
Scaffold protein x11α interacts with kalirin-7 in dendrites and recruits it to Golgi outposts
Upload
univ-paris5Category
view
1download
0
h’euroscience Vol. 42, No. 1, pp. 19-28, 1991 hinted in Great Britain
0306-4522/91 $3.00 + 0.00 Pergamon Press plc
0 1991 IBRO
SPECIFIC ROLE OF N-ACETYL-ASPARTYL-GLUTAMATE IN THE IN VW0 REGULATION OF DOPAMINE RELEASE
FROM DENDRITES AND NERVE TERMINALS OF NIGROSTRIATAL DOPAMINERGIC NEURONS IN
THE CAT
T. GALLI,* G. GODEHEU, F. ARTAUD, J. M. DESCE, A. PITTALUGA, L. BARBEITO, J. GLOWSINKI and A. CHI%RAMY
College de France INSERM U 114, Chaire de Neuropharmacologie, 11 place Marcelin Eerthelot, 75231 Paris Cedex 05. France
Abstract-Levels of N-acetyl-aspartyl-glutamate measured by high-pressure liquid chromatography were found to be very high in the cat substantia nigra, particularly in the pars compacta, while those in the caudate nucleus were much lower. In halothane-anaesthetized cats implanted with push-pull cannulae, N-acetyl-aspartyl-glutamate (lo-*M) induced a marked and prolonged release of newly synthesized [3H]dopamine, when infused into the posterior but not into the anterior part of the caudate nucleus. In contrast, in the presence of tetrodotoxin (10e6M), N-acetyl-aspartyl-glutamate (lo-‘M) reduced the residual release of [3H]dopamine; this effect was also more pronounced in the posterior than in the anterior part. In the conditions used, as indicated by experiments with [3H]N-acetyl-aspartyl-glutamate no glutamate was formed from the infused N-acetyl-aspartyl-glutamate. Ibotenate (lo-’ M) induced changes in [3H]dopamine release in both the absence and presence of tetrodotoxin, which were closely similar to those observed with N-acetyl-aspartyl-glutamate. Responses induced by either N-acetyl-aspartyl-gluta- mate or ibotenate were not mediated by N-methyl-n-aspartate receptors since N-methyl-n-aspartate stimulated the release of [3H]dopamine only when used in a high concentration (1O-4 M) and applied in a magnesium-free superfusion medium in both the presence of glycine (10m6 M) and strychnine (10d6 M). In addition, the stimulatory effect of iV-methyl-D-aspartate persisted in the presence of tetrodotoxin; it was of similar amplitude in both parts of the caudate nucleus and of shorter duration than that evoked by either N-acetyl-aspartyl-glutamate or ibotenate alone. N-Acetyl-aspartyl-glutamate interacted with dopaminergic neurons not only presynaptically in the caudate nucleus but also in the substantia nigra since a marked increase in [3H]dopamine release was observed both from local dendrites and from nerve terminals in the ipsilateral caudate nucleus when N-acetyl-aspartyl-glutamate (lo-’ M) was infused locally into the substantia nigra pars compacta. No effect could be seen in contralateral structures.
The isomer of natural N-acetyl-aspartyl-glutamate, B-N-acetyl-aspartyl-glutamate (lo-’ M), had no effect on [‘Hldopamine release when applied similarly in the substantia nigra, thus confirming the specificity of the action of N-acetyl-aspartyl-glutamate.
The striatum is innervated by cortical neurons rich in glutamate and/or aspartate”,” and several studies made either in vitro or in vivo have indicated that glutamate stimulates the release of dopamine (DA) in the rat or the cat striatum.*SJ4~28~” The stimulatory effect of glutamate on DA release is mediated, at least in part, by receptors located on dopaminergic nerve terminals. Indeed, the glutamate-evoked release of DA still occurs in the presence of tetrodotoxin,*~yJ4,28 a neurotoxin which blocks sodium channels and which should abolish indirect effects. In addition, recent experiments have revealed that glutamate also stimulates the release of DA from purified synapto- somes from the rat striatum.22 As shown recently in vivo in the cat, receptors of the amino-3-hydroxy-5-
*To whom correspondence should be. addressed. Abbreviations: AMPA, amino-3-hydroxy-5-methylisoxa-
zole-Cproprionate; DA, dopamine; NAAG, N-acetyl- aspartyl-glutamate; NMDA, N-methyl-n-aspartate.
methylisoxazole4propionate (AMPA) subtype are involved in the stimulation of DA release evoked by a low concentration of glutamate.* Several studies performed in the rat have indicated that N-methyl-D- aspartate (NMDA) receptors also contribute to the presynaptic regulation of DA release. For instance, as shown in rat striatal slices, in the absence of magnesium, NMDA stimulates the release of DA, an effect which is potentiated by glycine through a strychnine-insensitive site. 27 Magnesium or NMDA receptor antagonists completely abolish the response evoked by NMDA. These effects were still observed in the presence of tetrodotoxin or using purified synaptosomes, providing further evidence for the localization of NMDA receptors on dopaminergic nerve terminals.yJ2~28~”
The present study was undertaken to determine whether or not the acetylated dipeptide. N-Acetyl- aspartyl-glutamate (NAAG) also presynaptically modulates the in vivo release of DA in the cat caudate
19

20 T. GALLI et ai.
nucleus. Although it is still a matter of controversy, there is evidence suggesting that NAAG acts as a neurotransmitter in the CNS possibly by acting as an agonist of NMDA receptors.24*26~M*37 In addition, both biochemical and immunohistoche~cal studies have demonstrated the presence of NAAG in the rat striatum’5.23 and NAAG levels were shown to be reduced following cerebral decortication or excito- toxic lesion of the striatum.” NAAG has also been found in high amounts in the rat substantia nigrai5 and NAAG has been suspected to be partly co-local- ized with DA in dopaminergic neurons.23 This led us to examine the effects of local application of NAAG in the cat substantia nigra on the release of DA from dendrites and nerve terminals of the nigrostriatal dop~inergic neurons.
EXPERIMENTAL PROCEDURES
Materials
Cats of either sex were purchased from IFFA-CREDO, France. NAAG was obtained from Bachem, r.13,5-3Hltyro- sine (50 Ci/mmol) and ~-a~tyl-~~rtyl-L~3,4-3-H~ut~ate ~~.7Ci/~ol~ from NEN: AG 5OW-X8 and AG 1-X2 . , resins from Bio-Rad, France; NMDA, glycine, tetrodotoxin and strychnine from Sigma and the fi isomer of NAAG was a generous gift of V. Serval (Laboratoire de Chimie Organique Biologique, Universite Pierre et Marie Curie, Paris, France). All other compounds were obtained from Prolabo, France.
In each animal and for each cannula, rH]DA in each successive fraction was expressed as a percentage of average s~n~n~us release calculated from the live fractions col- lected before the treatment. Results are the mean + S.E.M. of data obtained with n animals when compared with corresponding control values obtained in untreated animals using the two-tailed Student’s r-test.
Superfusion with [3H]N-acetyl-aspartyl-giutamate
Mecsaremenr of N-acetyl-aspnrtyl-gruramate in tissues
Two cats were kiiled under deep halothane anaesthesia and their brains removed. Serial frontal sections (SOO~m) were made at - 15°C using a Leitz Kryomat microtome. Microdiscs of tissue were punched out in the caudate nucleus with a cooled stainless steel tube (0.9 mm diameter) while pieces of tissue from the substantia nigra were microdissected. Both types of sample were introduced into tubes containing 200 ~1 of perchloric acid (0.1 N), sonicated and the homogenates were centrifuged (lOmin, 12000g). Pellets were kept for the estimation of proteins6 and super- natants were passed through microcolumns of AG 5OW-X8 cation exchange resin (200-400 mesh, hydrogen form). An aliquot (loo@) of each sample was injected into a Beckman Gold high-pressure liquid c~omato~aphy system using a Beckman ODS Cl8 Ult~phe~ column @pm, 4.6 x 250mm). The mobile phase (llow rate, 1 mljmin) consisted of a phosphate buffer 0.1 M KH,PG, adjusted to pH 3.25 with phosphoric acid. Retention time for NAAG was 9 min 30 s, NAAG being detected by U.V. absorption (210 nm). As determined using appropriate standards and peak area integration, the limit of detection for NAAG was 10 pmol. Overall recovery was 90%.
A putative hydrolysis of NAAG during its application into the caudate nucleus was tested on three occasions. Halothane-anaestbetized cats implanted with a push-pull cannula into the caudate nucleus were used in conditions exactly similar to those in the DA release experiments except that [3H]tyrosine was omitted from the artificial CSF and the treatment was performed using 13H]NAAG (labelled on glutamate). Successive lO-min fractions of superfusates were collected into tubes ~ntaining 10~1 of phosphate buffer (KH,PO,, 0.8 M, pH 5.25) used in order to inhibit any N-acetylated a-linked acidic dipeptidase activity. An aliquot (150 ~1) of each fraction was mixed with cold NAAG (6.7 x 10e4 M in 10 ~1) and applied on AGl-X2 (200400 mesh formate form) micro-columns. Columns were washed three times with 1 ml of water, then [‘H]NAAG was eluted with 1 ml of HCGGH 0.1 M and lyophilixed. Samples were resuspended with 200~1 of the mobile phase and 36~1 were injected into the high-pressure liquid chromatography system. The radioactivity of the eflhtent mobile phase was measured in successive 30-s fractions by liquid scintillation spectrometry. Aspartate, glutamate, and [3H]glutamate were also estimated in superfusates according to the method of Barbeito et aL3
Dqmnine release experiments RESULTS
Experiments were carried out on 154 adult cats of either sex. After tracheotomy, animals, under deep halothane anaesthesia (3%), were held in a stereotaxic frame using two bars fixed with an acrylic resin to the frontal and occipital bones. This fixation maintained the head in stereo&n& position and avoided the nociceptive stim~tions of classi- cal restraints. This allowed the concentration of halothane to be reduced to 1.5% at the end of surgery. Alveolar CO, EKG, and rectal temperature were monitored. In some experiments, one push-pull cannula (1.4 mm diameter) was implanted into the left caudate nucleus (A: 13-A: 19, L:4.5, H : 5). In other experiments, four push-pull cannulae were implanted into caudate nuclei (A: i6, L:4.S H: 5) and sub-
N-Acetyl-aspartyl-glutamate levels in the caudate nucleus and the substantia nigra of the cat
NAAG was measured in pieces of tissue punched out in different areas of the caudate nuclei or microdissected from the substantiae nigrae of successive frontal sections taken at different anterior positions.
NAAG could easily be measured in different areas of the caudate nucleus, higher IeveIs being found in the lateral (C) than in the dorsomedial (B) or ventral
stantiae nigrae (A : 4, L : 3, H : - 5) (according to the atlas of Jasper and Aimone-Marsan). ” Each cannula was continu- ously perfused with an artificial cerebrospinal fluid (in mM): NaCl 126.5. NaHCG, 27.5. KC1 2.4. KHPG, 0.5. CaCl, 1.1. MgCl, 0.83, Na$OJa0.5, ‘glucose 5.9, adjufted’to pH 7.3 with an O&O,, 95: 5 v/v mixture) containing 50 pCi of t[3,5-3H]tyrosine at the rate of 25O~l/lOmin using peri- staltic pumps. At the end of each experiment, the animal was perfused through the carotid artery with 10% formalin in isotonic saline solution under deep halothane anaesthesia. Serial frontal sections were stained with Cresyl Violet and submitted to histological control.
One hour after the onset of superfusion with rH]tyrosine, successive IO-min fractions were collected in tubes con- taining an ice-cold stabilizing thioglycolic acid solution (20 mM in ethanol, 1 ml). Samples were stored at - 20°C until biochemical analysis which was performed as pre- viously described.’ Briefly, PH]DA was separated from [‘Hltyrosine, [3H]metabolites, and [‘Hhadiolysis compounds in superfusate fractions using liquid chromato~phy on amber&e (CG 50, type II, 20&400 mesh bulTered with a NaH,PG.,/K,HPO, solution pH 6.1) then on alumina (neutral aluminium oxide, grade 1). Radioactivity was estimated by liquid scintillation spectrometry. Data were corrected for recovery (about 75%).
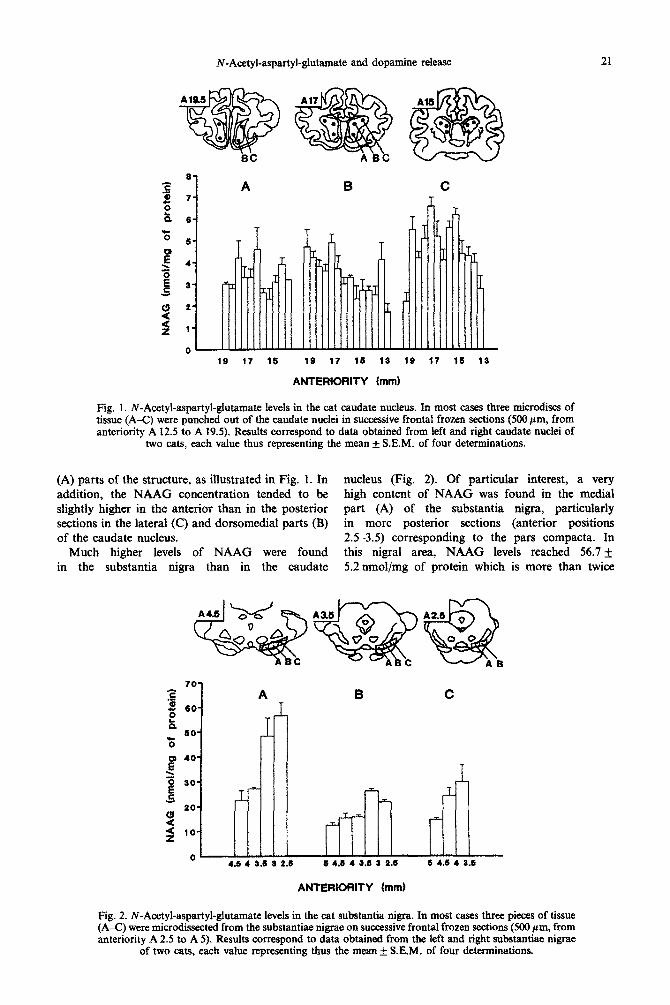
N-Acetyl-aspartyl-glutamate and dopamine release 21
19 17 1s 19 I? 15 13 19 17 15 13
ANTERIORITY (mm)
Fig. 1. ~“A~tyl-as~rtyl-curate levels in the cat caudate nucleus. In most cases three microdiscs of tissue (A-C) were punched out of the caudate nuclei in successive frontal frozen sections (500 pm, from anteriority A 12.5 to A 19.5). Results correspond to data obtained from ieft and right caudate nuclei of
two cats, each value thus representing the mean f S.E.M. of four determinations.
(A) parts of the structure, as illustrated in Fig. 1. In nucleus (Fig. 2). Of particular interest, a very addition, the NAAG con~ntration tended to be high content of NAAG was found in the medial slightly higher in the anterior than in the posterior part (A) of the substantia nigra, p~icularly sections in the lateral (C) and dorsomedial parts (B) in more posterior sections (anterior positions of the caudate nucleus. 2.5-3.5) corresponding to the pars compacta. In
Much higher levels of NAAG were found this nigral area, NAAG levels reached 56.7 It in the substantia nigra than in the caudate 5.2 nmol/mg of protein which is more than twice
70
A 60
60
30
20
10
0 ~ 44 4 3.5 8 2.6 S 4.5 4 3.6 3 2-S s 46 4 3.S
ANTERIORITY (mm)
Fig. 2. N-Acetyl-aspartyl-glutamate levels in the cat substantia nigra. In most cases three pieces of tissue (AC) were microdissected from the substantiae nigrae on successive frontal frozen sections (500 pm, from anteriority A 2.5 to A 5). Results correspond to data obtained from the left and right substantiae nigrae
of two cats, each value representing thus the mean + S.E.M. of four determinations.

22 T. GALLI et al.
the amount estimated in the pars reticulata (B) or lateralis (C).
For comparison, experiments were also made with male Spragu~Dawley rats in both the striatum and the substantia nigra (eight animals). NAAG levels found in the striatum (1.9 + 0.2 nmol/mg protein) as well as in the medial (7.9 &- 0.6 nmol/mg protein) and lateral (6.8 + 0.6 nmol/mg protein) parts of the substantia nigra were in the range of those found by Guarda et al. If Therefore, nigral levels of NAAG appear to be much higher in the cat than in the rat.
Stimulatory effect of N-acetyl-aspartyl-glutamate on [3H]dopamine release in the caudate nucleus: compari- son with ibotenate and N-methyl-D-aspartate
Cats were implanted with a push-pull cannula in each caudate nucleus in either the anterior (At619) or posterior (A13-15.5) parts. Since, as previously shown,’ glutamate stimulates the release of [3H]DA continuously synthesized from [3H]tyrosine in the cat caudate nucleus when used at a concentration as low as 10Y8 M, this low concentration was chosen for experiments using NAAG. In addition, prebminary experiments were made in order to verify that, at low8 M, NAAG hydrolysis into N-acetyl-aspartate and glutamate was limited. When [3H]NAAG (1O-8 M) was applied for 30 min into the caudate nucleus, the radioactivity recovered in the high- pressure liquid chromatography peak of NAAG represented, respectively, 83.0, 98.7, and 99.4% of the total quantity of [3H]NAAG delivered in each successive IO-mm fraction. Since low amounts of [“H]NAAG were still recovered in the two following fractions (15.2 and 1.3%), the total recovery of [HJNAAG was very close to 100%. Furthermore, the radioactivity estimated in the high-pressure liquid chromatography peak of glutamate was very low and represented about 0.24% (about 24 x lo-I’M) of the total amount of rH]NAAG delivered to the tissue. The total amount of glutamate and aspartate in the superfusates was not modi- fied by the presence of f3H]NAAG in the super- fusion fluid, the basal levels of glutamate and aspartate being 0.53 x 10T6 M and 0.23 x 10m6 M, respectively.
NAAG (IO-’ M) markedly stimulated the release of [“HIDA in the posterior but not in the anterior part of the caudate nucleus, as illustrated in Fig. 3. The stimulatory effect of NAAG on [3H]DA release was seen not only during its appli- cation but also thereafter up to the end of the superfusion.
Since it has been suggested that NAAG could act on NMDA receptors,” experiments were also made with NMDA and with ibotenate, which according to some studies may act as NMDA agonists. I3 As previously reported, when used at a concentration of lO--‘M, NMDA alone does not si~ificantly modify the release of f3H]DA in the
cat caudate nucleus.2 However, in the present experiments, a stimulatory effect of NMDA on [3H]DA release was observed when NMDA was applied at 10e4 M but not IO-’ M (data not shown) in the presence of glycine (lO-‘j M). Under these conditions, the NMDA-evoked release of [3H]DA occurred mainly during the drug application and was seen both in the anterior and posterior parts of the caudate nucleus. Ibotenate (lO-s M) alone stimulated the release of [-‘HIDA, but its effect was much more pronounced in the posterior than in the anterior part of the caudate nucleus (Fig. 3). Previously, we have reported that NMDA (10e5 M) alone increases markedly the efflux of both glutamate and aspartate in the cat caudate nucleus2 In contrast, in the present study, ibotenate (lOms M) did not signifi~ntly affect the efflux of these excitatory amino acids (data not shown).
Inhibitory effect of N-acetyl-aspartyl-glutamate on [3H]dopamine release in the presence of tetrodotoxin in the caudate nucleus: comparison with ibotenate and N-methyl-D-~partate
Previousfy, we have shown that the stimulatory effect of glutamate (1O-8 M) on [3H]DA release still occurred in the presence of tetrodotoxin8 For comparison, similar experiments were made with NAAG (lo-’ M) and ibotenate (lo-’ M).
As previously reported, the continuous delivery of tetrodotoxin (10m6 M) markedly decreased the spontaneous release of [‘HIDA ( -69%).8 Sur- prisingly, both NAAG (10e8 M) and ibotenate ( 10m5 M) reduced the tetrodotoxin-resistant release of [3H]DA. These effects were seen both during and after each drug appli~tion and were more prominent in the posterior than in the anterior part of the caudate nucleus (Fig. 4). In contrast, in the presence of tetrodotoxin, NMDA (10m4 M, in the absence of Mg2+ and presence of glycine 10w6M) stimulated [3H]DA release in both the anterior and posterior parts of the caudate nucleus (Fig. 4).
Efiects of the unilateral application of N-acetyl- aspartyl -glutamate and of its b isomer in the substantiu nigra on the release of [3H]dopamine from dendrites and nerve terminals of dopaminergic neurons
Cats were implanted with four push-pull cannulae (one in each substantia nigra pars compacta and one in each caudate nucleus) and the release of [3H]DA continuously formed from [3H]tyrosine was estimated in each structure. A slightly higher concentration of NAAG (lo-’ M) was used in these experiments in order to activate, as much as possible, a large population of dopaminergic cells.
The application of NAAG (lo-’ M) into the pars compacta of the left substantia nigra for 30 min induced a stimulation of [‘HIDA release from dopa- minergic dendrites, this effect occurring both during
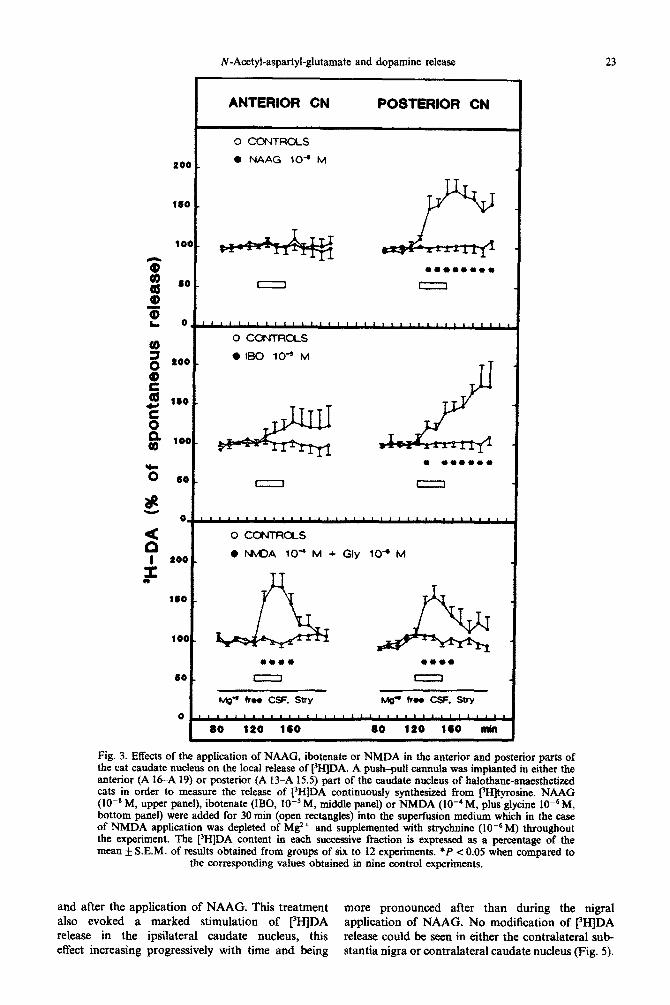
N-Acetyl-aspartyl-glutamate and dopamine release 23
600
160
1OQ
f 8 60
z 0,
3 0 600
z! 0 E 160
zi a 100
‘i3 60
s!! 0.
s I too
T
160
100
60
0
ANTERIOR CN POSTEFNOR CN
0 CONTROLS
0 NAAG lo-’ M
Fig. 3. Effects of the application of NAAG, ibotenate or NMDA in the anterior and posterior parts of the cat cat&ate nucleus on the local release of [3H]DA. A push-pull canmtla was implanted in either the anterior (A 16-A 19) or posterior (A 13-A 15.5) part of the caudate nucleus of ~Ioth~e-~~s~e~ cats in order to measure the release of rI-IjDA continuously synthesized from t3Hjtyrosine. NAAG (lOeN M, upper panel), ibotenate (IBG, 10m5 M, middle panel) or NMDA (lo-‘M, plus glycine 10m6 M, bottom panel) were added for 30 mm (open rectangles) into the super-fusion medium which in the case of NMDA application was depleted of M2+ and supplemented with strychnine (lo-” M) throughout the experiment. The [)H]DA content in each successive fraction is expressed as a percentage of the mean f S.E.M. of results obtained from groups of six to I2 experiments. *P e 0.05 when compared to
the corresponding values obtained in nine control experiments.
and after the application of NAAG. This treatment more pronounced after than during the nigral also evoked a marked stimulation of [3H]DA application of NAAG. No modification of 13H]DA release in the ipsilateral caudate nucleus, this release could he seen in either the contralateral sub- effect increasing progressively with time and being stantia nigra or contralateral caudate nucleus (Fig. 5).
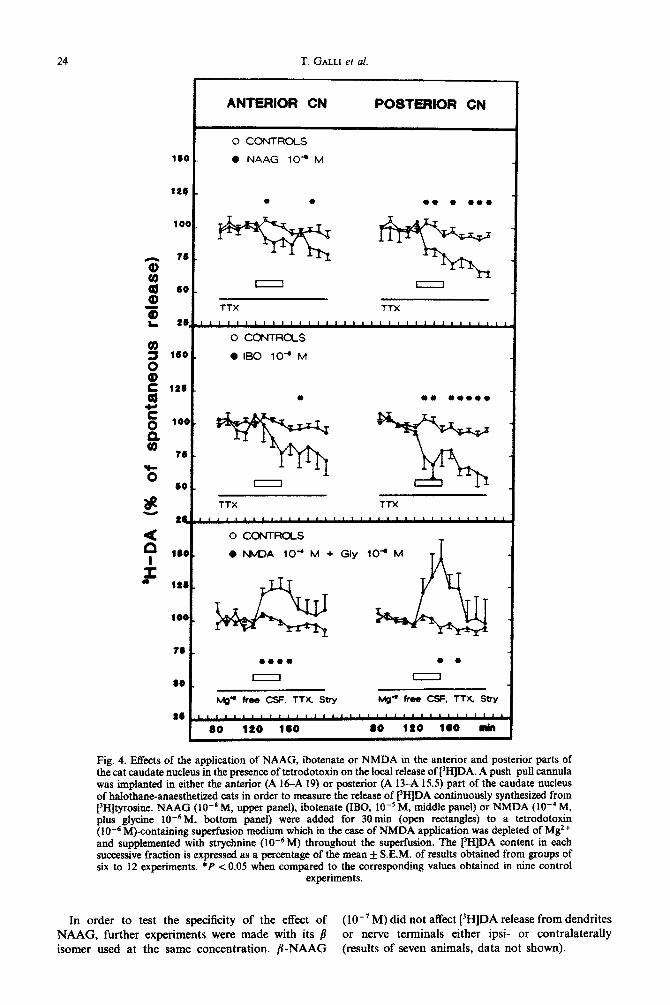
24 T. GALLI et al.
160
f
76
kd 66 a3
v 66
$ 160
$
s 126
E
B
100
160
166
60
Fig. 4. Effects of the application of NAAG, ibotenate or NMDA in the anterior and posterior parts of the cat caudate nucleus in the presence of tetrodotoxin on the local release of [“HIDA. A push-pull cannula was implanted in either the anterior (A l&A 19) or posterior (A 13-A 15.5) part of the caudate nucleus of halothane-anaesthetized cats in order to measure the release of [3wDA continuously synthesized from [3H]tyrosine. NAAG (lOPB M, upper panel), ibotenate (IBO, 10-r M, middle panel) or NMDA (IO-’ M, plus gIycine IOe6M, bottom panel) were added for 30min (open rectangles) to a tetrodotoxin ( 10e6 M)~n~ining superfusion medium which in the case of NMDA application was depleted of Mg2 + and supplemented with StiphAiAe (10e6M) throughout the superfusion. The t31-IlDA content in each successive fraction is expressed as a percentage of the mean & S.E.M. of results obtained from groups of six to 12 experiments. *P co.05 when compared to the corresponding values obtained in nine control
experiments.
In order to test the specificity of the effect of (lo-‘M) did not affect [‘HIDA release from dendrites NAAG, further experiments were made with its @ or nerve terminals either ipsi- or contralaterally isomer used at the same concentration. p-NAAG (results of seven animals, data not shown).
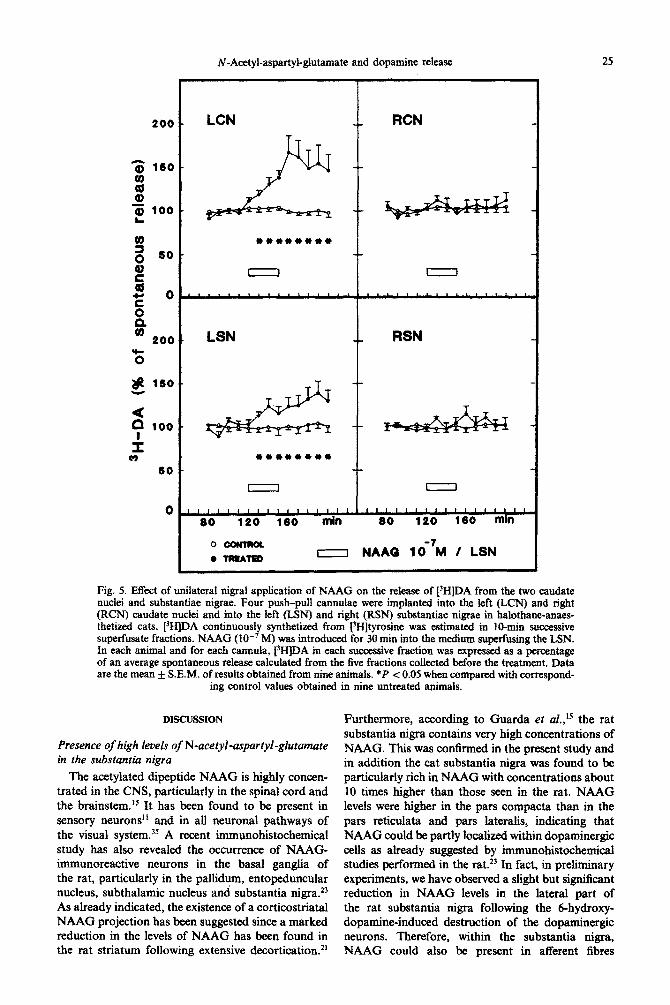
~-A~tyl-asp~yl-~u~mate and dopamine release 2s
Fig. 5. Effect of unilateral nigral application of NAAG on the release of [‘HIDA from the two caudate nuclei and substantiae nigrae. Four push-pull cannulae were implanted into the left (LCN) and right (RCN) caudate nuclei and into the left (LSN) and right (RSN) substantiae nigrae in halothane-anaes- thetixed cats. PH]DA ~ntinuously synthetized from PEfltyrosine was estimated in 10&n successive superfusate fractions. NAAG (IO-’ M) was introduced for 30 min into the medium sugaring the LSN. in each animal and for each cannula, PHJDA in each successive fraction was expressed as a percentage of an average spontaneous release calculated from the five fractions collected before the treatment. Data are the mean _L S.E.M. of results obtained from nine animals. *P < 0.05 when compared with correspond-
ing control values obtained in nine untreated animals.
DISCUSSION
Presence of high levels of N-acetyl-mpartyl-glutamate in the substantia nigra
The acetylated dipeptide NAAG is highly concen- trated in the CNS, particularly in the spinal cord and the brainstem.Iz It has been found to be present in sensory neurons” and in ail neuronal pathways of the visual system. 32 A recent i~unohist~he~~l study has also revealed the occurrence of NAAG- immunoreactive neurons in the basal ganglia of the rat, particularly in the pallidum, entopeduncular nucleus, subthalamic nucleus and substantia nigra.” As already indicated, the existence of a corticostriatal NAAG projection has been suggested since a marked reduction in the levels of NAAG has been found in the rat striatum following extensive decortication.21
Furthermore, according to Guarda et al.,15 the rat substantia nigra contains very high concentrations of NAAG. This was confirmed in the present study and in addition the cat substantia nigra was found to be particularly rich in NAAG with concentrations about 10 times higher than those seen in the rat. NAAG levels were higher in the pars compacta than in the pars reticulata and pars lateralis, indicating that NAAG could be partly localized within dopaminergic cells as already suggested by immunohistochemical studies performed in the rat.23 In fact, in preliminary experiments, we have observed a slight but significant reduction in NAAG levels in the lateral part of the rat substantia nigra following the 6-hydroxy dopamine-induced destruction of the dopaminergic neurons. Therefore, within the substantia nigra, NAAG could also be present in afferent fibres

26 T. GALLI et at.
ori~nating from structures whose identity has yet to be determined.
Specificity of the effects of N-acetyf-aspartyl-gluta- mate on [‘Hldopamine release from dendrites and nerve terminals of the dopaminergic neurons
Several groups have reported that NAAG is released either in vitro2S,3’,39 or in vivo 36 upon depolar- ization through a calcium-dependent process and that it is inactivated by a specific enzyme: i.e. N-acetylated g-linked acidic dipeptidase.“s3* Electrophysiological studies have indicated that NAAG modifies the activity of various populations of cells in the CNS. ‘“~“~‘8~~ All these observations are in favour of a transmitter or modulatory role of NAAG. However, the specificity of the electrophysiological responses can be questioned since NAAG is rapidly hydrolysed to N-acetyl-aspartate and glutamate which could be responsible for its excitatory action. Moreover, when used in high concentrations, as it is often the case, NAAG may act as a chelating agent of cations such as magnesium.
Since nonspecific effects of NAAG have been demonstrated under some conditions, Whittemore ef aL3* have even suggested that “NAAG may not exhibit intrinsic activity as an excitant in the CNS”. Nevertheless our results provide further evidence for a specific role of NAAG. Indeed, as verified by experiments made with rH]NAAG, at low concen- trations (lo-‘, lo-* M) and in conditions in which its hydrolysis is limited or negligible, NAAG was shown to modulate the release of [‘HIDA from dendrites and nerve terminals of dopaminergic neurons when applied in vivo in either the pars compacta or the caudate nucleus, respectively. The specificity of the effect of NAAG was further demonstrated since no change in i3H]DA release was observed when the /I isomer of NAAG was used.
E#ect of N-a~etyl-~partyl~glutamate on do~min- ergic neurons in the substantia nigra
As already discussed, within the substantia nigra, NAAG could be released from either afferent fibres and/or from dopaminergic dendrites taking into account the possible co-localization of DA and NAAG in dopaminergic cells. Our results suggest that NAAG modulates the activity of dopaminergic cells either directly or indirectly since a prolonged stimulation of [‘HJDA release was seen in the ipsilateral caudate nucleus when NAAG (IO-’ M) was applied for 30 min into the substantia nigra pars compacta. In addition, this effect was associated with a sustained stimulation of rH]DA release from dopaminergic dendrites. These responses are similar to those induced by both cholecystokinin-8-sulphate and neurokinin-A which similarly stimulated the release of DA from dopaminergic dendrites and nerve terminals when applied in low concentrations into the substantia nigra. ‘,4 This is of particular interest since cholecystokinin-8-sulphate and neurokinin-A which,
in the cat, are contained in dopaminergic dendrites and in striatonigral afferent fibres, respectively, have been shown to stimulate the activity of nigral dopaminergic cell~.‘~~~~~’
Direct and indirect presynaptic regulation by N-acetyl- aspartyl-glutamate of dopamine release in the caudate nucleus
NAAG interacts not only with dopaminergic neurons in the substantia nigra but also with dopa- minergic fibres in the caudate nucleus. Indeed, as shown by the effects of local application of NAAG into the caudate nucleus, NAAG seems to be involved in the presynaptic regulation of DA release by acting both through direct (tetrodotoxin-resistant) and indirect (tetrodotoxin-sensitive) processes.
As does glutamate, NAAG modulates the release of [3H]DA when used at a low concentration. How- ever, NAAG seems to act through receptors which are distinct from those of the AMPA or NMDA subtypes. Finally, since the responses evoked by ibotenate were closely similar to those induced by NAAG, ibotenate could act as an agonist at NAAG- sensitive receptors.
As shown previously, when applied locally at 10e8 M into the caudate nucleus, glutamate stimu- lates the release of 13H]DA both in the presence and the absence of tetrodotoxin.’ In addition, the tetrodo- toxin-resistant stimulatory effect of glutamate was reproduced by kainate. The glutamate or kainate- evoked responses were prevented by co-adminis- tration of quisqualate which rapidly desensitizes glutamate receptors of the AMPA subtype.3 In contrast to glutamate, at a concentration of lo-’ M, NAAG does not seem to act on receptors of the AMPA subtype. Indeed, although NAAG stimulated the release of [3H]DA in the absence of tetrodotoxin, it did not enhance but instead reduced the release of t3H]DA in the presence of the neurotoxin.
Studies made on rat striatat slices have indicated that in the absence of magnesium, NMDA stimulates the release of DA; this effect is potentiated by glycine through a strychnine-insensitive site22*27 indicating that glycine plays a critical role in the efficacy of the NMDA-mediated response. In these conditions, the NMDA-evoked response persists in the presence of tetrodotoxin and can even be demonstrated on purified synaptosomes indicating that NMDA recep- tors are located on dopaminergic nerve termi- na1s.9+22,28*” Our results, in vivo, confirm that in high concentration (10e4 M), NMDA stimulates markedly 13H]DA release in both the absence and presence of tetrodotoxin when experiments are performed in the absence of magnesium and the presence of glycine. An effect of NAAG on NMDA receptors can be excluded for several reasons. (1) The stimulatory effect of NAAG on [3H]DA release was seen with a low concentration (lo-* M) of the N-acetylated dipeptide in the presence of magnesium which has been shown to inhibit the NMDA-evoked response.

N-Acetyl-aspartyl-glutamate and dopamine release 27
(2) As already emphasized, in contrast to NMDA, NAAG did not stimulate but instead reduced the tetrodotoxin-resistant release of [‘HJDA. (3) The stimulatory effect of NAAG on [‘HIDA release was seen exclusively in the posterior part of the caudate nucleus while NMDA-evoked responses of similar amplitude occurred in both the anterior and posterior parts of the structure. (4) The response evoked by NAAG was of long duration while those induced by NMDA were of short duration. (5) Interestingly, NAAG (IO-* M) did not significantly increase the efflux of glutamate or aspartate in contrast to that observed with NMDA (lo-’ M) alone as reported previously.*
According to electrophysiological and binding studies, ibotenate may act as an agonist of NMDA receptors. I3 However, in our study, at the concen- tration used, ibotenate modulates presynaptically the release of 13HjDA by m~hanisms distinct from those involved in the NMDA-evoke responses. In fact, ibotenate could act on NAAG-sensitive receptors since, like NAAG, it induced opposite effects on [‘H]DA release in the absence and presence of tetrodotoxin. Moreover, both the tetrodotoxin-sensi- tive stimulatory and tetrodotoxin-resistant inhibitory effects of ibotenate were much more pronounced in the posterior than in the anterior part of the caudate nucleus. Finally, as NAAG, ibotenate did not modify the efflux of either glutamate or aspartate.
To our knowledge, NAAG and ibotenate presently are the only known compounds which have the capacity to reduce si~ifi~ntly the in viva tetrodo- torn-~sistant spontaneous release of CJmDA. Further studies are required to characterize the recep-
tors involved in this process. Similarly, the identity of the local neuronal circuit responsible for the tetrodotoxin-sensitive stimulatory effects of NAAG and ibotenate remains to be elucidated. Since endogeneous levels of NAAG were slightly higher in the anterior than in the posterior part of the caudate nucleus, it was rather surprising to see that both the tetrodotoxin-~ist~t and te~odotoxin-~nsitive effects of NAAG and ibotenate were much more prominent in the posterior than in the anterior part of the structure. Whatever the mechanisms involved, this demonstrates that the direct and/or indirect presynaptic regulation of DA release may be different in various parts of the cat caudate nucleus. This is in agreement with recent in vitro studies of our laboratory indicating differences in the presynaptic regulation of DA release in striosomal and matrix compartments,‘9
CONCLUSION
Our study indicates that NAAG interacts in several ways with nigrostriatal dopaminergic neurons by inducing changes in DA release from dendrites and nerve terminals and provides further strong evidence for a transmitter or modulatory role of NAAG in the CNS.
Acknowledgements-This study was supported by INSERM, Rhone-Poulenc Sante and a grant from the CEE (CI 1-0330-F). A.C. is a research fellow of RhSne-Poulenc Sante. The authors would like to thank Dr M. L. Kernel who kindly contributed to some ex~~ments. The excel- lent technical assistance of A. M. Godeheu is gratefully acknowledged.
REFERENCES
1. Artaud F., Baruch P., Stutzmann J. M., Saffroy M., Godeheu G., Barbeito L., HervC D., Studier J. M., Glowinski J. and Cheramy A. (1989) Chole-cystokinin: co-release with dopamine from nigro-striatal neurones in the cat. Eur. J. Neurosci. 1, 162-l 7 1.
2. Barbeito L., Cheramy A., Godeheu G., Desce J. M. and Glowinski J. (1990) Glutamate receptors of a quisqualate/kainate subtype are involved in the presynaptic regulation of dopamine release in the cat caudate nucleus in viva. Eur. J. Neurosci. 2, 304-31 I.
3. Barbeito L., Girault J. A., Godeheu G., Pittaluga A., Glowinski J. and Cheramy A. (1989) Activation of the bilateral corticostriatal glutamate@ projection by infusion of GABA into thalamic motor nuclei in the cat: an in t&o release study. Neuroscience 28, 365-374.
4. Baruch P., Artaud F., Godeheu G., Barbeito L., Glowinski J. and Cheramy A. (1988) Substance P and neurokinin A contribute by different mechanisms to the regulation of dopamine release from dendrites and nerve terminals of the nigrostriatal dopaminergie neurons. Neuroscience U, 889-898.
5. Blakely R. D. and Coyle J. T. (1988) The neurobiology of N-acetylaspartylglutamate. Znt. Rat. Neurobiof. 30,39-100. 6. Bradford M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the
principle of protein dye binding. Anolyt. Biochem. 7, 248-254. 7. Cheramy A., Leviel V., Daudet F., Guibert B., Chesselet M. F. and Glowinski J. (1981) Involvement of the thalamus
in the reciprocal regulation of the two nigro-striatal dopaminergic pathways. Neuroscience 6, 2657-2668. 8. Cheramy A., Romo R., Godeheu G., Baruch P. and Glowinski J. (1986) In viuo presynaptic control of dopamine release
in the cat caudate nucleus. II. Facilitatory or inhibitory influence of L-glutamate. Neuroscience 19, 1081-1090. 9. Clow D. W. and Jhamandas K. (1989) Characterization of L-glutamate action on the release of endogenous dopamine
from rat caudateputamen. J. Pharmac. exp. Ther. 248, 722-728. 10. Curatolo A., Marchetti M., Brancati A. and Sallco A. (1967) Aziome degli acidi N-ace&aspartico e N-acetil-aspartil-
glutamico sull’activita elettrica spontanea della cortecchia cerebrale di gatto. Arch. Ski. Biol. 51, 98403. li. ffrench-Mullen J. M. H., KolIer K., Zacxek R., Coyle J. T., Hori N. and Carpenter D. 0. (1985) N-A&y1 aspartyl
glutamate: possible role as the neurot~nsmitter of the lateral olfactory tract. Proc. nutn. Acud. Sci. U.S.A. 82, 3897-3900.
12. Fonnum F., Storm-Mathisen J. and Divac I. (1981) Biochemical evidence for glutamate as neurotransmitter in corticostriatal and corticothalamic fibers in rat brain. Neuroscience 6. 863-873.

28 T. GALLI et al.
13. Foster A. C. and Fagg G. (1984) Acidic amino acid binding sites in mammalian neuronal membranes: their characteristics and relationship to synaptic receptors. Brain Res. Rev. 7, 103-164.
14. Giorguieff M. F., Kernel M. L. and Glowinski J. (1977) Presynaptic effect of L-glutamic acid on the release of dopamine in rat striatal slices. Neurosci. Lett. 6, 73-77.
15. Guarda A., Ory-Lavollee L., Robinson M. B., Forloni G., Blakely R. D. and Coyle J. T. (1988) Quantitation of N-acetyl-aspartyl-glutamate in microdissected rat brain nuclei and peripheral tissues: findings with a novel liquid phase radioimmunoassay. Molec. Bruin Rex 3, 223-232.
16. Innis R. B., Andrade R. and Aghajanian G. K. (1985) Substance K excites dopaminergic and non dopaminergic neurons in rat substantia nigra. Bruin Res. 335, 381-383.
17. Jasper H. H. and Ajmone-Marsan C. (1954) A Stereotaxic Atlas of the Diencephalon of the Cat. The National Research Council of Canada, Ottawa.
18. J&ls M., Van Veldhuizen M., Urban I. J. A. and De Kloet E. R. (1987) N-Acetyl-aspartyl-glutamate: binding sites and excitatory action in the dorsal septum of the rat. Brain Res. 403, 192-197.
19. Kernel M. L., Desban M., Glowinski J. and Gauchy C. (1989) Distinct presynaptic control of dopamine release in striosomal and matrix areas of the cat caudate nucleus. Proc. nam Acad. Sci. U.S.A. 86, 9006-9010.
20. Kim J. S., Hassler R., Haug P. and Paik K. S. (1977) Effect of frontal cortex ablation on striatal glutamic acid levels in the rat. Brain Res. 132, 370-374.
21. Koller K. J., Zaczek R. and Coyle J. T. (1984) N-Acetyl-aspartyl-glutamate: regional levels in rat brain and the effects of brain lesions as determined by a new HPLC method. J. Neurochem. 43, 11361142.
22. Krebs M. O., Desce J. M., Kernel M. L., Gauchy C., Godeheu G., Chbramy A. and Glowinski J. (1991) Glutamatergic control of dopamine release in the rat striatum: evidence for presynaptic NMDA receptors on dopaminergic nerve terminals. J. Neurochem. 56, 8185.
23. Moffet J. R., Cassidy M. and Namboodiri M. A. A. (1989) Selective distribution of N-acetyl aspartyl glutamate immunoreactivity in the extrapyramidal system of the rat. Brain Res. 494, 255-266.
24. Mori-Okamoto J., Okamoto K. and Sekiguchi M. (1987) Electrophysiological and pharmacological actions of N-acetylaspartylghrtamate intracellularly studied in cultured. chick cerebellar neurons. B&n Res. 401, 60-67.
25. Pittaluza A.. Barbeito L.. Serval V.. Godeheu G.. Artaud F.. Glowinski J. and Chbramv A. (1988) Denolarization- evoked-release of N-acetyl-L-aspartyl-L-glutamate from rat brain synaptosomes. Eur. J.~Pharmac. ‘158,.263-266.
26. Puttfarcken P. S. and Coyle J. T. (1990) N-Acetylaspartylglutamate (NAAG): a possible modulator of [‘H]MKSOl binding in rat hippocampal membranes. Neurosci. Absrr. 15, 372411.
27. Ransom R. W. and Deschenes N. L. (1989) Glycine modulation of NMDA-evoked release of [3H]acetylcholine and [)H]dopamine from rat striatal slices. Neurosci. Lert. 96, 323-328.
28. Roberts P. J. and Anderson S. D. (1979) Stimulatory effect of L-glutamate and related amino acids on [3H]dopamine release from rat striatum: an in vitro model for glutamate actions. J. Neurochem. 32, 1539-1545.
29. Robinson M. B., Blakely R. D. and Coyle J. T. (1986) Quisqualate selectively inhibits a brain peptidase which cleaves N-acetyl-L-aspartyl-L-glutamate in vitro. Eur. J. Pharmuc. 130, 345-347.
30. Sekiguchi M., Okamoto K. and Sakai Y. (1987) Excitatory action of N-acetyl-aspartyl-glutamate on Purkinje cells in guinea pig cerebellar slices: an intrasomatic study. Brain Res. 423, 23-33.
31. Sekiguchi M., Okamoto K. and Sakai Y. (1989) Release of endogeneous N-acetyl-aspartyl-glutamate (NAAG) and uptake of [rH]NAAG in guinea pig slices. Brain Res. 482, 78-86.
32. Serval V., Barbeito L., Pittaluga A., ChCramy A., Lavielle S. and Glowinski J. (19%) Competitive inhibition of N-acetylated-alpha linked-acidic-dipeptidase activity by N-acetyl-aspartyl beta linked-glutamate. J. Neurochem. 55, 3946.
33. Skirboll L. R., Grace A. A., Hommer D. W., Rehfeld J., Goldstein M., Hiikfelt T. and Bunney B. S. (1981) Peptide monoamine coexistence: studies of the actions of cholecystokinin-like peptide on the electrical activity of midbrain dopamine neurons. Neuroscience 6, 2111-2124.
34. Snell L. D. and Johnson K. M. (1987) Characterization of the inhibition of excitatory amino acid-induced neurotransmitter release in the rat striatum by phencyclidine-like drugs. J. Pharmac. exp. Ther. 238, 938-945.
35. Tieman S. B., Cangro C. B. and Neale J. H. (1987) N-Acetyl-aspartylglutamate immunoreactivity in neurons of the cat’s visual system. Brain Res. 420, 188-193.
36. Tsai’ G., Forloni G., Robinson M. B., Stauch B. L. and Coyle J. T. (1988) Calcium-dependent evoked release of N-[3H]-acetyl-aspartyl-ghttamate from the optic pathway. J. Neurochem. 51, 19561959.
37. Westbrook G. L., Mayer M. L., Namboodiri M. A. A. and Neale J. H. (1986) High concentrations of N-acetyl-aspartyl- glutamate (NAAG) se.lectively activate NMDA receptors on mouse spinal cord neurons in cell culture. J. Neurosci. 6, 3385-3392.
38. Whittemore E. R. and Koemer J. F. (1989) An explanation for the purported excitation of piriform cortical neurons by N-acetyl-L-aspartyl-L-glutamic acid (NAAG). Proc. natn. Acud.-&. U.S.A. 86, 9602-9605.
39. Zollinger M., Amsler U., Do K. Q., Streit P. and Cdnod M. (1988) Release of N-acetyl-aspartyl glutamate on depolarization of rat brain slices. J. Neurochem. 5, 19191923.
(Accepted 22 November 1990)
Copyright © 2022 FDOKUMEN




















