
Pyruvate protects cerebellar granular cells from 6-hydroxydopamine-induced cytotoxicity by...
12
Pyruvate protects cerebellar granular cells from 6-hydroxydopamine-induced cytotoxicity by activating the Akt signaling pathway and increasing glutathione peroxidase expression F.J. Fernandez-Gomez, a,e,f M.D. Pastor, a,b,f E.M. Garcia-Martinez, c,e R. Melero-Fernandez de Mera, a,e,f M. Gou-Fabregas, d M. Gomez-Lazaro, a,e,f S. Calvo, a,b,f R.M. Soler, d M.F. Galindo, a,f and J. Jordán a,e,f, ⁎ a Departamento de Ciencias Médicas, Facultad de Medicina, Universidad Castilla-La Mancha, Spain b Grupo Glia e Isquemia, Universidad Castilla-La Mancha, Spain c Servicio de Farmacia, Complejo Hospitalario Universitario de Albacete, Spain d Departament de Ciències Mèdiques Bàsiques, Facultat de Medicin, Universitat de Lleida, Spain e Grupo Neurofarmacología, Universidad de Castilla-La Mancha, Spain f Centro Regional de Investigaciones Biomédicas, Albacete, Spain Received 19 May 2006; revised 7 July 2006; accepted 16 July 2006 Available online 15 September 2006 Parkinson disease (PD) is the second-most common age-related neurodegenerative disease and is characterized by the selective destruction of dopaminergic neurons. Increasing evidence indicates that oxidative stress plays a crucial role in the pathogenesis of idiopathic PD. Anti-oxidant agents including catalase, manganese porphyrin and pyruvate confer cytoprotection to different cell cultures when challenged with 6-hydroxydopamine (6-OHDA). Herein we used rat cerebellar granular cell cultures to ascertain the plausible cellular pathways involved in pyruvate-induced cytoprotection against 0.1 mM 6-OHDA. Pyruvate provided cytoprotection in a concentration-dependent manner (2–10 mM). Consistent with its well-established anti-oxidant capacity, pyruvate (10 mM) prevented 6-OHDA-induced lipid peroxidation by blocking the rise in intracellular peroxides and maintaining the intracellular reduced glutathione (GSH) levels. Further experiments revealed that pyruvate increased Akt, but not extracellular signal- regulated kinase phosphorylation. Moreover, phosphatidylinositol 3- kinase (PI3K) inhibitors attenuated pyruvate-induced cytoprotection indicating that PI3K-mediated Akt activation is necessary for pyruvate to induce cytoprotection. On the other hand, pyruvate also up-regulated glutathione peroxidase mRNA levels, but not those of the anti-oxidant enzymes superoxide dismutase-1 and -2, catalase or the anti-apoptotic oncogenes Bcl-2 or Bcl-x L . In summary, our results strongly suggest that pyruvate, besides the anti-oxidant properties related to its struc- ture, exerts cytoprotective actions by activating different anti-apoptotic routes that include gene regulation and Akt pathway activation. © 2006 Elsevier Inc. All rights reserved. Parkinson’ s disease (PD) is a neurodegenerative disorder characterized by a progressive loss of dopaminergic neurons in the substantia nigra pars compacta (Noelker et al., 2005), with the resulting loss of nerve terminals accompanied by a dopamine (DA) deficiency in the striatum (Hornykiewicz, 1966). 6- Hydroxydopamine (6-OHDA), an endogenously generated meta- bolite of dopamine oxidation (Fornstedt et al., 1986), is extensively utilized in experimental models of PD because it causes cell death in different cell types, including human neuroblastoma SK-N-SH (Shimizu et al., 2002), SH-SY5Y (Von Coelln et al., 2001; Jordan et al., 2004) and pheochromocytoma PC12 cells (Nie et al., 2002), besides of being a specific neurotoxin of dopaminergic neurons in vivo (Noelker et al., 2005; Blum et al., 2001). Indeed, if reactive oxygen species (ROS) are not properly detoxified, they can damage cell lipids, proteins or DNA, impairing normal cell function. Uncoupling of mitochon- drial oxidative phosphorylation resulting in energy deprivation and by-products resulting from 6-OHDA auto-oxidation such as quinones and hydrogen peroxide (H 2 O 2 ) are involved in these cytotoxic processes (Galindo et al., 2003; Thakar and Hassan, 1988). Interestingly, both mitochondrial dysfunction and oxida- tive stress appear to play a central role in the pathogenesis of PD (Beal, 2003; Blum et al., 2001; Mazzio et al., 2004). In fact, anti- oxidant agents, such as catalase, vitamin E, N-acetyl cysteine, ascorbic acid and pyruvate are known to elicit cytoprotection against 6-OHDA in experimental models (Jordan et al., 2004; Galindo et al., 2003; Lai and Yu, 1997). Alpha-ketoacid structures like pyruvate have been shown to confer cytoprotection against different noxious stimuli including transient forebrain ischemia, hemorrhagic shock, β-amyloid, exogenous H 2 O 2 and menadione (2-methyl-1,4-naphthoquinone) www.elsevier.com/locate/ynbdi Neurobiology of Disease 24 (2006) 296 – 307 ⁎ Corresponding author. Grupo de Neurofarmacología, Facultad de Medicina, Universidad de Castilla-La Mancha-CRIB, Avda. Almansa, 14, 02006 Albacete, Spain. Fax: +34 967599327. E-mail address: [email protected] (J. Jordan). Available online on ScienceDirect (www.sciencedirect.com). 0969-9961/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.nbd.2006.07.005
Transcript of Pyruvate protects cerebellar granular cells from 6-hydroxydopamine-induced cytotoxicity by...

www.elsevier.com/locate/ynbdi
Neurobiology of Disease 24 (2006) 296–307Pyruvate protects cerebellar granular cells from6-hydroxydopamine-induced cytotoxicity by activating the Aktsignaling pathway and increasing glutathione peroxidase expression
F.J. Fernandez-Gomez,a,e,f M.D. Pastor,a,b,f E.M. Garcia-Martinez,c,e
R. Melero-Fernandez de Mera,a,e,f M. Gou-Fabregas,d M. Gomez-Lazaro,a,e,f
S. Calvo,a,b,f R.M. Soler,d M.F. Galindo,a,f and J. Jordána,e,f,⁎
aDepartamento de Ciencias Médicas, Facultad de Medicina, Universidad Castilla-La Mancha, SpainbGrupo Glia e Isquemia, Universidad Castilla-La Mancha, SpaincServicio de Farmacia, Complejo Hospitalario Universitario de Albacete, SpaindDepartament de Ciències Mèdiques Bàsiques, Facultat de Medicin, Universitat de Lleida, SpaineGrupo Neurofarmacología, Universidad de Castilla-La Mancha, SpainfCentro Regional de Investigaciones Biomédicas, Albacete, Spain
Received 19 May 2006; revised 7 July 2006; accepted 16 July 2006Available online 15 September 2006
Parkinson disease (PD) is the second-most common age-relatedneurodegenerative disease and is characterized by the selectivedestruction of dopaminergic neurons. Increasing evidence indicatesthat oxidative stress plays a crucial role in the pathogenesis of idiopathicPD. Anti-oxidant agents including catalase, manganese porphyrin andpyruvate confer cytoprotection to different cell cultures when challengedwith 6-hydroxydopamine (6-OHDA). Herein we used rat cerebellargranular cell cultures to ascertain the plausible cellular pathwaysinvolved in pyruvate-induced cytoprotection against 0.1 mM 6-OHDA.Pyruvate provided cytoprotection in a concentration-dependentmanner(2–10 mM). Consistent with its well-established anti-oxidant capacity,pyruvate (10 mM) prevented 6-OHDA-induced lipid peroxidation byblocking the rise in intracellular peroxides and maintaining theintracellular reduced glutathione (GSH) levels. Further experimentsrevealed that pyruvate increased Akt, but not extracellular signal-regulated kinase phosphorylation. Moreover, phosphatidylinositol 3-kinase (PI3K) inhibitors attenuated pyruvate-induced cytoprotectionindicating that PI3K-mediated Akt activation is necessary for pyruvateto induce cytoprotection. On the other hand, pyruvate also up-regulatedglutathione peroxidase mRNA levels, but not those of the anti-oxidantenzymes superoxide dismutase-1 and -2, catalase or the anti-apoptoticoncogenes Bcl-2 or Bcl-xL. In summary, our results strongly suggestthat pyruvate, besides the anti-oxidant properties related to its struc-ture, exerts cytoprotective actions by activating different anti-apoptoticroutes that include gene regulation and Akt pathway activation.© 2006 Elsevier Inc. All rights reserved.
⁎ Corresponding author. Grupo de Neurofarmacología, Facultad deMedicina, Universidad de Castilla-La Mancha-CRIB, Avda. Almansa, 14,02006 Albacete, Spain. Fax: +34 967599327.
E-mail address: [email protected] (J. Jordan).Available online on ScienceDirect (www.sciencedirect.com).
0969-9961/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.nbd.2006.07.005
Parkinson’s disease (PD) is a neurodegenerative disordercharacterized by a progressive loss of dopaminergic neurons inthe substantia nigra pars compacta (Noelker et al., 2005), with theresulting loss of nerve terminals accompanied by a dopamine(DA) deficiency in the striatum (Hornykiewicz, 1966). 6-Hydroxydopamine (6-OHDA), an endogenously generated meta-bolite of dopamine oxidation (Fornstedt et al., 1986), isextensively utilized in experimental models of PD because itcauses cell death in different cell types, including humanneuroblastoma SK-N-SH (Shimizu et al., 2002), SH-SY5Y (VonCoelln et al., 2001; Jordan et al., 2004) and pheochromocytomaPC12 cells (Nie et al., 2002), besides of being a specificneurotoxin of dopaminergic neurons in vivo (Noelker et al., 2005;Blum et al., 2001). Indeed, if reactive oxygen species (ROS) arenot properly detoxified, they can damage cell lipids, proteins orDNA, impairing normal cell function. Uncoupling of mitochon-drial oxidative phosphorylation resulting in energy deprivationand by-products resulting from 6-OHDA auto-oxidation such asquinones and hydrogen peroxide (H2O2) are involved in thesecytotoxic processes (Galindo et al., 2003; Thakar and Hassan,1988). Interestingly, both mitochondrial dysfunction and oxida-tive stress appear to play a central role in the pathogenesis of PD(Beal, 2003; Blum et al., 2001; Mazzio et al., 2004). In fact, anti-oxidant agents, such as catalase, vitamin E, N-acetyl cysteine,ascorbic acid and pyruvate are known to elicit cytoprotectionagainst 6-OHDA in experimental models (Jordan et al., 2004;Galindo et al., 2003; Lai and Yu, 1997).
Alpha-ketoacid structures like pyruvate have been shown toconfer cytoprotection against different noxious stimuli includingtransient forebrain ischemia, hemorrhagic shock, β-amyloid,exogenous H2O2 and menadione (2-methyl-1,4-naphthoquinone)

Table 1
Sequences of the oligonucleotide primer pairs used for real-time PCR
SOD1-F 5′-TGCAGGGCGTCATTCACTT-3′SOD1-R 5′-CACAACTGGTTCACCGCTT-3′SOD2-F 5′-TTAACGCGCAGATCATGCA-3′SOD2-R 5′-GTAGGTCGCGTGGTGCTT-3′CATALASE-F 5′-CACACCTACGTACAGGCCG-3′CATALASE-R 5′-TTAGCTTTTCCCTTGGCAGC-3′Gpx1-F 5′-GCAGATACACCAGGCGCTTT-3′Gpx1-R 5′-GGCTTCTATATCGGGTTCGA-3′GAPDH-F 5′-CCAGCCTCGTCTCATAGACA-3′GAPDH-R 5′-GTAACCAGGCGTCCGATACG-3′RBCL-XL-F 5′-CTGCCTTGTTGGTGGGAC-3′RBCL-XL-R 5′-AAAGCATTCCCGAGAGGCT-3′BCL2-F 5′-ACGGTGGTGGAGGAACTCTT-3′BCL2-R 5′-CACAATCCTCCCCCAGTTC-3′
297F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
(Thor et al., 1982; Doroshow et al., 1990; Alvarez et al., 2003;Salahudeen et al., 1991; Slovin et al., 2001; Lee et al., 2001). Theunderlying mechanism of this cytoprotective effect is notcompletely understood. Pyruvate is considered a cytoprotectiveagent in experiments involving an attenuation of metabolicdysfunction and situations in which cellular glucose oxidation isrequired (Chung et al., 2004). In this manner, pyruvate seems to bethe critical component to explain the trophic activity of glia-conditioned media for central nervous system neurons (Selak et al.,1985). On the other hand, pyruvate might act as a ROS scavengerdue to its anti-oxidant capacity. For example, pyruvate is able toundergo nonenzymatic decarboxylation in the presence of H2O2
and thus prevent •OH formation by the so-called Fenton reaction(Alvarez et al., 2003).
Anti-oxidant systems and intracellular signaling pathways,including phosphatidylinositol 3-kinase (PI3K) and extracellularsignal-regulated kinase (ERK), have been shown to be importantin the regulation of cell survival. Superoxide dismutase, catalase,glutathione peroxidase and non-enzymatic molecules such asvitamin E, glutathione and alpha-ketoacid compounds are anti-oxidant systems that neutralize and eliminate ROS and confercytoprotection versus 6-OHDA-induced cell death (Jordan et al.,2004; Love et al., 2002; Lai et al., 1997). The PI3K pathway hasemerged as one of the critical factors in anti-apoptotical signaltransduction (Song et al., 2005; D’Mello et al., 1997; Franke etal., 2003). The activation of this pathway is known to protectcells from several apoptotical stimuli (Kennedy et al., 1997). TheERK (p42/p44 mitogen-activated protein kinase; MAPK) cascadeis a central pathway that transmits signals from manyextracellular agents to regulate cellular processes. It is thoughtto play a pivotal role in the integration and transmission oftransmembrane signals required for growth and differentiation.ERK activation is essential for cell growth and plays a crucialrole in apoptosis inhibition (Yoon and Seger, 2006; Pages et al.,1993).
Here we analyzed the possible mechanisms underlying thecytoprotective effects of pyruvate on 6-OHDA-induced toxicity ingranular cell cultures from rat cerebellum, a region wherestructural reorganization, alterations in neuronal activity andarchitecture were detectable following unilateral striatal 6-OHDAlesion (Perovic et al., 2005). We have found that pyruvate blocks6-OHDA-induced toxicity in a concentration-dependent mannerwithout affecting the auto-oxidation process of 6-OHDA. Furtherexperiments revealed that pyruvate prevented the 6-OHDA-induced, intracellular peroxide rise and blocked the depletion ofGSH levels. The activation of the Akt pathway and the increase inthe expression of glutathione peroxidase also participated in thecytoprotective mechanism activated by this alpha-ketoacid.Finally, the metabolic product, malate, did not protect cellcultures against 6-OHDA toxicity and failed to activate theabovementioned cytoprotective pathways.
Materials and methods
Cell culture
Primary cultures of cerebellar granular neurons were obtainedfrom dissociated cerebella of 7- to 8-day-old rats (Fernandez-Gomez et al., 2005a). Dissection and dissociation were carried outin Basal Medium Eagle (BME; Life Technology). Tissues wereincubated with trypsin for 20 min at 37°C and dissociated by
trituration in a medium containing DNase and trypsin. Cells wereplated on 96 plastic well dishes or on 60-mm plastic Petri dishespre-coated with poly-L-lysine (10 g/ml) at a concentration of8×106 cells/ml in BME containing 25 mM KCl, 10% de-complemented fetal calf serum (FCS; Life Technology), glutamineand antibiotics. Cytosine-β-D-arabino-furanoside (Ara-C) (10 μM)was added at 3 days in vitro (DIV) to prevent the growth of non-neuronal cells. All experiments were carried out after 7 days inculture.
Intracellular generation of reactive oxygen species
We used the oxidation-sensitive fluorescent dye 2′,7′-dichlor-odihydrofluorescein diacetate (DCFH-DA) to measure the produc-tion of reactive oxygen species (ROS), mainly hydrogen peroxideand hydroxyl radicals. DCFH-DA is deacetylated by esterases todichlorofluorescein (DCFH). This non-fluorescent product is thenconverted by reactive species into DCF, which can easily bevisualized by strong fluorescence at 530 nm when excited at485 nm. Cells seeded in 96-well culture plates were incubated withDCFC-DA (10 μg/ml) for 5 min, treated with or without 10 mMpyruvate before adding either 0.1 mM 6-OHDA or vehicle and 4 hlater fluorescence intensity was measured in a Spectra Max GeminiXS (Molecular Devices). The average relative percent ROSproduction from four wells of at least three separate cultures wasdetermined. Results are expressed as mean±SD values, andsignificance was determined by Student’s t test. Statisticalsignificance was considered at the p<0.05 level.
Measurement of glutathione levels
Levels of glutathione were determined by using monochlor-obimane (mBCl) fluorescence. Glutathione (GSH) is specificallyconjugated with mBCl to form a fluorescent bimane-GSHadduct in a reaction catalyzed by glutathione S-transferases(Shrieve et al., 1988). The concentration of the bimane-GSHadduct increases during the initial 10 to 12 min period of thisreaction with first order kinetics, before leveling off (Young etal., 1994). Culture medium was removed and cells were washedtwice with 0.2 ml Krebs and incubated for 1 h at 37°C in0.2 ml fresh Krebs containing 160 μM mBCl. After incubation,cells were washed twice with Krebs and fluorescence was
298 F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
measured at an excitation wavelength of 340 nm and emissionwavelength of 460 nm in a Spectra Max Gemini XS (MolecularDevices). The average relative percent reduced GSH levels fromat least three separate cultures was determined. Results areexpressed as mean±SD values, and significance was determinedby Student’s t test. Statistical significance was considered at thep < 0.05 level.
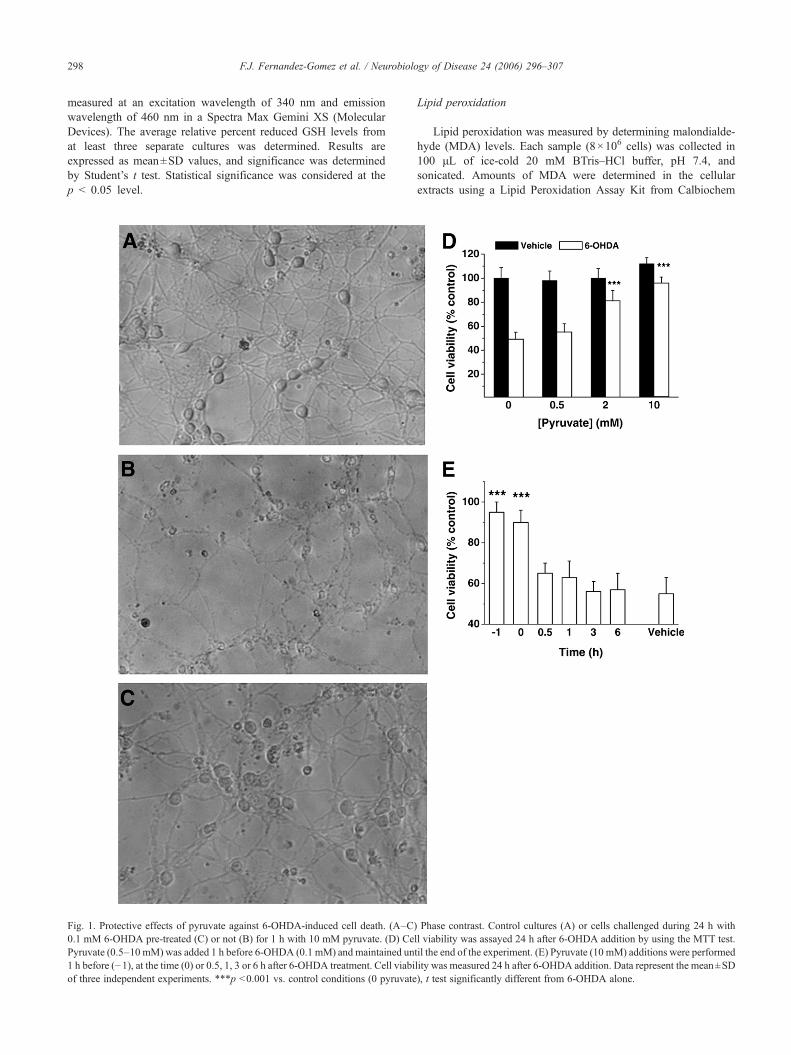
Fig. 1. Protective effects of pyruvate against 6-OHDA-induced cell death. (A–C)0.1 mM 6-OHDA pre-treated (C) or not (B) for 1 h with 10 mM pyruvate. (D) CePyruvate (0.5–10 mM) was added 1 h before 6-OHDA (0.1 mM) and maintained un1 h before (−1), at the time (0) or 0.5, 1, 3 or 6 h after 6-OHDA treatment. Cell viabiof three independent experiments. ***p <0.001 vs. control conditions (0 pyruvate
Lipid peroxidation
Lipid peroxidation was measured by determining malondialde-hyde (MDA) levels. Each sample (8×106 cells) was collected in100 μL of ice-cold 20 mM BTris–HCl buffer, pH 7.4, andsonicated. Amounts of MDA were determined in the cellularextracts using a Lipid Peroxidation Assay Kit from Calbiochem
Phase contrast. Control cultures (A) or cells challenged during 24 h withll viability was assayed 24 h after 6-OHDA addition by using the MTT test.til the end of the experiment. (E) Pyruvate (10 mM) additions were performedlity was measured 24 h after 6-OHDA addition. Data represent the mean±SD), t test significantly different from 6-OHDA alone.
299F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
(No. 437634) based on the condensation reaction of thechromogene 1-methyl-2-phenylindole with MDA. The stablechromophores were determined using a VERSAmax Lunablemicroplate reader (Molecular Devices) with absorbance at 586 nm.Results are expressed as micromole MDA per mg protein.
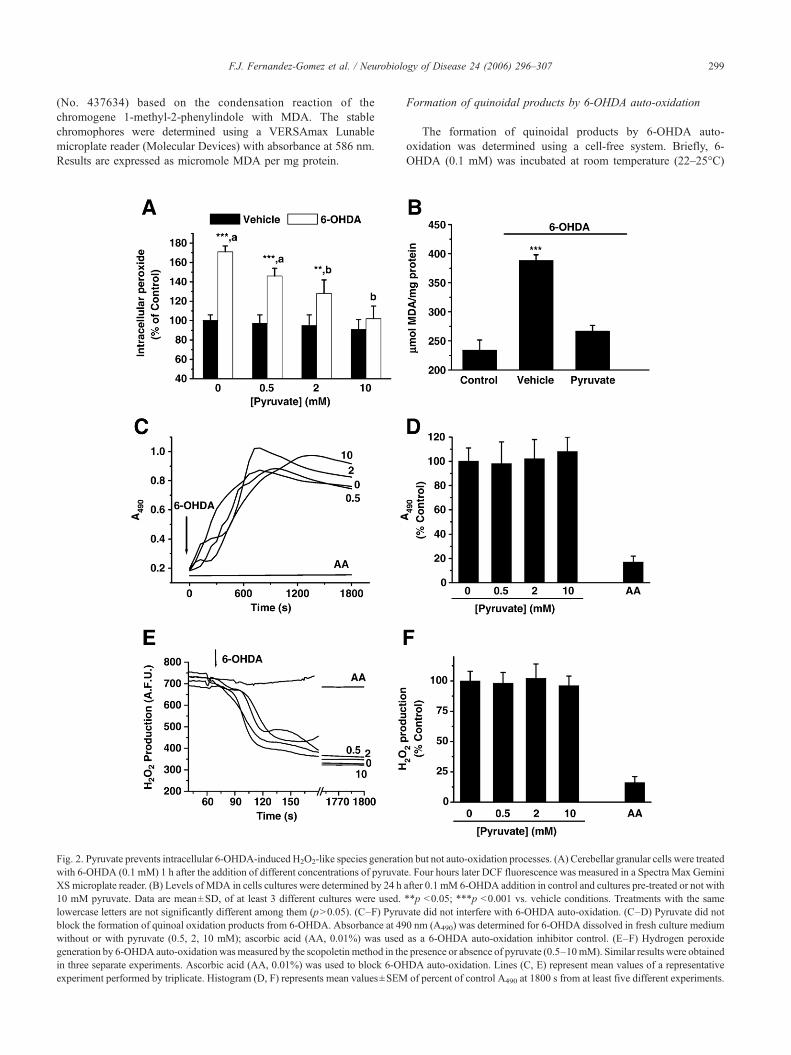
Fig. 2. Pyruvate prevents intracellular 6-OHDA-induced H2O2-like species generatiowith 6-OHDA (0.1 mM) 1 h after the addition of different concentrations of pyruvatXS microplate reader. (B) Levels of MDA in cells cultures were determined by 24 h10 mM pyruvate. Data are mean±SD, of at least 3 different cultures were used.lowercase letters are not significantly different among them (p>0.05). (C–F) Pyrublock the formation of quinoal oxidation products from 6-OHDA. Absorbance at 49without or with pyruvate (0.5, 2, 10 mM); ascorbic acid (AA, 0.01%) was usedgeneration by 6-OHDA auto-oxidation wasmeasured by the scopoletin method in thin three separate experiments. Ascorbic acid (AA, 0.01%) was used to block 6-OHexperiment performed by triplicate. Histogram (D, F) represents mean values±SEM
Formation of quinoidal products by 6-OHDA auto-oxidation
The formation of quinoidal products by 6-OHDA auto-oxidation was determined using a cell-free system. Briefly, 6-OHDA (0.1 mM) was incubated at room temperature (22–25°C)
n but not auto-oxidation processes. (A) Cerebellar granular cells were treatede. Four hours later DCF fluorescence was measured in a Spectra Max Geminiafter 0.1 mM 6-OHDA addition in control and cultures pre-treated or not with**p <0.05; ***p <0.001 vs. vehicle conditions. Treatments with the samevate did not interfere with 6-OHDA auto-oxidation. (C–D) Pyruvate did not0 nm (A490) was determined for 6-OHDA dissolved in fresh culture mediumas a 6-OHDA auto-oxidation inhibitor control. (E–F) Hydrogen peroxidee presence or absence of pyruvate (0.5–10mM). Similar results were obtainedDA auto-oxidation. Lines (C, E) represent mean values of a representativeof percent of control A490 at 1800 s from at least five different experiments.
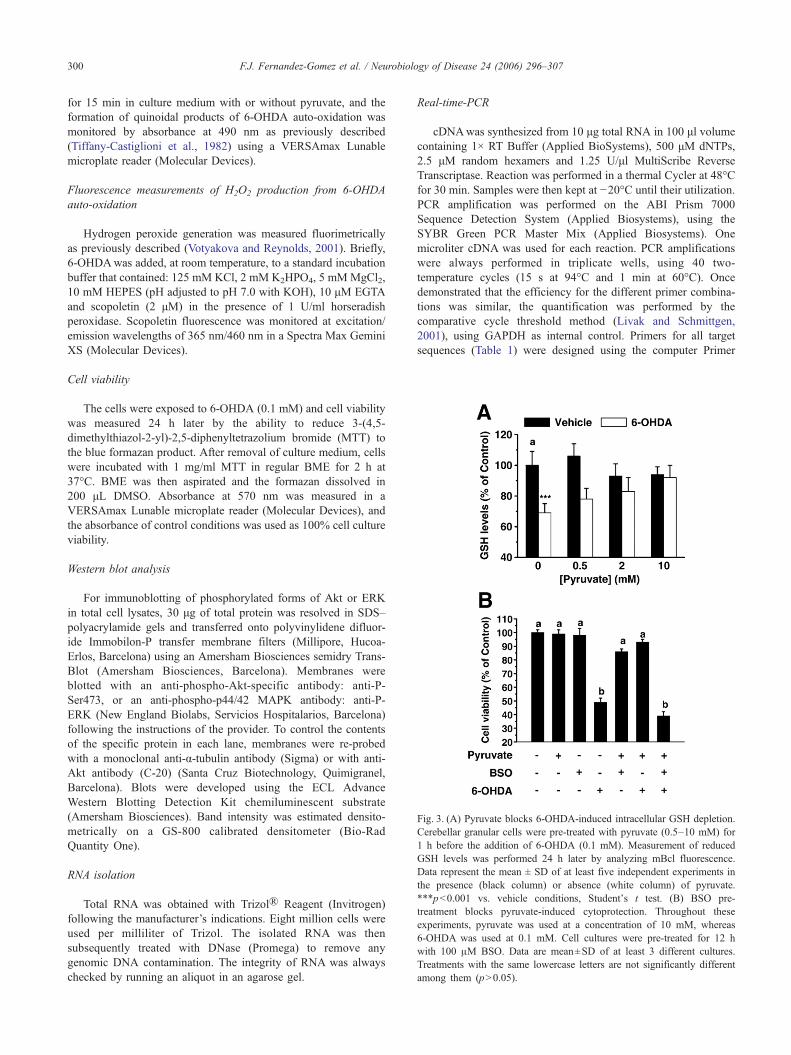
Fig. 3. (A) Pyruvate blocks 6-OHDA-induced intracellular GSH depletion.Cerebellar granular cells were pre-treated with pyruvate (0.5–10 mM) for1 h before the addition of 6-OHDA (0.1 mM). Measurement of reducedGSH levels was performed 24 h later by analyzing mBcl fluorescence.Data represent the mean ± SD of at least five independent experiments inthe presence (black column) or absence (white column) of pyruvate.***p<0.001 vs. vehicle conditions, Student's t test. (B) BSO pre-treatment blocks pyruvate-induced cytoprotection. Throughout theseexperiments, pyruvate was used at a concentration of 10 mM, whereas6-OHDA was used at 0.1 mM. Cell cultures were pre-treated for 12 hwith 100 μM BSO. Data are mean±SD of at least 3 different cultures.Treatments with the same lowercase letters are not significantly differentamong them (p>0.05).
300 F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
for 15 min in culture medium with or without pyruvate, and theformation of quinoidal products of 6-OHDA auto-oxidation wasmonitored by absorbance at 490 nm as previously described(Tiffany-Castiglioni et al., 1982) using a VERSAmax Lunablemicroplate reader (Molecular Devices).
Fluorescence measurements of H2O2 production from 6-OHDAauto-oxidation
Hydrogen peroxide generation was measured fluorimetricallyas previously described (Votyakova and Reynolds, 2001). Briefly,6-OHDAwas added, at room temperature, to a standard incubationbuffer that contained: 125 mM KCl, 2 mM K2HPO4, 5 mMMgCl2,10 mM HEPES (pH adjusted to pH 7.0 with KOH), 10 μM EGTAand scopoletin (2 μM) in the presence of 1 U/ml horseradishperoxidase. Scopoletin fluorescence was monitored at excitation/emission wavelengths of 365 nm/460 nm in a Spectra Max GeminiXS (Molecular Devices).
Cell viability
The cells were exposed to 6-OHDA (0.1 mM) and cell viabilitywas measured 24 h later by the ability to reduce 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) tothe blue formazan product. After removal of culture medium, cellswere incubated with 1 mg/ml MTT in regular BME for 2 h at37°C. BME was then aspirated and the formazan dissolved in200 μL DMSO. Absorbance at 570 nm was measured in aVERSAmax Lunable microplate reader (Molecular Devices), andthe absorbance of control conditions was used as 100% cell cultureviability.
Western blot analysis
For immunoblotting of phosphorylated forms of Akt or ERKin total cell lysates, 30 μg of total protein was resolved in SDS–polyacrylamide gels and transferred onto polyvinylidene difluor-ide Immobilon-P transfer membrane filters (Millipore, Hucoa-Erlos, Barcelona) using an Amersham Biosciences semidry Trans-Blot (Amersham Biosciences, Barcelona). Membranes wereblotted with an anti-phospho-Akt-specific antibody: anti-P-Ser473, or an anti-phospho-p44/42 MAPK antibody: anti-P-ERK (New England Biolabs, Servicios Hospitalarios, Barcelona)following the instructions of the provider. To control the contentsof the specific protein in each lane, membranes were re-probedwith a monoclonal anti-α-tubulin antibody (Sigma) or with anti-Akt antibody (C-20) (Santa Cruz Biotechnology, Quimigranel,Barcelona). Blots were developed using the ECL AdvanceWestern Blotting Detection Kit chemiluminescent substrate(Amersham Biosciences). Band intensity was estimated densito-metrically on a GS-800 calibrated densitometer (Bio-RadQuantity One).
RNA isolation
Total RNA was obtained with Trizol® Reagent (Invitrogen)following the manufacturer’s indications. Eight million cells wereused per milliliter of Trizol. The isolated RNA was thensubsequently treated with DNase (Promega) to remove anygenomic DNA contamination. The integrity of RNA was alwayschecked by running an aliquot in an agarose gel.
Real-time-PCR
cDNAwas synthesized from 10 μg total RNA in 100 μl volumecontaining 1× RT Buffer (Applied BioSystems), 500 μM dNTPs,2.5 μM random hexamers and 1.25 U/μl MultiScribe ReverseTranscriptase. Reaction was performed in a thermal Cycler at 48°Cfor 30 min. Samples were then kept at −20°C until their utilization.PCR amplification was performed on the ABI Prism 7000Sequence Detection System (Applied Biosystems), using theSYBR Green PCR Master Mix (Applied Biosystems). Onemicroliter cDNA was used for each reaction. PCR amplificationswere always performed in triplicate wells, using 40 two-temperature cycles (15 s at 94°C and 1 min at 60°C). Oncedemonstrated that the efficiency for the different primer combina-tions was similar, the quantification was performed by thecomparative cycle threshold method (Livak and Schmittgen,2001), using GAPDH as internal control. Primers for all targetsequences (Table 1) were designed using the computer Primer
301F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
Express software program specially provided with the ABI Prism7000 Sequence Detection System.
Statistical analysis
The data are expressed as the mean±SD and were analyzedusing the two-tailed unpaired Student’s t test. Treatment differenceswere considered statistically significant at p <0.05. Data analyseswere performed using the Statistical Program for the SocialSciences (SPSS for Windows, 11.0).
Results
Pyruvate protects against 6-OHDA-induced cell death
The results shown here confirm and expand the observationsthat 6-OHDA induces cell death in different models (Galindo etal., 2003; Mazzio et al., 2004; Blum et al., 2001). As shown in
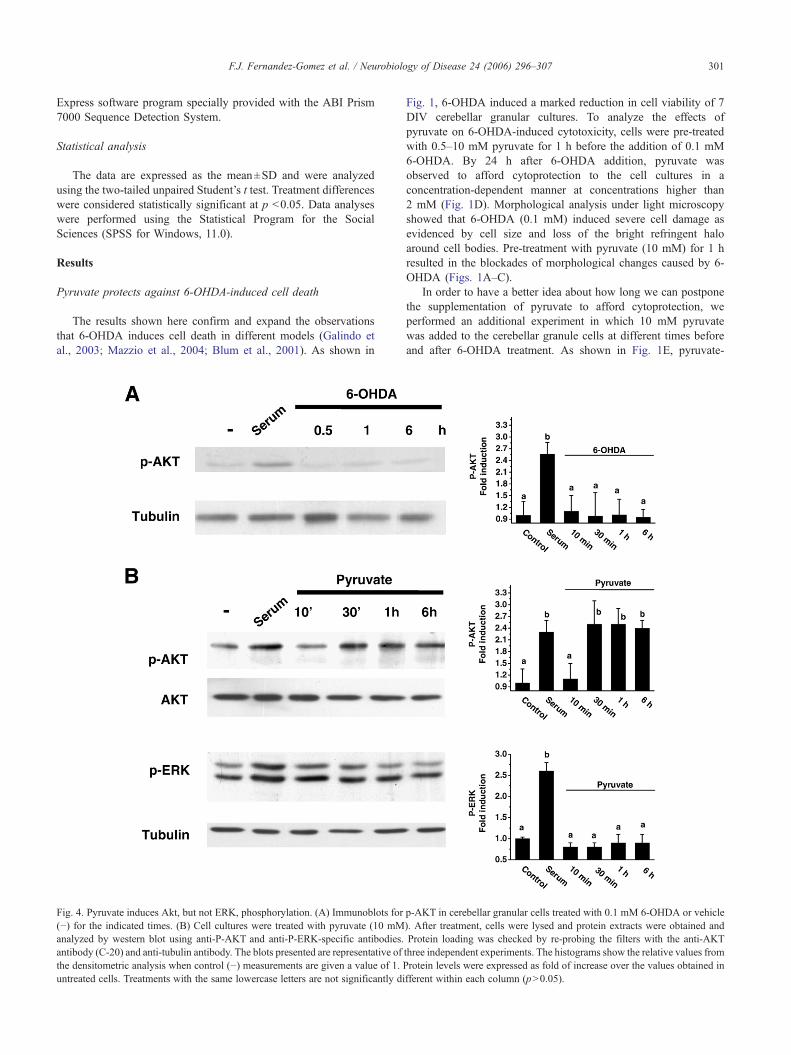
Fig. 4. Pyruvate induces Akt, but not ERK, phosphorylation. (A) Immunoblots for(−) for the indicated times. (B) Cell cultures were treated with pyruvate (10 mManalyzed by western blot using anti-P-AKT and anti-P-ERK-specific antibodies.antibody (C-20) and anti-tubulin antibody. The blots presented are representative ofthe densitometric analysis when control (−) measurements are given a value of 1. Puntreated cells. Treatments with the same lowercase letters are not significantly di
Fig. 1, 6-OHDA induced a marked reduction in cell viability of 7DIV cerebellar granular cultures. To analyze the effects ofpyruvate on 6-OHDA-induced cytotoxicity, cells were pre-treatedwith 0.5–10 mM pyruvate for 1 h before the addition of 0.1 mM6-OHDA. By 24 h after 6-OHDA addition, pyruvate wasobserved to afford cytoprotection to the cell cultures in aconcentration-dependent manner at concentrations higher than2 mM (Fig. 1D). Morphological analysis under light microscopyshowed that 6-OHDA (0.1 mM) induced severe cell damage asevidenced by cell size and loss of the bright refringent haloaround cell bodies. Pre-treatment with pyruvate (10 mM) for 1 hresulted in the blockades of morphological changes caused by 6-OHDA (Figs. 1A–C).
In order to have a better idea about how long we can postponethe supplementation of pyruvate to afford cytoprotection, weperformed an additional experiment in which 10 mM pyruvatewas added to the cerebellar granule cells at different times beforeand after 6-OHDA treatment. As shown in Fig. 1E, pyruvate-
p-AKT in cerebellar granular cells treated with 0.1 mM 6-OHDA or vehicle). After treatment, cells were lysed and protein extracts were obtained andProtein loading was checked by re-probing the filters with the anti-AKTthree independent experiments. The histograms show the relative values fromrotein levels were expressed as fold of increase over the values obtained infferent within each column (p>0.05).
302 F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
conferred protection was evident only when pyruvate was addedto the cells either before or at the same time as 6-OHDA, butdelayed treatment with pyruvate was no longer able to reverse thecytotoxicity. Moreover, if pyruvate was removed from the culturemedium just before the 6-OHDA addition, it lacked itscytoprotective effects.
Pyruvate exerts anti-oxidant actions in cerebellar granular cellcultures
Previous studies have shown that pyruvate might affordcytoprotection through a mechanism that implies a ROS scavengercapacity (Alvarez et al., 2003; Salahudeen et al., 1991).Additionally, it has also been proposed that 6-OHDA requires
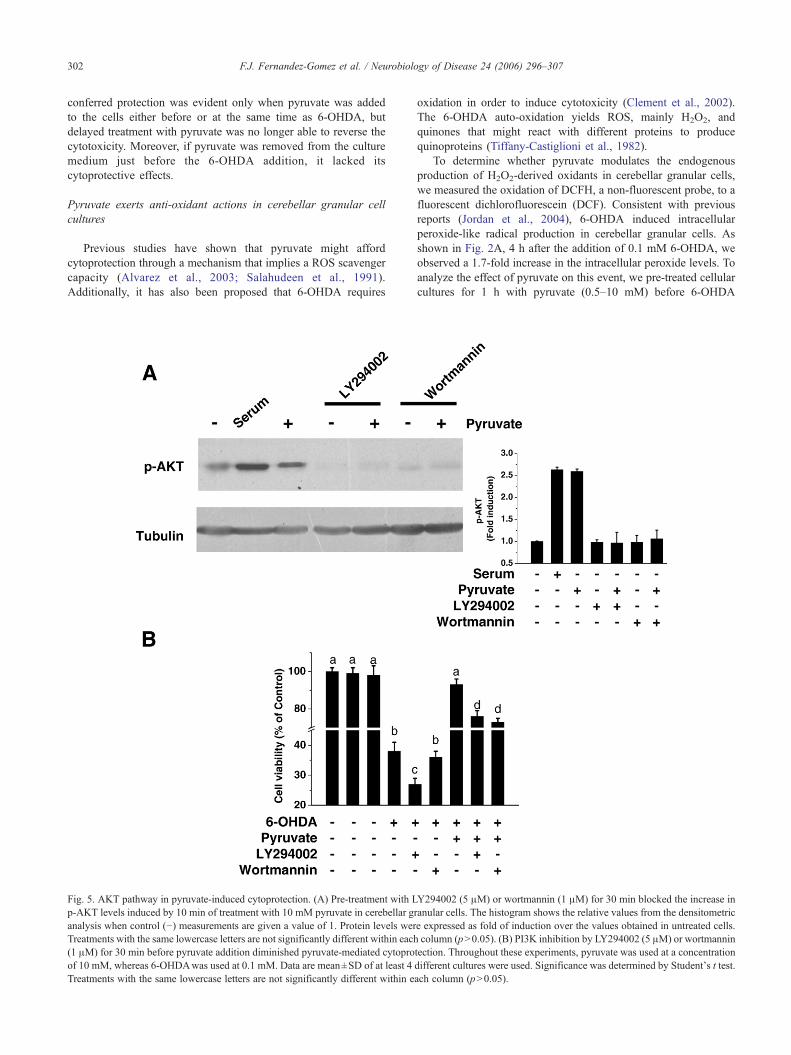
Fig. 5. AKT pathway in pyruvate-induced cytoprotection. (A) Pre-treatment with Lp-AKT levels induced by 10 min of treatment with 10 mM pyruvate in cerebellar ganalysis when control (−) measurements are given a value of 1. Protein levels werTreatments with the same lowercase letters are not significantly different within each(1 μM) for 30 min before pyruvate addition diminished pyruvate-mediated cytoproof 10 mM, whereas 6-OHDAwas used at 0.1 mM. Data are mean±SD of at least 4Treatments with the same lowercase letters are not significantly different within e
oxidation in order to induce cytotoxicity (Clement et al., 2002).The 6-OHDA auto-oxidation yields ROS, mainly H2O2, andquinones that might react with different proteins to producequinoproteins (Tiffany-Castiglioni et al., 1982).
To determine whether pyruvate modulates the endogenousproduction of H2O2-derived oxidants in cerebellar granular cells,we measured the oxidation of DCFH, a non-fluorescent probe, to afluorescent dichlorofluorescein (DCF). Consistent with previousreports (Jordan et al., 2004), 6-OHDA induced intracellularperoxide-like radical production in cerebellar granular cells. Asshown in Fig. 2A, 4 h after the addition of 0.1 mM 6-OHDA, weobserved a 1.7-fold increase in the intracellular peroxide levels. Toanalyze the effect of pyruvate on this event, we pre-treated cellularcultures for 1 h with pyruvate (0.5–10 mM) before 6-OHDA
Y294002 (5 μM) or wortmannin (1 μM) for 30 min blocked the increase inranular cells. The histogram shows the relative values from the densitometrice expressed as fold of induction over the values obtained in untreated cells.column (p>0.05). (B) PI3K inhibition by LY294002 (5 μM) or wortmannin
tection. Throughout these experiments, pyruvate was used at a concentrationdifferent cultures were used. Significance was determined by Student's t test.ach column (p>0.05).
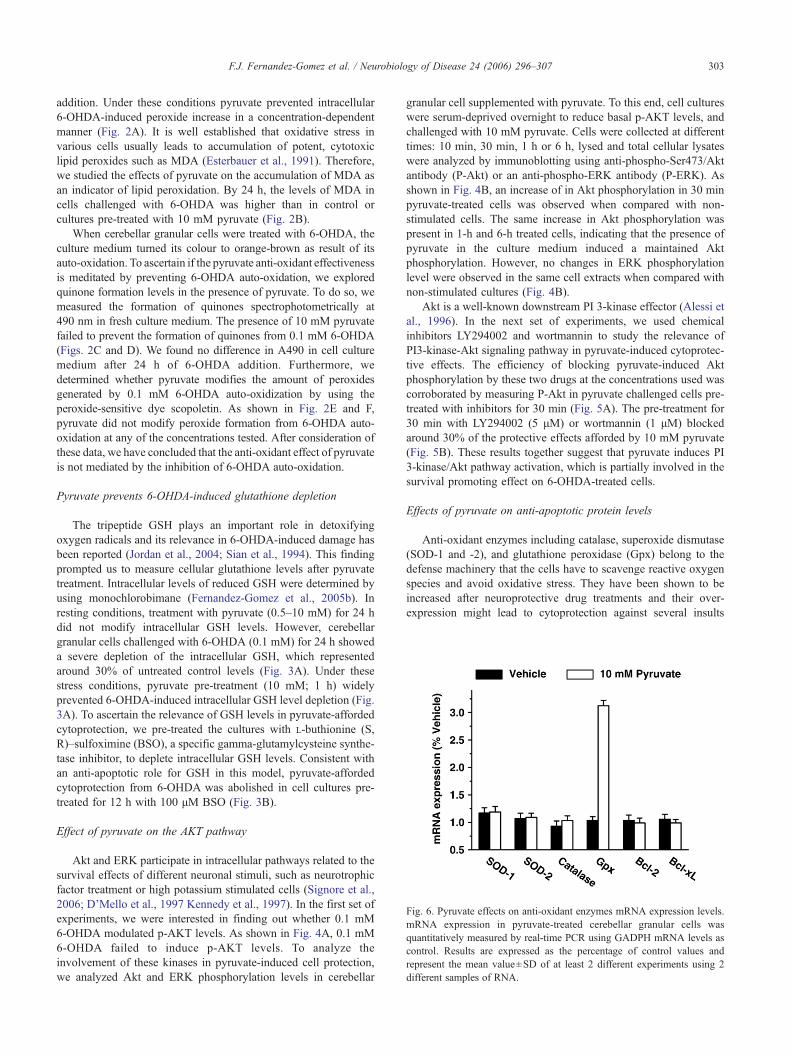
Fig. 6. Pyruvate effects on anti-oxidant enzymes mRNA expression levels.mRNA expression in pyruvate-treated cerebellar granular cells wasquantitatively measured by real-time PCR using GADPH mRNA levels ascontrol. Results are expressed as the percentage of control values andrepresent the mean value±SD of at least 2 different experiments using 2different samples of RNA.
303F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
addition. Under these conditions pyruvate prevented intracellular6-OHDA-induced peroxide increase in a concentration-dependentmanner (Fig. 2A). It is well established that oxidative stress invarious cells usually leads to accumulation of potent, cytotoxiclipid peroxides such as MDA (Esterbauer et al., 1991). Therefore,we studied the effects of pyruvate on the accumulation of MDA asan indicator of lipid peroxidation. By 24 h, the levels of MDA incells challenged with 6-OHDA was higher than in control orcultures pre-treated with 10 mM pyruvate (Fig. 2B).
When cerebellar granular cells were treated with 6-OHDA, theculture medium turned its colour to orange-brown as result of itsauto-oxidation. To ascertain if the pyruvate anti-oxidant effectivenessis meditated by preventing 6-OHDA auto-oxidation, we exploredquinone formation levels in the presence of pyruvate. To do so, wemeasured the formation of quinones spectrophotometrically at490 nm in fresh culture medium. The presence of 10 mM pyruvatefailed to prevent the formation of quinones from 0.1 mM 6-OHDA(Figs. 2C and D). We found no difference in A490 in cell culturemedium after 24 h of 6-OHDA addition. Furthermore, wedetermined whether pyruvate modifies the amount of peroxidesgenerated by 0.1 mM 6-OHDA auto-oxidization by using theperoxide-sensitive dye scopoletin. As shown in Fig. 2E and F,pyruvate did not modify peroxide formation from 6-OHDA auto-oxidation at any of the concentrations tested. After consideration ofthese data, we have concluded that the anti-oxidant effect of pyruvateis not mediated by the inhibition of 6-OHDA auto-oxidation.
Pyruvate prevents 6-OHDA-induced glutathione depletion
The tripeptide GSH plays an important role in detoxifyingoxygen radicals and its relevance in 6-OHDA-induced damage hasbeen reported (Jordan et al., 2004; Sian et al., 1994). This findingprompted us to measure cellular glutathione levels after pyruvatetreatment. Intracellular levels of reduced GSH were determined byusing monochlorobimane (Fernandez-Gomez et al., 2005b). Inresting conditions, treatment with pyruvate (0.5–10 mM) for 24 hdid not modify intracellular GSH levels. However, cerebellargranular cells challenged with 6-OHDA (0.1 mM) for 24 h showeda severe depletion of the intracellular GSH, which representedaround 30% of untreated control levels (Fig. 3A). Under thesestress conditions, pyruvate pre-treatment (10 mM; 1 h) widelyprevented 6-OHDA-induced intracellular GSH level depletion (Fig.3A). To ascertain the relevance of GSH levels in pyruvate-affordedcytoprotection, we pre-treated the cultures with L-buthionine (S,R)–sulfoximine (BSO), a specific gamma-glutamylcysteine synthe-tase inhibitor, to deplete intracellular GSH levels. Consistent withan anti-apoptotic role for GSH in this model, pyruvate-affordedcytoprotection from 6-OHDA was abolished in cell cultures pre-treated for 12 h with 100 μM BSO (Fig. 3B).
Effect of pyruvate on the AKT pathway
Akt and ERK participate in intracellular pathways related to thesurvival effects of different neuronal stimuli, such as neurotrophicfactor treatment or high potassium stimulated cells (Signore et al.,2006; D’Mello et al., 1997 Kennedy et al., 1997). In the first set ofexperiments, we were interested in finding out whether 0.1 mM6-OHDA modulated p-AKT levels. As shown in Fig. 4A, 0.1 mM6-OHDA failed to induce p-AKT levels. To analyze theinvolvement of these kinases in pyruvate-induced cell protection,we analyzed Akt and ERK phosphorylation levels in cerebellar
granular cell supplemented with pyruvate. To this end, cell cultureswere serum-deprived overnight to reduce basal p-AKT levels, andchallenged with 10 mM pyruvate. Cells were collected at differenttimes: 10 min, 30 min, 1 h or 6 h, lysed and total cellular lysateswere analyzed by immunoblotting using anti-phospho-Ser473/Aktantibody (P-Akt) or an anti-phospho-ERK antibody (P-ERK). Asshown in Fig. 4B, an increase of in Akt phosphorylation in 30 minpyruvate-treated cells was observed when compared with non-stimulated cells. The same increase in Akt phosphorylation waspresent in 1-h and 6-h treated cells, indicating that the presence ofpyruvate in the culture medium induced a maintained Aktphosphorylation. However, no changes in ERK phosphorylationlevel were observed in the same cell extracts when compared withnon-stimulated cultures (Fig. 4B).
Akt is a well-known downstream PI 3-kinase effector (Alessi etal., 1996). In the next set of experiments, we used chemicalinhibitors LY294002 and wortmannin to study the relevance ofPI3-kinase-Akt signaling pathway in pyruvate-induced cytoprotec-tive effects. The efficiency of blocking pyruvate-induced Aktphosphorylation by these two drugs at the concentrations used wascorroborated by measuring P-Akt in pyruvate challenged cells pre-treated with inhibitors for 30 min (Fig. 5A). The pre-treatment for30 min with LY294002 (5 μM) or wortmannin (1 μM) blockedaround 30% of the protective effects afforded by 10 mM pyruvate(Fig. 5B). These results together suggest that pyruvate induces PI3-kinase/Akt pathway activation, which is partially involved in thesurvival promoting effect on 6-OHDA-treated cells.
Effects of pyruvate on anti-apoptotic protein levels
Anti-oxidant enzymes including catalase, superoxide dismutase(SOD-1 and -2), and glutathione peroxidase (Gpx) belong to thedefense machinery that the cells have to scavenge reactive oxygenspecies and avoid oxidative stress. They have been shown to beincreased after neuroprotective drug treatments and their over-expression might lead to cytoprotection against several insults
304 F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
(Thiruchelvam et al., 2005). To ascertain whether pyruvate mightmodulate the expression of some of these enzymes, we utilizedquantitative real time PCR to measure the level of expression ofthose enzymes in cerebellar granular cells treated with 10 mMpyruvate for 1 h, time by which pyruvate treatment prevents 6-OHDA-induced cell death. The results, shown in Fig. 6, revealedthat pyruvate induces a strong up-regulation of GPX1, whereas nomodifications were observed in catalase or SOD family memberlevels.
In a previous study we observed that the overexpression ofBcl-xL prevented SH-SY5Y cells from 6-OHDA toxicity (Jordan etal., 2004). Thus, we tested whether pyruvate cytoprotective effectswere mediated throughout changes in the expression levels of Bcl-2or Bcl-xL. As shown in Fig. 6, by 1 h after 10 mM pyruvate addition,
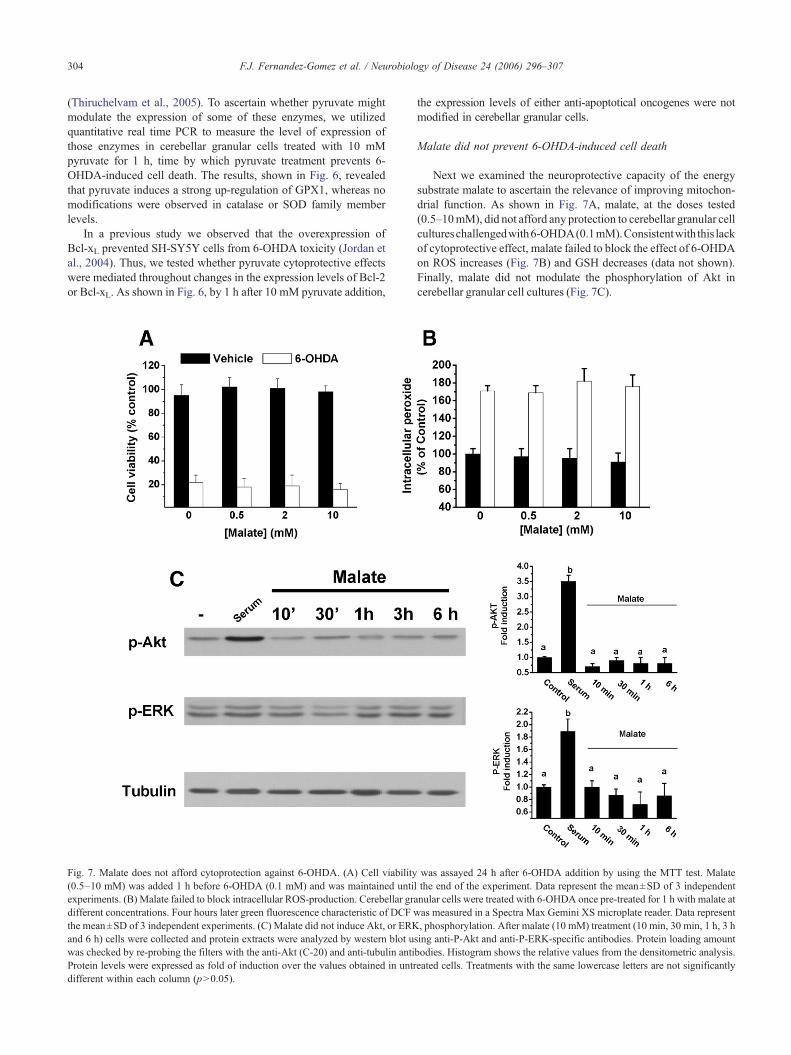
Fig. 7. Malate does not afford cytoprotection against 6-OHDA. (A) Cell viability(0.5–10 mM) was added 1 h before 6-OHDA (0.1 mM) and was maintained untiexperiments. (B) Malate failed to block intracellular ROS-production. Cerebellar gradifferent concentrations. Four hours later green fluorescence characteristic of DCF wthe mean±SD of 3 independent experiments. (C) Malate did not induce Akt, or ERKand 6 h) cells were collected and protein extracts were analyzed by western blot uwas checked by re-probing the filters with the anti-Akt (C-20) and anti-tubulin antiProtein levels were expressed as fold of induction over the values obtained in untrdifferent within each column (p>0.05).
the expression levels of either anti-apoptotical oncogenes were notmodified in cerebellar granular cells.
Malate did not prevent 6-OHDA-induced cell death
Next we examined the neuroprotective capacity of the energysubstrate malate to ascertain the relevance of improving mitochon-drial function. As shown in Fig. 7A, malate, at the doses tested(0.5–10mM), did not afford any protection to cerebellar granular cellcultureschallengedwith6-OHDA(0.1mM).Consistentwith this lackof cytoprotective effect, malate failed to block the effect of 6-OHDAon ROS increases (Fig. 7B) and GSH decreases (data not shown).Finally, malate did not modulate the phosphorylation of Akt incerebellar granular cell cultures (Fig. 7C).
was assayed 24 h after 6-OHDA addition by using the MTT test. Malatel the end of the experiment. Data represent the mean±SD of 3 independentnular cells were treated with 6-OHDA once pre-treated for 1 h with malate atas measured in a Spectra Max Gemini XS microplate reader. Data represent, phosphorylation. After malate (10 mM) treatment (10 min, 30 min, 1 h, 3 h
sing anti-P-Akt and anti-P-ERK-specific antibodies. Protein loading amountbodies. Histogram shows the relative values from the densitometric analysis.eated cells. Treatments with the same lowercase letters are not significantly

305F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
Discussion
The results presented in this study are consistent with thehypothesis that pyruvate affords cytoprotection by an anti-oxidantmechanism. Although this is a well-known fact, we havedemonstrated for the first time that this alpha-ketoacid conferscytoprotection to cerebellar granular cell cultures challenged with6-OHDA through a mechanism that involves the up-regulation ofglutathione peroxidase expression and the activation of the PI3K/Akt signaling pathway.
Consistent with the cytoprotective capacity of pyruvate shownin several experimental models where reactive oxygen species areinvolved, pyruvate, in a concentration-dependent manner, pro-tected rat cerebellar granular cell cultures and prevented the rise inlipid peroxidation induced by 24 h exposure to 6-OHDA (0.1 mM).According to our data, this anti-oxidant action of pyruvate isrelated to its ability to block the increase of intracellular peroxidelevels induced by 6-OHDA rather than by avoiding 6-OHDA auto-oxidation. Thus, pyruvate at the highest dose tested, 10 mM, failedto scavenge the peroxides or to prevent the quinone formationresulting from 6-OHDA auto-oxidation in a cell-free system.Although pyruvate did not increase GSH levels in restingconditions, it maintained the reduced glutathione levels after 6-OHDA treatment, revealing a possible mechanism for theprevention of peroxide formation. Consistent with a key role forthis tripeptide in our model, pyruvate-induced protection against 6-OHDA toxicity disappeared when we depleted intracellular GSHlevels by using the GSH synthesis inhibitor L-buthioninesulfoximine. GSH is an efficient anti-oxidant and a free radicalscavenger whose levels have been found to be altered in the basalganglia of PD patients (Sian et al., 1994). How pyruvate avoidsGSH depletion after 6-OHDA remains unknown. Although notaddressed in this study, it has been demonstrated that pyruvateinduces ATP and NAD(P)H formation (De Andrade et al., 2004). Itis therefore conceivable that pyruvate in cerebellar granular cellsincreases NAD(P)H levels and subsequently GSH levels.
The fact that pyruvate lost its cytoprotective effect when addedafter the 6-OHDA insult together with the inability of malate toafford cytoprotection to cerebellar granular cells prompted us toinvestigate the requirement of the activation of an intracellularsignaling pathway. To achieve our goal, we used quantitative real-time PCR to measure the levels of expression of several anti-oxidant enzymes 1 h after the addition of pyruvate (10 mM). Underour experimental conditions, mRNA levels of glutathione perox-idase increased by three folds. By contrast, no apparent changeswere found in the expression of other anti-oxidant enzymes such ascatalase, superoxide dismutase-1 and -2, or the anti-apoptoticproteins Bcl-2 and Bcl-xL which are known to prevent 6-OHDAtoxicity by avoiding GSH depletion (Jordan et al., 2004). Gpx is acentral enzyme that participates in maintaining intracellularperoxide homeostasis. Thus, it is responsible for detoxifying lipidhydroperoxides, hydrogen peroxide and hydroxyl radicals (Halli-well, 1992). In mitochondria, removal of H2O2 is catalyzedprimarily by mitochondrial Gpx at the expense of GSH.Accordingly, pyruvate, by keeping the levels of intracellularGSH in its reduced state in combination with increased glutathioneperoxidase levels, might optimize cellular peroxide scavengercapacity. In line with these results, Gpx has been shown toparticipate in the cytoprotective pathways activated by nervegrowth factor (NGF) in PC12 cells (Sampath et al., 1994) and inthe aged rat brain (Nistico et al., 1991). Further, increased levels of
Gpx renders cells resistant to 6-OHDA (Ridet et al., 2006) andother quinone-containing drugs such as doxorubicin, daunorubicin,mitomycin C, diaziquone and menadione (Doroshow et al., 1990).
Our findings also show that pyruvate increases Akt but notERK phosphorylation in cerebellar granular cells, suggesting thatthe PI3K-Akt signaling pathway may participate in the mechan-isms by which pyruvate confers cytoprotection. Further experi-ments using the upstream kinase phosphatidylinositol 3-kinaseinhibitors LY294002 (5 μM) or wortmannin (1 μM) demonstratedthat this pathway is in part responsible for pyruvate-inducedcytoprotection because both inhibitors decreased by around 30% ofthe protective effect of pyruvate. The implications of PI3K-Aktactivation in response to different anti-apoptotic agents has beenwell documented by many groups (D’Mello et al., 1997, Kennedyet al., 1997). For instance, it has been recently reported thaterythropoietin protects against 6-OHDA-induced dopaminergiccell death via the PI3K pathway (Signore et al., 2006). Thus, byusing the constitutively active Akt, these authors demonstrated theimportance of Akt in preserving dopaminergic cells exposed to 6-OHDA. It is noteworthy that pre-treatment with LY294002, but notwortmannin, predisposes cell cultures to 6-OHDA-induced toxi-city, suggesting that the PI3K-Akt pathway participates in thephysiological anti-apoptotic cellular machinery. Although bothinhibitors are considered to be specific PI3K inhibitors, it has beenrecently reported that LY294002 might also exert other effectsdespite inhibiting PI3K (Yamaguchi et al., 2006). We do not knowwhy these two inhibitors present different effects. Furtherinvestigation is warranted to delineate the likely distinct mechan-isms by which these drugs act.
In summary, our results appear to indicate that pyruvate-induced cytoprotection against 6-OHDA is mediated by anincrease in the anti-oxidant capacity of cells due to the up-regulation of glutathione peroxidase expression rather than to itsanti-oxidant capacity derived from its alpha-ketoacid structure or toits energetic substrate role. Furthermore, our data also show thatthe activation of the PI3K-Akt signaling pathway plays asignificant role in the protection afforded by pyruvate against 6-OHDA toxicity.
Acknowledgments
We thank Ichiro Ikuta for careful reading and comments on themanuscript. We are grateful to Dr. Maria Ramos and Prof. NorbertoAgurirre for discussions and Remedios Sanchis for technicalassistance. This study was supported by SAF2002-04721 andSAF2005-07919-C02-01 from CICYT and Sanidad. 04005-00from JCCM to J.J. PAI-05-017 from JCCM to SC and from Fondode Investigaciones Sanitarias (FIS) PI051445 to RMS. F.J. F-G, M.G-L and M.D.P. are fellows from JCCM and M.G-F from FIS.
References
Alessi, D.R., Andjelkovic, M., Caudwell, B., Cron, P., Morrice, N., Cohen,P., Hemmings, B.A., 1996. Mechanism of activation of protein kinase Bby insulin and IGF-1. EMBO J. 15, 541–551.
Alvarez, G., Ramos, M., Ruiz, F., Satrustegui, J., Bogonez, E., 2003.Pyruvate protection against beta-amyloid-induced neuronal death: roleof mitochondrial redox state. J. Neurosci. Res. 73, 260–269.
Beal, M.F., 2003. Bioenergetic approaches for neuroprotection in Parkin-son's disease. Ann Neurol 53 (Suppl 3), S39–S47 (discussion S47–8).
Blum, D., Torch, S., Lambeng, N., Nissou, M., Benabid, A.L., Sadoul, R.,

306 F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
Verna, J.M., 2001. Molecular pathways involved in the neurotoxicity of6-OHDA, dopamine and MPTP: contribution to the apoptotic theory inParkinson's disease. Prog. Neurobiol. 65, 135–172.
Chung, S.J., Lee, S.H., Lee, Y.J., Park, H.S., Bunger, R., Kang, Y.H., 2004.Pyruvate protection against endothelial cytotoxicity induced by block-ade of glucose uptake. J. Biochem. Mol. Biol. 37, 239–245.
Clement, M.V., Long, L.H., Ramalingam, J., Halliwell, B., 2002. Thecytotoxicity of dopamine may be an artefact of cell culture.J. Neurochem. 81, 414–421.
D'Mello, S.R., Borodezt, K., Soltoff, S.P., 1997. Insulin-like growthfactor and potassium depolarization maintain neuronal survival bydistinct pathways: possible involvement of PI 3-kinase in IGF-1signaling. J. Neurosci. 17, 1548–1560.
de Andrade, P.B., Casimir, M., Maechler, P., 2004. Mitochondrial activationand the pyruvate paradox in a human cell line. FEBS Lett. 578, 224–228.
Doroshow, J.H., Akman, S., Chu, F.F., Esworthy, S., 1990. Role of theglutathione-glutathione peroxidase cycle in the cytotoxicity of theanticancer quinones. Pharmacol. Ther. 47, 359–370.
Esterbauer, H., Schaur, R.J., Zollner, H., 1991. Chemistry and biochemistryof 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic.Biol. Med. 11, 81–128.
Fernandez-Gomez, F.J., Gomez-Lazaro, M., Pastor, D., Calvo, S., Aguirre,N., Galindo, M.F., Jordan, J., 2005a. Minocycline fails to protectcerebellar granular cell cultures against malonate-induced cell death.Neurobiol. Dis. 20, 384–391.
Fernandez-Gomez, F.J., Galindo, M.F., Gomez-Lazaro, M., Yuste, V.J.,Comella, J.X., Aguirre, N., Jordan, J., 2005b. Malonate induces celldeath via mitochondrial potential collapse and delayed swelling throughan ROS-dependent pathway. Br. J. Pharmacol. 144, 528–537.
Fornstedt, B., Rosengren, E., Carlsson, A., 1986. Occurrence anddistribution of 5-S-cysteinyl derivatives of dopamine, dopa and dopacin the brains of eight mammalian species. Neuropharmacology 25,451–454.
Franke, T.F., Hornik, C.P., Segev, L., Shostak, G.A., Sugimoto, C., 2003.PI3K/Akt and apoptosis: size matters. Oncogene 22, 8983–8998.
Galindo, M.F., Jordan, J., Gonzalez-Garcia, C., Cena, V., 2003. Chromaffincell death induced by 6-hydroxydopamine is independent of mitochon-drial swelling and caspase activation. J. Neurochem. 84, 1066–1073.
Halliwell, B., 1992. Reactive oxygen species and the central nervous system.J. Neurochem. 59, 1609–1623.
Hornykiewicz, O., 1966. Dopamine (3-hydroxytyramine) and brainfunction. Pharmacol. Rev. 18, 925–964.
Jordan, J., Galindo, M.F., Tornero, D., Gonzalez-Garcia, C., Cena, V., 2004.Bcl-x L blocks mitochondrial multiple conductance channel activationand inhibits 6-OHDA-induced death in SH-SY5Y cells. J. Neurochem.89, 124–133.
Kennedy, S.G., Wagner, A.J., Conzen, S.D., Jordan, J., Bellacosa, A.,Tsichlis, P.N., Hay, N., 1997. The PI 3-kinase/Akt signaling pathwaydelivers an anti-apoptotic signal. Genes Dev. 11, 701–713.
Lai, C.T., Yu, P.H., 1997. Dopamine- and L-beta-3,4-dihydroxyphenylala-nine hydrochloride (L-Dopa)-induced cytotoxicity towards catecholami-nergic neuroblastoma SH-SY5Y cells. Effects of oxidative stress andantioxidative factors. Biochem. Pharmacol. 7, 363–372.
Lee, J.Y., Kim, Y.H., Koh, J.Y., 2001. Protection by pyruvate againsttransient forebrain ischemia in rats. J. Neurosci. 21, RC171.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T))method. Methods 25, 402–408.
Love, R.M., Branton, R.L., Karlsson, J., Brundin, P., Clarke, D.J., 2002.Effects of antioxidant pretreatment on the survival of embryonicdopaminergic neurons in vitro and following grafting in an animalmodel of Parkinson's disease. Cell Transplant 11, 653–662.
Mazzio, E.A., Reams, R.R., Soliman, K.F., 2004. The role of oxidativestress, impaired glycolysis and mitochondrial respiratory redox failure inthe cytotoxic effects of 6-hydroxydopamine in vitro. Brain Res. 1004,29–44.
Nie, G., Cao, Y., Zhao, B., 2002. Protective effects of green tea polyphenols
and their major component, (−)-epigallocatechin-3-gallate (EGCG), on6-hydroxydopamine-induced apoptosis in PC12 cells. Redox. Rep. 7,171–177.
Nistico, G., Ciriolo, M.R., Fiskin, K., Iannone, M., De Martino, A., Rotilio,G., 1991. NGF restores decrease in catalase and increases glutathioneperoxidase activity in the brain of aged rats. Neurosci. Lett. 130,117–119.
Noelker, C., Bacher, M., Gocke, P., Wei, X., Klockgether, T., Du, Y., Dodel,R., 2005. The flavanoide caffeic acid phenethyl ester blocks 6-hydroxydopamine-induced neurotoxicity. Neurosci. Lett. 383, 39–43.
Pages, G., Lenormand, P., L'Allemain, G., Chambard, J.C., Meloche, S.,Pouyssegur, J., 1993. Mitogen-activated protein kinases p42mapk andp44mapk are required for fibroblast proliferation. Proc. Natl. Acad. Sci.U. S. A. 90, 8319–8323.
Perovic, M., Mladenovic, A., Rakic, L., Ruzdijic, S., Kanazir, S., 2005.Increase of GAP-43 in the rat cerebellum following unilateral striatal6-OHDA lesion. Synapse 56, 170–174.
Ridet, J.L., Bensadoun, J.C., Deglon, N., Aebischer, P., Zurn, A.D., 2006.Lentivirus-mediated expression of glutathione peroxidase: neuroprotec-tion in murine models of Parkinson's disease. Neurobiol. Dis. 21,29–34.
Salahudeen, A.K., Clark, E.C., Nath, K.A., 1991. Hydrogen peroxide-induced renal injury: a protective role for pyruvate in vitro and in vivo.J. Clin. Invest. 88, 1886–1893.
Sampath, D., Jackson, G.R., Werrbach-Perez, K., Perez-Polo, J.R., 1994.Effects of nerve growth factor on glutathione peroxidase and catalase inPC12 cells. J. Neurochem. 62, 2476–2479.
Selak, I., Skaper, S.D., Varon, S., 1985. Pyruvate participation in the lowmolecular weight trophic activity for central nervous system neurons inglia-conditioned media. J. Neurosci. 5, 23–28.
Shimizu, E., Hashimoto, K., Komatsu, N., Iyo, M., 2002. Roles ofendogenous glutathione levels on 6-hydroxydopamine-induced apopto-tic neuronal cell death in human neuroblastoma SK-N-SH cells.Neuropharmacology 43, 434–443.
Shrieve, D.C., Bump, E.A., Rice, G.C., 1988. Heterogeneity of cellularglutathione among cells derived from a murine fibrosarcoma or a humanrenal cell carcinoma detected by flow cytometric analysis. J. Biol. Chem.263, 14107–14114.
Sian, J., Dexter, D.T., Lees, A.J., Daniel, S., Agid, Y., Javoy-Agid, F.,Jenner, P., Marsden, C.D., 1994. Alterations in glutathione levels inParkinson's disease and other neurodegenerative disorders affectingbasal ganglia. Ann. Neurol. 36, 348–355.
Signore, A.P., Weng, Z., Hastings, T., Van Laar, A.D., Liang, Q., Lee, Y.J.,Chen, J., 2006. Erythropoietin protects against 6-hydroxydopamine-induced dopaminergic cell death. J. Neurochem. 96, 428–443.
Slovin, P.N., Huang, C.J., Cade, J.R., Wood, C.E., Nasiroglu, O., Privette,M., Orbach, P., Skimming, J.W., 2001. Sodium pyruvate is better thansodium chloride as a resuscitation solution in a rodent model of profoundhemorrhagic shock. Resuscitation 50, 109–115.
Song, G., Ouyang, G., Bao, S., 2005. The activation of Akt/PKB signallingpathway and cell survival. J. Cell Mol. Med. 9, 59–71.
Thakar, J.H., Hassan, M.N., 1988. Effects of 6-hydroxydopamine onoxidative phosphorylation of mitochondria from rat striatum, cortex, andliver. Can. J. Physiol. Pharmacol. 66, 376–379.
Thiruchelvam, M., Prokopenko, O., Cory-Slechta, D.A., Richfield, E.K.,Buckley, B., Mirochnitchenko, O., 2005. Overexpression of superoxidedismutase or glutathione peroxidase protects against the paraquat+maneb-induced Parkinson disease phenotype. J. Biol. Chem. 280,22530–22539.
Thor, H., Smith, M.T., Hartzell, P., Bellomo, G., Jewell, S.A., Orrenius, S.,1982. The metabolism of menadione (2-methyl-1,4-naphthoquinone) byisolated hepatocytes. A study of the implications of oxidative stress inintact cells. J. Biol. Chem. 257, 12419–12425.
Tiffany-Castiglioni, E., Saneto, R.P., Proctor, P.H., Perez-Polo, J.R., 1982.Participation of active oxygen species in 6-hydroxydopamine toxicityto a human neuroblastoma cell line. Biochem. Pharmacol. 31,181–188.

307F.J. Fernandez-Gomez et al. / Neurobiology of Disease 24 (2006) 296–307
von Coelln, R., Kugler, S., Bahr, M., Weller, M., Dichgans, J., Schulz, J.B., 2001. Rescue from death but not from functional impairment:caspase inhibition protects dopaminergic cells against 6-hydroxydo-pamine-induced apoptosis but not against the loss of their terminals.J. Neurochem. 77, 263–273.
Votyakova, T.V., Reynolds, I.J., 2001. DeltaPsi(m)-dependent and-independent production of reactive oxygen species by rat brainmitochondria. J. Neurochem. 79, 266–277.
Yamaguchi, K., Lee, S.H., Kim, J.S., Wimalasena, J., Kitajima, S., Baek, S.
J., 2006. Activating transcription factor 3 and early growth response 1are the novel targets of LY294002 in a phosphatidylinositol 3-kinase-independent pathway. Cancer Res. 66, 2376–2384.
Yoon, S., Seger, R., 2006. The extracellular signal-regulated kinase: multiplesubstrates regulate diverse cellular functions. Growth Factors 24, 21–44.
Young, P.R., ConnorsWhite, A.L., Dzido, G.A., 1994. Kinetic analysis ofthe intracellular conjugation of monochlorobimane by IC-21 murinemacrophage glutathione-S-transferase. Biochim. Biophys. Acta 1201,461–465.




















