
Radiofrequency Denervation for Facet Joint Low Back Pain A Systematic Review
Upload
independentCategory
view
2download
0
Consequences of unilateral nigrostriatal denervation on thethalamostriatal pathway in rats
M. S. Aymerich,1,2 P. Barroso-Chinea,1 M. Perez-Manso,1 A. M. Munoz-Patino,3 M. Moreno-Igoa,4 T. Gonzalez-Hernandez5 and J. L. Lanciego1
1Neuromorphology-Tracing Laboratory, Department of Neurosciences, Center for Applied Medical Research, University of NavarraMedical College, Pio XII Avenue no. 55, 31008 Pamplona, Spain2Department of Biochemistry, University of Navarra Medical College, Pamplona, Spain3Department of Morphological Sciences, University of Santiago de Compostela Medical College, Santiago de Compostela, Spain4LAGENBIO-13A, Department of Anatomy, Embryology and Animal Genetics, University of Zaragoza, Zaragoza, Spain5Department of Anatomy, University of La Laguna Medical College, La Laguna, Tenerife, Spain
Keywords: 6-OHDA, basal ganglia, Fluoro-Gold, parafascicular nucleus, Parkinson’s disease, vGLUT2
Abstract
The position of the caudal intralaminar nuclei within basal ganglia circuitry has largely been neglected in most studies dealing withbasal ganglia function. During the past few years, there has been a growing body of evidence suggesting that the thalamicparafascicular nucleus in rodents (PF) exerts a multifaceted modulation of basal ganglia nuclei, at different levels. Our aim was tostudy the activity of the thalamostriatal pathway in rats with unilateral dopaminergic depletion. The experimental approach comprisedfirst unilateral delivery of 6-OHDA in the medial forebrain bundle. Thirty days post-lesioning, animals showing a clear asymmetry werethen subjected to bilateral injection of Fluoro-Gold (FG) within the striatum. Subsequently, expression of the mRNA encoding thevesicular glutamate transporter 2 (vGLUT2) was detected within thalamostriatal-projecting neurons (FG-labeled) by in situhybridization and the results were confirmed by laser-guided capture microdissection microscopy followed by real-time PCR. Thedata showed that there was a marked neuronal loss restricted to PF neurons projecting to the dopamine-depleted striatum. Moreover,PF neurons innervating the dopamine-depleted striatum were intensely hyperactive. These neurons showed a marked increase onthe expression of vGLUT2 mRNA as well as for the mRNA encoding the subunit I of cytochrome oxidase as compared with thoseneurons projecting to the striatum with normal dopamine content. Thus, the selective neurodegeneration of PF neurons innervatingthe striatum together with the increased activity of the thalamostriatal pathway coexist after nigrostriatal denervation.
Introduction
The caudal intralaminar nuclei are the main source of glutamatergicthalamic inputs to the striatum (Smith et al., 2004), and are known toexert a widespread influence on the basal ganglia. Besides striatalinnervation, thalamic inputs arising from the parafascicular nucleus ofthe rodent thalamus (PF) can gain direct access to the globus pallidus(GP; Kincaid et al., 1991; Yasukawa et al., 2004) and the subthalamicnucleus (STN; Feger et al., 1994; Deschenes et al., 1996; Marini et al.,1999; Gonzalo et al., 2002; Lanciego et al., 2004; Castle et al., 2005).Indeed, innervation of the contralateral STN and substantia nigra parsreticulata (SNr) by PF afferents has also been reported (Marini et al.,1999; Castle et al., 2005). Although single tracing studies usingjuxtacellular injections have shown that a subpopulation of PF neuronsprojects simultaneously to both the striatum and STN, via axoncollaterals (Deschenes et al., 1996), thalamostriatal and thalamosub-thalamic projections mostly arise from different PF neurons (Fegeret al., 1994).
There is ongoing debate regarding the role played by the caudalintralaminar nuclei on the pathophysiology of Parkinson’s disease(PD). On the one hand, significant neurodegeneration of the
centromedian-parafascicular thalamic complex (CM-PF) has beendetected in PD (Henderson et al., 2000a, b). This neurodegeneration ishighly specific to the CM-PF complex, given that adjacent thalamicnuclei remain unaffected. These results are in accordance with earlierreports showing similar thalamic degeneration phenomena afterstriatal injection of 1-methyl-4-phenylpyridinium ion (MPP+) in rats(Ghorayeb et al., 2002) or as a consequence of the administration ofN-methyl-4-phenyl-1,2,3,6-tetrahydropiridine (MPTP) to mice (Fre-yaldenhoven et al., 1997). On the other hand, it has also been reportedthat PF neurons projecting to the STN are highly hyperactive inunilaterally dopamine-depleted rats (estimated increase in activity ofup to 62%; Hirsch et al., 2000; Orieux et al., 2000). Indeed, ibotenate-induced lesioning of the PF is highly effective in preventing theincrease in the activity within both the STN and the basal gangliaoutput nuclei that is typically observed after unilateral dopaminedepletion in rats (Bacci et al., 2004). When considering healthyanimals with normal dopamine content, PF lesions induced a number ofchanges in neurotransmitter-related gene expression within most basalganglia-related structures. These changes are probably due to theremoval of direct glutamatergic thalamic innervation (Bacci et al.,2002). After nigrostriatal damage, the existence of selective neurode-generation together with increased activity of the caudal intralaminarnuclei are difficult to conciliate. Our aim was to elucidate further thepotential impact of unilateral dopaminergic depletion on the rodent
Correspondence: Dr J. L. Lanciego, as above.E-mail: [email protected]
Received 9 November 2005, revised 16 February 2006, accepted 16 February 2006
European Journal of Neuroscience, Vol. 23, pp. 2099–2108, 2006 doi:10.1111/j.1460-9568.2006.04741.x
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

thalamostriatal system. The present study sheds some light on this issue,showing that the PF neurons giving rise to the thalamostriatal pathway in6-OHDA-lesioned rats are suffering from a marked neurodegeneration,while simultaneously the surviving neurons are particularly active,probably as a result of a self-compensatory mechanism.
Materials and methods
Male Wistar rats (230–260 g) were used. At all times, animals werehandled according to the European Council Directive 86 ⁄ 609 ⁄ EEC aswell as in agreement with the Society for Neuroscience Policy on theUse of Animals in Neuroscience Research. The experimental designwas approved by the Ethical Committee for Animal Testing at theUniversity of Navarra (Ref. 037 ⁄ 2000).
Surgical procedure and rotational testing
Animals were deeply anesthetized via an i.p. injection of a mixture offour parts of Ketaset (1% solution of ketamine, 75 mg ⁄ kg) and threeparts of Rompun (2% solution of xylazine, 10 mg ⁄ kg). The rats werethen placed in a stereotaxic frame (David Kopf, Tujunga, CA, USA)and a total of 8 lg of the neurotoxin 6-OHDA (2 lg ⁄ lL) wasdelivered at a rate of 0.5 lL ⁄min into the medial forebrain bundle(MFB).Twenty-eight days post-lesioning, the rotational behavior of the
lesioned animals was tested on a computerized rotometer (Panlab,Barcelona, Spain) following i.p. injection of 1 mg ⁄ kg apomorphine(Sigma, St. Louis, MO, USA). Circling behavior exhibited afterapomorphine treatment is directly related to the extent of thenigrostriatal damage induced with 6-OHDA (Carman et al., 1991;Hudson et al., 1993). In this regard, only animals showing more than400 net turns over a period of 90 min (n ¼ 35) were considered to bevalid for the ongoing study.The animals selected were subjected to a second surgical procedure
(after anesthesia with a mixture of ketamine and xylacine, as explainedabove) involving the bilateral delivery of the retrograde tracer Fluoro-Gold (FG) in mirror-like areas within the dorsolateral striatum (Fig. 1)according to the atlas of Paxinos & Watson (1998). The tracer wasiontophoretically delivered as a 2% solution in 0.1 m cacodylate buffer,pH 7.3, using a glass micropipette (inner tip diameter 25–40 lm) and apositive-pulsed direct current (5 lA, 7 s on ⁄ off). In order to depositonly small amounts of FG, the tracer was injected for 5 min and, onceterminated, the micropipette was left in place for 10 min beforewithdrawal. During removal of the micropipette, the current wasreversed to minimize tracer uptake through the injection tract.Animals were divided into two experimental groups. The first
group comprised ten animals that were killed by perfusion (underdeep anesthesia with 10% chloral hydrate in distilled water) andused for dual immunocytochemistry of FG and tyrosine hydroxylase(TH). The second group consisted of 25 animals, killed bydecapitation and then used for the immunofluorescent detection oftransported FG together with the fluorescent, non-radioactive in situhybridization for vesicular glutamate transporter 2 (vGLUT2)mRNA, as well as for laser-guided capture microdissection followedby real-time PCR, as in our hands the use of fresh tissue givesbetter efficiency in mRNA amplification.
Dual immunocytochemical detection of FG and TH
One week post-FG injection (5 weeks after the initial delivery of6-OHDA), ten animals were anesthetized with an overdose of 10%chloral hydrate and then perfused transcardially with saline Ringer’s
solution followed by 500 mL of a cold fixative solution containing 4%paraformaldehyde and 0.1% glutaraldehyde on 0.125 m phosphatebuffer (PB), pH 7.4. After perfusion, the skull was opened, the brainwas removed and it was stored in a cryoprotective solution containing20% glycerin and 2% dimethylsulfoxide in 0.125 m PB, pH 7.4(Rosene et al., 1986). Frozen coronal sections (40 lm thick) wereobtained in a sliding microtome and collected in 0.125 m PB, pH 7.4,in ten series of adjacent sections. Sections containing representativelevels of the striatum and the substantia nigra were used for dualimmunocytochemical processing to detect the retrogradely transportedFG and TH. The sections were first incubated for 60 h at 4 �C in acocktail of primary antisera containing 1 : 2000 rabbit anti-FG(Chemicon, Temecula, CA, USA) and 1 : 1000 mouse anti-TH(Sigma). The sections were then exposed for 2 h at room temperatureto a cocktail of bridge antisera containing 1 : 50 swine anti-rabbit IgG(Dako, Copenhagen, Denmark) and 1 : 50 goat anti-mouse IgG(Dako). The sections were then incubated in a peroxidase–anti-peroxidase (PAP) complex raised in mouse (1 : 600 mouse-PAP,Dako) and TH immunoreactivity was visualized using a regulardiaminobenzidine solution (DAB, Sigma – brown). Sections werefurther exposed to a PAP complex raised in rabbit (1 : 600 rabbit-PAP,Dako) for 90 min at room temperature, and FG-labeled neurons werevisualized following 5 min of incubation in Vector� VIP substrate(Vector Laboratories, Burlingame, CA, USA – purple). The sectionswere mounted on glass slides using a 2% solution of gelatin (Merck,Darmstadt, Germany) in 0.05 m Tris ⁄HCl pH 7.6, and were dried atroom temperature, rapidily dehydrated in toluene and coverslippedwith Entellan (Merck). All antisera used in this procedure were dilutedin 0.05 m TBS, pH 8.0, with 0.5% Triton X-100 (TBS-Tx; Tx fromSigma) and all the incubations with primary antisera were performedin the presence of 2% bovine serum albumin (Merck). Extensivewashing with 0.05 m TBS-Tx, pH 8.0, was carried out at each stage ofthe procedure. Several rinses with 0.05 m Tris ⁄HCl, pH 7.6, wereperformed prior to and after the reactions with the different chromogensolutions. For counting purposes, the neurons displaying FG labelingwithin different thalamic nuclei such as the centrolateral (CL),paracentral (PC) and PF were plotted using a camera lucida. Statisticalanalysis was performed by using Student’s t-test.
Preparation of the riboprobes
Sense and antisense riboprobes were synthesized from a double-stranded DNA fragment corresponding to nucleotides 494–3866 ofrat vGLUT2 (Stornetta et al., 2002). The plasmid was linearizedand transcribed using reagents from Roche (Mannheim, Germany).The 50-lL reaction mixture included 1 lg of template cDNA,1 mm of each ATP, CTP and GTP, 0.7 mm UTP and 0.3 mm
digoxigenin-UTP; 10 mm dithiothreitol; 50 U of RNase inhibitor;and 1 U of either T7 or SP6 RNA polymerase. After 2 h at 37 �C,the template cDNA was digested with 2 U of RNase-free DNAsefor 30 min at 37 �C. The riboprobes were then precipitated by theaddition of 100 lL of 4 m ammonium acetate and 500 lL ofethanol and centrifuged at 4 �C for 30 min. The quality of thesynthesis was tested by dot blot.
In situ hybridization for vGLUT2 combined with the detectionof transported FG
Animals (n ¼ 25) were killed by decapitation 1 week post-FGinjection (5 weeks after initial delivery of 6-OHDA). The brains wereremoved from the skull and quickly frozen in isopentane cooled with
2100 M. S. Aymerich et al.
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108
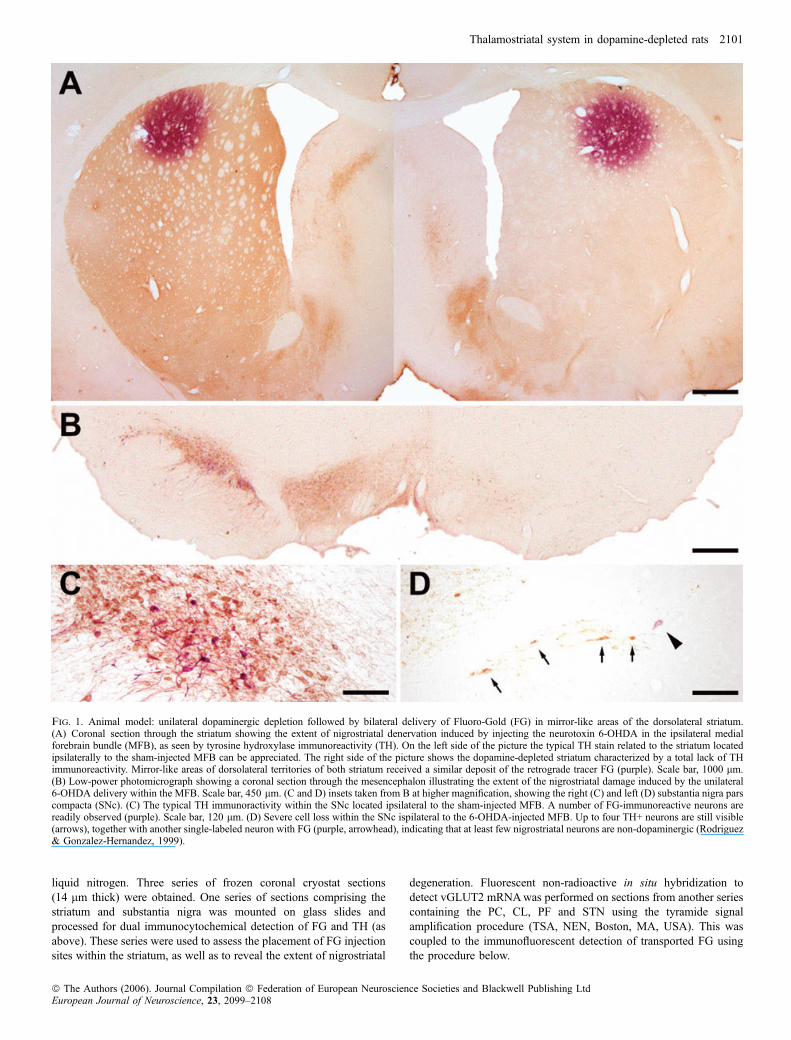
liquid nitrogen. Three series of frozen coronal cryostat sections(14 lm thick) were obtained. One series of sections comprising thestriatum and substantia nigra was mounted on glass slides andprocessed for dual immunocytochemical detection of FG and TH (asabove). These series were used to assess the placement of FG injectionsites within the striatum, as well as to reveal the extent of nigrostriatal
degeneration. Fluorescent non-radioactive in situ hybridization todetect vGLUT2 mRNAwas performed on sections from another seriescontaining the PC, CL, PF and STN using the tyramide signalamplification procedure (TSA, NEN, Boston, MA, USA). This wascoupled to the immunofluorescent detection of transported FG usingthe procedure below.
Fig. 1. Animal model: unilateral dopaminergic depletion followed by bilateral delivery of Fluoro-Gold (FG) in mirror-like areas of the dorsolateral striatum.(A) Coronal section through the striatum showing the extent of nigrostriatal denervation induced by injecting the neurotoxin 6-OHDA in the ipsilateral medialforebrain bundle (MFB), as seen by tyrosine hydroxylase immunoreactivity (TH). On the left side of the picture the typical TH stain related to the striatum locatedipsilaterally to the sham-injected MFB can be appreciated. The right side of the picture shows the dopamine-depleted striatum characterized by a total lack of THimmunoreactivity. Mirror-like areas of dorsolateral territories of both striatum received a similar deposit of the retrograde tracer FG (purple). Scale bar, 1000 lm.(B) Low-power photomicrograph showing a coronal section through the mesencephalon illustrating the extent of the nigrostriatal damage induced by the unilateral6-OHDA delivery within the MFB. Scale bar, 450 lm. (C and D) insets taken from B at higher magnification, showing the right (C) and left (D) substantia nigra parscompacta (SNc). (C) The typical TH immunoractivity within the SNc located ipsilateral to the sham-injected MFB. A number of FG-immunoreactive neurons arereadily observed (purple). Scale bar, 120 lm. (D) Severe cell loss within the SNc ispilateral to the 6-OHDA-injected MFB. Up to four TH+ neurons are still visible(arrows), together with another single-labeled neuron with FG (purple, arrowhead), indicating that at least few nigrostriatal neurons are non-dopaminergic (Rodriguez& Gonzalez-Hernandez, 1999).
Thalamostriatal system in dopamine-depleted rats 2101
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

After fixation for 10 min in 4% paraformaldehyde, the sectionswere incubated twice in 0.1% active DEPC ⁄ PB for 15 min. Afterpre-equilibrating in 5· SSC (0.75 m NaCl, 0.0075 m Na citrate), thesections were prehybridized at 58 �C for 1 h in the hybridizationsolution (50% formamide, 5· SSC, 40 lg ⁄mL denatured salmonDNA). The vGLUT2 probe was denatured for 5 min at 75 �C beforeadding it to the hybridization mix at a concentration of 400 ng ⁄mL,and the sections were then hybridized in this solution at 58 �C for16 h. The sections were then washed in 2· SSC at room temperaturefor 30 min, 2· SSC at 65 �C for 1 h and 0.1· SSC at 65 �C for 1 h.The slides were then equilibrated for 5 min in TN buffer (100 mm
Tris ⁄HCl, 150 mm NaCl, pH 7.5) and incubated with a peroxidase-conjugated anti-digoxigenin antibody for 30 min at room temperature(1 : 100 in TNB blocking buffer, Roche). Fluorescent labeling wasthen visualized using streptavidin coupled with Alexa� 546 (Molecu-lar Probes, Eugene, OR, USA). Once the in situ hybridization hadterminated, we carried out the FG detection protocol using a primaryantiserum against FG (1 : 2000 rabbit anti-FG, Chemicon) and abridge antiserum coupled with Alexa� 488 (1 : 50 goat anti-rabbitIgG Alexa� 488, Molecular Probes). After several rinses in PB, theslides were left to dry at room temperature overnight, before they werecleared in toluene and coverslipped with DPX (BDH Chemicals,Poole, UK) and finally analysed by confocal laser-scanning micros-copy (Zeiss 510 META). Neurons showing co-localization betweenvGLUT2 mRNA and FG were randomly selected (up to 20 neuronswithin each PF nucleus), and the intensity of vGLUT2 labeling wasvisualized using pseudocolor bidimensional densitometry (see Fig. 3).In order to ensure the labeled elements were appropriately visualizedand to avoid false positive results, the emission from the argon laser at488 nm was filtered through a bandpass filter of 505–530 nm, thisemission being color-coded in green. The emission following theexcitation with the helium laser at 543 was filtered through a bandpassfilter of 560–615 nm, and colour-coded in red. The sections were firstexamined using low-magnification lenses (·10 and ·20), andphotographs were then taken at higher magnification (·40 oil-immersion objective, NA 1.3, and ·63 oil-immersion objective,NA 1.4).
Laser-guided capture microdissection followed by real-timePCRThe third series of cryostat sections from the thalamic levels similar tothose subjected to in situ hybridization were used for microdissectionpurposes. These sections were mounted on plastic-coated slides(Leica). Thalamic areas showing FG labeling were visualized underdirect epifluorescent UV illumination (see Fig. 4), and were dissectedout and collected in 0.5-mL tubes containing the lysis buffer for RNAextraction. Total RNA was extracted using the Absolutely RNA�nanoprep kit (Stratagene, La Jolla, CA, USA) according to themanufacturer’s instructions, including the optional DNase I digestionstep. The RNAwas eluted in a final volume of 10 lL and was all usedin reverse transcription reactions. The cDNA was obtained by adding1 mm dNTP mix, 2 mm DTT, 5 pm hexamers, 40 U RNase inhibitor(Promega, Madison, WI, USA) and 200 U M-MLV reverse transcrip-tase (Promega) in a final volume of 30 lL and incubated at 37 �C for1 h. PCR reactions were carried out in a final volume of 24 lLcontaining 150 nm of each primer, 12 lL of 2· SYBR Green PCRMaster Mix (Applied Biosystems, Foster City, CA, USA) and 1 lLper reaction of cDNA for the amplification of the reference generps18, 2.5 lL per reaction for the amplification of vGLUT2 and0.5 lL for the subunit I of cytochrome oxidase (CO-I). All PCRreactions were performed in triplicate using an Opticon2TM (MJResearch, Waltham, MA, USA) cycler for real-time detection ofamplified dsDNA with SYBR Green. The thermal cycling parameterswere as follows: incubation at 95 �C for 10 min, 40 cycles of 95 �Cfor 15 s and 60 �C for 1 min, and a melting curve from 60 to 95 �C toensure unique product amplification. The data obtained were analysedusing the Q-Gene Core Module (Muller et al., 2002). The relativeexpression of vGLUT2 and CO-I was normalized using rps18 as areference gene and taking into account the amplification efficiencies ofeach pair of primers (see Table 1). The normalized levels of vGLUT2and CO-I from thalamic nuclei located on the lesioned side were thencompared with the normalized levels from the thalamic nucleiipsilateral to the sham-injected side. Statistical analysis was accom-plished by using the Wilcoxon test, as the data did not follow a normaldistribution.
Table 1. Primers used for real-time PCR
Gene Accession no. Sequences 5¢3¢Fragmentsize (bp)
Amplificationefficiency
Ribosomalprotein 18S
NM213557 F-CCTTCGCTATCACTGCCATT 222 2.705R-TGGCCAGAACCTGGCTATAC
vGLUT2 AF271235 F-AAGAAACGGGGGACATCACT 137 2.78R-GTCTTGCGCACTTTCTTGC
CO-I X14848 F-GAGGCTTCGGAAACTGACTTGT 145 2.67R-TGTTCCAGCTCCAGCTTCTACT
Fig. 2. Degeneration of the thalamostriatal pathway after dopaminergic depletion. (A) Histogram showing the number of FG-positive neurons within theparafascicular (PF), centrolateral (CL) and paracentral (PC) thalamic intralaminar nuclei. FG-labeled cells from PF, CL and PC of the sham side and the lesioned sidewere plotted with a camera lucida and counted. Statistical analysis was performed using a Student’s t-test (n ¼ 7). Highly significant changes were only found in PF(***P < 0.001). Cell counts performed in CL and PC failed to reach statistical significance (n.s.). (B) Coronal section through the right thalamus at the level of thelateral intralaminar (CL and PC) and midline nuclei (CM), showing the number of thalamostriatal neurons retrogradely labeled after the FG deposit shown inFig. 1A. Scale bar, 360 lm. (C) Photomicrograph taken from the left thalamus, at a similar level to the one shown in B. After an FG deposit in dopamine-depletedareas of the dorsolateral striatum (as seen in Fig. 1A), no differences in the number of FG-labeled neurons were apparent. Scale bar, 360 lm. (D) Coronal sectionthrough the right PF nucleus, showing the number of thalamostriatal neurons retrogradely labeled after the FG deposit shown in Fig. 1A. Scale bar, 360 lm.(E) Photomicrograph taken from the left PF nucleus, at a similar level to the one shown in D. After an FG deposit in dopamine-depleted areas of the dorsolateralstriatum (as seen in Fig. 1A), a decrease on the number of FG-labeled neurons was apparent, indicating a cell loss of thalamostriatal projecting neurons undercircumstances of dopaminergic depletion. Scale bar, 360 lm. A small portion of the fasciculus retroflexus (fr) is delineated for reference. Abbreviations for thalamicnuclei: CL (centrolateral), PC (paracentral), CM (central medial), MD (mediodorsal), VM (ventromedial), Po (posterior thalamic nuclear group).
2102 M. S. Aymerich et al.
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

Thalamostriatal system in dopamine-depleted rats 2103
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

Results
Degeneration of the thalamostriatal pathway in6-OHDA-treated rats
Initially, ten rats were lesioned with 6-OHDA. After conducting theapomorphine test, one animal failed to show any clear rotationalbehavior and was not studied further. In another two animals, thebilateral FG deposits were located in different striatal territories and
these animals were not also considered for the study. The datapresented below are for remaining seven animals. An example of thebilateral delivery of FG in mirror-like areas of the dorsolateral striatumis illustrated in Fig. 1. All injections led to the appearance ofretrogradely labeled cell bodies in their respective ipsilateral intrala-minar nuclei (Fig. 2B–E). As expected, the neurons labeled by tracerdelivery in the dorsolateral striatum were preferentially located in thedorsolateral territories of the PF nucleus, in accordance with the
Fig. 3. Fluorescent, non-radioactive in situ hybridization for vGLUT2. (A and B) Detection of vGLUT2 mRNA in the right (A) and left (B) PF nucleus. The PFneurons ipsilateral to the dopamine-depleted side of the brain displayed a higher expression of the vGLUT2 mRNA. Scale bars, 120 lm. (C and D) Expression ofvGLUT2 mRNA in the right (C) and left (D) subthalamic nucleus (STN) used as a reference nucleus. STN neurons ipsilateral to the 6-OHDA injection in the MFBexpress more vGLUT2 mRNA. Scale bars, 120 lm. (E–H) Color-coded, bi-dimensional densitometry of panels A–D, respectively, to illustrate better the differencesin vGLUT2 mRNA expression between left and right PFs and STNs. Scale bars, 120 lm. (I–L) Expression of vGLUT2 mRNA (I) within a single PF neuronprojecting to the striatum with normal dopamine content, retrogradely labeled with FG (J). (K) Pseudocolor bi-dimensional densitometry for vGLUT2 mRNA;(L) the co-localization between the ‘green’ and the ‘red’ channels. Scale bars, 20 lm. (M–P) Expression of vGLUT2 mRNA (M) within a single PF neuroninnervating the dopamine-depleted striatum, identified by means of retrograde transport of FG (N). The pseudocolor bi-dimensional densitometry is illustrated Oand the co-localization of FG and vGLUT2 mRNA is shown in P. Scale bars, 15 lm.
2104 M. S. Aymerich et al.
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

established topography of the thalamostriatal pathway arising from thePF nucleus (Lanciego et al., 2004). Retrograde-labeled cell bodieswere also found throughout other midline and intralaminar nuclei, forexample in the central medial (CM), PC and CL thalamic nuclei(Groenewegen & Berendse, 1994). Indeed, a few neurons were alsofound in the ventral thalamic nuclei (McFarland & Haber, 2001).
The number of FG-labeled neurons within the PF nucleus ipsilateralto the 6-OHDA lesion site was smaller than the number of PF-efferentneurons projecting to the striatum with a normal dopamine content(Figs 2D and E, and 4). The average cell loss was estimated to be 69.3%of the PF neurons sending afferents to the dopamine-depleted striatum(Fig. 2A). This is in accordance with the cell loss induced in the caudalintralaminar nuclei after nigrostriatal damage by using differenttechniques in rodents (Freyaldenhoven et al., 1997; Ghorayeb et al.,2002), as well as in PD patients (Henderson et al., 2000a, b). Nodifferences in the number of thalamostriatal-projecting neurons werefound when considering the other lateral intralaminar nuclei underexamination, such as CL and PC (Fig. 2A).
Expression of vGLUT2 mRNA in the cells of originof the thalamostriatal pathway
Another important aim of this study was to assess the changes ofneuronal activity in cells that originate the thalamostriatal pathway. Asthe PF is a glutamatergic nucleus, it was of interest to use a molecularmarker related to glutamate metabolism to evaluate the activity ofthese neurons. The expression of vGLUT2 mRNAwas chosen for thispurpose, as vGLUT2 is the specific vesicular glutamate transporterused by thalamic neurons at these levels (Fremeau et al., 2001; Herzoget al., 2001; Hur & Zaborszky, 2005).
Twenty-five animals were subjected to unilateral lesioning with6-OHDA and bilateral injections of FG in the striatum. Seven rats hadto be excluded from the study due to either the misplacement of thebilateral FG injections within similar striatal levels (n ¼ 4) or corticalcontamination by track labeling (n ¼ 3). Thus, the data below weregenerated from the analysis of 18 rats.
The tissue obtained was processed for either in situ hybridization orfor laser-guided capture microdissection followed by real-time PCR.Expression of vGLUT2 mRNA determined by in situ hybridizationwas considerably greater in the PF from the lesion side than the PFfrom the sham side (Fig. 3A, B, E and F). As an internal positivecontrol of the experiment we used the STN present in the same sectionas the PF nucleus. The intensity of the vGLUT2 mRNA in situhybridization in the dopamine-depleted side was much higher than inthe sham-injected side (Fig. 3C, D, G and H). Hyperactivity of theSTN upon dopamine depletion is widely accepted to be one of thecornerstones of the current basal ganglia model (Albin et al., 1989;DeLong, 1990). All FG-labeled neurons also expressed vGLUT2mRNA, confirming the glutamatergic nature of the thalamostriatalpathway (Fig. 3I–P). By using pseudocolor bidimensional densitom-etry, the intensity of vGLUT2 mRNA expression was observed withinthe neuronal profiles labeled by retrograde transport with FG (Fig. 3Kand O). In this regard, FG-labeled neurons projecting to the dopamine-depleted striatum were typically characterized by higher levels ofvGLUT2 mRNA expression than FG-labeled neurons projecting to thestriatum with normal dopamine content.In order to quantify these changes in gene expression accurately,
microdissected tissue samples containing projection neurons wereanalysed via real-time PCR. The typical appearance of thalamostriatal-projecting neurons within the PF (FG-labeled) as seen under the laser-guided capture microdissection microscope is illustrated in Fig. 4.Small PF territories comprising the thalamostriatal-projecting neurons(Fig. 4) were dissected out with the laser beam, and collected in a tubefor RNA extraction and real-time PCR analysis. From each nucleus(PF, CL and PC), we amplified vGLUT2, CO-I and the ribosomalprotein S18 (rps18). The rps18 was the reference gene used tonormalize the cycle threshold (Ct) values obtained for CO-I andvGLUT2. CO-I mRNA has been widely used as a general marker ofneuronal activity (Wong-Riley, 1989; Hevner & Wong-Riley, 1991,1993; Vila et al., 1996, 1997, 2000), and therefore we used CO-Iexpression as an internal marker of activity changes within the PF orthe other nuclei studied. Changes in vGLUT2 mRNA are interesting
Fig. 4. The use of the laser-guided capture microdissection microscope to dissect out small PF territories containing thalamostriatal-projecting neurons. Underdirect epifluorescent illumination with the appropriate filter combination, FG-positive neurons are visible (green fluorescent neurons in A and B), and these neuronswere taken as the reference to define the boundaries of the PF areas to be dissected out (red contours). The existence of ongoing degeneration of PF neuronsinnervating the dopamine-depleted striatum was also apparent by comparing the amount of labeled neurons in A, neurons projecting to a striatum with normaldopamine content, with those FG-positive neurons in B, neurons innervating the dopamine-depleted striatum. Taking advantage of the laser-guided capturemicrodissection microscope, only PF territories displaying FG labeling were dissected out while other PF territories containing unlabeled neurons remaineduntouched.
Thalamostriatal system in dopamine-depleted rats 2105
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

as they might correlate with changes in excitatory activity of thenuclei. Although all of the nuclei studied, PF, CL and PC, containedneurons projecting to the striatum (Fig. 5), only the PF displayedhighly significant changes in the levels of vGLUT2 and CO-I mRNAexpression. In the dopamine-depleted side there was a clear increasefor both markers, a two-fold increase in CO-I and a three-fold increasein vGLUT2 mRNA expression. By contrast, the differences observedin the CL and the PC nuclei, which project to the same regions of thestriatum, were not significant. It is important to note that changes inexpression of vGLUT2 correlate with changes in CO-I, and are evenhigher than this marker, suggesting that mRNA levels of vGLUT2could be a good marker of glutamatergic activity. These results, inconjunction with the observations by in situ hybridization labeling,indicate that the PF and not other intralaminar thalamic nuclei areoveractive in a rat model of PD.
Discussion
The current study has assessed the impact that nigrostriatal denerva-tion has on the thalamostriatal projections arising from the caudalintralaminar nuclei. On the one hand, this denervation causes a loss ofPF neurons innervating the dopamine-depleted striatum and on theother there is a marked increase in the metabolic activity ofthalamostriatal-projecting neurons. This increase in activity could bea compensatory mechanism as surviving neurons may try to maintainthalamic glutamate outflow to the striatum within normal levels. At thestriatal level, the balance between glutamate and dopamine plays animportant role in synaptic transmission. Striatal glutamatergic functionis enhanced after nigrostriatal damage (Meshul et al., 2000), resultingin a reorganization of corticostriatal projections (Cho et al., 2002) andreinforcing specific sets of corticostriatal synaptic connections (Bam-ford et al., 2004). Although the reactive changes in striatal glutamatetransmission described in response to dopamine denervation are often
interpreted in terms of changes in corticostriatal transmission, it mightalso be worth considering that the thalamostriatal system could play akey role.
Degeneration and increased activity of the thalamostriatalpathway after dopamine depletion: implications for basalganglia function in PD
The role of the thalamostriatal system in the basal ganglia circuitryunder normal and pathological conditions remains poorly under-stood. Brain circuits linking the caudal intralaminar nuclei with thebasal ganglia are well characterized (Smith et al., 2004),demonstrating that the PF nucleus is connected with virtually allipsilateral basal ganglia-related nuclei (Kincaid et al., 1991; Fegeret al., 1994; Deschenes et al., 1996; Marini et al., 1999;Gonzalo et al., 2002; Lanciego et al., 2004; Yasukawa et al.,2004; Castle et al., 2005), as well as with the contralateral STNand SNr (Marini et al., 1999; Castle et al., 2005). Thus, theincreased PF activity may induce a cascade of events related toglutamate overflow reaching the cortex, striatum, both segments ofthe globus pallidus, STN and SNr. In this regard, the caudalintralaminar nuclei could be considered as a potential candidate toexplain the increased activity of the STN in PD. Another candidatemight be the pedunculopontine nucleus (PPN; Orieux et al., 2000),at least in the initial stages of the disease (Gonzalo et al., 2002). Itis important to determine whether the hyperactivity of the PFnucleus is the result of a self-compensatory mechanism to maintainnormal thalamic glutamate levels in target structures or whether it isdriven by external influences, such as excitatory inputs comingfrom the PPN. The PPN projects massively to the PF, and pontineafferents are closely associated with PF neurons that innervate thestriatum (Erro et al., 1998). Neurons within the PPN innervating theSTN are known to be highly active in dopamine-depleted rats(Orieux et al., 2000), and this might also be the case forPPN neurons afferentiating thalamostriatal-projecting neurons withinthe PF.Neurodegeneration phenomena involving the caudal intralaminar
nuclei have been observed in several neurodegenerative diseases, suchas PD, progressive supranuclear palsy and Huntington’s disease(Heinsen et al., 1996; Henderson et al., 2000a, b; Kassubek et al.,2005). When considering animal models of PD, cell loss has beendetected in the intralaminar nuclei of MPTP-treated mice (Freyal-denhoven et al., 1997) as well as in the PF of dopamine-depleted ratsafter the intrastriatal delivery of MPP+ (the so-called ‘double toxinmodel’; see Ghorayeb et al., 2002). Efferent PF neurons that innervatethe STN in rats do not seem to degenerate after nigrostriataldenervation (Orieux et al., 2000). Therefore, according to the datapresented here, cell loss is most likely to be restricted to PF efferentneurons that target the striatum. The data available from human studiesprovide evidence regarding the existence of a primary, non-dopam-inergic neurodegeneration restricted to the caudal intralaminar nuclei.Thus, it is now important to define whether this thalamic degenerationappears before or after the nigrostriatal damage, as well as to ascertainif there is a relationship between both kinds of neuronal loss. In humanPD brains, it appears that intralaminar thalamic degeneration is alreadypresent by Hoehn and Yahr stages 2–3 PD, and it remains apparentlyunchanged in stages 4–5 PD (Henderson et al., 2005). In the presentstudy, thalamostriatal degeneration is obviously a retrograde change tonigrostriatal damage, and this is also the case for the intralaminarthalamic cell loss observed after MPTP delivery in mice (Freyaldenh-oven et al., 1997).
Fig. 5. Histogram showing changes in vGLUT2 and CO-I mRNA expression.After microdissection of different intralaminar thalamic nuclei (PF, parafasci-cular; CL, centrolateral; PC, paracentral), changes in expression of CO-I andvGLUT2 mRNA were analysed by real-time PCR and Ct values normalizedwith respect to the control gene, the ribosomal protein S18 (rps18). Resultsobtained are expressed as relative amounts with respect to the sham side.Statistical analysis was performed using a Wilcoxon test: ***P < 0.001,**P < 0.05, comparing values from the lesioned side with sham values. PF,n ¼ 14; CL, n ¼ 8; PC, n ¼ 8.
2106 M. S. Aymerich et al.
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

Are the caudal intralaminar nuclei a potential surgicaltarget for PD?
Over the past few years, evidence from both animal models andhumans suggests that the caudal intralaminar nuclei could beconsidered as a potential surgical target for movement disorders ofbasal ganglia origin. It was demonstrated that PF neurons projecting tothe STN are hyperactive in unilaterally dopamine-depleted rats(Orieux et al., 2000), indicating that PF neurons may well play akey role in the characteristic increase of activity observed in the STNunder pathological conditions. In a similar animal model of PD, Bacciet al. (2004) showed that the chemical lesioning of PF is capable ofpreventing the increase in metabolic activity typically observed in boththe output nuclei and the STN. Taken together, these data showinghyperactivity of the PF under circumstances of dopaminergic deple-tion could be considered as the rationale sustaining the potential use ofCM-PF for functional surgery in PD. In this regard, when consideringpioneer clinical experiences carried out in PD patients, several studieshave reported that approaching the CM-PF complex with a deep brainstimulation electrode alleviated several cardinal symptoms of thedisease, such as levodopa-induced dyskinesias and resting tremor(Caparros-Lefebvre et al., 1994, 1999a, b; Krauss et al., 2002; Weigel& Krauss, 2004). This approach has also been shown to have apositive influence on severe forms of Tourette’s syndrome (Houetoet al., 2005) as well as in the management of chronic pain (Krausset al., 2002; Weigel & Krauss, 2004). Although there are a number ofreports showing beneficial effects of targeting the CM-PF complexwith a DBS electrode, we should keep in mind that we are dealingwith a thalamic target experiencing ongoing degeneration phenomena.In this regard, a case report showing the deteriorating effects afterDBS in CM-PF was published by Henderson et al. (2001).
In conclusion, the data presented here indicate that thalamostriatal-projecting neurons suffer marked degeneration following dopaminedepletion. Furthermore, surviving PF neurons innervating the striatumbecome hyperactive. This is probably the result of a self-compensatorymechanism, although the role played by external influences, such asPPN excitatory projections targeting the PF, needs to be clarified.Although more effort is needed to determine the feasibility of usingthe caudal intralaminar nuclei as a surgical target for movementdisorders of basal ganglia origin, the data presented here, together withevidence from the literature, call for a reappraisal of the role played bythe caudal intralaminar nuclei in basal ganglia function, both in normaland in pathological conditions.
Acknowledgements
We thank Drs Patrice Guyenet and Ruth L. Stornetta from the Department ofPharmacology, University of Virginia, Charlottesville, VA, for kindly supplyingus with the vGLUT2 plasmid. Expert technical assistance was supplied by MsElvira Roda and by Ms Carmen Molina. This work was supported by grantsfrom the Spanish Department of Science and Technology, ref. BFI2003-02033,Fondo de Investigaciones Sanitarias, ref. P1051037, Department of Educationof the Government of Navarra, ref. 18 ⁄ 2005, and by the UTE-project ⁄ Foun-dation for Applied Medical Research (F.I.M.A).
Abbreviations
CL, centrolateral; CM, central medial; CM-PF, centromedian-parafascicularthalamic complex; CO-I, subunit I of cytochrome oxidase; FG, Fluoro-Gold;GP, globus pallidus; MFB, medial forebrain bundle; MPP+, 1-methyl-4-phenylpyridinium ion; MPTP, N-methyl-4-phenyl-1,2,3,6-tetrahydropiridine;PAP, peroxidase–anti-peroxidase; PB, phosphate buffer; PC, paracentral; PD,Parkinson’s disease; PF, thalamic parafascicular nucleus; PPN, pedunculopon-tine nucleus; SNr, substantia nigra pars reticulata; STN, subthalamic nucleus;TH, tyrosine hydroxylase; vGLUT2, vesicular glutamate transporter 2.
References
Albin, R.L., Young, A.B. & Penney, J.B. (1989) The functional anatomy ofbasal ganglia disorders. Trends Neurosci., 12, 366–375.
Bacci, J.J., Kachidian, P., Kerkerian-Le Goff, L. & Salin, P. (2004) Intralaminarthalamic nuclei lesions: widespread impact on dopamine denervation-mediated cellular defects in the rat basal ganglia. J. Neuropathol. Exp.Neurol., 63, 20–31.
Bacci, J.J., Kerkerian-Le Goff, L. & Salin, P. (2002) Effects of intralaminarthalamic nuclei lesion on glutamic acid decarboxylase (GAD65 and GAD67)and cytochrome oxidase subunit I mRNA expression in the basal ganglia ofthe rat. Eur. J. Neurosci., 15, 1918–1928.
Bamford, N.S., Zhang, H., Schmitz, Y., Wu, N.P., Ccepeda, C., Levine, M.S.,Schmauss, C., Zakharenkko, S.S., Zablow, L. & Sulzer, D. (2004)Heterosynaptic dopmaine neurotransmission selects sets of corticostriatalterminals. Neuron, 42, 653–663.
Caparros-Lefebvre, D., Blond, S., Feltin, M.P., Pollak, P. & Benabid, A.L.(1999b) Improvement of levodopa induced dyskinesias by thalamic deepbrain stimulation is related to slight variations in electrode placement:possible involvement of the centre median parafascicularis complex.J. Neurol. Neurosurg. Psychiatr., 67, 308–314.
Caparros-Lefebvre, D., Pollak, P., Feltin, M.P., Blond, S. & Benabid, A.L.(1999a) Effet de la stimulation thalamique sur les dyskinesies induites par laL dopa. Evaluation d’une cibe nouvelle: le centre median parafasciculaire.Rev. Neurol., 155, 543–550.
Caparros-Lefebvre, D., Ruchoux, M.M., Blond, S., Petit, H. & Percheron, G.(1994) Long-term thalamic stimulation in Parkinson’s disease: postmortemanatomoclinical study. Neurology, 44, 1856–1860.
Carman, L.S., Gage, F.H. & Shults, C.W. (1991) Partial lesion of the substantianigra: relation between extent of lesion and rotational behavior. Brain Res.,553, 275–283.
Castle, M., Aymerich, M.S., Sanchez-Escobar, C., Gonzalo, N., Obeso, J.A. &Lanciego, J.L. (2005) Thalamic innervation of the direct and indirect basalganglia pathways in the rat: ipsi- and contralateral projections. J. Comp.Neurol., 483, 143–153.
Cho, J., Duke, D., Manzino, L., Sonsalla, P.K. & West, M.O. (2002) Dopmainedepletion causes fragmented clustering of neurons in the sensorimotorstriatum: evidence of lasting reorganization of corticostriatal input. J. Comp.Neurol., 452, 24–37.
DeLong, M.R. (1990) Primate models of movement disorders of basal gangliaorigin. Trends Neurosci., 13, 266–271.
Deschenes, M., Bourassa, J., Van Diep, D. & Parent, A. (1996) A single-cellstudy of axonal projections arising from the posterior intralaminar nuclei inthe rat. Eur. J. Neurosci., 8, 140–143.
Erro, E., Lanciego, J.L. & Gimenez-Amaya, J.M. (1998) Relationships betweenthalamostriatal neurons and pedunculopontine projections to the thalamus: aneuroanatomical tract-tracing study in the rat. Exp. Brain Res., 127, 162–170.
Feger, J., Bevan, M. & Crossman, A.R. (1994) The projections from theparafascicular thalamic nucleus to the subthalamic nucleus and the striatumarise from separated neuronal populations: a comparison with thecorticostriatal and corticosubthalamic efferents in a retrograde fluorescentdouble-labelling study. Neuroscience, 60, 125–132.
Fremeau, R.T. Jr, Troyer, M.D., Pahner, I., Nygaard, G.O., Tran, C.H., Reimer,R.J., Bellochio, E.E., Fortin, D., Storm-Mathisen, J. & Edwards, R.H. (2001)The expression of vesicular glutamate transporters defines two classes ofexcitatory synapses. Neuron, 31, 247–260.
Freyaldenhoven, T.E., Ali, S.F. & Schmued, L.C. (1997) Systemicadministration of MPTP induces thalamic degeneration in mice. BrainRes., 759, 9–17.
Ghorayeb, I., Fernagut, P.O., Hervier, L., Labattu, B., Bioulac, B. & Tison, F.(2002) A ‘single toxin-double lesion’ rat model of striatonigral degenerationby intrastriatal 1-methyl-4-phenylpyridinium ion injection: a motor beha-vioural analysis. Neuroscience, 115, 533–546.
Gonzalo, N., Lanciego, J.L., Castle, M., Vazquez, A., Erro, E. & Obeso, J.A.(2002)The parafascicular thalamic complex and basal ganglia circuitry: furthercomplexity to the basal ganglia model. Thalamus Rel. Sys., 1, 341–348.
Groenewegen, H.J. & Berendse, H.W. (1994) The specificity of the ‘nonspe-cific’ midline and intralaminar nuclei. Trends Neurosci., 17, 52–57.
Heinsen, H., Rub, U., Gangnus, D., Jungkunz, G., Bauer, M., Ulmar, G.,Bethke, B., Schuler, M., Bocker, F., Eisenmenger, W., Gotz, M. & Strik, M.(1996) Nerve cell loss in the thalamic centromedian-parafascicular complexin patients with Huntington’s disease. Acta Neuropathol., 91, 161–168.
Henderson, J.M., Carpenter, K., Cartwright, H. & Halliday, G.M. (2000a) Lossof thalamic intralaminar nuclei in progressive supranuclear palsy andParkinson’s disease: clinical and therapeutic implications. Brain, 123, 1410–1421.
Thalamostriatal system in dopamine-depleted rats 2107
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108

Henderson, J.M., Carpenter, K., Cartwright, H. & Halliday, G.M. (2000b)Degeneration of the centre median-parafascicular complex in Parkinson’sdisease. Ann. Neurol., 47, 345–352.
Henderson, J.M., O’Sullivan, D.J., Fung, V.S., Hely, M.A., Morris, J.G. &Halliday, G.M. (2001) Lesion of thalamic centromedian-parafascicularcomplex after chronic deep brain stimulation. Neurology, 56, 1576–1579.
Henderson, J.M., Schleimer, S.B., Allbutt, H., Dabholkar, V., Abela, D., Jovic,J. & Quinlivan, M. (2005) Behavioral effects of parafascicular thalamiclesions in an animal model of parkinsonism. Behav. Brain Res., 162, 222–232.
Herzog, E., Bellenchi, G.C., Gras, C., Bernard, V., Ravassard, P., Bedet, C.,Gasnier, B., Giroz, B. & El Mestikawy, S. (2001) The existence of a secondvesicular glutamate transporter specifies subpopulations of glutamatergicneurons. J. Neurosci., 21, RC181 (1–6).
Hevner, R.F. & Wong-Riley, M.T. (1991) Neuronal expression of nuclear andmitochondrial genes for cytochrome oxidase (CO) subunits analyzed byin situ hybridization: comparison with CO activity and protein. J. Neurosci.,11, 1942–1958.
Hevner, R.F. & Wong-Riley, M.T. (1993) Mitochondrial and nuclear geneexpression for cytochrome oxidase subunits are disproportionately regulatedby functional activity in neurons. J. Neurosci., 13, 1805–1819.
Hirsch, E.C., Perier, C., Orieux, G., Francois, C., Feger, J., Yelnik, J., Vila, M.,Levy, R., Tolosa, E.S., Marin, C., Herrero, M.T., Obeso, J.A. & Agid, Y.(2000) Metabolic effects of nigrostriatal denervation in basal ganglia. TrendsNeurosci., 23, 78–85.
Houeto, J.L., Karachi, C., Mallet, L., Pilon, B., Yelnik, J., Mesnage, V., Welter,M.L., Navarro, S., Pelissolo, A., Damier, P., Pidoux, B., Dormont, D., Cornu,P. & Agid, Y. (2005) Tourette’s syndrome and deep brain stimulation.J. Neurol. Neurosurg. Psychiatry, 76, 992–995.
Hudson, J.L., van Horne, C.G., Stromberg, I., Brock, S., Clayton, J., Masserano,J., Hoffer, B.J. & Gerhardt, G.A. (1993) Correlation of apomorphine- andamphetamine-induced turningwith nigrostriatal dopamine content in unilateral6-hydroxydopamine lesioned rats. Brain Res., 626, 167–174.
Hur, E.E. & Zaborszky, L. (2005) Vglut2 afferents to the medial prefrontal andprimary somatosensory cortices: a combined retrograde tracing in situhybridization. J. Comp. Neurol., 483, 351–373.
Kassubek, J., Juengling, F.D., Ecker, D. & Landwehrmeyer, G.B. (2005)Thalamic atrophy in Huntington’s disease co-varies with cognitive perform-ance: a morphometric MRI analysis. Cereb. Cortex, 15, 846–853.
Kincaid, A.E., Penney, J.B. Jr, Young, A.B. & Newman, S.W. (1991) Theglobus pallidus receives a projection from the parafascicular nucleus in therat. Brain Res., 553, 18–26.
Krauss, J.K., Pohle, T., Weigel, R. & Burgunder, J.M. (2002) Deep brainstimulation of the centre median-parafascicular complex in patients withmovement disorders. J. Neurol. Neurosurg. Psychiatr., 72, 546–548.
Lanciego, J.L., Gonzalo, N., Castle, M., Sanchez-Escobar, C., Aymerich,M.S. &Obeso, J.A. (2004) Thalamic innervation of striatal and subthalamic neuronsprojecting to the rat entopeduncular nucleus.Eur. J. Neurosci., 19, 1267–1277.
Marini, G., Pianca, L. & Tredici, G. (1999) Descending projections arisingfrom the parafascicular nucleus in rats: trajectory of fibers, projection patternand mapping of terminations. Somatosen. Mot. Res., 16, 207–222.
McFarland, N.R. & Haber, S.N. (2001) Organization of thalamostriatalterminals from the ventral motor nuclei in the macaque. J. Comp. Neurol.,429, 321–336.
Meshul, C.K., Cogen, J.P., Cheng, H.W., Moore, C., Krentz, L. & McNeil, T.H.(2000) Alteration in rat striatal glutamate synapses following a lesion of thecortico- and ⁄ or nigrostriatal pathway. Exp. Neurol., 165, 191–206.
Muller, P.Y., Janovjak, H., Miserez, A.R. & Dobbie, Z. (2002) Processing ofgene expression data generated by quantitative real time RT-PCR.Biotechniques, 32, 1371–1379.
Orieux, G., Francois, C., Feger, J., Yelnik, J., Vila, M., Ruberg, M., Agid, Y. &Hirsch, E.C. (2000) Metabolic activity of excitatory parafascicular andpedunculopontine inputs to the subthalamic nucleus in a rat model ofParkinson’s disease. Neuroscience, 97, 79–88.
Paxinos, G. & Watson, C. (1998) Atlas of the Rat Brain in StereotaxicCoordinates, 4th edn. Academic Press, New York.
Rodriguez, M. & Gonzalez-Hernandez, T. (1999) Electrophysiological andmorphological evidence for a GABAergic nigrostriatal pathway.J. Neurosci., 19, 4682–4694.
Rosene, D.L., Roy, N.J. & Davis, B.J. (1986) A cryoprotection method thatfacilitates cutting frozen sections of whole monkey brain for histological andhistochemical processing without freezing artifact. J. Histochem. Cytochem.,34, 1301–1316.
Smith, Y., Raju, D.V., Pare, J.F. & Sidibe, M. (2004) The thalamostriatalsystem: a highly specific network of the basal ganglia circuitry. TrendsNeurosci., 27, 520–527.
Stornetta, R., Sevigny, C.P., Schreihofer, A.M., Rosin, D.L. & Guyenet, P.(2002) Vesicular glutamate transporter DNPI ⁄ VGLUT2 is expressed by bothC1 adrenergic and nonaminergic presympathetic vasomotor neurons of therat medulla. J. Comp. Neurol., 444, 207–220.
Vila, M., Levy, R., Herrero, M.T., Faucheux, B., Obeso, J.A., Agid, Y. &Hirsch, E.C. (1996) Metabolic activity of the basal ganglia in parkinsoniansyndromes in human and non-human primates: a cytochrome oxidasehistochemistry study. Neuroscience, 71, 903–912.
Vila, M., Levy, R., Herrero, M.T., Ruberg, M., Faucheux, B., Obeso, J.A.,Agid, Y. & Hirsch, E.C. (1997) Consequences of nigrostriatal denervationon the functioning of the basal ganglia in human and nonhuman primates: anin situ hybridization study of cytochrome oxidase subunit I mRNA.J. Neurosci., 17, 765–773.
Vila, M., Perier, C., Feger, J., Yelnik, J., Faucheux, B., Ruberg, M., Raisman-Vozari, R., Agid, Y. & Hirsch, E.C. (2000) Evolution of changes in neuronalactivity in the subthalamic nucleus of rats with unilateral lesion of thesubstantia nigra assessed by metabolic and electrophysiological measure-ments. Eur. J. Neurosci., 12, 337–344.
Weigel, R. & Krauss, J.K. (2004) Center median-parafascicular complex andpain control. Review from a neurosurgical perspective. Stereotact. Funct.Neurosurg., 82, 115–126.
Wong-Riley, M.T. (1989) Cytochrome oxidase: an endogenous metabolicmarker for neuronal activity. Trends Neurosci., 12, 94–101.
Yasukawa, T., Kita, T., Xue, Y. & Kita, H. (2004) Rat intralaminar nucleiprojections to the globus pallidus: a biotinylated dextran amine anterogradetracing study. J. Comp. Neurol., 471, 153–167.
2108 M. S. Aymerich et al.
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 23, 2099–2108
Copyright © 2022 FDOKUMEN




















