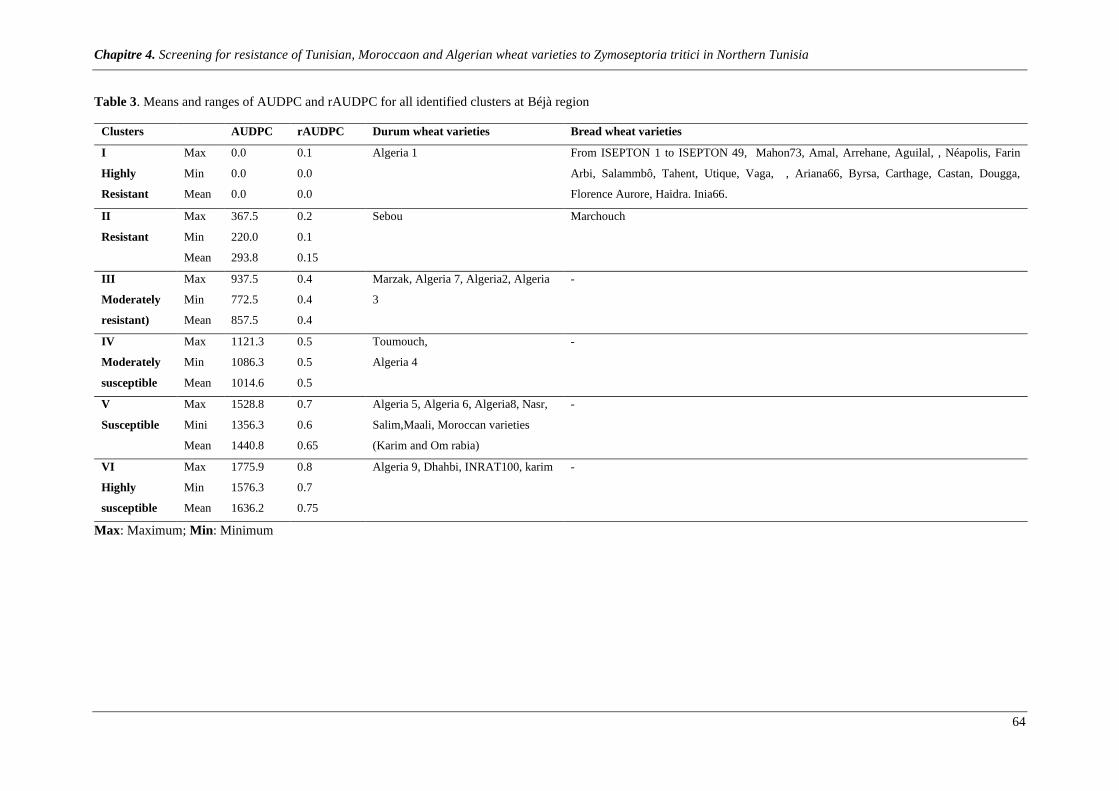
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat varieties to Zymoseptoria tritici in Northern Tunisia
64
Table 3. Means and ranges of AUDPC and rAUDPC for all identified clusters at Béjà region
Max: Maximum; Min: Minimum
Clusters AUDPC rAUDPC Durum wheat varieties Bread wheat varieties
I
Highly
Resistant
Max 0.0 0.1 Algeria 1 From ISEPTON 1 to ISEPTON 49, Mahon73, Amal, Arrehane, Aguilal, , Néapolis, Farin
Arbi, Salammbô, Tahent, Utique, Vaga, , Ariana66, Byrsa, Carthage, Castan, Dougga,
Florence Aurore, Haidra. Inia66.
Min 0.0 0.0
Mean 0.0 0.0
II
Resistant
Max 367.5 0.2 Sebou Marchouch
Min 220.0 0.1
Mean 293.8 0.15
III
Moderately
resistant)
Max 937.5 0.4 Marzak, Algeria 7, Algeria2, Algeria
3
-
Min 772.5 0.4
Mean 857.5 0.4
IV
Moderately
susceptible
Max 1121.3 0.5 Toumouch,
Algeria 4
-
Min 1086.3 0.5
Mean 1014.6 0.5
V
Susceptible
Max 1528.8 0.7 Algeria 5, Algeria 6, Algeria8, Nasr,
Salim,Maali, Moroccan varieties
(Karim and Om rabia)
-
Mini 1356.3 0.6
Mean 1440.8 0.65
VI
Highly
susceptible
Max 1775.9 0.8 Algeria 9, Dhahbi, INRAT100, karim -
Min 1576.3 0.7
Mean 1636.2 0.75
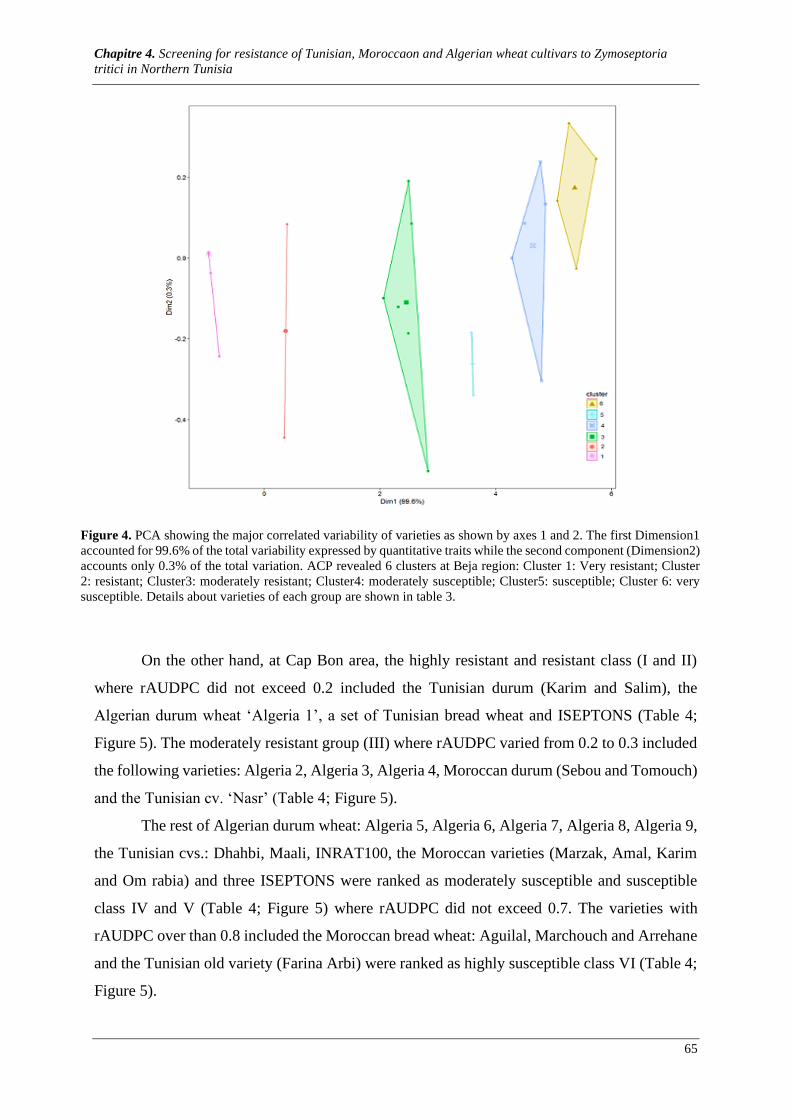
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria
tritici in Northern Tunisia
65
On the other hand, at Cap Bon area, the highly resistant and resistant class (I and II)
where rAUDPC did not exceed 0.2 included the Tunisian durum (Karim and Salim), the
Algerian durum wheat ‘Algeria 1’, a set of Tunisian bread wheat and ISEPTONS (Table 4;
Figure 5). The moderately resistant group (III) where rAUDPC varied from 0.2 to 0.3 included
the following varieties: Algeria 2, Algeria 3, Algeria 4, Moroccan durum (Sebou and Tomouch)
and the Tunisian cv. ‘Nasr’ (Table 4; Figure 5).
The rest of Algerian durum wheat: Algeria 5, Algeria 6, Algeria 7, Algeria 8, Algeria 9,
the Tunisian cvs.: Dhahbi, Maali, INRAT100, the Moroccan varieties (Marzak, Amal, Karim
and Om rabia) and three ISEPTONS were ranked as moderately susceptible and susceptible
class IV and V (Table 4; Figure 5) where rAUDPC did not exceed 0.7. The varieties with
rAUDPC over than 0.8 included the Moroccan bread wheat: Aguilal, Marchouch and Arrehane
and the Tunisian old variety (Farina Arbi) were ranked as highly susceptible class VI (Table 4;
Figure 5).
Figure 4. PCA showing the major correlated variability of varieties as shown by axes 1 and 2. The first Dimension1
accounted for 99.6% of the total variability expressed by quantitative traits while the second component (Dimension2)
accounts only 0.3% of the total variation. ACP revealed 6 clusters at Beja region: Cluster 1: Very resistant; Cluster
2: resistant; Cluster3: moderately resistant; Cluster4: moderately susceptible; Cluster5: susceptible; Cluster 6: very
susceptible. Details about varieties of each group are shown in table 3.
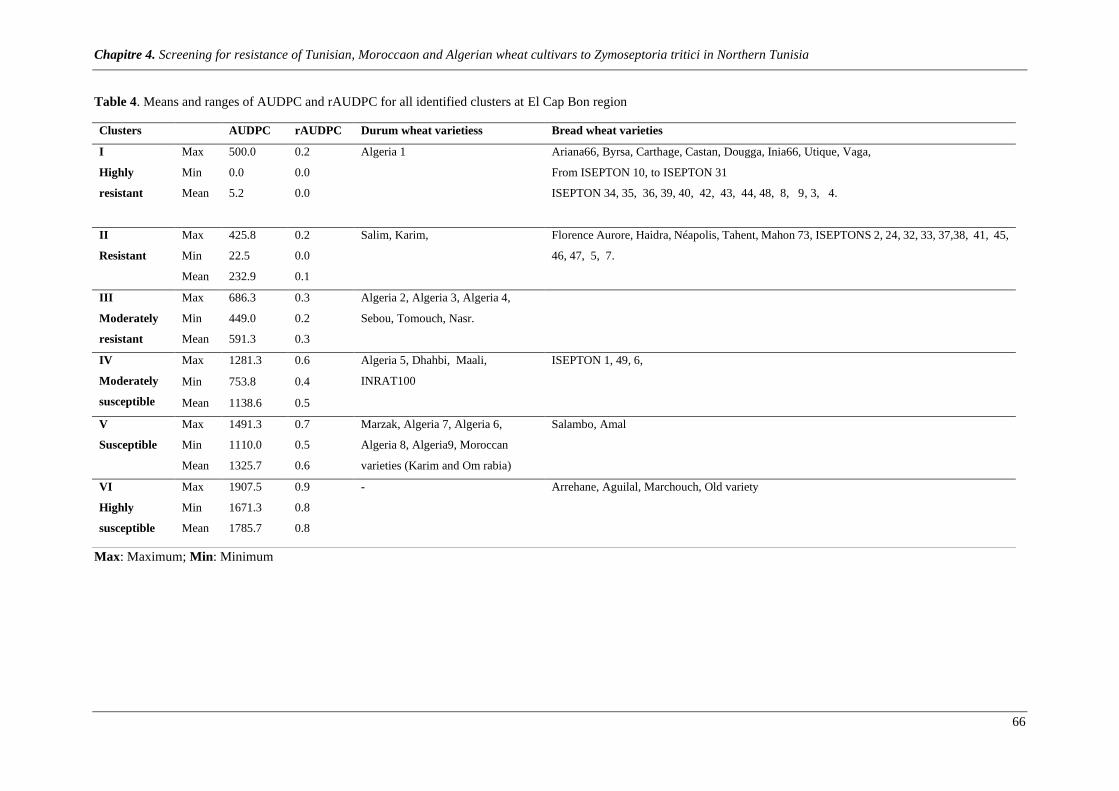
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria tritici in Northern Tunisia
66
Table 4. Means and ranges of AUDPC and rAUDPC for all identified clusters at El Cap Bon region
Max: Maximum; Min: Minimum
Clusters AUDPC rAUDPC Durum wheat varietiess Bread wheat varieties
I
Highly
resistant
Max 500.0 0.2 Algeria 1 Ariana66, Byrsa, Carthage, Castan, Dougga, Inia66, Utique, Vaga,
From ISEPTON 10, to ISEPTON 31
ISEPTON 34, 35, 36, 39, 40, 42, 43, 44, 48, 8, 9, 3, 4.
Min 0.0 0.0
Mean 5.2 0.0
II
Resistant
Max 425.8 0.2 Salim, Karim, Florence Aurore, Haidra, Néapolis, Tahent, Mahon 73, ISEPTONS 2, 24, 32, 33, 37,38, 41, 45,
46, 47, 5, 7. Min 22.5 0.0
Mean 232.9 0.1
III
Moderately
resistant
Max 686.3 0.3 Algeria 2, Algeria 3, Algeria 4,
Sebou, Tomouch, Nasr.
Min 449.0 0.2
Mean 591.3 0.3
IV
Moderately
susceptible
Max 1281.3 0.6 Algeria 5, Dhahbi, Maali,
INRAT100
ISEPTON 1, 49, 6,
Min 753.8 0.4
Mean 1138.6 0.5
V
Susceptible
Max 1491.3 0.7 Marzak, Algeria 7, Algeria 6,
Algeria 8, Algeria9, Moroccan
varieties (Karim and Om rabia)
Salambo, Amal
Min 1110.0 0.5
Mean 1325.7 0.6
VI
Highly
susceptible
Max 1907.5 0.9 - Arrehane, Aguilal, Marchouch, Old variety
Min 1671.3 0.8
Mean 1785.7 0.8
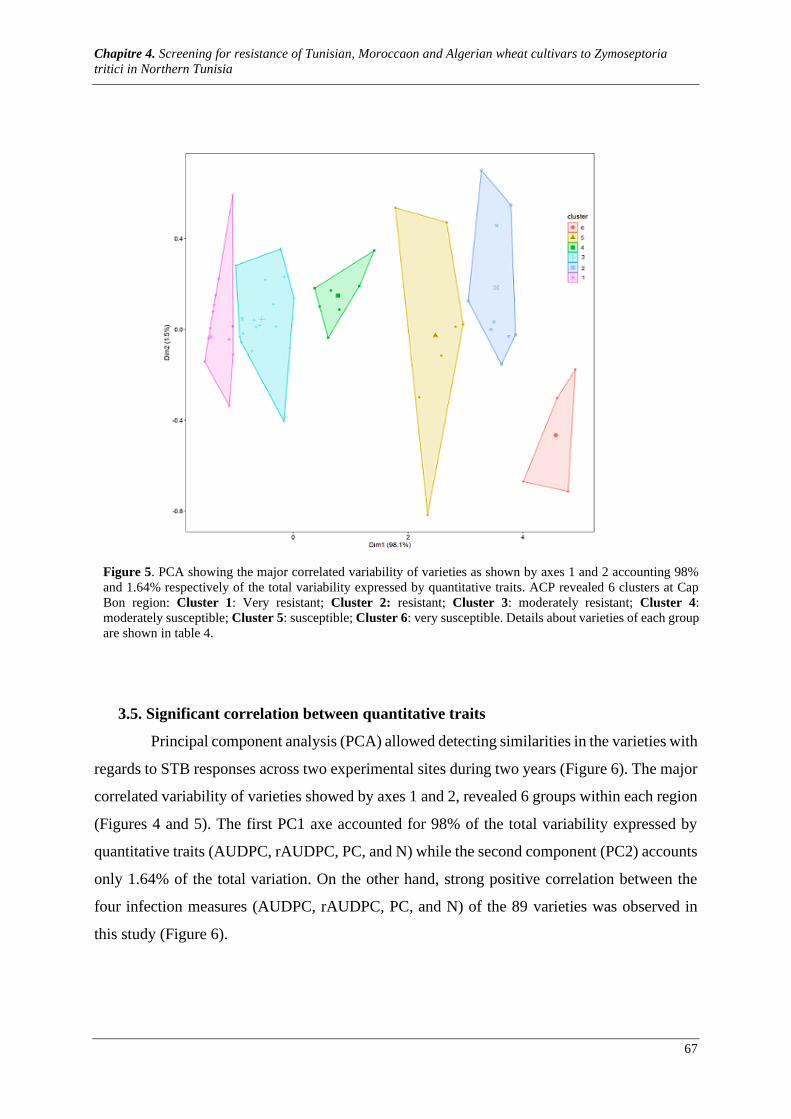
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria
tritici in Northern Tunisia
67
3.5. Significant correlation between quantitative traits
Principal component analysis (PCA) allowed detecting similarities in the varieties with
regards to STB responses across two experimental sites during two years (Figure 6). The major
correlated variability of varieties showed by axes 1 and 2, revealed 6 groups within each region
(Figures 4 and 5). The first PC1 axe accounted for 98% of the total variability expressed by
quantitative traits (AUDPC, rAUDPC, PC, and N) while the second component (PC2) accounts
only 1.64% of the total variation. On the other hand, strong positive correlation between the
four infection measures (AUDPC, rAUDPC, PC, and N) of the 89 varieties was observed in
this study (Figure 6).
Figure 5. PCA showing the major correlated variability of varieties as shown by axes 1 and 2 accounting 98%
and 1.64% respectively of the total variability expressed by quantitative traits. ACP revealed 6 clusters at Cap
Bon region: Cluster 1: Very resistant; Cluster 2: resistant; Cluster 3: moderately resistant; Cluster 4:
moderately susceptible; Cluster 5: susceptible; Cluster 6: very susceptible. Details about varieties of each group
are shown in table 4.
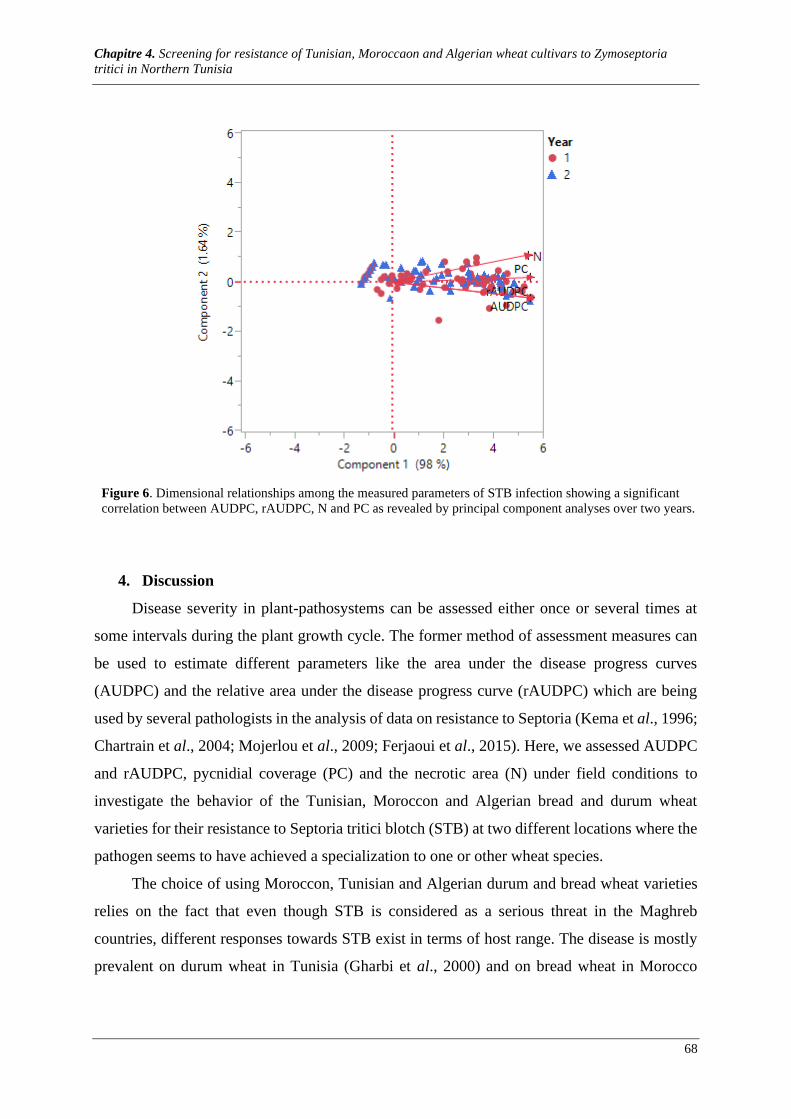
Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria
tritici in Northern Tunisia
68
4. Discussion
Disease severity in plant-pathosystems can be assessed either once or several times at
some intervals during the plant growth cycle. The former method of assessment measures can
be used to estimate different parameters like the area under the disease progress curves
(AUDPC) and the relative area under the disease progress curve (rAUDPC) which are being
used by several pathologists in the analysis of data on resistance to Septoria (Kema et al., 1996;
Chartrain et al., 2004; Mojerlou et al., 2009; Ferjaoui et al., 2015). Here, we assessed AUDPC
and rAUDPC, pycnidial coverage (PC) and the necrotic area (N) under field conditions to
investigate the behavior of the Tunisian, Moroccon and Algerian bread and durum wheat
varieties for their resistance to Septoria tritici blotch (STB) at two different locations where the
pathogen seems to have achieved a specialization to one or other wheat species.
The choice of using Moroccon, Tunisian and Algerian durum and bread wheat varieties
relies on the fact that even though STB is considered as a serious threat in the Maghreb
countries, different responses towards STB exist in terms of host range. The disease is mostly
prevalent on durum wheat in Tunisia (Gharbi et al., 2000) and on bread wheat in Morocco
Figure 6. Dimensional relationships among the measured parameters of STB infection showing a significant
correlation between AUDPC, rAUDPC, N and PC as revealed by principal component analyses over two years.

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria
tritici in Northern Tunisia
69
(Mazzouz et al., 1995) while it is a major threat for both durum and bread wheat in Algeria
(Ayad et al., 2014).
Results of this study showed a good STB development at the two locations and analysis
of variance revealed the presence of important and significant variability of the experimental
materials used to evaluate the susceptibility and resistance of wheat varieties against STB. On
the other hand, PCA results showed that the four quantitative variables (N, PC, rAUDPC and
AUDPC) contributed in the total with 99 % of variance, and high positive correlation was
recorded between these measured parameters. These results are in agreement with Odilbekov
et al. (2018) finding which reported that, STB measured parameters affected by the disease
increased upon disease progression. Similar studies conducted by Karisto et al. (2017) showed
a positive correlation between quantitative variables (AUDPC and PC) and STB infection.
More important, a significant interaction between Variety: Region was observed in this
study suggesting a possible physiological specialization of the pathogen across studied regions
(Boughalleb et Harrabi, 1997). STB was nearly absent on ISEPTONS and the Tunisian
commercial bread wheat at Béjà where they were ranked as immune, resistant and very resistant
groups (Class I, II and III; respectively) which confirms previous conclusions about the great
level of resistance of bread wheat in Tunisia and the adaptation of Z. tritici isolates to durum
wheat in Tunisia (Gharbi et al., 2000; Fakhfakh et al., 2011). Interestingly, STB was also absent
on Moroccan bread wheat varieties at Béjà showing again that Béjà is a clear hot spot region
for STB on durum wheat.
In Cap Bon region, STB was present on Tunisian bread wheat but still insignificant
compared to the bread wheat landrace known as “Farina Arbi” which was considered as a
susceptible check in our study. In this context, Holloway (2014) and McDonald and Mundt
(2016) reported that when a very susceptible variety becomes widely grown, STB will be more
severe in the next season on the susceptible wheat. However, insignificant infection was
reported on Tunisian durum wheat Karim, Salim and Nasr which were ranked as resistant class
in this region. This could be mainly associated with the limited cultivated areas of durum wheat
which represent only 20 % of the total cereal growing areas at El Haouaria region (Cap Bon
area) compared to the old variety ‘Farina Arbi’ that cover more than 60%.
Surprisingly, STB was strongly present at Cap Bon region on Moroccan bread and durum
wheat varieties and at Béjà only on Moroccan durum wheat with the highest mean of PC, N and
rAUDPC and they were ranked as very susceptible class VII. This data contrasted with the
finding of Zahri and collaborators (2014) that highlighted the resistance of Moroccan durum

Chapitre 4. Screening for resistance of Tunisian, Moroccaon and Algerian wheat cultivars to Zymoseptoria
tritici in Northern Tunisia
70
wheat and the susceptibility of bread wheat to STB (Mazouz et al., 1995; Jilbene, 1996) when
cultivated in Morocco. This could be related to the great adaptation of Z. tritici population to
wheat species in each geographic area (Aouini, 2018).
5. Conclusion
As previously reported, STB populations from Cap Bon region seem to be mostly specific
to bread wheat genotypes (Bel Hadj Chedli et al., 2018) whereas STB population from Béjà
seems to be more adapted to durum wheat varieties where it occurs annually (Gharbi et al.,
2000). At the country level, this opposite situation between Morocco and Tunisia could be
explained by the existence of host-species specificity in Septoria (Kema et al. 1996; Kema et
al., 2018). The resistance bread wheat varieties identified in this study may possess different
resistance genes that can be used in developing cultivars with and durable resistance to Septoria
diseases (Aouini, 2018; Medini et al., 2014) and may prove useful in breeding efforts to
improve STB resistance in wheat (Zhang et al., 2001).
Future research on the genetic diversity and population structure of durum and bread
wheat adapted to Z. tritici in Tunisia is underway. Finally, the analysis of population dynamics
of Z. tritici with respect to diversity and frequency distribution of the resistance sources is
essential to guide decisions on developing strategies for durable resistance.

Chapter 5. Effect of Host-wheat species on Genetic differentiation
of ‘Zymoseptoria tritici’ at single field in Northern Tunisia

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
71
Objectifs et démarche
La diversité génétique des populations de Z. tritici a été bien documentée. Toutefois,
aucune information n’a été publiée en ce qui concerne la variation de la diversité génétique et
de la structure des populations en fonction de l’espèce et de la variété choisie.
Dans ce chapitre, une première approche moléculaire complémentaire à l’approche
phénotypique adoptée précédemment (chapitre 4) a été appliquée. En partant de l’hypothèse
que les populations de Z. tritici collectées de différentes espèces de blé soient génétiquement
distinctes, une caractérisation moléculaire de 65 isolats mono-pycnidiaux issus de 22 variétés
de blé tendre, blé dur et triticale a été suivie.
L’expérience a été conduite durant la saison agricole 2016-2017 en « Augmented
design » en plein champ et sous l’infection naturelle dans la région du Cap Bon. Un
échantillonnage aléatoire a eu lieu et uniquement trois isolats ont été retenus de chaque
génotype. L’extraction d’ADN a été réalisée moyennant le Kit ‘Sbeadex® mini plant kit’ alors
que le génotypage a été assuré par le Kit ‘Type-it microsatellite kit (Qiagen)’ moyennant 12
marqueurs microsatellites. La présence des deux mating types et l’occurrence d’un cycle sexuel
régulier de Z. tritici ont été aussi vérifiées.
Dans une deuxième étape, une approche bio-informatique a été adoptée pour déterminer
la diversité génétique et la stucture des populations de Z. tritici au sein de l’espèce Triticum et
aussi pour la construction de l’arbre phylogénétique par le biais de plusieurs logiciels, à savoir
Genalex, Structure, DARwin 6…

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
72
Abstract
Zymoseptoria tritici is currently one of the most devastating fungal diseases of durum
wheat in Tunisia. Up to date, few studies have compared the genetic diversity of Z. tritici
populations sampled from different wheat-host species. In this context, 65 isolates were
collected during 2016-2017 cropping season, from naturally infected wheat species: bread
wheat, durum wheat and triticale grown under rainfed conditions and naturally infected at at
the Cap Bon Area located North western Tunisia. The genotyping of the 65 Z. tritici isolates
using 12 microsatellite markers revealed significant levels of genetic diversity in the total
population and within species and cultivars populations. Results indicate an equal Nei’s gene
diversity (0.52), unbiased gene diversity (0.58) and allele richness (4.43) within Z. tritici-durum
and bread wheat populations. Therefore, weak population differentiation (0.13) between species
population explained by high levels of gene flow (3.26) and a lack of a genetic structuration
were then concluded. Furthermore, all studied populations displayed an unequal mating type
distribution with a predominance of Mat 1-2. Our data did not show a significant interaction
between the mating type distribution, the genetic diversity, the population structure of the Z.
tritici – host species specialization. Thus, a proper and an accurate Z. tritici population sampling
is required to better capture empirical genetic diversity between the well adapted Z. tritici
isolates to their corresponding hosts.
Key words: wheat species, Zymoseptoria tritici, genetic diversity and structure, Tunisia
1. Introduction
Zymoseptoria tritici (Z. tritici) the causal agent of Septoria tritici blotch (STB) has
become an imminent threat in several wheat-growing areas causing significant economic
impact worldwide (McDonald, 2016, Fones and Gurr, 2015) and has been reported as a severe
disease in many countries such as in : North and South America, Europe, the Central and West
Asia and North Africa (Linde et al., 2002; Zhan et al., 2003; Kabbage et al., 2008; Singh et al.,
2016). STB could reduce wheat yields by 30 to 50% under inducive conditions, increasing thus
fungicides use that costs global expenditure of hundreds of millions of dollars each year
(Torriani et al., 2015). This fungus has a heterothallic bipolar mating system with two mating
type alleles, mat1-1 and mat1-2 (Waalwijk et al. 2002), and actively sporulates through asexual
and sexual fructifications that release splash-borne pycnidisopores and air-borne ascospores,
respectively (Ponomarenko et al., 2011), both contributing to epidemics. The ascospores
constitute the primary inoculum released from wheat debris and can be formed year-round

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
73
while asexual pycnidiospores are splash-dispersed over short distances and act as the main
secondary inoculum and hence ensure the disease progress during the growing season (Eyal et
al., 1987; Kema et al., 1996; Eyal 1999; Hunter et al., 1999; McDonald and Linde, 2002;
Ponomarenko et al., 2011). This pathogen can accomplish up to six cycles on the wheat crop
(Fones and Gurr, 2015). Due to the Z. tritici's mixed reproductive system, Z. tritici populations
are extremely diverse (Linde et al., 2002; Hartmann et al., 2018) where the sexual reproduction
plays a key role in shaping the genetic structure (Chen and McDonald 1996; Linde et al., 2002;
Zhan et al., 2003). Hence, knowledge of the genetic diversity and the structure of Z. tritici
populations is essential to predict the evolutionary potential of the pathogen which could better
direct the host resistance deployment and the fungicide management strategies (McDonald and
Linde, 2002).
Previous studies reported that the Middle East was probably the origin of Z. tritici, and
the domestication of wheat-Z. tritici happened ~11.000 years ago (Stukenbrock et al., 2007).
This pathogen can infect durum wheat (Triticum turgidum L. subsp. durum (Desf.), bread wheat
(Triticum aestivum L. subsp. aestivum) and triticale (Triticosecale spp) and can also infect other
grass species (Ponomarenko et al., 2011; Stukenbrock et al., 2011). The long co-evolution with
wheat has resulted into a highly specialized pathogen of wheat that is difficult to control (Poppe
et al., 2014). The physiological specialization of Z. tritici on bread wheat and durum wheat was
for a long time open to various speculations especially in some regions such as the Maghreb
region where STB is a major threat (Medini and Hamza, 2008).
In Algeria, STB was reported on the entire country cereal zones on both cultivated wheat
species durum wheat and bread wheat (Harrat and Bouznad, 2018). In Morocco, bread wheat
is the major crop affected by Septoria while durum wheat seems to be more resistant to this
disease (Jilbène et al., 1995; Mazzouz et al., 1995) which might suggest a great specialization
of the pathogen (Kema et al., 1996). The inverse situation exists in Tunisia where severe
incidence occurs annually and mainly on durum wheat (Fakhfakh et al., 2011) which could be
related to the proportion of cultivated durum wheat area as compared to that of bread wheat in
each country (Medini and Hamza, 2008). Recently, STB has intensified in some bread wheat-
growing area of Northern Tunisia especially at “El Haouaria” region, where severe epidemic
occurs annually mainly because of the extensive monoculture of the susceptible wheat “Farina
Arbi” in that area (Bel Hadj Chedli et al., 2018).
Genetic variation and structure of Z. tritici populations have been intensively investigated
in recent years at a large geographic scales using several type of markers (Razavi and Hughes

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
74
2004; Abrinbana et al., 2010; El Chartouni et al., 2011; Gurung et al., 2011; Boukef et al.,
2012; Drabešová et al., 2013; Dalvant et al., 2018) and also at small spatial scale (Linde et al.,
2002; Kabbage et al., 2008; El Chartouni et al., 2012; Welch et al., 2017; Siah et al., 2018;
Morais et al., 2019). Recently, Gibriel (2019) highlighted the important genetic variation and
population structure among bread and durum wheat isolates from many countries. In Tunisia,
very little is known about the epidemiology, the genetic diversity and structure of Z. tritici
populations harvested from different wheat species and cultivars in the field mostly conducted
on durum wheat (Boukef et al., 2012; Berraies et al., 2013; Naouari et al., 2016).
Hence, this study is the primary investigation of genetic diversity and stucture of Z. tritici
sampled from different species (durum wheat, bread wheat and triticale) in Tunisia. First, our
primary goal was to test the hypothesis that Z. tritici collected from different wheat species
would be genetically differentiated. Second, this study aims to characterize the genetic diversity
between the different Z. tritici- populations collected from bread wheat (Triticum aestivum L.
subsp. aestivum), durum wheat (Triticum turgidum L. subsp. durum (Desf.) and triticale
(Triticosecale spp.) cultivated in a single field at Cap Bon region and to assess the effect of the
host on the pathogen genetic diversity.
2. Materiel and Methods
2.1. Wheat varieties and Z. tritici sampling
In this study, a total of 22 wheat varieties including commercial durum and bread wheat
and triticale cultivars from Algeria, Morocco and Tunisia, previously classified from resistant,
moderately resistant, moderately susceptible, susceptible to highly susceptible (chapitre 3) were
used to study the genetic diversity of Z. tritici under natural conditions (Table 1). The
experiment was carried out during the 2016-17 cropping season at the farmer field located at
Cap Bon Area (36°47’47’’N, 11°0’8’’E, governorate of Nabeul, northwest of Tunisia) which
was reported recently as a primary hot-spot for STB on bread wheat (Bel Hadj Chedli et al.,
2018).
The trial was conducted in an augmented design and each cultivar was grown in
individual sub-plot. All plots were naturally infected by Z. tritici and where not treated with
fungicides. Within each sub-plot, five infected flag leaves from different plants were randomly
collected at growth stage 31 (Zadok Scale) and subsequently one mono-pycnidial isolate was
obtained per leaf following the protocol published by Siah et al. (2010). Only three isolates per
cultivar were analyzed in this study (Table 1). Isolates from each specie and cultivar are
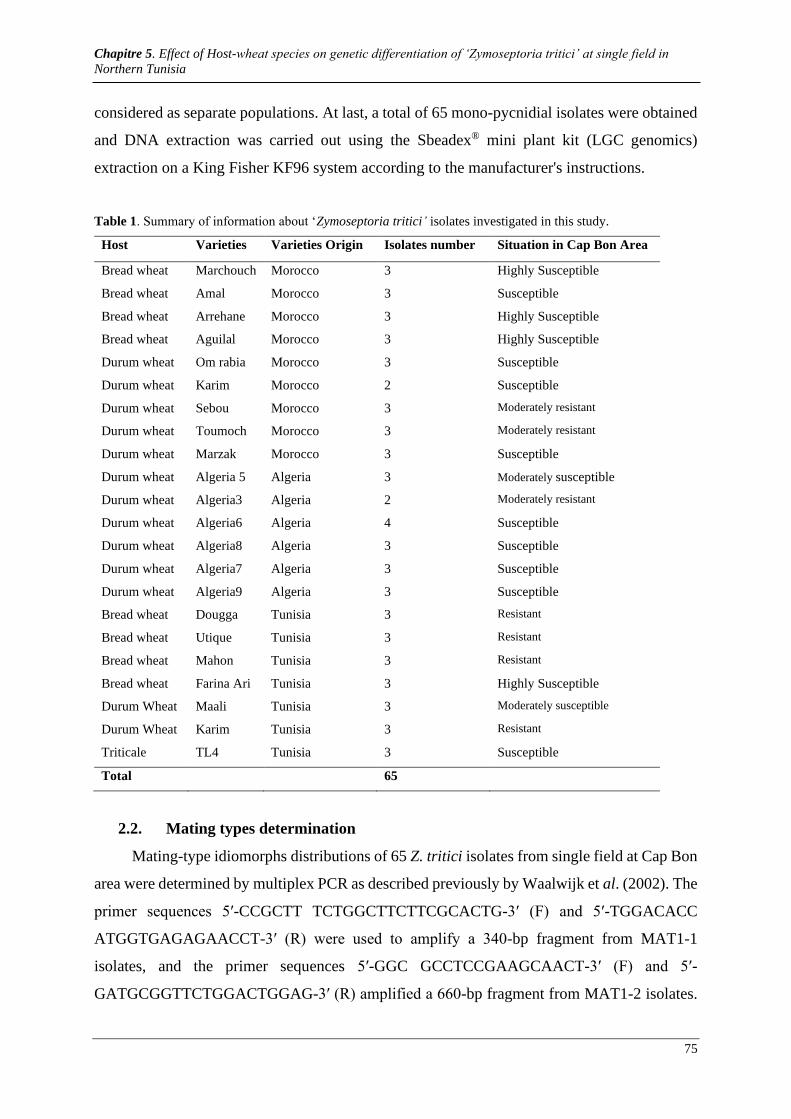
Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
75
considered as separate populations. At last, a total of 65 mono-pycnidial isolates were obtained
and DNA extraction was carried out using the Sbeadex® mini plant kit (LGC genomics)
extraction on a King Fisher KF96 system according to the manufacturer's instructions.
Table 1. Summary of information about ‘Zymoseptoria tritici’ isolates investigated in this study.
Host Varieties Varieties Origin Isolates number Situation in Cap Bon Area
Bread wheat Marchouch Morocco 3 Highly Susceptible
Bread wheat Amal Morocco 3 Susceptible
Bread wheat Arrehane Morocco 3 Highly Susceptible
Bread wheat Aguilal Morocco 3 Highly Susceptible
Durum wheat Om rabia Morocco 3 Susceptible
Durum wheat Karim Morocco 2 Susceptible
Durum wheat Sebou Morocco 3 Moderately resistant
Durum wheat Toumoch Morocco 3 Moderately resistant
Durum wheat Marzak Morocco 3 Susceptible
Durum wheat Algeria 5 Algeria 3 Moderately susceptible
Durum wheat Algeria3 Algeria 2 Moderately resistant
Durum wheat Algeria6 Algeria 4 Susceptible
Durum wheat Algeria8 Algeria 3 Susceptible
Durum wheat Algeria7 Algeria 3 Susceptible
Durum wheat Algeria9 Algeria 3 Susceptible
Bread wheat Dougga Tunisia 3 Resistant
Bread wheat Utique Tunisia 3 Resistant
Bread wheat Mahon Tunisia 3 Resistant
Bread wheat Farina Ari Tunisia 3 Highly Susceptible
Durum Wheat Maali Tunisia 3 Moderately susceptible
Durum Wheat Karim Tunisia 3 Resistant
Triticale TL4 Tunisia 3 Susceptible
Total 65
2.2. Mating types determination
Mating-type idiomorphs distributions of 65 Z. tritici isolates from single field at Cap Bon
area were determined by multiplex PCR as described previously by Waalwijk et al. (2002). The
primer sequences 5′-CCGCTT TCTGGCTTCTTCGCACTG-3′ (F) and 5′-TGGACACC
ATGGTGAGAGAACCT-3′ (R) were used to amplify a 340-bp fragment from MAT1-1
isolates, and the primer sequences 5′-GGC GCCTCCGAAGCAACT-3′ (F) and 5′-
GATGCGGTTCTGGACTGGAG-3′ (R) amplified a 660-bp fragment from MAT1-2 isolates.

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
76
PCR reactions were performed in 2.5 µl volumes containing (6µM) of each primer, 5µl (600
µM) of dNTPs, 5 µl of 10 X reaction buffer, 0.25 unit of Taq polymerase (Promega) and 10 ng
of genomic DNA. The total PCR reaction volume was adjusted with sterile distilled water to 50
µl per PCR reaction. PCR amplifications were performed using a ‘Biorad’ thermocycler with
the following thermal cycling conditions: 1 min at 94°C for initial denaturation, 30 s at 68°C
for annealing, 1 min at 72°C for extension, followed by a final 10-min extension at 72°C. PCR-
amplified products were separated in 1% (wt/vol) agarose gels electrophoresis in 0.5× Tris-
Borate-EDTA (TBE) buffer at 100 V for 45 min.
2.3. Microsatellite analysis
Twelve pairs of single locus microsatellite primers (SSR) markers (St1, St2, St3A, St3B,
St3C St4, St5, St6, St7, St9, St10, and St12) described by Gautier et al. (2014) were used to
study the genetic variability of 65 Z. tritici isolates (details are shown in chapter 3). Two bread
wheat isolates from Netherlands (IPO323 and IPO94269) and one durum wheat isolate from
Algeria (IPO95052) were used as refences in this study. However, 20μL PCR reactions were
performed using the Type-it microsatellite kit (Qiagen) in accordance with manufacture
recommendations. PCR reactions were performed in 25µL volumes with the following
components: 2.5μL deionized water, 2.5μL Q-solution, 2.5μL of the primer mix (containing
2μM of each primer), 12.5μL of the Type-it mix and 5μL of DNA (10ng.µL). Reactions were
run at 95°C for 5 min, followed by 35 cycles of 95°C for 30s, 55°C for 90s and 72°C for 30s,
with a final extension step of 60°C for 30 min, using a PT100 thermocyler (Biorad). The PCR
products were run on 3130xl instrument (Life Technologies) using the Liz500 size standard.
2.4. Data analysis
Mating types distribution was determined by calculating the ratio of mating-type alleles
(Mat 1–1/Mat1–2) and χ2 tests were performed to determine whether the frequencies of the two
mating types within different species-populations departed from the null hypothesis of a 1:1
ratio (Waalwijk et al. 2002).
Genetic diversity in the total population, the total number of alleles and allele frequencies
at each SSR locus, the polymorphism (P%), Shannon information index (I), genetic diversity
(H) and unbiased diversity (uh) were investigated within each population using GENALEX 6.5
software (Peakall and Smouse, 2012). The magnitude of genetic differentiation among
populations was assessed using Wright’s FST statistic (Nei, 1973). Analysis of Molecular
Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
77
Variance (AMOVA) and gene flow index (Nm) were further investigated using GENALEX 6.5
within and among each population.
Structure analyses were carried out to investigate the genetic structure of Z. tritici
populations using Structure software version 2.3.4 (Pritchard et al., 2000). Population structure
analyses were based on Twenty-seven multilocus genotypes which were identified out of the
initial 65 populations. The relationship between individuals was calculated using principal
coordinate analysis (PCoA) to detect genetic divergence among subpopulations (Sun et al.,
2013). A neighbor-joining phylogenetic tree based on distance between subpopulations was
established with DARwin6 software with 1000 bootstraps (Perrier and Jacquemoud-Collet,
2006).
3. Results
3.1. Distribution of mating-type alleles at single field
Among 65 Z. tritici isolates sampled from the three wheat species, three isolates that did
not amplify were not included in mating type frequencies. The two mating types, Mat1-1 and
Mat1-2, were found within the three studied species. However, the χ2 test detected a significant
deviation from the 1:1 ratio at the significance level of P = 0.05 with a great predominance of
Mat1-2 compared to Mat1-1(Table 2).
Table 2. Distribution of Z. tritici mating types within wheat species from single field at Cap Bon region.
Wheat Species Isolates Number MAT 1-1 (%) MAT1- 2 (%) χ2
Durum Wheat 39 30.76 69.23 2.88
Bread wheat 20 35 65 0.9
Triticale 3 0 100 1.5
Total population 62* 30.64 70.49 4.64
* Three isolates that did not amplify were not included in mating type determination
3.2. Genetic variability in core chromosome according to host species
Twenty-seven multilocus genotypes (MLG) were identified from the initial population.
However, high and equal genetic diversity (0.7), Shannon’s index (0.4) and unbiased diversity
(0.4) were recorded for durum and bread wheat Z. tritici populations (Figure 1). Similarly, great
genetic diversity (0.5) was observed for the Z. tritici in triticale population (Figure 1). The most
important level of polymorphism (91%), private alleles number (14) and multilocus genotypes
number (17) were observed for the durum wheat derived Z. tritici isolates (Table 3) followed
by the bread wheat derived isolates population and subsequently by the triticale derived isolate
population with 83% and 75%, of polymorphism level respectively (Table 3).
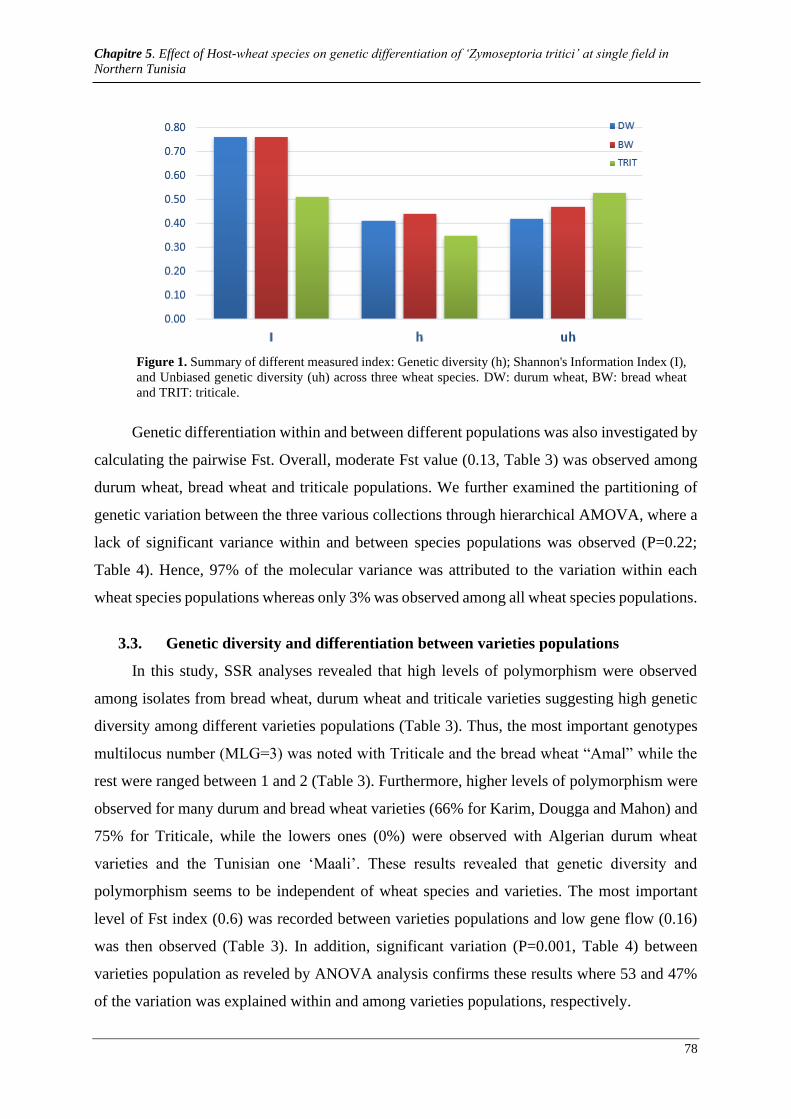
Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
78
Genetic differentiation within and between different populations was also investigated by
calculating the pairwise Fst. Overall, moderate Fst value (0.13, Table 3) was observed among
durum wheat, bread wheat and triticale populations. We further examined the partitioning of
genetic variation between the three various collections through hierarchical AMOVA, where a
lack of significant variance within and between species populations was observed (P=0.22;
Table 4). Hence, 97% of the molecular variance was attributed to the variation within each
wheat species populations whereas only 3% was observed among all wheat species populations.
3.3. Genetic diversity and differentiation between varieties populations
In this study, SSR analyses revealed that high levels of polymorphism were observed
among isolates from bread wheat, durum wheat and triticale varieties suggesting high genetic
diversity among different varieties populations (Table 3). Thus, the most important genotypes
multilocus number (MLG=3) was noted with Triticale and the bread wheat “Amal” while the
rest were ranged between 1 and 2 (Table 3). Furthermore, higher levels of polymorphism were
observed for many durum and bread wheat varieties (66% for Karim, Dougga and Mahon) and
75% for Triticale, while the lowers ones (0%) were observed with Algerian durum wheat
varieties and the Tunisian one ‘Maali’. These results revealed that genetic diversity and
polymorphism seems to be independent of wheat species and varieties. The most important
level of Fst index (0.6) was recorded between varieties populations and low gene flow (0.16)
was then observed (Table 3). In addition, significant variation (P=0.001, Table 4) between
varieties population as reveled by ANOVA analysis confirms these results where 53 and 47%
of the variation was explained within and among varieties populations, respectively.
Figure 1. Summary of different measured index: Genetic diversity (h); Shannon's Information Index (I),
and Unbiased genetic diversity (uh) across three wheat species. DW: durum wheat, BW: bread wheat
and TRIT: triticale.

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
79
Table 3. Genetic diversity of Zymoseptoria tritici populations, genetic differentiation between populations and
gene flow.
N: Isolate Number; Ne = No. of Effective Alleles; P%: polymorphic loci; Fst: Wright’s F index indicating genetic differentiation between
populations; Nm: gene flow; Pa: private allele; SE: Standard Error.
Table 4. Analysis of Molecular Variance (AMOVA) for 65 Zymoseptoria tritici isolates.
Source df SS MS Est. Var. % variation P
Species
populations
Among subpopulations 2 32399.633 16199.817 297.185 3% 0.229
Within subpopulations 62 707596.643 11412.849 11412.849 97%
Total 64 739996.277
11710.034 100%
Cultivars
populations
Among subpopulations 21 474712.444 22605.354 5567.020 47% 0.001
Within subpopulations 43 265283.833 6169.391 6169.391 53%
Total 64 739996.277
11736.411 100%
df: degree of freedom; SS: sum of square; MS: mean of square; P: probability
Population Sub-popluations N Ne (SE) %P MLG Fst (SE) Nm (SE)
Species
populations
Durum wheat 41 2.02(0.29) 91.67 17
Bread wheat 21 2.09(0.22) 83.33 10
Triticale 3 1.7(0.15) 75.00 3
Total 65 1.94(0.13) 83.33 27 0.13(0.03) 3.26 (1.12)
Varieties
populations
Toumouch 3 1.33(0.11) 41.67 2
Marzak 3 1.06(0.06) 8.33 1
Om rabia 3 1.26(0.11) 33.33 2
Sebou 3 1(0) 0.00 1
Karim 2 1.53(0.11) 66.67 2
Marchouch 3 1(0) 0.00 1
Amal 3 1.56(0.17) 58.33 3
Arrehane 3 1.6(0.10) 75.00 2
Aguilal 3 1.75(0.31) 75.00 2
Algeria5 3 1(0) 0.00 1
Algeria3 2 1.08(0.83) 8.33% 2
Algeria6 4 1.1(0.06) 16.67 1
Algeria7 3 1(0) 0.00 1
Algeria8 3 1(0) 0.00 1
Algeria9 3 1.33(0.11) 41.67 2
Dougga 3 1.53(0.11) 66.67 2
Mahon 3 1.53(0.11) 66.67 2
Farina Arbi 3 1.4(0.12) 50.00 2
Utique 3 1.96(0.2) 83.33 2
Karim 3 1.53(0.11) 66.67 2
Maali 3 1(0) 0.00 1
TL4 3 1.7(0.15) 75.00 3
Total 65 1.33(0.02) 37.88 27 0.63 (0.04) 0.16 (0.02)

Chapitre 5. Effect of Host-wheat species on genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
80
3.4. Lack of genetic structure within total population
The PCoA analysis showed a lack of a genetic divergence among different wheat-species
populations (Figure 2). Some durum wheat isolates which were more aggregated and also some
bread wheat isolates have a divergent position without the formation of a distinct group.
Besides, genetic similarity was observed between two Tunisian bread wheat isolates and both
bread wheat isolates from the Netherlands (IPO323 and IPO94269). Therefore, the Algerian
durum wheat isolate (IPO95052), the four-bread wheat and one triticale isolates were grouped
together (Figure 3). Few exceptions were observed with a set of 12 durum wheat isolates that
were located in separate clade from all other Z. tritici investigated populations (Figure 3).
Overall, most of the bread wheat, the durum wheat and the triticale Z. tritici derived
populations were grouped together as revealed by the dendrogram (Figure 3). Structure analysis
confirmed these results where a lack of a genetic structure neither within wheat species
populations nor within cultivars populations was revealed (Figure 4).
Figure 2. PCoA analysis of 65 Z. tritici isolates sampled from three wheat species cropped in single
field at Cap Bon region during 2016-2017 cropping season.

Chapitre 5. Effect of Host-wheat species on Genetic differentiation of ‘Zymoseptoria tritici’ at single field in Northern Tunisia
81
Figure 3. Genetic clustering and relationships between 65 Z. tritici isolates sampled from bread wheat, durum wheat and triticale from 22 wheat varieties cultivated
in Northern Tunisia. The tree was constructed using the weighted neighbor-joining method implemented in DARwin 6 software. Isolates from bread wheat, durum
wheat and triticale were colored, black, blue and green respectively. References-isolates (IPO323 and IPO94269 and IPO95052), we designed with the red color.
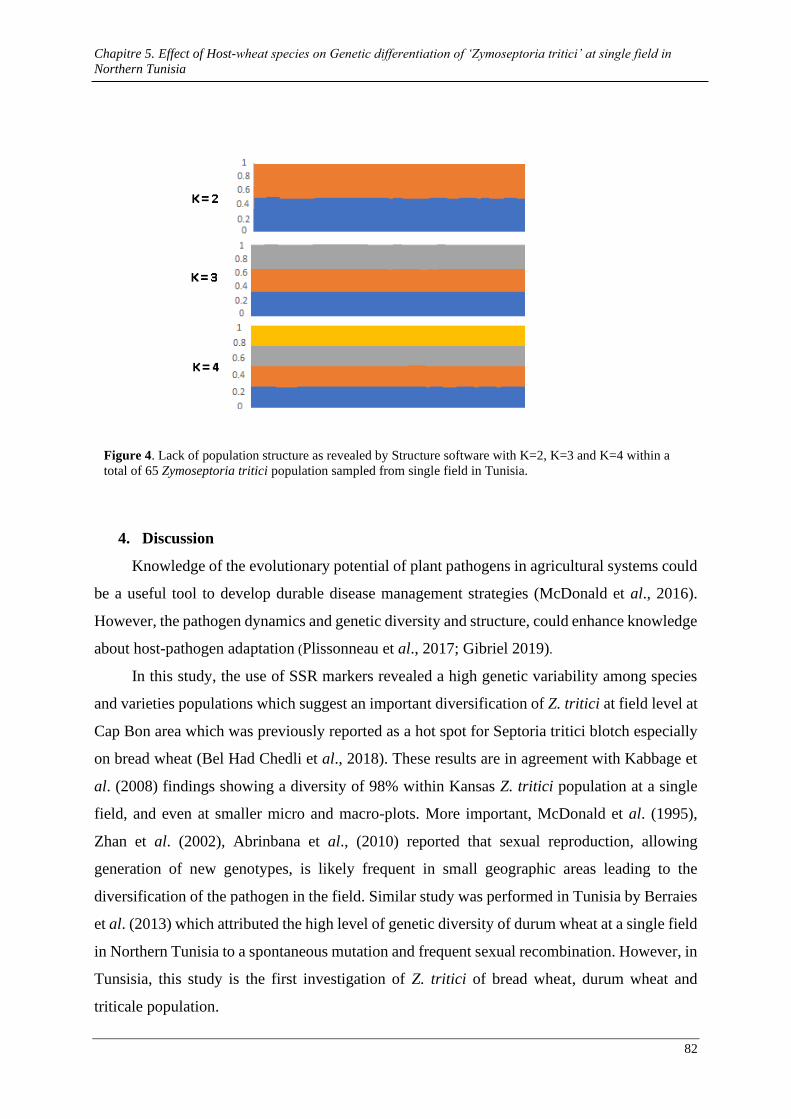
Chapitre 5. Effect of Host-wheat species on Genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
82
4. Discussion
Knowledge of the evolutionary potential of plant pathogens in agricultural systems could
be a useful tool to develop durable disease management strategies (McDonald et al., 2016).
However, the pathogen dynamics and genetic diversity and structure, could enhance knowledge
about host-pathogen adaptation (Plissonneau et al., 2017; Gibriel 2019).
In this study, the use of SSR markers revealed a high genetic variability among species
and varieties populations which suggest an important diversification of Z. tritici at field level at
Cap Bon area which was previously reported as a hot spot for Septoria tritici blotch especially
on bread wheat (Bel Had Chedli et al., 2018). These results are in agreement with Kabbage et
al. (2008) findings showing a diversity of 98% within Kansas Z. tritici population at a single
field, and even at smaller micro and macro-plots. More important, McDonald et al. (1995),
Zhan et al. (2002), Abrinbana et al., (2010) reported that sexual reproduction, allowing
generation of new genotypes, is likely frequent in small geographic areas leading to the
diversification of the pathogen in the field. Similar study was performed in Tunisia by Berraies
et al. (2013) which attributed the high level of genetic diversity of durum wheat at a single field
in Northern Tunisia to a spontaneous mutation and frequent sexual recombination. However, in
Tunsisia, this study is the first investigation of Z. tritici of bread wheat, durum wheat and
triticale population.
Figure 4. Lack of population structure as revealed by Structure software with K=2, K=3 and K=4 within a
total of 65 Zymoseptoria tritici population sampled from single field in Tunisia.

Chapitre 5. Effect of Host-wheat species on Genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
83
The occurrence of a sexual cycle was further revealed by the presence of two mating types
Mat1-1 and Mat1-2 on bread wheat, durum wheat and triticale species which indicate that Z.
tritici isolates undertake sexual reproduction irrespectively to the host species (Waalwijk et al.,
2002; Elbekali et al., 2012; Ayad et al., 2014). The even mating type distribution seems to be
not affected by wheat varieties from which the isolates were harvested. This finding could be
related to the lack of specificity between mating types and wheat varieties during host pathogen
interactions wheat-Z. tritici pathosystem (Kema et al., 1996; Brading et al., 2002; Siah et al.,
2010). The predominance of Mat1-2, as revealed in this study could be due to the small
population size. Hence, a proper sampling is definitely required to avoid a mating type ratio
distortion (Leslie and Klein, 1996; Abrinbana et al., 2010).
In this study, twenty-seven unique genotypes were detected among the total sixty-five
investigated Z. tritici isolates, presenting thus 41% of the total number of isolates. Detecting
unique genotypes is common in Z. tritici population genetic studies with different proportions.
In 2010, Castillo and collaborators, identified 35 and 39 multilocus haplotypes among the 58
isolates of “Los Hornos” and 62 of “Balcarce” locality respectively. The greater number of
identified clones or may be due to the shorter distance between sampling points and the higher
conidial splash-dispersal in the field (Cordo et al., 2006).
On the other hand, an equal genetic diversity, Shannon’s index and Unbiased diversity
were observed within bread and durum wheat derived Z. tritici populations which could prove
that diversity in the field seems to be not affected by wheat species. Therefore, the private allele
number, the proportion of multilocus genotypes (MLG) and the polymorphism level were
relatively high for the durum wheat derived Z. tritici population, but slightly smaller for the
bread wheat and the triticale Z. tritici populations. These findings could be explained by the
unbiased sampling of the diverse Z. tritici populations where we have a higher number of the
durum wheat derived Z. tritici isolates (41) compared to the numbers of isolates collected from
bread wheat (21) and triticale (3).
Interestingly, minor differentiation and important gene flow were detected among wheat
species populations. Similar studies were conducted by Siah et al. (2013) highlighting a low
population differentiation (GST = 0.08) and a high degree of gene flow (Nm = 5.64) of local
French Z. tritici population. In this context, Oğuz et al. (2019) reported that when the pathogen
undergoes asexual reproduction, the risk of epidemic induction through gene flow is high.
Furthermore, Zhan and McDonald (2004) reported that during the long period of co-evolution
of Z. tritici and wheat, gene flow could decrease population differentiation. At Cap Bon Area,

Chapitre 5. Effect of Host-wheat species on Genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
84
the old bread wheat, a tall low yielding bread wheat landrace ‘Farina Arbi’, is cultivated
annually for over a century, and the seeds are maintained by local farmers and not
commercialized. Consequently, the monoculture of wheat ‘Farina Arbi’ landrace at Cap Bon
area could explain the lack of differentiation between populations as the pathogen is hardly
defeated by the local host on which it is well-adapted and established. More importantly
Dalvant et al. (2018) assumed that movement of infected grains might have contributed to such
gene flow. Wind-born ascospores released during sexual reproduction more likely participate
to this homogenization and significantly increases gene flow within the field as well as among
different fields and regions (Zhan et al., 2003; El-Chartouni et al., 2011).
Furthermore, AMOVA analysis highlighted the lack of a significant differentiation
observed among and within durum wheat, bread wheat and triticale derived Z. tritici
populations while 47 and 53% of the genetic variation was explained by differences among and
within cultivars populations, respectively. These results are in agreement with Linde et al.,
(2002) study which reported that the most important level of genetic diversity (94%) occurs
within individual fields. In Tunisia, the existence of a strong gene flow and a lack of
differentiation between populations were previously reported at both regional (Boukef et al.,
2012; Nouari et al., 2016) and field levels (Berraies et al., 2013). However, the genetic variation
between and among different wheat species and cultivars at filed level still not well
documented. Recently, Gibriel (2019) investigated the genetic and genomic diversity of a
worldwide Z. tritici collection sampled from bread and durum wheat in the major global wheat
producing regions. He demonstrated an important genetic variation as revealed by the high
Single Nucleotide Polymorphisms (SNP) rate especially in the Middle East population. Thus,
important structural variation on accessory chromosomes and genome-wide differences
between durum and bread wheat isolates were further reported contributing to the dynamic
nature of Z. tritici populations (McDonald et al., 2016; Hartmann et al., 2017; Gibriel, 2019).
Bayesian statistical and PCoA analyses confirms a lack of a genetic structuration for Z.
tritici population within wheat species in local field in Tunisia. Recently, Welch et al. (2017)
reported an absence of clear structuration of Irish Z. tritici population sampled from different
wheat cultivars and grown at two experimental fields under different levels of diseases pressure.
Nevertheless, significant rates of population structure were reported in many previous studies
at region and country levels (Abrinbana et al., 2010; Drabešová et al., 2012; Siah et al., 2018)
whereas the level of the genetic differentiation seems to vary depending on the markers used
and the geographical locations studied (Nouari et al., 2016).

Chapitre 5. Effect of Host-wheat species on Genetic differentiation of ‘Zymoseptoria tritici’ at single field in
Northern Tunisia
85
The dendrogram constructed using the unweighted neighbor-joining method revealed that
the majority of isolates from different wheat species were grouped together. The two references
bread wheat isolates from the Netherlands (IPO323 and IPO94269) were grouped together with
a set of bread wheat isolates in the same group. Therefore, the Algerian durum wheat isolate
(IPO95052), four bread wheat and one triticale isolates were grouped together which revealed
a genetic similarity between theses isolates. On the other hand, a set of 12 durum wheat isolates
were aggregated and constitutes mostly a separate clade. This study revealed that no genetic
variation and structure was observed within durum and bread wheat species and cultivars. These
results contrasted Gibriel (2019) finding which aimed to assess genetic variation between
durum and bread wheat isolates in order to detect polymorphisms. He reported a large-scale
structural variation in accessory chromosomes and conserved core chromosomes in bread and
durm wheat Z. tritici population worldwide.
Nevertheless, the unbiased sampling due to a small population size investigated during
this study hampered an accurate comparison between the durum and bread wheat derived Z.
tritici isolates. Moreover, a genetic structure was expected and that could provide an insight
regarding the Z. tritici- host specificity (ŠÍP et al., 2015).
5. Conclusion
This study constitutes a preliminary characterization of the Z. tritici population sampled
from different wheat species at a single field in Northern Tunisia. Additional large collections
of isolates need to be analyzed in order to further investigate the population differentiation
observed in three wheat species and also to identify the different causes that could influence the
lack of the genetic structure of Z. tritici in Tunisia such as: varieties adaptation; disease
pressure, geographic spreads and meteorological conditions. The use of other marquers such as
SNPs could be helpful to detect a genetic stucutre between durum and bread wheat isolates.
Examination of cultivar resistance under conditions of natural infection and further field
sampling and pathogen population analyses are needed to assess the population biology and the
genetic structure of STB, to develop new wheat varieties in national breeding programs and
improve disease management strategies.

Discussion générale et conclusions

Discussion Générale et Conclusion
86
Discussion Générale et Conclusion
Les premiers blés tétraploïdes ont été cultivés en Syrie plus particulièrement à Damas. Le
Blé dur (Triticiumtu rgidium spp durum) est apparu pour la première fois à Can Hasan III en
Turquie il y a 7500 ans (Ren et al., 2013). La migration de l’Homme a conduit très vite à
l’expansion géographique du blé (Baloch et al., 2017) disséminé initialement vers l’Europe et
le Nord de l’Afrique durant la période Néolithique (Zapata et al., 2004). Aujourd’hui, le blé
s’étend sur environ 17 millions d’ha dans le Grand Maghreb représentant ainsi la première
source alimentaire. En Tunisie, la production du blé reste tributaire des conditions climatiques
et des pratiques culturales (Rastoin et Benabderrazek, 2014). Toutefois, l’utilisation excessive
des fertilisants, les semis précoces, l’augmentation de la densité du semis et le recours aux
variétés semi-naines ont augmenté considérablement les attaques des bio-agresseurs notamment
les maladies foliaires (Ben Mouhamed et al., 2000).
Causée par Zymoseptoria tritici, Septoria tritici blotch (STB) est considérée parmi les
principales maladies fongiques qui entrave annuellement les champs de blé et particulièrement
le blé dur (Ben Mohamed et al., 2000 ; Ammar et al., 2011). Les premiers résultats obtenus
après deux saisons agricoles successives 2015-16 et 2016-17, ont montré la gravité de la
septoriose qui a été enregistrée dans pratiquement toutes les régions céréalières prospectées
(Bizerte, Béjà, Le Kef, Jendouba, Zaghouan et la région du Cap Bon-Sud comme ‘Soliman,
Grombalia et Beni khalled’) chez toutes les variétés de blé dur et principalement ‘Maali, Karim
et Razzak’. Ces dernières sont les plus utilisées par les agriculteurs du Nord-Ouest qui ont
recours au minimum à deux traitements fongicides pour lutter contre la septoriose (Allagui et
al., 2014).
Ainsi, pour faire face à cette situation, un programme national d’amélioration pour le
développement des variétés à la fois dotée d’un haut rendement et résistantes à la septoriose,
parait essentiel (Sbei et al., 2009 ; El Falleh et Gharbi, 2014). D’où l’intérêt d’installer des
essais variétaux à travers des plateformes afin d’identifier de nouvelles sources de résistance.
Par ailleurs, certains chercheurs ont proposé le recours aux mélanges variétaux comme
une alternative pour réduire les attaques de STB (Gigot, 2013). Dans le cadre des essais
variétaux installés à la « Plateforme Septoriose » au sein de la station « Kodia », Ben M’Barek
et ses collaborateurs (2019) ont pu déduire que l’ajout des variétés résistantes ‘Salim’ et

Discussion Générale et Conclusion
87
‘Monastir’ à raison de 25% avec la variété sensible ‘Karim’ a diminué de manière significative
les attaques de Z. tritici d’une part et amélioré le rendement d’autre part.
Dans un autre contexte de réduction des attaques de STB, les études de Ferjaoui (2015)
ont conduit à l’identification du premier QTL majeur chez l’accession ‘Agili 39’ qui a été
cultivée durant plusieurs années successives dans la station expérimentale du CRRGC à Oued
Béjà connu comme un hot-spot de STB chez le blé dur. Cette accession a prouvé sa résistance
vis-à-vis de plusieurs isolats aux stades plantule et adulte. Un peu plus tard, Aouini (2018) a
montré que les analyses de résistance dans la population recombinante générée ont révélé que
la résistance à large spectre de ‘Agili 39’ résulte du pyramidage naturel de plusieurs QTLs à
effets mineurs. Elle a ajouté que les QTLs identifiés sur les régions chromosomiques 2BL et
2BS ont un effet majeur sur la résistance de ‘Agili 39’ à Z. tritici.
Tandis que la résistance du blé tétraploïde à Z. tritici n’a pas suscité beaucoup d’attention
et peu de travaux ont traité l’interaction Z. tritici-blé dur (Aouini, 2018), la résistance chez le
blé hexaploïde a été bien documentée et jusqu’à présent 21 gènes majeurs ou stb et 167 loci
quantitatifs (QTLs) ont été identifiés et déployés dans les travaux de sélection dans le monde
(Ghaffary et al., 2011 ; Brown et al., 2015 ; Mirzadi et al., 2015). En Tunisie, les variétés
commerciales de blé tendre sont dotées d’un excellent niveau de résistance sous l’infection
naturelle dans la région d’El Haouria comme il a été démontré dans le chapitre 2. Cultivées à
Béjà, connue par des attaques sévères de septoriose presque chaque année, ces variétés ont
connu une absence totale de la maladie. Cependant, il serait très important de chercher des
sources de résistances dans les variétés Tunisiennes de blé tendre, qui seront utiles pour les
programmes de sélection et d’amélioration. L’introgression des gènes de résistances du blé
tendre au blé dur serait d’un intérêt majeur.
Toutefois, malgré la bonne résistance des variétés de blé tendre à STB en Tunisie, cette
étude vient de prouver pour la première fois, une grande sensibilité d’une variété de blé tendre
cultivée exclusivement dans un petit village appartenant à la région d’El Haouaria avec des
incidences et des sévérités les plus importantes durant les deux saisons agricoles (2016 et 2017).
Depuis des décennies, pour la désigner, les agriculteurs de ce village utilisent la dénomination
générale ‘Farina arbi’ et ils procèdent à leur propre multiplication tout en refusant d’introduire
ou de cultiver d’autres variétés commerciales. En occupant des superficies très importantes dans
la région d’El Haouaria, cette variété ancienne sert particulièrement à la fabrication du « kaak »
qui fait partie des traditions de cette région.
De ce fait, et suite à la monoculture de la variété ‘Farina Arbi’, les agriculteurs se trouvent
face à une situation alarmante et un nouveau « hot-spot de STB » chez le blé tendre qui vient

Discussion Générale et Conclusion
88
de se créer dans la région d’El Haouaria. Devant cette situation, il est fortement recommandé
aux agriculteurs d’El Haouaria de pratiquer la rotation des cultures pour briser le cycle de
reproduction sexuée de Z. tritici qui persiste d’une année à une autre à cause de l’intensification
et la monoculture de la variété ‘Farina Arbi’ pour éliminer toutes structures vivantes (débris et
chaumes) pouvant héberger les ascospores.
De plus, ce qui a été surprenant dans cette recherche, c’est que la variété ‘Farina Arbi’
classée comme sensible dans la région d’El Haouaria a montré une résistance exceptionnelle à
ce pathogène lorsqu’elle a été cultivée à Béjà et classée comme immune. En plus, une infection
non significative a été observée chez les variétés commerciales de blé dur (Karim, Salim et
Nasr) cultivées à El Haouaria. Ceci peut être attribué à la dominance de la culture de blé dur
par rapport à celle de blé tendre dans la région de Béjà et de la culture de blé tendre à El
Haouaria tout en sachant que ‘Farina Arbi’ domine à peu près 60% des cultures céréalières dans
cette région (Bel Hadj Chedli et al., 2018). Ces résultats laissent à suggérer une spécialisation
physiologique de Z. tritici envers l’espèce T. aestivum dans cette région.
Toutefois, bien que la spécificité du pathosystème Z. tritici-blé envers l’espèce a attiré
l’attention de plusieurs chercheurs et a créé des débats depuis une cinquantaine d’années (Eyal
et al., 1973), cette interaction reste toujours non claire. De plus, la spécificité de ce pathogène
a été rapportée aussi bien chez les hexaploïdes (T. aestivm, AABBDD, 2n=42) (Kema et al.,
1996a ; Kema et al., 1996b; Kema et Van Silfout, 1997) que chez les tétraploïdes (T. turgidum
L. subp durum, AABB, 2n=28) (Ghaneie et al., 2012; Medini et Hamza, 2008) suggérant ainsi
une interaction gène pour gène (GFG) entre le blé et Z. tritici (Kema et al., 2018). Cette
constatation a été précédemment signalée par Eyal et al. (1973), Van Ginkel et Scharen (1988),
Johnson (1992), qui ont référé la spécificité de Z. tritici à la virulence/avirulence envers des
cultivars particuliers de blé dur ou de blé tendre. D’autre part, une évidence de l’existence d’une
division dans la population de Z. tritici qui sépare les isolats de blé tendre, virulents uniquement
sur blé tendre, et les isolats de blé dur adaptés uniquement au blé dur, a été rapportée (Kema et
al., 1996). Ainsi, certains auteurs ont attribué ceci à la dominance d’une espèce par rapport à
une autre dans certains pays comme dans le cas du blé dur en Tunisie, en Algérie et au Maroc
(Sayoud, 1995 ; Yahyaoui et al., 2000 ; Hamza et al., 2008).
Cette hypothèse vient d’être confirmée dans cette étude qui a dévoilé une grande
sensibilité des variétés Marocaines de blé tendre, résistantes au Maroc, dans la région d’El
Haouaria. Cultivées à Béjà, toutes les variétés Tunisiennes et Marocaines de blé tendre étaient
résistantes et toutes les variétés de blé dur quelle que soit l’origine étaient sensibles. Ces
constatations supposeraient que les populations de Z. tritici ont développé un pouvoir pathogène

Discussion Générale et Conclusion
89
plus élevé envers le T. durum à Béjà et envers le blé tendre T. aestivum à El Haouria comme
une réaction d’adaptation (Kema et al., 1996 a et b; Medini et Hamza, 2008). Ces résultats ne
pourront que confirmer d’une part le phénomène de la spécificité de Z. tritici envers l’espèce
hôte en Tunisie et d’autre part dévoiler la haute sensibilité des variétés de blé tendre et de blé
dur d’origines Marocaine et Algérienne lorsqu’elles sont cultivées en Tunisie. Il parait donc
essentiel de déterminer le niveau de spécificité de l’espèce hôte des isolats de blé tendre et blé
dur, en effectuant des inoculations croisées sur un panel de variétés de blé dur et blé tendre
Tunisiens et d’installer des essais variétaux à travers les plateformes dans les pays de l’Afrique
du Nord d’une façon particulière.
Par ailleurs, le génotypage d’une collection de 65 isolats de blé tendre, blé dur et triticale
d’origines Marocaine, Algérienne et Tunisienne cultivées à proximité dans la région du Cap
Bon durant la saison agricole 2016-17 en utilisant 12 marqueurs SSR, a révélé que la diversité
génétique, la richesse allélique, la diversité non biaisée et l’indice de Shannon chez les deux
populations de blé tendre et de blé dur sont similaires et des valeurs relativement basses pour
la population de Triticale. Ceci prouve en partie que la diversité au sein du champ semble ne
pas être affectée par l’espèce cultivée dans cette étude et dévoile le faible potentiel de l’espèce
et de la variété dans la variation génétique de Z. tritici (Welch et al., 2017).
En tenant compte de l’espèce mise en jeu, l’étude de la composante principale (ACP) et
l’analyse Bayésienne ont montré que les isolats de blé tendre, blé dur et triticale sont groupés
ensemble à l’exception d’un nombre limité d’isolats qui ont montré une certaine parenté. Une
divergence bien claire a été observée chez quelques isolats de blé tendre qui ont été groupés
ensemble et autres de blé dur qui ont montré une similarité génétique entre eux. A l’échelle
régionale la majorité des isolats de blé tendre ont été groupés ensemble avec les trois isolats de
référence (IPO323, IPO94269, IPO95052). Des constations similaires ont été obtenues par
Berraies et al. (2013) qui ont pensé que lorsque les sources d’inoculum sont originaires d’une
population locale de Z. tritici, les spores conservent ainsi une structure familiale limitée.
L’étude de la variabilité génétique et phénotypique des populations de l’agent pathogène
constitue un défi majeur pour le développement d’une gestion durable avec des cultivars
résistants et facilite la mise en place des programmes de sélection et des stratégies de lutte
(McDonald, 1997 ; McDonald et Linde, 2002 ; McDonald et al., 2016). Toutefois, la diversité
génétique de Z. tritici a été bien documentée et plusieurs travaux ont été élaborés dans ce sujet.
En 2003, une étude a porté sur une collection de 1673 isolats à l’échelle continentale, (Algérie,
Californie, Canada, Danemark, Australie, Allemagne, Israel, Mexico, Oregon, Syrie, Texas,
Uruguay, Royaume-Uni) montrant ainsi que la plus grande diversité génétique a été observée

Discussion Générale et Conclusion
90
en Israel ce qui a conduit à constater que le croissant fertile est le centre d’origine de Z. tritici
(Linde et al., 2000 ; Zhan et al., 2003). Toutefois, une importante diversité génétique du
génome nucléaire a été notée au niveau du champ (~12%) comparée à celle entre les régions
(~5%) et entre les continents (~3%). En plus, une faible différentiation entre les différentes
populations due à un important flux de gène a été ainsi rapportée. Des marqueurs AFLP ont été
utilisés pour étudier la diversité génétique de ce pathogène à plusieurs niveaux : micro et macro-
plots (Kabbage et al., 2008). Ces mêmes marqueurs ont été investigués pour montrer une
importante diversité génétique chez des isolats de blé dur de l’Algérie, de la Tunisie, et du
Canada (Medini and Hamza, 2008). Des comparaisons de la diversité génétique d’une
population Irlandaise de Z. tritici collectée à partir de différentes variétés (Welch et al., 2017)
et sur plusieurs années en France (El Chartrouni et al., 2011 et 2012 ; Morais et al., 2019) ont
été aussi abordées moyennant différents marqueurs moléculaires (SSR et SSCP).
En Tunisie, très peu de travaux ont traité la diversité génétique de ce pathogène et
uniquement trois travaux ont été élaborés dans ce sujet. La première étude de la génétique des
populations de Z. tritici en Afrique du Nord et en Tunisie particulièrement a été menée par
Boukef (2012) qui, en se basant sur 11 marqueurs microsatellites, a signalé une grande diversité
génétique de Z. tritici à partir d’une large collection de plusieurs pays (218 isolats). Récemment,
Nouari et al. (2016) ont étudié le génome nucléaire et mitochondrial de ce pathogène chez 108
isolats de blé dur collectés de Bizerte, Béjà, kef et Jendouba. Ainsi, une grande diversité
génétique, un important flux de gène et une faible différentiation entre les populations et une
absence de structuration ont été notés. A l’échelle parcellaire, des résultats similaires ont été
signalés par Berraies et al. (2013) moyennant une population de 45 isolats de blé dur (Karim)
collectée d’un seul champ à Béjà en utilisant 7 marqueurs SSR.
Dans cette étude, l’approche moléculaire basée sur 12 marqueurs microsatellites, a révélé
une importante diversité dans une population Tunisienne de Z. tritici collectée à partir du blé
tendre à travers trois regions (Cap Bon, Bizerte et Béjà). Ainsi, le grand nombre d’allèles privés,
de génotypes multilocus (MLG) et l’indice de Shannon les plus importants ont été enregistrés
dans tous les champs prospectés à El Haouaria. Une telle diversité génétique élevée qui a été
trouvée au sein de toutes les populations pourrait, d’une part être à l’origine d’une capacité
d’adaptation du pathogène aux différentes conditions climatiques des régions humide et semi-
aride de la Tunisie (Boukef, 2012). D’autre part, ceci peut être lié à l’occurrence régulière d’un
cycle sexué de Z. tritici dans cette région ce qui a été prouvé par la présence des proportions
presque égales de MAT1-1 et MAT1-2 (Bel Hadj Chedli et al., 2019). Les ascospores
transportés par le vent sur de longues distances génèrent ainsi de nouveaux allèles et la

Discussion Générale et Conclusion
91
recombinaison génétique entre ces types sexuels compatibles lors du cycle sexué aboutit à
l’apparition de nouveaux génotypes conduisant par conséquent à une grande diversification
(Zhan et al., 2003; Siah et al., 2013; Siah et al., 2018). Par contre, la dominance du MAT1-2
observée dans la région de Goubellat et les faibles pourcentages ainsi notés à Béjà et Bizerte ne
peuvent en aucun cas négliger la présence d’un cycle sexué dans ces régions. Par ailleurs, ceci
peut être attribué au nombre réduit des isolats collectés à partir de ces endroits et l’inadéquation
de l’échantillonnage (El Chatrouni et al., 2012) du fait que Z. tritici était rarement présent dans
les champs de blé tendre en Tunisie.
Le polymorphisme ainsi observé à El Haouaria laisse suggérer que probablement cette
population de Z. tritici possède naturellement de nouveaux gènes à échanger ce qui a augmenté
considérablement la diversité génétique et a influencé la pathogénicité de la population
(McDonald et Mundt, 2016 ; Dalvant et al., 2018). Malgré qu’un important flux de gène a été
signalé dans les différentes populations étudiées quelle que soit l’échelle considérée : région,
parcelle, espèce et variété, une faible différentiation génétique ainsi qu’une absence de
structuration ont été notées. Ces résultats peuvent être liés à la dispersion des spores qui se
produiraient probablement sur une large zone géographique et par conséquent un flux de gène
sur une longue distance (McDonald et al., 1999 ; Linde et al., 2002 ; Zhan et al., 2003). Les
mêmes constatations ont révélé une absence de structuration chez d’autres agents pathogènes
en Tunisie à savoir Fusarium culmorum agent causal de la Fusariose du Blé (Oufensou et al.,
2019). Par contre une structuration bien claire a été observée dans une population clonale de
Puccinia striiformis, agent causal de la rouille jaune, dans les régions méditerranéennes
(Tunisie, Maroc, Algérie…), en Europe (France, Allemagne, Belgique…) et aussi à travers une
population recombinante en Chine et au Sud de l’Asie (Népal et Pakistan) (Thach et al., 2016).
En Tunisie, la taille réduite de la population étudiée de Z. tritici, la limitation des zones
d’échantillonnage et le type de marqueur utilisé pourraient aussi expliquer l’absence d’une
structure claire des populations (Siah et al., 2018). Toutefois, il est connu que les différents
marqueurs moléculaires peuvent causer des différences même en utilisant les mêmes
échantillons (Drabešová et al., 2012). De leur part, Väli et ses collaborateurs (2008) pensaient
que les marqueurs SSR ne sont pas capables de capturer les petites variations génomiques qui
peuvent se reproduire au niveau de chaque nucléotide. Cette constatation a été confirmée
recemment par Gibriel (2019) qui, en utilisant des ‘Single nucleotide polymorphisms (SNPs)’,
a pu détecter une grande diversité génétique et une importante structuration d’une collection de
Z. tritici collectée à partir du blé tendre et du blé dur tout en soulignant ainsi une specialisation
de Z. tritici -blé.

Discussion Générale et Conclusion
92
Le génotypage moyennant des (SNPs) pourrait probablement détecter une stucturation
pareille en Tunisie. L’utilisation d’un plus grand nombre de marqueurs pourrait être ainsi
recommandée. D’autres travaux sont nécessaires pour consolider les résultats trouvés dans cette
investigation, et plusieurs études doivent être aussi effectuées non seulement au niveau régional
et parcellaire mais aussi au niveau de la même feuille et la même lésion sur plusieurs années.
La recherche des sources de variations de ces isolats génétiquement distincts reste à déterminer.
Le croisement des différentes souches de Z. tritici collectées à partir du blé tendre ‘Farina Arbi’
de la region du Cap Bon, et l’installation des series d’inoculations artificielles dans des
chambres de culture avec une caractérisation phénotypique et génotypique et le séquençage du
génome de plusieurs souches de blé tendre issues de cette région sont ainsi recommandés pour
trouver des explications génétiques de cette specialisation signalée dans la region d’El Haouria.

Références
93
Références Bibliographiques
Abrinbana M., Mozafari J., Shams-Bakhsh M., Mehrabi R. 2010. Genetic structure of
Mycosphaerella graminicola populations in Iran. Plant Pathology, 59: 829-838.
Allagui M. B., Sebei A., et Sakkouhi L. 2014. Importance de l’association de variétés de blé
dur dans la réduction des maladies fongiques foliaires et l’amélioration de la productivité.
Journée Nationale sur la valorisation des résultats de la Recherche dans le domaine des
Grandes Cultures, 8-10.
Ammar K., Gharbi M.S., and Deghais M. 2011. Wheat in Tunisia. In Bonjean, A.P.Angus, and
W.J. van Ginkel, M. (eds.) The world wheat book: A history of wheat breeding. Lavoisier,
Paris, France, (2):443-463.
Anonyme 1. Ministère de l’agriculture, des ressources hydrauliques et de la pêche, 2015.
Anonyme 2. FAO, Food and Agriculutre Policy decision Analysis. Country fact sheet on food
and agriculture policy trends 2017,1-6.
Anonyme 3. Valorisation des produits forestiers au Nord-Ouest. Ministère de développement
et de coopération internationale. Office de développement du Nord-Ouest, 2004.
Aouini L. 2018. Durum wheat and septoria tritici blotch : gene and prospects for breeding. PhD
thesis, Wageningen University, Wageningen, Netherlands: 225p.
Ayad D., Sayoud R., Benbelkacem K., et Bouznad Z. 2014. La tâche septorienne du blé:
Première signalisation de la présence en Algérie des deux Mating types du téleomorphe
Mycosphaerella graminicola (Fuckel) Schröter, (anamorphe: Septoria tritici Rob. Ex
Desm.) et diversité phénotypique de l’agent pathogène. “Nature & Technologie ". B-
Sciences Agronomiques et Biologiques: 34 -45.
Bachta M.S. 2011. La céréaliculture en Tunisie Une politique de régulation à repenser, note
d’analyse du CIHEAM, 64: 3.
Balotch F.S., Alsaleh A., Shahid M.Q., Ciftci V., et al. 2017. A hole genome DArTseq and SNP
analysis for genetic diversity assessment in durum wheat from central fertile crescent.
PloS one, 12: e 0167821.
Banke S., Peschon A., et McDonald B A.2004. Phylogenetic analysis of globally distributed
Mycrosphaerella graminicola populations based on three DNA sequence loci. Fungal
Genet. Biol,41:226-23.
Banke S., et McDonald B.A, 2005. Migration patterns among global populations of the
pathogenic fungus Mycosphaerella graminicola. Molecular Ecology, 14: 1881-1896.

Références
94
Bannon F., and Cook B.M. 1998. Studies on dispersal of Septoria tritici pycnidiospores in
wheat-clover intercrops. Plant Pathol, 47:49-56
Bathgate J.A., et Loughman R. 1994. Are ascospores a significant source of primary inoculum
of leptoshaeria nodorum, L. avenariaf. sp avenaria and Mycosphaerella graminicola in
the Northern wheat belt of Western Australia. In proceeding of the 4th international
workshop on septoria of cereals. pp :271-275.
Bel Hadj Chedli R., Ben M’Barek S., Yahyaoui A., Kehel Z., Rezgui S.2018. Occurrence of
Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and bread
wheat in Northern Tunisia. Chliean Journal of Agricultural Research, 78(4):559-568.
Bel Hadj Chedli R., Aouini, L., Ben M’Barek S., Yahyaoui, A., Rezgui, S., et Chaabène, H.
2019. Distribution of mating types in Zymoseptoria tritici populations collected from
bread wheat at El Haouaria region. Journal of New sciences, 61(5): 3892-3898.
Benbelkacem K., Bouznad Z., Benslimane H., Salhi N.L. 2011. Occurrence of septoria leaf
blotch in Algéria and assessment of wheat resistance. 8ème International Symposium on
Mycrosphaerella and Stagnospora Diseases of Cereals, Mexico City September: 10-14.
Ben Hamouda H., Ben Youssef S., Sonia Bedhiaf S., Mohamed Annabi M., Zied Draief Z., and
Gharbi M.S. 2016. Status of agricultural innovations, innovation platforms, and
innovations investment. Tunisia. 2015 PARI project country report: Republic of Tunisia.
Forum for Agricultural Research in Africa (FARA), Accra, Ghana.
Ben Hassine A., & Hamada W. 2014. First isolation of the Mycosphaerella graminicola
teleomorph stage causing Septoria leaf blotch on wheat in Tunisia. New Disease Reports
29, 18.
Ben Jabeur M., Somai-Jemmali L., et Hamada W. 2017. Thyme essential oil as an alternative
mechanism: biofungicide-causing sensitivity of Mycosphaerella graminicola. Journal of
Applied Microbiology, 122:932-939.
Ben M’Barek S., Karisto P., Fakhfakh M., Mikaberidze A., Yahyaoui A. 2019. Improved
control of Septoria trtici blotch in durum wheat using cultivar mixtures. bioRxiv: 1-23.
Ben Mohamed L., Rouaissi M., Sebei A., Hamza S., and Harrabi M. 2000. Effet du génotype,
de la date de semis, de la fertilisation azotée et potassique et des fongicides sur le
développement de Septoria tritici. p. 350-356. In Royo, C., Nachit, M., Di Fronzo, N.,
and Araus, J.L. (eds.) Durum wheat improvement in the Mediterranean region: New
challenges. CHIEHAM- options méditerranéennes : Série A. Séminaires Méditerranéens,
40: 350-356.

Références
95
Ben Salem M., Daaloul A., et Ayadi A. 1995. Le blé dur en Tunisie. In : Di Fonzo N. (ed.),
Kaan F. (ed.), Nachit M. (ed.). Durum wheat quality in the Mediterranean region
Zaragoza. CIHEAM Options Méditerranéennes : Série A. Séminaires Méditerranéens,
22 :81-91.
Berraies S., Gharbi M.S., Belzile F., Yahyaoui A., Hajlaoui M.R., Trifi M., Jean, M., et Rezgui
S. 2013. High genetic diversity of Mycospaherella graminicola (Zymoseptoria tritici)
from a single wheat field in Tunisia as revealed by SSR markers. African Journal of
Biotechnology, 12: 1344-1349.
Berraies S., Ammar K., Gharbi M.S., Amor Y, Rezgui, S. 2014. Quantitative inheritance of
resistance to Septoria tritici blotch in durum wheat in Tunisia. Chilean Journal of
Agricultural Research, 74: 35-40.
Bœuf F. 1936. Rôle du Service Botanique et Agronomique dans l’Economie Agricole de la
Tunisie. Résultats acquis, Programme en cours de réalisation. La Tunisie Agricole N°3 :
203-221
Boughalleb N., and Harrabi M. 1997. Physiologic specialization of Septoria tritici in North-
Africa. Tropicultura 15(1) :33-36.
Boukef, S. 2012. Structure des populations, spécificité parasitaire et évaluation de la résistance
aux fongicides chez Zymoseptoria tritici, agent de la septoriose du blé en Tunisie. Thèse
de doctorat de L’Institut National Agronomique de Tunisie, 1-102.
Brading P.A., Vertrappen E.C.P., Kema G.H.J., Brown J.K.M. 2002. A gene for gene
relationship between wheat and Mycosphaerella graminicola, the Septoria tritici blotch
pathogen. Phytophathology 92:439-445.
Brown J.K.M., Chartrain L., Lasserre-Zuber P., Saintenac C. 2015.Genetics of resistance to
Zymoseptoria tritici and applications to wheatbreeding. Fungal Genetics and Biology,
79 :33-41.
Brunner P.C., Stefano F.F., Croll T.D., Stukenbrock E., McDonald B.A. 2013. Coevolution and
Life Cycle Specialization of Plant Cell Wall Degrading Enzymes in a Hemibiotrophic
Pathogen. Mol. Biol. Evol. doi:10.1093/molbev/mst04.
Castillo N., Cordo C., Simón M. R. 2010. Molecular variability among isolates of
Mycosphaerella graminicola, the causal agent of septoria tritici blotch, in Argentina.
Chartrain L., Brading P.A., Makepeace J.C., Brown J.K.M. 2004. Sources of resistance to
Septoria tritici blotch and implications for wheat breeding. Plant Pathology, 53: 454-460.

Références
96
Chartrain L., Berry S.T., & Brown J.K.M., 2005. Resistance of wheat line Kavkaz-K4500
L.6.A.4 to septoria tritici blotch controlled by isolate-specific resistance genes.
Phytopathology, 95: 664-71.
Chebil A., & Frija A. 2016. Impact of improving water-use efficiency on its valuation: The case
of irrigated wheat production in Tunisia. African Journal of Agricultural and Resource
Economics, 11 (2) : 131-140.
Chen R.S, &McDonald B.A.1996. Sexual reproduction plays a major role in the genetic
structure of populations of the fungus Mycosphaerella graminicola. Genetics 142:1119-
1127.
Cordo C.A., Linde C.C., Zhan J., & McDonald B. 2006. Genotypic diversity of the wheat leaf
blotch pathogen (Septoria tritici) in Buenos Aires province. Sociedad Argentina Botánica,
41: 293–305.
Cordo C.A., Monaco C.I., Altamirano R., Perello A.E., Larran S., Kripelz N.I., and Simon M.R.
2017. Wheat Conditions Associated with the Release and Dispersal of Zymoseptoria
tritici Spores in the Argentine Pampas Region. International journal of Agronomy: 1-13.
Coppin E., Debuchy R., Arnaise S., et Picard M. 1997. Mating type and sexual developpement
in filamentous Ascomycetes. Microbiology and Molecular Biology Reviews, 61 :441-
428.
Cuthbert R.D. 2011. Molecular mapping of septoria tritici blotch resistance in hexaploid wheat
(Triticum aestivum L.). PhD thesis, University of Manitoba, Canada.
Dalvand M., Zafar D., Soleimani Pari M.J., Roohparvar R., and Tabib Ghafari S.M. 2018.
Studying Genetic Diversity in Zymoseptoria tritici, Causal Agent of Septoria Tritici
Blotch, by Using ISSR and SSR Markers.J. Agr. Sci. Tech, 20: 1307-1316.
Duncan K.E., Howard R.J. 2000. Cytological analysis of wheat infection by the leaf blotch
pathogen Mycosphaerella graminicola - Mycological research, 104( 9) : 1074-1082.
De Wolf E.2008. Septoria Tritici Blotch. Plant pathology. EP-133.
Djerbi M., et Ghodhbane A. 1975. Contribution à l’étude du Septoria tritici Rob. ex Desm.,
conservation et résistance variétale. Annales de l’INRAT, 48, Fasc. 1.
Drabešová J., Ryšánek P., Brunner P., McDonald B.A., and Croll D. 2013. Population genetic
structure of Mycosphaerella graminicola and quinone outside inhibitor (QoI) resistance
in the Czech Republic. European Journal of Plant Pathology: 135, 211.
Drummond J.B., Craigie R.A., Braithwaite M., Gillum A.T. McCloy B.L. 2015.The effect of
fungicide dose rate and mixtures on Zymoseptoria tritici in two cultivars of autumn sown
wheat. New Zealand Plant Protection 68: 420-427.

Références
97
Duda R. O., and Hart P. E. 1973. Pattern Classification and Scene Analysis. Wiley, New York.
Elbekali A.Y., Siah A., Ramdani A., El Chartouni L., Tisserant B., Deweer C., Hafidi M.,
Reignault, P., Halama P. 2012. First report of the mating types MAT1-1 and MAT1-2 of
Mycosphaerella graminicola in Morocco. J. Plant Pathol. 94:243–247.
El Chartouni L., Tisserant B., Siah A., Duyme F., Leducq J.-B., Deweer C., et al. 2011. Genetic
diversity and population structure in French populations of Mycosphaerella graminicola.
Mycologia, 103, 764.
El Chartouni L., Tisserant B., Siah A., Duyme F., Durand R., Halama P., & Reignault P. 2012.
Evolution of Mycosphaerella graminicola at the wheat leaf and field levels.
Phytopathologia Mediterranea, 51(3):32.
El Felah M., Gharbi M.S. 2014. Les céréales en Tunisie : Historique, contraintes de
développement et perspectives. Journée Nationale sur la valorisation des résultats de la
Recherche dans le domaine des Grandes Cultures : 1-7.
EL Felah M., Gharbi M.S., Ben Ghanem H., et Elloumi M. 2015. Les céréales en Tunisie entre
mythe et réalité. Annales de l’INRAT, 2ème Numéro Spécial Centenaire, 88 :1-17.
Estep L.K., Torriani S.F.F., Zala M., Anderson N.P., Flower M.D., McDonald B.A., Mundt
C.C. et Brunner P.C. 2015. Emergence and early evolution of fungicide resistance in
North American populations of Zymoseptoria tritici, 64: 961-971.
Eyal Z., Amiri Z., Wahl I. 1973. Physiologic specialization of Septoria tritici. Phytopathology
63:1087-1091.
Eyal Z., Scharen A.L., Huffman M.D., et Prescott J.M. 1985. Global insights into virulence
frequencies of Mycosphaerella graminicola. Phytopathology, 75: 1456-1462.
Eyal Z., Scharen A.L., Prescott J.M., et Vanginkel M. 1987. The Septoria Diseases of Wheat
Concepts and Methods of Disease Management. CIMMYT, Mexico, D.F, 10-35.
Eyal Z. 1999. The Septoria/ Stagonospora blotch diseases of wheat: past, peresent, and future.
p. 177–182. In: “Septoria and Stagonospora Disease of Cereal: A Compilation of Global
Research”. Mexico, D.F.: CIMMYT: 177–182.
Fakhfakh M.M., Rezgui S., Jarrahi T., Zouani R., Bousalmi A., Sgatni K., et Yahyaoui A. 2011.
Mapping distribution of Septoria allowed targeting management options of the disease.
8th International symposium on Mycosphaerella and Stagonospora diseases of cereals.
CIMMYT, Mexico.
Feldman M., et Sears R.E. 1981. The wild gene sources of wheat. Scientific American, 244:
102-112.

Références
98
Ferjaoui S., M’Barek S.B., Bahri B., Slimane R.B., and Hamza S. 2015. Identification of
resistance sources to Septoria tritici blotch in old durum wheat germplasm applied for the
analysis of the Zymoseptoria tritici-durum wheat interaction. Journal of Plant Pathology
97 (3): 471- 481.
Fones H., & Gurr S. 2015. The impact of Septoria tritici blotch disease on wheat: An EU
perspective. Fungal Genetics and Biology, 79:3.
Fones H.N., Eylesa C.J., Kaya W., Cowpera J, Gurr S.J. 2017. A role for random, humidity-
dependent epiphytic growth prior to invasionof wheat by Zymoseptoria tritici. Fungal
Genetics and Biology, 106:51-60
Garcia C., et Marshall D.1992. Observations on the ascogenous stage of Septoria tritici in
Texas. Mycological Research, 96: 65-70.
Garrett K.A., Nita M., De Wolf E.D., Gomez L., and Sparks A.H. 2009. Plant Pathogens as
Indicators of Climate Change. Elsevier B.V:425-436.
Gautier A., Marcel T., Confais J., Crane C., Kema G., Suffert F., et Walker A.S. 2014.
Development of a rapid multiplex SSR genotyping method to study populations of the
fungal plant pathogen Zymoseptoria tritici. BMC Research Notes, 7(373): 1-10.
Ghaffary S.M.T, Robert O., Laurent V., Lonnet P., et al.2011. Genetic analysis of resistance to
septoria tritici blotch in the French winter wheat cultivars Balance and Apache. Theor
Appl Genet., 123:741–754.
Ghaffary S.M.T., Faris J.D., Friesen T.L., Visser R.J.F., et al. 2012. New broad-spectrum
resistance to septoria tritici blotch derived from synthetic hexaploid wheat. Theor Appl
Genet., 124:125–142.
Gharbi M.S., et Deghaies M. 1997. Principaux enseignements à tirer de la campagne céréalière
1995/1996. Actes des 3èmes journées nationales sur les acquis de la recherche agronomique
halieutique et vétérinaire, 29/11-01/12/1996, Nabeul, Tunisie.
Gharbi M.S., Deghais M., Ben Amar F. 2000. Breeding for resistance to Septoria tritici in
durum wheat. In: Royo C. (ed.), Nachit M. (ed.), Di Fonzo N. (ed.), Araus J.L. (ed.).
Durum wheat improvement in the Mediterranean region : New challenges. Options
Méditerranéennes : Série A. Séminaires Méditerranéens,40 : 397- 401.
Gharbi M.S., Berraiess Fakhar S., Ammar K., Hajlaoui M.R., Yahyaoui A., et Trifi M. 2008.
Breeding durum wheat with resistance to Septoria leaf blotch for sustainable production
under Mediterranean conditions. International durum wheat symposium: from seed to
pasta, 30/06-03/07/2008, Bologne, Italie.

Références
99
Gharbi M.S., Berraies S., Ammar K., Yahyaoui A. 2011. Delivering disease resistant cultivars
to enhance sustainability of durum wheat production in Tunisia. p. 60. In Duveiller E, Singh
PK, et al. (eds.) 8th International Symposium on Mycosphaerella and Stagonospora
Diseases of Cereals, Mexico City. 10-14 September 2011. Book of Abstracts. CIMMYT,
Mexico.
Gharbi M.S., et El Faleh M. 2013. Les céréales en Tunisie : Plus d’un siècle de recherche
variétale. Annales de l'INRAT, Numéro Spécial Centenaire, 86 :45-68.
Ghaneie A., Mehrabi R., Safaie N., Abrinbana M., Saidi A., Aghaee M. 2012. Genetic variation
for resistance to Septoria tritici blotch in Iranian tetraploid wheat landraces. European
Journal of Plant Pathology, 132:191-202.
Gibriel H. 2019.Comparative pathogenomics in the ascomycete plant pathogens Zymoseptoria
tritici and Verticillium dahlia. Wageningen, Netherland.
Gigot C. 2013. Potentialités des associations de variétés pour limiter la progression épidémique
de la septoriose du blé: rôle des mécanismes de dispersion des spores par la pluie dans un
couvert végétal hétérogène. AgroParisTech – Institut National de la Recherche
Agronomique Unité Environnement et Grandes Cultures, 17-27.
Gilmour R., Gogel B.J., Cullis B.R., Welham S.J., and Thompson R. 2002. ASReml user guide
release 1.0. VSN International Ltd., Hemel Hempstead, UK.
Gladders P., Paveley N.D., Barrie I.A., Hardwick N.V., et al. 2001. Agronomic and
meteorological factors affecting the severity of leaf blotch caused by Mycosphaerella
graminicola in commercial wheat crops in England. Annals of Applied Biology, 138, 301–
311.
Gohari A. 2015. Identification and functional characterization of putative (a) virulence factors
in the fungal wheat pathogen Zymoseptoria tritici. PhD Thesis, Wageningen University, The
Netherlands
Goodwin S.B. 2007.Back to basics and beyond: increasing the level of resistance to Septoria
tritici blotch in wheat. Australasian Plant Pathology, 36: 532–538.
Gouache D., Bensadoum A., Brun F., Pagé C., et al. 2013. Modeling climate change impact on
Septoria tritici Blotch (STB) in France Accounting for climate model and disease model
uncertainty. Agricultural and Forest Meteorology, 170:242.252.
GrapheAgri 2016. Céréales - Agreste :120-127.
http://agreste.agriculture.gouv.fr/IMG/pdf/Gaf16p120-131.pdf
GrapheAgri 2016. Céréales - Agreste :117-128.
http://agreste.agriculture.gouv.fr/IMG/pdf/Gaf2017p117-128.pdf

Références
100
Gurung S., Goodwin S.B., Kabbage M., Bockus W., et B. Adhikari T. 2011. Genetic
Differentiation at Microsatellite Loci Among Populations of Mycosphaerella graminicola
from California, Indiana, Kansas, and North Dakota. Phytopathology, 101: 1251-1257.
Hailu E., et Woldeab G. 2015. Survey of Rust and Septoria Leaf Blotch Diseases of Wheat in
Central Ethiopia and Virulence Diversity of Stem Rust Puccinia graminis f. sp. tritici.
Advances in Crop Sciences Technology, (3):2.
Halama P. 1996. The occurrence of Mycosphaerella graminicola, teleomorph of Septoria tritici
in France. Plant Pathology, 45: 135-138.
Hanson S. 2016. Tunisia Grain and Feed Annual. USDA Foreign Agricultural service, GAIN
Report Number: 1-8.
Harrabi M., et Cherif M. 1990. Diagnostic des maladies de grandes cultures. Document
technique de l’INAT, 50.
Harrat W., and Bouznad Z. 2018. Prevalence, cultural and pathogenic characterization of
Zymoseptoria tritici, agent of wheat septoria leaf blotch, in Algeria. African Journal of
Agricultural Research, 13(40): 2146-2153.
Hartman F.E. 2017. Population Genomics of adaptation in the fungal wheat pathogen
Zymoseptoria tritici. Thèse de Doctorat, ETH Zurich.
Henze M., Beyer M., Klink H., et Verreet J.A. 2007. Characterizing Meteorological Scenarios
Favorable for Septoria tritici Infections in Wheat and Estimation of Latent Periods. Plant
disease, 91(11):1445-1449.
Hunter T., Coker R.R., et Royle D.J. 1999. The teleomorph stage, Mycosphaerella graminicola,
in epidemics of Septoria tritici blotch on winter wheat in the UK. Plant Pathology, 48:
51–57.
Holloway G.2014. Septoria tritici blotch of wheat. DEPI Information Note Series May 2014. 2
p. Department of Environment and Primary Industries (DEPI), Melbourne, Australia.
Jilbene, 1996. Etat des recherches sur la septoriose au Maroc. Proceeding du Symposium
régional sur les Maladies des céréales et les Légumineuses Alimentaires Edit. INRA
Rabat, 11-14.
Johnson R.1992. Past, present and future opportunities in breeding for disease resistance, with
examples from wheat. Breeding for disease resistance. Springer: 3-22
Jürgens T., Linde C.C., and McDonald B.A. 2006. Genetic structure of Mycosphaerella
graminicola populations from Iran, Argentina and Australia. European Journal of Plant
Pathology 115: 223-233.

Références
101
Kabbage M., Leslie J.F., Zeller K.A., Hulbert H., et Bockus W.W. 2008. Genetic diversity of
Mycosphaerella graminicola, the causal agent of Septoria tritici blotch, in Kansas winter
wheat. Jounral of Agricultural, Food and Environmental Sciences, 2 (1):1934-7235.
Karisto P., Hund A., Yu K., Anderegg J., Walter A., Mascher F., McDonald B.A, et
Mikaberidze A. 2017. Ranking quantitative resistance to Septoria tritici blotch in elite
wheat cultivars using automated image analysis. bioRxiv: 3-44.
Kellner R., Bhattacharyya A., Poppe S., Hsu T.Y. et al. 2014. Expression Profiling of the Wheat
Pathogen Zymoseptoria tritici Reveals Genomic Patterns of Transcription and Host-
Specific Regulatory Programs. Genome Biology and Evaluation. 6(6):1353–1365.
Kema G.H.J., Annone J.G., Sayoud R., Van Silfhout C.H, Van Ginkel M., and De Bree J. 1996
a. Genetic variation for virulence and resistance in the wheat–Mycosphaerella graminicola
pathosystem I. Interactions between pathogen isolates and host cultivars. Phytopathology,
86: 200- 212.
Kema, G.H.J., Sayoud R., Annone, J.G., Silfhout C.H. van. 1996 b. Genetic variation for
virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem II.
Analysis of interactions between pathogen isolates and host cultivars Phytopathology 86,
213-220. Kronstard, J.W., et Staben,C. 1997. Mating type in filamentous fungi. Annual
Review of Genetics. 31:245-276.
Kema, G.H.J., Verstappen, E.C.P., Todorova, M., and Waalwijk, C. 1996. Successful crosses
and molecular tetrad and progeny analyses demonstrate heterothallism in Mycosphaerella
graminicola. Curr. Genet. 30: 251-258.
Kema G.H.J., et Van Silfhout C.H. 1997. Genetic variation for virulence and resistance in the
wheat–Mycosphaerella graminicola pathosystem III. Comparative seedling and adult plant
experiments. Phytopathology, 87:266-272.
Kema G. H. J., Goodwin S. B., Hamza S., Verstappen E. C. P ET AL. 2002. A Combined AFLP
and RAPD Genetic Linkage Map of Mycosphaerella graminicola, the Septoria tritici Leaf
Blotch Pathogen of Wheat. Genet., 161: 1497–1505.
Kema G.H.J., Gohari A.M., Aouini L., Gibriel H.A.Y, Ware S.B., et al. 2018. Stress and sexual
reproduction affect the dynamics of the wheat pathogen effector AvrStb6 and strobilurin
resistance. Nature Genetics, 50:375-380.
Kettles G.J., et Kanyuka K. 2016. Dissecting the Molecular Interactions between Wheat and
the Fungal Pathogen Zymoseptoria tritici. Frontiers in plant sciences, 7(508): 1-7.

Références
102
Kidane Y.G., Hailemariam B.N., Mengistu D.K., Fadda C., Pè M.E., et Dell’Acqua M. 2017.
Genome-Wide Association Study of Septoria tritici Blotch Resistance in Ethiopian
Durum Wheat Landraces. Frontiers in plant sciences, 8:1-3.
Krupinsky J. M., and Tanaka D.L. 2001. Leaf Spot Diseases on Winter Wheat Influenced by
Nitrogen, Tillage, and Haying after a Grass–Alfalfa Mixture in the Conservation Reserve
Lasram M. 2013. INRAT History: a hundred years serving Tunisian Agriculture. Annales de
l’INRAT, Numéro Spécial Centenaire, 86 :6-42.
Leslie J.F., Klein K.K. 1996. Female fertility and mating type effects on effective population
size and effective population size and evolution in filamentous fungi. Genetics, 144:557-
567.
Linde C.C., Zhan J., McDonald B.A. 2002. Population structure of Mycosphaerella
graminicola: from lesions to continents. Phytopathology, 92: 946-955.
Ltifi A., et Sakkouhi L. 2008. La septoriose du blédur : Résistance Variétale et impact de la
maladie sur le rendement. Annales de l’INRAT :76-84.
Lounis-Lalou F. 2005. Bilan d’activité 2004/2005. Amélioration des céréales. Ferme
expérimentale de démonstration et production de semences d’Oued Smar, ITGC, 35.
Lawson J. 2016. daewr: Design and Analysis of Experiments with R. R package version 1:1-7.
https://CRAN.R-project.org/package=daewr
Macauley H. 2015. Document de référence : Les cultures céréalières : riz, maïs, millet, sorgho
et blé. Nourrir l’Afrique. Centre international de référence Abdou Diouf :1-13.
Madariaga B.R.1986. Occurrence in Chile of Mycosphaerella graminicola (Fuckel) Schroeter,
teleomorph of Septoria tritici Rob. Ex Desm. Agricultura Tecnica,46, 209-211.
Mamluk A.F., Gilchrist D.G., et Singh M. 1995. Variartion in virulence in isolates of
Mycrosphaerella graminicola from six geographical wheat growing regions.
Phytopathology. Medit, 34:45-51.
Marshall R., Kombrink A., Motteram J., Loza-Reyes E., et al. 2011. Analysis of two in planta
expressed LysM effector homologs from the fungus Mycosphaerella graminicola reveals
novel properties and varying contricbution to virulence on wheat. Plant physiology,
156(2): 756-769.
Mazzouz H., Saadaoui M., Jilbène M., Lyamani A. 1995. Importance des septorioses du blé au
Maroc en 1991. Al Awamia, 91 :63-69.
McDonald B.A., Martinez J.P., 1990. Restriction fragment length polymorphisms in Septoria
tritici occur at a high frequency. Current Genetics, 17:133–138.

Références
103
McDonald B.A., Pettway R.E., Chen R.S., Boeger J.M., Martinez, J.P. 1995. The population
genetics of Septoria tritici (teleomorph Mycosphaerella graminicola). Canadien Journal
of Botany, 73:292-301.
McDonald B.A. 1997. The Population Genetics of Fungi: Tools and Techniques.
Phythopathology, 87(4):448-453.
McDonald B.A., Zhan J., et Burdon J.J. 1999. Genetic Structure of Rhynchosporium secalis in
Australia. Ecologie and Population Biology, 89(8): 639-644
McDonald B.A., Linde C.C. 2002. The population genetics of plant pathogens and breeding
strategies for durable resistance. Euphytica, 124: 163-180.
McDonald B.A. 2014. Using dynamic diversity to achieve durable disease resistance in
agricultural ecosystems. Tropical Plant Pathology, 39(3):191-196.
McDonald B.A. 2015. How can research on pathogen population biology suggest disease
management strategies? The example of barley scaled (Rhynchosporium commune). Plant
Pathology, 64: 1005-1013.
McDonald B.A., et Mundt C.C. 2016. How knowledge of Pathogen Population Biology Informs
Management of Septoria Tritici Blotch. Phytopathology, 106(09): 948-955.
Meamiche H.N., Aouini L., Bouznad Z., Gerrit H.J. 2018. Equal distribution of mating type
alleles and the presence of strobilurin resistance in Algerian Zymoseptoria tritici field
populations. Plant Disease :1-27.
Medini M., and Hamza S. 2008. Pathotype and molecular characterization of Mycosphaerella
graminicola isolates collected from Tunisia, Algeria, and Canada. Journal of Plant
Pathology, 90:65–73.
Medini M., Ferjaoui S., Bahri B., Mhri W., Hattab S., Hamza S. 2014. Bulk segregant Analysis
and marker-trait association reveal common AFLP markers for resistance to Septoria leaf
blotch in Tunisian old durum wheat. BASE Biotechnology, Agronomy, Society.
Environment,18(1):3-10.
Mizradi G.A.M., Ware S, Wittenberg H.J., Mehrabi R., et al. 2015. Effector discovery in the
fungal wheat pathogen Zymoseptoria tritici. Molecular Plant Pathology: 1-15.
Mojerlou S., Safaie N., Alizadeh A., Khelghatibana F. 2009. Measuring and modeling crop
loss of wheat caused by septoria leaf blotch in seven cultivars and lines in Iran. Journal
of Plant Protection Research 49: 257-262.
Morais D., Laval V., Sache I., et Suffert F. 2015. Comparative pathogenicity of sexual and
asexual spores of Zymoseptoria tritici (septoria tritici blotch) on wheat leaves. Plant
pathology:1-11.

Références
104
Morais D., Duplaix C., Sache I., Laval V., Suffert F., and Walker., A.S. 2019. Overall stability
in the genetic structure of a Zymoseptoria tritici population from epidemic to
interepidemic stages at a small spatial scale. European Journal of Plant Pathology.
doi.org/10.1007/s10658-018-01666-y.
Naouari M., Siah A., Elgazzah M., Reignault P., and Halama P. 2016. Mitochondrial DNA-
based genetic diversity and population structure of Zymoseptoria tritici in Tunisia.
European Journal of Plant Pathology,146:305-314.
Nasraoui B. 2008. Principales maladies fongiques des céréales et légumineuses en Tunisie,
Nei M.1973. Analysis of gene diversity in subdivided populations. Proceedings of the National
Academy of Sciences of the United States of America, 70: 3321-3323.
Nelson S.C., 1996. A Simple analysis of disease foci. Phytopathology, 86:332-339.
Odlibekov F., Armoniené R., Henriksson T., Chwade A. 2018. Proximal Phenotyping and
Machine Learning Methods to Identify Septoria Tritici Blotch Disease Symptoms in
wheat. Frontier in Plant Science, 9:1-11.
O’Driscoll A., Kildea S., Doohan F., Spink J., & Mullins E. 2014. The wheat–Septoria conflict:
A new front opening up? Trends in Plant Science, 19(9): 602-610.
Oğuz A.C., Ölmez F., and Karakaya A. Genetic Diversity of Net Blotch Pathogens of Barley
in Turkey. International Journal of Agriculture & Biology: 1814-9596.
Oufensou S., Balmas V., Scherm B., Rau D., Camiolo S. 2019. Genetic variability, chemotype
distribution, and aggressiveness of Fusarium culmorum on durum wheat in Tunisia.
Phytopathologia Mediterranea, 58:103-113.
Palma-Guerrero J., Torriani S.F.F., Zala M., Carter D., et al. 2016. Comparative transcriptomic
analyses of Zymoseptoria tritici strains show complex lifestyle transitions and
intraspecific variability in transcription profiles. Molecular Plant Pathology, 17(6), 845-
859.
Peakall R., and Smouse P.E. 2012. GenAlEx 6.5: genetic analysis in Excel. Population genetic
software for teaching and research-an update. Bioinformatics, 28:2537-2539.
Perrier X., and Jacquemoud-Collet J.P. 2006. DARwin6 software.
Pietravalle S., Shaw M.W., Parker S.R., et Van Den Bosch F. 2003. Modeling of relationships
between weather and Septoria tritici epidemics on winter wheat: a critical approach.
Phytopathology, 93, 1329–1339.
Plissonneau C., Benevenuto J., Mohd-Assaad N., Fouché S., Hartmann F. E., and Croll, D.
2017. Using population and comparative genomics to understand the genetic basis of
effector-driven fungal pathogen evolution. Frontiers in Plant Sciences, 8:119.

Références
105
Ponomarenko A., Goodwin S.B., and Kema G.H.J. 2011. Septoria tritici blotch (STB) of wheat
Septoria tritici blotch (STB) of wheat. Plant Heal. Instr. :1–7.
Poppe S. 2014. Host specialization in the fungal plant pathogen Zymoseptoria tritici.
Dissertation, Dem Fachbereich Biologie der Philipps-Universität Marburg vorgelegt von:
2-73.
Pritchard J.K., Stephens M., et Donnelly P. 2000. Inference of population structure using
multilocus genotype data. Genetics, 155: 945.
Quadvlieg W., Kema, G.H.J., Groenewald J.Z., Verkley, et al. 2011. Zymoseptoria gen. nov.:
a new genus to accommodate Septoria-like species occurring on graminicolous hosts.
Persoonia 26, 57–69.
Ramdani A., Halama P., Elbekali1 A.Y., Siah A., et al. 2011. Septoria tritici blotch of wheat in
Morocco: current status and perspective. 8th International Symposium on
Mycosphaerella and Stagonospora Diseases of Cereals Mexico City September.
Rapilly F. 1991. L’épidémiologie en pathologie végétale : mycoses aériennes. Institut National
de la recherche agronomique. Paris,11-36.
Rastoin J.L., et Ben Abderrazik H. 2014. Céréales et oléoprotéagineux au Maghreb : Pour un
co-développement de filières territorialisées. Tunisie : le poids des institutions. Editions
Institut de Prospective Economique du Monde Méditerranéen (IPEMED), Maroc, 37.
Razavi M., & Hughes G. 2004. Molecular variability of Mycosphaerella graminicola as
detected by RAPD markers. Journal of Phytopathology, 152(10), 543–548.
R Core Team. 2017. R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Ren J., Sun D., Chen L., You F.M., Peng Y., et al. 2013. Genetic Dversity revealed By Single
Nucleotide Polymorphism Markers in a Worldwide Germplasm Collection of Durum
Wheat. International Journal of Molecular Sciences, 14:7061-7088.
Rezgui S., Fakhfakh M.M., Boukef S., Rhaiem A., Chérif Mo., Chérif Ma et Yahyaoui A.H.
2008. Effect of common cultural practices on Septoria leaf blotch disease and grain yield
of irrigated durum wheat. Tunisian Journal of Plant Protection, 3: 59-67.
Ross McKenzie.1998. Wheat nutrition and fertiliser requirements-Nitrogen.
Roohparvar R. 2007. Drug transporters of the fungal wheat pathogen Mycosphaerella
graminicola. Thèse de doctorat, Université de Wageningen, Netherland, 182.
Rudd J.J, Kanyuka K., Hassani-Pak K., Derbyshire M., et al. 2015. Transcriptome and
Metabolite Profiling of the Infection Cycle of Zymoseptoria tritici on Wheat Reveals a
Biphasic Interaction with Plant Immunity Involving Differential Pathogen Chromosomal

Références
106
Contributions and a Variation on the Hemibiotrophic Lifestyle Definition. Plant
Physiology, 167: 1158–1185
Russell V. Lenth. 2016. Least-Squares Means: The R Package lsmeans. Journal of Statistical
Software, 69(1): 1-33.
Saade M.E. 1996. Adoption and impact of high yielding wheat varieties in Northern Tunisia.
CIMMYT. Economics Working Paper 96-03. CIMMYT, Mexico D.F.
Saari E.E., Wilcoxcon R.D.1974. Plant situation of high-yielding dwarf wheats in Asia and
Africa. Annu.Rev. Phytopathology.12, 49-68.
Saari E.E, Prescott J.M.1975. A scale for appraising the foliar intensity of wheat diseases. Plant
Disease Reporter, 59:377-380.
Saidi A., Safaie N., and Palizi P. 2012. Study on mating types and sensitivity to strobilurin
fungicide in fungal wheat pathogen Mycosphaerella graminicola. Crop Protection, 1(4):
271-277.
Sánchez-Vallet A., McDonald M.C., Solomon P.S., McDonald B.A.2015. Is Zymoseptoria
tritici a hemibiotroph?. Fungal Genetics and Biology, 79:29-32.
Sanderson FR, 1972. A Mycosphaerella species as the ascogenous state of Septoria tritici
Rob.and Desm. New Zealand Journal of Botany,10 :707-709.
SAS Institute 2014. ‘JMP® 11 scripting guide.’ 2nd edn (SAS Institute Inc.: Cary, NC, USA)
Sayoud R., Ezzahiri B., et Bouznad Z. 1999. Les maladies de céréales et de légumineuses
alimentaires au Maghreb. I.T.G.C.
Sbei A., 2009. Contribution à l’étude de la variabilité de la résistance à la septoriose chez le blé
dur et la virulence chez Septoria tritici : Conséquences sur le rendement. Thèse de
Doctorat, Institut National Agronomique de Tunisie : 129.
Schneider F., Koch G., Jung C., and Verreet J. A. 2001. Genotypic diversity of the wheat leaf
blotch pathogen Mycosphaerella graminicola (anamorph) Septoria tritici in Germany.
European Journal. Plant Pathology, 107: 285-290.
Seifbarghi S., Razavi M., Aminian H., Zare R., and Etebarian H.E. 2009. Studies on the host
range of Septoria species on cereals and some wild grasses in Iran. Phytopathologia
Mediterranea, 48, 422–429
Siah A., Tisserant B., El Chartouni L., Duyme F., et al. 2010. Mating type idiomorph from
French population of the wheat pathogen Mycrosphaerella graminicola : widespread
equal distribution and low but distinct levels of molecular polymorphism. Fungal
Biology, 114: 980-990.

Références
107
Siah A., Reignault P., & Halama P.2013. Genetic diversity of Mycosphaerella graminicola
isolates from a single field. Communications in Agricultural and Applied Biological
Sciences, 78, 437.
Siah, A., cedex, L., Elbekali, A.Y., Ramdani, A., Reignault, P., Torriani, S.F.F., Brunner, P.C,
Halama P. 2015. QoI Resistance and Mitochondrial Genetic Structure of Zymoseptoria
tritici in Morocco. Plant Disease, 98 (8): 1138-1144.
Siah A., Bomble M., Tisserant B., Cadalen T., Holvoet M., Hilbert R., Reignault P., Halama P.
2018. Genetic structure of Zymoseptoria tritici in Northern France at region, field, plant
and leaf layer scales. Phytopathology :1-42.
Singh R.P., Singh P.K., Rutkoski J., Hodson D.P., He X., Jørgensen L.N., Hovmøller M.S. and
Huerta-Espino J. 2016. Disease Impact on Wheat Yield Potential and Prospects of
Genetic Control.
ŠÍP V., Chropava J., and Palicova J. 2015. Response of Selected Winter Wheat Cultivars to
Inoculation with different Mycosphaerella graminicola Isolates. Czech J. Genet. Plant
Breed., 51, 2015 (3): 86–95.
Simko I., Piepho H.P. 2012. The Area Under the Disease Progress Stairs: Calculation,
Advantage, and Application. Phythopathology, 102(4) : 381-389.
Slama A., Ben Salem M., Ben Naceur M., Zid E. 2005.Les céréales en Tunisie : production
effet de la sécheresse et mécanismes de résistance. Sécheresse, 16, (3) :225-229.
Somai-Jemmali L. 2015. Stimulateurs des défenses naturelles du blé dur en Tunisie et du blé
tendre en France contre la septoriose causée par Zymoseptoria tritici. Thèse de Doctorat
en cotutelle INAT-ULCO.
Somai-Jemmali L., Randoux B., Siah A., Magnin-Robert M., et al. 2016. Similar infection
process and induced defense patterns during compatible interactions between
Zymoseptoria tritici and both bread and durum wheat species. European Journal of plant
pathology,
Steinberg G. 2015. Cell Biology of Zymoseptoria tritici. Pathogen cell organization and wheat
infection. Fungal Genetic and Biology, 79 :17-23.
Stewart T.M., Perry A.J., and Evans, M.J. 2014. Resistance of Zymoseptoria tritici to
azoxystrobin and epoxiconazole in the lower North Island of New Zealand. New Zealand
Plant Protection, 67: 304-313.
Stukenbrock E.H., Banke S., Javan-Nikkhah M., and McDonald B. A. 2007. Origin and
domestication of the fungal wheat pathogen Mycosphaerella graminicola via sympatric
speciation. Mol Biol Evol. 24:398–411.

Références
108
Stukenbrock E. H., Frank G. J., Marcello Z., Hansen T.T., McDonald B.A., Schierup M.H.
2010. Whole-Genome and Chromosome Evolution Associated with Host Adaptation and
Speciation of the Wheat Pathogen Mycosphaerella graminicola. Recent Genome
Evolution of M. graminicola. PLoS Genetics, 6 (12) :1-10.
Stukenbrock E.H., Bataillon T., Dutheil J.Y., Hansen, T.T., et al. 2011. The making of a new
pathogen: insights from comparative population genomics of the domesticated wheat
pathogen Mycosphaerella graminicola and its wild sister species. Genome Res. 21:2157-
2166.
Suffert F., et Sache I. 2011. Relative importance of different types of inoculum to the
establishment of Mycosphaerella graminicola in wheat crops in north-west Europe. Plant
Pathology,60: 878-889.
Suffert F., Sache I., Lannou C. 2013. Assessment of quantitative traits of aggressiveness in
Mycosphaerella graminicola on adult wheat plants. Plant Pathology, 62:1330-1341.
Suffert F., Ravigné V., et Sache I. 2015. Can short-term selection for fitness traits in a fungal
plant pathogen be driven by seasonal changes? A case study in the wheat pathogen
Zymoseptoria tritici. Appl. Environ. Microbiol:1-27.
Suffert F., Morais D., Delestre G., Gélisse S., Laval V., et Sache I. 2016. Investigation of the
mechanisms of sexual reproduction in Zymoseptoria tritici and their consequences on
STB dynamics. INRA BIOGER, ISSDC9 Paris.
Suffert F., Delestre G., Gélisse S.2018. Sexual Reproduction in the Fungal Foliar Pathogen
Zymoseptoria tritici Is Driven by Antagonistic Density Dependence Mechanisms.
Microbial Ecology.
Sun X., Kang,S., Zhang Y., Tan X., Yu Y., et al. 2013. Genetic diversity and population
structure of rice pathogen Ustilaginoidea virens in China. PLoS ONE, 8(9), e76879.
Taher K., Graf S., Fakhfakh M.M., Salah H.B.H., et al. 2014. Sensitivity of Zymoseptoria tritici
Isolates from Tunisia to Pyraclostrobin, Fluxapyroxad, Epoxiconazole, Metconazole,
Prochloraz and Tébuconazoles. Journal of Phytopathology, 162:442-448.
Thach T., Ali S., Vallavieille-Pope C., Justesen A.F., et Hovmøller M.S. 2016. Worldwide
population structure of the wheat rust fungus Puccinia striiformis in the past. Fungal
Genetics and Biology, 87 :1-8.
Takele A., Lencho A., W/Ab G., Hailu E., Kassa B. 2015. Status of Wheat Septoria Leaf Blotch
(Septaria tritici Roberge in Desmaz) in South West and Western Shewa Zones of Oromiya
Regional State, Ethiopia. Research in Plant Sciences, 3(3): 43-48.

Références
109
Teferi T.A., Gebreslassie Z.S. 2015. Occurrence and intensity of wheat Septoria tritici blotch
and host response in Tigray, Ethiopia. Crop protection, 68:76-71.
Torriani S.F.F., Brunner P.C., McDonald B.A., & Sierotzki H. 2009. Qoi resistance emerged in
dependently at least 4times in European populations of Mycosphaerella graminicola. Pest
Management Science, 65(2), 155–162.
Torriani S.F.F., Melichar J.P.E., Mills C., Pain N., Sierotzki H., et Courbot M. 2015.
Zymoseptoria tritici: A major threat to wheat production, integrated approaches to
control. Fungal Genetics and Biology 79: 8–12.
Tunisian Ministry of Agriculture Water Resources and Fisheries. 2015. Statistics Service of the
General Direction of Agricultural Production, Ministry of Agriculture, Water Resources
and Fishing, Tunis, Tunisia.
Ünal G., Kayim M., Ay T., et Yones A. 2017. Biological Control Research Institute, Yüreğir,
Adana, Turkey. Turkish Journal of Agriculture and Forestry:1-9.
Waalwijk C., Mendes O., Verstappen E.C.P., de Waard M.A, et Kema G.H.J. 2002. Isolation
and characterization of the mating type idiomorphs from the wheat septoria leaf blotch
fungus Mycosphaerella graminicola. Fungal Genetics and Biology, 35 :277-286.
Valade R., Vitry C., Leilte S., Stragaliati J., Senellart P., et al. 2010. Constitution d’une
mycothèque des champignons pathogènes du blé tendre et mise au point d’outils
permettant la caractérisation et la quantification de ces espèces. FSOV :1-13.
Väli Ü., Einarsson A., Waits L., & Ellegren H. 2008. To what extent do microsatellite markers
reflect genome‐wide genetic diversity in natural populations? Molecular ecology, 17(17),
3808-3817.
Van Beuningen L.T., et Kohli M.M. 1990. Deviation from the regression of infection heading
and height as a measure of resistance to septoria tritici blotch in wheat. Plant dis.74
(7):488-493.
Ware B.S. 2006. Aspects of sexual reproduction in Mycosphaerella species on wheat and barley
genetic studies on specificity, mapping, and fungicide resistance. Thèse de Doctorat,
Wageningen niversity.
Welch T., Feechan, A., et Kildea, S. 2017. Effect of host resistance on genetic structure of core
and accessory chromosomes in Irish Zymoseptoria tritici. Eur. J. Plant Pathology. DOI
10.1007/s10658-017-1259-9.
Yahyaoui A., Hakim S., Al-Naimi M., Nachit M.M .2000. Multiple disease resistance in durum
wheat (Triticum turgidum L. var. durum). p. 387-392. In Royo C, Nachit M, Di Fronzo
N, Araus JL (eds.). Durum wheat improvement in the Mediterranean region: New

Références
110
challenges. CIHEAM, Zaragoza, Spain. Options Méditerranéennes. Série A : Séminaires
Méditerranéens ; n. 40.
Zadoks J.C., Chang T.T, Konzak C.F. 1974. “A decimal code for the growth stages of cereals,”
Weed Research, 14 (6): 415-421.
Zapata L., Pena Chocarro L., PerezJorda G., Stika H.P. 2004. Early Neolithic Agriclture in the
Iberian Peninsula. Journal of world Perihistory 18:283-325
Zahri S., Farih A, Douira A. 2014. Statut des principales maladies cryptogamiques foliaires du
blé au Maroc en 2013. J. Appl. Biosci: 6543-6549.
Zahri S., Farih A., Badoc A., Douira A. 2008. Importance des septorioses dans les champs de
blés Marocains. Bull. Soc. Pharm. Bordeaux, 147: 29-38
Zhan J., Mundt C.C., Mc Donald B.A. 1998. Measuring immigration and sexual reproduction
in field of Mycosphaerella graminicola. Phytopathology. 88:1330-1337.
Zhan J., Mundt C., Hoffer M., & McDonald B.A. 2002. Local adaptation and effect of host
genotype on the rate of pathogen evolution: An experimental test in a plant pathosystem.
Journal of Evolutionary Biology, 15(4): 634-647.
Zhan J., Pettway R.E., McDonald BA. 2003. The global genetic structure of the wheat pathogen
Mycosphaerella graminicola is characterized by high nuclear diversity, low
mitochondrial diversity, regular recombination, and gene flow. Fungal Gene. Biol.
38:286-297.
Zhan J., et McDonald B.A. 2004. Global genetic structure in natural population of
Mycosphaerella graminicola. Phytopathology, 91:12-14.
Zhang X., Haley S.D, Jin Y. 2001. Inheritance of Septoria Tritici Blotch Resistance in Winter
Wheat. Crop Sciences, 41:323–326.

Productions Scientifiques
111
Productions scientifiques
Articles scientifiques
Bel Hadj Chedli, R., Ben M’Barek, S., Yahyaoui, A., Kehel, Z., Rezgui, S.2018. Occurrence
of Septoria tritici blotch (Zymoseptoria tritici ) disease on durum wheat, triticale, and bread
wheat in Northern Tunisia. Chliean Journal of Agricultural Research, 78(4):559-568. (Publié)
Bel Hadj Chedli R., Aouini, L., Ben M’Barek S., Yahyaoui, A., Rezgui, S., et Chaabène, H.
2019. Distribution of mating types in Zymoseptoria tritici populations collected from bread
wheat at El Haouaria region. Journal of New sciences, 61(5) : 3892-3898. (Publié)
Bel Hadj Chedli R., Ben M’Barek S., Souissi, A., Yahyaoui, A., Rezgui, S., et Chaabène, H.
2019. Screening for resistance of Tunisian, Moroccan and Algerian wheat cultivars to
Zymoseptoria tritici in Northern Tunisia. (Soumis dans: Journal of Plant Pathology)
Bel Hadj Chedli R., Aouini L., Ben M’Barek S., Bochra Bahri A., Verstappen E, Gerrit K.H.J.,
Rezgui S, Yahyaoui A., Chaabane H. 2019. Genetic differentiation between ‘Zymoseptoria
tritici’ populations sampled from bread wheat in Tunisia revealed by SSR markers. (Soumis
dans: European Journal of Plant Pathology)
Conférences:
Bel Hadj Chedli R., Akrouti, W., Yahyaoui, A., Rezgui, S. Incidence evaluation of
Zymoseptoria tritici in treated and untreated durum wheat fields in Northern Tunisia.
13thEuropean conférence on fungal genetics ECFG13, Paris, April 3-6,2016.
Bel Hadj Chedli R., Yahyaoui, A., Rezgui, S. Distribution de la septoriose chez le blé tendre
selon les étages bioclimatiques en Tunisie. Conférence scientifique internationale sur
l’environnement et l’agriculture, Hammamet, 24 et 25 Avril 2017.
Bel Hadj Chedli R., Aouini, L., Ben M’Barek S., Bahri, B.A., Els, V., Gerrit, K.H.J.,Yahyaoui,
A., Rezgui, S., et Chaabène, H. 2019. High genetic diversity among Zymoseptoria tritici isolates
from bread wheat in Northern Tunisia. The International Symposium on Cereal Leaf Blights
2019 in University College Dublin 22 -24 May 2019.

Annexes
112
Annexes
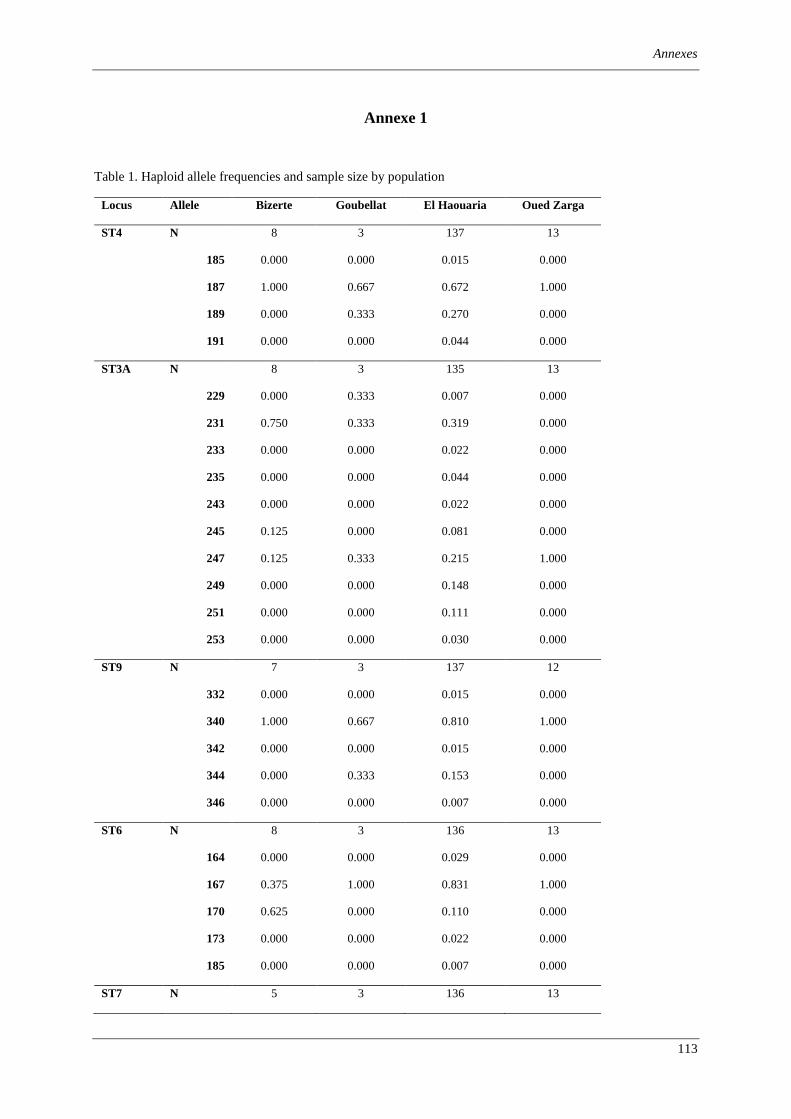
Annexes
113
Annexe 1
Table 1. Haploid allele frequencies and sample size by population
Locus Allele Bizerte Goubellat El Haouaria Oued Zarga
ST4 N 8 3 137 13
185 0.000 0.000 0.015 0.000
187 1.000 0.667 0.672 1.000
189 0.000 0.333 0.270 0.000
191 0.000 0.000 0.044 0.000
ST3A N 8 3 135 13
229 0.000 0.333 0.007 0.000
231 0.750 0.333 0.319 0.000
233 0.000 0.000 0.022 0.000
235 0.000 0.000 0.044 0.000
243 0.000 0.000 0.022 0.000
245 0.125 0.000 0.081 0.000
247 0.125 0.333 0.215 1.000
249 0.000 0.000 0.148 0.000
251 0.000 0.000 0.111 0.000
253 0.000 0.000 0.030 0.000
ST9 N 7 3 137 12
332 0.000 0.000 0.015 0.000
340 1.000 0.667 0.810 1.000
342 0.000 0.000 0.015 0.000
344 0.000 0.333 0.153 0.000
346 0.000 0.000 0.007 0.000
ST6 N 8 3 136 13
164 0.000 0.000 0.029 0.000
167 0.375 1.000 0.831 1.000
170 0.625 0.000 0.110 0.000
173 0.000 0.000 0.022 0.000
185 0.000 0.000 0.007 0.000
ST7 N 5 3 136 13
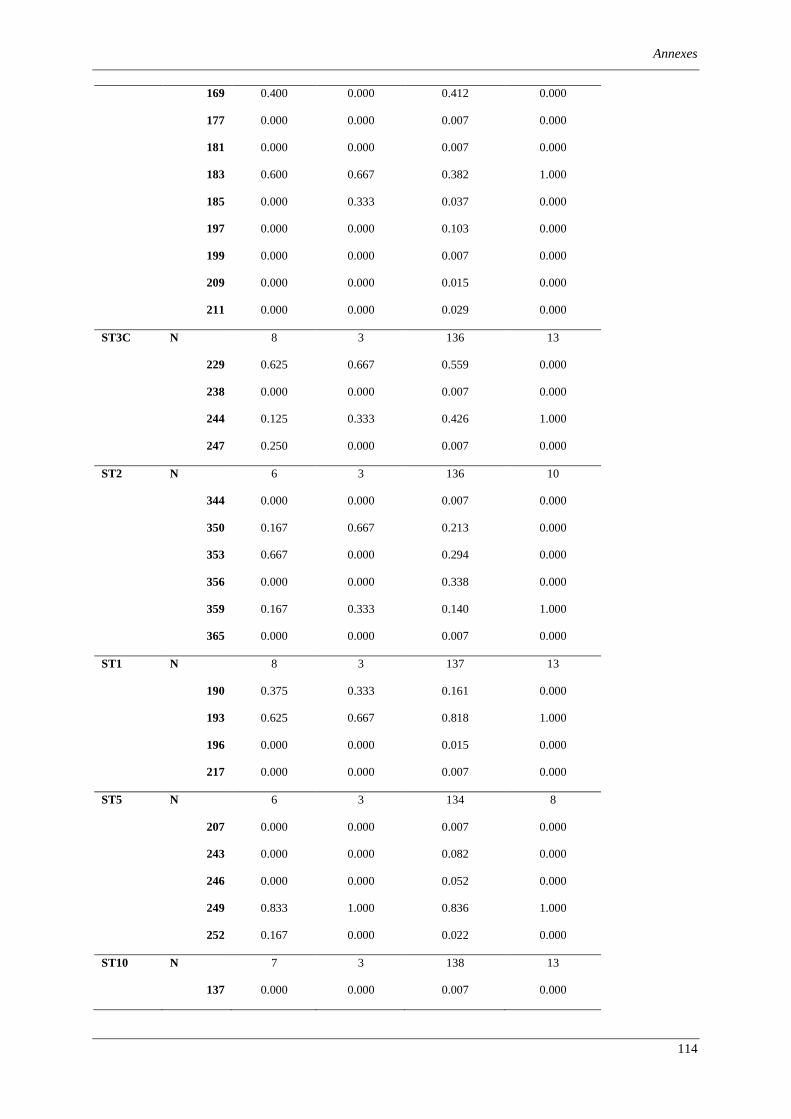
Annexes
114
169 0.400 0.000 0.412 0.000
177 0.000 0.000 0.007 0.000
181 0.000 0.000 0.007 0.000
183 0.600 0.667 0.382 1.000
185 0.000 0.333 0.037 0.000
197 0.000 0.000 0.103 0.000
199 0.000 0.000 0.007 0.000
209 0.000 0.000 0.015 0.000
211 0.000 0.000 0.029 0.000
ST3C N 8 3 136 13
229 0.625 0.667 0.559 0.000
238 0.000 0.000 0.007 0.000
244 0.125 0.333 0.426 1.000
247 0.250 0.000 0.007 0.000
ST2 N 6 3 136 10
344 0.000 0.000 0.007 0.000
350 0.167 0.667 0.213 0.000
353 0.667 0.000 0.294 0.000
356 0.000 0.000 0.338 0.000
359 0.167 0.333 0.140 1.000
365 0.000 0.000 0.007 0.000
ST1 N 8 3 137 13
190 0.375 0.333 0.161 0.000
193 0.625 0.667 0.818 1.000
196 0.000 0.000 0.015 0.000
217 0.000 0.000 0.007 0.000
ST5 N 6 3 134 8
207 0.000 0.000 0.007 0.000
243 0.000 0.000 0.082 0.000
246 0.000 0.000 0.052 0.000
249 0.833 1.000 0.836 1.000
252 0.167 0.000 0.022 0.000
ST10 N 7 3 138 13
137 0.000 0.000 0.007 0.000
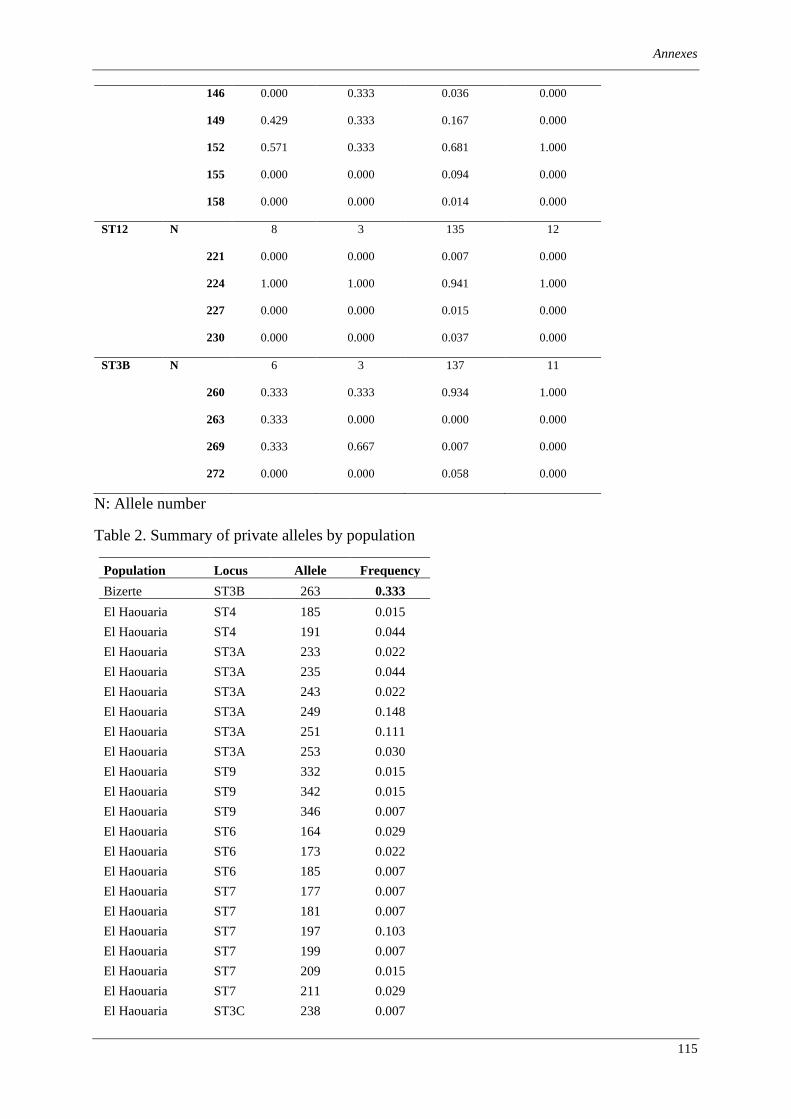
Annexes
115
146 0.000 0.333 0.036 0.000
149 0.429 0.333 0.167 0.000
152 0.571 0.333 0.681 1.000
155 0.000 0.000 0.094 0.000
158 0.000 0.000 0.014 0.000
ST12 N 8 3 135 12
221 0.000 0.000 0.007 0.000
224 1.000 1.000 0.941 1.000
227 0.000 0.000 0.015 0.000
230 0.000 0.000 0.037 0.000
ST3B N 6 3 137 11
260 0.333 0.333 0.934 1.000
263 0.333 0.000 0.000 0.000
269 0.333 0.667 0.007 0.000
272 0.000 0.000 0.058 0.000
N: Allele number
Table 2. Summary of private alleles by population
Population Locus Allele Frequency
Bizerte ST3B 263 0.333
El Haouaria ST4 185 0.015
El Haouaria ST4 191 0.044
El Haouaria ST3A 233 0.022
El Haouaria ST3A 235 0.044
El Haouaria ST3A 243 0.022
El Haouaria ST3A 249 0.148
El Haouaria ST3A 251 0.111
El Haouaria ST3A 253 0.030
El Haouaria ST9 332 0.015
El Haouaria ST9 342 0.015
El Haouaria ST9 346 0.007
El Haouaria ST6 164 0.029
El Haouaria ST6 173 0.022
El Haouaria ST6 185 0.007
El Haouaria ST7 177 0.007
El Haouaria ST7 181 0.007
El Haouaria ST7 197 0.103
El Haouaria ST7 199 0.007
El Haouaria ST7 209 0.015
El Haouaria ST7 211 0.029
El Haouaria ST3C 238 0.007

Annexes
116
El Haouaria ST2 344 0.007
El Haouaria ST2 356 0.338
El Haouaria ST2 365 0.007
El Haouaria ST1 196 0.015
El Haouaria ST1 217 0.007
El Haouaria ST5 207 0.007
El Haouaria ST5 243 0.082
El Haouaria ST5 246 0.052
El Haouaria ST10 137 0.007
El Haouaria ST10 155 0.094
El Haouaria ST10 158 0.014
El Haouaria ST12 221 0.007
El Haouaria ST12 227 0.015
El Haouaria ST12 230 0.037
El Haouaria ST3B 272 0.058

Annexes
117
Annexe 2
Iseptons Stb genes
1 Stb 1- Stb 4- Stb 10- Stb 17
2 Stb 1- Stb 7- Stb 10- Stb 17
3 Stb 1- Stb 9- Stb 15- Stb 17
4 Stb 7- Stb 10- Stb 17
5 Stb 9- Stb 10- Stb 17
6 Stb 4- Stb 10
7 Stb 10
8 Stb 7- Stb 9
9 Stb 7- Stb 8-Stb 9- Stb 17
10 Stb 4- Stb 8-Stb 9- Stb 11
11 Stb 7
12 Stb 1- Stb 4- Stb 7- Stb 8- Stb17
13 Stb 4- Stb 7- Stb 8- Stb 13
14 Stb 4- Stb 7- Stb 8
15 Stb 4- Stb 7
15 Stb 9
16 Stb 1- Stb 9
17 Stb 1- Stb 7- Stb 17
18 Stb 11- Stb 17
19 Stb 10- Stb 11- Stb 17
20 Stb 7- Stb 17
21 Stb 3- Stb 7- Stb 17
22 Stb 11- Stb 17
23 Stb 1- Stb 4- Stb 17
24 Stb 1- Stb 4- Stb 9- Stb 17
25 Stb 1- Stb 9
26 Stb 4
27 Stb 7
28 Stb 7- Stb 10- Stb 17
29 Stb 3- Stb 7- Stb 8- Stb 17
30 Stb 1- Stb 3- Stb 4- Stb 7- Stb 9- Stb
17
31 Stb 4- Stb 9- Stb 17
32 Stb 4- Stb 7- Stb 9- Stb 11-Stb17
33 Stb 1- Stb 4- Stb 7- Stb 10- Stb 17
34 Stb 4- Stb 7- Stb 8

Annexes
118
35 Stb 4- Stb 7- Stb 8
36 Stb 3- Stb 7- Stb 9
37 Stb 3- Stb 4- Stb 7- Stb 17
38 Stb 4- Stb 6- Stb 17
39 Stb 3- Stb 4- Stb 7- Stb 17
40 Stb 3- Stb 4- Stb 7 Stb 10- Stb 17
41 Stb 9- Stb 11
42 Stb 4- Stb 7- Stb 9- Stb 17
43 Stb 1- Stb 4- Stb 7- Stb 8-
44 Stb 1- Stb 4- Stb 7- Stb 10- Stb 17
45 Stb 4- Stb 7- Stb 8- Stb 9- Stb 17
46 Stb 4- Stb 7- Stb 8- Stb 9- Stb 10
47 Stb 9- Stb 10
48 Stb 1- Stb 7
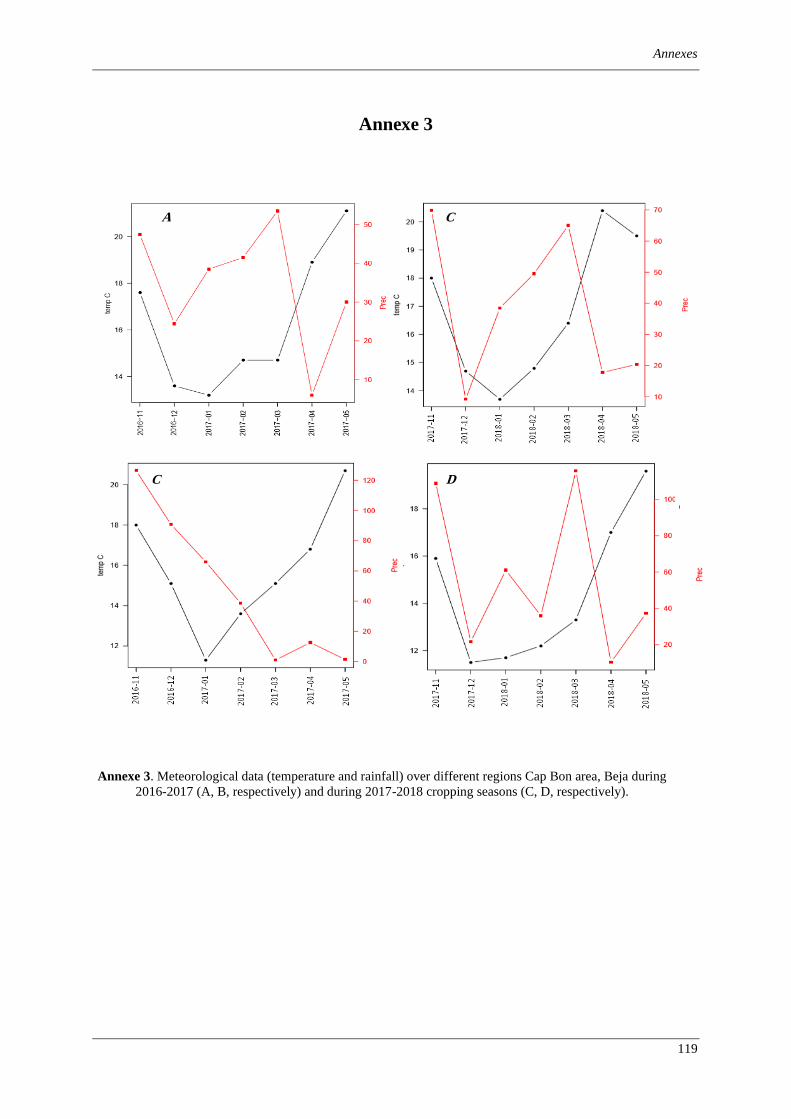
Annexes
119
Annexe 3
Annexe 3. Meteorological data (temperature and rainfall) over different regions Cap Bon area, Beja during
2016-2017 (A, B, respectively) and during 2017-2018 cropping seasons (C, D, respectively).
A C
C D

Annexes
120

559CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
RESEARCH
Occurrence of Septoria tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and bread wheat in Northern TunisiaRim Bel Hadj Chedli1, Sarrah Ben M’Barek2, Amor Yahyaoui3*, Zakaria Kehel4, and Salah Rezgui1
1National Agronomic Institute of Tunisia (INAT), 43 Avenue Charles Nicolle, 1002 Tunis, Tunisia.2Regional Field Crop Research Center of Beja (CRRGC) BP 350, 9000 Beja, Tunisia.3Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), km 45 Carretera México-Veracruz El Batán, Texcoco, Estado de México, México. *Corresponding author ([email protected]).4International Center for Agricultural Research in the Dry Areas (ICARDA), Rue Hafiane Cherkaoui, Agdal Rabat Po Box 6299 PC: 10112, Morocco
Received: 13 August 2018; Accepted: 19 October 2018; doi:10.4067/S0718-58392018000400559
ABSTRACT
Wheat (Triticum aestivum L.) is the most important cereal crop in Tunisia, nonetheless production is highly affected by drought and diseases mainly Septoria tritici blotch (STB) caused by Zymoseptoria tritici (Roberge ex Desm.) Quaedvl. & Crous anamorph and Mycosphaerella graminicola (Fuckel) J. Schröt. 1894 teleomorph; that has become an inherent disease of durum wheat (Triticum turgidum L. subsp. durum (Desf.) van Slageren) but rarely observed on bread wheat (Triticum aestivum L. subsp. aestivum) and on triticale (×Triticosecale spp.) The main objective of this work was to study the prevalence and geographical distribution of Z. tritici on triticale, durum wheat and particularly on bread wheat in different cereal growing regions of North and Northwestern Tunisia to confirm its presence/absence on bread wheat. For this study, 126 wheat fields were surveyed in North and Northwestern Tunisia during 2015-2016 and 2016-2017 cropping seasons. STB on durum wheat was present in the majority of inspected durum wheat fields, where high mean incidence (60%) and severity (40%) were recorded at Jendouba, Bizerte, Beja, and Kef. The survey data revealed low risk on bread wheat with an incidence of 23% and 29% at Bizerte and Beja, respectively. However high incidence of 84% and 52% was recorded at Cap Bon in 2016 and 2017, respectively and mainly at El Haouaria where STB severity was relatively high on bread wheat landrace of unknown origin but called by local farmers as ‘Farina arbi’. Sporadic incidence was recorded on Triticale of 100% at Jendouba (in 2016), and 33% at Bizerte (in 2016 and 2017) and absence at all other surveyed sites; likewise for severity at same locations where 13% and 42% were recorded in 2016. The survey data revealed low risk on bread wheat except at El Haouaria where STB severity was relatively high on a bread wheat landrace; while it was considered as high risk at all durum wheat fields in Beja, Bizerte, Jendouba, Zaghouan and Kef regions, such distinct occurrence could lead to clarify host specificity in Z. tritici.
Key words: Farina Arbi, survey, Triticum, Tunisia, wheat, Zymoseptoria.
INTRODUCTION
The cereal sector is of major economic importance in Tunisia. It provides major staple food commodities for most Tunisian households. Cereals are cultivated on almost one third of agricultural land (1.5 million hectares) (Tunisian Ministry of Agriculture, Water Resources and Fishing, 2015), 58% are located in the Northern and Western regions where durum wheat (Triticum turgidum L. subsp. durum (Desf.) van Slageren) represents 54%, against 36% for bread wheat (Triticum aestivum L. subsp. aestivum) and 10% for barley (FAO, 2017). Average production is around 1.05 million tons, which

560CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
represent approximately 80% of the country needs (Gharbi et al., 2000). However, cereal production in Tunisia faces many challenges of which drought is the most limiting abiotic stress in semi arid zones (Slama et al., 2005), while biotic stress, mainly leaf rust and Septoria tritici blotch (STB), cause important yield losses particularly on durum wheat in sub-humid regions (Ben Mohamed et al., 2000; Gharbi et al., 2000) of North and Northwestern Tunisia. STB caused by the ascomycete fungus Zymoseptoria tritici (Roberge ex Desm.) Quaedvl. & Crous became more important in Tunisia since the introduction of early maturing, semi dwarf, high yielding varieties. It has become an inherent disease of durum wheat, and thus a significant challenge for breeders to release varieties which combine good resistance and higher yields (Ammar et al., 2011). In contrast to durum wheat, bread wheat varieties grown in Tunisia are almost indemn of Septoria. High incidence of STB on durum compared to bread wheat in Tunisia suggests either an adaptation of Z. tritici isolates to durum rather than bread wheat (Yahyaoui et al., 2000) or high levels of resistance in bread wheat. The observed levels of resistance amongst cultivated bread wheat varies from year to year, most likely based on the environmental conditions and the dynamics of STB populations. Although Septoria was observed on durum wheat annually, up to now not much is known on the occurence of STB on bread wheat. Hence, the main objective of this paper was to study the prevalence and geographical distribution of Z. tritici on Triticum species and particularly on T. aestivum in different cereal growing regions of North and Northwestern Tunisia to eventually confirm its presence/absence on bread wheat.
MATERIALS AND METHODS
Study area description and climatic conditions of the surveyed regionsSurveys were conducted during two cropping seasons (2015-2016; 2016-2017) at seven major wheat-growing areas in North and Northwestern Tunisia (Figure 1). Fifty-seven fields were surveyed in Cap Bon North (El Haouaria), Cap Bon South, Bizerte, Manouba, Beja, and Jendouba during 2016 and sixty-nine fields were surveyed in Cap Bon regions, Bizerte, Manouba, Beja, Jendouba, Zaghouan, and Kef during 2017 (Figure 1). Certain varieties were more predominant than others rendering therefore inter region comparison rather difficult to make. Meteorological data (temperature and rainfall, Table 1) and geographical coordinates (altitude, longitude and latitude, Table 2) over different climatic regions for each survey areas were recorded. The average rainfall varied from 500 to 800 mm and the temperature ranged between 6 and 33 °C in the sub-humid region (Cap Bon North, Bizerte and Beja). Precipitation and temperature rates varied from 400 to 600 mm and from 5 to 37 °C respectively in the semi-arid regions (Cap Bon south, Manouba, Zaghouan, Jendouba, and Kef).
Figure 1. Map of Tunisia showing the location of survey areas across different climatic regions during 2016 and 2017 cropping seasons. Sub-humid: Cap Bon North (A), Bizerte (B) and Beja (C). Semi-arid region of Northern Tunisia: Cap Bon South (D), Manouba (E), Zaghouan (F), Jendouba (G), and El Kef (H).
561CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
Cereal crops The survey covered the major cereal growing areas in North and Northwestern Tunisia where commercial durum wheat varieties (‘Karim’, ‘Razzak’, ‘Maali’) occupy over 60% of the area compared to the introduced varieties (‘Saragola’, ‘Carioca’, ‘Sculpture’, ‘Soudaine’) that cover so far less than 10%. Commercial bread wheat varieties (‘Salambo’, ‘Utique’, ‘Haidra’) and introduced varieties (‘Zanzibar’) occupy no more than 20% of the area; while triticale (‘Bienvenue’ and others) covers about 1%-3%; the rest of the area is covered by barley and oats. A unique situation in Cap Bon region where a landrace bread wheat (‘Farina arbi’) occupies over 60% of the area, the rest is covered by commercial barley, durum and bread wheat varieties. ‘Farina arbi’, a tall low yielding bread wheat (landrace) of unknown origin is cultivated annually for over a century, according to local farmers, in the same region and exclusively used for pastry known as “Kaak”. The seed is maintained by local farmers and not commercialized. Bread wheat landrace (‘Farina arbi’) and Septoria tritici blotch (STB) differentials (comprised within CIMMYT’s ISEPTON) were phenotyped at experimental station of Bou Salem (Tunisia) under artificial inoculation with Zymoseptoria tritici ([Roberge ex Desm.)] Quaedvl. & Crous) populations sampled from durum wheat. Inoculation was performed at tillering stage using bulk isolates at a rate of 106 spores mL-1 according to Ferjaoui et al. (2015) with slight modifications.
Septoria leaf blotch disease assessmentField surveys were conducted during flowering stage of bread and durum wheat, each field was visited once. STB prevalence was assessed within and between regions based on number of fields surveyed and the presence/absence of Septoria at each location. The incidence was reported on this study based on Saari-Prescott modified “0-9” Cobb-scale (Saari and Prescott, 1975). In this survey, we designed five field classes (Table 3) to assess STB within each region where prevalence, severity and incidence were the main criteria. The relative importance of STB was based on its prevalence at each location where incidence and severity were assessed and averaged at each surveyed field. In this study, prevalence
mm °CCap Bon North1 500-800 9-31Cap Bon South2 400-500 7-34Bizerte 600-800 7-33Beja 500-600 6-32Manouba 400-600 8-34Zaghouan 400-600 4-34Jendouba 400-500 5-37El Kef 300-400 2-38
Rainfall
1Cap Bon North: El Haouaria.2Cap Bon South: Soliman, Beni Khalled, and Grombalia.
Table 1. Climatic conditions of inspected regions during the survey period.
Regions
Weather conditions (range)
Temperature (min-max)
Table 2. Geographical coordinates of inspected regions during the survey period.
m a.s.l. Bizerte 16-354 09°03’1” 09°69’83” 32°50’56” 37°14’12”Cap bon North1 11-876 10°02’73” 10°10’95” 36°52’41” 36°67’82”Cap bon South2 24-446 10°45’27” 10°49’10” 36°47’47” 36°92’68”Manouba 58-244 09°59’1” 09°91’41” 36°30’70” 36°85’04”Zaghouan 81-320 09°45’43” 10°4’42” 36°21’18” 36°30’53”Beja 18-290 09°09’01” 10°78’62” 36°22’26” 36°68’31”Jendouba 21-341 08°32’35” 08°42’45” 36°28’4” 36°32’48”Kef 100-327 08’39°12” 08°48’50” 36°22’26” 36°11’44”
TreatmentsAltitude(range)
Longitude (N)(range)
Latitude (E)(range)
1Cap Bon North: El Haouaria.2Cap Bon South: Soliman, Beni Khalled, and Grombalia.m a.s.l.: Meters above sea level.
562CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
indicates how wide spread is STB, whereas incidence conveys information on the risk of the disease within a severity range. In the survey protocol of the CIMMYT septoria phenotyping platform four classes (Class I-V) were adopted. Class I: Low prevalence (%), severity (0-9), and low incidence indicating insignificant risk. Class II: low prevalence and incidence indicating low risk. Class III: moderate prevalence and incidence indicating moderate risk to be monitored. Class IV: fields where STB was wide spread and apparent high severity observed at flag leaf, indicating high risk of the disease. Class V: includes fields heavily infested by STB; situation where the disease is obvious at each field surveyed and where the severity is at its most, i.e. severe symptom on flag leaf and spikes, this is a situation where STB is a high risk. Relevant agronomic data such as variety name, sowing date, fertilization, crop density and spatial pattern and previous crops were recorded. Altitude, longitude and latitude were also recorded using Global Positioning System (GPS).
Data analysisLinear mixed model was used to analyze disease data (incidence and severity) collected during the survey using ASReml-R software (Gilmour et al., 2002). The years, regions, species, varieties together with their interactions were assumed to be fixed.
RESULTS
Incidence of wheat Septoria tritici blotchSurvey results showed that STB incidence on bread wheat was very limited across surveyed areas in Northern Tunisia Triticum species and wheat varieties identified during the survey during the two cropping seasons are presented in Table 4. STB prevalence during the two cropping seasons (2015-2016 and 2016-2017) was insignificant to low on commercial bread wheat varieties in the majority of the surveyed regions (Tables 5 and 6). It was ranked as class I or II except at
I 0-10 0-3 0-2 InsignificantII 10-20 3-5 2-3 LowIII 20-40 5-6 3-5 ModerateIV 40-60 6-8 5-7 HighV 60-100 8-9 7-9 Severe
Prevalence1
%
1(Number of infected fields (STB present)/total number of fields surveyed) × 100.2H: Level of plant infection.3Percentage of STB within each class level at surveyed field.
Table 3. Survey designated Classes for Septoria tritici blotch (STB) prevalence, severity and incidence.
Class
Severity2
(Cobb-Scale: 0-9)
SeverityH Incidence3
Table 4. Triticum species and wheat varieties identified during the survey during the two cropping seasons.
Bread wheat Zanzibar Bizerte Utique Bizerte, Beja, Zaghouan Haïdra Bizerte, Beja, Zaghouan Vaga Bizerte, Jendouba Salambo Cap Bon North (El Haouaria), Jendouba Bread wheat landrace Cap Bon North (El Haouaria)Durum wheat Karim Bizerte, Beja, Jendouba, Manouba, Cap Bon North, Cap Bon South, Kef Maali Bizerte, Beja, Cap Bon North, Cap Bon South, Jendouba, Manouba, Kef Razzak Bizerte, Beja, Cap Bon North, Cap Bon South, Jendouba, Zaghouan, Kef Monastir Cap Bon North Carioca Bizerte, Jendouba, Zaghouan Saragolla Bizerte Soudaine Bizerte Sculpture Bizerte, Beja, JendoubaTriticale TL4 Bizerte, Cap Bon North, Bienvenue Bizerte, Manouba
Species Varieties Surveyed regions
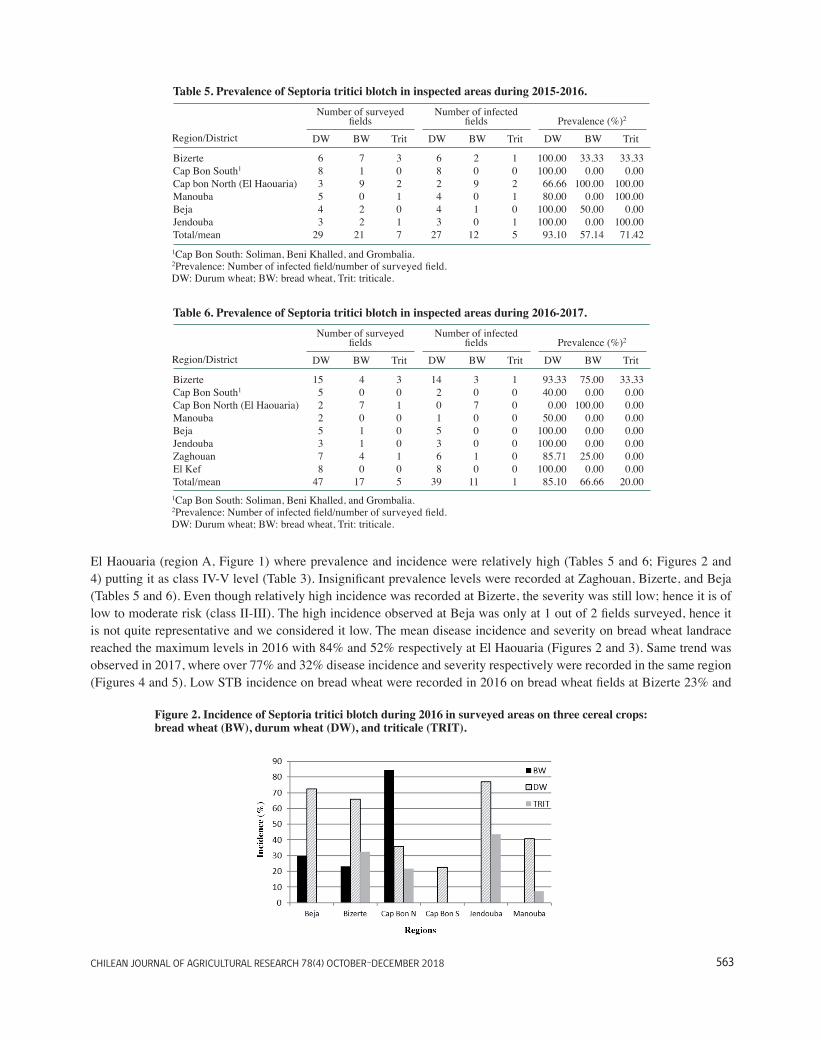
563CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
El Haouaria (region A, Figure 1) where prevalence and incidence were relatively high (Tables 5 and 6; Figures 2 and 4) putting it as class IV-V level (Table 3). Insignificant prevalence levels were recorded at Zaghouan, Bizerte, and Beja (Tables 5 and 6). Even though relatively high incidence was recorded at Bizerte, the severity was still low; hence it is of low to moderate risk (class II-III). The high incidence observed at Beja was only at 1 out of 2 fields surveyed, hence it is not quite representative and we considered it low. The mean disease incidence and severity on bread wheat landrace reached the maximum levels in 2016 with 84% and 52% respectively at El Haouaria (Figures 2 and 3). Same trend was observed in 2017, where over 77% and 32% disease incidence and severity respectively were recorded in the same region (Figures 4 and 5). Low STB incidence on bread wheat were recorded in 2016 on bread wheat fields at Bizerte 23% and
1Cap Bon South: Soliman, Beni Khalled, and Grombalia.2Prevalence: Number of infected field/number of surveyed field.DW: Durum wheat; BW: bread wheat, Trit: triticale.
Table 5. Prevalence of Septoria tritici blotch in inspected areas during 2015-2016.
Bizerte 6 7 3 6 2 1 100.00 33.33 33.33Cap Bon South1 8 1 0 8 0 0 100.00 0.00 0.00Cap bon North (El Haouaria) 3 9 2 2 9 2 66.66 100.00 100.00Manouba 5 0 1 4 0 1 80.00 0.00 100.00Beja 4 2 0 4 1 0 100.00 50.00 0.00Jendouba 3 2 1 3 0 1 100.00 0.00 100.00Total/mean 29 21 7 27 12 5 93.10 57.14 71.42
Region/District Trit
Prevalence (%)2
BWDWTrit
Number of infected fields
BWDWTrit
Number of surveyed fields
BWDW
1Cap Bon South: Soliman, Beni Khalled, and Grombalia.2Prevalence: Number of infected field/number of surveyed field.DW: Durum wheat; BW: bread wheat, Trit: triticale.
Table 6. Prevalence of Septoria tritici blotch in inspected areas during 2016-2017.
Bizerte 15 4 3 14 3 1 93.33 75.00 33.33Cap Bon South1 5 0 0 2 0 0 40.00 0.00 0.00Cap Bon North (El Haouaria) 2 7 1 0 7 0 0.00 100.00 0.00Manouba 2 0 0 1 0 0 50.00 0.00 0.00Beja 5 1 0 5 0 0 100.00 0.00 0.00Jendouba 3 1 0 3 0 0 100.00 0.00 0.00Zaghouan 7 4 1 6 1 0 85.71 25.00 0.00El Kef 8 0 0 8 0 0 100.00 0.00 0.00Total/mean 47 17 5 39 11 1 85.10 66.66 20.00
Region/District Trit
Prevalence (%)2
BWDWTrit
Number of infected fields
BWDWTrit
Number of surveyed fields
BWDW
Figure 2. Incidence of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops: bread wheat (BW), durum wheat (DW), and triticale (TRIT).

564CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
Beja 29%, and in 2017 at Bizerte 17% and Zaghouan 5% (Figures 2 and 4). The severity percentages in these regions did not exceed 5% during the two cropping seasons (Figures 3 and 5). These data showed that STB level was at class I and II ratings except at El Haouaria, where it was rated class IV and V. Unlike the situation on bread wheat, STB was widely distributed on durum wheat and was highly prevalent at Bizerte, Beja, Jendouba, and El Kef where it ranked from class III to V (Table 3). The overall prevalence of the disease was about 50%, 85.71%, 93.3% at Manouba, Zaghouan, and Bizerte, respectively, in 2017 (Table 6). More than 65% and 47% of the disease incidence and severity respectively were recorded in the majority of prospected areas (Beja, Bizerte, Jendouba and
Figure 3. Severity of Septoria tritici blotch during 2016 in surveyed areas on three cereal crops: bread wheat (BW), durum wheat (DW), and triticale (TRIT).
Figure 4. Incidence of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops species: bread wheat (BW), durum wheat (DW), and triticale (TRIT).
Figure 5. Severity of Septoria tritici blotch during 2017 in surveyed areas on three cereal crops: bread wheat (BW), durum wheat (DW), and triticale (TRIT).
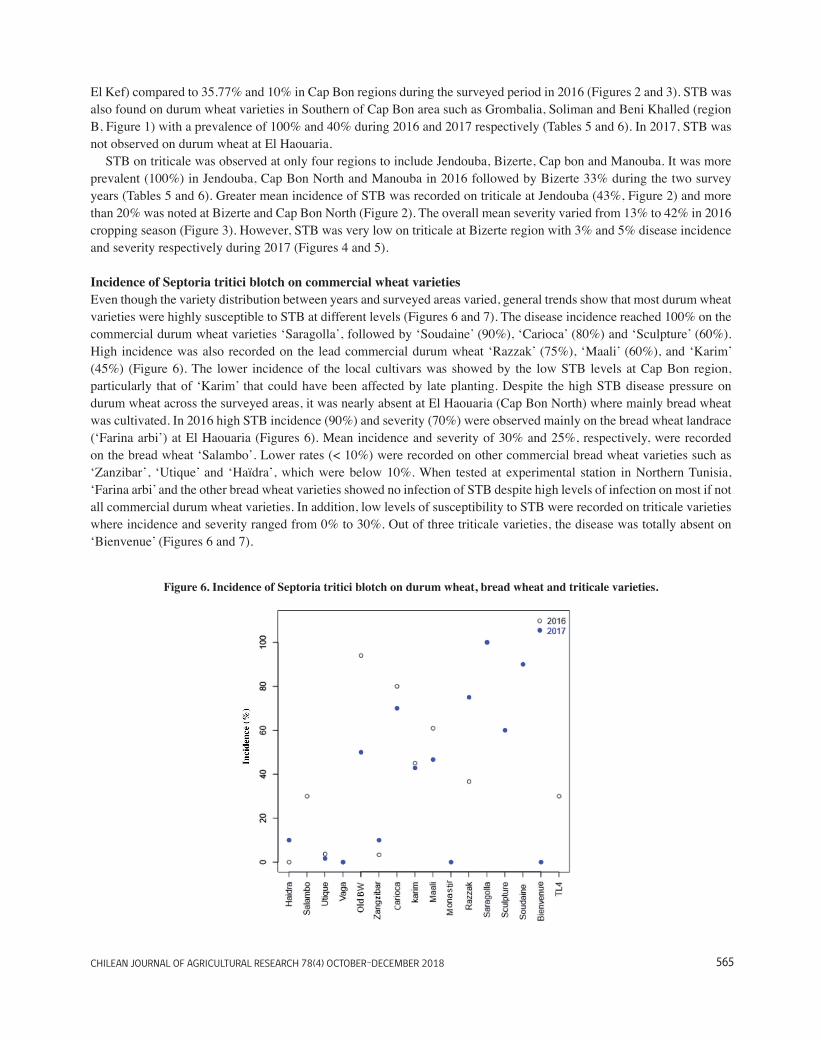
565CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
Figure 6. Incidence of Septoria tritici blotch on durum wheat, bread wheat and triticale varieties.
El Kef) compared to 35.77% and 10% in Cap Bon regions during the surveyed period in 2016 (Figures 2 and 3). STB was also found on durum wheat varieties in Southern of Cap Bon area such as Grombalia, Soliman and Beni Khalled (region B, Figure 1) with a prevalence of 100% and 40% during 2016 and 2017 respectively (Tables 5 and 6). In 2017, STB was not observed on durum wheat at El Haouaria. STB on triticale was observed at only four regions to include Jendouba, Bizerte, Cap bon and Manouba. It was more prevalent (100%) in Jendouba, Cap Bon North and Manouba in 2016 followed by Bizerte 33% during the two survey years (Tables 5 and 6). Greater mean incidence of STB was recorded on triticale at Jendouba (43%, Figure 2) and more than 20% was noted at Bizerte and Cap Bon North (Figure 2). The overall mean severity varied from 13% to 42% in 2016 cropping season (Figure 3). However, STB was very low on triticale at Bizerte region with 3% and 5% disease incidence and severity respectively during 2017 (Figures 4 and 5).
Incidence of Septoria tritici blotch on commercial wheat varietiesEven though the variety distribution between years and surveyed areas varied, general trends show that most durum wheat varieties were highly susceptible to STB at different levels (Figures 6 and 7). The disease incidence reached 100% on the commercial durum wheat varieties ‘Saragolla’, followed by ‘Soudaine’ (90%), ‘Carioca’ (80%) and ‘Sculpture’ (60%). High incidence was also recorded on the lead commercial durum wheat ‘Razzak’ (75%), ‘Maali’ (60%), and ‘Karim’ (45%) (Figure 6). The lower incidence of the local cultivars was showed by the low STB levels at Cap Bon region, particularly that of ‘Karim’ that could have been affected by late planting. Despite the high STB disease pressure on durum wheat across the surveyed areas, it was nearly absent at El Haouaria (Cap Bon North) where mainly bread wheat was cultivated. In 2016 high STB incidence (90%) and severity (70%) were observed mainly on the bread wheat landrace (‘Farina arbi’) at El Haouaria (Figures 6). Mean incidence and severity of 30% and 25%, respectively, were recorded on the bread wheat ‘Salambo’. Lower rates (< 10%) were recorded on other commercial bread wheat varieties such as ‘Zanzibar’, ‘Utique’ and ‘Haïdra’, which were below 10%. When tested at experimental station in Northern Tunisia, ‘Farina arbi’ and the other bread wheat varieties showed no infection of STB despite high levels of infection on most if not all commercial durum wheat varieties. In addition, low levels of susceptibility to STB were recorded on triticale varieties where incidence and severity ranged from 0% to 30%. Out of three triticale varieties, the disease was totally absent on ‘Bienvenue’ (Figures 6 and 7).
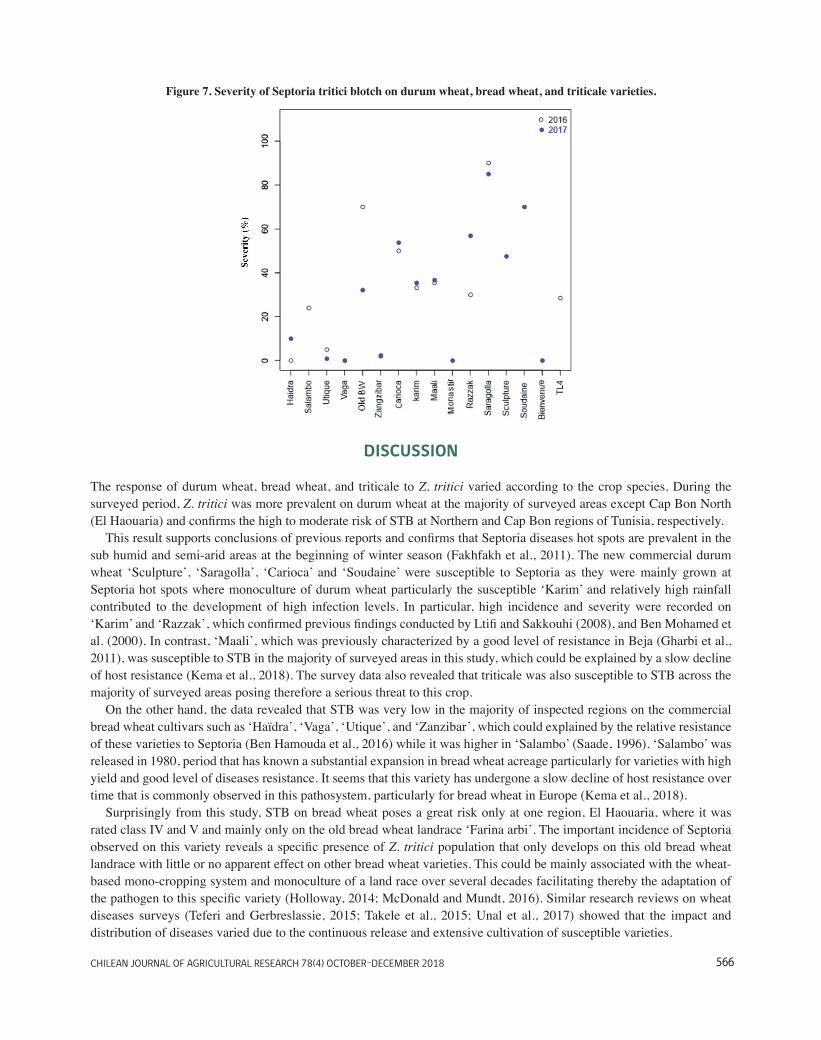
566CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
DISCUSSION
The response of durum wheat, bread wheat, and triticale to Z. tritici varied according to the crop species. During the surveyed period, Z. tritici was more prevalent on durum wheat at the majority of surveyed areas except Cap Bon North (El Haouaria) and confirms the high to moderate risk of STB at Northern and Cap Bon regions of Tunisia, respectively. This result supports conclusions of previous reports and confirms that Septoria diseases hot spots are prevalent in the sub humid and semi-arid areas at the beginning of winter season (Fakhfakh et al., 2011). The new commercial durum wheat ‘Sculpture’, ‘Saragolla’, ‘Carioca’ and ‘Soudaine’ were susceptible to Septoria as they were mainly grown at Septoria hot spots where monoculture of durum wheat particularly the susceptible ‘Karim’ and relatively high rainfall contributed to the development of high infection levels. In particular, high incidence and severity were recorded on ‘Karim’ and ‘Razzak’, which confirmed previous findings conducted by Ltifi and Sakkouhi (2008), and Ben Mohamed et al. (2000). In contrast, ‘Maali’, which was previously characterized by a good level of resistance in Beja (Gharbi et al., 2011), was susceptible to STB in the majority of surveyed areas in this study, which could be explained by a slow decline of host resistance (Kema et al., 2018). The survey data also revealed that triticale was also susceptible to STB across the majority of surveyed areas posing therefore a serious threat to this crop. On the other hand, the data revealed that STB was very low in the majority of inspected regions on the commercial bread wheat cultivars such as ‘Haïdra’, ‘Vaga’, ‘Utique’, and ‘Zanzibar’, which could explained by the relative resistance of these varieties to Septoria (Ben Hamouda et al., 2016) while it was higher in ‘Salambo’ (Saade, 1996). ‘Salambo’ was released in 1980, period that has known a substantial expansion in bread wheat acreage particularly for varieties with high yield and good level of diseases resistance. It seems that this variety has undergone a slow decline of host resistance over time that is commonly observed in this pathosystem, particularly for bread wheat in Europe (Kema et al., 2018). Surprisingly from this study, STB on bread wheat poses a great risk only at one region, El Haouaria, where it was rated class IV and V and mainly only on the old bread wheat landrace ‘Farina arbi’. The important incidence of Septoria observed on this variety reveals a specific presence of Z. tritici population that only develops on this old bread wheat landrace with little or no apparent effect on other bread wheat varieties. This could be mainly associated with the wheat-based mono-cropping system and monoculture of a land race over several decades facilitating thereby the adaptation of the pathogen to this specific variety (Holloway, 2014; McDonald and Mundt, 2016). Similar research reviews on wheat diseases surveys (Teferi and Gerbreslassie, 2015; Takele et al., 2015; Unal et al., 2017) showed that the impact and distribution of diseases varied due to the continuous release and extensive cultivation of susceptible varieties.
Figure 7. Severity of Septoria tritici blotch on durum wheat, bread wheat, and triticale varieties.

567CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
Thus, the magnitude of virulence and disease incidence are variable and closely related to the frequency of the variety used in a particular area as well as the proportion of durum wheat area as compared to that of bread wheat (Yahyaoui et al., 2000). Testing ‘Farina arbi’ land race for its resistance/susceptibility to Z. tritici at other Northern regions where durum wheat is mostly cultivated showed no STB infection. This unique bread wheat landrace, completely susceptible at El Haouaria (North eastern Tunisia) and completely resistant at Beja Northwestern Tunisia, could be that we are definitely dealing with two distinct Z. tritici populations and could give more highlight on STB specificity. Further studies will be conducted to characterize the STB populations from El Haouaria that are mostly specific to the bread wheat land races (‘Farina arbi’) and have no effect on other bread and durum wheat varieties. Such phenomenon has not been observed before and could lead to further understanding of STB host specificity.
CONCLUSIONS
The survey data revealed low risk of Zymoseptoria tritici on bread wheat except at Cap Bon region especially at El Haouaria where Septoria tritici blotch (STB) severity was relatively high on the old bread wheat landrace, while rare occurrence at other sites was observed on some commercial bread wheat varieties. Sporadic incidence and high severity were observed on triticale across the surveyed fields. Although Tunisia is primarily a durum-wheat producing country with Z. tritici being mostly prevalent on durum wheat; bread wheat is of great economic importance, even though it occupies small areas. The occurrence of STB on the landrace could lead to development of Septoria population that could become of major importance on bread wheat as is the case in Morocco and other regions. The presence of an STB population at one site and infecting a single cultivar will be further investigated and will possibly lead to better understanding of Z. tritici population dynamics that could become an important tool in screening for disease resistance.
ACKNOWLEDGEMENTS
This research was supported by “CRP WHEAT Tunisia Septoria Precision Phenotyping Platform”.
REFERENCES
Ammar, K., Gharbi, M.S., and Deghais, M. 2011. Wheat in Tunisia. p. 443-463. In Bonjean, A.P., Angus, W.J, and van Ginkel, M. (eds.) The world wheat book: A history of wheat breeding. Vol. 2. Lavoisier, Paris, France.
Ben Hamouda, M., Ben Youssef, S., Bedhiaf, S., Annabi, M., Draief, Z., and Gharbi, M.S. 2016. Status of agricultural innovations, innovation platforms, and innovations investment. Tunisia. 2015 PARI project country report: Republic of Tunisia. Forum for Agricultural Research in Africa (FARA), Accra, Ghana.
Ben Mohamed, L., Rouaissi, M., Sebei, A., Hamza, S., et Harrabi, M. 2000. Effet du génotype, de la date de semis, de la fertilisation azotée et potassique et des fongicides sur le développement de Septoria tritici. p. 350-356. In Royo, C., Nachit, M., Di Fronzo, N., and Araus, J.L. (eds.) Durum wheat improvement in the Mediterranean region: New challenges. CIEHAM, Zaragoza, Spain. (Options Méditerranéenne: Série A. Séminaires Méditerranéens; n. 40).
Fakhfakh, M.M., Rezgui, S., Jarrahi, T., Zouani, R., Bousalmi, A., Sgatni K., and Yahyaoui, A. 2011. Mapping distribution of Septoria allowed targeting management options of the disease. 8th International Symposium on Mycosphaerella and Stagonospora Diseases of Cereals, Mexico City. 10-14 September 2011. Book of Abstracts. Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), Mexico.
FAO. 2017. Country fact sheet on food and agriculture policy trends. Socio-economic context and role of agriculture. p. 1-6. Food and Agriculture Policy Decision Analysis (FAPDA), FAO, Rome, Italy. Available at http://www.fao.org/3/a-i7738e.pdf.
Ferjaoui, S., M’Barek, S.B., Bahri, B., Slimane, R.B. and Hamza, S. 2015. Identification of resistance sources to septoria tritici blotch in old durum wheat germplasm applied for the analysis of the Zymoseptoria tritici-durum wheat interaction. Journal of Plant Pathology 97(3):471-481.
Gharbi, M.S., Berraies, S., Ammar, K., and Yahyaoui, A. 2011. Delivering disease resistant cultivars to enhance sustainability of durum wheat production in Tunisia. p. 60. In Duveiller E., Singh, P.K., et al. (eds.) 8th International Symposium on Mycosphaerella and Stagonospora Diseases of Cereals, Mexico City. 10-14 September. Book of Abstracts. Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), Mexico.

568CHILEAN JOURNAL OF AGRICULTURAL RESEARCH 78(4) OCTOBER-DECEMBER 2018
Gharbi, M.S., Deghais, M., and Ben Amar, F. 2000. Breeding for resistance to Septoria tritici in durum wheat. p. 397-401. In Royo, C., Nachit, M., Di Fronzo, N., and Araus, J.L. (eds.) Durum wheat improvement in the Mediterranean region: New challenges. CIHEAM, Zaragoza, Spain. (Options Méditerranéennes. Série A: Séminaires Méditerranéens; n. 40).
Gilmour, R., Gogel, B.J., Cullis, B.R., Welham, S.J., and Thompson, R. 2002. ASReml user guide release 1.0. VSN International Ltd., Hemel Hempstead, UK.
Holloway, G. 2014. Septoria tritici blotch of wheat. DEPI Information Note Series May 2014. 2 p. Department of Environment and Primary Industries (DEPI), Melbourne, Australia.
Kema, G.H.J., Gohari, A.M., Aouini, L., Gibriel, H.A.Y., Ware, S.B., et al. 2018. Stress and sexual reproduction affect the dynamics of the wheat pathogen effector AvrStb6 and strobilurin resistance. Nature Genetics 50:375-380.
Ltifi, A., et Sakkouhi, L. 2008. La septoriose du blédur: Résistance variétale et impact de la maladie sur le rendement. Annales de l’INRAT 81:76-84.
McDonald, B.A., and Mundt, C.C. 2016. How knowledge of pathogen population biology informs management of Septoria tritici blotch. Phytopathology 106(9):948-955.
Saade, M.E. 1996. Adoption and impact of high yielding wheat varieties in Northern Tunisia. CIMMYT Economics Working Paper 96-03. Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), Mexico D.F.
Saari, E.E., and Prescott, J.M. 1975. A scale for appraising the foliar intensity of wheat diseases. Plant Disease Reporter 59:377-380. Slama, A., Ben Naceur, M., Ben Salem, M., et Zied, E. 2005. Les céréales en Tunisie: production, effet de la sécheresse et
mécanismes de résistance. Sécheresse 16(3):225-229.Takele, A., Lencho, A., Getaneh, W.A., Hailu, E., and Kassa, B. 2015. Status of wheat Septoria leaf blotch (Septaria tritici
Roberge in Desmaz) in South West and Western Shewa Zones of Oromiya Regional State, Ethiopia. Research in Plant Sciences 3(3):43-48.
Teferi, T.A., and Gebreslassie, Z.S. 2015. Occurrence and intensity of wheat Septoria tritici blotch and host response in Tigray, Ethiopia. Crop Protection 68:76-71.
Tunisian Ministry of Agriculture, Water Resources and Fishing. 2015. Statistics Service of the General Direction of Agricultural Production. Ministry of Agriculture, Water Resources and Fishing, Tunis, Tunisia.
Unal, G., Kayim, M., Ay, T., and Yones, M.A. 2017. Evaluation of disease intensity and molecular identification of Zymoseptoria tritici causing Septoria leaf blotch on wheat in the Eastern Mediterranean Region of Turkey. Turkish Journal of Agriculture and Forestry 41(6):405-413.
Yahyaoui, A., Hakim, S., Al-Naimi, M., and Nachit, M.M. 2000. Multiple disease resistance in durum wheat (Triticum turgidum L. var. durum). p. 387-392. In Royo, C., Nachit, M., Di Fronzo, N., and Araus, J.L. (eds.) Durum wheat improvement in the Mediterranean region: New challenges. CIHEAM, Zaragoza, Spain (Options Méditerranéennes. Série A: Séminaires Méditerranéens; n. 40).

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3892
Distribution of mating types in Zymoseptoria tritici populations
collected from bread wheat at El Haouaria region
Distribution des types sexuels dans une population de Zymoseptoria
tritici collectée du blé tendre dans la région d’El Haouaria
R. BELHADJ CHEDLI1,4*
, L. AOUINI 2, S. BEN M’BAREK
3,4, A. YAHYAOUI
4,5, S. REZGUI
1,
H. CHAABENE1
1 National Agronomic Institute of Tunisia (INAT), 43 Avenue Charles Nicolle, 1002 Tunis, Tunisia. 2 Department of Agronomy, Purdue University, 915 West State Street, West Lafayette, IN 47907 3 Regional Field Crops Research Center of Beja (CRRGC) BP 350, 9000 Beja, Tunisia. 4 CRP Wheat Septoria Phenotyping Platform, Tunisia. 5 Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), km 45 Carretera México-Veracruz
El Batán, Texcoco.
*Corresponding author: [email protected]
Abstract – Bread wheat is one of the most important cereal crop in Tunisia, nonetheless production is
highly affected by drought and diseases mainly Septoria tritici blotch (STB) caused by the fungus Zymoseptoria tritici. The main objective of this study is to assess the mating types distribution of 103
bread-wheat-derived Z. tritici isolates collected during 2015-2016 cropping season from El Haouaria
region where STB occurs annually on a bread wheat landrace that is called by farmers ‘Farina Arbi’. For this study, a multiplex PCR was conducted using two pairs of mating type-specific primers for
MAT1-1 and MAT1-2. The results showed that on average both mating types occurred with an equal
frequency of MAT1-1 and MAT1-2 (44.7% and 55.3% respectively), except for one field where the
MAT1-2 was predominant with 74%, which could be certainly related to the reduced population size in this field. Overall, the equal mating type distribution observed at El Haouaria region suggests a frequent
sexual reproduction of bread-wheat derived Z. tritici isolates in Tunisia.
Keywords: Farina Arbi, Septoria, sexual reproduction
Résumé - Le blé tendre est l’une des céréales les plus importantes en Tunisie. Sa production reste
hautement affectée par la sècheresse et les maladies fongiques notamment la septoriose, causée par le
champignon Zymoseptoria tritici qui attaque principalement le blé dur. L’objectif majeur de cette étude est la détermination de la distribution des deux mating types (MAT1-1 and MAT1-2) d’une population
de 103 isolats de Z. tritici colléctés à partir du blé tendre ancien ‘Farina Arbi’ au niveau de quatre
champs, dans la région d’El Haouaria là où une grande incidence de septoriose a été notée durant la
saison agricole 2015-2016. Une PCR multiplexe utilisant deux paires d’amorces spécifiques MAT 1-1 et MAT1-2 a été effectuée. Les résultats de cette étude ont montré une distribution égale des deux types
sexuels (44.7% vs 55.3%). Cependant, une prédominance du MAT1-2 avec 74% a été notée au sein d’un
seul champ, ce qui pourrait s’expliquer par la taille réduite de la population de Z. tritici dans ce champ. D’une façon générale, la distribution équilibrée des deux types sexuels observée dans la région d’El
Haouaria suggère une reproduction sexuée fréquente des isolats de blé tendre en Tunisie.
Mots clés : Farina Arbi, Septoriose, reproduction sexuée
1. Introduction
Vers la fin du XIXeme siècle, la culture du blé tendre (Triticum aestivum L. subsp. aestivum) en Tunisie
existait sous forme de mélange dans les champs avec l'orge (Hordeum vulgare), le blé dur cultivé (Triticum turgidum L. subsp. durum (Desf.)), et d’autres espèces reconnaissables par la forme, la
couleur et la taille des épis, et aussi par les barbes et les grains (Gharbi et al., 2013). A cette époque, les
agriculteurs ne connaissaient même pas les noms vernaculaires aidant à la distinction de différents

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3893
cultivars de blé tendre et de blé dur, et n’utilisaient ainsi que la dénomination générale ‘Farina’ et ‘Gameh’ pour les deux espèces respectivement. Ce n'est que depuis la colonisation française que la
culture du blé tendre a commencé à prendre de la place dans l’agriculture Tunisienne (Ammar et al.,
2011 ; El Falleh et al., 2014). Au début des années 1930-1940, un développement rapide et relativement considérable des emblavures
de blé tendre a eu lieu grâce à l’introduction de la variété Florence Aurore. Dotée d’un bon rendement,
d’une excellente adaptabilité à la sécheresse dans les zones semi-arides et d’une bonne résistance aux différentes maladies (Septoria, Oïdium, Rouille…), cette dernière a pris, progressivement de
l’importance dans tous les pays du bassin méditerranéen jusqu’à l’introduction du blé semi-nain vers la
fin des années 1960 par CIMMYT (Saade, 1996). Malgré leur performance en termes de qualité, ces
variétés et beaucoup d’autres introduites après, ont été disparu très vite à cause de leurs susceptibilités à plusieurs maladies telles que la rouille et la septoriose. Jusqu’à ce jour, la sole nationale est prédominée
par les variétés de blé tendre, Salambo (1980), Utique (1996), Haidra (2004) et plus récemment Tahent
(2009). En effet, toutes ces variétés ont été sélectionnées pour leur bon rendement et leur résistance aux maladies à savoir la rouille jaune, rouille brune et la septoriose (Ammar et al., 2011 ; Gharbi et al., 2013 ;
Ben Hamouda et al., 2016).
Etant donné leur contribution dans la production en grain du blé, l’introduction des variétés semi-naines
a augmenté considérablement les attaques des pathogènes notamment la septoriose (Gharbi et al., 2013). En effet la progression verticale de la maladie à partir des feuilles basales là où l’infection commence
vers la feuille drapeau est devenue très rapide avec les nouveaux cultivars grâce à la hauteur réduite de
la plante (Ben Mohamed et al., 2000). Dans le cas de Zymoseptoria tritici, l’agent causal de la Septoriose du blé, la reproduction sexuée est possible uniquement lorsque les deux idiomorphes sont présents au
même niveau et à la même localité géographique et celle-ci ne sera initiée que par la rencontre et la
fusion de ces deux gamètes qui sont génétiquement compatibles (Zhan et al., 2002). L’identification, le clonage et le séquençage des idiomorphes de Z. tritici ont été réalisés à partir des deux isolats de
référence IPO323 et IPO94269 (Waalwijk et al., 2002). Ces idiomorphes, lors du stade sexué de Z.
tritici, donnent naissance aux pseudothèces qui sont produits sous certaines conditions durant l’année
(Hunter et al., 1999). Toutefois, la distribution géographique des types sexuels parait fortement liée à l’évolution et la biologie des populations des champignons hétérothalliques (McDonald et Mundt,
2016).
En Tunisie, cette maladie s’est intensifiée dans les zones humides de Bizerte et Béja engendrant des pertes des rendements dépassant 60% chez les variétés de blé dur sensibles comme ‘Karim’ (Gharbi et
al., 2008). La sévérité d’attaque par Z. tritici dans les champs de blé dur est influencée par la sensibilité
élevée des variétés améliorées ‘Maali’, ‘Khiar’ et ‘Razzek’. Cependant, la variété Salim reconnue comme résistante à la rouille brune est considérée également comme la plus résistante à Z. tritici en
Tunisie ((Ben Mohamed et al., 2000 ; Gharbi et al., 2013).
Toutefois, des études antérieures ont mentionné que la plus grande incidence de ce pathogène a été
observée sur le blé dur que sur le blé tendre (Djerbi et Ghodhbane, 1975). Cette hypothèse a été aussi supportée par Gharbi et al. (2000), qui ont signalé que le blé tendre en Tunisie a montré depuis toujours
un haut niveau de résistance à ce pathogène. C’est pour cette raison que seuls les travaux s’intéressant
à l’équilibre gamétique, la diversité génétique, les gènes de résistance et l’importance de Z. tritici en Tunisie n’ont été effectués que sur le blé dur (Boukef et al., 2012 ; Ferjaoui et al., 2015 ; Aouini, 2018).
Bien que la septoriose fût observée sur le blé dur annuellement, une incidence surprenante a été signalée
ces dernières années, particulièrement dans la région d’El Haouaria chez une ancienne variété appelée
par les agriculteurs de cette région : ‘Farina Arbi’ (Bel Hadj Chedli et al., 2018). Les objectifs majeurs de ce travail sont : (i) d’étudier la distribution des types sexuels (MAT1-1 et MAT 1-2) et d’évaluer le
potentiel de la reproduction sexuée de Z. tritici à partir d’une collection de 103 isolats obtenue à partir
de la variété de blé tendre ‘Farina Arbi’ durant la saison agricole 2015-2016, (ii) réévaluer l'importance relative de cette maladie en Tunisie.
2. Matériel et Méthodes
2.1. Echantillonnage
Des feuilles de blé tendre infectées par Zymoseptoria tritici ont été échantillonnées pendant la saison
agricole 2015/2016. Tous les échantillons ont été collectés à partir des champs de blé tendre ‘Farina
Arbi’ infectés naturellement dans la région de El Haouariavers la fin de la saison de croissance (GS 70 selon l’echelle Zadok (Zadoks et al., (1974)). L’échantillonnage a été réalisé selon la méthode

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3894
hiérarchique décrite par McDonald et al. (1999) (Figure 1). Après réception au laboratoire, des procédures du stockage des feuilles et d’isolement ont été adoptées.
Figure 1. Méthode d’échantillonnage adaptée : dans chaque point circulaire, 10 feuilles de blé tendre ont été échantillonnées
à des intervalles réguliers de 10 m entre les différents points selon des lignes parallèles distants de 10 m (Méthode adaptée
par McDonald et al.1999)
2.2. Préparation des isolats et multiplication sporale
Cette étape a consisté à préparer au total 103 souches mono-pycnidiales conformément à la méthode adaptée par Siah et al. (2010). En effet il s’agit d’incuber, des feuilles fraiches de blé tendre présentant
des pycnides, une nuit dans une boîte de Pétri préalablement stérilisée contenant du papier filtre imbibé
d’eau. L’environnement humide à l’intérieur de la boite a permis la libération des cirrhes contenant des
spores qui ont été récupéré sur milieu PDA. Ainsi, la masse fongique obtenue a été déposée sur milieu liquide pour la multiplication sporale puis récoltée dans des tubes Eppendorf et stockée au (-80) pour
servir par la suite à l’extraction d’ADN. Les détails du nombre d’isolats et leurs origines sont décrits
dans le tableau 1.
Tableau 1. Collection de Z. tritici utilisées dans cette étude : site, variété, hôte et nombre d’isolats.
Région Site Variété Hôte Lattitude
(intervalle)
Longitude
(intervalle)
Nombre de
champs
Nombre
d’isolats
Cap Bon Haouaria Farina Arbi Blé tendre 36.84-36.94 10.93-11.02 4 103
2.3. Extraction d’ADN
La première étape d’extraction a consisté à lyophiliser les échantillons qui ont été broyée par la suite
moyennant un Tissue Lyser II (Qiagen cat.no. 85300) jusqu'à l’obtention d’une poudre qui a servi à
l’extraction de l’ADN en utilisant le kit d’extraction ‘Sbeadex® mini plant kit (LGC genomics) sur un système KingFisher KF96 conformément aux instructions du fabricant. La qualité d’ADN a été par la
suite vérifiée sur gel d’agarose (1%).
2.4. Détermination des types sexuels
Les deux idiomorphes (MAT1-1 et MAT1-2) ont été déterminés avec une PCR multiplexe en combinant
les couples d’amorces spécifiques à chaque type sexuel (Waalwijk et al., 2002) (Tableau 2). Dans cette
étude, deux isolats de référence à savoir IPO95052 (660 bp, MAT1-2) et IPO323 (340 bp, MAT1-1), ont été utilisés comme témoin. Les produits PCR obtenus sont séparés par une électrophorèse sur gel
d’agarose (1% agarose, 0,5X TBE) à 100V pendant 45min. Le gel d’agarose est ensuite coloré avec du
bromure d’éthidium et visualisé sous lumière UV.

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3895
Tableau 2. Couple d’amorces spécifiques à chaque type sexuel
Mating Type Primer 5'to 3' longueur
Mat 1-1F CCGCTTTCTGGCTTCTTCGCACTG 660 bp
Mat 1-1R TGGACACCATGGTGAGAGAACCT
Mat 1-2F GGCGCCTCCGAAGCAACT 340 bp
Mat 1-2R GATGCGGTTCTGGACTGGAG
2.5. Analyse des données
Le potentiel de la reproduction sexuée a été évalué en utilisant le test Chi-deux (χ2). En considérant le
scenario des cycles réguliers d’une reproduction sexuée, une distribution égale (ratio 1:1) des idiomorphes à l’intérieur de chaque population est attendue.
Le Chi-deux a été calculé comme décrit ci-dessous et comparé avec la probabilité P dans le tableau de
χ2 à un degré de liberté. χ2 = Σ (O-E) 2/E Avec, O : la fréquence observée
E : la fréquence attendue
Σ : somme pour les deux types sexuels
3. Résultats et Discussion
Différentes régions céréalières dans le Nord du pays considéré comme des foyers chauds (hot-spot) de
septoriose ont été prospectées durant la saison agricole 2015-2016 afin d’étudier la distribution géographique de la septoriose chez le blé tendre (Triticum aestivum), le blé dur (Triticum durum) et le
triticale (Triticale secalis) (Bel Hadj chedli et al., 2018). En effet, une incidence surprenante de Z. tritici
a été observée exclusivement dans la région d’El Haouaria chez une variété ancienne de blé tendre.
Désignée par ‘Farina Arbi’, cette variété sert à la fabrication des biscuits traditionnels de cette région et plus particulièrement le ‘Kaak Arbi’.
Les résultats de cette étude ont montré que 46 isolats ont été amplifiés à 340 pb avec MAT1-1 et 57
isolats ont été amplifiés à 660 bp avec MAT1-2 avec des fréquences de 44,66 et 55,33 % respectivement (Figure 2). La détermination des types hétérothalliques (Mating type) a été bien documenté dans la
littérature et l’existence des deux types (MAT1-1 et MAT1-2) sur le blé a été prouvée dans plusieurs
études (Zhan et al., 2002 ; Siah et al., 2010 ; Ayad et al., 2013 ; Alioui et al., 2014 ; Harrat et Bouznad, 2018). Dans le cas de cette étude, les résultats du test Chi-deux (χ2) appliqué pour tester l’hypothèse
nulle (ratio 1 :1) pour une population se produisant au hasard a révélé une différence non significative
entre les fréquences des deux idiomorphes dans toute la population d’une façon générale dans les
champs 1, 2 et 4 particulièrement (Tableau 3).
Tableau 3. Fréquences des idiomorphes et test Chi-deux types sexuels MAT1-1 et MAT1-2 de Zymoseptoria tritici dans
la région d’El Haouaria
Région Parcelle Nombre d’isolats MAT1-1 MAT1-2 χ2
El Haouaria
1 29 14 (48,27%) 15 (51,72%) 0.03 2 31 18 (58,06%) 13 (41,93%) 0.80 3 23 6 (26,08%) 17 (73,91%) 5.26 4 20 8 (40%) 12 (60%) 0.8
Total 103 46 (44,66%) 57 (55,33%) 1.17
Ces résultats sont en accord avec ceux identifiés par Siah et al. (2010), El Chartouni et al. (2012) et
Morais et al. (2017), suggérant ainsi l’existence d’un cycle régulier de la forme sexuée de Z. trtici en conditions naturelles. Des fréquences approximativement égales des idiomorphes soulignent le rôle
important des ascospores impliqués dans l’initiation de la maladie et le maintien de l’épidémie dans
cette région (Abrindaba et al., 2010 ; Meamiche et al., 2017).
Contrairement, un déséquilibre gamétique a été observé dans le champ 3, avec un ratio déviant signification du 1 :1 à (P ≤ 0.05) entre les deux idiomorphes avec une prédominance du MAT 1-2. Des
études similaires ont signalé la présence des fréquences inégales des types sexuels à partir de plusieurs
populations de Z. tritici de différentes origines : Iran (Saidi et al., 2012) ; California et Kansas (Gurung et al., 2011) ; France (Morais et al., 2017). A partir d’une population syrienne de Z. tritici, Waalwijk et
al. (2002) ont signalé des fréquences inégales des deux types sexuels avec une dominance du MAT 1-
1, conséquente de la taille réduite de la population et de l’échantillonnage répété des clones. Certains

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3896
chercheurs ont attribué ce déséquilibre à l’effet de la sélection ou la dérive génétique qui favorise certains génotypes tout en augmentant la fréquence générant un déséquilibre jusqu’à ce que la
reproduction sexuée aura le temps d’atteindre un équilibre (McDonald et Linde, 2002 ; Boukef, 2012 ;
Abrindabaet al., 2010). Cette étude constitue une première approche dans la caractérisation moléculaire des isolats de Z. tritici collectés à partir du blé tendre en Tunisie. Ainsi, l’étude de l’équilibre gamétique
identifié dans les trois champs ne pourra que confirmer le déroulement de la phase sexuée dans la région
d’El Haouaria pour assurer un inoculum primaire dans le déclenchement des épidémies. La forme sexuée des isolats de blé tendre pourrait être ultérieurement confirmée par la présence des pseudothèces au
niveau des champs. Par ailleurs, une étude de la diversité génétique à l'aide de marqueurs microsatellites
permettra d’approfondir les connaissances sur la structure des populations de Z. tritici en Tunisie et
améliorera significativement nos connaissances sur l'impact de la reproduction sexuée sur l'évolution de ce pathogène en Tunisie.
Figure 2. PCR Multiplex pour Mat1-1 et Mat1-2 pour quelques isolats de Z. tritici chez le blé tendre de la région d’El
Haouaria (champs 1) avec les deux isolats de références : IPO95052 (660 bp, Mat1-2) et IPO323 (340 bp, Mat1-1).
3. Conclusion
L’identification des proportions égales des deux idiomorphes au sein des mêmes champs dans la région
du Cap Bon suggère que la forme téléomorphe se reproduit dans les champs de blé tendre et même dans
un espace réduit (même champs). D’autres travaux sont nécessaires pour consolider les résultats trouvés dans cette investigation. En effet, l’identification du cycle sexué, sa fréquence tout au long du cycle de
la plante et l’étude de la diversité génétique de Z. tritici chez le blé tendre en Tunisie restent à déterminer.
Remerciements
Les auteurs remercient vivement toute l’équipe de “CRP Wheat Septoria Precision Phenotyping
Platform, Tunisia” pour leur contribution dans ce travail. Egalement nos remerciements s’adressent à l’équipe du Pr. Kema du laboratoire de Phytopathologie à l’Université de Wageningen (Pays-Bas) pour
leur collaboration.
4. Références
Abrinbana M, Mozafari J, Shams-Bakhsh M, Mehrabi R (2010) Genetic structure of
Mycosphaerella graminicola populations in Iran. Plant Pathology 59:829-838.Doi: 10.1111/j.1365-
3059.2010.02309. Ammar K, Gharbi MS, Deghais M (2011) Wheat in Tunisia. In: The World Wheat Book: A History
of Wheat Breeding. Lavoisier, 2 pp: 443-463.
Allioui N, Siahb A, BrinisaL, Reignault P, et Halama P (2014) Mating type distribution provides evidence for sexual reproduction of Mycosphaerella graminicola in Algeria. Canadian Journal of
Plant Pathology36(4): 475-481.DOI: 10.1080/07060661.2014.964777
Aouini L (2018) Durum wheat and septoria tritici blotch : gene and prospects for breeding. PhD thesis,
Wageningen University, Wageningen, Netherlands: 1-225. Ayad D, Sayoud R, Benbelkacem K, Bouznad Z (2013) La tache septorienne du blé: Première
signalisation de la présence en Algérie des deux Mating types du téleomorphe Mycosphaerella
graminicola (Fuckel) Schröter, (anamorphe : Septoriatritici Rob. ex Desm.) et diversité phénotypique de l’agent pathogène. Nature and Technology :34-45.
Be lHadj Chedli R, Ben M’Barek S, Yahyaoui A, Kehel Z, Rezgui, S (2018) Occurrence of Septoria
tritici blotch (Zymoseptoria tritici) disease on durum wheat, triticale, and bread wheat in Northern

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3897
Tunisia. CHILEAN JOURNAL OF AGRICULTURAL RESEARCH, 78(4): 559-568.doi:10.4067/S0718-58392018000400559
Ben Hamouda M., Ben Youssef S, Bedhief S, Annabi M, Draief Z, Gharbi M S (2016) Status of
Agricultural Innovations, Innovation Platforms, and Innovations Investment in Tunisia. PARI project country report: Republic of Tunisia. Forum for Agricultural Research in Africa: 21-25.
Ben Mohamed L, Rouaissi M, Sebei A, Hamza S, Harrabi M (2000) Effet du génotype, de la date de
semis, de la fertilisation azotée et potassique et des fongicides sur le développement de Septoriatritici. In Royo C. (ed.), Nachit M. (ed.), Di Fonzo N. (ed.), Araus J.L. (ed.). Durum wheat
improvement in the Mediterranean region : New challenges. CHIEHAM- options
méditerranéenne : Série A. Séminaires Méditerranéens 40: 350-356.
Boukef S (2012) Structure des populations, spécificité parasitaire et évaluation de la résistance aux fongicides chez Zymoseptoriatritici, agent de la septoriose du blé en Tunisie. Institut National
Agronomique de Tunisie : 1-102.
Djerbi M, Ghodhbane A (1975) Contribution à l’étude du Septoriatritici Rob. ex Desm., conservation et résistance variétale. Ann INRAT 48:1.
EL Chartouni L, Tisseront B, Siah A, DuymeF, Durant R, Halama PP, Reignault P (2012)
Evolution of Mycosphaerella graminicola at the wheat leaf and field levels. Phytopathologia
Mediterranea 51 (52): 332−339. El Felah M, Gharbi MS, Ghanem HB, Elloumi M (2015) Les céréales en Tunisie entre mythe et
réalité. Annales de l’INRAT, 2ème Numéro Spécial Centenaire 88: 1-17.
Ferjaoui S, Ben M’Barek S, BahriAB, Slimane RB, Hamza S (2015) Identification of resistance sources to septoriatritici blotch in old durum wheat germplasm applied for the analysis of the
Zymoseptoria tritici-durum wheat interaction. Journal of Plant Pathology 97 (3): 471- 481.
Gharbi MS, et El Faleh M (2013) Les céréales en Tunisie : plus d’un siècle de recherche variétale. Ann INRAT, Numéro Spécial- Centenaire de l’INRAT86 :45-68.
Gharbi MS, Berraiess S, Fakhar S, Ammar K, Hajlaoui MR, Yahyaoui A, Trifi M (2008) Breeding
durum wheat with resistance to Septoria leaf blotch for sustainable production under Mediterranean
conditions. International durum wheat symposium: from seed to pasta, 30/06-03/07/2008, Bologne, Italie.
Gharbi MS, Deghais M, Ben Amar F (2000). Breeding for resistance to Septoria tritici in durum wheat.
In: Royo C. (ed.), Nachit M. (ed.), Di Fonzo N. (ed.) , Araus J.L. ( ed.). Durum wheat improvement in the Mediterranean region : New challenges. Options Méditerranéennes : Série A. Séminaires
Méditerranéens 40: 397- 401.
Gurung SB, Goodwin S, Kabbage MW, Bockus W, Adhikari T (2011). Genetic Differentiation at Microsatellite Loci Among Populations of Mycosphaerella graminicola from California, Indiana,
Kansas, and North Dakota. Phytopathology 101(10): 1251-1257.doi:10.1094/PHYTO-08-10-0212
Harrat W, Bouznad Z (2018) Prevalence, cultural and pathogenic characterization of
Zymoseptoriatritici, agent of wheat septoria leaf blotch, in Algeria. African Journal of Agricultural Research 13(40): 2146-2153.doi:10.1094/PHYTO-08-10-0212
Hunter T, Coker RR, et Royal DJ (1999) The teleomorph stage, Mycosphaerella graminicola, in
epidemic of Septoriatritici blotch on winter wheat in the UK. Plant Pathol 48:51-57. McDonald BA, Mundt CC (2016) How knowledge of Pathogen Population Biology Informs
Management of SeptoriaTritici Blotch. Phytopathology 106(09): 948-955.
McDonald BA, Linde, CC (2002) The population genetics of plant pathogens and breeding strategies
for durable resistance. Euphytica, 124: 163-180.Annu. Rev. Phytopathol. 2002. 40:349–79 doi: 10.1146/annurev.phyto.40.120501.101443
McDonald BA, Zhan J, et Burdon J J (1999) Genetic Structure of Rhynchosporium secalis in
Australia. Ecologie and Population Biologie89(8): 639-644.DOI:10.1094/PHYTO.1999.89.8.639 Meamiche HN, Aouini L, Bouznad Z, Gerrit HJ (2018) Equal distribution of mating type alleles and
the presence of strobilurin resistance in Algerian Zymoseptoria tritici field populations. Plant
Disease: 1-27. doi.org/10.1094/PDIS-03-16-0298-RE Morais D, Duplaix C, Sache I, Laval V, Suffert F, Walker AS (2017). Stability in the genetic structure
of a Zymoseptoria tritici population from epidemic to interepidemic stages at a small spatial scale.
bioRxiv: 1-37.
Saade, M.E. 1996. Adoption and impact of high yielding wheat varieties in Northern Tunisia. CIMMYT, Mexico D.F. Economics Working Paper: 03-96.

Volume 61 (5). Published January, 01, 2019 www.jnsciences.org E-ISSN 2286-5314
BELHADJ et al. (2019) / Journal of new sciences, Agriculture and Biotechnology, 61 (5), 3892-3898 3898
Saidi A, Safaie N, Palizi P (2012) Study on mating types and sensitivity to strobilurin fungicide in fungal wheat pathogen Mycosphaerella graminicola.J. Crop Prot. 1 (4): 271-277.
Siah A, Bomble M, TisserantB, Cadalen T, Holvoet M, Hilbert R, Reignault P, Halama P (2018)
Genetic structure of Zymoseptoria tritici in northern France at region, field, plant and leaf layer scales. Phytopathology: 1-42. doi.org/10.1094/PHYTO-09-17-0322-R
Siah A, Tisserant B, El Chartouni L, Duyme F, Deweer C, Fichter C, Sanssené J, Durand R,
Reignault P, Halama P (2010). Mating type idiomorphsfrom French population of the wheat pathogen Mycosphaerella graminicola: widespread equal distribution and low but distinctlevels of
molecular polymorphism. Fungal Biology114:980-990. doi :10.106/j.funbio.2010.09.008
Waalwijk C, Mendes O, Verstappen ECP, de Waard MA, Kema GHJ (2002.) Isolation and
characterization of the mating type idiomorphs from the wheat septoria leaf blotch fungus Mycosphaerella graminicola. Fungal Genetics and Biology 35:277-286.
doi:10.1006/fgbi.2001.1322
Zhan J, Kema GH J, Waalwijk C, McDonald BA (2002) Distribution of mating type alleles in the wheat pathogen Mycosphaerella graminicola over spatial scales from lesions to continents. Fungal
Genetics and Biology 36:128-136.

ORIGINAL ARTICLE
Screening for resistance of Tunisian, Moroccan and Algerian wheatcultivars to Zymoseptoria tritici in Northern Tunisia
Rim Bel Hadj Chedli1,2 & Sarrah Ben M’Barek3,2 & Amir Souissi1 & Amor Yahyaoui4,2 & Salah Rezgui1 &
Hanène Chaabane5
Received: 22 May 2019 /Accepted: 24 April 2020# Società Italiana di Patologia Vegetale (S.I.Pa.V.) 2020
AbstractSeptoria tritici blotch (STB) disease caused by Zymoseptoria tritici is the most predominant disease on durum wheat inTunisia, while its occurrence on bread wheat is rare. In this study, we investigated the performance of 89 wheat cultivarsfrom Morocco, Algeria and Tunisia, screened in Tunisia for their relative resistance to STB. Field experiments werecarried out in an augmented design, during 2016–17 and 2017–18 cropping seasons at two locations in NorthernTunisia: Beja (Oued Beja station) and Cap Bon regions (Menzel Temim and El Haouaria). All trials were conductedunder natural infection. Visual disease assessments were quantified using the percentage of leaf area covered by pycnidia(PC), necrotic area (N), area under disease progress curve of each genotype (AUDPC) and the relative area under diseaseprogress curve (rAUDPC). Results indicated that the majority of Tunisian, Algerian and Moroccan durum wheat popula-tions (TDM, ADW and MDW) were susceptible to STB at both locations in Tunisia ranging from susceptible to highlysusceptible, with the rAUDPC, N and PC ranging from 0.5 to 0.8, 30 to 65% and 40 to 75% respectively. On the otherhand, the Moroccan bread wheat genotypes (MBW) were susceptible in Cap Bon area and resistant at Beja. Tunisian breadwheat genotypes (TBW) were resistant at both locations; with levels varying from immune to resistant classes whererAUDPC, PC and N did not exceed 0.2 and 10% respectively, with the exception of the local bread wheat variety known as“Farina Arbi” which was susceptible at Cap Bon and resistant at Beja.
Keywords Wheat genotypes . Zymoseptoria tritici . Resistance . Susceptibility
Introduction
North Africa has been the cradle of wheat production for cen-turies and was the bread basket for the Romain Empire(Bachta 2011). Nowadays, the Maghreb zone of NorthAfrica is still the major durum wheat producer which is thebasis for their traditionnal dishes such as couscous and pasta(Rastoin and Benabdrazik 2014). Tunisia is among the coun-tries with high cereal consumption and the average per capitaconsumption reached 259 kg (Rastoin and Ben Abderrazik2014; Hanson 2016). Durum wheat Triticum turgidum L.subsp. durum (Desf.)) is commonly cultivated in Tunisia prob-ably since the Roman era, while cultivation of bread wheat(Triticum aestivum L. subsp. aestivum) was introduced byFrench colonists in the early 1900s (El Felah et al. 2015).Hence, the cultivation of bread wheat in Tunisia started withthe selection of the cultivar Florence-Aurore that covered upto 80% of the Tunisian bread wheat acreage until 1952 then
Electronic supplementary material The online version of this article(https://doi.org/10.1007/s42161-020-00563-w) contains supplementarymaterial, which is available to authorized users.
* Rim Bel Hadj [email protected]
1 Laboratory of genetics and plant breeding, National AgronomicInstitute of Tunisia (INAT), University of Carthage, 43 AvenueCharles Nicolle, 1002 Tunis, Tunisia
2 CRPWheat Septoria Precision Phenotyping Platform, Tunis, Tunisia3 Regional Field Crops Research Center of Beja (CRRGC) BP 350,
9000 Beja, Tunisia4 Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT),
km 45 Carretera México-Veracruz El Batán, Texcoco, Mexico5 Laboratory of Bioagressors and Integrated protection in agriculture,
National Agronomic Institute of Tunisia (INAT), University ofCarthage, 43 Avenue Charles Nicolle, 1002 Tunis, Tunisia
Journal of Plant Pathologyhttps://doi.org/10.1007/s42161-020-00563-w

dropped to about 50% by 1959 (Ammar et al. 2011; BenHamouda et al. 2016).
After the introduction of the semi-dwarf high yieldingbread wheats lines selected from CIMMYT nurseries knownas « Mexican wheat », the bread wheat lines such as Baroota52, Sonora 63, Inia 66, Tobari 66, Ariana 66 and Mahon 73,selected from introduced populations were releasedandcovered large areas due to their high-yield performances(Saade 1996). These varieties were soon replaced by manywheats line such as ‘Soltane 72’, but also rapidly abandoneddue to their succeptibility tomany diseases namely yellow rustand septoria. During the late 1980’s and early 1990’s,‘Salammbô 80’ was the most grown bread wheat variety thatcovered 70% of the bread wheat acreage and was subsequent-ly followed by ‘Byrsa 87’. These two varieties became themost popular because of their high-yield potentiel and resis-tance to rust and septoria (Saade 1996). More recent releasesuch as Utique (1996), Haidra (2004), and Tahent (2009) hadgood performances and acceptable diseases resistance mainlyto powdery mildew, yellow rust, leaf rust, and septoria(Ammar et al. 2011).
Across the Magreb countries, the most prevalent foliardisease is Septoria tritici blotch (STB) caused by theascomycete fungus Zymoseptoria tritici (Roberge ex Desm.)Quaedvl. & Crous anamorph and Mycosphaerellagraminicola (Fuckel) J. Schröt.1894 teleomorph. This diseasecan reach epidemic levels on early planted cereals particularlywhen rainfall occurs in late winter and/or spring which iscommon in the Mediterranean-type environments (Chartrainet al. 2005).
In Tunisia, STB causes major yield losses on durumwheat (Ben Mohamed et al. 2000) where most commer-cial durum wheat cultivars are highly susceptible to thispathogen while bread wheat has been tolerant to STB(Gharbi and Deghaies 1997; Gharbi et al. 2000). Underfavorable growing conditions STB disease could reduceyield by 40% (Gharbi et al. 2011; Berraies et al. 2014).The prevailing strains seem to have developed a uniqueaggressiveness towards durum wheat (Yahyaoui et al.2000). The opposite situation exists in Morocco, wherebread wheat is the major crop affected by STB(Mazzouz et al. 1995; Zahiri et al. 2014). It has beenpreviously reported that isolates of Z. tritici exhibit bothcultivar specificity (ability to infect only some cultivars ofeither durum or bread-wheat) and host species specificity[ability to only infect one or the other wheat (Kema et al.1996; Kema and Silfhout 1997)]. This contrast betweenMorocco and Tunisia may explain further that specificityof Z. tritici isolates exists in wheat (Yahyaoui et al. 2000).However, in Algeria, the STB represents the major threatto both durum and bread wheat on coastal and sub-littoralregions of the country (Benbelkacem et al. 2011; Ayadet al. 2014).
Recent research provided better insight on the epidemiolo-gy of Z. tritici in the Maghreb countries. Siah et al. (2015)reported a high level of genetic diversity within the MoroccanZ. tritici population. The occurrence of the teleomorph stageof Z. tritici has been confirmed in Algeria and Tunisia(Meamiche et al. 2018; Ben Hassine and Hamada 2014)where the two mating types were found at equal frequencies(Boukef 2012; Ayad et al. 2014). Thus, knowledge of thehost-pathogen relationship and understanding the basis ofhost-specificity and resistance in the Mediterranean area areessential for successful genetic control of STB both on durumand bread wheat in the Maghreb region.
The objectives of this study were: (i) to evaluate bread anddurum wheat varieties from Tunisia, Algeria and Morocco forSTB resistance under natural infection in Tunisia, (ii) to eval-uate the phenotypic differences in resistance of wheat geno-types to STB using the area under disease progress curve(AUDPC), the relative AUDPC (rAUDPC), pycnidial cover-age (PC) and necrosis (N) in order to compare STB develop-ment between wheat species and finally (iii) to investigate therelationships between quantitative traits.
Materials and methods
Description of the study areas and experimentaldesign
Field experiments were conducted during 2016–2017 and2017–2018 wheat-growing seasons, in two regions under dif-ferent sub-humid zones where STB epidemics regularly occur(Fig. 1). The first experiment was carried out at the CRPWheat Septoria Precision Phenotyping Platform- experimen-tal station of the CRRGC at Oued-Beja (36°44′05”N, 9°13″35”E, governorate of Beja, northwest of Tunisia) located inthe sub-humid bioclimatic zone where the average annualrainfall ranges from 500 to 850 mm and a daily mean temper-ature varies between 10 and 28 °C. This area is particularlyknown to be a hot spot for STB especially on durum wheat.The second experimental was carried out at a farmer fieldlocated at Cap Bon Area (36°47′47”N, 11°0′8″E, governorateof Nabeul, northwest of Tunisia) with precipitation and tem-perature rates varying from 400 to 600 mm and from 6 to33 °C, respectively. This region has been recently identifiedas a hot spot for STB especially on bread wheat (Bel HadjChedli et al. 2018).
The experiments were set in two trials and set up in anaugmented design. During 2016–2017, the trials were sownon November 17 and 18 at Beja and Cap Bon, respectivelywhile during the 2017–2018, these fields were sown onNovember 6 and 11 at Cap Bon and Beja respectively. Eachgenotype was sown in paired rows of 2 mwith 4 g of seeds perrow. Standard experimental station plot maintenance was
J Plant Pathol

applied (herbicide, hand weeding and fertilizer application) toensure adequate crop development.
Plant materials
A set of 89 wheat genotypes including bread wheat fromCIMMYT’s International Septoria Observation Nursery(ISEPTON); bread and durum wheat varieties from Tunisia,Morocco and Algeria (Table 1), were screened for their rela-tive resistance to STB. Details about ISEPTON’S stb genesare reported in supplementary data 2. Two susceptible checkswere used in this study: the bread wheat landrace “FarinaArbi” identified as susceptible in Cap Bon region during asurvey conducted in 2015–2016 cropping season (Bel HadjChedli et al. 2018) and the most susceptible durum wheatcultivar ‘Karim’ (Deghaïs et al. 2007; Ammar et al. 2011).
Evaluation of disease severity and area under diseaseprogress curve
Visual disease assessment based on the leaf area covered withpycnidia (PC) and the necrotic area (N) were estimated as apercentage of the uppermost-infected leaves either on Flagleaf or Flag leaf −1 at the end of the growth stage, i.e. GS70.In addition, the symptoms and lesion development over theassessment period were summarized by the area under diseaseprogress curve (AUDPC) that allows the identification of dif-ferent classes of resistance. Disease severity was scored foreach plot using the double-digit scale (Saari and Prescott1975). The first digit (D1) indicates disease progress on theinfected plants, and the second digit (D2) refers to severity ofinfection. Three consecutive evaluations weremade at 10 daysinterval, at GS51, GS59 and GS65 respectively according toZadok’s scales (Zadoks et al. 1974).
The AUDPC and the relative area under disease progresscurve (rAUDPC) were subsequently calculated according toSimko and Piepho (2012) formula:
AUDPC ¼ ∑n−1
i¼1
yi þ yiþ1
2� tiþ1−tið Þ
Where:Yi: STB severity at time ti,t(i + 1)-ti = time interval (days) between two disease scores,n = number of times when STB was recorded.
rAUDPC ¼ AUDPC g2notypeð ÞAUDPC Karimð Þ
Where: Karim is the most susceptible variety used as acheck.
Statistical analysis
All the observations in the experimental field and dependentvariables were subjected to analysis of variance (ANOVA)using ‘aov’ function from R package ‘daewr’ (Lawson 2016)implemented in R software v3.4.2 (R Core Team 2017) andleast-Squares Means using R package ‘lsmeans’ (V. Lenth2016). Distances between wheat genotypes using hierarchicalclustering method and correlations coefficients between char-acters were calculated for all traits analyzed in the study. Aweighted clustering algorithm K-mean (K = 6) (Duda andHart 1973) was performed using JMP®11.0 in order to groupthe different genotypes in classes. Subsequently, Principal com-ponent analysis (PCA) was performed using the JMP®11.0statistical software (SAS Institute Inc., Cary, NC, USA) withComponent analysis procedure (SAS Institute 2014).
Results
Meteorological conditions during the crop cycle
Meteorological data (temperature and rainfall) over differentclimatic regions and during the two cropping seasons was
Fig. 1 Map of Tunisia showing the location of study area (Beja and Cap Bon area) during 2017–2018 cropping season. The STB experiments were set inan augmented design at both locations
J Plant Pathol
Table 1 Wheat cultivars screened for resistance to Septoria tritici blotch disease during 2016–2017 and 2017–2018 cropping seasons
BW/DW
Cultivar name Number Country Provenance Origin/Year
BW Néapolis 1 Tunisia CRRGCB CRRGC & INRAT
BW Mahon 73 1 Tunisia CRRGCB Algeria, 1910
BW Inia 66 1 Tunisia CRRGCB INRAT/CIMMYT, 1970
BW Castan 1 Tunisia CRRGCB France, 1976
BW Dougga 74 1 Tunisia CRRGCB CIMMYT, 1974
BW Florence Aurore 1 Tunisia CRRGCB France, 1974
BW Carthage 74 1 Tunisia CRRGCB Mexico, 1974
BW Ariana66 1 Tunisia CRRGCB France, 1970
BW Tahent 1 Tunisia CRRGCB INRAT/CIMMYT, 2010
BW Haïdra 1 Tunisia CRRGCB INRAT, 2004
BW Utique 96 1 Tunisia CRRGCB INRAT/CIMMYT, 1996
BW Salammbô 80 1 Tunisia CRRGCB INRAT/CIMMYT, 1980
BW Vaga 92 1 Tunisia CRRGCB INRAT/CIMMYT, 1992
BW Byrsa 87 1 Tunisia CRRGCB INRAT/ CIMMYT, 1987
DW Maali 1 Tunisia INGC Tunisia, 2007
DW Nasr 1 Tunisia INGC INRAT/ICARDA, 2004
DW INRAT 100 1 Tunisia INGC INRAT, 2017
DW Dhahbi 1 Tunisia INGC INRAT, 2017
DW Razzak 1 Tunisia INGC INRAT, 1987
DW Salim 1 Tunisia INGC INRAT, 2009
DW Karim 1 Morocco Morocco INRA Morocco, 1985
DW Marzak 1 Morocco Morocco INRA Morocco, 1984
DW Sebou 1 Morocco Morocco INRA Morocco, 1987
DW Omrabia 1 Morocco Morocco INRA Morocco, 1988
BW Amal 1 Morocco Morocco INRA Morocco, 1993
BW Arrehane 1 Morocco Morocco INRA Morocco, 1996
BW Aguilal 1 Morocco Morocco INRA Morocco, 1996
BW Marchouch 1 Morocco Morocco INRA Morocco 1984
DW Tomouch 1 Morocco Morocco INRA Morocco, 1997
DW Algeria1 1 Algeria Algeria DZ/CCB
DW Algeria 2 1 Algeria Algeria DZ/CCB
DW Algeria 3 1 Algeria Algeria DZ/CCB
DW Algeria 4 1 Algeria Algeria DZ/CCB
DW Algeria 5 1 Algeria Algeria DZ/CCB
DW Algeria 6 1 Algeria Algeria DZ/CCB
DW Algeria 7 1 Algeria Algeria DZ/CCB
DW Algeria 8 1 Algeria Algeria DZ/CCB
DW Algeria 9 1 Algeria Algeria DZ/CCB
BW ISEPTONS 49 CIMMYT CIMMYT CIMMYT
Checks Karim 1 Tunisia INGC INRAT/CIMMYT, 1980
Farina Arbi 1 Tunisia Farmers Landrace/El Haouaria farmers
Total: 89
DW: durum wheat, BW: bread wheat
Durum accessions Algerian (DZ) crossing block (CCB)
INGC: Institut National des Grandes Cultures (National Institute of Field Crops)
CRRGCB: Centre Régional de Recherche des Grandes Cultures de Béja (Regional Field Crops Research Center of Béja)
J Plant Pathol
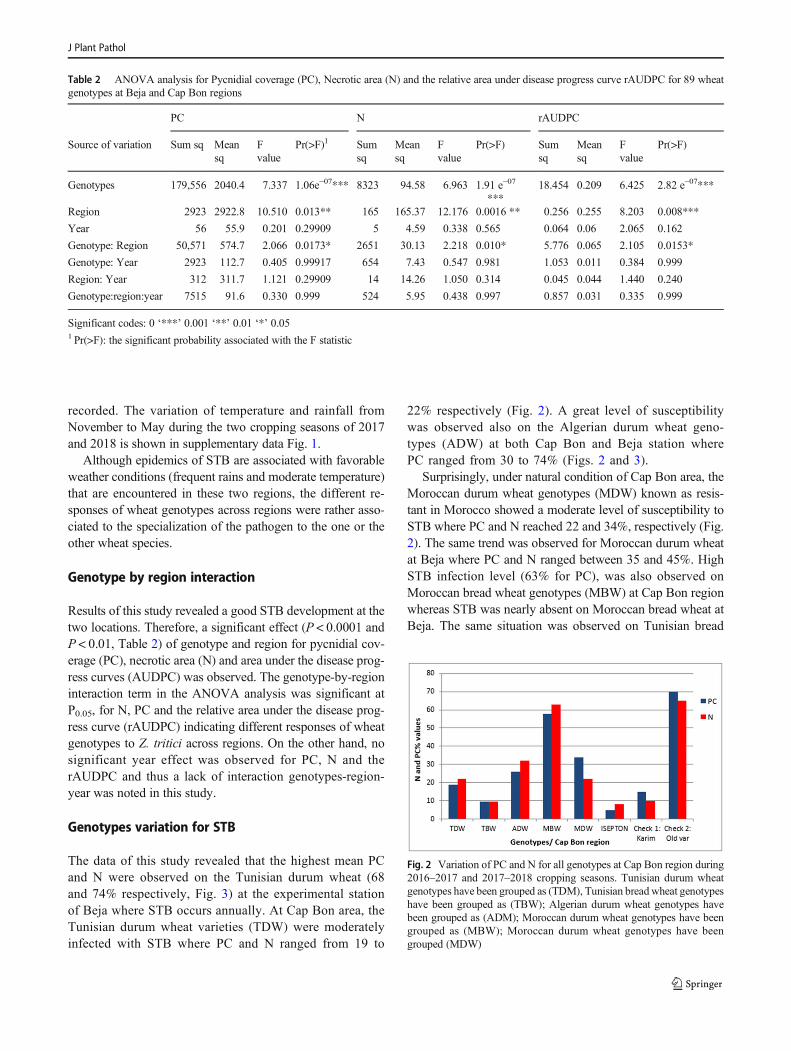
recorded. The variation of temperature and rainfall fromNovember to May during the two cropping seasons of 2017and 2018 is shown in supplementary data Fig. 1.
Although epidemics of STB are associated with favorableweather conditions (frequent rains and moderate temperature)that are encountered in these two regions, the different re-sponses of wheat genotypes across regions were rather asso-ciated to the specialization of the pathogen to the one or theother wheat species.
Genotype by region interaction
Results of this study revealed a good STB development at thetwo locations. Therefore, a significant effect (P < 0.0001 andP < 0.01, Table 2) of genotype and region for pycnidial cov-erage (PC), necrotic area (N) and area under the disease prog-ress curves (AUDPC) was observed. The genotype-by-regioninteraction term in the ANOVA analysis was significant atP0.05, for N, PC and the relative area under the disease prog-ress curve (rAUDPC) indicating different responses of wheatgenotypes to Z. tritici across regions. On the other hand, nosignificant year effect was observed for PC, N and therAUDPC and thus a lack of interaction genotypes-region-year was noted in this study.
Genotypes variation for STB
The data of this study revealed that the highest mean PCand N were observed on the Tunisian durum wheat (68and 74% respectively, Fig. 3) at the experimental stationof Beja where STB occurs annually. At Cap Bon area, theTunisian durum wheat varieties (TDW) were moderatelyinfected with STB where PC and N ranged from 19 to
22% respectively (Fig. 2). A great level of susceptibilitywas observed also on the Algerian durum wheat geno-types (ADW) at both Cap Bon and Beja station wherePC ranged from 30 to 74% (Figs. 2 and 3).
Surprisingly, under natural condition of Cap Bon area, theMoroccan durum wheat genotypes (MDW) known as resis-tant in Morocco showed a moderate level of susceptibility toSTB where PC and N reached 22 and 34%, respectively (Fig.2). The same trend was observed for Moroccan durum wheatat Beja where PC and N ranged between 35 and 45%. HighSTB infection level (63% for PC), was also observed onMoroccan bread wheat genotypes (MBW) at Cap Bon regionwhereas STB was nearly absent on Moroccan bread wheat atBeja. The same situation was observed on Tunisian bread
Table 2 ANOVA analysis for Pycnidial coverage (PC), Necrotic area (N) and the relative area under disease progress curve rAUDPC for 89 wheatgenotypes at Beja and Cap Bon regions
PC N rAUDPC
Source of variation Sum sq Meansq
Fvalue
Pr(>F)1 Sumsq
Meansq
Fvalue
Pr(>F) Sumsq
Meansq
Fvalue
Pr(>F)
Genotypes 179,556 2040.4 7.337 1.06e−07*** 8323 94.58 6.963 1.91 e−07
***18.454 0.209 6.425 2.82 e−07***
Region 2923 2922.8 10.510 0.013** 165 165.37 12.176 0.0016 ** 0.256 0.255 8.203 0.008***
Year 56 55.9 0.201 0.29909 5 4.59 0.338 0.565 0.064 0.06 2.065 0.162
Genotype: Region 50,571 574.7 2.066 0.0173* 2651 30.13 2.218 0.010* 5.776 0.065 2.105 0.0153*
Genotype: Year 2923 112.7 0.405 0.99917 654 7.43 0.547 0.981 1.053 0.011 0.384 0.999
Region: Year 312 311.7 1.121 0.29909 14 14.26 1.050 0.314 0.045 0.044 1.440 0.240
Genotype:region:year 7515 91.6 0.330 0.999 524 5.95 0.438 0.997 0.857 0.031 0.335 0.999
Significant codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.051 Pr(>F): the significant probability associated with the F statistic
Fig. 2 Variation of PC and N for all genotypes at Cap Bon region during2016–2017 and 2017–2018 cropping seasons. Tunisian durum wheatgenotypes have been grouped as (TDM), Tunisian breadwheat genotypeshave been grouped as (TBW); Algerian durum wheat genotypes havebeen grouped as (ADM); Moroccan durum wheat genotypes have beengrouped as (MBW); Moroccan durum wheat genotypes have beengrouped (MDW)
J Plant Pathol

wheat genotypes (TBW) and ISEPTON with insignificant PCand N levels where they did not exceed 10% (Fig. 2) at CapBon, whereas the susceptible check bread wheat landrace“Farina Arbi” showed high level of susceptibility (70 and65% for PC and N respectively, Fig. 2). Despite the highSTB disease pressure on bread wheat landrace “Farina Arbi”observed in previous survey at Cap Bon region, this landraceseems to be immune at Beja station (Fig. 3).
Genotypes classification
K-means classification proved the presence of significant dif-ferences between all tested genotypes related to the percentageof diseases infection observed in wheat areas included in thestudy. The analysis ranged wheat genotypes in six differentclasses.
At Beja site, an immune reaction (class I: rAUDPC = 0)was recorded for all ISEPTON accessions, and all Tunisianand Moroccan bread wheat genotypes (Table 3). TwoMoroccan wheat genotypes (Sebou and Marchouch) wereranked as resistant (class II; Table 3, Fig. 4) where rAUDPCdid not exceed 0.1. Two Moroccan durum wheat (Marzak,Toumouh) and four Algerian durum wheat genotypes(Algeria2, Algeria 3, Algeria 4, Algeria 7) were ranked asclass III and IV (moderately resistant to moderately suscepti-ble). At Beja site, the genotypes with rAUDPC higher than 0.6such as Algerian durum genotypes: Algeria 5, Algeria 6,Algeria 8, and Algeria 9, Tunisian durum wheat: Salim,Maali, Karim, Dhahbi, INRAT100, and Nasr ranked as classVI and VII, susceptible and highly susceptible cultivars(Table 3, Fig. 4).
On the other hand, at Cap Bon area, the highly resistant andresistant class (I and II) where rAUDPC did not exceed 0.2included the Tunisian durum genotypes (Karim and Salim),
the Algerian durum wheat ‘Algeria 1’, a set of Tunisian breadwheat and ISEPTONS (Table 4; Fig. 5). The moderately re-sistant group (III) where rAUDPC varied from 0.2 to 0.3 in-cluded the following genotypes: Algeria 2, Algeria 3, Algeria4, Moroccan durum genotypes (Sebou and Tomouch) and theTunisian cv. ‘Nasr’ (Table 4; Fig. 5).
The rest of Algerian durum genotypes: Algeria 5, Algeria6, Algeria 7, Algeria 8, Algeria 9, the Tunisian cvs.: Dhahbi,Maali, INRAT100, the Moroccan varieties (Marzak, Amal,Karim and Om rabia) and three ISEPTON accessions wereranked as moderately susceptible and susceptible class IVand V (Table 4; Fig. 5) where rAUDPC did not exceed 0.7.The genotypes with rAUDPC over than 0.8 included theMoroccan bread wheat genotypes: Aguilal, Marchouch andArrehane and the Tunisian landrace (Farina Arbi) were rankedas highly susceptible class VI (Table 4; Fig. 5).
Significant correlation between quantitative traits
Principal component analysis (PCA) allowed detecting simi-larities in the genotypes with regard to STB responses acrosstwo experimental sites during two years (Fig. 6). The majorcorrelated variability of genotypes showed by axes 1 and 2,revealed 6 groups within each region (Figs. 4 and 5). The firstPC1 axe accounted for 98% of the total variability expressedby quantitative traits (AUDPC, rAUDPC, PC, and N) whilethe second component (PC2) accounts for only 1.64% of thetotal variation. On the other hand, strong positive correlationbetween the four infection measures (AUDPC, rAUDPC, PC,and N) of the 89 genotypes was observed in this study (Fig. 6).
Discussion
Disease severity in plant-pathosystems can be assessed eitheronce or several times at some intervals starting from diseaseinitiation until the end of the epidemic. The former method ofassessment measures can be used to estimate different param-eters such as the area under the disease progress curves(AUDPC) and the relative area under the disease progresscurve (rAUDPC) which are used by several pathologists inthe analysis of data on resistance to Septoria (Kema et al.1996; Chartrain et al. 2004; Mojerlou et al. 2009; Ferjaouiet al. 2015). Here, we assessed AUDPC and rAUDPC, pyc-nidial coverage (PC) and the necrotic area (N) under fieldconditions to investigate the behavior of the Tunisian,Moroccon and Algerian bread and durum wheat varieties fortheir resistance to Septoria tritici blotch (STB) at two differentlocations where the pathogen seems to have achieved a spe-cialization to one or other wheat species.
The choice of using Moroccon, Tunisian and Algeriandurum and bread wheat genotypes relies on the fact that eventhough STB poses a serious threat in the Maghreb countries,
Fig. 3 Variation of PC and N across genotypes at Beja station regionduring 2016–2017 and 2017–2018 cropping seasons. Tunisian durumwheat genotypes have been grouped as (TDM), Tunisian bread wheatgenotypes have been grouped as (TBW); Algerian durum wheat geno-types have been grouped as (ADM); Moroccan durum wheat genotypeshave been grouped as (MBW); Moroccan durum wheat genotypes havebeen grouped (MDW)
J Plant Pathol

Table 3 Means and ranges of AUDPC and rAUDPC for all identified clusters at Beja region
Clusters AUDPC rAUDPC Durum wheat genotypes Bread wheat genotypes
IHighlyresistant
Maximum 0.0 0.0 Algeria 1 From ISEPTON 1 to ISEPTON 49, Mahon73,Amal, Arrehane, Aguilal,, Néapolis, Farina Arbi,Salammbô, Tahent, Utique, Vaga,, Ariana66, Byrsa,Carthage, Castan, Dougga, Florence Aurore, Haidra.Inia66.
Minimum 0.0 0.0
Mean 0.0 0.0
IIResistant
Maximum 367.5 0.2 Sebou MarchouchMinimum 220.0 0.1
Mean 293.8 0.1
IIIModerately
resistant
Maximum 937.5 0.4 Marzak, Algeria 7, Algeria2, Algeria 3 –Minimum 772.5 0.4
Mean 857.5 0.4
IVModerately
suscepti-ble
Maximum 1121.3 0.5 Toumouch, Algeria 4 –Minimum 1086.3 0.5
Mean 1014.6 0.5
VSusceptible
Maximum 1528.8 0.7 Algeria 5, 6, 8, Nasr, Salim,Maali,Moroccan varieties (Karim and Omrabia)
–Minimum 1356.3 0.6
Mean 1440.8 0.7
VIHighly
suscepti-ble
Maximum 1775.9 0.8 Algeria 9, Dhahbi, INRAT100, karim –Minimum 1576.3 0.7
Mean 1636.2 0.8
Fig. 4 PCA showing the majorcorrelated variability of genotypesas shown by axes 1 and 2. Thefirst Dimension1 accounted for98% of the total variabilityexpressed by quantitative traitswhile the second component(Dimension2) accounts only1.64% of the total variation. ACPrevealed 6 clusters at Beja region:Cluster 1: Very resistant; Cluster2: resistant; Cluster 3: moderatelyresistant; Cluster 4: moderatelysusceptible; Cluster 5: suscepti-ble; Cluster 6: very susceptible.Details about genotypes of eachgroup are shown in Table 3
J Plant Pathol

Table 4 Means and ranges of AUDPC and rAUDPC for all identified clusters at Cap Bon region
Clusters AUDPC rAUDPC Durum wheat genotypes Bread wheat genotypes
IHighly resistant
Maximum 500.0 0.2 Algeria 1 Ariana66, Byrsa, Carthage, Castan, Dougga,Inia66, Utique, Vaga,
From ISEPTON 10, to ISEPTON 31ISEPTON 34, 35, 36, 39, 40, 42, 43, 44,
48, 8, 9, 3, 4.
Minimum 0.0 0.0
Mean 5.2 0.0
IIResistant
Maximum 425.8 0.2 Salim, Karim, Florence Aurore, Haidra, Néapolis, Tahent,Mahon 73, ISEPTON 2, 24, 32, 33, 37,38,41, 45, 46, 47, 5, 7.
Minimum 22.5 0.0
Mean 232.9 0.1
IIIModerately
resistant
Maximum 686.3 0.3 Algeria 2, 3, 4,Sebou, Tomouch, Nasr.Minimum 449.0 0.2
Mean 591.3 0.3
IVModerately
susceptible
Maximum 1281.3 0.6 Algeria 5, Dhahbi, Maali, INRAT100 ISEPTON 1, 6, 49.Minimum 753.8 0.4
Mean 1138.6 0.5
VSusceptible
Maximum 1491.3 0.7 Marzak, Amal, Algeria 6, 7,8,9, Moroccan varieties(Karim and Om rabia)
Salambo, AmalMinimum 1110.0 0.5
Mean 1325.7 0.6
VIHighly
susceptible
Maximum 1907.5 0.9 – Arrehane, Aguilal, Marchouch, Farina ArbiMinimum 1671.3 0.8
Mean 1785.7 0.8
Fig. 5 PCA showing the majorcorrelated variability of genotypesas shown by axes 1 and 2accounting 98% and 1.64%respectively of the total variabilityexpressed by quantitative traits.ACP revealed 6 clusters at CapBon region: Cluster 1: Veryresistant; Cluster 2: resistant;Cluster 3: moderately resistant;Cluster 4: moderately susceptible;Cluster 5: susceptible; Cluster 6:very susceptible. Details aboutgenotypes of each group areshown in Table 4
J Plant Pathol
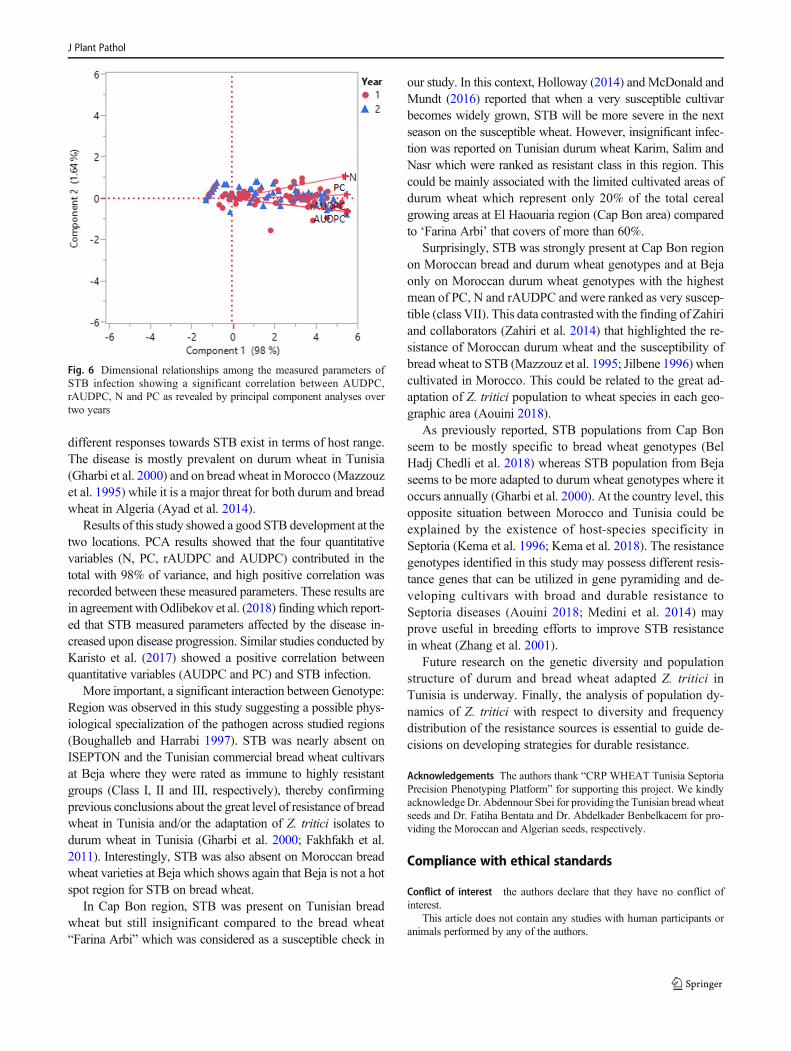
different responses towards STB exist in terms of host range.The disease is mostly prevalent on durum wheat in Tunisia(Gharbi et al. 2000) and on bread wheat inMorocco (Mazzouzet al. 1995) while it is a major threat for both durum and breadwheat in Algeria (Ayad et al. 2014).
Results of this study showed a good STB development at thetwo locations. PCA results showed that the four quantitativevariables (N, PC, rAUDPC and AUDPC) contributed in thetotal with 98% of variance, and high positive correlation wasrecorded between these measured parameters. These results arein agreement with Odlibekov et al. (2018) findingwhich report-ed that STB measured parameters affected by the disease in-creased upon disease progression. Similar studies conducted byKaristo et al. (2017) showed a positive correlation betweenquantitative variables (AUDPC and PC) and STB infection.
More important, a significant interaction between Genotype:Region was observed in this study suggesting a possible phys-iological specialization of the pathogen across studied regions(Boughalleb and Harrabi 1997). STB was nearly absent onISEPTON and the Tunisian commercial bread wheat cultivarsat Beja where they were rated as immune to highly resistantgroups (Class I, II and III, respectively), thereby confirmingprevious conclusions about the great level of resistance of breadwheat in Tunisia and/or the adaptation of Z. tritici isolates todurum wheat in Tunisia (Gharbi et al. 2000; Fakhfakh et al.2011). Interestingly, STB was also absent on Moroccan breadwheat varieties at Beja which shows again that Beja is not a hotspot region for STB on bread wheat.
In Cap Bon region, STB was present on Tunisian breadwheat but still insignificant compared to the bread wheat“Farina Arbi” which was considered as a susceptible check in
our study. In this context, Holloway (2014) and McDonald andMundt (2016) reported that when a very susceptible cultivarbecomes widely grown, STB will be more severe in the nextseason on the susceptible wheat. However, insignificant infec-tion was reported on Tunisian durum wheat Karim, Salim andNasr which were ranked as resistant class in this region. Thiscould be mainly associated with the limited cultivated areas ofdurum wheat which represent only 20% of the total cerealgrowing areas at El Haouaria region (Cap Bon area) comparedto ‘Farina Arbi’ that covers of more than 60%.
Surprisingly, STB was strongly present at Cap Bon regionon Moroccan bread and durum wheat genotypes and at Bejaonly on Moroccan durum wheat genotypes with the highestmean of PC, N and rAUDPC and were ranked as very suscep-tible (class VII). This data contrastedwith the finding of Zahiriand collaborators (Zahiri et al. 2014) that highlighted the re-sistance of Moroccan durum wheat and the susceptibility ofbread wheat to STB (Mazzouz et al. 1995; Jilbene 1996) whencultivated in Morocco. This could be related to the great ad-aptation of Z. tritici population to wheat species in each geo-graphic area (Aouini 2018).
As previously reported, STB populations from Cap Bonseem to be mostly specific to bread wheat genotypes (BelHadj Chedli et al. 2018) whereas STB population from Bejaseems to be more adapted to durum wheat genotypes where itoccurs annually (Gharbi et al. 2000). At the country level, thisopposite situation between Morocco and Tunisia could beexplained by the existence of host-species specificity inSeptoria (Kema et al. 1996; Kema et al. 2018). The resistancegenotypes identified in this study may possess different resis-tance genes that can be utilized in gene pyramiding and de-veloping cultivars with broad and durable resistance toSeptoria diseases (Aouini 2018; Medini et al. 2014) mayprove useful in breeding efforts to improve STB resistancein wheat (Zhang et al. 2001).
Future research on the genetic diversity and populationstructure of durum and bread wheat adapted Z. tritici inTunisia is underway. Finally, the analysis of population dy-namics of Z. tritici with respect to diversity and frequencydistribution of the resistance sources is essential to guide de-cisions on developing strategies for durable resistance.
Acknowledgements The authors thank “CRP WHEAT Tunisia SeptoriaPrecision Phenotyping Platform” for supporting this project. We kindlyacknowledgeDr. Abdennour Sbei for providing the Tunisian bread wheatseeds and Dr. Fatiha Bentata and Dr. Abdelkader Benbelkacem for pro-viding the Moroccan and Algerian seeds, respectively.
Compliance with ethical standards
Conflict of interest the authors declare that they have no conflict ofinterest.
This article does not contain any studies with human participants oranimals performed by any of the authors.
Fig. 6 Dimensional relationships among the measured parameters ofSTB infection showing a significant correlation between AUDPC,rAUDPC, N and PC as revealed by principal component analyses overtwo years
J Plant Pathol

Informed consent Informed consent was obtained from all individualparticipants included in the study.
References
Ammar K Gharbi MS, and Deghais M (2011) Wheat in Tunisia. p. 443–463. In Bonjean, APAngus, andWJ van Ginkel,M (eds.) The worldwheat book: a history of wheat breeding. Vol. 2. Lavoisier, Paris,France
Aouini L (2018) Durum wheat and septoria tritici blotch : gene andprospects for breeding. PhD thesis, Wageningen University,Wageningen, Netherlands: 1–225
Ayad D, Sayoud R, Benbelkacem K, et Bouznad Z (2014) La tâcheseptorienne du blé: Première signalisation de la présence enAlgérie des deux Mating types du téleomorphe Mycosphaerellagraminicola (Fuckel) Schröter, (anamorphe: Septoria tritici Rob.Ex Desm.) et diversité phénotypique de l’agent pathogène.“Nature & Technologie ". B- Sciences Agronomiques etBiologiques: 34–45
Bachta MS (2011) La céréaliculture en Tunisie Une politique derégulation à repenser, note d’analyse du CIHEAM, 64:3
Bel Hadj Chedli R, Ben M’Barek S, Yahyaoui A, Kehel Z, Rezgui S(2018) Occurrence of Septoria tritici blotch (Zymoseptoria tritici)disease on durum wheat, triticale, and bread wheat in NorthernTunisia. Chil J Agric Res 78(4):559–568. https://doi.org/10.4067/S0718-58392018000400559
Ben Hamouda H, Ben Youssef S, Sonia Bedhiaf S, Mohamed Annabi M,Zied Draief Z, Gharbi MS (2016) Status of agricultural innovations,innovation platforms, and innovations investment. Tunisia. 2015PARI project country report: republic of Tunisia. Forum for agricul-tural research in Africa (FARA), Accra, Ghana
Ben Hassine A, Hamada W (2014) First isolation of theMycosphaerellagraminicola teleomorph stage causing Septoria leaf blotch on wheatin Tunisia. New Dis Rep 29:18
BenMohamed L, Rouaissi M, Sebei A, Hamza S, Harrabi M (2000) Effetdu génotype, de la date de semis, de la fertilisation azotée etpotassique et des fongicides sur le développement de Septoria tritici.p. 350-356. In Royo, C., Nachit, M., Di Fronzo, N., and Araus, J.L.(eds.) Durumwheat improvement in the Mediterranean region: Newchallenges. CIEHAM, Zaragoza, Spain.(Options Méditerranéenne:Série A. Séminaires Méditerranéens; n. 40
Benbelkacem K, Bouznad Z, Benslimane H, Salhi NL (2011).Occurrence of septoria leaf blotch in Algéria and assessment ofwheat res is tance . 8ème in ternat ional symposium onMycrosphaerella and Stagnospora diseases of cereals, MexicoCity September: 10-14
Berraies S, Ammar K, Gharbi MS, Amor Y, Rezgui S (2014)Quantitative inheritance of resistance to Septoria tritici blotch indurum wheat in Tunisia. Chil J Agric Res 74:35-40
Boughalleb N, Harrabi M (1997) Physiologic specialization of Septoriatritici in North-Africa. Tropicultura 15(1):33–36
Boukef S (2012) Structure des populations, spécificité parasitaire etévaluation de la résistance aux fongicides chez Zymoseptoria triticiagent de la septoriose du blé en Tunisie. Institut NationalAgronomique de Tunisie:1–102
Chartrain L, Brading PA, Makepeace JC, Brown JKM (2004) Sources ofresistance to Septoria tritici blotch and implications for wheat breed-ing. Plant Pathol 53:454–460
Chartrain L, Berry ST, Brown JKM (2005) Resistance of wheat lineKavkaz-K4500 L.6.A.4 to septoria tritici blotch controlled byisolate-specific resistance genes. Phytopathology 95:664–671
Deghaïs M, Kouki M, Gharbi M.S, et El Felah M. 2007. Les Variétés decéréales cultivées en Tunisie. Editions INRAT, 445p
Duda RO, Hart PE (1973) Pattern classification and scene analysis.Wiley, New York
El Felah M, Gharbi MS, Ben Ghanem H, Elloumi M (2015) Les céréalesen Tunisie entre mythe et réalité. Annales de l’INRAT, 2èmeNuméro Spécial Centenaire, 88 :1–17
Fakhfakh MM, Rezgui S, Jarrahi T, Zouani, R, Bousalmi A, Sgatni K,Yahyaoui A (2011) Mapping distribution of Septoria allowedtargeting management options of the disease. 8th InternationalSymposium on Mycosphaerella and Stagonospora Diseases ofCereals, Mexico City 10–14 September 2011
Ferjaoui S, M’Barek SB, Bahri B, Slimane RB, Hamza S (2015)Identification of resistance sources to Septoria tritici blotch in olddurum wheat germplasm applied for the analysis of theZymoseptoria tritici-durum wheat interaction. J Plant Pathol 97(3):471–481
Gharbi MS, et Deghaies M (1997) Principaux enseignements à tirer de lacampagne céréalière 1995/1996. Actes des 3èmes journéesnationales sur les acquis de la recherche agronomique halieutiqueet vétérinaire, 29/11–01/12/1996, Nabeul, Tunisie
Gharbi MS, Deghais M, Ben Amar F (2000) Breeding for resistance toSeptoria tritici in durum wheat :397-401. In Royo C, Nachit M, DiFronzo N, and Araus JL (2000) durum wheat improvement in theMediterranean region: new challenges.: CIHEAM, Zaragoza, Spain.Options Méditerranéennes. Série a: Séminaires Méditerranéens; n.40
GharbiMS, Berraies S, Ammar K, Yahyaoui A (2011) Delivering diseaseresistant cultivars to enhance sustainability of durum wheat produc-tion in Tunisia. p. 60. In Duveiller E, Singh PK, et al. (eds.) 8thInternational Symposium on Mycosphaerella and StagonosporaDiseases of Cereals, Mexico City. 10–14 September 2011. Bookof abstracts. CIMMYT, Mexico
Hanson S (2016) Tunisia grain and feed annual. USDA ForeignAgricultural service, GAIN Report Number, pp 1–8
Holloway G (2014) Septoria tritici blotch of wheat. DEPI informationnote series may 2014. 2 p. Department of Environment and PrimaryIndustries (DEPI), Melbourne, Australia
Jilbene (1996) Etat des recherches sur la septoriose auMaroc. Proceedingdu Symposium régional sur les Maladies des céréales et lesLégumineuses Alimentaires Edit. INRA Rabat : 11–14
Karisto P, Hund A, Yu K, Anderegg J, Walter A, Mascher F, McDonaldBA, et Alexey Mikaberidze A (2017) Ranking quantitative resis-tance to Septoria tritici blotch in elite wheat cultivars using automat-ed image analysis.bioRxiv: 3-44
Kema GHJ, Silfhout CH (1997) Genetic variation for virulence and re-sistance in the wheat–Mycosphaerella graminicola pathosystem III.Comparative seedling and adult plant experiments. Phytopathology87:266–272
Kema GHJ, Annone JG, Sayoud R, Van Silfhout CH, Van Ginkel M, DeBree J (1996) Genetic variation for virulence and resistance in thewheat–Mycosphaerella graminicola pathosystem I. Interactions be-tween pathogen isolates and host cultivars. Phytopathology 86:200–212
Kema GHJ, Gohari AM, Aouini L, Gibriel HAY, Ware SB et al (2018)Stress and sexual reproduction affect the dynamics of the wheatpathogen effector AvrStb6 and strobilurin resistance. Nat Genet50:375–380
Lawson J (2016) Daewr: design and analysis of experiments with R. Rpackage version 1:1–7 https://CRAN.R-project.org/package=daewr
Russell V. Lenth (2016) Least-squares means: The R Package lsmeans.Journal of Statistical Software, 69(1), 1–33. https://doi.org/10.18637/jss.v069.i01
Mazzouz H, Saadaoui M, Jilbène M, Lyamani A (1995) Importance desseptorioses du blé au Maroc en 1991. Al Awamia 91:63–69
McDonald BA, Mundt CC (2016) How knowledge of pathogen popula-tion biology informs management of Septoriatritici blotch.Phytopathology 106(9):948–955
J Plant Pathol

Meamiche HN, Aouini L, Bouznad Z, Gert HJ (2018) Equal distributionof mating type alleles and the presence of strobilurin resistance inAlgerian Zymoseptoria tritici field populations. Plant Disease :1-27
MediniM, Ferjaoui S, Bahri B, MhriW, Hattab S, Sonia Hamza S (2014)Bulk segregant Analysis and marker-trait association reveal com-mon AFLP markers for resistance to Septoria leaf blotch inTunisian old durum wheat. BASE Biotechnol Agron Soc Environ,18(1):3–10
Mojerlou S, Safaie N, Alizadeh A, Khelghatibana F (2009) Measuringand modeling crop loss of wheat caused by septoria leaf blotch inseven cultivars and lines in Iran. J Plant Prot Res 49:257–262
Odlibekov F, Armoniené R, Henriksson T, Chwade A (2018) ProximalPhenotyping and machine learning methods to identify SeptoriaTritici blotch disease symptoms in wheat. Front Plant Sci 9:1–11
R Core Team (2017) R: a language and environment for statistical com-puting. R Foundation for Statistical Computing, Vienna, Austriahttps://www.R-project.org/
Rastoin JL, et Ben Abderrazik H (2014) Céréales et oléoprotéagineux auMaghreb : Pour un co-développement de filières territorialisées.Tunisie : le poids des institutions. Editions Institut de ProspectiveEconomique du Monde Méditerranéen (IPEMED), Maroc, 37p.Rapilly F. 1991. L’épidémiologie en pathologie végétale : mycosesaériennes. Institut National de la Recherche Agronomique 11–36
SaadeME (1996) Adoption and impact of high yieldingwheat varieties innorthern Tunisia. CIMMYT. Economics working paper 96-03.CIMMYT, Mexico D.F
Saari EE, Prescott JM (1975) A scale for appraising the foliar intensity ofwheat diseases. Plant Dis Rep 59:377–380
SAS Institute 2014. ‘JMP® 11 scripting guide.’ 2nd edn (SAS InstituteInc.: Cary, NC, USA)
Siah A, cedex L, Elbekali AY, Ramdani A, Reignault P, Torriani SFF,Brunner PC, Halama P (2015) QoI resistance and mitochondrialgenetic structure of Zymoseptoria tritici in Morocco. Plant Dis, 98(8): 1138–1144
Simko I, Piepho HP (2012) The area under the disease progress stairs:calculation, advantage, and application. PHYTO 102(4):381–389
Yahyaoui A, Hakim S, Al-Naimi M, Nachit MM (2000) Multiple diseaseresistance in durum wheat (Triticum turgidum L. var. durum). p.387-392. In Royo C, Nachit M, Di Fronzo N, Araus JL (eds.).Durum wheat improvement in the Mediterranean region: new chal-lenges. CIHEAM, Zaragoza, Spain. Options Méditerranéennes.Série a: Séminaires Méditerranéens; n. 40
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for the growthstages of cereals. Weed Res 14(6):415–421
Zahiri S, Farih A, Douira A (2014) Statut des principales maladiescryptogamiques foliaires du blé au Maroc en 2013. J. Appl, Biosci
Zhang X, Haley SD, Jin Y (2001) Inheritance of Septoria Tritici blotchresistance in winter wheat. Crop Sci 41:323–326
Publisher’s note Springer Nature remains neutral with regard to jurisdic-tional claims in published maps and institutional affiliations.
J Plant Pathol




















