
Consequences of unilateral nigrostriatal denervation on the thalamostriatal pathway in rats
Upload
independentCategory
view
6download
0
www.elsevier.com/locate/yexnr
Experimental Neurology 189 (2004) 78–93
Sparing of behavior and basal extracellular dopamine after
6-hydroxydopamine lesions of the nigrostriatal pathway
in rats exposed to a prelesion sensitizing
regimen of amphetamine
Isabella Anna Moroz,a,1 Susana Pecina,b,1 Timothy Schallert,c,d and Jane Stewarta,*
aCenter for Studies in Behavioral Neurobiology, Concordia University, Montreal, PQ, CanadabDepartment of Psychology, University of Michigan, Ann Arbor, MI 48109, USA
cDepartment of Psychology and Institute for Neuroscience, University of Texas, Austin, TX 78712, USAdDepartment of Neurosurgery, University of Michigan, Ann Arbor, MI 48109, USA
Received 6 December 2003; revised 1 May 2004; accepted 7 May 2004
Abstract
Repeated administration of amphetamine leads to enduring augmentation of its behavioral-activating effects, enhanced dopamine (DA)
release in striatal regions, and morphological changes in DA target neurons. Here we show that exposure to a 2-week escalating-dose regimen
of amphetamine prevents behavioral asymmetries of forelimb use and spontaneous (drug-independent) turning behavior following unilateral
6-hydroxydopamine (6-OHDA) lesions of the nigrostriatal pathway made 7–14 days after termination of amphetamine treatment
(Experiments 1–3). Exposure to three nonescalating injections of amphetamine 7 days before 6-OHDA lesions had no effect (Experiment 2).
Prelesion amphetamine treatment led to normalization of basal extracellular levels of striatal DA as measured by microdialysis on days 11–
14 and 25–28 after lesioning (Experiment 3). However, there were no significant differences between treatment groups in postmortem tissue
levels of DA and its metabolites, indicating a dissociation between the DA depletion and the extracellular levels of DA as measured by
microdialysis. Finally, rats exposed to the escalating amphetamine regimen had reduced lesion-induced loss of TH-IR cells in the ipsilateral
DA cell body regions (Experiment 3). Thus, prelesion exposure to the escalating doses of amphetamine may render the cells resistant to the
consequences of damage after subsequent 6-OHDA lesions, possibly by accelerating the development of compensatory changes in the DA
neurons that typically accompany behavioral recovery. The potential role of amphetamine-induced endogenous neurotrophic factors in the
behavioral sparing and normalization of basal extracellular DA levels observed after subsequent 6-OHDA lesions is discussed.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Amphetamine; Prelesion; Behavioral sparing; Striatal dopamine; 6-OHDA; Nigrostriatal pathway; Neurotrophic factors; Exercise
Introduction activating effects that are associated with heightened
Exposure to psychostimulant drugs leads to changes in
brain and behavior that outlast the drugs’ acute neurophar-
macological effects. Repeated administration of amphet-
amine results in an enduring enhancement of its behavioral-
0014-4886/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.expneurol.2004.05.012
* Corresponding author. Center for Studies in Behavioral Neurobiology,
Department of Psychology, Concordia University, 7141 Sherbrooke Street.
W., Montreal, Quebec, Canada H4B 1R6. Fax: +1-514-848-2817.
E-mail address: [email protected] (J. Stewart).1 These authors contributed equally to these independent experiments.
Isabella Anna Moroz is currently a postdoctoral fellow at the Centre for
Stroke Recovery, Ontario Health Research Institute, University of Ottawa,
Ottawa, Ontario, Canada.
dopamine (DA) overflow in striatal regions in response
to acute drug challenges (Kalivas and Stewart, 1991;
Robinson and Becker, 1986). This phenomenon of ‘‘be-
havioral sensitization’’ is very different from ‘‘amphetamine
neurotoxicity’’ that develops after continuous infusion or
frequent injection of multiple high doses of amphetamine
and leads to depletion of striatal DA presumably due to
terminal degeneration (for a review, see Robinson and
Becker, 1986). The behavioral and neurochemical effects
of repeated administration of amphetamine develop gradu-
ally and have been observed for months after the termination
of drug treatment (Paulson et al., 1991). Enduring enhance-
ment of function within the midbrain DA system has also

Table 1
Two-week escalating-dose treatment with amphetamine
Monday Tuesday Wednesday Thursday Friday Saturday Sunday
Week
1
1 1 2 2 2 – –
Week
2
3 4 4 4 4 – –
The numbers represent mg/kg of d-amphetamine sulfate per injection on
each day. All animals received two injections per day.
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 79
been observed after partial unilateral 6-hydroxydopamine (6-
OHDA) lesions of the nigrostriatal neurons. In these models,
basal extracellular levels of dopamine released from the
surviving terminals in the lesioned striatum, as measured
using in vivo microdialysis, normalize gradually, corres-
ponding in time to behavioral recovery from such lesions
(Abercrombie et al., 1990; Altar et al., 1994; Castaneda et al.,
1990; Robinson and Whishaw, 1988; Robinson et al., 1994).
The gradual and long-lasting nature of behavioral sensitiza-
tion and recovery from partial lesions has been proposed to
occur as a result of neurochemical (Robinson and Becker,
1986; Robinson et al., 1994) and structural modifications
(Finkelstein et al., 2000; Robinson and Kolb, 1997) in neural
circuitry, and alterations in patterns of synaptic connectivity,
most probably brought about by the actions of neurotrophic
factors (Flores and Stewart, 2000a; Moroz et al., 2003). We
hypothesized that repeated administration of amphetamine
before 6-OHDA lesions of the nigrostriatal DA system may
mobilize neurotrophic factors and other precursors to struc-
tural adaptations at the time of lesioning that could, in turn,
enhance DA neuron functioning and thus reduce the impact
of the lesions on behavior, that is, facilitate behavioral
sparing after such lesions.
In intact rats, repeated treatment with amphetamine has
been shown to increase dendritic length, density of dendritic
spines, and the number of branched spines on the major DA
output neurons in the nucleus accumbens (NAcc), striatum,
and prefrontal cortex (Li et al., 2003; Robinson and Kolb,
1997, 1999). Furthermore, repeated administration of am-
phetamine has also been shown to induce increases in
expression of neurotrophic factors known to affect the
survival, maintenance, and morphological plasticity of adult
neurons. Treatments with amphetamine have resulted in
increases in astrocytic basic fibroblast growth factor (FGF-
2) in the DA cell body regions, ventral tegmental area (VTA)
and substantia nigra pars compacta (SNc; Flores et al., 1998),
and DA striatal terminal regions (Flores and Stewart, 2000b),
as well as increases in brain-derived neurotrophic growth
factor (BDNF) in the basolateral amygdala and its target
regions including medial NAcc and dorsal medial striatum
(Meredith et al., 2002). More importantly, the number of
FGF-2-immunoreactive astrocytes in the VTA and SNc has
been strongly and positively correlated with the magnitude
of behavioral sensitization, and infusions of an FGF-2-
neutralizing antibody into the VTA before amphetamine
administration have been shown to prevent the development
of sensitization (Flores et al., 2000). Thus, the long-lasting
behavioral and neurochemical changes induced by repeated
administration of amphetamine could be mediated through
the actions of neurotrophic factors such as FGF-2 and BDNF.
In view of findings that increases in neurotrophic factor
expression before brain injury have the potential to protect
against subsequent insults (Altar et al., 1994; Choi-Lund-
berg et al., 1998; Mandel et al., 1997; Shults et al., 1996,
2000), we hypothesized that exposure to amphetamine
before 6-OHDA lesions might serve to attenuate behavioral
and neurochemical deficits after these lesions. We report
here two independent experiments, conducted in separate
laboratories, using different strains of rats and different
placements of 6-OHDA infusions within the nigrostriatal
pathway, both showing that exposure to an escalating-dose
regimen of amphetamine prevents subsequent 6-OHDA
lesion-induced asymmetries of forelimb use and turning
behavior. In addition, we report that this behavioral sparing
coincides with normalization of basal extracellular DA
levels in the striatum as assessed using in vivo microdialysis.
Materials and methods
Experiment 1
Subjects
Adult male Long–Evans rats (Charles River, Wilming-
ton, MA; 250–300 g), housed on a reversed light–dark
cycle with free access to food and water served as subjects.
Amphetamine regimen
d-Amphetamine sulfate (Sigma, St. Louis, MO), dis-
solved in physiological saline, was given in a 2-week
escalating-dose regimen (esc/amph; n = 10), as described
previously (Flores and Stewart, 2000b). This regimen in-
volved two daily intraperitoneal (ip) injections of amphet-
amine in the colony room, 7–8 h apart, 5 days a week, for 2
weeks. The dose of amphetamine began with 1 mg/kg and
escalated to 4 mg/kg for the last 4 days of treatment (see
Table 1). The saline-treated control group (sal; n = 12)
received 0.9% saline (1 ml/kg). To control for amphetamine-
induced increases in motor activity, four rats from the sal
group were housed 24 h/day in cages with running wheels
(34 cm diameter) during the amphetamine administration
period (Robinson and Kolb, 1999).
Intrastriatal 6-OHDA lesions
Unilateral 6-OHDA (6-hydroxydopamine hydrochloride,
Sigma) infusions (4 Al of 10 Ag/4 Al in 0.05% ascorbic acid
solution; 0.5 Al/min) into the striatum (stereotaxic coordi-
nates with flat skull: 1.7 mm anterior, 2.9 mm lateral to
bregma, and 4.0 mm ventral to skull) were performed under
Equithesin (25 mg/kg pentobarbital; Sigma) and 150 mg/kg
chloral hydrate anesthesia (0.35 ml/100 g ip) followed by
atropine sulfate (0.1 mg/kg ip; Sigma).

tal Neurology 189 (2004) 78–93
Behavioral testing and limb-use observation
Use of each forelimb for upright support and for landing
when descending from a rearing position was analyzed both
pre- and postoperatively in the cylinder test, as previously
described (Tillerson et al., 2001). Occurrences of forelimb
use for wall exploration and landing were determined sep-
arately, and each was expressed in terms of (1) the percent
ipsilateral limb use [ipsi / (ipsi + contra + both) � 100] and
(2) the percent contralateral limb use [contra / (ipsi + contra +
both) � 100]. The percent contralateral limb use was then
subtracted from the percent ipsilateral limb use for both the
wall behavior and landing. These two scores (wall and
landing) were averaged to obtain a single overall limb-use
asymmetry score that corrected for variability in the absolute
number of landing movements versus wall movements
among animals or between groups. This asymmetry score
has been shown previously to be highly correlated with the
degree of 6-OHDA-induced striatal DA depletion (Schallert
and Tillerson, 1999).
Designs and procedures
The timing of the experimental manipulations in Exper-
iment 1 is outlined in Fig. 1A. Briefly, 2 weeks after
termination of the 2-week escalating-dose regimen of am-
phetamine, rats received unilateral intrastriatal 6-OHDA
I.A. Moroz et al. / Experimen80
Fig. 1. Diagram outlining the timing of the experimental manipulations in
Experiments 1, 2, and 3.
infusions. Asymmetry of forelimb use was tested in the
cylinder test before 6-OHDA lesioning (pretest) and 1, 3, 7,
and 14 days after lesioning.
Statistical analyses
Data were analyzed by mixed-factor analysis of variance
(ANOVA) with drug (sal, esc/amph) as the between factor
and day (pretest, d1, d3, d7, d14) as the repeated factor.
Experiments 2 and 3
Subjects
Adult male Wistar rats (Charles River, QC; 325–350 g),
housed on a reversed light–dark cycle with free access to
food and water served as subjects.
Amphetamine regimens
d-Amphetamine sulfate (SmithKline Beecham Pharma,
Oakville, ON), dissolved in physiological saline, was given
either in a 2-week escalating-dose regimen (esc/amph, see
Table 1; Experiment 2, n = 5; Experiment 3, n = 7), or once
a day, every second day, for a total of three injections (3
amph; 3 mg/kg ip, n = 5). Saline-treated animals for each
amphetamine regimen (sal, Experiment 2, n = 4/group;
Experiment 3, n = 5) received 0.9% saline (1.0 ml/kg).
Intra-MFB 6-OHDA lesions
Unilateral 6-OHDA (6-hydroxydopamine hydrochloride,
Sigma, Oakville, ON) infusions (2 Al of 8 Ag/4 Al of 0.9%saline containing 0.05% ascorbic acid solution; rate: 0.2 Al/min) into the medial forebrain bundle (MFB; stereotaxic
coordinates with flat skull: 2.9 mm posterior, 1.7 mm
lateral to bregma, and 7.6 mm ventral to dura) were
performed under sodium pentobarbital anesthesia (30 mg/
kg ip) supplemented with isoflurane (Biomeda MTC, Cam-
bridge, ON). Atropine sulfate (0.5 mg/ml, 0.2 ml/rat sc)
was given to reduce bronchial secretions. Desmethyli-
mipramine (15 mg/kg ip; RBI Biochemicals, Oakville,
ON), a norepinephrine reuptake inhibitor, was given 30
min before infusion of 6-OHDA to protect the noradre-
nergic cells from the neurotoxin.
In Experiment 3, after 6-OHDA infusion, guide cannulae
(22-gauge stainless steel) were implanted bilaterally into the
striatum (stereotaxic arms angled 10j from the vertical
plane, stereotaxic coordinates with flat skull: 1.2 mm
anterior, 3.0 mm lateral to bregma, and 3.4 mm ventral to
skull surface). The cannulae were anchored to the skull with
stainless steel screws and secured to the surface with dental
cement.
Behavioral testing and limb-use observation
Forelimb use for upright support and for landing when
descending from a rearing position was analyzed pre- and
postoperatively in the cylinder test, as described in Exper-
iment 1. Rats were also tested for spontaneous ipsiversive
turning (turning toward the side of the lesion) in a novel

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 81
environment, as previously described (Moroz et al., 2003).
The number of compact (within the diameter of approxi-
mately 20 cm) 360j turns and 180j half turns ipsilateral and
contralateral to the side of the lesion was recorded and
summed across 5-min testing sessions. The number of
ipsilateral turns was presented as a percent of the total
number of turns displayed by an animal [ipsi / (ipsi +
contra) � 100%].
Microdialysis
Microdialysis was conducted as described previously
(Emmi et al., 1996, 1997) in hexagonal testing chambers
(42 � 39 � 33.5 cm) built from Plexiglas with wooden
ceilings and stainless steel rod floors. The cages were
individually enclosed in wooden cubicles where lighting
was provided on a reversed cycle by overhead light bulbs
(15 W). The dialysis probe consisted of a 2.2-mm length of
semipermeable dialysis membrane (Fisher Scientific, 240
Am OD, 13000 M.W. cutoff), closed at one end and attached
to a 21-mm-long, 26-gauge stainless steel length of tubing.
A 40- to 50-cm length of PE-20 tubing connected the other
end of the stainless steel shaft to an infusion swivel
stationed above the testing chamber, which was connected
via PE-20 tubing to a variable speed infusion pump. A small
diameter, fused silica tube extended internally through the
probe with one end resting 0.5 mm from the tip of the probe
and the other end exiting the PE tubing 35 cm below the
infusion swivel. The probes were secured by brass collars
that screwed onto the guide cannulae. The external length of
PE-20 tubing was protected from damage by steel spring
casings. The probes were designed so that the entire length
of semipermeable membrane extended below the guide
cannula tip.
The probes were inserted on the evening preceding the
microdialysis testing. To prevent occlusion, artificial cereb-
rospinal fluid (ACSF, 145 mM Na+, 2.7 mM K+, 1.22 mM
Ca2+, 1.0 mM Mg2+, 150 mM Cl�, 0.2 mM ascorbate, 2
mM Na2HPO4, pH, 7.4) was perfused overnight at a rate
of 0.25 Al/min. Dialysate sampling and activity monitoring
began the next morning. Half of the animals from each
treatment condition were dialyzed on the lesioned side on
the first day of dialysis and on the intact side on the
second day of dialysis; for the other animals, the con-
ditions were reversed. The dialysate flow rate was in-
creased to 0.6 Al/min, and baseline dialysate samples
(approximately 12 Al/sample) were collected every 20
min. A 10-Al volume of dialysate was extracted from each
sample and immediately analyzed using one of two similar
high-performance liquid chromatography systems with
electrochemical detection (HPLC-EC). The samples were
loaded onto reverse-phase columns (15 � 4.6 mm; Hasil
C18, 5 Am; S.P.E. Limited, Concord, ON, Canada) through
manual injection ports (Rheodyne 7125; 20 Al loop);
reduction and oxidation currents for DA and its metabo-
lites, dihydroxyphenylacetic acid (DOPAC) and homova-
nillic acid (HVA), were measured with dual-channel ESA
coulometric detectors (Coulochem 5100, with a model
5011 analytical cell). The currents for DA were measured
independent of those for DOPAC and HVA using separate
channels of the Coulochem detectors. The mobile phases
[15% acetonitrile, 0.076 M SDS, 0.1 M ethylenediamine-
tetraacetic acid (EDTA), 0.058 M NaPO4, 0.27 M citric
acid, pH = 3.35] were circulated through each closed
system at a flow rate of 1.3 ml/min by Waters 515 HPLC
pumps. The peaks obtained for DA, DOPAC, and HVA
were integrated and quantified by EZChrom Chromatog-
raphy Data System (S.P.E. Limited). Dialysate samples
from individual rats always were analyzed with the same
HPLC-EC system, and the assignment of animals to each
system was counterbalanced across all treatment groups.
Food was removed from the chambers before sampling,
but a water drinking tube was available.
Postmortem tissue analysis
Animals were killed by decapitation and their brains
were rapidly removed, placed in isopentane, cooled on dry
ice, and frozen overnight at �80jC. The following day, the
brains were sliced on a cryostat into 200 Am sections.
Punches were taken from the dorsal striatum (1 � 2 mm
diameter) of the lesioned and nonlesioned hemisphere, from
two sections about 1.2 and 1.5 mm anterior to bregma, and
from the SN (2 � 1 mm diameter) of the lesioned and
nonlesioned hemisphere, from two sections about 5.2 and
5.5 posterior to bregma. The neurochemical assessment of
DA, DOPAC, and HVA was then performed as described
previously (Moroz et al., 2003). Punches were suspended in
phosphate buffer (PB) and frozen overnight. The following
day, samples were thawed and centrifuged at 4000 rpm for
15 min. Pellets were suspended in 0.1 M NaCl and analyzed
for protein content. The supernatant was removed and
assayed for DA, DOPAC, and HVA using HPLC-EC. The
supernatant was injected into a 15-cm C18 column (5 mm
particle size, Scientific Products and Equipment, Ontario).
The mobile phase consisted of 30 mM citric acid, 60 mM
sodium phosphate monobasic, 0.10 mM EDTA, 14% ace-
tonitrile, and 0.08 mM sodium dodecyl sulphate, pH 3.35.
The mobile phase was pumped through the system at 1.2 ml/
min using a Waters 515 HPLC pump. Compounds were
detected and quantified with an ESA coulochem detector
(model 5100A) equipped with an analytical cell (model
5011; E1 = +0.35 V, E2 = �0.3 V, ESA, Inc.). The
concentrations were estimated from peak heights by com-
parison with injections of known amounts of pure standards
(Sigma) and expressed as micrograms per milligram of
protein.
TH immunohistochemistry
Brains were removed and stored overnight in the
fixative solution [paraformaldehyde (w/v) and 15% picric
acid (v/v) in PB (pH 6.9)] at 4jC. Coronal 50 Am sections
were cut on a vibratome and stored overnight in PB at
4jC. Before slicing, a small mark was cut in each brain to

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9382
allow discrimination of the hemispheres in each section.
Sections were then processed for TH immunoreactivity, as
described previously (Moroz et al., 2003). Free-floating
tissue sections were preincubated in 0.3% Triton X-100 PB
and 1% normal goat serum (NGS) for 1 h at room
temperature. They were then incubated for 24 h at 4jCwith the rabbit anti-TH polyclonal antibody diluted to
1:5000 (Chemicon) in PB and 1% NGS. After incubation
in the primary antibody, sections were rinsed three times
for 5 min in cold PB and incubated for 1 h at room
temperature (RT) in a solution of rat adsorbed biotinylated
anti-rabbit antibody (Vector) diluted to 1:200 with PB and
1% NGS. After three 5-min washes in cold PB, sections
were incubated in an avidin–horseradish peroxidase com-
plex (Vectastain Elite ABC Kit, Vector, Burlington Canada)
for 30 min at RT and rinsed again three times for 5-min in
cold PB. Next, sections were incubated for 10 min at room
temperature and under constant agitation in a solution of
0.05% 3,3V-diaminobenzidine (Sigma) in PB. Then, with-
out washing, the sections were transferred to a 2,3V-diaminobenzidine-PB solution, pH 7.8, with 0.01% H2O2
to catalyze the reaction. To obtain an orange-brown reac-
tion product, NiCl2 was withheld from the 3,3V-diamino-
benzidine-PB-H2O2 solution. This incubation was
terminated 5 min later by washing the sections three times
for 10 min in cold PB. Tissue from all groups was included
in each batch of immunocytochemical processing. Sections
were then mounted on gelatin-coated slides, dried for at
least 24 h, hydrated in distilled water (1 min), and
gradually dehydrated in 70%, 95%, and 100% ethanol.
Slides were then cleared in Hemo-De and cover slipped
with Permount (Fisher Scientific).
Image analysis
Immunostained sections were analyzed under a Leica
microscope (Leitz DMRB). The number of TH-immunore-
active (IR) cells per squared millimeter was estimated from
digitized images of sample areas within SNc and VTA using
a computerized image analysis system (NIH Image 1.6).
Structure boundaries were defined according to the Paxinos
and Watson (1997) stereotaxic atlas. TH-IR cells were
manually counted in a minimum of eight images taken from
SNc and four from VTA at two different levels from
bregma: �5.2 and�5.3 mm, in each hemisphere. Only
sections in which the medial and lateral parts of the SNc
were clearly separated by the medial terminal nucleus of the
accessory optic tract were selected. The number of TH-IR
cells calculated from these levels from bregma has been
previously shown to accurately represent 6-OHDA-induced
degeneration of the SNc and VTA neurons (Carman et al.,
1991; Gordon et al., 1997). No attempt was made to
estimate the total TH-IR neurons number in three dimen-
sions. The images were assigned code names, and the FGF-
2 and TH-IR cells were counted by an individual blind to
the code assignment. The cell counts from the areas sampled
in each hemisphere of each brain region were summed and
divided by the total area examined.
Design and procedures
The timing of the treatments and experimental mani-
pulations in Experiments 2 and 3 is outlined in Figs. 1B
and C.
Experiment 2
Seven days after the last amphetamine injection of each
dose regimen, rats received unilateral 6-OHDA infusions
into the MFB. An additional group of rats given compa-
rable saline injections before the 6-OHDA lesions received
postlesion exercise treatment (exercise, n = 8) to compare
efficacy of amphetamine pretreatment to the well-estab-
lished beneficial effects of postinjury exercise intervention
(Tillerson et al., 2001, 2002, 2003). Postlesion exercise
consisted of walking or running on the floor of the
laboratory while inside commercially available transparent,
ventilated, shatter-resistant plastic balls (30 cm diameter)
that permitted a 360j range of motion (Jumbo Kritter
Krawler, Lee’s Aquarium and Pet Products, San Marcos,
CA). If rats initially did not move voluntarily when placed
inside the balls, the experimenter moved the ball in
random directions. For those rats that moved voluntarily,
the dimensions of the space available for exercise sessions
(about 4 m2) did not allow for the maintenance of higher
velocities over longer periods of time. Furthermore, the
speed of movement of each animal varied within each
exercise session. Typically, the rats were more energetic at
the beginning of the session, thus spending most time
running, after which they would switch to walking or even
stop moving. In the latter event, the experimenter would
immediately move the ball to ensure constant movement of
each animal throughout the 30-min sessions. Care was
taken to reproduce the various speeds and patterns of
movement displayed by rats that ran voluntarily. In addi-
tion, side-to-side and front-to-back rocking motions of the
ball were used on all rats intermittently throughout each
exercise session in an attempt to impose balance chal-
lenges and thus serving to strengthen the muscles of the
limbs. The exercise sessions began 4–5 h after lesioning
and continued for the next 14 days. The first session lasted
for 15 min, the next 7 for 30 min, and the final 7 for 15
min/day. Rats were tested once a week for forelimb use in
the cylinder test and for turning behavior in the novel
environment, starting 7 days after lesioning. The preoper-
ative test for asymmetry of forelimb use in the cylinder test
was conducted before the initiation of prelesion amphet-
amine treatments. Rats were killed and their brains were
taken for DA, DOPAC, and HVA tissue assays 28 days
after lesioning.
Experiment 3
Seven days after the last amphetamine or saline injection,
rats received unilateral 6-OHDA infusions into the MFB

Fig. 2. Experiment 1: Asymmetry of limb-use (mean F SEM) in the
cylinder test assessed 1, 3, 7, and 14 days after intrastriatal 6-OHDA lesions
was prevented by the esc/amph treatment [ F(1,109) = 13.51, P < 0.01].
The saline-treated rats housed in cages with free access to running wheels
(data not shown) did not differ from the saline-treated rats that did not run
[F(1,59) = 0.17, ns]; thus, these rats were combined to form the sal group.
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 83
followed immediately by bilateral intrastriatal guide cannulae
implantation.Microdialysis was performed at two time points
after lesioning—between 11 and 14 days and again between
25 and 28 days. Each rat was tested over 2 days at each time
point (either on days 11 and 12 and subsequently on days 25
and 26, or on days 13 and 14 and subsequently on days 27–
28, as determined by the number of microdialysis boxes
available). On the evening preceding each microdialysis
session, rats were moved to the microdialysis chambers
where the probes were inserted and perfused overnight at a
slow rate of < 0.25 Al/min. Dialysate sampling began the
following morning when half of the animals from each
treatment condition were dialyzed on either the lesioned or
the intact side. The same evening, a probe was inserted into
the other striata, and dialysate sampling of the opposite side
began the next morning. Half of the animals from each
treatment condition were dialyzed on days 11 and 12, and
half on days 13 and 14; similarly for the later time point after
lesioning, half of the animals from each treatment was
dialyzed on days 25 and 26, and 27 and 28 at the later time
point. The day after the microdialysis for both hemispheres
was completed, the rats were administered behavioral tests.
Following the last behavioral test conducted after micro-
dialysis at 28 days postlesion, the rats were killed and their
brains were removed and cut into anterior and posterior
portions. The anterior portion was taken for tissue analysis
of DA and its metabolites; sections from the posterior portion
were processed for TH immunoreactivity.
Statistical analyses
Behavioral data in Experiment 2 were analyzed by
mixed-factor ANOVAs with group (sal, esc/amph, 3 amph,
exercise) as the between factor and day (d7, d14, d21, and
d28) as the within factor. Pretest data for the forelimb
asymmetry test were analyzed with one-way ANOVA.
Tissue levels of DA, DOPAC, and HVA in Experiment 2
were analyzed by mixed-factor ANOVAs with group (sal,
esc/amph, 3 inj/amph, exercise) as the between factor and
side [nonlesioned (NL), lesioned (L)] as the within factor.
Behavioral data in Experiment 3 were analyzed using one-
way ANOVAs as required; post hoc comparisons were
made using the Fisher’s PLSC test (P < 0.05). Data from
two animals (one from the sal group and one from the esc/
amph group) were excluded from all analyses in Experi-
ment 3 due to lack of lesion as indicated by postmortem
analysis of tissue levels of DA and its metabolites. Rats
that performed fewer than five landings and fewer than 10
wall movements during the cylinder tests (criterion less
stringent than the typical 10 landings and 20 wall move-
ments criterion recommended by Schallert and Tillerson,
1999, employed here to compensate for general lower
levels of motor activity in Wistar than in Long–Evans
rats) and those that did not display tight turns of less than
30 cm in diameter (Fornaguera et al., 1994) during the
asymmetry of turning tests were not included in the
analyses. This resulted in slightly different degrees of
freedom for the F values on the two behavioral tests
conducted at two different time points after lesioning in
Experiment 3. Since none of these animals was inactive
during both behavioral tests, thus providing a measure of
their behavioral performance on at least one of the tests,
they were all included in the subsequent neurochemical
and cell counts analyses. Neurochemical analyses of tissue
and extracellular levels of DA, DOPAC, and HVA and
estimated numbers of TH-IR cells per squared millimeter
for each brain region in Experiment 3 were conducted
using mixed-factor ANOVAs with drug (sal, esc/amph) as
the between factor and side [nonlesioned (NL), lesioned
(L)] as the within factor. Tests for simple main effects were
used to determine the source of the significant Group �Side interactions; Bonferroni correction was used to keep
the overall error rate at 0.05.
Results
Behavior
Exposure to the 2-week escalating-dose regimen of
amphetamine before intrastriatal (Experiment 1) or intra-
MFB infusions of 6-OHDA (Experiments 2 and 3) pre-
vented the preferential use of the ipsilateral forelimb for
vertical exploration (Figs. 2–4A) and ipsilateral turning
(Figs. 3B and 4B) after 6-OHDA lesions. Remarkably, the
esc/amph groups from all three experiments displayed no
asymmetry of limb use or turning when compared to the sal
group (Figs. 2–4). In addition, in Experiment 2, the esc/
amph group, but not the 3 amph group, did not differ from
the postlesion exercise group (Fig. 3). In Experiment 1, rats

Fig. 3. Experiment 2: Mean (FSEM) asymmetry of (A) limb use and (B) spontaneous turning tested 7, 14, 21, and 28 days after 6-OHDA lesions of the MFB.
ANOVAs revealed a significant main effect of group for the limb-use asymmetry test [F(3,54) = 3.1, P < 0.05] and a similar tendency for the turning asymmetry
test [F(3,54) = 2.50, P = 0.09]. The esc/amph and the exercise groups displayed no asymmetry of limb use and did not differ from each other; both differed
significantly from the sal group (*P < .05). There were no differences between the groups in forelimb asymmetry during the pretest [F(3,18) = 1.00, ns]. Note in
(B), a dashed line at 50% represents no asymmetry or an equal number of ipsilateral and contralateral turns.
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9384
housed in cages with free access to running wheels before
lesioning showed high levels of motor activity, running an
average of 9.4 km/24 h, but their postlesion forelimb use
asymmetry was not different from that of the sal group.
In vivo microdialysis
Dopamine (DA)
Basal levels of DA from the intact and lesioned striata
of animals treated with the escalating-dose regimen of
amphetamine before intra-MFB infusions of 6-OHDA
(Experiment 3) and then tested using microdialysis be-
tween days 11–14 and 25–28 are shown in Figs. 5A and
D, respectively. It can be seen at both time points that
the esc/amph group showed normalization of basal DA
levels on the lesioned side, whereas the sal group had
significantly lower basal levels of DA in the striatum on
the lesioned side than on the intact side. This effect is
reflected in a significant Group � Side interaction ob-
served 11–14 days after lesioning.
DOPAC and HVA
Basal levels of both DOPAC and HVA measured on
days 11–14 and 25–28 after lesioning (Experiment 3) are
shown in Figs. 5B, C, E, and F, respectively. The
ANOVAs revealed a significant effect of side, but no
Group � Side interaction. It can be seen that levels of
both metabolites were significantly lower on the lesioned
side in both amph/esc and sal groups. See Fig. 5 legend for
results of statistical analyses.

Fig. 4. Experiment 3: Mean (F SEM) asymmetry of (A) limb use and (B) spontaneous turning, tested 11–14 (Test 1) and 25–28 (Test 2) days after 6-OHDA
lesions of the MFB. ANOVAs on the data revealed significant group effects [limb-use asymmetry, Test 2, F(1,8) = 6.05, P < 0.05; turning, Test 1, F(1,9) =
8.13, P < 0.05, Test 2, F(1,10) = 10.93, P < 0.05]. The esc/amph group displayed no asymmetry of limb use on Test 2 or turning on either Test 1 or 2. Note in
ipsilateral and contralateral turns.
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 85
Postmortem tissue analysis
The tissue levels of DA, DOPAC, and HVA in the dorsal
striatum and in SN observed 28 days after lesioning are
shown in Figs. 6(Experiment 2) and 7(Experiment 3). In all
groups, the postmortem levels of DA and its metabolites were
significantly depleted on the side of the lesion, indicating
extensive degeneration of terminals and cell bodies. It can be
seen in Figs. 6B, D, and F that the 3 amph and the esc/amph
group appear to have higher levels of DA and its metabolites
in the nonlesioned SN. One-way ANOVAs conducted on data
expressed as the percentage of DA, DOPAC, and HVA
remaining on the lesioned side [(amount remaining on the
lesioned side / amount on the nonlesioned side) � 100%]
revealed no significant differences between the groups on any
of these measures.
TH immunoreactivity in SNc and VTA
The number of TH-positive cells remaining on the
lesion and no-lesion side in the SNc and VTA of rats
treated with the escalating-dose regimen of amphetamine
before intra-MFB infusions of 6-OHDA is shown in
(B), a dashed line at 50% represents no asymmetry or an equal number of
Fig. 8 (Experiment 3). It can be seen that 28 days after
lesioning, the number of TH-IR cells was reduced on the
side of the lesion in both the SNc and VTA. It is readily
apparent, however, that the esc/amph group had a greater
number of TH-IR cells remaining on the side of the lesion
than the sal group. This effect is reflected in a significant
Group � Side interaction. See Fig. 8 legend for results of
the statistical analyses.
Discussion
The purpose of this investigation was to determine
whether exposure to a sensitizing regimen of amphetamine
before 6-OHDA lesions of the nigrostriatal DA system
would facilitate behavioral sparing after such lesions. The
present experiments, conducted independently in separate
laboratories, using different strains of rats and different
placements of 6-OHDA infusions within regions of the
nigrostriatal pathway, show that exposure to the 2-week
escalating-dose regimen of amphetamine before lesioning

Fig. 5. Experiment 3: Mean (F SEM) basal concentrations of extracellular DA, DOPAC, and HVA in six dialysate samples taken at 20 min intervals on the
lesioned (L) and nonlesioned (NL) side of the striatum on days 10–14 (A, C, E) and 24–28 (B, D, F) after 6-OHDA lesions of theMFB. ANOVAs (Drug � Side)
yielded the following significant effects on Test 1: DA, side, F(1,10) = 17.06, P < 0.05, Drug � Side interaction, F(1,10) = 6.29, P < 0.05; DOPAC, side,
F(1,10) = 52.87, P < 0.05; HVA, side, F(1,10) = 80.48, P < 0.05, and on Test 2: DA, side, F(1,10) = 7.36, P < 0.05; DOPAC, side, F(1,9) = 28.86, P < 0.05;
HVA, side, F(1,10) = 48.41, P < 0.05. Paired sample t tests conducted to analyze simple main effects revealed that 11–14 days after lesioning, DA levels in the
dialysate were lower on the lesioned than on the nonlesioned side only in the sal group (P < 0.05, Bonferroni corrected).
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9386
prevented the asymmetries of forelimb use and turning
behavior seen in rats given saline during the same period.
Furthermore, it was shown in Experiment 2 that exposure to
three nonescalating injections of amphetamine before
lesioning did not lead to behavioral sparing. The prophy-
lactic effect of the escalating-dose regimen was comparable
to the effect of postlesion exercise as seen here in Experi-
ment 2 and as reported previously (Tillerson et al., 2001,
2002, 2003). Note, however, that the effects seen here were
obtained with a novel and less intense exercise program than
the ones used by Tillerson et al.
The major strength of the present investigation is
derived from the fact that our primary findings of behav-
ioral sparing after prelesion treatment with the escalating
doses of amphetamine are consistent in all experiments,
despite methodological differences, such as the site of 6-
OHDA administration: intrastriatal (Experiment 1) versus
intra-MFB (Experiments 2 and 3). Even though the
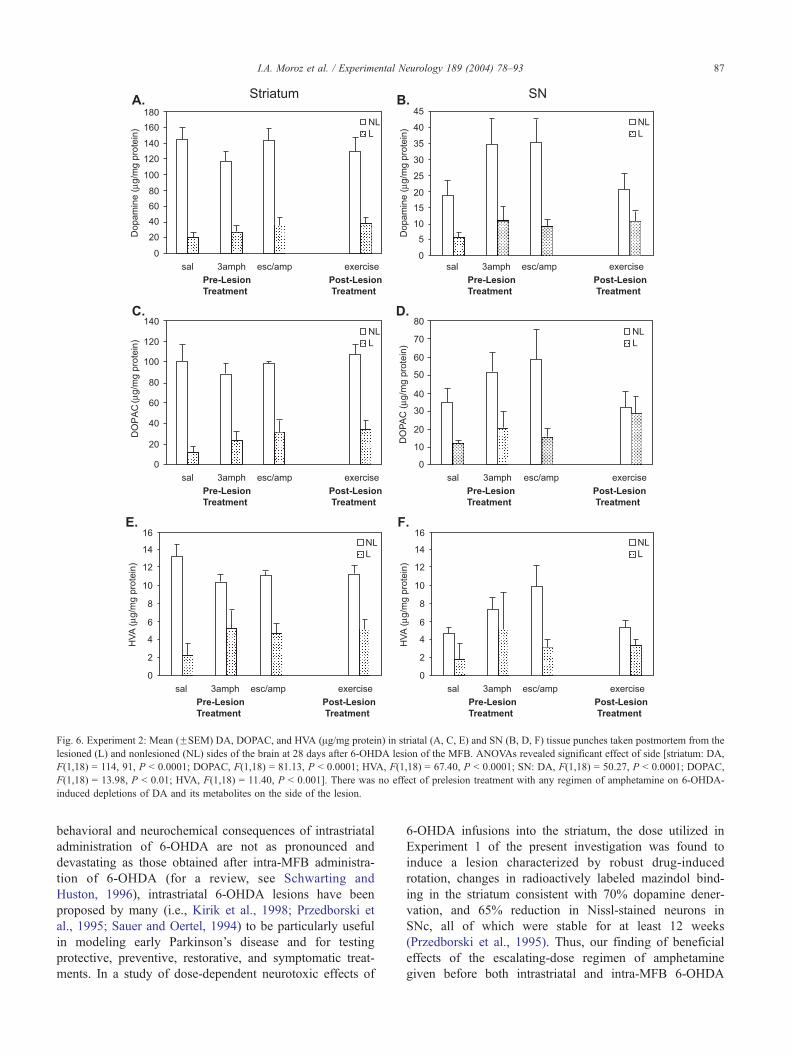
Fig. 6. Experiment 2: Mean (FSEM) DA, DOPAC, and HVA (Ag/mg protein) in striatal (A, C, E) and SN (B, D, F) tissue punches taken postmortem from the
lesioned (L) and nonlesioned (NL) sides of the brain at 28 days after 6-OHDA lesion of the MFB. ANOVAs revealed significant effect of side [striatum: DA,
F(1,18) = 114, 91, P < 0.0001; DOPAC, F(1,18) = 81.13, P < 0.0001; HVA, F(1,18) = 67.40, P < 0.0001; SN: DA, F(1,18) = 50.27, P < 0.0001; DOPAC,
F(1,18) = 13.98, P < 0.01; HVA, F(1,18) = 11.40, P < 0.001]. There was no effect of prelesion treatment with any regimen of amphetamine on 6-OHDA-
induced depletions of DA and its metabolites on the side of the lesion.
I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 87
behavioral and neurochemical consequences of intrastriatal
administration of 6-OHDA are not as pronounced and
devastating as those obtained after intra-MFB administra-
tion of 6-OHDA (for a review, see Schwarting and
Huston, 1996), intrastriatal 6-OHDA lesions have been
proposed by many (i.e., Kirik et al., 1998; Przedborski et
al., 1995; Sauer and Oertel, 1994) to be particularly useful
in modeling early Parkinson’s disease and for testing
protective, preventive, restorative, and symptomatic treat-
ments. In a study of dose-dependent neurotoxic effects of
6-OHDA infusions into the striatum, the dose utilized in
Experiment 1 of the present investigation was found to
induce a lesion characterized by robust drug-induced
rotation, changes in radioactively labeled mazindol bind-
ing in the striatum consistent with 70% dopamine dener-
vation, and 65% reduction in Nissl-stained neurons in
SNc, all of which were stable for at least 12 weeks
(Przedborski et al., 1995). Thus, our finding of beneficial
effects of the escalating-dose regimen of amphetamine
given before both intrastriatal and intra-MFB 6-OHDA

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9388
insult has significant implications for understanding of
mechanisms involved in early and more advanced stages
of nigrostriatal degeneration.
Fig. 8. Experiment 3: TH-IR. Mean (FSEM) number of TH-IR cells per
square millimeter in the SNc (A) and VTA (B), 28 days after 6-OHDA lesion
of theMFB. ANOVAs (Drug� Side) revealed significant main effect of drug
[SNc, F(1,10) = 7.25, P = .02; VTA, F(1,10) = 5.22, P = 0.04], side [SNc,
F(1,10) = 53.26, P < 0.0001; VTA, F(1,10) = 196.17, P < 0.0001], and Drug
� Side interaction [SNc, F(1,10) = 4, 11, P = 0.07; VTA, F(1,10) = 6.46, P =
0.03]. Unpaired t tests conducted to analyze simple main effects revealed that
the number of TH-IR cells remaining on the side of the lesion was higher in
esc/amph group (*P < 0.05; Bonferroni corrected).
Fig. 7. Experiment 3:Mean (FSEM)DA,DOPAC, andHVA (Ag/mg protein)
in striatal tissue punches taken postmortem from the lesioned (L) and
nonlesioned (NL) sides of the brain at 28 days after 6-OHDA lesion of the
MFB. ANOVAs revealed significant effect of side [DA, F(1,10) = 74, 36, P <
0.001; DOPAC, F(1,10) = 86.97, P < 0.0001; HVA, F(1,10) = 56.87, P <
0.0001]. There was no effect of prelesion treatment with the escalating-dose
regimen of amphetamine on 6-OHDA-induced depletions of DA and its
metabolites on the side of the lesion.

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 89
Acutely, amphetamine, unlike other stimulants such as
cocaine, induces DA release in both cell body and terminal
regions through its actions at the dopamine transporter (DAT;
Povlock and Schenk, 1997; Seiden et al., 1993). The
importance of DAT in the acute effects of amphetamine on
dopamine release has raised previously the possibility that
adaptations in DAT function might underlie the development
of sensitization after repeated drug exposure. Similarly, in
light of findings showing that DAT plays a crucial role in the
neurodegenerative responses to dopaminergic neurotoxins,
and that processes that inhibit DAT activity can ameliorate
this neurotoxicity (for a review, see Fleming et al., in press;
Miller et al., 1999), it is tempting to speculate that the
behavioral sparing observed in the group exposed to the 2-
week escalating-dose regimen of amphetamine before
lesioning might have occurred as an effect of sustained
down-regulation of DAT resulting from the repeated expo-
sure of this group to amphetamine before lesioning. Inter-
estingly, however, unlike cocaine (Letchworth et al., 1997,
2001; Pilotte et al., 1994), repeated amphetamine adminis-
tration has been reported not to affect striatal DA uptake or
DAT binding (Allard et al., 1990; Mintz et al., 1994; Persico
et al., 1993), and on the contrary modest increases in DAT
mRNA have been observed (Lu and Wolf, 1997; Shilling et
al., 1997). It can be noted, as well, that in studies done in
vitro in PC12 cells, amphetamine has been found both to
sensitize dopamine release and to cause neurite outgrowth,
but not to have any residual effects on either dopamine
uptake or binding to the transporter (Kantor et al., 2002; Park
et al., 2002). Furthermore, unlike the findings showing
reduced nigrostriatal neurodegeneration when neurotoxins
were administered in the presence of DAT-uptake inhibitors
(Clarke and Reuben, 1995; Ricaurte et al., 1985) or in DAT
knockout mice (Gainetdinov et al., 1997; Sanghera et al.,
1997), the results of Experiments 2 and 3 revealed no
neuroprotection of striatal or SN tissue levels of DA or its
metabolites in the group of rats exposed to the escalating-
dose regimen of amphetamine administered 7 days before
lesioning. Thus, the behavioral sparing observed in that
group cannot be attributed to a different degree of striatal
tissue DA depletion.
In Experiment 3, using in vivo microdialysis, it was
found that behavioral sparing in rats exposed to the esca-
lating-dose regimen of amphetamine before lesioning was
accompanied by normalization of basal extracellular levels
of DA in terminal striatal regions observed as early as 11–
14 days after lesioning. Specifically, the concentration of
DA in dialysate from the striatum ipsilateral to the 6-OHDA
lesion was as high as that from the contralateral striatum in
rats treated with escalating doses of amphetamine before
lesioning but was significantly lower in rats treated with
saline during the same period. At the same time, significant
reduction of the basal levels of DOPAC and HVA in the
ipsilateral striatum, as well as depletion of striatal tissue
levels of DA, clearly indicated the presence of lesions in
both groups. The finding that behavioral sparing was not
accompanied by higher tissue levels of DA in Experiments 2
and 3 is in agreement with earlier microdialysis studies
showing that the gradual process of behavioral recovery
from partial 6-OHDA lesions is accompanied by normali-
zation of extracellular DA levels, but not tissue DA levels in
the striatum (Robinson and Whishaw, 1988; Robinson et al.,
1994). In particular, extracellular DA levels in the striatum
were reduced 4 days after the lesion when ipsilateral turning
predominated but were normalized 3–4 weeks later when
turning was no longer asymmetric. The significance of this
finding lies in the fact that even though a number of
presynaptic compensatory adaptations within the DA neu-
rons, such as increased DA metabolism, synthesis, and
release, proposed to play a role in behavioral recovery have
been found to be maximal within 3 days following such
lesions, behavioral recovery was not complete until 3–4
weeks after lesioning (Altar et al., 1994). Thus, the gradual
normalization of extracellular DA demonstrated by Robin-
son et al. (1994) seemed more critical for behavioral
recovery. The present findings of behavioral sparing and
normalization of extracellular DA being evident as early as
11–14 days after lesioning in rats treated with an escalating-
dose regimen of amphetamine before lesioning suggest that
exposure to this regimen may accelerate the development of
compensatory changes in the DA neurons that typically
accompany behavioral recovery. The present results are
particularly striking since, unlike in Robinson et al.
(1994), the lesions in Experiments 2 and 3 were made in
the MFB and not in the SN. In contrast to SN lesions, after
which behavioral recovery has been reported to occur
spontaneously (Emmi et al., 1996, 1997; Robinson and
Whishaw, 1988; Robinson et al., 1994), MFB lesions are
known to produce more extensive striatal DA depletions
(Costall et al., 1976), greater losses of TH-IR neurons in the
SN (Carman et al., 1991; Perese et al., 1989), and more
devastating behavioral deficits from which animals never
recover (Carman et al., 1991; Kirik et al., 1998; Lee et al.,
1996) unless interventions such as forced use of the im-
paired side of the body (Tillerson et al., 2001, 2002) or
wheel running (Tillerson et al., 2003) are employed.
Further, the results of Experiment 3 revealed that rats
exposed to the escalating-dose regimen of amphetamine
before lesioning had reduced lesion-induced loss of TH-IR
cells in the ipsilateral DA cell body regions, SNc and VTA,
compared to rats exposed to saline, indicating a partial
neuroprotective effect of prelesion treatment with escalating
doses of amphetamine. It is important to note, however, that
at the same time, these rats had comparably large depletions
of striatal tissue DA (Experiment 2 and 3) and SN tissue DA
(Experiment 3) as rats exposed to saline during the same
period. Another possible explanation for the reduced loss of
TH-IR cells that we observed 28 days after lesioning is that
it may reflect the return of the dopaminergic phenotype
observed 32 weeks after lesioning by Bowenkamp et al.
(1996) and Finkelstein et al. (2000). Although the functional
significance of this reduced loss of TH-IR cells is unknown

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9390
at the present time, it may reflect compensatory changes in
the functioning of the remaining DA cells. As mentioned
earlier, the enduring enhancement of function within the
remaining DA neurons after partial lesions of the nigros-
triatal DA system is reminiscent of the enhanced DA
response seen in sensitization to amphetamine. Specifically,
behavioral sensitization is accompanied by augmented ex-
tracellular DA overflow in striatal regions in response to
subsequent challenges, such as exposure to drugs or stres-
sors, in the absence of changes in tissue levels of DA
(Robinson and Becker, 1986). Importantly, the events that
lead to enhanced DA functioning are initiated by the actions
of psychostimulants, such as amphetamine, in the DA cell
body regions (Cador et al., 1995; Kalivas and Weber, 1988,
Vezina, 1993; Vezina and Stewart, 1990) and may include,
among other factors, sustained increase in astrocytic FGF-2
expression in these regions (Flores and Stewart, 2000a,b;
Flores et al., 1998). Thus, possibly the reduced loss of TH-
IR cells in the SNc and VTA of rats exposed to the prelesion
treatment with escalating doses of amphetamine reflects a
local protection of these cells as a result of the ability of this
drug treatment to induce increases in the expression of one
or more neurotrophic factors such as FGF-2.
The idea that manipulations given before brain injury
can be protective is not unprecedented. Numerous studies
have shown that exogenous delivery of FGF-2, BDNF, or
GDNF before 6-OHDA lesions prevents the development
of lesion-induced behavioral deficits and protects against
loss of DA neurons and striatal DA innervation (Altar et
al., 1994; Choi-Lundberg et al., 1998; Mandel et al., 1997;
Shults et al., 1996, 2000). Recently, it was found that
forced use of one forelimb for 7 days before 6-OHDA
lesions led to sparing of limb use and attenuation of DA
loss in striatal tissue (Cohen et al., 2003). Interestingly,
forced use of one forelimb in itself increases FGF-2-IR in
the SNc and VTA in both hemispheres (unpublished
observations; Moroz et al., 2002) and GDNF in the
striatum contralateral to the overused forelimb (Cohen et
al., 2003). In the present experiment, exposure to the
escalating-dose regimen of amphetamine known to in-
crease the expression of FGF-2 in both the DA cell bodies
and terminals (Flores and Stewart, 2000b) led to behav-
ioral sparing after 6-OHDA lesions, whereas the exposure
to three injections of amphetamine, found previously to
increase FGF-2 expression only in the cell body regions of
DA neurons (Flores et al., 1998), did not. This finding
appears to suggest that increased expression of FGF-2, and
possibly other neurotrophic factors, in the terminal regions
of DA neurons may be more critical for behavioral
recovery. Support for this idea comes from studies show-
ing that preservation of normal motor functions in the 6-
OHDA lesion model requires intrastriatal, but not nigral,
administration of GDNF (Kirik et al., 2000; Shults et al.,
1996; Sullivan et al., 1998). We emphasize, however, that
the direct involvement of neurotrophic factors in the
beneficial effects of prelesion exposure to the escalating-
dose regimen of amphetamine or prelesion forced use of
one limb remains to be demonstrated.
Another explanation for the behavioral sparing induced by
previous exposure to the escalating-dose regimen of amphet-
amine is its effect on the dendritic morphology of postsyn-
aptic neurons (Li et al., 2003; Robinson and Kolb, 1997,
1999). Such morphological changes could represent a reor-
ganization of synaptic inputs onto these neurons. Conse-
quently, the prophylactic effects of exposure to the escalating-
dose regimen of amphetamine before 6-OHDA lesions of the
nigrostriatal DA neurons found in the present experiments
could be mediated by the amphetamine-induced increases in
neurotrophic factors (Flores and Stewart, 2000b) and the
enhanced synaptic transmission between the surviving DA
input neurons and their striatal and cortical targets.
Interestingly, postinjury administration of amphetamine
has been shown to enhance functional recovery in patients
suffering from stroke and in animals following cortical
lesions or ischemia (Feeney, 1997; Gladstone and Black,
2000; Goldstein, 2000). Multiple mechanisms have been
proposed to underlie this therapeutic effect of amphetamine,
including neuritogenesis, synaptogenesis (Stroemer et al.,
1998), and enhancement of noradrenergic transmission
(Feeney, 1997; Gladstone and Black, 2000). Importantly,
behavioral recovery observed in ischemic animals treated
with amphetamine correlates with increased expression of
markers of neuronal remodeling, such as growth associated
protein-43 (GAP-43) and synaptophysin (Stroemer et al.,
1995). As argued earlier, modifications of neuronal archi-
tecture and synaptic connectivity are likely to be mediated
via actions of neurotrophic factors. In fact, there is evidence
that exogenous administration of FGF-2 after the onset of
ischemia enhances behavioral recovery without reducing the
infarct volume (Kawamata et al., 1996). Instead, FGF-2
administration is associated with a selective increase in
GAP-43 expression (Kawamata et al., 1997).
In summary, we report that exposure to a 2-week
escalating-dose regimen of amphetamine before unilateral
6-OHDA lesions prevents asymmetries of forelimb use and
turning behavior and leads to normalization of the basal
extracellular DA in striatal dialysate on the side of a
unilateral 6-OHDA lesion in the absence of significant
protection of striatal tissue DA levels. Although the mech-
anisms mediating this effect remain to be elucidated, pos-
sibly this prelesion exposure to the escalating doses of
amphetamine leads to behavioral sparing after subsequent
6-OHDA lesions via acceleration of the development of
compensatory changes in the DA neurons that typically
accompany behavioral recovery. Furthermore, the endoge-
nous increases in the expression of neurotrophic growth
factors previously observed after exposure to the 2-week
escalating-dose regimen of amphetamine may play a key
role in the enhanced responsiveness of the DA system to a
subsequent 6-OHDA insult and the resulting behavioral
sparing. Although the findings do not speak directly to the
therapeutic use of amphetamine to facilitate recovery after

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 91
injury, they point to mechanisms that may be involved. For
example, in agreement with epidemiological data, emerging
evidence from animal models of neurogenerative diseases,
including Parkinson’s disease (PD), strongly suggest that
experiential interventions, such as exercise, enriched envi-
ronment housing, and dietary restriction, offer substantial
protection against the behavioral and neurochemical deficits
observed in these models (see Bezard et al., 2003; and
reviews by Mattson, 2000; Smith and Zigmond, 2003).
Interestingly, this protection has been demonstrated to be
due, at least in part, to the ability of these interventions to
increase availability of neurotrophic factors, such as BDNF,
GNDF, and FGF-2, in the areas affected by the injury. Along
similar lines, epidemiological data reporting an apparent
protective effect of cigarette smoking on the risk of PD have
been extended by experimental findings from animal mod-
els of PD suggesting that regional increases in neurotrophic
factors, BDNF and FGF-2, serve as a possible mechanism
by which (�)-nicotine protects from experimental parkin-
sonism (Belluardo et al., 1998; Blum et al., 1996; Maggio et
al., 1997). Thus, with the progress made in early detection
of neurodegenerative diseases, experiential and pharmaco-
logical interventions known to increase levels of neuro-
trophic factors in the brain may be used prophylactically to
slow down or even prevent the neurodegenerative cascade
and the consequent motor dysfunction associated with PD.
Acknowledgments
We thank Susan Ajersch and Heshmat Rajabi for their
assistance with behavioral testing and tissue processing in
Experiments 2 and 3. We also thank Gabriela Redwine for
editorial help. This project was supported by grants to JS from
Canadian Institutes of Health Research and Le Fonds
Quebecois de la recherche sur la nature et les technologies
(FRNTQ) and by the National Institutes of Health grant to TS.
References
Abercrombie, E.D., Bonatz, A.E., Zigmond, M.J., 1990. Effects of L-dopa
on extracellular dopamine in striatum of normal and 6-hydroxydop-
amine-treated rats. Brain Res. 525, 36–44.
Allard, P., Eriksson, K., Ross, S.B., Marcusson, J.O., 1990. Unaltered
[3H]GBR-12935 binding after chronic treatment with dopamine active
drugs. Psychopharmacology 102, 291–294.
Altar, C.A., Boylan, C.B., Fritsche, M., Jones, B.E., Jackson, C., Wiegard,
S.J., Lindsay, R.M., Hyman, C., 1994. Efficacy of brain-derived neuro-
trophic factor and neurotrophin-3 on neurochemical and behavioral
deficits associated with partial nigrostriatal dopamine lesions. J. Neuro-
chem. 63, 1021–1032.
Belluardo, N., Blum, M., Mudo, G., Andbjer, B., Fuxe, K., 1998. Acute
intermittent nicotine treatment produces regional increases of basic fi-
broblast growth factor messenger RNA and protein in the tel- and
diencephalon of the rat. Neuroscience 83, 723–740.
Bezard, E., Dovero, S., Belin, D., Duconger, S., Jackson-Lewis, V., Przed-
borski, S., Piazza, P.V., Gross, C.E., Jaber, M., 2003. Enriched environ-
ment confers resistance to phenyl-1,2,3,6-Tetrahydropyridine and
cocaine: involvement of dopamine transporter and trophic factors. J.
Neurosci. 23, 10999–11007.
Blum, M., Wu, G., Mudo, G., Belluardo, N., Andersson, K., Agnati, L.F.,
Fuxe, K., 1996. Chronic continuous infusion of (�) nicotine reduces
basic fibroblast growth factor messenger RNA levels in the ventral
midbrain of the intact but not of the 6-hydroxydopamine-lesioned rat.
Neuroscience 70, 169–177.
Bowenkamp, K.E., David, D., Lapchak, P.L., Henry, M.A., Granholm,
A.C., Hoffer, B.J., Mahalik, T.J., 1996. 6-hydroxydopamine induces
the loss of the dopaminergic phenotype in substantia nigra neurons
of the rat. A possible mechanism for restoration of the nigrostriatal
circuit mediated by glial cell line-derived neurotorphic factor. Expl.
Brain Res. 111, 1–7.
Cador, M., Bjijou, Y., Stinus, L., 1995. Evidence of complete independence
of the neurobiological substrates for the induction and expression of
behavioral sensitization to amphetamine. Neuroscience 65, 385–395.
Carman, L.S., Gage, F., Shults, C.W., 1991. Partial lesions of the substantia
nigra: relation between extent of lesion and rotational behavior. Brain
Res. 533, 275–283.
Castaneda, E., Whishaw, I.Q., Robinson, T.E., 1990. Changes in striatal
dopamine neurotransmission assessed with microdialysis following re-
covery from a bilateral 6-OHDA lesion: variation as a function of lesion
size. J. Neurosci. 10, 1847–1854.
Choi-Lundberg, D.L., Lin, Q., Schallert, T., Crippens, D., Davidson, B.L.,
Chang, Y.-N., Chiang, Y.L., Qian, J., Bardwaj, L., Bohn, M.C., 1998.
Behavioral and cellular protection of rat dopaminergic neurons by an
adenoviral vector encoding glial cell line-derived neurotrophic factor.
Exp. Neurol. 154, 261–275.
Clarke, P.B., Reuben, M., 1995. Inhibition by dizocilpine (MK-801) of
striatal dopamine release induced by MPTP and MPP+: possible action
at the dopamine transporter. Br. J. Pharmacol. 114, 315–322.
Cohen, A.D., Tillerson, J.L., Smith, A.D., Schallert, T., Zigmond, M.J.,
2003. Neuroprotective effects of prior limb use in 6-hydroxydopamine-
treated rats: possible role of GDNF. J. Neurochem. 85, 299–305.
Costall, B., Marsden, C.D., Naylor, R.J., Pycock, C.J., 1976. The relation-
ship between striatal and mesolimbic dopamine dysfunction and the
nature of circling responses following 6-hydroxydopamine and electro-
lytic lesions of the ascending dopamine systems of rat brain. Brain Res.
118, 87–113.
Emmi, A., Rajabi, H., Stewart, J., 1996. Behavioral and neurochemical
recovery from partial 6-hydroxydopamine lesions of the substantia
nigra is blocked by daily treatment with glutamate receptor antagonists
MK-801 and CPP. J. Neurosci. 16, 5216–5224.
Emmi, A., Rajabi, H., Stewart, J., 1997. Behavioral and neurochemical
recovery from partial 6-hydroxydopamine lesions of the substantia
nigra is blocked by daily treatment with D1/D5, but not D2, dopamine
receptor antagonists. J. Neurosci. 17, 3840–3846.
Feeney, D.M., 1997. From laboratory to clinic: noradrenergic enhance-
ment of physical therapy for stroke or trauma patients. Adv. Neurol.
73, 383–394.
Finkelstein, D.I., Stanic, F., Parish, C.L., Thomas, D., Dickson, K., Horne,
M.K., 2000. Axonal sprouting following lesions of the rat substantia
nigra. Neuroscience 97, 99–112.
Fleming, S.M., Delville, Y., Schallert, T., 2003. An intermittent, controlled-
rate, slow progressive degenerationmodel of Parkinson’s disease suitable
for evaluating neuroprotective therapies: effects of methylphenidate.
Behav. Brain Res. (in press)..
Flores, C., Stewart, J., 2000a. Basic fibroblast growth factor as a mediator
of the effects of glutamate in the development of long lasting sensiti-
zation to stimulant drugs: studies in the rat. Psychopharmacology 151,
152–165.
Flores, C., Stewart, J., 2000b. Changes in astrocytic basic fibroblast growth
factor expression during and after prolonged exposure to escalating
doses of amphetamine. Neuroscience 298, 287–293.
Flores, C., Rodaros, D., Stewart, J., 1998. Long-lasting induction of astro-
cytic basic fibroblast growth factor by repeated injections of amphe-

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–9392
tamine: blockade by concurrent treatment with a glutamate antagonist. J.
Neurosci. 18, 9547–9555.
Flores, C., Samaha, A.-N., Stewart, J., 2000. Requirement of endogenous
basic fibroblast growth factor for sensitization to amphetamine. J. Neu-
rosci. 20, 1–5.
Fornaguera, J., Carey, R.J., Dai, H., Huston, J.P., Schwarting, R.K.W.,
1994. Differentiation of motor inactivation from movement asymmetry
effects in an animal model of hemi-parkinsonism. NeuroReport 6,
173–194.
Gainetdinov, R.R., Fumagalli, F., Jones, S.R., Caron, M.G., 1997. Dopa-
mine transporter is required for in vivo MPTP neurotoxicity: evidence
from mice lacking the transporter. J. Neurochem. 69, 1322–1325.
Gladstone, D.J., Black, S.E., 2000. Enhancing recovery after stroke with
noradrenergic pharmacology: a new frontier? Can. J. Neurol. Sci. 27,
97–105.
Goldstein, L.B., 2000. Effects of amphetamines and small related mole-
cules on recovery after stroke in animals and man. Neuropharmacology
39, 852–859.
Gordon, M.N., Schreier, W.A., Ou, X., Holcomb, L.A., Morgan, D.G.,
1997. Exaggerated astrocyte reactivity after nigrostriatal deafferentia-
tion in the aged rat. J. Comp. Neurol. 388, 106–119.
Kalivas, P.W., Stewart, J., 1991. Dopamine transmission in the initiation
and expression of drug- and stress-induced sensitization of motor ac-
tivity. Brain Res. Rev. 16, 223–244.
Kalivas, P.W., Weber, B., 1988. Amphetamine injection into the A10
dopamine region sensitizes rats to peripheral amphetamine and co-
caine. J. Pharmacol. Exp. Ther. 245, 1095–1102.
Kantor, L., Park, Y.H., Wang, K.K.W., Gnegy, M.E., 2002. Enhanced
amphetamine-mediated dopamine release develops in PC12 cells
after repeated amphetamine treatment. Eur. J. Pharmacol. 451,
27–35.
Kawamata, T., Alexis, N.E., Dietrich, W.D., Finklestein, S.P., 1996. Intra-
cisternal basic fibroblast growth factor (bFGF) enhances behavioral
recovery following focal cerebral infarction in the rat. J. Cereb. Blood
Flow Metab. 16, 542–547.
Kawamata, T., Dietrich, W.D., Schallert, T., Gotts, J.E., Cocke, R.R., Beno-
witz, L.L., Finkelstein, S.P., 1997. Intracisternal basic fibroblast growth
factor enhances functional recovery and up-regulates the expression of a
molecular marker of neuronal sprouting following focal cerebral infarc-
tion. Proc. Natl. Acad. Sci. 94, 8179–8184.
Kirik, D., Rosenblad, C., Bjorklund, A., 1998. Characterization of beha-
vioral and neurodegenerative changes following partial lesions of the
nigrostriatal dopamine system induced by intrastriatal 6-hydroxydop-
amine in the rat. Exp. Neurol. 152, 259–277.
Kirik, D., Rosenblad, C., Bjorklund, A., 2000. Preservation of a functional
nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA
lesion model depends on the site of administration of the trophic factor.
Eur. J. Neurosci. 12, 3871–3882.
Lee, C.S., Sauer, H., Bjorklund, A., 1996. Dopaminergic neuronal degen-
eration and motor impairments following axon terminal lesion by intra-
striatal 6-hydroxydopamine in the rat. Neuroscience 72, 641–653.
Letchworth, S.R., Daunais, J.B., Hedgecock, A.A., Porrino, L.J., 1997.
Effects of chronic cocaine administration on dopamine transporter
mRNA and protein in the rat. Brain Res. 750, 214–222.
Letchworth, S.R., Nader, M.A., Smith, H.R., Friedman, D.P., Porrino, L.J.,
2001. Progression of changes in dopamine transporter binding site
density as a result of cocaine self-administration in rhesus monkeys.
J. Neurosci. 21, 2799–2807.
Li, Y., Kolb, B., Robinson, T.E., 2003. The location of persistent amphet-
amine-induced changes in the density of dendritic spines on medium
spiny neurons in the nucleus accumbens and caudate-putamen. Neuro-
psychopharmacology 28, 1082–1085.
Lu, W., Wolf, M.E., 1997. Expression of dopamine transporter and
vesicular monoamine transporter 2 mRNAs in rat midbrain after
repeated amphetamine administration. Brain Res. Mol. Brain Res. 49,
137–148.
Maggio, R., Riva, M., Vaglini, F., Fornai, F., Racagni, G., Corsini, G.U.,
1997. Striatal increase of neurotrophic factors as a mechanism of nic-
otine protection in experimental parkinsonism. J. Neural Transm. 104,
1113–1123.
Mandel, R.J., Spratt, S.K., Snyder, R.O., Left, S.E., 1997. Midbrain injec-
tion of recombinant adeno-associated virus encoding rat glial cell line-
derived neurotrophic factor protects nigral neurons in a progressive 6-
hydroxydopamine-induced degeneration model of Parkinson’s disease
in rats. Proc. Natl. Acad. Sci. 94, 14083–14088.
Mattson, M.P., 2000. Neuroprotective signaling and the aging brain: take
away my food and let me run. Brain Res. 886, 47–53.
Meredith, G.E., Callen, S., Scheuer, D.A., 2002. Brain-derived neurotro-
phic factor expression is increased in the rat amygdala, piriform cortex
and hypothalamus following repeated amphetamine administration.
Brain Res. 949, 218–227.
Miller, G.W., Gainetdinov, R.R., Levey, A.I., Caron, M.G., 1999. Dopa-
mine transporters and neuronal injury. Trends Pharmacol. Sci. 20,
424–429.
Mintz, M., Gordon, I., Roz, N., Rehavi, M., 1994. The effect of repeated
amphetamine treatment on striatal DA transporter and rotation in rats.
Brain Res. 668, 239–242.
Moroz, I.A., Cohen, A.D., Tillerson, J.L., Maxwell, K., Martinez, E.,
Schallert, T., Stewart, J., 2002. Effects of forced limb use on behavioral
outcome and FGF-2-IR after partial unilateral 6-OHDA lesions of the
nigrostriatal dopamine neurons. Program No. 885.5.2002 Abstract
Viewer/Itinerary Planner. Society for Neuroscience, Washington, D.C.
(Online).
Moroz, I.A., Rajabi, H., Rodaros, D., Stewart, J., 2003. Effects of sex and
hormonal status on astrocytic FGF-2 and TH-immunoreactivity after
medial forebrain bundle 6-hydroxydopamine lesions of the midbrain
dopamine neurons. Neuroscience 118, 463–476.
Park, Y.H., Kantor, L., Wang, K.K.W., Gnegy, M.E., 2002. Repeated in-
termittent treatment with amphetamine induces neurite outgrowth in rat
pheochromocytoma cells (PC12 cells). Brain Res. 951, 43–52.
Paulson, P.E., Camp, D.M., Robinson, T.E., 1991. Time course of transient
behavioral depression and persistent behavioral sensitization in relation
to regional brain monoamine concentrations during amphetamine with-
drawal in rats. Psychopharmacology 103, 480–492.
Paxinos, G., Watson, C., 1997. The Rat Brain in Stereotaxic Coordinates.
Academic Press, New York.
Perese, D.A., Ulman, J., Viola, J., Ewing, S.E., Bankiewicz, K.S., 1989. A
6-hydroxydopamine-induced selective parkinsonian rat model. Brain
Res. 494, 285–293.
Persico, A.M., Schindler, C.W., Brannock, M.T., Gonzalez, A.M., Sur-
ratt, C.K., Uhl, G.R., 1993. Dopaminergic gene expression during
amphetamine withdrawal. NeuroReport 4, 41–44.
Pilotte, N.S., Sharpe, L.G., Kuhar, M.J., 1994. Withdrawal of repeated
intravenous infusions of cocaine persistently reduces binding to dopa-
mine transporters in the nucleus accumbens of Lewis rats. J. Pharmacol.
Exp. Ther. 269, 963–969.
Povlock, S.L., Schenk, J.O., 1997. A multisubstrate kinetic mechanism of
dopamine transport in the nucleus accumbens and its inhibition by
cocaine. J. Neurochem. 69, 1093–1105.
Przedborski, S., Levivier, M., Jiang, H., Ferreira, M., Jackson-Lewis, V.,
Donaldson, D., Togasaki, D.M., 1995. Dose-dependent lesions of the
dopaminergic nigrostriatal pathway induced by intrastriatal injection of
6-hydroxydopamine. Neuroscience 67, 631–647.
Ricaurte, G.A., Langston, J.W., DeLanney, L.E., Irwin, I., Brooks, J.D.,
1985. Dopamine uptake blockers protect against the dopamine deplet-
ing effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in
the mouse striatum. Neurosci. Lett. 59, 59–64.
Robinson, T.E., Becker, J.B., 1986. Enduring changes in brain and behav-
ior produced by chronic amphetamine administration: a review and
evaluation of animal models of amphetamine psychosis. Brain Res.
Rev. 396, 157–198.
Robinson, T.E., Kolb, B., 1997. Persistent structural modifications in nu-
cleus accumbens and prefrontal cortex neurons produced by previous
experience with amphetamine. J. Neurosci. 17, 8491–8497.

I.A. Moroz et al. / Experimental Neurology 189 (2004) 78–93 93
Robinson, T.E., Kolb, B., 1999. Alterations in the morphology of den-
drites and dendritic spines in the nucleus accumbens and prefrontal
cortex following repeated treatment with amphetamine or cocaine. Eur.
J. Neurosci. 11, 1598–1604.
Robinson, T.E., Whishaw, I.Q., 1988. Normalization of extracellular dopa-
mine in striatum following recovery from a partial unilateral 6-OHDA
lesion of the substantia nigra: a microdialysis study in freely moving
rats. Brain Res. 450, 209–224.
Robinson, T.E., Mocsary, Z., Camp, D.M., Whishaw, I.Q., 1994. Time
course of recovery of extracellular dopamine following partial damage
to the nigrostriatal dopamine system. J. Neurosci. 14, 2687–2696.
Sanghera, M.K., Manaye, K., McMahon, A., Sonsalla, P.K., German, D.C.,
1997. Dopamine transporter mRNA levels are high in midbrain neurons
vulnerable to MPTP. NeuroReport 8, 3327–3331.
Sauer, H., Oertel, W.H., 1994. Progressive degeneration of nigrostriatal
dopamine neurons following intrastriatal terminal lesions with 6-
hydroxydopamine: a combined retrograde tracing and immunocyto-
chemical study in the rat. Neuroscience 59, 401–415.
Schallert, T., Tillerson, J., 1999. Intervention strategies for degeneration of
dopamine neurons in Parkinsonism: optimizing behavioral assessment of
outcome. In: Emerich, D.F., Dean III, R.L., Sanberg, P.R. (Eds.), Central
Nervous System Diseases. Humana, Totowa, NJ, pp. 131–151.
Schwarting, R.K.W., Huston, J.P., 1996. Unilateral 6-hydroxydopamine
lesions of the meso-striatal dopamine neurons and their physiological
sequelae. Prog. Neurobiol. 49, 215–266.
Seiden, L.S., Sabol, K.E., Ricaurte, G.A., 1993. Amphetamine: effects on
catecholamine systems and behavior. Annu. Rev. Pharmacol. Toxicol.
33, 639–677.
Shilling, P.D., Kelsoe, J.R., Segal, D.S., 1997. Dopamine transporter
mRNA is up-regulated in the substantia nigra and the ventral teg-
mental area of amphetamine-sensitized rats. Neurosci. Lett. 236,
131–134.
Shults, C.W., Kimber, T., Martin, D., 1996. Intrastriatal injection of
GDNF attenuates the effects of 6-hydroxydopamine. NeuroReport 7,
627–631.
Shults, C.W., Ray, J., Tsuboi, K., Gage, F.H., 2000. Fibroblast growth
factor-2-producing fibroblasts protect the nigrostriatal dopaminergic
system from 6-hydroxydopamine. Brain Res. 883, 192–204.
Smith, A.D., Zigmond, M.J., 2003. Can the brain be protected through
exercise? Lessons from an animal model of parkinsonism. Exp. Neurol.
184, 31–39.
Stroemer, R.P., Kent, T.A., Hulsebosch, C.E., 1995. Correlation of cortical
plasticity with behavioral recovery following stroke with amphetamine
administration. J. Cereb. Blood Flow Metab. 15, S182.
Stroemer, R.P., Kent, T.A., Hulsebosch, C.E., 1998. Enhanced neocorti-
cal neural sprouting, synaptogenesis, and behavioral recovery with
D-amphetamine therapy after neocortical infarction in rats. Stroke 29,
2381–2393.
Sullivan, A.M., Opacka-Juffry, J., Blunt, S.B., 1998. Long-term protection
of the rat nigrostriatal dopaminergic system by glial cell line-derived
neurotrophic factor against 6-hydroxydopamine in vivo. Eur. J. Neuro-
sci. 10, 57–63.
Tillerson, J.L., Cohen, A.D., Philhower, J., Miller, G.W., Zigmond, M.J.,
Schallert, T., 2001. Forced limb-use effects on the behavioral and neu-
rochemical effects of 6-hydroxydopamine. J. Neurosci. 21, 4427–4435.
Tillerson, J.L., Cohen, A.D., Caudle, W.M., Zigmond, M.J., Schallert, T.,
Miller, G.W., 2002. Forced nonuse in unilateral parkinsonian rats exac-
erbates injury. J. Neurosci. 22, 6790–6799.
Tillerson, J.L., Caudle, W.M., Reveron, M.E., Miller, G.W., 2003. Exer-
cise induces behavioral recovery and attenuates neurochemical defi-
cits in rodent models of Parkinson’s disease. Neuroscience 119,
899–911.
Vezina, P., 1993. Amphetamine injected into the ventral tegmental area
sensitizes the nucleus accumbens dopaminergic response to systemic
amphetamine: an in vivo microdialysis study in the rat. Brain Res. 605,
332–337.
Vezina, P., Stewart, J., 1990. Amphetamine administered to the ventral
tegmental area but not to the nucleus accumbens sensitizes rats to
systemic morphine: lack of conditioned effects. Brain Res. 516,
99–106.
Copyright © 2022 FDOKUMEN




















