
Bicyclic RGD peptides: Novel high-affinity ligands for selective ...
Upload
independentCategory
view
2download
0
New functional ligands for ficolin-3 amonglipopolysaccharides of Hafnia alvei
Anna Swierzko2, Jolanta Lukasiewicz1,3,Maciej Cedzynski2, Anna Maciejewska3,Wojciech Jachymek3, Tomasz Niedziela3,Misao Matsushita4, and Czeslaw Lugowski3,5
2Laboratory of Immunobiology of Infections, Institute for Medical Biology,Polish Academy of Sciences, Lodowa 106, PL-93-232 Lodz, Poland;3Department of Immunochemistry, Ludwik Hirszfeld Institute of Immunologyand Experimental Therapy, Polish Academy of Sciences, R. Weigla 12,PL-53-114 Wroclaw, Poland; 4Department of Applied Biochemistry, TokaiUniversity, 4-1-1 Kitakaname, Hiratsuka-shi, Kanagawa 259-1292, Japan; and5Department of Biotechnology and Molecular Biology, University of Opole,PL-45-035 Opole, Poland
Received on May 24, 2011; revised on July 29, 2011; accepted on August 19, 2011
Ficolin-1 (M), ficolin-2 (L), ficolin-3 (H) and mannan-binding lectin (MBL) activate the complement system andhave opsonic activity. The specificity of ficolin-3 is poorlycharacterized and currently limited to a few ligands only.We present new specific targets for human ficolin-3, ident-ified among lipopolysaccharides (LPSs, endotoxin) ofHafnia alvei. The interaction was restricted to LPSs offour strains: 23, Polish Collection of Microorganisms(PCM) 1200, PCM 1203 and PCM 1205 and limited totheir O-specific polysaccharides (O-specific PSs) composedof different numbers of oligosaccharide (OS) repeatingunits (RUs). Moreover, these LPS/ficolin-3 complexesactivated the lectin pathway of complement in a C4b-depo-sition assay in a calcium- and magnesium-dependent way.A neoglycoconjugate of the O-specific PS fraction of H.alvei 1200 LPS with bovine serum albumin (BSA) was pre-pared and used as a tool for the determination of ficolin-3concentration and activity in serum. To confirm a struc-ture of the O-specific PS 1200 selected for the conjugatepreparation, structural analysis was performed on a seriesof O-specific PSs released by the mild acid hydrolysis ofthe LPS. The isolated O-specific PSs, showing the differentlength distributions, were devoid of a major part of thecore OS region and had Hep-Kdo disaccharide at a redu-cing end. The neoglycoconjugate was a highly selectivetool for the determination of ficolin-3 concentration andactivity in serum (lectin pathway activation in the C4bdeposition assay) and was not affected by MBL, ficolin-1and ficolin-2 or natural antibodies.
Keywords: complement / endotoxin / ficolin /Hafnia /lipopolysaccharide
Introduction
Many of the mechanisms of innate immunity depend on fast,flexible and straightforward interactions between soluble orcell-associated pattern-recognition molecules (PRMs) andpathogen-associated molecular patterns (PAMPs)–conservativestructures common for the variety of pathogens. Examples ofsuch PRM–PAMP are the collagen-associated lectins, i.e.ficolin-1 (M), ficolin-2 (L), ficolin-3 (H, Hakata antigen) andmannan-binding lectin (MBL), together with their microbialligands. These molecules are involved in the complement acti-vation, via the lectin pathway, an important element of innateimmune response. This unique feature depends on the abilityof these lectins to form complexes with MBL-associatedserine proteases (MASPs; Matsushita 2007). The C-terminalfibrinogen-like region of ficolins constitutes thetarget-recognizing domain (MacDonald and Kilpatrick 2007;Matsushita 2007; Endo et al. 2011). Identification of ligandsfor ficolins is a prerequisite to understand the role of thecollagen-related lectins in innate immunity and to design diag-nostic tools to measure concentration and the activity of thesekey complement components in body fluids, e.g. serum.Ficolin-3 is an oligomer of subunits consisting of three
identical polypeptide chains. Despite similar molecular organ-ization, biochemical properties and activities, it differs fromother proteins of that family as the amino acid sequence hom-ology between ficolin-3 and ficolin-1 or ficolin-2 is 48%(Matsushita 2007; Thiel 2007; Hummelshoj et al. 2008;Matsushita 2009; Garred et al. 2010; Endo et al. 2011).Ficolin-3 is synthesized in the liver by hepatocytes and bileduct epithelial cells and in lungs by type II alveolar andciliated bronchial epithelial cells. It is secreted into blood, bileducts, bronchi and alveoli (Matsushita 2007); thus, it mayparticipate in both systemic and local innate immuneresponse.The average concentration of ficolin-3 in human sera is 18
µg/mL, the highest among ficolins and significantly exceedingthe MBL level (Yae et al. 1991). The importance of serumficolin-3 was indicated by several reports concerning diseaseassociations with abnormally low or high concentrations.Lower levels of ficolin-3 were observed among patients suf-fering from systemic lupus erythematosus (Inaba et al. 1990)and sarcoidosis (Svendsen et al. 2008). A reduced ficolin-3
1To whom correspondence should be addressed: Tel: +48-71-370-99-27; Fax:+48-71-337-2171; e-mail: [email protected]
Glycobiology vol. 22 no. 2 pp. 267–280, 2012doi:10.1093/glycob/cwr119Advance Access publication on September 2, 2011
© The Author 2011. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] 267
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

level in hepatic cirrhosis may be a marker of impaired liverfunction (Fukutomi et al. 1996). Schlapbach et al. demon-strated recently an enhanced risk of febrile neutropenia(especially with bacteremia) in ficolin-3-deficient pediatriccancer patients, treated with chemotherapy (Schlapbach et al.2009) as well as necrotizing enterocolitis among neonates(Schlapbach et al. 2010). Wang et al. (2007) reported highconcentrations of ficolin-3 in the placentas from preeclampticpregnant women, accompanied by low serum levels.Considering human ficolins, the specificity of ficolin-3 is
rather poorly characterized. Only a few ligands have beenidentified to date. The first communication concerning targetsfor ficolin-3 dealt with its ability to agglutinate erythrocytescoated with lipopolysaccharide (LPS, endotoxin, O-antigen)of Salmonella Typhimurium, S. Minnesota and Escherichiacoli O111 (Sugimoto et al. 1998). Tsujimura et al. (2002)identified an exopolysaccharide (EPS) of Aerococcus viridansas a ligand for ficolin-3, but its detailed structure has not beenelucidated to date. No ligands were identified among numer-ous strains of streptococci, staphylococci and E. coli (Krarupet al. 2005). N-Acetylated BSA (BSA-NAc) was the lastligand identified to date. This artificial ligand was reported tobe useful for the determination of ficolin-3-dependent comp-lement activity (Lacroix et al. 2009; Munthe-Fog et al. 2009;Hein et al. 2010) and was used to confirm the first case officolin-3 deficiency in a patient suffering from recurrent infec-tions (Munthe-Fog et al. 2009).Herein we present a part of our continuous studies of struc-
ture–biological activity relationships for LPSs, that have ledto the identification of several Hafnia alvei LPSs as newligands bound by ficolin-3.Hafnia alvei, a Gram-negative bacterium, is a causative
agent of bacteremia and septicemia in humans and animals. Itis also associated with respiratory diseases and mixed hospitalinfections in humans. Sepsis developed due to H. alvei infec-tion is also a serious clinical problem in the animal productionindustry of commercial laying hens, pullets and rainbow trout(Janda et al. 1999). LPSs of H. alvei are major surface anti-gens, composed of three main regions lipid A, core oligosac-charide (OS) and the O-specific PS, consisting of differentnumbers of OS repeating units (RUs). The biosynthesis andassembly of LPSs is a complex multi-step process comprisingthe polymerization of the O-specific chain and its subsequentligation onto the outer core domain of the lipid A–core OSacceptor (Raetz and Whitfield 2002). The O-specific PS isusually characterized by different degrees of polymerizationdue to the varying number of RUs attached to the core OS.The degree of polymerization is a main reason of heterogen-eity observed among LPSs isolated from one particular strain.Preparations of smooth-type LPS usually constitute a mixtureof molecules composed of lipid A substituted by core OS anddifferent numbers of RUs as well as lipid A substituted bycore OS only. The O-antigens of H. alvei are subdivided into40 O-serotypes (Baturo and Raginskaya 1978; Romanowska2000). The structures of the O-specific PS from more than 30serologically different H. alvei strains have been elucidated(Romanowska 2000; Katzenellenbogen et al. 2005;Lukasiewicz et al. 2009). In the case of core OS, only fourtypes have been identified to date. Unlike almost all of
enterobacterial endotoxins, Kdo-containing motifs were ident-ified in the outer core region of H. alvei LPS (Lukasiewiczet al. 2009).Herein, we present newly identified interactions between
ficolin-3 and some of H. alvei LPSs. We have identifiedregions of LPS recognized by this lectin and present theseligands as a potential diagnostic tool for the determination officolin-3 concentration and ficolin-3-dependent lectin pathwayactivity measurements in body fluids.
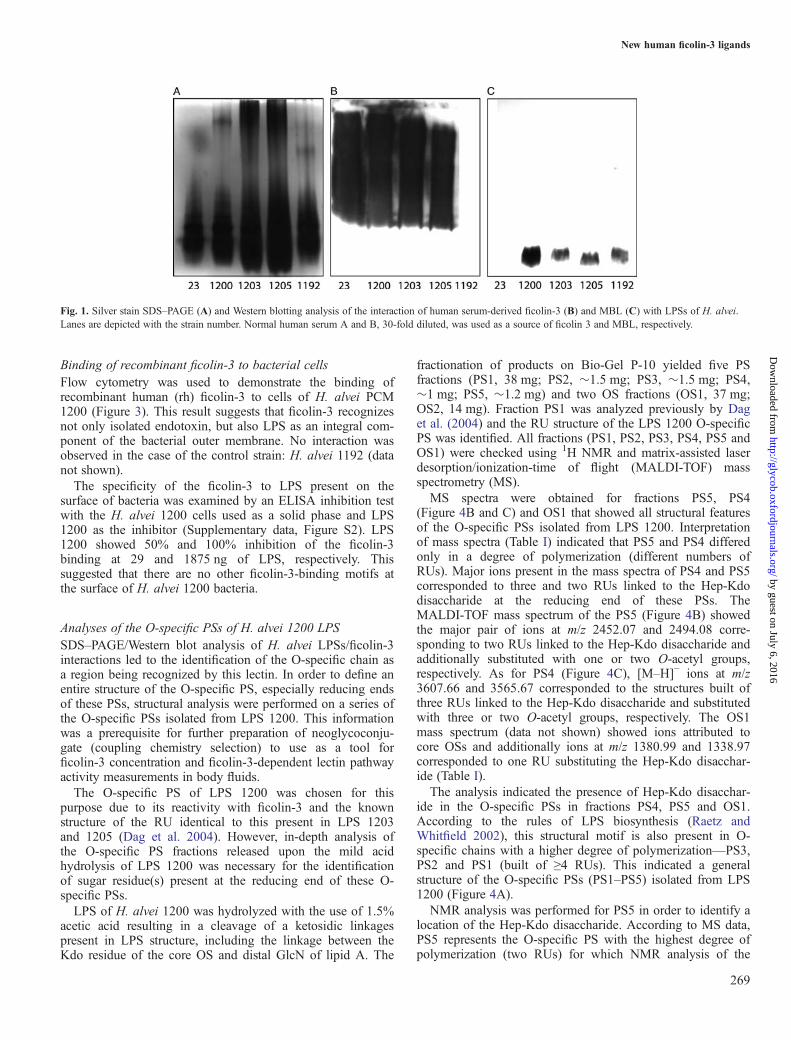
ResultsInteraction of serum ficolin-3 with H. alvei LPSsForty-five H. alvei LPSs were tested in a dot-blot assay to findthese being recognized by serum-derived ficolin-3 (data notshown). Four LPSs isolated from strains, 23, PCM 1200, PCM1203, PCM 1205, were reactive and were further analyzed insodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS–PAGE)/Western blot using human sera as the source officolin-3 or ficolin-2 (serum A), MBL (serum B) or ficolin-1(pooled A and B sera). This was done to identify a regionwithin LPSs recognized by ficolin-3. SDS–PAGE combinedwith silver staining of smooth-type LPS usually gives a charac-teristic ladder-like multi-band pattern. The fast-migrating frac-tions originate from lipid A substituted with core OS. Theslower-migrating fractions show the length distribution of thepolymer built of lipid A–core OS substituted with differentnumbers of RUs (Figure 1A). The ficolin-3 bound tohigh-molecular-weight fractions of LPS, which indicated theO-specific PS (with different numbers of RUs) as a regionrecognized by this lectin (Figure 1A and B). LPS isolated fromH. alvei PCM 1192 was used as a negative control. In contrast,MBL interacted with low-molecular-weight LPS fractions of1192, 1200, 1203 and 1205 strains, corresponding to the lipidA-core region (Figure 1A and C). The reactivity of ficolin-2and ficolin-1 with LPSs tested was negligible (data notshown). Efficient binding of ficolin-3 to the aforementionedLPSs was also confirmed by ELISA, with a detectable (A405
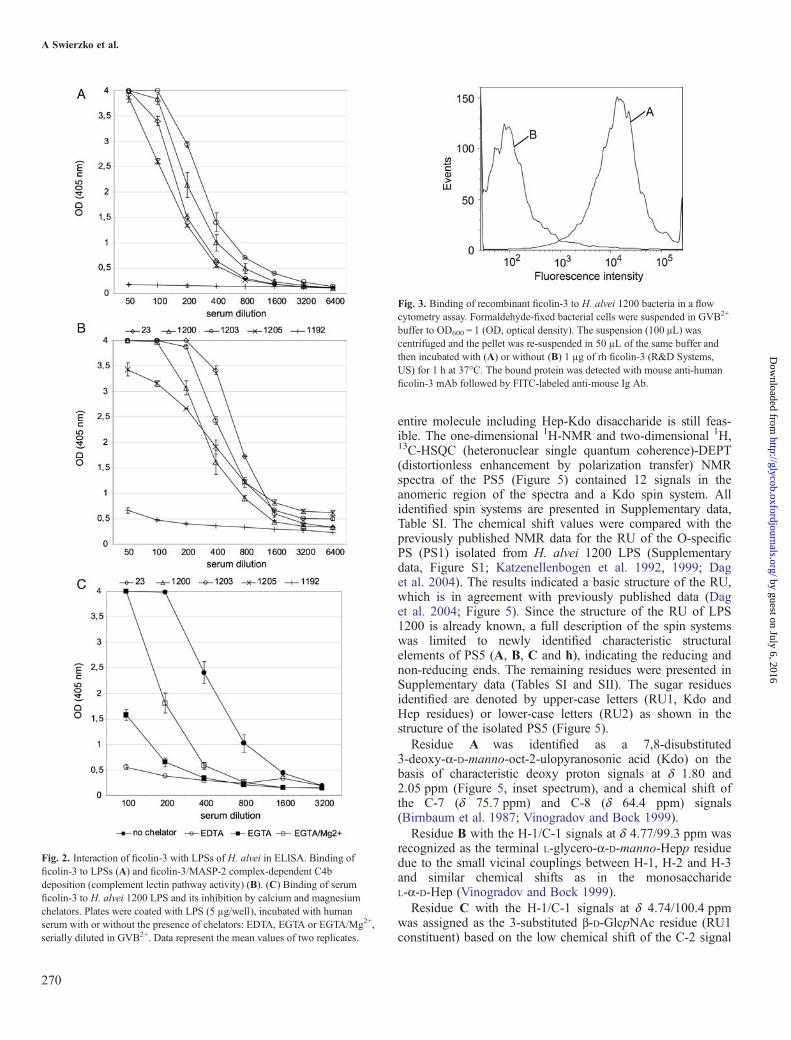
nm > 0.2) reaction for serum dilutions 1:800–1:1600(Figure 2A).
Complement activationAbility of LPS 23, 1200, 1203, 1205 to initiate the lectinpathway of the complement activation through a formation ofLPS/ficolin-3/MASP-2 complexes and cleavage of C4 wasdemonstrated by a C4b deposition assay (Figure 2B). LPS iso-lated from H. alvei PCM 1192 was used as a negative control.
Inhibition of ficolin-3 binding to LPS 1200 by ion chelatorsLPS 1200 was used as a solid phase in ELISA and thebinding of ficolin-3 was measured in the presence or theabsence of EDTA (ethylenediaminetetraacetic acid) or EGTA(ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraaceticacid). The observed interaction between ficolin-3 and LPSH. alvei 1200 was calcium- and magnesium-dependent. It wasstrongly inhibited by both EDTA and EGTA and, to someextent, was reconstituted by Mg2+ supplementation (EGTA/Mg2+; Figure 2C).
A Swierzko et al.
268
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Binding of recombinant ficolin-3 to bacterial cellsFlow cytometry was used to demonstrate the binding ofrecombinant human (rh) ficolin-3 to cells of H. alvei PCM1200 (Figure 3). This result suggests that ficolin-3 recognizesnot only isolated endotoxin, but also LPS as an integral com-ponent of the bacterial outer membrane. No interaction wasobserved in the case of the control strain: H. alvei 1192 (datanot shown).The specificity of the ficolin-3 to LPS present on the
surface of bacteria was examined by an ELISA inhibition testwith the H. alvei 1200 cells used as a solid phase and LPS1200 as the inhibitor (Supplementary data, Figure S2). LPS1200 showed 50% and 100% inhibition of the ficolin-3binding at 29 and 1875 ng of LPS, respectively. Thissuggested that there are no other ficolin-3-binding motifs atthe surface of H. alvei 1200 bacteria.
Analyses of the O-specific PSs of H. alvei 1200 LPSSDS–PAGE/Western blot analysis of H. alvei LPSs/ficolin-3interactions led to the identification of the O-specific chain asa region being recognized by this lectin. In order to define anentire structure of the O-specific PS, especially reducing endsof these PSs, structural analysis were performed on a series ofthe O-specific PSs isolated from LPS 1200. This informationwas a prerequisite for further preparation of neoglycoconju-gate (coupling chemistry selection) to use as a tool forficolin-3 concentration and ficolin-3-dependent lectin pathwayactivity measurements in body fluids.The O-specific PS of LPS 1200 was chosen for this
purpose due to its reactivity with ficolin-3 and the knownstructure of the RU identical to this present in LPS 1203and 1205 (Dag et al. 2004). However, in-depth analysis ofthe O-specific PS fractions released upon the mild acidhydrolysis of LPS 1200 was necessary for the identificationof sugar residue(s) present at the reducing end of these O-specific PSs.LPS of H. alvei 1200 was hydrolyzed with the use of 1.5%
acetic acid resulting in a cleavage of a ketosidic linkagespresent in LPS structure, including the linkage between theKdo residue of the core OS and distal GlcN of lipid A. The
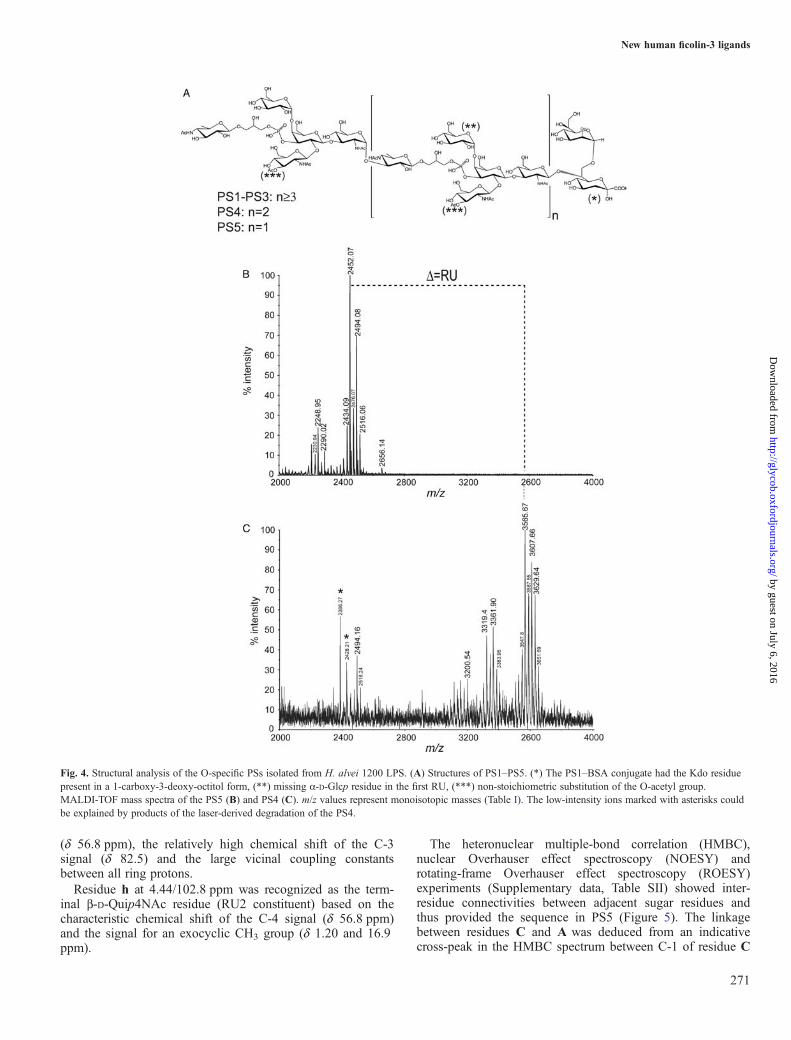
fractionation of products on Bio-Gel P-10 yielded five PSfractions (PS1, 38 mg; PS2, �1.5 mg; PS3, �1.5 mg; PS4,�1 mg; PS5, �1.2 mg) and two OS fractions (OS1, 37 mg;OS2, 14 mg). Fraction PS1 was analyzed previously by Daget al. (2004) and the RU structure of the LPS 1200 O-specificPS was identified. All fractions (PS1, PS2, PS3, PS4, PS5 andOS1) were checked using 1H NMR and matrix-assisted laserdesorption/ionization-time of flight (MALDI-TOF) massspectrometry (MS).MS spectra were obtained for fractions PS5, PS4
(Figure 4B and C) and OS1 that showed all structural featuresof the O-specific PSs isolated from LPS 1200. Interpretationof mass spectra (Table I) indicated that PS5 and PS4 differedonly in a degree of polymerization (different numbers ofRUs). Major ions present in the mass spectra of PS4 and PS5corresponded to three and two RUs linked to the Hep-Kdodisaccharide at the reducing end of these PSs. TheMALDI-TOF mass spectrum of the PS5 (Figure 4B) showedthe major pair of ions at m/z 2452.07 and 2494.08 corre-sponding to two RUs linked to the Hep-Kdo disaccharide andadditionally substituted with one or two O-acetyl groups,respectively. As for PS4 (Figure 4C), [M–H]− ions at m/z3607.66 and 3565.67 corresponded to the structures built ofthree RUs linked to the Hep-Kdo disaccharide and substitutedwith three or two O-acetyl groups, respectively. The OS1mass spectrum (data not shown) showed ions attributed tocore OSs and additionally ions at m/z 1380.99 and 1338.97corresponded to one RU substituting the Hep-Kdo disacchar-ide (Table I).The analysis indicated the presence of Hep-Kdo disacchar-
ide in the O-specific PSs in fractions PS4, PS5 and OS1.According to the rules of LPS biosynthesis (Raetz andWhitfield 2002), this structural motif is also present in O-specific chains with a higher degree of polymerization—PS3,PS2 and PS1 (built of ≥4 RUs). This indicated a generalstructure of the O-specific PSs (PS1–PS5) isolated from LPS1200 (Figure 4A).NMR analysis was performed for PS5 in order to identify a
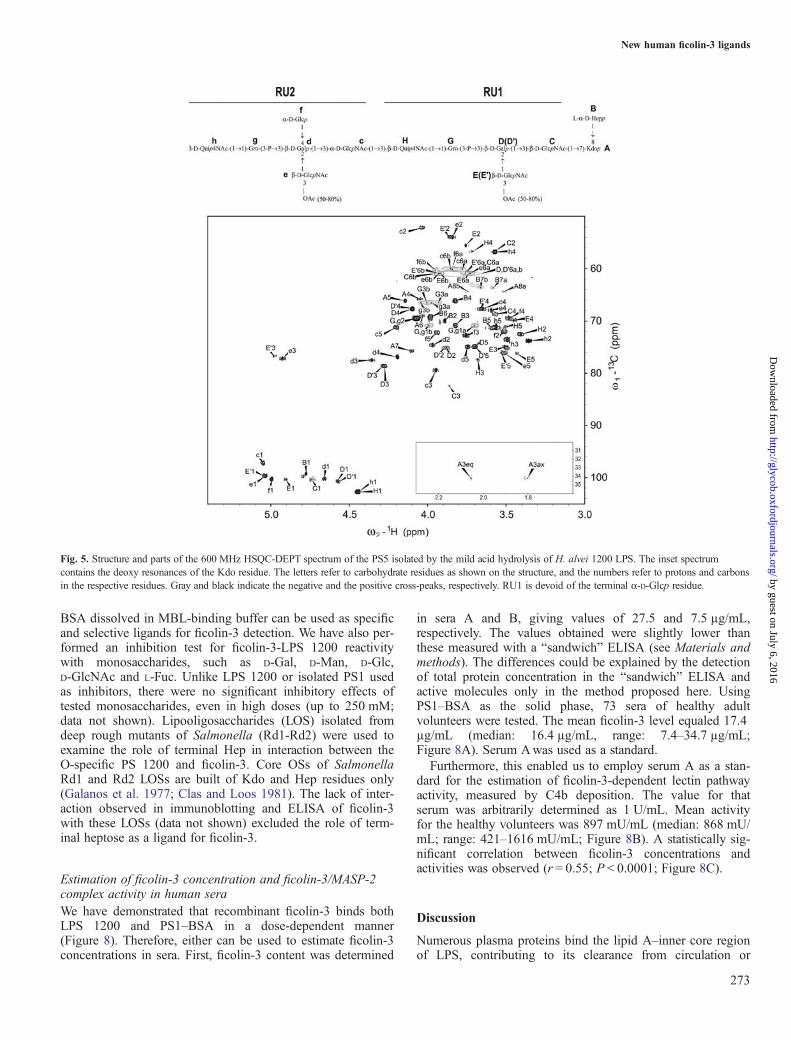
location of the Hep-Kdo disaccharide. According to MS data,PS5 represents the O-specific PS with the highest degree ofpolymerization (two RUs) for which NMR analysis of the
Fig. 1. Silver stain SDS–PAGE (A) and Western blotting analysis of the interaction of human serum-derived ficolin-3 (B) and MBL (C) with LPSs of H. alvei.Lanes are depicted with the strain number. Normal human serum A and B, 30-fold diluted, was used as a source of ficolin 3 and MBL, respectively.
New human ficolin-3 ligands
269
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
entire molecule including Hep-Kdo disaccharide is still feas-ible. The one-dimensional 1H-NMR and two-dimensional 1H,13C-HSQC (heteronuclear single quantum coherence)-DEPT(distortionless enhancement by polarization transfer) NMRspectra of the PS5 (Figure 5) contained 12 signals in theanomeric region of the spectra and a Kdo spin system. Allidentified spin systems are presented in Supplementary data,Table SI. The chemical shift values were compared with thepreviously published NMR data for the RU of the O-specificPS (PS1) isolated from H. alvei 1200 LPS (Supplementarydata, Figure S1; Katzenellenbogen et al. 1992, 1999; Daget al. 2004). The results indicated a basic structure of the RU,which is in agreement with previously published data (Daget al. 2004; Figure 5). Since the structure of the RU of LPS1200 is already known, a full description of the spin systemswas limited to newly identified characteristic structuralelements of PS5 (A, B, C and h), indicating the reducing andnon-reducing ends. The remaining residues were presented inSupplementary data (Tables SI and SII). The sugar residuesidentified are denoted by upper-case letters (RU1, Kdo andHep residues) or lower-case letters (RU2) as shown in thestructure of the isolated PS5 (Figure 5).Residue A was identified as a 7,8-disubstituted
3-deoxy-α-D-manno-oct-2-ulopyranosonic acid (Kdo) on thebasis of characteristic deoxy proton signals at δ 1.80 and2.05 ppm (Figure 5, inset spectrum), and a chemical shift ofthe C-7 (δ 75.7 ppm) and C-8 (δ 64.4 ppm) signals(Birnbaum et al. 1987; Vinogradov and Bock 1999).Residue B with the H-1/C-1 signals at δ 4.77/99.3 ppm was
recognized as the terminal L-glycero-α-D-manno-Hepp residuedue to the small vicinal couplings between H-1, H-2 and H-3and similar chemical shifts as in the monosaccharideL-α-D-Hep (Vinogradov and Bock 1999).Residue C with the H-1/C-1 signals at δ 4.74/100.4 ppm
was assigned as the 3-substituted β-D-GlcpNAc residue (RU1constituent) based on the low chemical shift of the C-2 signal
Fig. 2. Interaction of ficolin-3 with LPSs of H. alvei in ELISA. Binding officolin-3 to LPSs (A) and ficolin-3/MASP-2 complex-dependent C4bdeposition (complement lectin pathway activity) (B). (C) Binding of serumficolin-3 to H. alvei 1200 LPS and its inhibition by calcium and magnesiumchelators. Plates were coated with LPS (5 µg/well), incubated with humanserum with or without the presence of chelators: EDTA, EGTA or EGTA/Mg2+,serially diluted in GVB2+. Data represent the mean values of two replicates.
Fig. 3. Binding of recombinant ficolin-3 to H. alvei 1200 bacteria in a flowcytometry assay. Formaldehyde-fixed bacterial cells were suspended in GVB2+
buffer to OD600 = 1 (OD, optical density). The suspension (100 μL) wascentrifuged and the pellet was re-suspended in 50 µL of the same buffer andthen incubated with (A) or without (B) 1 µg of rh ficolin-3 (R&D Systems,US) for 1 h at 37°C. The bound protein was detected with mouse anti-humanficolin-3 mAb followed by FITC-labeled anti-mouse Ig Ab.
A Swierzko et al.
270
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
(δ 56.8 ppm), the relatively high chemical shift of the C-3signal (δ 82.5) and the large vicinal coupling constantsbetween all ring protons.Residue h at 4.44/102.8 ppm was recognized as the term-
inal β-D-Quip4NAc residue (RU2 constituent) based on thecharacteristic chemical shift of the C-4 signal (δ 56.8 ppm)and the signal for an exocyclic CH3 group (δ 1.20 and 16.9ppm).
The heteronuclear multiple-bond correlation (HMBC),nuclear Overhauser effect spectroscopy (NOESY) androtating-frame Overhauser effect spectroscopy (ROESY)experiments (Supplementary data, Table SII) showed inter-residue connectivities between adjacent sugar residues andthus provided the sequence in PS5 (Figure 5). The linkagebetween residues C and A was deduced from an indicativecross-peak in the HMBC spectrum between C-1 of residue C
Fig. 4. Structural analysis of the O-specific PSs isolated from H. alvei 1200 LPS. (A) Structures of PS1–PS5. (*) The PS1–BSA conjugate had the Kdo residuepresent in a 1-carboxy-3-deoxy-octitol form, (**) missing α-D-Glcp residue in the first RU, (***) non-stoichiometric substitution of the O-acetyl group.MALDI-TOF mass spectra of the PS5 (B) and PS4 (C). m/z values represent monoisotopic masses (Table I). The low-intensity ions marked with asterisks couldbe explained by products of the laser-derived degradation of the PS4.
New human ficolin-3 ligands
271
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

and H-7 of residue A (Supplementary data, Table SII) and theKdo substitution pattern. The newly identified element,→3)-β-D-GlcpNAc-[L-α-D-Hepp-(1→ 8)-]-(1→ 7)-Kdo (resi-dues C, B and A), indicated the reducing end of the PS5, thusthe linkage between the first RU of the O-specific PS andthe Hep-Kdo disaccharide. The presence of terminalβ-D-Quip4NAc (h) indicated the structure of the biologicalRU of LPS 1200 (Figures 4A and 5). Additionally, in agree-ment with studies of Katzenellenbogen et al. (2005), anomericconfiguration of D-GlcpNAc (residue C) in RU1 is alteredfrom α to β, compared with that of the subsequent RUs of theO-specific PS (e.g. RU2). The herein presented structural dataindicated that regardless of the number of RUs, there wasalways the Hep-Kdo disaccharide at the reducing end as alinking element between the O-specific chain and the core OSof the LPS.
Conjugation of the reduced PS1 with BSADiscovered interaction prompted us to synthesize a glycocon-jugate of PS1 isolated from LPS 1200 with BSA (PS1-BSA)in order to prepare a tool for ficolin-3 concentration andficolin-3-dependent lectin pathway activity measurements inbody fluids. The conjugation was performed according toLonngren and Goldstein (1978), through the Kdo residue ofPS1 reduced to 1-carboxy-3-deoxy-octitol. The reduced PS1was checked by one-dimensional NMR. The data confirmed
the PS structure but also revealed de-O-acetylation, whichoccurred during the reduction procedure (data not shown).The PS in the conjugate PS1–BSA represented the O-specificPS devoid of a major part of the core OS region. The totalcarbohydrate:protein ratio in the PS1–BSA was 1:1 and wasdetermined with the use the phenol-sulfuric acid procedure(Fox and Robyt 1991) combined with the protein concen-tration measurement (A280 nm). Molecular weight of the conju-gate was estimated by SDS–PAGE analysis with the use ofprotein molecular weight markers within the range 40–200kDa (Supplementary data, Figure S3). Two methods wereused for the detection of the PS1–BSA: Coomassie stainingfor the detection of BSA (weak staining due to the PS substi-tution) and immunoblotting with the use of polyclonal anti-bodies (Abs) specific for the O-specific PS of LPS 1200 forthe detection of carbohydrate moiety. Obtained PS1–BSAconjugate was heterogeneous mixture due to the heterogeneityof used PSs (the number of RUs). Estimated mass of theglycoconjugate was within a range of 72 to >200 kDa withstronger diffused bands around 130, 170 and >200 kDa(Supplementary data, Figure S3). Due to the described hetero-geneity of the PSs obtained results allowed only to estimatethe number of O-specific PS RUs (molecular weight �1071Da) per conjugate molecule (molecular weight of BSA �67kDa). In the case of PS1–BSA, the calculated number wasaround 5 RUs (the conjugate of 72 kDa), 59 RUs (the conju-gate of 130 kDa), 96 RUs (the conjugate of 170 kDa) and>120 RUs (the conjugate of >200 kDa).
Binding of recombinant ficolin-3 to the PS1–BSAThe conjugate of the reduced PS1 and BSA (PS1–BSA) wasused to assess binding between PS1 and human serum-derived or rh ficolin-3 in ELISA. The PS1 used for the prep-aration of the conjugate was also devoid of O-acetyl groups aswas determined by NMR analysis (data not shown). Thebinding was compared with that of native and de-O-acetylatedLPS 1200. Similar dose-dependent binding of both serum andrh ficolin-3 was demonstrated for all ligands with the strongestbinding to de-O-acetylated LPS (Figure 6A and B). Thisshowed that the presence or the lack of O-acetyl groups didnot affect significantly the interaction between ficolin-3 andthe O-specific PS of LPS 1200.
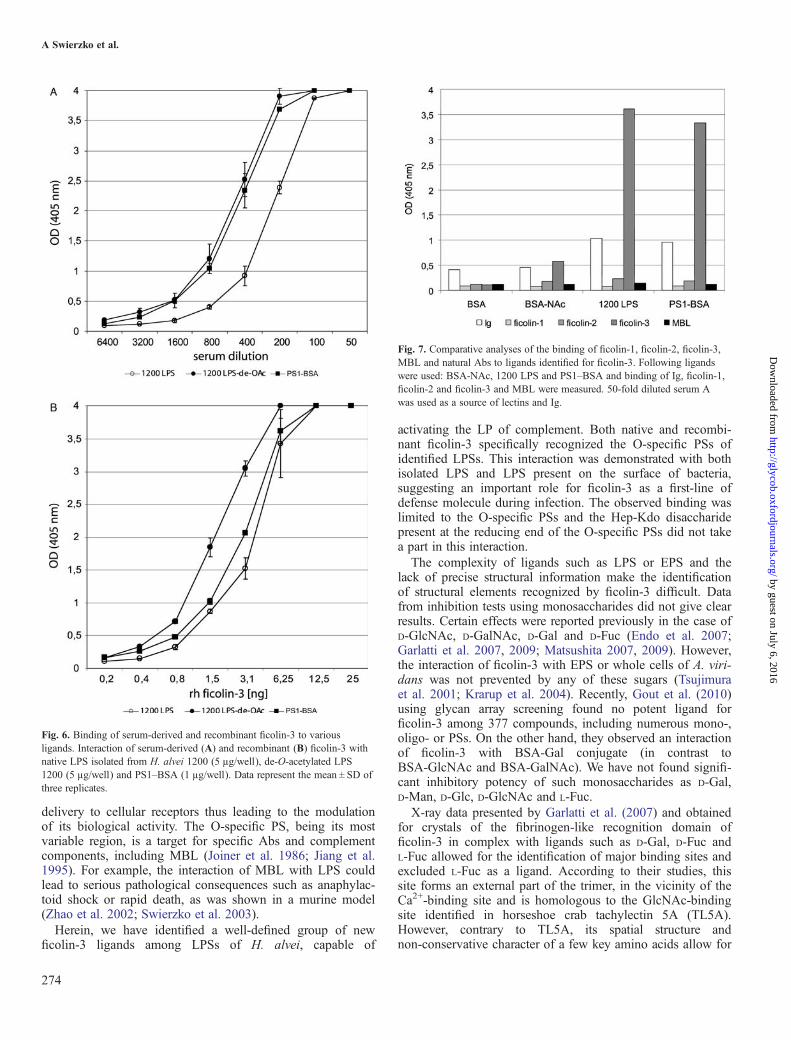
Ficolin-3 selectivity for interaction with PS1–BSAand LPS 1200In order to compare the specificity of binding of ficolin-3 toH. alvei 1200 LPS, PS1–BSA and BSA-NAc, an ELISAanalysis was performed (Figure 7). A very strong reactivity officolin-3 with H. alvei 1200 LPS and PS1–BSA was con-firmed. These products were not recognized by other ficolins(1 and 2) or MBL. However, the human sera used in thisexperiment contained some natural Abs reacting with nativeLPS and PS1–BSA. None of the lectins recognized BSA, andonly ficolin-3 reacted weakly with BSA-NAc (Figure 7).Slightly higher A405 nm values for the ficolin-3/BSA-NAcinteraction were recorded when the N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) (HEPES) buffer rec-ommended by Munthe-Fog et al. (2009) was used (data notshown). These results showed that H. alvei 1200 LPS or PS1–
Table I. Interpretation of MALDI-TOF mass spectra of fractions PS4, PS5and OS1 isolated from LPS H. alvei 1200
Observed iona
(m/z)Interpretationof ion
Schematic structure of molecularion (M)
Fraction OS11380.99 [M–H]− RUi-[Hep-Kdo] + 1 × OAc1363.00 [M–H2O–H]
− RUi-[Hep-Kdo] + 1 × OAc1338.98 [M–H]− RUi-[Hep-Kdo]1320.97 [M–H2O–H]
− RUi-[Hep-Kdo]1420.88b [M–H]− Core OS1402.90b [M–H2O–H]
− Core OS1336.92b [M–H2O–H]
− Core OS − EtnFraction PS52656.14 [M–H]− RU-RU-[Hep-Kdo] + 2 × OAc2494.08 [M–H]− RU-RUi-[Hep-Kdo] + 2 × OAc2452.07 [M–H]− RU-RUi-[Hep-Kdo] + 1 × OAc2476.07 [M–H2O–H]
− RU-RUi-[Hep-Kdo] + 2 × OAc2434.09 [M–H2O–H]
− RU-RUi-[Hep-Kdo] + 1 × OAc2290.02 [M–H]− RU-RUi-[Hep-Kdo] + 1 × OAc2248.95 [M–H]− RU-RUi-[Hep-Kdo]
Fraction PS43607.66 [M–H]− RU-RU-RUi-[Hep-Kdo] + 3 × OAc3565.67 [M–H]− RU-RU-RUi-[Hep-Kdo] + 2 × OAc3629.64 [M +Na–H]− RU-RU-RUi-[Hep-Kdo] + 3 × OAc3587.55 [M +Na–H]− RU-RU-RUi-[Hep-Kdo] + 2 × OAc3651.69 [M + 2Na–H]− RU-RU-RUi-[Hep-Kdo] + 3 × OAc3589.67 [M–H2O–H]
− RU-RU-RUi-[Hep-Kdo] + 3 × OAc3547.80 [M–H2O–H]
− RU-RU-RUi-[Hep-Kdo] + 2 × OAc3361.90 [M–H]− RU-RUi-RUi-[Hep-Kdo] + 1 × OAc3200.54 [M–H]− RUi-RUi-RUi-[Hep-Kdo] + 1 × OAc
am/z values correspond to monoisotopic mass; RU, a complete repeating unitof the O-specific PS; RUi, an incomplete RU devoid of terminal α-D-Glcp;OAc, O-acetyl group.bIons corresponded to core OSs (core OS = 2 × Hex + 3 × Hep + Kdo + 3P +Etn; Jachymek et al. 1995).
A Swierzko et al.
272
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
BSA dissolved in MBL-binding buffer can be used as specificand selective ligands for ficolin-3 detection. We have also per-formed an inhibition test for ficolin-3-LPS 1200 reactivitywith monosaccharides, such as D-Gal, D-Man, D-Glc,D-GlcNAc and L-Fuc. Unlike LPS 1200 or isolated PS1 usedas inhibitors, there were no significant inhibitory effects oftested monosaccharides, even in high doses (up to 250 mM;data not shown). Lipooligosaccharides (LOS) isolated fromdeep rough mutants of Salmonella (Rd1-Rd2) were used toexamine the role of terminal Hep in interaction between theO-specific PS 1200 and ficolin-3. Core OSs of SalmonellaRd1 and Rd2 LOSs are built of Kdo and Hep residues only(Galanos et al. 1977; Clas and Loos 1981). The lack of inter-action observed in immunoblotting and ELISA of ficolin-3with these LOSs (data not shown) excluded the role of term-inal heptose as a ligand for ficolin-3.
Estimation of ficolin-3 concentration and ficolin-3/MASP-2complex activity in human seraWe have demonstrated that recombinant ficolin-3 binds bothLPS 1200 and PS1–BSA in a dose-dependent manner(Figure 8). Therefore, either can be used to estimate ficolin-3concentrations in sera. First, ficolin-3 content was determined
in sera A and B, giving values of 27.5 and 7.5 µg/mL,respectively. The values obtained were slightly lower thanthese measured with a “sandwich” ELISA (see Materials andmethods). The differences could be explained by the detectionof total protein concentration in the “sandwich” ELISA andactive molecules only in the method proposed here. UsingPS1–BSA as the solid phase, 73 sera of healthy adultvolunteers were tested. The mean ficolin-3 level equaled 17.4µg/mL (median: 16.4 µg/mL, range: 7.4–34.7 µg/mL;Figure 8A). Serum Awas used as a standard.Furthermore, this enabled us to employ serum A as a stan-
dard for the estimation of ficolin-3-dependent lectin pathwayactivity, measured by C4b deposition. The value for thatserum was arbitrarily determined as 1 U/mL. Mean activityfor the healthy volunteers was 897 mU/mL (median: 868 mU/mL; range: 421–1616 mU/mL; Figure 8B). A statistically sig-nificant correlation between ficolin-3 concentrations andactivities was observed (r = 0.55; P < 0.0001; Figure 8C).
Discussion
Numerous plasma proteins bind the lipid A–inner core regionof LPS, contributing to its clearance from circulation or
Fig. 5. Structure and parts of the 600 MHz HSQC-DEPT spectrum of the PS5 isolated by the mild acid hydrolysis of H. alvei 1200 LPS. The inset spectrumcontains the deoxy resonances of the Kdo residue. The letters refer to carbohydrate residues as shown on the structure, and the numbers refer to protons and carbonsin the respective residues. Gray and black indicate the negative and the positive cross-peaks, respectively. RU1 is devoid of the terminal α-D-Glcp residue.
New human ficolin-3 ligands
273
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
delivery to cellular receptors thus leading to the modulationof its biological activity. The O-specific PS, being its mostvariable region, is a target for specific Abs and complementcomponents, including MBL (Joiner et al. 1986; Jiang et al.1995). For example, the interaction of MBL with LPS couldlead to serious pathological consequences such as anaphylac-toid shock or rapid death, as was shown in a murine model(Zhao et al. 2002; Swierzko et al. 2003).Herein, we have identified a well-defined group of new
ficolin-3 ligands among LPSs of H. alvei, capable of
activating the LP of complement. Both native and recombi-nant ficolin-3 specifically recognized the O-specific PSs ofidentified LPSs. This interaction was demonstrated with bothisolated LPS and LPS present on the surface of bacteria,suggesting an important role for ficolin-3 as a first-line ofdefense molecule during infection. The observed binding waslimited to the O-specific PSs and the Hep-Kdo disaccharidepresent at the reducing end of the O-specific PSs did not takea part in this interaction.The complexity of ligands such as LPS or EPS and the
lack of precise structural information make the identificationof structural elements recognized by ficolin-3 difficult. Datafrom inhibition tests using monosaccharides did not give clearresults. Certain effects were reported previously in the case ofD-GlcNAc, D-GalNAc, D-Gal and D-Fuc (Endo et al. 2007;Garlatti et al. 2007, 2009; Matsushita 2007, 2009). However,the interaction of ficolin-3 with EPS or whole cells of A. viri-dans was not prevented by any of these sugars (Tsujimuraet al. 2001; Krarup et al. 2004). Recently, Gout et al. (2010)using glycan array screening found no potent ligand forficolin-3 among 377 compounds, including numerous mono-,oligo- or PSs. On the other hand, they observed an interactionof ficolin-3 with BSA-Gal conjugate (in contrast toBSA-GlcNAc and BSA-GalNAc). We have not found signifi-cant inhibitory potency of such monosaccharides as D-Gal,D-Man, D-Glc, D-GlcNAc and L-Fuc.X-ray data presented by Garlatti et al. (2007) and obtained
for crystals of the fibrinogen-like recognition domain officolin-3 in complex with ligands such as D-Gal, D-Fuc andL-Fuc allowed for the identification of major binding sites andexcluded L-Fuc as a ligand. According to their studies, thissite forms an external part of the trimer, in the vicinity of theCa2+-binding site and is homologous to the GlcNAc-bindingsite identified in horseshoe crab tachylectin 5A (TL5A).However, contrary to TL5A, its spatial structure andnon-conservative character of a few key amino acids allow for
Fig. 6. Binding of serum-derived and recombinant ficolin-3 to variousligands. Interaction of serum-derived (A) and recombinant (B) ficolin-3 withnative LPS isolated from H. alvei 1200 (5 µg/well), de-O-acetylated LPS1200 (5 µg/well) and PS1–BSA (1 µg/well). Data represent the mean ± SD ofthree replicates.
Fig. 7. Comparative analyses of the binding of ficolin-1, ficolin-2, ficolin-3,MBL and natural Abs to ligands identified for ficolin-3. Following ligandswere used: BSA-NAc, 1200 LPS and PS1–BSA and binding of Ig, ficolin-1,ficolin-2 and ficolin-3 and MBL were measured. 50-fold diluted serum Awas used as a source of lectins and Ig.
A Swierzko et al.
274
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

some flexibility of binding site (Garlatti et al. 2007). Thecrucial role of Ca2+ suspected previously by Garlatti et al.(2007) was further evidenced by Hein et al. (2010) for abinding of ficolin-3 to BSA-NAc and subsequent complementactivation. However, an addition of magnesium ions did notallow ficolin-3 to recognize BSA-NAc (Hein et al. 2010).According to our observation, the interaction betweenficolin-3 and LPS 1200 requires the presence of both calciumand magnesium ions, as was shown by a strong inhibition ofbinding of ficolin-3 to LPS by EDTA or EGTA and muchweaker in the case of EGTA/Mg2+ (Figure 2C).Crystallographic analysis done by Garlatti et al. (2007) was
also performed for ficolin-2 and monosaccharide ligands.Further studies of ficolin-2 binding to more complex ligand(1,3-β-D-glucan, built of four Glc residues) revealed a novelextended recognition area with three additional binding sitesinvolved. The shape of the recognition area allowed forbinding of PSs and provided the observed plasticity of recog-nition by ficolin-2. Due to the lack of a proper, structurallycharacterized ligand for ficolin-3, similar analysis has notbeen performed for ficolin-3 and its natural ligands such asEPS of A. viridans. Despite of some differences in structure,amino acids composition and ligand specificity betweenficolin-2 and ficolin-3, ficolin-3 can also have similar affinitytoward complex ligands characterized by repetition pattern,especially at the surface of microorganism.According to the previous studies, it could be assumed that
the presence of D-GlcNAc, D-GalNAc, D-Gal and D-Fuc maydetermine the binding specificity of ficolin-3 (Endo et al.2007; Garlatti et al. 2007, 2009; Matsushita 2007, 2009).Moreover, the example of BSA-NAc supports a conclusionabout an important role of the N-acetyl groups. However, theaffinity for acetylated compounds, including BSA-NAc andHSA-NAc (acetylated-human serum albumin), was alsoreported for ficolin-2 and ficolin-1 (Frederiksen et al. 2005;Ng et al. 2007; Lacroix et al. 2009; Munthe-Fog et al. 2009).The advantage of PS1 is a much stronger LP activation incomparison to BSA-NAc. Moreover, the O-specific PSs ident-ified represent components of the natural target for this lectinand are not recognized by other ficolins or MBL.Some structures of H. alvei O-specific PSs, identified here
as exclusive ligands for ficolin-3, are known (Supplementarydata, Figure S1; Katzenellenbogen et al. 1992, 1999;Dag et al. 2004). Their comparison clearly shows that thepresence of D-GlcpNAc, D-Quip4NAc and O-acetyl groups isa common feature of the O-specific PSs isolated from LPS1200, 1203, 1205 and 23. However, no substantial differencein the reactivity of ficolin-3 with native and de-O-acetylatedLPS 1200 was observed. Therefore, the O-acetyl groups donot constitute a factor for this interaction. It might be specu-lated that the disaccharide, →3)-α-D-GlcNAc-(1→3)-β-D-Qui4NAc-(1→, could be a common fragment in 23,1200, 1203 and 1205 LPSs, which is recognized by ficolin-3.Moreover, the presence of N-acetyl groups is the only struc-tural feature shared by all presented here O-specific PSs andBSA-NAc. Therefore, it suggests that the structural arrange-ment of N-acetyl groups may determine the pattern recog-nition. Recently, Thomsen et al. (2011) have hypothesized
Fig. 8. An employment of the PS1–BSA conjugates as a diagnostic tool. Anestimation of ficolin-3 concentration (A), ficolin-3/MASP-2 complex activity(B) in normal human sera of 73 healthy adult volunteers and correlationbetween concentration and activity (C).
New human ficolin-3 ligands
275
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

differences in binding patterns of all human ficolins resultfrom different spacing of acetyl groups in the used ligands orare due to oligomerization defects observed for recombinantproteins. It should be stressed that there are some other“N-acetylated” LPSs among H. alvei LPSs which were notbound by ficolin-3 (e.g. LPS 1192) as was shown by dot blot-ting, ELISA and flow cytometry.An advantage of the described PSs is their selectivity
toward ficolin-3. We have shown previously that LPSs of H.alvei belong to the group of atypical endotoxins, due to thepresence of the Kdo residue in the outer core region(Lukasiewicz et al. 2009). This feature, characteristic for mostof H. alvei LPSs, including LPS 1200, 1203 and 1205(Lukasiewicz et al., unpublished results), makes possible easyisolation of the O-specific PS devoid of large part of the coreregion, which could be bound by other immune factors suchas MBL (Figure 1C) or immunoglobulins. This featureprompted us to prepare PS1–BSA neoglycoconjugate as a toolto measure ficolin-3 concentration in human serum as well asficolin-3/MASP-2-dependent complement LP activity (in theC4b deposition assay), without interference from MBL,ficolin-2 and ficolin-1 or natural Abs. It may also be used forthe isolation of ficolin-3 from body fluids. The employmentof hypertonic MBL-binding buffer (Petersen et al. 2001) forboth concentration and complement activation assays seems tobe more reasonable than HEPES buffer (Munthe-Fog et al.2009), due to higher values of absorbance. Moreover, theMBL-binding buffer prevents classical complement pathwayactivation (Petersen et al. 2001), which is extremely importantwhen C4b deposition is determined. According to previousstudies, the average concentration of ficolin-3 in human serais within the range 20–30 µg/mL (Krarup et al. 2005;Munthe-Fog et al. 2008, 2009; Andersen et al. 2009). Themethods previously used employed monoclonal Abs (mAbs)(“sandwich ELISA”; Krarup et al. 2005; Munthe-Fog et al.2008, 2009; Andersen et al. 2009) or A. viridans (Kilpatricket al. 2003) EPS as a solid phase for ELISA. Until now thereis no “golden standard” for the determination of both ficolin-3level and activity. We present an alternative, functional assayuseful for both scientific and diagnostic purposes.The exploration of the relationship between innate immu-
nity mechanisms and disease/risk factors depends on appro-priate experimental tools allowing the isolation, purification aswell as the measurement of concentration and specific activity.This is difficult in the case of such a complex system ascomplement, involving several key molecules (immunoglobu-lin G (IgG)/immunoglobulin M (IgM), MBL, ficolin-1,ficolin-2 and ficolin-3 etc.) with wide specificity. Genesencoding MBL and ficolins are characterized by polymorph-isms, which are correlated with function and also changes in aspecific lectin concentration. Thus, other approaches based ona specific, functional ligand for ficolin-3 could give resultscorrelated with functional and effective form of these lectins.
Materials and methodsBacterial strainsHafnia alvei strains PCM 1, 1M, 2, 17, 23, 31, 32, 37, 38, 39,399, 481, 537, 600, 744, 974, 2694, 1187, 1188, 1190, 1191,
1192, 1195, 1196, 1198, 1200, 1203, 1204, 1205, 1206, 1207,1208, 1209, 1210, 1211, 1212, 1213, 1214, 1215, 1218, 1220,1221, 1222, 1224 and 114-60 were obtained from the PCM atthe Ludwik Hirszfeld Institute of Immunology andExperimental Therapy (Wroclaw, Poland). The bacteria weregrown in Davis medium, killed with 0.5% phenol and centri-fuged using a CEPA flow laboratory centrifuge (Peterssonet al. 1997).
Abs and seramAbs, mouse anti-human MBL (clone HYB 131-01) andmouse anti-human ficolin-1 (clone ABS-036-01), came fromBioPorto, Denmark. Mouse anti-human ficolin-3 mAb (clone4H5) and anti-human ficolin-2 mAb (clone 2F5) wereprepared by Misao Matsushita. Polyclonal rabbit anti-humanC4 Abs were obtained from Sigma, US. Secondary Abs,horseradish peroxidase (HRP)- or fluorescein isothiocyanate(FITC)-conjugated rabbit anti-mouse Ig, HRP-conjugatedrabbit anti-human IgG, A and M and HRP-conjugated goatanti-rabbit Ig, were obtained from DAKO, Denmark.Normal human sera (A and B) were collected from healthy
adult volunteers. Serum A had normal ficolin-3 and ficolin-2levels (32.7 and 3.5 µg/mL, respectively) and low MBL con-centration (0.2 µg/mL). Serum B had low ficolin-3 andficolin-2 levels (12 and 2 µg/mL, respectively) and normalMBL concentration (2 µg/mL). The levels of ficolin-3 andficolin-2 in these sera were kindly measured (by “sandwich”ELISA) by Prof. Jens C. Jensenius (Department of MedicalMicrobiology and Immunology, University of Aarhus,Denmark) and Dr. David C. Kilpatrick (Scottish NationalBlood Transfusion Service, National Science Laboratory,Edinburgh, Scotland), respectively. MBL concentrations weredetermined as described previously (Cedzynski et al. 2004).Ficolin-1 was not determined in either serum. Normal humansera tested for ficolin-3 concentration and ficolin-3/MASPcomplex-dependent complement activity were obtained from73 healthy adults. The local ethical committee approved thestudy and samples were obtained with written informedconsent of the volunteers.
Preparation of the BSA-NAcThe BSA-NAc was prepared as described by Munthe-Foget al. (2009).
Preparation of LPSs and O-specific PSLPSs were extracted from bacterial cells by the hot phenol/water method (Westphal and Jann 1965) and purified asdescribed previously (Petersson et al. 1997). Poly- and OSswere isolated by mild acidic hydrolysis and fractionated asdescribed elsewhere (Dag et al. 2004).
De-O-acetylation of LPSLPS 1200 was suspended in 12.5% NH3 in water and incu-bated for 16 h at 22°C, followed by dilution with water andlyophilization.
A Swierzko et al.
276
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Mass spectrometryMALDI-TOF MS was carried out on an Applied Biosystems4800 MALDI-TOF/TOF™ instrument. 2,4,6-Trihydroxyaceto-phenone (25 mg/mL, acetonitrile:water, 1:1, v/v) was used as amatrix. Spectra were scanned in the range m/z 1500–6000.External calibration in the negative-ion mode was appliedusing the MSCAL2 calibration kit (ProteoMass™).
NMR spectroscopyNMR spectra were obtained on a Bruker Avance 600 MHzspectrometer at the Laboratory of Structural Analyses,Wroclaw University of Technology. NMR spectra of the O-specific PS were obtained for 2H2O solutions at 30°C usingacetone (δH 2.225 and δC 31.05) as internal reference. In 31PNMR spectroscopy experiments, phosphorus resonances werereferenced to an external 85% phosphoric acid (δ 0 ppm) in aseparate experiment. The data were acquired and processedusing standard Bruker software and assigned with the use ofSPARKY software (Goddard and Kneller 2001). The signalswere assigned by two-dimensional experiments (correlationspectroscopy (COSY), clean-total correlation spectroscopy(TOCSY), NOESY, ROESY, HMBC, HSQC-DEPT andHSQC with and without carbon decoupling). In theclean-TOCSY experiments, the mixing times used were 30,60 and 100 ms. The delay time in HMBC was 60 or 65 ms,and the mixing time for NOESY and ROESY was 200 and500 ms, respectively.
Reduction of the fraction PS1The PS1 (22 mg/mL of H2O) was reduced with NaBH4
(10 mg) for 16 h at 37°C. The solution was acidified withCH3COOH to pH 4.5 and purified by gel permeation chrom-atography, performed on a Bio-Gel P-2 column equilibratedwith 0.05 M pyridine/acetic acid buffer at pH 5.6. The frac-tion containing reduced PS1 was freeze-dried, checked byone-dimensional NMR analysis and used for conjugationwith BSA.
Conjugation of the reduced PS1 with BSAThe conjugation was performed according to Lonngren andGoldstein (1978). Briefly, the reduced PS1 was dissolved inH2O, and the pH of the solution was adjusted to 4.75, followedby the addition of 1-ethyl-3-(dimethylaminopropyl)-carbodiimide hydrochloride. The reaction was carried out for1 h at 22°C, and pH 4.75 was maintained throughout by theaddition of HCl. After 1 h of additional incubation, 0.4 mL ofBSA solution (5 mg/mL) was added to the solution and theconjugation was carried out for 4 h at 22°C. The conjugate(PS1–BSA) was purified by gel permeation chromatography,using a Sephadex G-100 column equilibrated with 5% ethanol.The fraction of the conjugate obtained was checked for thepresence of protein (A280 nm measurement) and PS1 (a dot blot-ting with the use of polyclonal rabbit serum against H. alvei1200). The total carbohydrate:protein ratio in the PS1–BSAwas determined with the use of protein concentration measure-ment (A280 nm) combined with the phenol-sulfuric acid pro-cedure (Fox and Robyt 1991). Reduced fraction PS1 was usedas a standard for phenol-sulfuric acid total carbohydrate
determination. Molecular weight of the conjugate was esti-mated by SDS–PAGE according to the method of Laemmli(1970). The conjugate fractions were visualized by Coomassiestaining and immunoblotting performed on the SDS–PAGE-separated (7.5% gel) conjugate fractions with the use ofrabbit polyclonal serum against H. alvei 1200, as describedpreviously (Lukasiewicz et al. 2009). Following protein mol-ecular weight markers were used: M1 (prestained, 161-0309,Bio-Rad, US), M2 (prestained, SM0671, Fermentas,Lithuania) and M3 (unstained, SM0661, Fermentas,Lithuania).
Western blotting and dot-blot analysisHafnia alvei LPSs were separated on 15% polyacrylamidegels (Laemmli 1970) and then transferred onto polyvinylidenefluoride membranes (Bio-Rad). Membranes were blockedwith Superblock solution (ThermoScientific, US) and incu-bated overnight (at 4°C) with serum A (for ficolin-3 andficolin-2 detection) or serum B (for MBL detection), 30-folddiluted in a high-ionic strength MBL-binding buffer (20 mMTris/HCl, 10 mM CaCl2, 1 M NaCl, pH 7.4; Petersen et al.2001), supplemented with 0.1% BSA or a pool of sera A andB (1:1, each prediluted 1:15 in the same buffer, for ficolin-1detection). Then, primary Ab (anti-MBL or anti-ficolin-3or anti-ficolin-2 or anti-ficolin-1, respectively) was added,followed by the HRP-conjugated rabbit anti-mouse Ig, as asecondary Ab. Membranes were incubated with electrochemi-cal luminescence (ECL) substrate (Santa Cruz Biotechnology,US) and then exposed to an X-ray film. For dot-blot analysis,10 µg of the tested LPS was applied directly on the nitrocellu-lose membrane (Schleicher and Schuell, Germany). Afterblocking, the membrane was incubated with serum A as asource of ficolin-3 and the binding of ficolin-3 to LPS wasdetected as for Western blotting.
Binding of ficolin-3 to bacterial cellsBacterial cultures were washed out from the solid mediumwith PBS, centrifuged and fixed with 1.5% formaldehyde.After washing with cold PBS, cell pellets were suspended inveronal-buffered saline supplemented with 0.1% gelatin, 0.3mM CaCl2 and 2 mM MgCl2 (GVB2+, Gelatin VeronalBuffer) to OD = 1 (A600 nm). The suspension (100 μL) wascentrifuged. The pellet was re-suspended in 50 µL of GVB2+
buffer and then incubated with or without 1 µg of humanrecombinant ficolin-3 (R&D Systems, US) for 1 h at 37°C.The bound protein was detected with mouse anti-humanficolin-3 mAb followed by FITC-labeled anti-mouse Ig Ab.The reactivity was estimated using Cytomics FC 500 MPLBeckman-Coulter flow cytometer.
Inhibition of binding of ficolin-3 to bacterial cellsELISA inhibition was performed by analogy to the methoddescribed in section Inhibition of ficolin-3 interaction withLPS by calcium and magnesium chelators with the followingmodifications. Human serum (serum A, prediluted 1:50) wasincubated for 30 min at 28°C with different amounts of H.alvei 1200 LPS. Next, the mixture was transferred to the wellsof microtiter plate coated with suspension of H. alvei 1200
New human ficolin-3 ligands
277
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

cells in PBS (10 μg/well). After 2 h incubation at 37°C,the bound H ficolin was detected with the use ofH-ficolin-specific mAb and HRP-labeled anti-mouse Ig.Absorbance (A405 nm) was measured with the use of aBenchmark Plus microplate spectrophotometer (Bio-Rad).
Binding of lectins and immunoglobulins to various ligandsMicrotitre MaxiSorp U96 plates (NUNC, Denmark) werecoated with LPS (5 µg/well), PS1–BSA (1 µg/well), BSA(0.5 µg/well) or BSA-NAc (0.5 µg/well) dissolved in PBS.After overnight incubation at 4°C, plates were washed withtris buffered saline (TBS)-Ca2+ containing 0.05% Tween-20and blocked with 0.1% BSA in TBS-Ca2+ (2 h, 37°C).Human serum (A or B) or human recombinant ficolin-3 wasdiluted in MBL-binding buffer containing 0.1% BSA, withoutTriton X-100. Plates were incubated overnight at 4°C. Then,to estimate ficolin-3 binding, specific mAbs (0.5 µg/mL in0.1% BSA/TBS-Ca2+) were added and after 2 h at 37°C fol-lowed by detection with HRP-conjugated rabbit anti-mouseIg. ABTS (Sigma) was employed as a substrate for peroxi-dase. Absorbance (A405 nm) was measured with the use of aBenchmark Plus microplate spectrophotometer (Bio-Rad).Alternatively, to test the binding of ficolin-2, ficolin-1, MBLand immunoglobulins appropriate primary and correspondingHRP-conjugated secondary Abs were employed.
Inhibition of ficolin-3 interaction with LPS by calciumand magnesium chelatorsInhibition of ficolin-3 interaction with LPS by Ca2+ and Mg2+
chelators was tested according to the modified proceduredescribed by Hein et al. (2010). Briefly, microtitre MaxiSorpU96 plates (NUNC) were coated with H. alvei 1200 LPS (5µg/well), dissolved in PBS. After overnight incubation at 4°C,plates were washed with TBS-Ca2+ containing 0.05%Tween-20 and blocked with 0.1% BSA in TBS-Ca2+ (2 h, 37°C). Human serum (A), prediluted 1:25 was mixed with anequal volume of PBS, 20 mM solutions of EDTA, EGTA or20 mM EGTA/10 mM Mg2+ (EGTA/Mg2+) and then seriallydiluted in GVB2+. Plates were incubated for 1 h at 37°C.Then, ficolin-3-specific mAbs (clone 4H5, Hycult,Netherlands; 0.5 µg/mL in 0.1% BSA/TBS-Ca2+) were addedand after 1 h at 37°C followed by detection withHRP-conjugated rabbit anti-mouse Ig (DAKO). ABTS(Sigma) was employed as a substrate for peroxidase.Absorbance (A405 nm) was measured with the use of aBenchmark Plus microplate spectrophotometer (Bio-Rad).
Ficolin-3 concentration in human seraIn general, the procedure followed that described in thesection Binding of lectins and immunoglobulins to variousligands. Briefly, the plates were coated with PS1–BSA dilutedin PBS. After overnight incubation at 4°C, plates were washedwith TBS-Ca2+/0.05% Tween-20 and blocked with 0.1%BSA/TBS-Ca2+. Tested sera were 200-fold diluted in 0.1%BSA/MBL-binding buffer and transferred in triplicate into thewells. Plates were incubated overnight at 4°C. Subsequentlyanti-human ficolin-3 Abs (0.5 µg/mL in 0.1% BSA/TBS-Ca2+)were added and after incubation for 2 h at 37°C followed by
detection with HRP-conjugated rabbit anti-mouse Ig. Humanserum A or rh ficolin-3 were used as standards.
C4b deposition assayFicolin-3-dependent lectin pathway activity was estimated in amodified C4 deposition assay [described for MBL byPetersen et al. (2001)]. Briefly, the microtiter plates werecoated with PS1–BSA conjugate (1 µg/well) and incubatedovernight at 4°C. The plates were washed with TBS-Ca2+/0.05% Tween-20 and blocked with 0.1% BSA/TBS-Ca2+.Sera to be tested (100-fold diluted in 0.1% BSA/MBL-binding buffer) were added and incubated overnight(4°C). Next, low ficolin-3 serum (B), prediluted 1:3000 in0.1% BSA/TBS-Ca2+, was added as a source of C4 factor.After incubation for 2 h at 37°C, any deposited C4b wasdetected with the use of anti-human C4 Ab and HRP-labeledgoat anti-rabbit Ig. The MBL-binding buffer was used in thisexperiment to prevent the activation of complement via theclassical pathway (Petersen et al. 2001).
Statistical analysisCorrelation was tested with Spearman’s test. P-values of<0.05 were considered as statistically significant.
Supplementary data
Supplementary data for this article is available online athttp://glycob.oxfordjournals.org/.
Funding
This work was supported by Polish Ministry of Sciences andHigher Education (N401 084 32/1944 and N401 267339).
Acknowledgements
Prof. Jens C Jensenius (Department of Medical Microbiologyand Immunology, University of Aarhus, Denmark) andDr. David C Kilpatrick (Scottish National Blood TransfusionService, National Science Laboratory, Edinburgh, Scotland)are kindly acknowledged for measurements of ficolin-3 andficolin-2 concentrations in sera A and B. We kindly thankDr. Piotr Jakimowicz from Department of Biotechnology atthe University of Wroclaw for MALDI-TOF MS measure-ments. We thank Dr. DC Kilpatrick for critically reading andhelpful discussion.
Conflict of interest
A.S., J.L., M.C, A.M., W.J., T.N., C.L. and Ludwik HirszfeldInstitute of Immunology and Experimental Therapy andInstitute for Medical Biology have a patent pending on theuse of identified ficolin-3 ligands as a tool for both scientificand diagnostic purposes. The authors have no additionalfinancial interests.
A Swierzko et al.
278
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Abbreviations
BSA, bovine serum albumin; BSA-NAc, N-acetylated BSA;COSY, correlation spectroscopy; DEPT, distortionlessenhancement by polarization transfer; ECL, electrochemicalluminescence; EDTA, ethylenediaminetetraacetic acid; EGTA,ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraaceticacid; FITC, fluorescein isothiocyanate; GVB, Gelatin VeronalBuffer; HEPES, N-(2-Hydroxyethyl)piperazine-N′-(2-ethane-sulfonic acid); HMBC, heteronuclear multiple-bond corre-lation; HRP, horseradish peroxidase; HSA, human serumalbumin; HSQC, heteronuclear single quantum coherence;IgG, immunoglobulin G; LOS, lipooligosaccharide; LPS, lipo-polysaccharide; MALDI-TOF, matrix-assisted laser deso-rption/ionization-time of flight; MASP, MBL-associated serineprotease; MBL, mannan-binding lectin; MS, mass spec-trometry; NOESY, nuclear Overhauser effect spectroscopy;OD, optical density; OS, oligosaccharide; PAMP, pathogen-associated molecular pattern; PCM, Polish Collection ofMicroorganisms; PRM, pattern-recognition molecule; PS,polysaccharide; rh, recombinant human; ROESY, rotating-frame Overhauser effect spectroscopy; RU, repeating unit;TBS, tris buffered saline; TL5A, tachylectin 5A; TOCSY,total correlation spectroscopy.
References
Andersen T, Munthe-Fog L, Garred P, Jacobsen S. 2009. Serum levels officolin-3 (Hakata antigen) in patients with systemic lupus erythematosus. JRheumatol. 36:757–759.
Baturo AP, Raginskaya VP. 1978. Antigenic scheme for Hafniae. Int J SystBacteriol. 28:126–127.
Birnbaum GI, Roy R, Brisson JR, Jennings HJ. 1987. Conformations ofammonium 3-deoxy-D-manno-2-octulosonate (KDO) and methyl α- andβ-ketopyranosides of KDO: X-ray structure and 1H NMR analyses. JCarbohydr Chem. 6:17–39.
Cedzynski M, Szemraj J, Swierzko AS, Bak-Romaniszyn L, Banasik M,Zeman K, Kilpatrick DC. 2004. Mannan-binding lectin insufficiency inchildren with recurrent infections of the respiratory system. Clin ExpImmunol. 136:304–311.
Clas F, Loos M. 1981. Antibody-independent binding of the first componentof complement (C1) and its subcomponent C1q to the S and R forms ofSalmonella minnesota. Infect Immun. 31:1138–1144.
Dag S, Niedziela T, Dzieciatkowska M, Lukasiewicz J, Jachymek W,Lugowski C, Kenne L. 2004. The O-acetylation patterns in the O-antigensof Hafnia alvei strains PCM 1200 and 1203, serologically closely relatedto PCM 1205. Carbohydr Res. 339:2521–2527.
Endo Y, Matsushita M, Fujita T. 2007. Role of ficolin in innate immunity andits molecular basis. Immunobiology. 212:371–379.
Endo Y, Matsushita M, Fujita T. 2011. The role of ficolins in the lectinpathway of innate immunity. Int J Biochem Cell Biol. 43:705–712.
Frederiksen PD, Thiel S, Larsen CB, Jensenius JC. 2005. M-ficolin, an innateimmune defence molecule, binds patterns of acetyl groups and activatescomplement. Scand J Immunol. 62:462–473.
Fox JD, Robyt JF. 1991. Miniaturization of three carbohydrate analyses usinga microsample plate reader. Anal Biochem. 195:93–96.
Fukutomi T, Ando B, Sakamoto S, Sakai H, Nawata H. 1996. Thermolabileβ-2 macroglycoprotein (Hakata antigen) in liver disease: Biochemical andimmunohistochemical study. Clin Chim Acta. 255:93–106.
Galanos C, Luderitz O, Rietschel ET, Westphal O. 1977. Newer aspects of thechemistry and the biology of bacterial lipopolysaccharides with specialreference to their lipid A component. In: Goodwin W, editor. Biochemistryof Lipids. 2nd ed. Baltimore: University Park Press. p. 239–335.
Garlatti V, Belloy N, Martin L, Lacroix M, Matsushita M, Endo Y, Fujita T,Fontecilla-Camps JC, Arlaud GJ, Thielens NM, et al. 2007. Structuralinsights into the innate immune recognition specificities of L- andH-ficolins. EMBO J. 26:623–633.
Garlatti V, Martin L, Lacroix M, Gout E, Arlaud GJ, Thielens NM,Gaboriaud C. 2009. Structural insights into the recognition properties ofhuman ficolins. J Innate Immun. 2:17–23.
Garred P, Honore C, Ma YJ, Rorvig S, Cowland J, Borregaard N,Hummelshoj T. 2010. The genetics of ficolins. J Innate Immun. 2:3–16.
Goddard TD, Kneller DG. 2001. SPARKY. 3rd ed. San Francisco: Universityof California.
Gout E, Garlatti V, Smith DF, Lacroix M, Dumestre-Perard C, Lunardi T,Martin L, Cesbron JY, Arlaud GJ, Gaboriaud C, et al. 2010. Carbohydraterecognition properties of human ficolins: Glycan array screening revealsthe sialic acid binding specificity of M-ficolin. J Biol Chem.285:6612–6622.
Hein E, Honore C, Skjoedt MO, Munthe-Fog L, Hummelshoj T, Garred P.2010. Functional analysis of ficolin-3 mediated complement activation.PLoS One. 5:e15443.
Hummelshoj T, Munthe-Fog L, Madsen HO, Sim RB, Garred P. 2008.Comparative study of the human ficolins reveals unique features officolin-3 (Hakata antigen). Mol Immunol. 45:1623–1632.
Inaba S, Okochi K, Yae Y, Niklasson F, de Verder CH. 1990. Serologicalstudies of an SLE-associated antigen-antibody system discovered as a pre-cipitation reaction in agarose gel: The HAKATA antigen-antibody system.Fukuoka Igaku Zasshi. 81:284–291.
Jachymek W, Petersson C, Helander A, Kenne L, Lugowski C, Niedziela T.1995. Structural studies of the O-specific chain and a core hexasaccharideof Hafnia alvei strain 1192 lipopolysaccharide. Carbohydr Res.269:125–138.
Janda JM, Abbott SL, Albert MJ. 1999. Prototypal diarrheagenic strains ofHafnia alvei are actually members of the genus Escherichia. J ClinMicrobiol. 37:2399–2401.
Jiang GZ, Sugiyama T, Kato Y, Koide N, Yokochi T. 1995. Binding ofmannose-binding protein to Klebsiella O3 lipopolysaccharide possessingthe mannose homopolysaccharide as the O-specific polysaccharide and itsrelation to complement activation. Infect Immun. 63:2537–2540.
Joiner KA, Grossman N, Schmetz M, Leive L. 1986. C3 binds preferentiallyto long-chain lipopolysaccharide during alternative pathway activation bySalmonella montevideo. J Immunol. 136:710–715.
Katzenellenbogen E, Kocharova NA, Zatonsky GV, Bogulska M, WitkowskaD, Shashkov AS, Knirel YA, Romanowska E. 1999. Structure of theO-specific polysaccharide of Hafnia alvei 23 having an oligosaccharide-phosphate repeating unit. J Carbohydr Chem. 18:545–558.
Katzenellenbogen E, Kocharova NA, Zatonsky GV, Shashkov AS, BogulskaM, Knirel YA. 2005. Structures of the biological repeating units in theO-chain polysaccharides of Hafnia alvei strains having a typical lipopoly-saccharide outer core region. FEMS Immunol Med Microbiol. 45:269–278.
Katzenellenbogen E, Romanowska E, Kocharova NA, Knirel YA, ShashkovAS, Kochetkov NK. 1992. The structure of a glycerol teichoic acid-likeO-specific polysaccharide of Hafnia alvei 1205. Carbohydr Res.231:249–260.
Kilpatrick DC, McLintock LA, Allan EK, Copland M, Fujita T, JordanidesNE, Koch C, Matsushita M, Shiraki H, Stewart K, et al. 2003. No strongrelationship between mannan binding lectin or plasma ficolins andchemotherapy-related infections. Clin Exp Immunol. 134:279–284.
Krarup A, Sorensen UB, Matsushita M, Jensenius JC, Thiel S. 2005. Effectof capsulation of opportunistic pathogenic bacteria on binding of thepattern recognition molecules mannan-binding lectin, L-ficolin, andH-ficolin. Infect Immun. 73:1052–1060.
Krarup A, Thiel S, Hansen A, Fujita T, Jensenius JC. 2004. L-ficolin is apattern recognition molecule specific for acetyl groups. J Biol Chem.279:47513–47519.
Lacroix M, Dumestre-Perard C, Schoehn G, Houen G, Cesbron JY, ArlaudGJ, Thielens NM. 2009. Residue Lys57 in the collagen-like region ofhuman L-ficolin and its counterpart Lys47 in H-ficolin play a key role inthe interaction with the mannan-binding lectin-associated serine proteasesand the collectin receptor calreticulin. J Immunol. 182:456–465.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature. 227:680–685.
Lonngren J, Goldstein I. 1978. Carbohydrate antigens coupling melibionicacid to bovine serum albumin using water-soluble carbodiimide. MethodsEnzymol. 50:160–162.
Lukasiewicz J, Niedziela T, Jachymek W, Kenne L, Lugowski C. 2009. TwoKdo-heptose regions identified in Hafnia alvei 32 lipopolysaccharide: Thecomplete core structure and serological screening of different Hafnia O ser-otypes. J Bacteriol. 191:533–544.
New human ficolin-3 ligands
279
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

MacDonald S, Kilpatrick D. 2007. Collagen-related defence proteins asanimal lectins. In: Kilpatrick D, editor. Collagen-Related Lectins in InnateImmunity. Kerala: Research Signpost. p. 1–16.
Matsushita M. 2007. The ficolin family: An overview. In: Kilpatrick D,editor. Collagen-Related Lectins in Innate Immunity. Kerala: ResearchSignpost. p. 17–32.
Matsushita M. 2009. Ficolins: Complement-activating lectins involved ininnate immunity. J Innate Immun. 2:24–32.
Munthe-Fog L, Hummelshoj T, Honore C, Madsen HO, Permin H, Garred P.2009. Immunodeficiency associated with FCN3 mutation and ficolin-3deficiency. N Engl J Med. 360:2637–2644.
Munthe-Fog L, Hummelshoj T, Ma YJ, Hansen BE, Koch C, Madsen HO,Skjodt K, Garred P. 2008. Characterization of a polymorphism in thecoding sequence of FCN3 resulting in a ficolin-3 (Hakata antigen)deficiency state. Mol Immunol. 45:2660–2666.
Ng PM, Le Saux A, Lee CM, Tan NS, Lu J, Thiel S, Ho B, Ding JL. 2007.C-reactive protein collaborates with plasma lectins to boost immuneresponse against bacteria. EMBO J. 26:3431–3440.
Petersen SV, Thiel S, Jensen L, Steffensen R, Jensenius JC. 2001. An assayfor the mannan-binding lectin pathway of complement activation. JImmunol Methods. 257:107–116.
Petersson C, Niedziela T, Jachymek W, Kenne L, Zarzecki P, Lugowski C.1997. Structural studies of the O-specific polysaccharide of Hafnia alveistrain PCM 1206 lipopolysaccharide containing D-allothreonine. Eur JBiochem. 244:580–586.
Raetz CR, Whitfield C. 2002. Lipopolysaccharide endotoxins. Annu RevBiochem. 71:635–700.
Romanowska E. 2000. Immunochemical aspects of Hafnia alvei O antigens.FEMS Immunol Med Microbiol. 27:219–225.
Schlapbach LJ, Aebi C, Hansen AG, Hirt A, Jensenius JC, Ammann RA.2009. H-ficolin serum concentration and susceptibility to fever and neutro-penia in paediatric cancer patients. Clin Exp Immunol. 157:83–89.
Schlapbach LJ, Thiel S, Kessler U, Ammann RA, Aebi C, Jensenius JC.2011. Congenital H-ficolin deficiency in premature infants with severenecrotising enterocolitis. Gut. 60:1438–1439.
Sugimoto R, Yae Y, Akaiwa M, Kitajima S, Shibata Y, Sato H, Hirata J,Okochi K, Izuhara K, Hamasaki N. 1998. Cloning and characterization ofthe Hakata antigen, a member of the ficolin/opsonin p35 lectin family. JBiol Chem. 273:20721–20727.
Svendsen CB, Hummelshoj T, Munthe-Fog L, Milman N, Garred P, LaursenIA, Christiansen M, Krogfelt KA. 2008. Ficolins and mannose-
binding lectin in Danish patients with sarcoidosis. Respir Med.102:1237–1242.
Swierzko AS, Cedzynski M, Kirikae T, Nakano M, Klink M, Kirikae F,Ziolkowski A, Vinogradov EV, Kawakami M. 2003. Role of thecomplement-lectin pathway in anaphylactoid reaction induced with lipopo-lysaccharide in mice. Eur J Immunol. 33:2842–2852.
Thiel S. 2007. Complement activating soluble pattern recognition moleculeswith collagen-like regions, mannan-binding lectin, ficolins and associatedproteins. Mol Immunol. 44:3875–3888.
Thomsen T, Schlosser A, Holmskov U, Sorensen GL. 2011. Ficolins andFIBCD1: Soluble and membrane bound pattern recognition molecules withacetyl group selectivity. Mol Immunol. 48:369–381.
Tsujimura M, Ishida C, Sagara Y, Miyazaki T, Murakami K, Shiraki H,Okochi K, Maeda Y. 2001. Detection of serum thermolabile beta-2 macro-glycoprotein (Hakata antigen) by enzyme-linked immunosorbent assayusing polysaccharide produced by Aerococcus viridans. Clin Diagn LabImmunol. 8:454–459.
Tsujimura M, Miyazaki T, Kojima E, Sagara Y, Shiraki H, Okochi K, MaedaY. 2002. Serum concentration of Hakata antigen, a member of the ficolins,is linked with inhibition of Aerococcus viridans growth. Clin Chim Acta.325:139–146.
Vinogradov E, Bock K. 1999. The structure of the core part of Proteus mir-abilis O27 lipopolysaccharide with a new type of glicosidic linkage.Carbohydr Res. 319:92–101.
Wang CC, Yim KW, Poon TC, Choy KW, Chu CY, Lui WT, Lau TK, RogersMS, Leung TN. 2007. Innate immune response by ficolin binding in apop-totic placenta is associated with the clinical syndrome of preeclampsia.Clin Chem. 53:42–52.
Westphal O, Jann K. 1965. Bacterial lipopolysaccharides: Extraction withphenol-water and further applications of the procedure. MethodsCarbohydr Chem. 5:83–89.
Yae Y, Inaba S, Sato H, Okochi K, Tokunaga F, Iwanaga S. 1991. Isolationand characterization of a thermolabile beta-2 macroglycoprotein (‘thermola-bile substance’ or ‘Hakata antigen’) detected by precipitating (auto) anti-body in sera of patients with systemic lupus erythematosus. BiochimBiophys Acta. 1078:369–376.
Zhao L, Ohtaki Y, Yamaguchi K, Matsushita M, Fujita T, Yokochi T,Takada H, Endo Y. 2002. LPS-induced platelet response and rapidshock in mice: Contribution of O-antigen region of LPS and involve-ment of the lectin pathway of the complement system. Blood.100:3233–3239.
A Swierzko et al.
280
by guest on July 6, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















