
Tumor-Associated Macrophages in the Cutaneous SCC Microenvironment Are Heterogeneously Activated
17
Tumor-associated macrophages in the cutaneous SCC microenvironment are heterogeneously activated Julia S. Pettersen 1 , Judilyn Fuentes-Duculan 2 , Mayte Suárez-Fariñas 2,3 , Katherine C. Pierson 2 , Alexander Pitts-Kiefer 1 , Linda Fan 1 , Daniel A. Belkin 1 , Claire Q.F. Wang 2 , Shivaprasad Bhuvanendran 4 , Leanne M. Johnson-Huang 2 , Mark J. Bluth 1 , James G. Krueger 2 , Michelle A. Lowes 2 , and John A. Carucci 1 1 Department of Dermatology, Weill Medical College of Cornell University, New York, NY 2 Laboratory of Investigative Dermatology, The Rockefeller University, New York, NY 3 The Center for Clinical and Translational Science, The Rockefeller University, New York, NY 4 Bio-Imaging Resource Center, The Rockefeller University, New York, NY Abstract Tumor-associated macrophages (TAMs) may play an important role in tumor immunity. We studied the activation state of TAMs in cutaneous SCC, the second most common human cancer. CD163 was identified as a more abundant, sensitive, and accurate marker of TAMs, compared to CD68. CD163 + TAMs produced pro-tumoral factors MMP9 and MMP11, at the gene and protein levels. Gene set enrichment analysis (GSEA) was used to evaluate M1 and M2 macrophage gene sets in the SCC genes and to identify candidate genes in order to phenotypically characterize TAMs. There was co-expression of CD163 and alternatively activated “M2” markers, CD209 and CCL18. There was enrichment for classically activated “M1” genes in SCC, which was confirmed in situ by co-localization of CD163 and phosphorylated STAT1, IL-23p19, IL-12/IL-23p40, and CD127. Also, a subset of TAMs in SCC was bi-activated as CD163 + cells expressed markers for both M1 and M2, shown by triple-label immunofluorescence. These data support heterogeneous activation states of TAMs in SCC, and suggest that a dynamic model of macrophage activation would be more useful to characterize TAMs. Keywords cutaneous squamous cell carcinoma; CD163; macrophages; skin INTRODUCTION Cutaneous squamous cell carcinoma (SCC) is the second most common human cancer, affecting more than 300,000 individuals in the United States annually (Brantsch et al., 2008; Weinberg et al., 2007). Although most cases can be treated successfully by surgical removal, certain aggressive cases can cause extensive tissue destruction and metastasize to local lymph nodes and distant organs. These aggressive cases are responsible for approximately 10,000 non-melanoma skin cancer deaths in the US each year. Aggressive behavior by SCC is observed in solid organ transplant recipients (Carucci, 2004). Based on the potential for Corresponding author: John A. Carucci, Weill Medical College of Cornell University, Department of Dermatology, 1305 York Ave, New York, NY, 10021, Ph: 646-962-5574, Fax: 646-962-0004, [email protected]. Conflict of Interest The authors do not have financial interests related to this work. NIH Public Access Author Manuscript J Invest Dermatol. Author manuscript; available in PMC 2012 April 23. Published in final edited form as: J Invest Dermatol. 2011 June ; 131(6): 1322–1330. doi:10.103/jid.2011.9. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
rockefeller -
Category
Documents
-
view
0 -
download
0
Transcript of Tumor-Associated Macrophages in the Cutaneous SCC Microenvironment Are Heterogeneously Activated

Tumor-associated macrophages in the cutaneous SCCmicroenvironment are heterogeneously activated
Julia S. Pettersen1, Judilyn Fuentes-Duculan2, Mayte Suárez-Fariñas2,3, Katherine C.Pierson2, Alexander Pitts-Kiefer1, Linda Fan1, Daniel A. Belkin1, Claire Q.F. Wang2,Shivaprasad Bhuvanendran4, Leanne M. Johnson-Huang2, Mark J. Bluth1, James G.Krueger2, Michelle A. Lowes2, and John A. Carucci11Department of Dermatology, Weill Medical College of Cornell University, New York, NY2Laboratory of Investigative Dermatology, The Rockefeller University, New York, NY3The Center for Clinical and Translational Science, The Rockefeller University, New York, NY4Bio-Imaging Resource Center, The Rockefeller University, New York, NY
AbstractTumor-associated macrophages (TAMs) may play an important role in tumor immunity. Westudied the activation state of TAMs in cutaneous SCC, the second most common human cancer.CD163 was identified as a more abundant, sensitive, and accurate marker of TAMs, compared toCD68. CD163+ TAMs produced pro-tumoral factors MMP9 and MMP11, at the gene and proteinlevels. Gene set enrichment analysis (GSEA) was used to evaluate M1 and M2 macrophage genesets in the SCC genes and to identify candidate genes in order to phenotypically characterizeTAMs. There was co-expression of CD163 and alternatively activated “M2” markers, CD209 andCCL18. There was enrichment for classically activated “M1” genes in SCC, which was confirmedin situ by co-localization of CD163 and phosphorylated STAT1, IL-23p19, IL-12/IL-23p40, andCD127. Also, a subset of TAMs in SCC was bi-activated as CD163+ cells expressed markers forboth M1 and M2, shown by triple-label immunofluorescence. These data support heterogeneousactivation states of TAMs in SCC, and suggest that a dynamic model of macrophage activationwould be more useful to characterize TAMs.
Keywordscutaneous squamous cell carcinoma; CD163; macrophages; skin
INTRODUCTIONCutaneous squamous cell carcinoma (SCC) is the second most common human cancer,affecting more than 300,000 individuals in the United States annually (Brantsch et al., 2008;Weinberg et al., 2007). Although most cases can be treated successfully by surgical removal,certain aggressive cases can cause extensive tissue destruction and metastasize to locallymph nodes and distant organs. These aggressive cases are responsible for approximately10,000 non-melanoma skin cancer deaths in the US each year. Aggressive behavior by SCCis observed in solid organ transplant recipients (Carucci, 2004). Based on the potential for
Corresponding author: John A. Carucci, Weill Medical College of Cornell University, Department of Dermatology, 1305 York Ave,New York, NY, 10021, Ph: 646-962-5574, Fax: 646-962-0004, [email protected].
Conflict of InterestThe authors do not have financial interests related to this work.
NIH Public AccessAuthor ManuscriptJ Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
Published in final edited form as:J Invest Dermatol. 2011 June ; 131(6): 1322–1330. doi:10.103/jid.2011.9.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the host immunity to regulate tumor behavior in SCC, it is important to characterize thetumor-associated immune microenvironment.
Macrophages are one of the major populations of tumor-infiltrating leukocytes associatedwith solid tumors (Gordon and Taylor, 2005). Macrophages that infiltrate and surroundtumor nodules are defined as tumor-associated macrophages (TAMs) (Wang et al., 2010),and different studies have shown that macrophages may either inhibit or stimulate tumorgrowth. Initially, TAMs were shown to participate in the early eradication of tumor cells invitro (Romieu-Mourez et al., 2006). However, other studies have suggested that TAMs maycontribute to carcinogenesis, as there is a positive correlation between increased numbers ofTAMs and poor prognosis in some human cancers (Bingle et al., 2002; El-Rouby, 2010;Leek et al., 1996; Lin and Pollard, 2007; Nonomura et al., 2010; Shabo et al., 2008; Sica etal., 2006; Steidl et al., 2010). TAMs can fail to recognize tumor antigens (Fadok et al., 1998)and may release factors that directly stimulate tumor growth and angiogenesis (Lin et al.,2006; Lin and Pollard, 2007). Furthermore, the tumor itself can create a dynamicmicroenvironment that can transform TAMs (Gocheva et al., 2010). Thus, TAMs in the SCCmicroenvironment may be associated with tumor growth.
Currently, the general classification of macrophage activation parallels the Th1/Th2paradigm, defining classically activated (M1) and alternatively activated (M2) cells(Mantovani et al., 2004; Mosser and Edwards, 2008). Classically activated macrophages areinduced by IFNγ and have a high capacity to present antigen. Alternatively activatedmacrophages are induced by the cytokine IL-4, which promotes Type 2 responses. As SCCsusually progress, in association with a Th2 microenvironment and low levels of IFNγ, it isthought that the net immune response is ineffective at suppressing tumor growth. Hence,TAMs have commonly been considered alternatively activated or strongly skewed to the M2phenotype (Biswas et al., 2006; Gordon and Martinez, 2010; Martinez et al., 2009; Siveenand Kuttan, 2009). However, there has been renewed debate over the phenotypic activationof TAMs as the physiology of these macrophages has been shown to change over time andto demonstrate remarkable plasticity (Mosser and Edwards, 2008).
Given the importance of TAMs contributing to tumor growth, and the current conflictingstate of the understanding of TAM activation, we set out to phenotypically characterizemacrophages in SCC. Initially, we used a non-biased genomic approach to guide our choiceof markers to further evaluate TAMs. “M1” and “M2” activated macrophage gene sets(Fuentes-Duculan et al., 2010) were analyzed in our SCC genomic phenotype (Haider et al.,2006) using gene set enrichment analysis (GSEA) (Bluth et al., 2009; Subramanian et al.,2005). We then identified candidate genes that were expressed in the macrophage and SCCgene sets and performed immunofluorescence on SCCs versus CD163, as our constitutivemacrophage marker.
Previously, we have shown that in normal skin and psoriasis, CD163 is the most usefulmarker of dermal macrophages (Zaba et al., 2007). We expanded on that work tocharacterize tumor-associated macrophages in human SCC. We found the following: (1)Compared to CD68, CD163 was a more abundant, sensitive, and accurate marker of TAMs;(2) There was an increase in the pro-tumoral factors MMP9 and MMP11 in SCC, andCD163+ TAMs produced MMP9 and MMP11; (3) There was co-expression of CD163 andalternatively activated “M2” markers, CD209 and CCL18; (4) There was enrichment forclassically activated “M1” genes in SCC, which was confirmed in situ by co-localization ofCD163 and phosphorylated STAT1, IL-23p19, IL-12/IL-23p40, and CD127; and (5) Asubset of TAMs in SCC was bi-activated as CD163+ cells expressed markers for both M1and M2, shown by triple-label immunofluorescence. These data support heterogeneousactivation states of TAMs in SCC, and suggest that a dynamic model of macrophage
Pettersen et al. Page 2
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

activation would be more useful to characterize TAMs. Furthermore, driving TAMactivation toward a more dominant anti-cancer phenotype might be a potential therapeuticstrategy.
RESULTSMacrophages were more abundant in SCC compared to normal skin
Macrophages were quantified in SCC and normal skin (n= 8–18) using CD163, which weconsider a reliable marker of macrophages in normal skin and psoriasis, and CD68, thewidely accepted macrophage marker (Bluth et al., 2009; Fuentes-Duculan et al., 2010; Zabaet al., 2007). Representative immunohistochemistry is shown for CD163 and CD68, and cellcounts of the cases are presented (Figure 1a–b). The vast majority of CD163+ and CD68+
macrophages were surrounding, rather than infiltrating, the SCC tumor nests, and bothCD163+ and CD68+ macrophages were significantly increased, approximately two-fold, inSCC compared to normal skin (p<0.001 for both). Additionally, using double-labelimmunofluorescence, CD163 co-localized with CD68 but there were CD163+ cells that didnot co-express CD68, suggesting that CD163 is a more robust and sensitive marker ofmacrophages in the skin than CD68 (Figure 1c). We also evaluated the co-expression ofCD163 with the well-known dendritic cell (DC) marker, CD11c (Bluth et al., 2009). As wehave previously shown in normal skin (Zaba et al., 2007), CD163+ cells in SCC also did notco-localize with this DC marker (Figure 1d), demonstrating that these are two distinctleukocyte populations. In contrast, CD68+ cells close to SCC tumor nests did show co-localization with CD11c (Figure 1e).
SCC TAMs expressed pro-tumoral products in the tumor microenvironmentTAMs may produce factors that encourage tumorigenesis. We have shown SCC TAMsproduce the pro-lymphangiogenic factor VEGF-C, which favors tumor growth anddevelopment (Moussai et al., 2011). Matrix metalloproteinases (MMPs), enzymes that maycontribute to tumor invasion by degrading the matrix surrounding tumor nodules, may alsobe produced by TAMs in SCC. MMP1, MMP10 and MMP13 genes have been shown to beupregulated in SCC (Haider et al., 2006). MMP9 (gelatinase B) and MMP11 (stromelysin-3)proteins correlate with increased tumor aggressiveness (Buergy et al., 2009; Pinto et al.,2003; Shah et al., 2010; Steidl et al., 2010; Zhao et al., 2010). Neither MMP9 nor MMP11have been classified as products representative of either state of macrophage activation. Weshowed that there was increased gene expression by RT-PCR of MMP9 in SCC compared toadjacent non-tumoral skin and normal skin (p=0.07 and 0.008, respectively) (Figure 2a).Similarly, MMP11 was also increased in SCC compared to adjacent non-tumoral skin andnormal skin (p=0.003 and 0.025, respectively) (Figure 2b). We then asked whether TAMscould be a possible source of the increased MMP9 and MMP11. There was abundant co-localization of MMP9 and MMP11 with CD163+ macrophages in SCCs compared to normalskin (Figure 2c–d). These findings suggest that TAMs secrete pro-tumoral products in theSCC microenvironment.
SCC TAMs expressed products of alternatively activated macrophagesWe evaluated expression of phosphorylated STAT6 (STAT6p) based on the association ofTh2 cells and the tumor microenvironment (de Oliveira et al., 2009; Todaro et al., 2008).STAT6 plays an important role in signalling pathways that lead to the differentiation of Th2cells, and STAT6p translocates to the nucleus in IL-4-activated cells (Forbes et al., 2010;Takeda et al., 1996). STAT6p co-localization with CD163 was abundant in theinflammatory infiltrate associated with SCC compared to normal skin (Figure 3a),suggesting the presence of IL-4 activation in TAMs.
Pettersen et al. Page 3
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

To further evaluate the tumor microenvironment, we used M1 and M2 gene sets to correlatewith the SCC genomic phenotype by using gene set enrichment analysis (GSEA) (Bluth etal., 2009; Lamb et al., 2006; Subramanian et al., 2005). We have used this approachpreviously (Bluth et al., 2009), and described it thoroughly in a prior publication (Suarez-Farinas et al., 2010). Fuentes-Duculan et al. recently published these sets of genes defining“M1” macrophages, induced with IFNγ, and “M2” macrophages, induced with IL-4,compared with control (Fuentes-Duculan et al., 2010). We hypothesized that there should begreater expression of M2 macrophage genes in the SCC genomic phenotype (Martinez et al.,2009; Siveen and Kuttan, 2009), defined by the SCC versus normal skin genes(Subramanian et al., 2007). However, the M2 gene set was not significantly enriched in SCCgenomic phenotype, which may reflect that the M2 gene set is similarly expressed in bothSCC and normal skin.
Despite the lack of enrichment of M2 genes in the SCC transcriptome, some published M2genes (Martinez et al., 2006) were upregulated in our M2 gene set (Table 1), includingCD209 (DC-SIGN) (Puig-Kroger et al., 2004; Soilleux et al., 2002), CCL17 (Bonecchi etal., 1998; Katakura et al., 2004) and CCL18 (Gustafsson et al., 2008; Kodelja et al., 1998;Kwan et al., 2008; Mantovani et al., 2004; Martinez et al., 2006). By double labelimmunofluorescence, CD163+ cells co-localized with CD209 and CCL18 in SCC and to alesser extent in normal skin (Figure 3b–c). In the SCC microenvironment, we have thusshown that TAMs expressed some surface markers (CD209) and chemokines (CCL18) ofM2-type macrophages. In addition and consistent with previous findings (Schutyser et al.,2005), we showed that in normal skin macrophages at steady state are in an alternativelyactivated state.
SCC TAMs expressed characteristics of classically activated macrophages in amicroenvironment with Type-1 activation
While there was not any significant enrichment of M2 macrophage genes in the SCCgenomic phenotype, the GSEA results did indicate that M1 macrophage gene sets weresignificantly enriched in the SCC genomic phenotype (Supplemental Table 1). Table 1 listsselected genes that were upregulated in both the M1 gene sets and the SCC versus normalskin transcriptome, indicating an M1-type macrophage activation pattern in the SCCmicroenvironment. It was perhaps surprising that there was such a marked M1 genomicsignature in SCC. However, several recent studies have shown that IFNγ-producing T cellscan be found in SCCs and other tumors (Huang et al., 2009; Koller et al.; Kryczek et al.,2009; Li-Weber and Krammer, 2003).
To confirm that macrophages in the SCC tumor microenvironment could be responsive toIFNγ, we performed double label immunofluorescence with CD163 and the two requisitechains of the IFNγ receptor. The IFNγ receptor 1 subunit (IFNγR1) that is internalizedupon binding with IFNγ (Schroder et al., 2004) was present on the majority of macrophagesin SCC compared to macrophages in normal skin, which did not express this receptor(Figure S1a). There is a second chain of IFNγ receptor (IFNγR2), which showed a similarpattern of co-localization with CD163+ cells in SCC compared to normal skin (Figure S1b).CD68+ macrophages in SCC also strongly expressed IFNγR1 and IFNγR2 compared tonormal skin (Figure S1c–d). These data indicate that SCC TAMs are capable of respondingto IFNγ. STAT1 and Mx-1, well-recognized IFNγ-induced genes (Landolfo et al., 1995;Saha et al.), were upregulated in the M1 gene set and SCC transcriptome (Table 1).Phosphorylated STAT1 (STAT1p), the form of STAT1 that translocates to the nucleus inIFNγ-activated cells, showed minimal expression in normal skin, but co-localized withCD163 in the juxtatumoral dermis of SCC (Figure 4a), indicating CD163+ macrophages inSCC were indeed responding to IFNγ.
Pettersen et al. Page 4
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

To further classify these SCC TAMs, the expression of markers and cytokines on TAMsconsidered to be representative of classical/M1 macrophage activation were evaluated(Table 1) (Martinez et al., 2006). There was increased gene expression of the marker CD127(IL-7 receptor) and the cytokine subunits, IL-23p19 and IL-12/23p40 in the M1 macrophagegene set and increased gene expression of IL-23p19 in the SCC transcriptome (Table 1). Theexpression of CD127 has been shown to be strongly down regulated by IL-4 (Crawley etal.), which supports its role as an M1 marker. We confirmed these findings at the proteinlevel by double label immunofluorescence. CD163+ TAMs abundantly co-expressed CD127and produced IL-23p19 and IL-12/IL-23p40 compared to normal skin CD163+ macrophages(Figure 4b–d). Thus, in the SCC microenvironment, TAMs express surface markers(CD127) and cytokines (IL-23 subunits) of M1-type activated macrophages.
A subset of SCC TAMs simultaneously expressed markers of both classical andalternative activation
We have shown that TAMs in the SCC environment exhibit characteristics of both classicaland alternative activation. However, it was not clear if there were two separate populationsof macrophages driven by each cytokine, or if one subset of macrophages was responding toboth cytokines simultaneously. Using triple label immunofluorescence and confocalimaging, some CD163+ cells were identified that co-localized with both STAT1p andSTAT6p (shown by white arrows, Figure 5a), indicating that some SCC TAMs respond toboth Th1 and Th2 signals and that there is a complex SCC tumor microenvironment ofmixed Th1 and Th2 activation.
To further evaluate the TAM phenotype in this setting, we also identified a subpopulation ofCD163+ macrophages that co-expressed both CD127 (upregulated in M1 macrophages) andCD209 (upregulated in M2 macrophages) (shown by white arrows, Figure 5b). There werealso CD163+ cells that co-localized with CD127 only (yellow cells) and CD163+ cells thatco-localized with CD209 (dark teal cells). This suggests that macrophage activation in SCCis heterogeneous, as we found several types of TAMs: M1 macrophages responding to Th1signals, M2 macrophages responding to Th2 signals, and bi-activated macrophagesresponding to both Th1 and Th2 cytokines simultaneously.
DISCUSSIONOur studies suggest that CD163 should be considered the superior marker to identify TAMsin SCC. First, CD163+ TAMs were prominent and more abundant in the cutaneous SCCtumor microenvironment and significantly increased compared to macrophages in normalskin. Second, CD163 is the most sensitive marker for TAMs in SCC, as it identifies moredermal macrophages surrounding the tumor nodules than CD68. Third, CD163 is a moreaccurate marker of TAMs, because it did not co-localize with CD11c+ DCs in SCC,compared to CD68, which showed some overlap. This is similar in psoriasis, where CD163had the least overlap with CD11c (Fuentes-Duculan et al., 2010). In addition, althoughCD163 was previously considered a macrophage marker solely for alternative activation, wehave shown that CD163 not only identifies alternatively activated macrophages, but alsoclassically and bi-activated macrophages in SCC. These findings suggest that future studiescould benefit from using CD163 as a pan-macrophage marker in the skin.
To understand the cellular state of macrophage activation in the cutaneous SCCmicroenvironment, we must consider the setting in which these TAMs exist (Figure 5c). Thetumor microenvironment is defined as a mixture of tumor and non-tumor cells at thedynamic interface of neoplasia (van Kempen et al., 2003). Prior studies have documented aninflux of IFNγ-producing cells in the tumor microenvironment (Kryczek et al., 2009). Wefound a strong IFNγ genomic signature in the SCC tumor microenvironment coupled with
Pettersen et al. Page 5
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

evidence of upregulated IFNγ receptors and abundant IFNγ-activation of the infiltratingTAMs, indicating a Th1 type immune environment. However, the tumor microenvironmentalso has increased levels of Th2 cytokines (IL-4 and IL-10) produced by both immune andtumor cells (Bluth et al., 2009; Lathers and Young, 2004; Pries and Wollenberg, 2006;Todaro et al., 2008). In our study, there was Th2 activation shown by phosphorylatedSTAT6 and CCL18 co-expression by CD163+ macrophages. Thus, these data suggest thatTAMs in SCC respond to a dynamic mix of Th1 and Th2 signals.
These studies show that in normal skin at steady state, there is a predominant Th2/M2environment. In contrast, SCC TAMs demonstrate features of both M1 and M2 activation.Conventionally, strong Th1/M1 responses should prevent tumor progression (Hung et al.,1998). Furthermore, in imiquimod treated SCC, increased levels of IFNγ in SCC inducedanti-tumor effects and inhibited tonic anti-inflammatory signals of IL-10 (Huang et al.,2009). However, despite the presence of a strong M1 signal, the natural history of SCC isusually tumor progression, and hence this M1 signal is not able to eradicate the tumor. Themechanism for the ineffectiveness of Th1/M1 macrophages in eradicating the tumor is notclear, but may be due to the imbalance of Th1 and Th2 cytokines and their effects on TAMs.
Our study provides insight into the heterogeneous phenotypes and functions of TAMs. Theeffect of M1 and M2 TAMs may be to amplify immune responses to the tumor by inducingchemotaxis and activation of infiltrating T cells. The abundant influx of macrophages intothe tumor microenvironment can also help promote tumor growth by stimulatingangiogenesis and tissue remodelling. The pro-tumoral role of TAMs in SCC is supported bythe production of pro-lymphangiogenic factors such as VEGF-C (Moussai et al., 2011). Ourobservation that TAMs produce MMP9 and MMP11 also supports their pro-tumoral role, asMMPs facilitate direct tumor spread and release of matrix-sequestered angiogenic factorsthat encourage tumor growth (Egeblad and Werb, 2002). Also, it is possible that tumors mayproduce factors that act on TAMs in a paracrine fashion (Balkwill and Mantovani, 2001). Apositive correlation between increased numbers of macrophages and poor prognosis invarious human cancers has been demonstrated (Bingle et al., 2002; El-Rouby, 2010; Lin andPollard, 2007; Nonomura et al., 2010; Shabo et al., 2008; Sica et al., 2006; Steidl et al.,2010). It is possible that the weaker classical activation state favors production of pro-carcinogenic growth factors by TAMs. Based on this, the potential to induce a moredominant M1 activation state in TAMs could be pursued as a promising target fortherapeutic interventions.
There has been much debate over the phenotypic activation of TAMs as the physiology ofmacrophages has been shown to demonstrate remarkable plasticity (Mosser and Edwards,2008; Siveen and Kuttan, 2009). Critics of the current linear model of macrophagepolarization argue that the model does not allow for the plethora of possible macrophageactivation states and capabilities. Our evidence suggests that the activation state of amacrophage cannot be pigeonholed into one category, and that the linear model ofmacrophage activation may not encompass all the potential roles of the macrophage.Therefore, we favor the dynamic color wheel model of macrophage activation proposed byMosser and Edwards (Mosser and Edwards, 2008) as it accommodates the chameleon-likeproperties of this cell (Stout and Suttles, 2004).
MATERIALS and METHODSInstitutional Review Board approval (Weill Cornell Medical College) and informed consentwas obtained before enrolling patients in this study, and the study adhered strictly toDeclaration of Helsinki Principles.
Pettersen et al. Page 6
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Samples used in studyFor immunohistochemistry and immunofluorescence, cutaneous nodular Stage 1 SCCsamples were obtained during Mohs micrographic surgery (n=3–18). Tumors were obtainedfrom sun-exposed regions, namely head, neck and dorsal hands. All tumors were < 2 cm onexamination and showed dermal invasion on light microscopy. Ten normal specimens wereobtained via punch biopsies from non-sun exposed areas of patients without skin cancer andnormal abdominoplasty tissue.
Immunohistochemistry and ImmunofluorescenceStandard procedures were used for immunohistochemistry as described (Bluth et al., 2009).Normal skin and SCC (n=8–18) were stained with macrophage markers CD163 and CD68(antibodies in Supplemental Table 2) and a counterstain. Normal papillary dermis,designated as the tissue extending from the epidermal-dermal border to 100μm deep to theepidermis, and SCC “juxtatumoral dermis,” defined as the dermis 100μm circumferential tothe tumor, were examined as previously described (Kaporis et al., 2007). Positive cells werecounted using NIH IMAGE J software, and cell counts per unit area (μm2 × 100,000) weredetermined. Immunofluorescence stains were carried out in a standard manner (Fuentes-Duculan et al., 2010) (Supplemental Table 2). Images were acquired using either ZeissAxioplan 2 widefield fluorescence microscope or upright confocal microscope. Dermalcollagen fibers gave green autofluorescence, and antibodies conjugated with fluorochromesoften gave background epidermal fluorescence. Single stain controls and isotype controlswere performed for the confocal images (Figure S2).
Gene Array AnalysisSCC microarray data have been previously published (Haider et al., 2006). To estimate thefold-change of SCC versus normal skin, a moderated t-test available in limma package fromR/Bioconductor was used, and genes were considered significant with a FCH>2 and FDR<0.05. GSEA was used to evaluate the enrichment of the macrophage transcriptomes in theSCC genomic phenotype (as defined by SCC vs Normal fold-change by microarray) as in aprevious publication by our group (Suarez-Farinas et al., 2010) using GSEA desktopapplication (Lamb et al., 2006; Suarez-Farinas et al., 2010). The genomic transcriptomes ofM1 and M2 macrophages derived from in vitro cytokine-stimulated macrophages have beenpublished (Fuentes-Duculan et al., 2010). Macrophages were polarized by adding IFNγ,IL-4, TNF-α, LPS, and LPS plus IFNγ, and then compared to control macrophages. Therewere 585 upregulated and 334 downregulated probes in the M1 macrophages respectively,and 132 upregulated and 29 downregulated probes in the M2 macrophages respectively(Fuentes-Duculan et al., 2010).
Reverse transcriptase-polymerase chain reaction (RT-PCR)RNA was extracted from paired SCC and NT samples meeting the above inclusion criteria(n=15) and normal skin from healthy volunteers (n=10), using the RNeasy Mini KIT(Qiagen, Valencia, CA, U.S.A.). RT-PCR was performed as previously described (Chamianet al., 2005; Fuentes-Duculan et al., 2010). The PCR was performed in two batches. The firstgroup was 10 paired SCC and NT, and 2 normals, and the second group was 5 paired SCCsamples (described in Moussai et al., 2011) and 8 normal skin RNA. The primers for MMP9and MMP11 were from Applied Biosystems (Hs00957562_m1 and Hs00171829_m1,respectively), normalized to HARP housekeeping gene. Since samples were obtained at twodifferent time points, the log2 data was adjusted using a linear model to account for the batcheffect.
Pettersen et al. Page 7
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

StatisticsCell counts were analyzed by Mann Whitney U test, and p-values reported. Logarithmic RT-PCR data was analyzed using paired t-test for SCC v NT and unpaired t-test to compare withnormal skin control.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsResearch was supported by the Dana Foundation (Human Immunology Consortium Grant), which supports JSP,KCP, LF, AP-K, and JAC. MJB is supported by National Institutes of Health (NIH) grant T32-HL07423; MAL issupported by NIH grant 1 K23AR052404 and the Doris Duke Foundation, LJ-H is supported by the Doris DukeFoundation; MS-F is partially supported by NIH grant UL1 RR024143 from the National Center for ResearchResources (NCRR) and the Milstein Medical Research Program, CQFW is supported by the Milstein MedicalResearch Program. We thank plastic surgeon DM Senderoff for the generous donation of abdominoplasty surgicalwaste.
ABBREVIATIONS
SCC squamous cell carcinoma
TAMs tumor-associated macrophages
GSEA gene set enrichment analysis
FDR false discovery rate
ReferencesBalkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001; 357:539–45.
[PubMed: 11229684]
Bingle L, Brown NJ, Lewis CE. The role of tumour-associated macrophages in tumour progression:implications for new anticancer therapies. J Pathol. 2002; 196:254–65. [PubMed: 11857487]
Biswas SK, Gangi L, Paul S, Schioppa T, Saccani A, Sironi M, et al. A distinct and uniquetranscriptional program expressed by tumor-associated macrophages (defective NF-kappaB andenhanced IRF-3/STAT1 activation). Blood. 2006; 107:2112–22. [PubMed: 16269622]
Bluth MJ, Zaba LC, Moussai D, Suarez-Farinas M, Kaporis H, Fan L, et al. Myeloid dendritic cellsfrom human cutaneous squamous cell carcinoma are poor stimulators of T-cell proliferation. JInvest Dermatol. 2009; 129:2451–62. [PubMed: 19387481]
Bonecchi R, Sozzani S, Stine JT, Luini W, D’Amico G, Allavena P, et al. Divergent effects ofinterleukin-4 and interferon-gamma on macrophage-derived chemokine production: anamplification circuit of polarized T helper 2 responses. Blood. 1998; 92:2668–71. [PubMed:9763548]
Brantsch KD, Meisner C, Schonfisch B, Trilling B, Wehner-Caroli J, Rocken M, et al. Analysis of riskfactors determining prognosis of cutaneous squamous-cell carcinoma: a prospective study. LancetOncol. 2008; 9:713–20. [PubMed: 18617440]
Buergy D, Weber T, Maurer GD, Mudduluru G, Medved F, Leupold JH, et al. Urokinase receptor,MMP-1 and MMP-9 are markers to differentiate prognosis, adenoma and carcinoma in thyroidmalignancies. Int J Cancer. 2009; 125:894–901. [PubMed: 19480010]
Carucci JA. Cutaneous oncology in organ transplant recipients: meeting the challenge of squamouscell carcinoma. J Invest Dermatol. 2004; 123:809–16. [PubMed: 15482465]
Chamian F, Lowes MA, Lin SL, Lee E, Kikuchi T, Gilleaudeau P, et al. Alefacept reduces infiltratingT cells, activated dendritic cells, and inflammatory genes in psoriasis vulgaris. Proc Natl Acad SciU S A. 2005; 102:2075–80. [PubMed: 15671179]
Pettersen et al. Page 8
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Crawley AM, Vranjkovic A, Young C, Angel JB. Interleukin-4 downregulates CD127 expression andactivity on human thymocytes and mature CD8+ T cells. Eur J Immunol. 2010; 40:1396–407.[PubMed: 20201040]
de Oliveira MV, Fraga CA, Gomez RS, Paula AM. Immunohistochemical expression of interleukin-4,-6, -8, and -12 in inflammatory cells in surrounding invasive front of oral squamous cellcarcinoma. Head Neck. 2009; 31:1439–46. [PubMed: 19424975]
Egeblad M, Werb Z. New functions for the matrix metalloproteinases in cancer progression. Nat RevCancer. 2002; 2:161–74. [PubMed: 11990853]
El-Rouby DH. Association of macrophages with angiogenesis in oral verrucous and squamous cellcarcinomas. J Oral Pathol Med. 2010; 39:559–64. [PubMed: 20412402]
Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, Henson PM. Macrophages that haveingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J Clin Invest. 1998; 101:890–8.[PubMed: 9466984]
Forbes E, van Panhuys N, Min B, Le Gros G. Differential requirements for IL-4/STAT6 signalling inCD4 T-cell fate determination and Th2-immune effector responses. Immunol Cell Biol. 2010;88:240–3. [PubMed: 20010912]
Fuentes-Duculan J, Suarez-Farinas M, Zaba LC, Nograles KE, Pierson KC, Mitsui H, et al. Asubpopulation of CD163-positive macrophages is classically activated in psoriasis. J InvestDermatol. 2010; 130:2412–22. [PubMed: 20555352]
Gocheva V, Wang HW, Gadea BB, Shree T, Hunter KE, Garfall AL, et al. IL-4 induces cathepsinprotease activity in tumor-associated macrophages to promote cancer growth and invasion. GenesDev. 2010; 24:241–55. [PubMed: 20080943]
Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity.2010; 32:593–604. [PubMed: 20510870]
Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2005; 5:953–64.[PubMed: 16322748]
Gustafsson C, Mjosberg J, Matussek A, Geffers R, Matthiesen L, Berg G, et al. Gene expressionprofiling of human decidual macrophages: evidence for immunosuppressive phenotype. PLoSOne. 2008; 3:e2078. [PubMed: 18446208]
Haider AS, Peters SB, Kaporis H, Cardinale I, Fei J, Ott J, et al. Genomic analysis defines a cancer-specific gene expression signature for human squamous cell carcinoma and distinguishesmalignant hyperproliferation from benign hyperplasia. J Invest Dermatol. 2006; 126:869–81.[PubMed: 16470182]
Huang SJ, Hijnen D, Murphy GF, Kupper TS, Calarese AW, Mollet IG, et al. Imiquimod enhancesIFN-gamma production and effector function of T cells infiltrating human squamous cellcarcinomas of the skin. J Invest Dermatol. 2009; 129:2676–85. [PubMed: 19516264]
Hung K, Hayashi R, Lafond-Walker A, Lowenstein C, Pardoll D, Levitsky H. The central role ofCD4(+) T cells in the antitumor immune response. J Exp Med. 1998; 188:2357–68. [PubMed:9858522]
Kaporis HG, Guttman-Yassky E, Lowes MA, Haider AS, Fuentes-Duculan J, Darabi K, et al. Humanbasal cell carcinoma is associated with Foxp3+ T cells in a Th2 dominant microenvironment. JInvest Dermatol. 2007; 127:2391–8. [PubMed: 17508019]
Katakura T, Miyazaki M, Kobayashi M, Herndon DN, Suzuki F. CCL17 and IL-10 as effectors thatenable alternatively activated macrophages to inhibit the generation of classically activatedmacrophages. J Immunol. 2004; 172:1407–13. [PubMed: 14734716]
Kodelja V, Muller C, Politz O, Hakij N, Orfanos CE, Goerdt S. Alternative macrophage activation-associated CC-chemokine-1, a novel structural homologue of macrophage inflammatory protein-1alpha with a Th2-associated expression pattern. J Immunol. 1998; 160:1411–8. [PubMed:9570561]
Koller FL, Hwang DG, Dozier EA, Fingleton B. Epithelial interleukin-4 receptor expression promotescolon tumor growth. Carcinogenesis. 2010; 31:1010–7. [PubMed: 20176658]
Pettersen et al. Page 9
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Kryczek I, Banerjee M, Cheng P, Vatan L, Szeliga W, Wei S, et al. Phenotype, distribution,generation, and functional and clinical relevance of Th17 cells in the human tumor environments.Blood. 2009; 114:1141–9. [PubMed: 19470694]
Kwan WH, Navarro-Sanchez E, Dumortier H, Decossas M, Vachon H, dos Santos FB, et al. Dermal-type macrophages expressing CD209/DC-SIGN show inherent resistance to dengue virus growth.PLoS Negl Trop Dis. 2008; 2:e311. [PubMed: 18827881]
Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ, et al. The Connectivity Map: usinggene-expression signatures to connect small molecules, genes, and disease. Science. 2006;313:1929–35. [PubMed: 17008526]
Landolfo S, Gribaudo G, Angeretti A, Gariglio M. Mechanisms of viral inhibition by interferons.Pharmacol Ther. 1995; 65:415–42. [PubMed: 7544016]
Lathers DM, Young MR. Increased aberrance of cytokine expression in plasma of patients with moreadvanced squamous cell carcinoma of the head and neck. Cytokine. 2004; 25:220–8. [PubMed:15036248]
Leek RD, Lewis CE, Whitehouse R, Greenall M, Clarke J, Harris AL. Association of macrophageinfiltration with angiogenesis and prognosis in invasive breast carcinoma. Cancer Res. 1996;56:4625–9. [PubMed: 8840975]
Li-Weber M, Krammer PH. Regulation of IL4 gene expression by T cells and therapeutic perspectives.Nat Rev Immunol. 2003; 3:534–43. [PubMed: 12876556]
Lin EY, Li JF, Gnatovskiy L, Deng Y, Zhu L, Grzesik DA, et al. Macrophages regulate the angiogenicswitch in a mouse model of breast cancer. Cancer Res. 2006; 66:11238–46. [PubMed: 17114237]
Lin EY, Pollard JW. Tumor-associated macrophages press the angiogenic switch in breast cancer.Cancer Res. 2007; 67:5064–6. [PubMed: 17545580]
Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverseforms of macrophage activation and polarization. Trends Immunol. 2004; 25:677–86. [PubMed:15530839]
Martinez FO, Gordon S, Locati M, Mantovani A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: new molecules and patterns of gene expression. JImmunol. 2006; 177:7303–11. [PubMed: 17082649]
Martinez FO, Helming L, Gordon S. Alternative activation of macrophages: an immunologicfunctional perspective. Annu Rev Immunol. 2009; 27:451–83. [PubMed: 19105661]
Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol.2008; 8:958–69. [PubMed: 19029990]
Moussai D, Mitsui H, Pettersen JS, Pierson KC, Shah KR, Suarez-Farinas M, et al. The HumanCutaneous Squamous Cell Carcinoma Microenvironment Is Characterized by Increased LymphaticDensity and Enhanced Expression of Macrophage-Derived VEGF-C. J Invest Dermatol. 2011;131:229–36. [PubMed: 20827282]
Nonomura N, Takayama H, Kawashima A, Mukai M, Nagahara A, Nakai Y, et al. Decreasedinfiltration of macrophage scavenger receptor-positive cells in initial negative biopsy specimens iscorrelated with positive repeat biopsies of the prostate. Cancer Sci. 2010; 101:1570–3. [PubMed:20384632]
Pinto CA, Carvalho PE, Antonangelo L, Garippo A, Da Silva AG, Soares F, et al. Morphometricevaluation of tumor matrix metalloproteinase 9 predicts survival after surgical resection ofadenocarcinoma of the lung. Clin Cancer Res. 2003; 9:3098–104. [PubMed: 12912961]
Pries R, Wollenberg B. Cytokines in head and neck cancer. Cytokine Growth Factor Rev. 2006;17:141–6. [PubMed: 16540364]
Puig-Kroger A, Serrano-Gomez D, Caparros E, Dominguez-Soto A, Relloso M, Colmenares M, et al.Regulated expression of the pathogen receptor dendritic cell-specific intercellular adhesionmolecule 3 (ICAM-3)-grabbing nonintegrin in THP-1 human leukemic cells, monocytes, andmacrophages. J Biol Chem. 2004; 279:25680–8. [PubMed: 15070901]
Romieu-Mourez R, Solis M, Nardin A, Goubau D, Baron-Bodo V, Lin R, et al. Distinct roles for IFNregulatory factor (IRF)-3 and IRF-7 in the activation of antitumor properties of humanmacrophages. Cancer Res. 2006; 66:10576–85. [PubMed: 17079482]
Pettersen et al. Page 10
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Saha B, Jyothi Prasanna S, Chandrasekar B, Nandi D. Gene modulation and immunoregulatory rolesof interferon gamma. Cytokine. 2010; 50:1–14. [PubMed: 20036577]
Schroder K, Hertzog PJ, Ravasi T, Hume DA. Interferon-gamma: an overview of signals, mechanismsand functions. J Leukoc Biol. 2004; 75:163–89. [PubMed: 14525967]
Schutyser E, Richmond A, Van Damme J. Involvement of CC chemokine ligand 18 (CCL18) innormal and pathological processes. J Leukoc Biol. 2005; 78:14–26. [PubMed: 15784687]
Shabo I, Stal O, Olsson H, Dore S, Svanvik J. Breast cancer expression of CD163, a macrophagescavenger receptor, is related to early distant recurrence and reduced patient survival. Int J Cancer.2008; 123:780–6. [PubMed: 18506688]
Shah SA, Spinale FG, Ikonomidis JS, Stroud RE, Chang EI, Reed CE. Differential matrixmetalloproteinase levels in adenocarcinoma and squamous cell carcinoma of the lung. J ThoracCardiovasc Surg. 2010; 139:984–90. discussion 90. [PubMed: 20304142]
Sica A, Schioppa T, Mantovani A, Allavena P. Tumour-associated macrophages are a distinct M2polarised population promoting tumour progression: potential targets of anti-cancer therapy. Eur JCancer. 2006; 42:717–27. [PubMed: 16520032]
Siveen KS, Kuttan G. Role of macrophages in tumour progression. Immunol Lett. 2009; 123:97–102.[PubMed: 19428556]
Soilleux EJ, Morris LS, Leslie G, Chehimi J, Luo Q, Levroney E, et al. Constitutive and inducedexpression of DC-SIGN on dendritic cell and macrophage subpopulations in situ and in vitro. JLeukoc Biol. 2002; 71:445–57. [PubMed: 11867682]
Steidl C, Lee T, Shah SP, Farinha P, Han G, Nayar T, et al. Tumor-associated macrophages andsurvival in classic Hodgkin’s lymphoma. N Engl J Med. 2010; 362:875–85. [PubMed: 20220182]
Stout RD, Suttles J. Functional plasticity of macrophages: reversible adaptation to changingmicroenvironments. J Leukoc Biol. 2004; 76:509–13. [PubMed: 15218057]
Suarez-Farinas M, Lowes MA, Zaba LC, Krueger JG. Evaluation of the psoriasis transcriptome acrossdifferent studies by gene set enrichment analysis (GSEA). PLoS One. 2010; 5:e10247. [PubMed:20422035]
Subramanian A, Kuehn H, Gould J, Tamayo P, Mesirov JP. GSEA-P: a desktop application for GeneSet Enrichment Analysis. Bioinformatics. 2007; 23:3251–3. [PubMed: 17644558]
Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene setenrichment analysis: a knowledge-based approach for interpreting genome-wide expressionprofiles. Proc Natl Acad Sci U S A. 2005; 102:15545–50. [PubMed: 16199517]
Takeda K, Tanaka T, Shi W, Matsumoto M, Minami M, Kashiwamura S, et al. Essential role of Stat6in IL-4 signalling. Nature. 1996; 380:627–30. [PubMed: 8602263]
Todaro M, Lombardo Y, Francipane MG, Alea MP, Cammareri P, Iovino F, et al. Apoptosis resistancein epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ. 2008;15:762–72. [PubMed: 18202702]
van Kempen LC, Ruiter DJ, van Muijen GN, Coussens LM. The tumor microenvironment: a criticaldeterminant of neoplastic evolution. Eur J Cell Biol. 2003; 82:539–48. [PubMed: 14703010]
Wang YC, He F, Feng F, Liu XW, Dong GY, Qin HY, et al. Notch signaling determines the M1 versusM2 polarization of macrophages in antitumor immune responses. Cancer Res. 2010; 70:4840–9.[PubMed: 20501839]
Weinberg AS, Ogle CA, Shim EK. Metastatic cutaneous squamous cell carcinoma: an update.Dermatol Surg. 2007; 33:885–99. [PubMed: 17661931]
Zaba LC, Fuentes-Duculan J, Steinman RM, Krueger JG, Lowes MA. Normal human dermis containsdistinct populations of CD11c+BDCA-1+ dendritic cells and CD163+FXIIIA+ macrophages. JClin Invest. 2007; 117:2517–25. [PubMed: 17786242]
Zhao ZS, Chu YQ, Ye ZY, Wang YY, Tao HQ. Overexpression of matrix metalloproteinase 11 inhuman gastric carcinoma and its clinicopathologic significance. Hum Pathol. 2010; 41:686–96.[PubMed: 20060156]
Pettersen et al. Page 11
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Macrophages were more abundant in SCC compared to normal skinRepresentative immunohistochemistry (10x) and cell counts of the macrophage markers (a)CD163 (with an inset of CD163+ cells at 20x) and (b) CD68, showing a significantlyincreased number of macrophages surrounding SCC tumor nests compared to normal skin.Each dot represents one patient. ***P <0.001. (c) CD163 (green) co-localized with CD68(red) shown as yellow, but there were CD163+ cells that did not co-express CD68. (d)CD163 (green) did not co-express CD11c (red), while (e) some CD68+ cells (green) did co-express CD11c (red) shown as yellow. Bar=100μm.
Pettersen et al. Page 12
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. SCC TAMs expressed pro-tumoral products in the tumor microenvironmentMean mRNA expression of (a) MMP9 and (b) MMP11 relative to HARP after adjustmentfor batch effect in normal skin (white bars), non-tumoral skin (NT, gray bars), and SCC(black bars) with standard error of the mean. *P <0.05, **P <0.01. CD163+ cells (green)demonstrated abundant co-localization with (c) MMP9 (red) and (d) MMP11 (red) in SCCcompared to normal skin. Double positive cells appear yellow. Bar=100μm.
Pettersen et al. Page 13
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. SCC TAMs expressed products of alternatively activated macrophagesMany CD163+ cells (green) co-expressed (a) phosphorylated STAT6 (STAT6p) (red), (b)CD209/DC-SIGN (red), and (c) CCL18 (red) compared to normal skin. Double positivecells appear yellow. Bar=100μm
Pettersen et al. Page 14
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
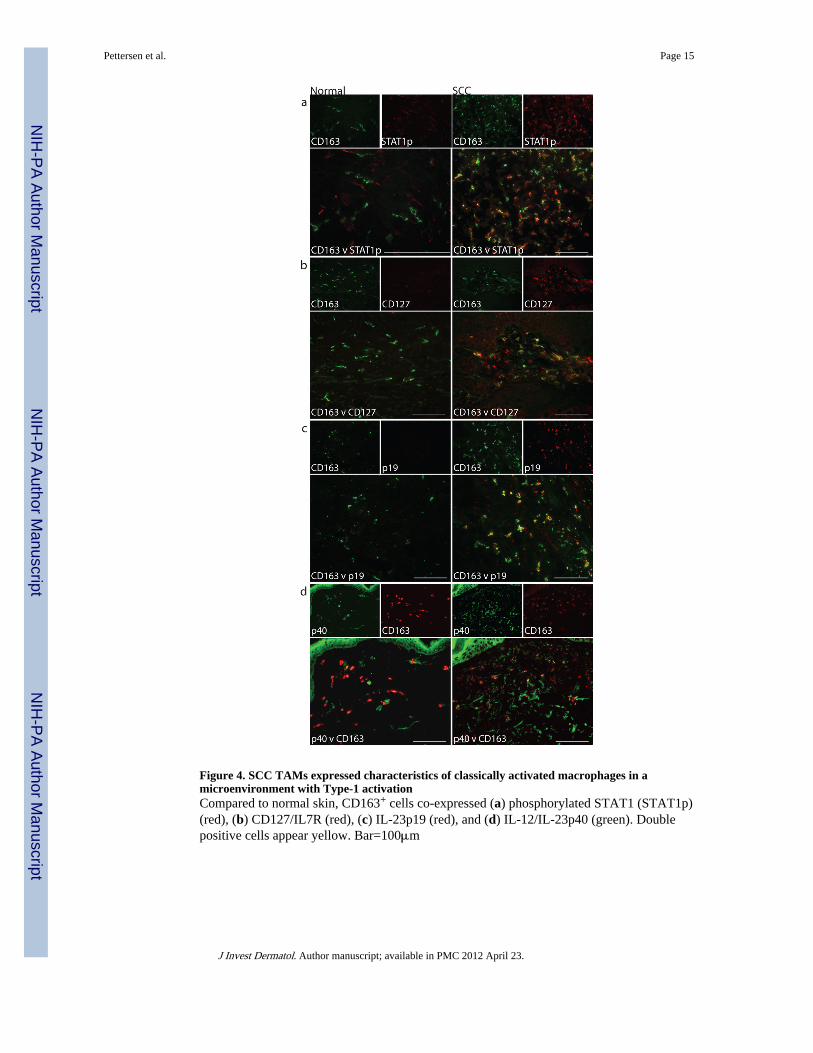
Figure 4. SCC TAMs expressed characteristics of classically activated macrophages in amicroenvironment with Type-1 activationCompared to normal skin, CD163+ cells co-expressed (a) phosphorylated STAT1 (STAT1p)(red), (b) CD127/IL7R (red), (c) IL-23p19 (red), and (d) IL-12/IL-23p40 (green). Doublepositive cells appear yellow. Bar=100μm
Pettersen et al. Page 15
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. A subset of SCC TAMs simultaneously expressed characteristics of both classical andalternative activationTriple-labeled confocal immunofluorescence revealed the presence of (a) CD163+ cells(blue) that simultaneously co-expressed STAT6p (green) and STAT1p (red) and (b) CD163+
cells (green) that simultaneously co-expressed the markers CD127 (red) and CD209 (blue)in SCC. Triple-positive cells (white) are indicated by arrows. Bar=100μm. (c) The proposedmodel of SCC macrophage polarization. Th1 and Th2 cells produce cytokines, IFNγ andIL-4, respectively, and act on resident CD163+ macrophages to polarize these cells inseveral directions. IFNγ stimulates the M1 phenotype (CD127 and IL-23), and IL-4stimulates towards the M2 phenotype (CD209 and CCL18). There is also production ofmediators that are not driven by known polarizing cytokines, such as MMP9, MMP11, andVEGF-C. The overall outcome is a poly-activated TAM.
Pettersen et al. Page 16
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Pettersen et al. Page 17
Table 1
Representative upregulated genes in the M1 and M2 macrophage gene sets* and the SCC transcriptome**.
Representative Genesa M1 Gene Set M2 Gene Set SCC Transcriptomed
M1
STAT1 8.51 c 2.47
Mx1 19.03 c 2.89
IL-23p19 3.10 c 2.34
IL-12/IL-23p40 3.36 c 1.11
CD127 4.44 c e
M2
CD209 b 9.25 e
CCL18 b 3.43 3.55f
CCL17 b 2.58 1.43
*Fuentes-Duculan et al., 2010
**Haider et al., 2006
aGenes representative of M1 and M2 macrophages were selected and their fold change in the M1 and M2 gene sets and the SCC transcriptome are
listed.
bGenes not differentially expressed in the M1 gene set compared to control.
cGenes not differentially expressed in the M2 gene set compared to control.
dSCC versus normal skin transcriptome, fold change and FDR <0.05.
eHU95 chip did not include this probe.
fFDR > 0.05
J Invest Dermatol. Author manuscript; available in PMC 2012 April 23.










![Douez v. Facebook, Inc., 2017 SCC 33, [2017] 1 SCR 751 ...](https://static.fdokumen.com/doc/165x107/6323c69fb104cba27a08d825/douez-v-facebook-inc-2017-scc-33-2017-1-scr-751-.jpg)









