
Tumor angiogenesis, macrophages and mast cell microdensities in endometrioid endometrial carcinoma

Molecular lmmunolo~~, Vol. 27, No. 8, pp. 751-761, 1990 Printed in Great Britain.
0161-5890190 $3.00 + 0.00 c 1990 Pergamon Press plc
SPECIFIC BINDING OF LIPOPOLYSACCHARIDES TO
MOUSE MACROPHAGES-I. CHARACTERISTICS OF THE INTERACTION AND INEFFICIENCY OF THE
POLYSACCHARIDE REGION
MOHAMED-ALI TAHRI-JOUTI and RICHARD CHABY*
Equipe “Endotoxines”, Unit& de Recherche Associte, 1116 du Centre National de la Recherche Scientifique, BPtiment 432, Universitt de Paris&d, 91405 Orsay, France
(Firsl rewiocd 7 October 1989; accepted in revised form 23 January 1990)
Abstract-Tritium-labeled lipopolysaccharide interacted specifically and reversibly with mouse peritoneal macrophages. The binding was higher at 22’C than at 4”C, but was no longer observable at 37’C. The specificity of the interaction (inhibition with unlabeled LPS) was strictly dependent on the presence of serum, and required divalent cations. The binding was saturable. The specific binding sites of peritoneal macrophages were saturated with 6-9 x IO6 LPS molecules/cell, and those of macrophage-like cell lines with 2-3 x 10” molecules/cell. The binding of LPS was not inhibited by ligands of scavenger receptors (maleylated BSA) or complement receptors (zymosan), but was strongly inhibited with dexamethasone, a glucocorticoid which is known to modulate the expression of other surface markers of macrophages. The polysaccharide region of the LPS, as well as 3-deoxy-2-octulosonic acid (KDO) coupled to bovine serum albumin, did not bind to macrophages, whereas a specific binding was observed with a lipid A-BSA conjugate.
INTRODUCTION
Lipopolysaccharide (LPS), a major constituent of the outer membrane of Gram-negative bacteria, causes a variety of beneficial (Johnson, 1983) and deleterious physiological effects (Morrison and Ulevitch, 1978) in humans and laboratory animals. These include fever, endotoxic shock, and acute-phase reaction. Although LPS is known to exert potent stimulatory effects on a number of animals cells including T lymphocytes (Milner et al., 1983) and B lymphocytes (Anderson et al., 1972), it is now generally recognized that the cell type primarily responsible for its physiologic effects is the macrophage, which synthesizes large amounts of immunoregularity molecules including interleukin-1 (IL-l) (Durum et al., 1985; Lasfargues et al., 1987), tumor necrosis factor (TNF) (Beutler et al., 1985; Lasfargues and Chaby, 1988), prostaglandins and leukotrienes (Aderem et al., 1986).
LPS is built up of a polysaccharide (PS) covalently bound to a phosphorylated glycolipid (lipid A). The PS region which emerges at the outside of the bac- terial outer membrane is freely accessible to host
*Author to whom correspondence should be addressed. Abbreviations: BSA, bovine serum albumin; BLA, lipid
fragment released during alkaline p-elimination of the periodate-treated endotoxin of Bordeteh pertussis; CM, culture medium; DEX, dexamethasone, FCS, fetal calf serum; HEPES, N-2-hvdroxvethvl-piperazine-N’-2- ethanesulfonic acid; HS, heat-inactivated horse serum; LPS, lipopolysaccharide; PBS, phosphate-buffered saline; PS- 1, polysaccharide fragment of Bordeteh pertussis endotoxin.
cells, and might mediate the binding of bacteria to macrophages observed by several investigators (Euteneuer et al., 1986; Tomita ef al., 1981). On the other hand, the release of LPS from bacteria may allow the lipid A region, initially immersed in the lipid bilayer of the bacterial membrane, to become accessible for interactions with macrophage sites, which initiates activation of these cells and leads to endotoxic symptoms. The presence of binding sites to both PS and lipid A regions of LPS on membranes of macrophages could therefore be reasonably envis- aged, inasmuch as a number of different plasma membrane receptors have been identified on mononuclear phagocytes (Gordon ef al.. 1988), among which are receptors for mannosyl, N-acetyl glucosaminyl and fucosyl glycoconjugates (Ezekovitz and Stahl, 1988), sulfated polysaccharides (Chong and Parish, 1986), sialyl-gangliosides (Cracker et al.,
1988), and lipoproteins (Gianturco et al., 1986). We established in a previous work (Haeffner-
Cavaillon et al., 1982) that a lectin-like receptor present on rabbit peritoneal macrophages recognizes the polysaccharide region of Bordetella pertussis LPS. Lectin-like receptors have been also identified on macrophages by other investigators (Ezekovitz and Stahl, 1988; Warner et al., 1985). In the current study, however, we show that there is no receptor for the polysaccharide region of the same LPS on mouse macrophages. In the accompanying paper (Tahri- Jouti et al., 1990) the role of lipid A substructures in the specific binding of LPS to mouse macrophages has been demonstrated.
751

752 M.-A. TAHRI-JOUTI and R. CHABV
MATERIALS AND METHODS
Animals. Swiss male mice (4-6 weeks old) were from Janvier (Le Genest Saint-Isle, France) and A,lJ female mice (8-10 weeks old) were from the Institut Pasteur (Paris, France).
Chemicals, reagents and media. Heat-inactivated horse serum (HS) was from Gibco (Glasgow, Scotland), and heat-inactivated fetal calf serum (FCS) was from Boehringer (Mannheim, F.R.G.). ULTROSER-G, an artificial serum substitute, was from IBF (Villeneuve-La-Garenne, France). Culture medium (CM) was RPMI- 1640 supplemented with L-glutamine (2 mM), streptomycin (100 pg/ml), penicillin (100 [U/ml) and 2-mercaptoethanol (5 x 10m5 M). Anti-human complement serum, heparin, Neutral Red, ionophore A23 187, dexamethasone, and cycloheximide were from Sigma Chemical Co. (St Louis, MO, U.S.A.). Thioglycollate broth and mini- mal essential medium (MEM) were from Diagnostic Pasteur (Ville d’Avray, France).
with anhydrous hydrazine (1 ml) for 24 hr at 22 C. The gel-like material which was formed was dissolved in ethanol, and the solvents were evaporated. Residual hydrazine was evaporated under vacuum in the presence of toluene. A solution of sodium nitrite (190~1, 22mgjml) was slowly added (7ml), under magnetic stirring, to a cold (O’C) suspension of the hydrazide (15 mg) in 0.2 M HCl. After 5 min, the suspension was slowly added to a solution of BSA (3 ml, 20mg/ml) in 0.2 M borate buffer JpH 9.2). During the addition, the pH was maintained at 9.2 with 1 M NaOH. The resultant mixture was then dialyzed overnight against 0.15 M NaCI. The conju- gate was purified by chromatography on a Sephadex Cl00 column. After dialysis against distilled water, the BSA-KDO conjugate (60 mg) was recovered by lyophilization. estimation of KDO by the periodate- thiobarbiturate assay (Chaby and Szabo, 1975) indicated the presence of 5-6 KDO residues per molecule of BSA.
LPSs and.~ragmenrs deriving thewfrom. Bordetella pertuss~s endotoxin was extracted from the bacteria by the phenol-water method (Westphal et af., 1952). LPSs from Escherichia coli J5 and Salmonella minne- sota Re-595 mutants were purchased from Sigma Chemical Co. (St Louis, MO, U.S.A.). The poly- saccharide (PSI) fragment of B. pertussis LPS was isolated by chromatog~phy on a Biogel PI0 column, after hydrolysis of the endotoxin with 10 mM sodium acetate (pH 4.5), for 2 hr at 100-C, in the presence of 1% SDS, according to Caroff et al. (1988). Monophosphoryl-lipid A was prepared from B. pertussis endotoxin as previously described (Le Dur et al., 1978).
Maleyl-BSA was prepared as described by Butler and Hartley (1972). According to the U.V. spectrum of the material (absorbance of the maleyl group at 250nm), 53% of the primary amino groups of the protein were maleylated.
Prepuratiu~ of‘ unlabeled f~ga~ds. The lipid frag- ment released from B. pertussis endotoxin after peri- odate treatment followed by alkaline [j-elimination of the sugar moiety @LA) was prepared as described previously (Lasfargues et al., 1989). BLA obtained from 5 mg of LPS was suspended in borate buffer (pIi 9.2,3 ml) and was added to a solution of BSA (350 ~1, lOmg/ml) in the same buffer, After incubation for 2 hr at 20 C, sodium borohydride (0.1 ml, 1 mg,/ml) was added, and the solution was kept at 4°C for an additional 18 hr. The conjugate consisting of BLA coupled to BSA (BSA-PLA) was isolated after centri- fugation (90 min; 100,000~) of the solution and lyophili~dtion of the supernatant. The BSA-/i LA preparation was purified by chromatography on a Sephadex Gl SO column, to eliminate aggregated and high molecular weight entities.
Preparation of labeled ligands. Two tritium-labeled LPS preparations ([‘HIOx-LPS and [“HIGlut-LPS) were obtained by reduction with tritiated boro- hydride of periodate-oxidized or gluta~dldehyde-sub- stituted LPS of B. pertassis, respectively. [3H]Ox-LPS was prepared by a modification of the procedure of Watson and Riblet (1975) as previously described. The specific activity was 120,000 cpmipg. Glutaralde- hyde-substituted LPS was isolated after incubation of B. pertussis LPS (4.6 mg) with a 0.2% glutaraldehyde solution (1. I5 ml) in 0.01 M phosphate buffer (PI-I 7), for 2.5 hr at 2O.C, followed by an ultracentrifugation (90 min, 50,OOOg). The pellet was suspended in 0.2 M borate buffer (0.5 ml, pH 8.9) and incubated for 18 hr at 4-C after addition of NaB[‘H], (13.5 mCi: 0.5 Ci/mmol; 250 ~1). After extensive dialysis and lyophilization, [‘HIGlut-LPS (3.8 mg. 44,000 cpmipg) was isolated. The tritium-labeled polysaccharide fragment of B. pertussis endotoxin was obtained by direct reduction of PS-1 (6.8 mg) with tritiated sodium borohydride (13.5 mCi; 0.5 Ci/mmol; 250 ~1). After purification by gel chromatography (Biogel PlO), [“HIPS-l (4.3 mg; 39~16Ocpm~~~g) was isolated.
“‘I-Labeling of BSA-BLA, BSA-KDO and
maleyl-BSA was performed by the chloramine-T procedure of Hunter and Greenwood (1962).
To obtain a conjugate of 3-deoxy-D-manno-2- Peritoneal macrophages and ceff fines. 5774 and octolosonic acid (KDO) covalently linked to BSA, P388-Dl lnonocyte~macrophage cell lines were grown the metl~oxycarbonyloctyl oside of KDO (a mixture in CM supplemented with IO and 5% FCS, respec- of r and fl anomers) was prepared (D. Charon, tively. Cells were cultured in 7S-cm’ flasks (Nunclon. unpublished results) by treatment of the benzyl ester Kamstrup, Denmark) at 37 C in a 5% COZ and of acetobromo-KDO with methoxycarbonyl octanol, 100% r.h. atmosphere, passaged every 2-4 days, and in the presence of mercuric salts. After removal of the released from the plastic surface by flushing with benzyl group, the material (50 mg) was incubated culture medium.

LPS binding to mouse macrophages-I 753
Mouse peritoneal exudates were harvested 5 days after i.p. injection of 1.7 ml of thioglycollate broth, by peritoneal lavage with MEM, containing 10 U/ml of sodium heparine. Suspension of exudate cells (IO6 Neutral Red-positive cells per well, in plates with 24 wells) in RPMI- 1640 buffered with 20 mM HEPES, were incubated for 2 hr at 37°C. Nonadherent cells were removed by four washings with MEM (500/~I/well). Adherent cells were incubated for an additional 18 hr in the same medium before use in the LPS-binding assay.
Binding of‘ labeled ligands to adherent cells. The assay was a modification of the method described previously (Haeffner-Cavaillon et al.. 1982). The labeled ligand (usually [‘HIOx-LPS at 1.33 pg/ml) alone, or with a 50-fold excess of B. pertussis LPS, was suspended in MEM supplemented with BSA (3.3 mgiml) and 10% heat-inactivated serum (usually horse serum, unless otherwise specified). Triplicate aliquots ( I.50 ~1) of the suspensions were incubated for I hr at 20 C (unless specified otherwise) with lo6 plastic-adherent macrophages which were preincu- bated for 18 hr in RPMI-1640 buffered with 20 mM HEPES. The supernatant was then discarded. The plates were washed three times by immersion in ice-cold 0.9% NaCl solution, the time of each wash- ing being of about 2 sec. The bound material was solubilized with ;a solution (0.5 ml) of SDS (1%) containing 2 mM EDTA. Radioactivities of ‘H- and “L1-labeled ligands were measured in a Rack Beta 1212 liquid scintillation counter, and a Mini Gamma
1275 gamma counter, respectively (both from LKB, Turku, Finland).
Statistical analysis. Graph lines were drawn by fitting binding equations to the experimental data, using a computer program of nonlinear regression, written in BASIC (Duggleby, 1981).
RESULTS
Specl$ic binding of [-‘H]Ox-LPS to mouse peritoneal macrophages
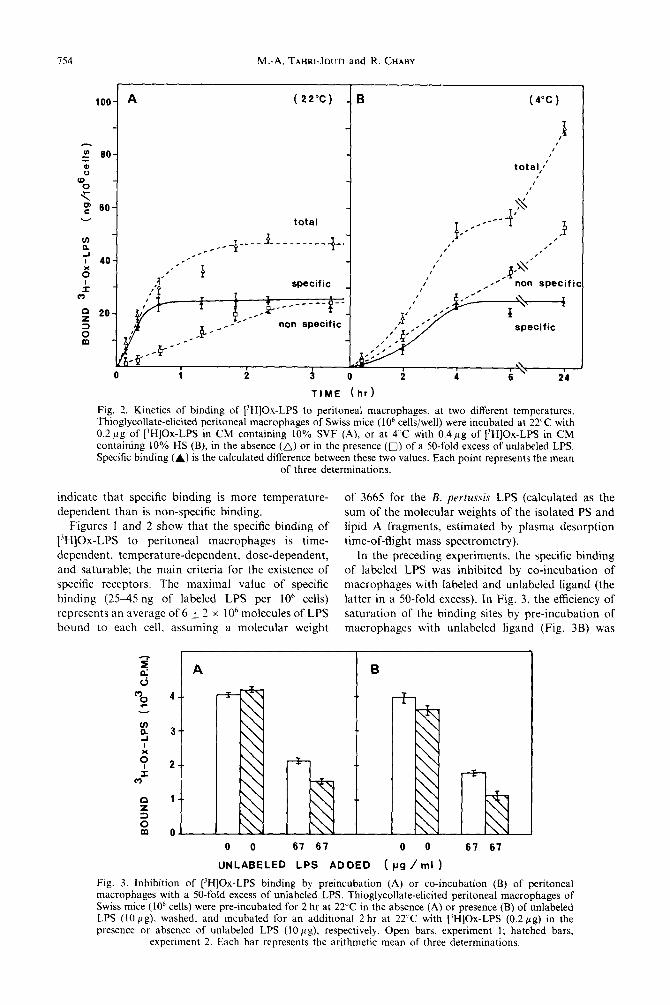
The dependence of the binding of labeled LPS to peritoneal macrophages as a function of ligand con- centration is shown in Fig. 1. The difference between the total binding (in the absence of unlabeled LPS) and the nonspecific binding (in the presence of a 50-fold excess of unlabeled LPS), reached a constant value, both at 22°C (Fig. 1A) and at 4°C (Fig. IB), which means that the specific binding of the ligand was saturable. The saturation value of this specific binding was very similar at 4 and 22’C.
On the other hand, the kinetics of the specific binding of [3H]Ox-LPS to the macrophages showed a considerable temperature dependence (Fig. 2). At 22’C, maximum binding of labeled LPS was observed after 30 min (Fig. 2A), whereas 4 hr were required at 4’C to reach a plateau of specific binding (Fig. 2B). It should be noted that both Figs I and 2 indicate that specific binding is lower than non-specific binding at 4’C, but is higher than the latter at 22°C. This may
loo- A (22°C) - 6 (4°C)
l-
80-
60-
total
d-------+-
,/N*ddd;,.,
I I
,’ , ,I ,’ ,’
I’ non specific
,‘ ,_-+-- I’ .
I’
1 f
specific
3H-Ox-LPS 019 /ml I
Fig. 1. Binding of [‘HIOx-LPS to peritoneal macrophages as a function of [‘HIOx-LPS concentration, at two different temperatures, Thioglycollate-elicited peritoneal macrophages of Swiss mice (10” cells/well) were incubated at 22°C for 2 hr (A), or at 4°C for 18 hr (B) in CM containing 10% HS, with increasing amounts of labeled LPS in the absence (A) or in the nresence (0) of a SO-fold excess of unlabeled LPS. _-/ Specific binding (A) is the calculated difference between these t&kalues. Each point represents the mean
of three determinations.
M.-A. TAHRI-JOUTI and R. CHABY
ioo A i
-P---- 4___
total,*’
,’
I-._=---+ ,p I’
I’ <’
I’ I’ **’
p’ ,’ .’
non specifi
TIME (hr)
Fig. 2. Kinetics of binding of [‘HIOx-LPS to peritoneal macrophages, at two different temperatures. Thioglycollate-elicited peritoneal macrophages of Swiss mice ( lo6 cells/well) were incubated at 22°C with 0.2~g of [‘HIOx-LPS in CM containing 10% SVF (A), or at 4 C with 0.4pg of [‘HIOx-LPS in CM containing 10% HS (B), in the absence (A) or in the presence (0) of a 50-fold excess of unlabeled LPS. Specific binding (A) is the calculated difference between these two values. Each point represents the mean
of three determinations.
dependent than is non-specific binding. indicate that specific binding is more temperature- of 3665 for the B. pertussis LPS (calculated as the
sum of the molecular weights of the isolated PS and lipid A fragments, estimated by plasma desorption time-of-flight mass spectrometry).
Figures 1 and 2 show that the specific binding of [‘HIOx-LPS to peritoneal macrophages is time- dependent, temperature-dependent, dose-dependent, and saturable; the main criteria for the existence of specific receptors. The maximal value of specific binding (2545 ng of labeled LPS per 10’ cells) represents an average of 6 + 2 x 10h molecules of LPS bound to each cell, assuming a molecular weight
In the preceding experiments, the specific binding of labeled LPS was inhibited by co-incubation of macrophages with labeled and unlabeled ligand (the latter in a 50-fold excess). In Fig. 3, the efficiency of saturation of the binding sites by pre-incubation of macrophages with unlabeled ligand (Fig. 3B) was
0 0 67 67 0 0 67 67
UNLABELED LPS ADDED ( pg /ml )
Fig. 3. Inhibition of [‘HIOx-LPS binding by preincubation (A) or co-incubation (B) of peritoneal macrophages with a 50-fold excess of unlabeled LPS. Thioglycollate-elicited peritoneal macrophages of Swiss mice (lo6 cells) were pre-incubated for 2 hr at 22°C in the absence (A) or presence (B) of unlabeled LPS (10 pg). washed, and incubated for an additional 2 hr at 22’C with [3H]Ox-LPS (0.2 pg) in the presence or absence of unlabeled LPS (10 pg), respectively. Open bars, experiment 1; hatched bars,
experiment 2. Each bar represents the arithmetic mean of three determinations.

LPS binding to mouse macrophages-I 75s
compared with that obtained in the co-incubation
procedure (Fig. 3A). Results of two independent
experiments (open and hatched bars) indicated that the two procedures were equally effective.
The reversibility of the binding of labeled LPS (the last criterion for the presence of a receptor) is illustrated by Fig. 4. After a binding step, the dis- sociation of LPS from the macrophages appears to be a rapid phenomenon. Even at 4-C, an amount of LPS equivalent to half of that specifically bound to the macrophages was dissociated from the cells in less than 5 min (Fig. 4) whereas the association of the same amount of LPS to the cells required 2-3 h at the same temperature (Fig. 2B).
Cornprison of the binding of’ LPSs labeled bll dl’jfkrent me this
It was important to check that the binding of [3H]Ox-LPS to mouse macrophages was not the result of the chemical modification of the polysaccharide region which occurs during the labeling procedure (after periodate oxidation and borohydride reduction, some of the heptose residues of the endotoxin are transformed in [?H]mannose). For this reason. we analyzed the binding of an LPS preparation ([“H]Glut-LPS) obtained by a different labeling technique (unsubstituted amino groups of the polysaccharide chain were coupled to glutaraldehyde, and the terminal aldehyde residues were reduced with tritiated borohydride). Results (not shown) indicated a specific and saturable binding of [“HIGlut-LPS to P388-DI cells (9.5 f 0.5 ngj106
cells) very similar to that of [3H]Ox-LPS (11 * 0.5 ng/lO’ cells).
Serum requirement
It has been reported already (Haeffner-Cavaillon et al., 1985) that the presence of serum in the incubation medium is required for the specific binding of LPS to macrophages. This finding is confirmed by the results of the present study. We observed that the specific binding of [3H]Ox-LPS increases as a function of the concentration of horse serum added in the medium. Optimal binding was reached with 10% HS (result not shown). In the absence of serum, or in the presence of an artificial serum substitute (ULTROSER-G from IBF, Villeneuve-La-Garenne. France), specific binding of LPS is not detectable (Table I). Results in Table I also show that them is no species specificity of the serum added in the incubation medium. However, all sera were not equally effective in this test. Horse serum was much more efficient than the homologous (Swiss) serum. On the other hand, it is noteworthy that the serum obtained from C3H/HeJ mice (which arc hyporesponsive to most of the biological effects of LPS) appeared significantly less efficient than the homologous serum in this test.
Inn requirement
Figure 5 shows that the specific binding of [‘HIOx-LPS to macrophages is considerably lower in the absence of divalent cations. When chelators of divalent cations (EDTA. EGTA) were added in the
t
Total binding
--- - - - - _ _ _ _ _ _ _ _ _ _ _ _ _ - _ _ _ _.
50
Specific binding
0 10 20 30 40 50 60
TIME (min)
Fig. 4. Kinetics of dissociation of [‘HIOx-LPS from peritoneal macrophages. Thioglycollate-elicited peritoneal macrophages of Swiss mice (IO6 cells/well) were incubated for 16 hr at 4-C with 0.2 pg of labeled LPS. in CM containing 10% HS. Cells were then rapidly washed with cold saline and incubated at 4 C for various periods in CM (500 ~1) containing 10% HS. Radioactivity released in the extracellular medium was measured. The dashed and dotted horizontal lines represent respectively the levels of total and specific
binding of labeled LPS, measured after the washing of the cells.

156 M.-A. TAHRI-JOUTI and R. CHABY
Table I. Bindmg” of (‘H]Ox-LPS to mouse pentoneal macrophagx in the presence of various serah
metabolic inhibitor of cells. Cycloheximide, which
Specdic binding inhibits protein biosynthesis, had no effect on non-
Serum (ng,‘lO” cells) specific binding. but induced some inhibition (36%)
None 0 of the specific binding. Colchicine, an inhibitor of the Artificial substitute 0 motile apparatus of cells (microtubules and micro- Mouse: SWIG\ 9.8 & 0.5
C?H:HeJ 3.‘) + 0.6 filaments) had a moderate effect on the specific
uor\e 22.5 +_ 0.1 binding (26% inhibition) but reduced considerably Fetal calf 8.2 + 2.0
7.3 + 0.4 (54%) the non-specific binding of LPS. It is note-
Human
“Bmdlng of labeled LPS (1.22 pcg;ml) to thmglycollate- &cited peritoneal macrophagea of Swiss mice ( IO6 cell\) wx meacursd after incubation for 2 hr at 22 ‘C
worthy that treatment of macrophages with LPS results in the expression of the intracellular precursor of IL- I/j’, which has been shown (Baldari and Telford,
,n the abaencc or presence of added (6.6pg/ml) unlabeled LPS.
Spwfic binding ~‘a, calculated as the difference between the meanb of triphcatr determination\ for these two conditions, and the standard dewatton of each result i\ the square root of the sum of the squares of each mc;m‘\ standard deviation.
‘All sera were heat-mactivated at 56 C for 30 min. and used at 10% 11, CM.
medium. a strong inhibition (75%) of the specific binding was observed (Fig. SA). Likewise, when the binding assay was performed in a medium devoid of divalent cations (PBS), the specific binding was 92% lower than that obtained in a medium which contained physiological concentrations of calcium and magnesium (RPMI) (Fig. 5B).
lr@ence of’ metabolic inhibitors and dexamethasone
We analyzed the effects of different metabolic inhibitors, or inhibitors of macrophage functions, on the non-specific and specific binding of [3H]Ox-LPS to macrophages. Results in Table 2 show that both non-specific and specific binding of LPS are poorly influenced by the presence of sodium azide. a general
1989) to be associated with the microtubules of the cytoskeleton. Further transport of IL-l to the cell surface may be mediated by the microtubular network. Inhibition of this network with colchicine may modify the balance between intracellular and membrane constituents of the cell and. therefore, modify the non-specific interactions between these components and LPS. Opposite effects were observed with dexamethasone. Results in Table 2 show that this glucocorticoid had no effect on the non-specific binding of [3H]Ox-LPS, but strongly reduced (60%) its specific binding.
Binding of LPS (0 mouse macrophages CI/ dijCerent origin
We compared the binding of [“HIOx-LPS to peri- toneal macrophages of Swiss and A,‘J mice, and to the macrophage cell lines P388-Dl and 5774. With the four cell populations, the specific binding of LPS was saturable, and reached its optimal value with 0.2-0.4/~g of labeled ligand per well (results not shown). However, as shown in Table 3. the levels of saturation of P388-Dl and 5774 cells with LPS (I 1.6
B
I * I ._ 0 5 10 RPM1 PBS
CHELATING AGENT (mM) MEDIUM
Fig. 5. Specific binding of [3H]Ox-LPS to peritoneal macrophages (A) in RPM1 supplemented with 10% SVF, in the presence of increasing concentrations of EGTA (a) or EDTA (0). and (B) in RPM1 or PBS
supplemented with 10% HS dialyzed against PBS.

LPS binding to mouse macrophages--I
Table 2. Influence of various pharmacological agents on the binding of [‘HIOx-LPS to macrophages
Non-specific binding Specific binding
Dose
Agent“ (M) c”m % Inhibition c”m % Inhibition
None 2029 + 103 4224 +_ 103
Sodium aride 10-1 2391 f 21 0 3782 +_ 28 13
Colchicine IO h 943 i 40 54 3120166 26
Cycloheximide 7x10 b 2752 + I42 0 2697 + 146 36
Dexamethasone 1.3 x 10-5 2428 t_ 44 0 1701 t_218 60
“Peritoneal macrophages of Swiss mice were incubated for 2 hr at 20 C with the agent and [‘HIOx-LPS
(0.2 pg). in the absence (total bindmg) or m the presence (non-specific binding) of unlabeled LPS (IO pg).
The specific bmding is the calculated difference between total and non-specific bmding. Incubations were
carried out in RPM1 (150~1) supplemented with BSA (0.5 mg) and 10% HS.
and 19 ng/lO’ cells) were markedly lower than those of peritoneal macrophages (41 and 54 ng/106 cells).
Absence of contribution of complement and scatlenger
receptors
It has beenestablished that maleylated-BSA (Somers et al., 1987) and fucoidan, a sulfated fucose-contain- ing algal polysaccharide (Brown et al., 1980) are recognized by the scavenger receptor present on the macrophage membrane, whereas zymosan is a ligand recognized by the receptor (CR3) for the C3Bi con- stituent of complement (Ross et al., 1985). The recog- nition by CR3 of particulate activators of the alterna- tive complement pathway is blocked by zymosan and by some long-chain polysaccharides such as b-glucan (Czop and Austen, 1985) and inulin, an unbranched J-D-(2 --) l)polyfructofuranosyl-cc-D-glucose, which is a storage carbohydrate of dahlias (Cooper and Carter, 1986). To determine whether the scavenger receptor and the complement receptor could contri- bute to the binding of LPS, we analyzed the influence of maleylated BSA, fucoidan, zymosan, and inulin on the binding of [‘HIOx-LPS to peritoneal macrophages of Swiss mice. Results in Table 4 show that zymosan and inulin (ligands of CR3 and inhibitors of alterna- tive pathway activators) fail to inhibit the binding of labeled LPS. On the other hand, some inhibition of LPS binding was observed with high doses (67 pgg!ml) of ligands of the scavenger receptor (maleylated-BSA and fucoidan), but the level of inhibition (10%) was much lower than that (54%) obtained with the same concentration of unlabeled LPS.
Inhibition qf‘ L PSs with diferent polysaccharide chain
lengths
We compared the efficiency of three lipopoly- saccharides (bearing polysaccharides of different
Table 3. Specific binding of [‘HIOx-LPS to mouse macrophages”
Specific binding
Macrophages (n&u06 cells)
Peritoneal macrophages. Swiss mice 41 i4
A/J mice 54 * 2
Cell lines: P388-DI I I.6 F 0.8
5774 I9 + 4
“The bindmg was measured after incubation (2 hr, 22-C) of the cells
(IO6 cells/well) with a saturating concentration (0.4 fig/well) of
[JH]Ox-LPS, in CM containmg 10% HS.
Non-specific binding was estimated by addition of unlabeled (50-fold
excess) LPS. Data represent the arithmetic means + standard
deviations of triplicate determinations.
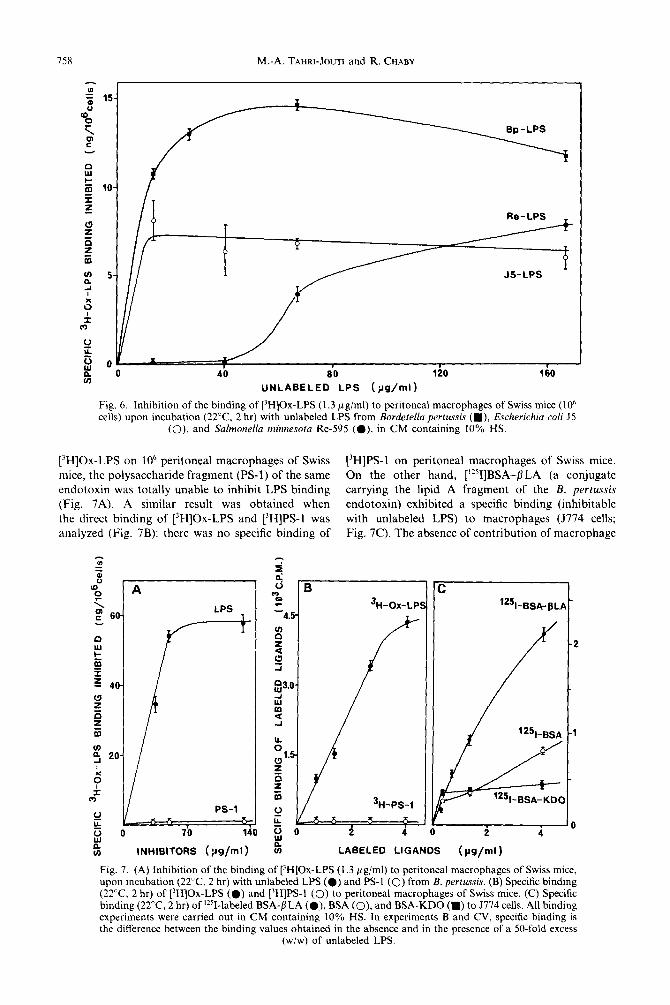
chain length) to inhibit the binding of [3H]Ox-LPS to macrophages. Bordetella pertussis endotoxin (Bp-LPS) carries a polysaccharide consisting of 12 sugar residues (Chaby and Caroff, 1988). The endotoxin of the E. co/i J.5 mutant (JS-LPS) bears an octasaccharide containing three KDO residues (Barclay and Scott, 1987) whereas the polysaccharide region of the endotoxin of an Re mutant (Re-LPS) of Salmonella minnesota (R595) consists exclusively of a KDO disaccharide (Brade and Rietschel, 1984). None of these three endotoxins contained O-repeating units in their polysaccharide chain. Results in Fig. 6 show that a low concentration (40pgg/ml) of unlabeled Re-LPS did not inhibit the binding of [3H]Ox-LPS. The inhibition obtained with JS-LPS was much lower than that induced by the homologous Bp-LPS. However, at high concentrations (167 ,ug/ml) similar inhibitions of the LPS-specific binding were induced by Re-LPS and JS-LPS.
The isolated polysaccharide region does not bind to
mouse macrophages
Whereas unlabeled LPS from B. pertussis
(50-100 @g/ml) inhibited the binding of 55 ng of
Table 4. Influence of ligands of the complement receptor and the
scavenger receptor, on the binding of [3H]Ox-LPS to macrophages”
Binding
Co”ce”tratio”s mhlbitionh
Inhibitors (@g/ml) (%)
LPS 33.3 35.1 + 7
66.7 53.7 _t x
133.4 56.8 * 7
Zymosan 150 O&O
Inulin 33.3 Ok0
66.7 O&O
133.4 o+_o
Maleylated BSA 6.7 058
13.3 0+_4
33.3 Ok3
66.7 IO.1 _+4
Fucoldan 33.3 o_to
66.7 10.5 i: 7
133.4 10.1 16
‘The binding was measured after incubation (2 hr. 22‘C) of thio-
glycollate-elicited peritoneal macrophages of Swiss mice (IO”
cellsjwell) with 0.2~g,kvell of [3H]Ox-LPS. in CM containing
10% HS.
“Inhibitions were calculated as the difference between the arithmetic
means of three determinations obtained in the absence and in the
presence of the inhibitor. Standard deviation of each inhibition
value is the square root of the sum of the squares of each mean’s
standard deviation used to determine the binding inhibition.
Results are expressed as the percentage of the binding observed
in the absence of inhibitor.
M.-A. TAHRI-JOUTI and R. CHABY
UNLABELED LPS (pg/ml)
Fig. 6. Inhibition of the binding of [‘HIOx-LPS (1.3 ~g/ml) to peritoneal macrophages of Swiss mice (lo6 cells) upon incubation (22°C. 2 hr) with unlabeled LPS from Bordetella perrussis (m), Escherichia co/i J5
(0). and Salmonella minnesota Re-595 (0). in CM containing 10% HS.
[‘HIOx-LPS on lo6 peritoneal macrophages of Swiss [‘HIPS-l on peritoneal macrophages of Swiss mice.
mice, the polysaccharide fragment (PS-1) of the same On the other hand, [r2’I]BSA-flLA (a conjugate endotoxin was totally unable to inhibit LPS binding carrying the lipid A fragment of the B. pertussis
(Fig. 7A). A similar result was obtained when endotoxin) exhibited a specific binding (inhibitable
the direct binding of [3H]Ox-LPS and [‘HIPS-l was with unlabeled LPS) to macrophages (5774 cells;
analyzed (Fig. 7B): there was no specific binding of Fig. 7C). The absence of contribution of macrophage
PS-1
P IA INHlalTORS (?e/mt)
‘H-Ox-LP!
: ‘*%BSA-j3LP
LABELED LIGANDS (m/ml 1
Fig. 7. (A) Inhibition of the binding of [-‘HIOx-LPS (1.3 pg/ml) to peritoneal macrophages of Swiss mice, upon incubation (22°C 2 hr) with unlabeled LPS (a) and PS-1 (0) from B. perfussis. (B) Specific binding (22°C 2 hr) of [‘HIOx-LPS (0) and [‘HIPS-l (0) to peritoneal macrophages of Swiss mice. (C) Specific binding (22°C 2 hr) of ‘251-labeled BSA-flLA (0) BSA (0) and BSA-KDO (u) to 5774 cells. All binding experiments were carried out in CM containing 10% HS. In experiments B and CV, specific binding is the difference between the binding values obtained in the absence and in the presence of a 50-fold excess
(w/w) of unlabeled LPS.

LPS binding to mouse macrophages--I 759
binding sites for BSA and KDO to the binding of LPS is assessed by the fact that the specific bindings of [?]BSA and of [‘251]BSA-KD0 were considerably lower than that of [‘251]BSA-PLA.
DISCUSSION
In this paper, we have demonstrated that labeled LPS binds to macrophages both by specific inter- actions (inhibited by unlabeled LPS) and non-specific interactions, probably as a result of the insertion of LPS into the lipid layer of the cellular membrane (Rothfield and Romeo, 1971; Benedetto et al., 1973). LPS-LPS associations, because of hydrophobic inter- actions between the lipid moieties of these molecules, may also contribute to this “non-specific” binding. The specific binding of LPS to mouse peritoneal macrophages appeared to be dose-dependent, time- dependent and saturable (Figs 1 and 2). The tem- perature had also a considerable effect on the kinetics of the specific binding (30 min at 22’ C; 4 hr at 4’C) but had no effect on the saturation value (6 x lo6 molecules/cell). On the other hand, non-specific binding was less temperature dependent than specific binding. It should be noted that above 22°C (and particularly at 37 C) specific binding was completely obscured. This could be because of increased non- specific interactions with the lipid layer, resulting from a modification of the fluidity of the cellular membrane at this temperature.
The number of LPS molecules specifically associ- ated to a cell differs significantly, depending on the type of the macrophages used. We found 6.7 and 8.8 x lOh LPS molecules on thioglycollate-elicited peritoneal macrophages of Swiss and A/J mice, but only 3.1 and 1.9 x 10h molecules on 5774 and P388-Dl macrophage-like cells, respectively. The last two values are very similar to the number of molecules of lipid IV, that bound to RAW 264.7 cells, as determined recently by Hampton et al.
(1988). This also proves that in spite of the presence of L-glycero-D-manno-heptose in all wild-type endo- toxins, LPS-binding is not a result of the presence of the mannose receptor, as this receptor is not expressed by 5774 and P388-D 1 macrophage-like cells (Diment et ul., 1987). The contribution of the “fucose receptor” to the binding of LPS is also very low, as the inhibiting activity of the polysaccharide fucoidan was very low. The number of LPS molecules bound per cell is high compared with the number of other receptors known to be present on macrophages, such as the CSF-1 receptor (5 x lo4 receptors/cell) (Bartelmez et al., 1985), or FcR-II receptors (2-6 x lo5 receptors/cell) (Mellman and Plutner, 1984). This high number of cell-bound LPS can be the result, however, of LPS-LPS associations (Shands et al., 1967). In this regard, specific binding was observed only in the presence of serum. Horse serum was the most effective additive. Recent results (not shown) suggest that the role of serum is to decrease
LPS-LPS interactions and, as a consequence, to increase LPS-membrane interactions.
The specific binding of LPS to macrophages was reversible. We found, however, that the kinetics of the dissociation of the LPS-receptor complex was 30-fold faster than that of the association: at 4’C, 5 min and 2.5 h were required, respectively, for the dissociation and the association of 50% of the optimal amount of cell-bound LPS.
It is well established that macrophages treated with IFN-1/ are more easily activated by LPS (Adams and Hamilton, 1984). On the other hand, Boraschi and Meltzer (1979) observed that peritoneal macrophages from A/J mouse were not activable by LPS, and Hamilton et al. (1986) reported that A/J macrophages, although expressing a receptor for IFN-y, were unresponsive to this ligand. It was therefore important to determine if A/J macrophages express LPS receptors. We found (Table 3) a high specific binding of LPS to A/J macrophages. This finding is in consonance with the observation (Boraschi and Meltzer, 1979) that the deficiency of A/J macrophages is independent of the LPS gene. and with our previous results which established that the levels of TNF secreted by A/J macrophages during stimulation with LPS were similar to those obtained with macrophages of conventional mice (Lasfargues and Chaby, 1988).
We also examined the influence of dexamethasone
(DEX), a glucocorticoid which has been shown to modulate certain macrophage functions, and particularly to block the production of interleukin 1 (Snyder and Unanue, 1982) tumor necrosis factor (Beutler ef al., 1986) and prostaglandins (Wahl and Wahl, 1985). As regards membrane components of macrophages, it has been established that DEX treat- ment inhibits the expression of Ia antigen (Warren and Vogel, 1985) but enhances the number of Fc receptors (Girard et a/., 1987) and scavenger recep- tors (Hirsch and Mazzone, 1986). As regards LPS- binding sites on macrophages, we observed in this study (Table 2) that DEX has no effect on the non-specific binding of LPS, but strongly reduced (60%) its specific binding. This result might explain the blocking effect of DEX on LPS-induced IL- I and TNF production mentioned above. It is noteworthy that DEX, which has been shown here to inhibit the expression of LPS-binding sites on macrophages, en- hances the expression of scavenger receptors (Hirsch and Mazzone, 1986), whereas LPS prevents the ex- pression of these receptors (Van Lenten et ul., 1985).
As regards the putative role of integrins (adhesion promoting receptors: CR3; LFA-1; ~150, 95) in LPS binding, although Wright and Jong (1986) have shown that integrins recognize LPS, our result suggest that at least one of these. CR3, is not a specific binding site for LPS, as zymosan (a ligand of CR3) and inulin (another activator of the alternative complement pathway) did not inhibit the specific binding of LPS to macrophages. The specific receptor

760 M.-A. TAHRI-JOUTI and R. CHABY
for LPS may be another membrane entity, as cells Brade H. and Rietschel E-T. (1984) ~(-2 + 4-Interlinked
that do not express CR3, LFA-I or ~150, 95, such 3-deoxy-D-manno-octulosonic acid disaccharide. A com-
as endothelial cells, do show marked physiological mon constituent of enterobacterial lipopolysaccharides. Eur. J. Biochem. 145, 231-236.
responses to LPS (Schleimer and Rutledge, 1986). B rown M. S.. Basu S. K.. Falck J. R., Ho Y. K. and
In this uaver we also show that the nolvsaccharide Goldstein J. L. (1980) The scavenger cell pathway for I . . -
region of LPS is not involved in the binding of this lipoprotein degradation: specificity of the binding site that
molecule to murine macrophages. This result conflicts mediates the uptake of negatively-charged LDL by
with a previous observation (Haeffner-Cavaillon et macrophages. J. .wpramolec. Struct. 13, 67- 8 1.
Butler P. J. G. and Hartlev B. S. (1972) Malevlation of al.. 1982) of a lectin-like receptor for B. pertussis LPS on rabbit peritoneal macrophages. It is, however, noteworthy that contrary to mice, rabbits are
very frequent hosts of B. bronchiseptica, a bacterium carrying a lipopolysaccharide which exhibits a strong serological cross-reactivitv with the B. aertussis
LPS. It is therefore reasonable to assume that in addition to an ubiquitous anti-lipid A receptor, the macrophages of some species (human, rabbit) may
Chaby R. and Caroff M. (1988) Lipopolysaccharides
Caroff M., T&ken A. and Szdbo L. (1988) Detergent amino groups. Melh. E&bm. 25B; 191-197.
accelerated hydrolysis of bacterial endotoxins and deter-
of Bordetella pertussis endotoxin. In Puthogenesis and
mination of the anomeric configuration of the glycosyl-
Immunity in Pertussis (Edited by Wardlaw A. C. and Parton R.). pp. 247-271. John Wilev. New York.
phosphate present in the “isolated Lipid A” fragment of the Borderella pertussis endotoxin. Carhohvdr. Res. 175. 273.-282.
I . . also express a lectin-like receptor which interacts Chaby R. and Szabo L. (1975) 3-Deoxy-2-octulosonic acid
with the polysaccharide region bf some, but not all, 5-phosphate: a component of the endotoxin of Bordetellu
endotoxins. perfussis. Eur. J. Biochem. 59, 2777280. Chong A. S. F. and Parish C. R. (1986) Cell surface
Acknowledgements-This work was supported by the Centre receptors for sulfated polysaccharides-a potential
National de la Recherche Scientifique. M. A. Tahri-Jouti marker for macrophage subsets. immunology 58,277-284.
was supported in part by a Morocco Government fellow- Cooper P. D. and Carter M. (1986) Anti-complementary
ship, and performed this work in fulfilment of the Ph.D. action of polymorphic “solubility forms” of particulate
requirements at the Universite de Paris-Sud, Orsay, France. inulin. Molec. Immun. 23, 895-901.
Cracker P. R.. Morris L. and Gordon S. (1988) Novel cell surface adhesion receotors involved in interactions between stromal macrophages and hematopoietic cells. J. Cell Sci. Suppl. 9, 185 -206.
Czop J. K. and Austen K. F. (1985) A beta-glucan inhibitable receptor on human monocytes: its identity with the phagocyte receptor for particulate activators of the alternative complement pathway. J. Immun. 134, 258882593.
REFERENCES
Adams D. 0. and Hamilton T. A. (1984) The cell biology of macrophage activation. Ann. Rec. Immun. 2, 283-318.
Aderem A. A., Cohen D. S., Wright S. D. and Cohn Z. A. (1986) Bacterial lipopolysaccharides prime macrophages for enhanced release of arachidonic acid metabolites. J. rsp. Med. 164, 1655179.
Anderson J., Sjoberg 0. and Moller G. (1972) Induction of immunoglobulin and antibody synthesis in vitro by lipopolysaccharides. Eur. J. Zmmun. 2, 349-353.
Baldari C. T. and Telford J. L. (1989) The intracellular precursor of IL-l/I is associated with microtubules in acivated U937 cells. J. Immun. 142, 785-791.
Barclay G. R. and Scott B. B. (1987) Serological relation- ships between Escherichiu co/i and Salmonella smooth- and rough-mutant lipopolysaccharides as revealed by enzyme-linked immunosorbent assay for human immuno- globulin G antiendotoxin antibodies. Infect. Immun. 55, 2706-2714.
Bartelmez S. H., Sacca R. and Stanley E. R. (1985) Lineage specific receptors used to identify a growth factor for developmentally early hematopoietic cells: assay for hemopoietm2. J. cell. Physiol. 12i, 362--369.
Benedetto D. A.. Shands 1. W. and Shah D. 0. (1973) Interaction of bacterial lipopolysaccharide with phdspho: lipid bilayers and monolayers. Biochim. biophys. Acta 298, 145-157.
Beutler B., Krochin N., Milsark I. W., Leudke C. and Cerami A. (1986) Control of cachectin (tumor necrosis factor) synthesis: mechanisms of endotoxin resistance. Science 232, 977-980.
Beutler B.. Mahoney J., LeTrang N., Pekala P. and Cerami A. (1985) Purification of cachectin, a lipoprotein lipase- suppressing hormone secreted by endotoxin-treated RAW 264.7 cells. J. exp. Med. 161, 984-995.
Boraschi D. and Meltzer M. S. (1979) Defective tumoricidal capacity of macrophages from A/J mice. II. Comparison of the macrophage cytotoxic defect of A/J mice with that of Lipid A-unresponsive C3HIHeJ mice. J. Immun. 122, 1592m-1597.
Diment S., Leech M. and Stahl P. D. (1987) Generation of macrophage variants with 5-azdcytidine: selection for mannose receptor expression. J. Leukoqte Biol. 42, 485490.
Duggleby R. G. (1981) A nonlinear regression program for small computers. Analyt. Biochem. 110, 9-18.
Durum S. K., Schmidt J. A. and Oppenheim J. J. (1985) Interleukin-1: an immunological perspective. Ann. Rev. Immun. 3, 263-287.
Euteneuer B., Storkel S. and Loos M. (1986) Contributions of Clq, bacterial lipopolysaccharide. and porins during attachment and ingestion phases of phagocytosis by murine macrophages. InJect. Immun. 51, X07-8 15.
Ezekovitz R. A. B. and Stahl P. D. (1988) The structure and function of vertebrate mannose lectin-like proteins. J. Cell Sci. Suppl. 9, 121-133.
Gianturco S. H., Brown S. A., Via D. P. and Bradley W. A. (1986) The b-VLDL receptor pathway of murine P388D, macrophages. J. Lipid Res. 27, 412430.
Girard M. T., Hjaltadottir S., Fejes-Toth A. N. and Guyre P. M. (1987) Glucocorticoids enhance the y-interferon augmentation of human monocyte immunoglobulin G Fc receptor expression. J. Immun. 138, 3235-3241.
Gordon S., Perry V. H., Rabinowitz S., Chung L-P. and Rosen H. (1988) Plasma membrane receptors of the mono- nuclear phagocyte system. J. Cell Sci. Suppl. 9, l-26.
Haeffner-Cavaillon N., Cavaillon J. M., Etievant M., Lebbar S. and Szabo L. (1985) Specific binding of endo- toxin to human monocytes and mouse macrophages: serum requirement. Cell. Immun. 91, 119- 131.
Haeffner-Cavaillon N., Chaby R.. Cavaillon J-M. and Szabo L. (1982) Lipopolysaccharide receptor on rabbit peritoneal macrophages. I. Binding characteristics. J. Immun. 128, 1950-1954.

LPS binding to mouse macrophages-I 761
Hamilton T. A., Somers S. D., Becton D. L., Celada A., Schleimer R. P. and Rutledge B. K. (1986) Cultured human Schreiber R. D. and Adams D. 0. (1986) Analysis of vascular endothelial cells acquire adhesiveness for neutro- deficiencies in IFN-y-mediated priming for tumor cyto- phils after stimulation with interleukin 1, endotoxin, and toxicity in peritoneal macrophages from A/J mice. tumor-promoting phorbol diesters. J. Zmmun. 136, J. Immun. 137, 3367-3371. 649-654.
Hampton R. Y., Golenbock D. T. and Raetz C. R. H. (1988) Lipid A binding sites in membranes of macrophage tumor cells. J. biol. Chem. 263, 4802-4807.
Hirsch L. J. and Mazzone T. (1986) Dexamethasone modulates lipoprotein metabolism in cultured human monocyte-derived macrophages. J. c/in. Invest. 77, 485490.
Shands J. W., Graham J. A. and Nath K. (1967) Morpho- logic structure of isolated bacterial lipopolysaccharide. J. molec. Biol. 25, 15-21.
Snyder D. S. and Unanue E. (1982) Corticosteroids inhibit macrophage Ia expression and interleukin 1 production. J. Immun. 129, 1803-1805.
Hunter W. M. and Greenwood F. C. (1962) Preparation of iodine-13 I labeled human growth hormone of high specific activity. Nature, Lond. 194, 495496.
Johnson A. G. (1983) Adjuvant action of bacterial endo- toxins on antibody formation. A historical perspective. In Beneficial Eftecr.7 of Endoloxins (Edited by Nowotny A.), p. 12. Plenum Press. New York.
Lasfareues A. and Chabv R. (1988) Endotoxin-induced
Somers S. D., Hamilton T. A. and Adams D. 0. (1987) Maleylated bovine serum albumin triggers cytolytic function in selected populations of primed murine macrophages. J. Immun. 139, 136 l-l 368.
Tahri-Jouti M-A., Mondange M. Le Dur A., Auzanneau F-I.. Charon D.. Girard R. and Chabv R. (1990) Soecific binding of lipopolysaccharides to mouse macrophages. II. Involvement of distinct lipid A substructures. Mo/ec. Zmmun. 27, 763-770.
tumor necrosis factor -(TNF): selective triggering of TNF and interleukin-1 production by distinct gluco- samine-derived lipids. CeN Immun. 115, 165-178.
Lasfargues A., Le Dur A., Charon D., Szabo L. and Chaby R. (1987) Induction by lipopolysaccharide of intra- and extracellular interleukin 1 production: analysis with synthetic models. J. Immun. 135, 429436. _
Lasfaraues A.. Tahri-Jouti M. A.. Pedron T.. Girard R. and Chalky R. (1989) Effects of lipopolysaccharide on macrophages analyzed with anti-Lipid A monoclonal antibodies and polymyxin B. Eur. J. Immun. 19, 22 19%2225.
Le Dur A., Caroff M., Chaby R. and Szabo L. (1978) A novel type of endotoxin structure present in Bordetellu pertussis. Isolation of two different lipopolysaccharides bound to lipid A. Eur. J. Biochem. 84, 579-589.
Mellman I. and Plutner H. (1984) Internalization and degradation of macrophage Fc receptors bound to poly- valent immune complexes. J. ceN. Biol. 98. 117&l 177.
Milner E. C. B., Rudbach J. A. and Von Eschen K. B. (1983) Cellular responses to bacterial lipopolysaccharide: T cells recognize LPS determinants. Stand. J. Zmmun. 18, 21-28.
Morrison D. C. and Ulevitch R. J. (1978) The effects of bacterial endotoxins on host mediating systems. Am. J. Path. 93, 527-536.
Ross G. D.. Cain J. A. and Lachmann P. J. (1985) Membrane complement receptor type three (CR,) has lectin-like properties analogous to bovine conglutinin and functions as a receptor for zymosan and rabbit erythrocytes as well as a receptor for iC3b. J. Immun. 134, 3307-3315.
Rothfield L. and Romeo D. (1971) Role of lipids in biosynthesis of bacterial cell envelope. Boct. Rev. 35, 14 -38.
Tomita T., Blumenstock E. and Kanegasaki S. (1981) Phagocytic and chemiluminescent responses of mouse peritoneal macrophages to living and killed Sabnonefiu typhimurium and other bacteria. Infect. Immun. 32, 124221248.
Van Lenten B. J., Fogelman A. M., Seager J., Ribi E., Haberland M. E. and Edwards P. A. (1985) Bacterial endotoxin selectively prevents the expression of scavenger- receptor activity on human monocyte-macrophages. J. Immun. 134, 3718-3721.
Wahl S. M. and Wahl L. M. (1985) Regulation of macrophage collagenase, prostaglandins and fibroblast- activating factor production by anti-inflammatory agents: different regulatory mechanisms for tissue injury and repair. Cell. Immun. 92, 302-312.
Warner J. C., Savage N. and Mitchell D. (1985) Character- istics of lipopolysaccharide interaction with human peripheral-blood monocytes. Biochem. J. 232, 379-383.
Warren M. K. and Vogel S. N. (1985) Opposing effects of glucocorticoids on interferon-y induced murine macro- phages Fc receptor and Ia antigen expression. J. immun. 134, 2462-2469.
Watson J. and Riblet R. (1975) Genetic control of responses to bacterial lipopolysaccharides in mice II. A gene that influences a membrane component involved in the activation of bone-marrow-derived lymphocytes by lipopolysaccharides. J. Immun. 114, 146221468.
Westphal O., Lilderitz 0. and Bister F. (1952) Uber die Extraktion von Bacterien mit Phenol-Wasser. Z. Naturforsch. 7b, 148-155.
Wright S. D. and Jong M. T. C. (1986) Adhesion-promoting receptors on human macrophages recognize E. coli by binding to lipopolysaccharide. J. exp. Med. 164, 18761888.
Copyright © 2022 FDOKUMEN




















