
Wir wünschen allen Bürgerinnen und Bürgern ein frohes und ...

Philosophische Fakultät der
Rheinischen Friedrich-Wilhelms-Universität Bonn
Masterarbeit zur Erlangung des akademischen Grades „Master of Arts (M.A.)“ / „Master of Science (M.Sc.)“
Thema: Späte Neandertaler und frühe anatomisch moderne Menschen in Europa. Zur Frage der Begegnung und des kulturellen Vergleichs.
vorgelegt von Jennifer Ligbado
Matrikelnummer: Studiengang: Master
Kernfach: Frühgeschichtliche Archäologie Europas und Archäologie der Römischen Provinzen
Ergänzungsfach: Kunstgeschichte des Mittelalters und Landschaftsgeschichte
6. Fachsemester
Erstgutachter/in: Privat Dozent Dr. Ralf W. Schmitz Zweitgutachter/in: Dr. Liana Giemsch

i
Inhaltsverzeichnis
1! Einleitung*..........................................................................................................................*1!2! Historischer*Hintergrund*............................................................................................*3!2.1! Das*Neandertal*....................................................................................................................*3!2.2! Eine*unverhoffte*Entdeckung*und*dessen*Auswirkung*auf*die*Nachwelt*.......*4!2.2.1!Befürworter!und!Gegner!einer!umstrittenen!Behauptung!............................................!6!
2.3! Homo*sapiens,*eine*Menschenform*mit*einer*vorsintflutlichen*Vergangenheit?*...............................................................................................................................*9!
3! Bearbeiteter*Zeitraum*und*geographische*Ausdehnung*...............................*10!4! Klima*................................................................................................................................*11!4.1! Einblicke*zum*jüngsten*Eiszeitalter*und*der*dazu*gehörigen*Bestimmungsmethoden*............................................................................................................*11!4.2! Klimatische*Bedingungen*während*der*letzten*Kaltzeit*und*des*Übergangs*vom*MittelPzum*Jungpaläolithikum*......................................................................................*13!4.3! Heinrich*Ereignisse*(HeinrichPEvents)*......................................................................*14!
5! Kampanischer*Ignimbrit,*(KI)/Y5*Eruption*.......................................................*15!6! Der*Neandertaler*.........................................................................................................*17!6.1! Verbreitungsraum*............................................................................................................*18!6.2! Welche*archäologischen*Kulturen*sind*von*Bedeutung?*....................................*19!6.2.1!Das!Moustérien!in!Europa!..........................................................................................................!20!6.2.2!Das!Châtelperronien!im!südwestlichen!Europa!(Frankreich!und!Spanien)!.........!23!6.2.3!Das!LincombienJRanisienJJerzmanowicien!im!Nordosten!Europas!(Großbritannien,!Belgien,!Deutschland!und!Polen)!....................................................................!25!6.2.4!Das!Szeletien!im!östlichen!Mitteleuropa!..............................................................................!29!6.2.5!Das!Bohunicien!im!östlichen!Mitteleuropa!.........................................................................!32!
6.3! Subsistenzverhalten*........................................................................................................*34!6.3.1!Lebensweise!......................................................................................................................................!35!6.3.2!Jagdwaffenspektrum!.....................................................................................................................!37!6.3.3!Nahrungsspektrum!........................................................................................................................!38!
7! Der*anatomisch*moderne*Mensch*.........................................................................*41!7.1! Verbreitungsraum*............................................................................................................*42!7.2! Welche*archäologischen*Kulturen*sind*von*Bedeutung?*....................................*43!7.2.1!Das!Uluzzien!im!südöstlichen!Europa!(Italien!und!Griechenland)!...........................!43!7.2.2!Das!ProtoJAurignacien/Aurignacien!in!WestJ.!MittelJ!OstJ!und!Südeuropa!.........!46!
7.3! Subsistenzverhalten*........................................................................................................*51!7.3.1!Lebensweise!......................................................................................................................................!52!7.3.2!Jagdwaffenspektrum!.....................................................................................................................!53!7.3.3!Nahrungsspektrum!........................................................................................................................!54!
8! Beispielplätze*...............................................................................................................*55!8.1! Abric*Romaní*de*Capellades,*Level*J*(ca.*50*ka)*.....................................................*56!8.2! Cueva*del*Esquilleu*in*Kantabrien*(spätes*Moustérien)*.....................................*60!8.3! Labeko*Koba*(Châtelperronien*und*ProtoPAurignacien)*....................................*62!8.4! SaintPCésaire*(Moustérien*denticulé,*Moustérien/Châtelperronien/Aurignacien)*......................................................................*64!8.5! Grotte*von*Spy*(Moustérien/LincombianPRanisianPJerzmanowice/Aurignacien)*...................................................................................................*67!8.6! Ranis*(LincombianPRanisianPJerzmanowice)*.........................................................*68!

ii
8.7! Vedrovice*V*(Szeletien)*...................................................................................................*68!8.8! BrnoPBohunice*(Bohunicien)*........................................................................................*69!8.9! Stránská*skála*(Bohunicien)*.........................................................................................*70!8.10! Grotta*del*Cavallo*(Moustérien/Uluzzien)*............................................................*70!8.11! Grotta*di*Fumane*(Moustérien/Uluzzien)*.............................................................*72!8.12! Klissoura*Grotte*(Uluzzien/Aurignacien)*..............................................................*73!8.13! RocPdePCombe*(Châtelperronien,*ProtoPAurignacien/Aurignacien)*...........*74!
9! Problematik*des*vorgestellten*Zeitraums*und*der*dazugehörigen*archäologischen*Kulturen*...............................................................................................*76!9.1! Neandertaler*oder*anatomisch*moderner*Mensch?*.............................................*77!9.2! Akkulturation,*Wunschdenken*oder*Realität?*Und*wie*wahrscheinlich*ist*eine*Koexistenz*verschiedener*Kulturgruppen,*mit*sporadischem*oder*regelmäßigem*Austausch*kultureller*Aspekte*sowie*„Tauschgut“?*..........................*79!9.3! Auf*welchem*Weg*kam*der*anatomisch*moderne*Mensch*nach*Westeuropa?* 80!9.4! Was*führte*zum*Aussterben*der*Neandertaler?*.....................................................*81!
10! Zusammenfassung*und*Fazit*.................................................................................*84!

iii
Abkürzungsverzeichnis
ABA Acid-Base-Acid = Chemische Vorbehandlung
von fast allen organischen Proben für die 14C
Methode
ABOx-SC acid-base-oxidation-stepped combustion =
Methode, um von alten Kohleproben
Verschmutzungen zu entfernen
AMS Accelerator mass spectrometry =
Beschleunigermassenspekrometrie
BC Before Christ
c. calculated = berechnet
uncal. BP oder BP uncalibrated Before Present = nicht kalibriert
vor heute (1950)
cal. BP Calibrated Before Present = kalibriert vor
heute (1950)
B2K oder b2k Before 2000 = vor dem Jahr 2000
ESR Electron spin resonance dating =
Elektronenspinresonanz
HE Heinrich Ereignis
Inf./inf. Inférieur = niedriger (als)
IRD Ice-rafted debis = Heinrich-Lagen
GRIP
NGRI
grönländisches Eisbohrprojekt
Nord grönländisches Eisbohrprojekt
GI und GS Grönland Interstadial und Grönland Stadial
ka kiloannum = ein Kilo Jahr
KI Kampanischer Ignimbrit
LRJ Lincombien-Ranisien-Jerzmanowicien
MIS 3/OIS 3 Marine Isotopic Stage/Oxygen Isotopic
Stage= Sauerstoff-Isotopenstufe/Sauerstoff-
Isotopenstadium
Ma Megannum = eine Million Jahre
MTA Moustérien „de tradition acheuléenne“
Sup./sup. Supérieur = höher (als)
TL Datierung Thermolumineszenz Datierung
TMJ Transition vom Mittel- zum Jungpaläolithikum
U/Th Datierung Uranium-Thorium Datierung
ü.d.M. über dem Meeresspiegel

1
1 Einleitung
Die Transition vom Mittel- zum Jungpaläolithikum (TMJ) ist ein Zeitraum, der
seit einem Jahrhundert von vielen Disziplinen –die Paläoanthropologie allen
voran– erforscht und kontrovers diskutiert wird. Trotz Fortschritten bleiben auch
heute noch viele Fragen offen. Wir befinden uns in einem Zeitraum großer
Veränderungen, die sowohl archäologischer als auch biologischer Natur sind.
Vor ∼60.000 bis 50.000 Jahren (Bosch et al. 2015 zufolge zwischen 50-40 ka)
beginnt der anatomisch moderne Mensch sich von Afrika aus über die ganze
Welt auszubreiten. Auf seinen Wanderwegen trifft er wahrscheinlich vor
∼45.000 bis 43.000 Jahren in Europa auf die dort lebenden Neandertaler,
welche sich vor knapp 300.000 Jahren auf dem europäischen Kontinent
entwickelten und zu diesem Zeitpunkt kurz vor dem Aussterben stehen (Nigst et
al. 2014). Aufgrund seiner Ausbreitung wurde des Öfteren vermutet, dass der
anatomisch moderne Mensch eine Schlüsselfigur im Bezug auf das Aussterben
des Homo neanderthalensis spielte (s.: Svoboda 2005; Benazzi et al. 2015),
wobei das Klima oftmals auch als möglicher Faktor in Betracht gezogen wurde
(s.: Stringer et al. 2003; van Andel et al. 2003; Jiménez-Espejo et al. 2007)
sowie deren Ernährung (Hockett/Haws 2005). Die eigentliche TMJ, die in
Westeuropa zwischen ∼40.000 und 30.000 14C BP eingegliedert wird
(Roebroeks 2008), bereitet in mancher Hinsicht Probleme. Zum einen befinden
wir uns in einem Zeitraum, in dem es zunehmend schwieriger ist korrekte
Datierungen zu erhalten, da die 14C Methode an ihre Grenzen gelangt. Und
zum anderen entstehen Europaweit fast parallel verlaufende Kulturen, wie das
Châtelperronien, Uluzzien, Bohunicien usw., die einerseits mittelpaläolithische
Technologien mit sich führen, andererseits aber jungpaläolithische Neuerungen
in ihren Inventaren mit sich bringen. Dies führte zu heftigen Debatten, in denen
darüber diskutiert wurde, wer die Macher jener Kulturen waren (Zilhão/ D’Errico,
2003; Mellars, 2004, 2005, 2006; Trinkaus, 2006; Zilhão et al., 2007;
Teyssandier, 2008, Bar-Yosef/Bordes 2010). Während die einen den
Neandertaler als Macher einiger dieser Technokomplexe sahen, trauten andere
dem Neandertaler eine solche Modernität nicht zu und vermuteten stattdessen,

2
dass nur der anatomisch moderne Mensch als Macher in Frage kommen kann.
Noch weitere sahen in diesen Kulturen den Beweis einer gelungen
Akkulturation beider Gattungen. Fakt ist, dass aufgrund eindeutiger
Knochenfunde einige dieser Kulturen dem Neandertaler zugeschrieben werden,
andere wiederum angesichts neuer Untersuchungen mittlerweile dem
anatomisch modernen Menschen.
In dieser Arbeit wird ein begrenzter Zeitraum von ca. 15.000 Jahren
bearbeitet, beginnend um ungefähr 50.000 v. Chr. im späten Mittelpaläolithikum
als das späte Moustérien in Europa verbreitet war und endend um ungefähr
35.000 v. Chr. im frühen Jungpaläolithikum, als der anatomisch moderne
Mensch als alleiniger Vertreter der Gattung Homo übrig blieb und dank seiner
Innovationen auf technologischer wie auch kultureller Ebene ein neues Zeitalter
einleitete. Um sich eine genauere Übersicht zu verschaffen, werden beide Arten
getrennt voneinander in zwei unterschiedlichen Kapiteln bearbeitet. Die
wichtigsten Kulturen mit ihren Technologien und Leitformen werden
zusammengefasst vorgestellt, als auch deren Verbreitungsraum und
Subsistenzverhalten. Anhand von ausgewählten Beispielen stechen eventuelle
Gemeinsamkeiten sowie die großen Unterschiede hervor und können somit
besser miteinander verglichen werden. In einer Diskussion werden
problematische Fragen über Zeit und Raum, wer die Macher jener Kulturen
waren, die möglichen Routen des anatomisch modernen Menschen nach
Europa, Akkulturation und Koexistenz nochmals aufgegriffen und erklärt. Das
Klima und geologische Aktivitäten der Altsteinzeit spielen ebenfalls eine
wichtige Rolle. Sie gelten als gute chronostratigraphische Marker und werden
parallel zur Archäologie und anderen Disziplinen eingesetzt, um Ereignisse
chronologisch einzurahmen. In einem letzten Kapitel wird eine
Zusammenfassung und ein Fazit mit den erhaltenen Kenntnissen vorgestellt.

3
2 Historischer Hintergrund
2.1 Das Neandertal
Im Neandertal –welches seinen Namen dem reformierten Prediger und
Dichter Joachim Neander1 (1650-1680) zu verdanken hat (Bosinski 2013)–
nahm Mitte des 19. Jahrhunderts die Geschichte des Neandertalers seinen
Lauf. Das Tal, welches unter den Einheimischen auch als „Gesteins“ oder
„Hundsklipp“ bekannt war (Bolus/Schmitz 2006), liegt dreizehn Kilometer östlich
von Düsseldorf2. Über die Jahrtausende schnitt der Fluss namens Düssel Täler
in die Region. Noch bis Anfang des 19. Jahrhunderts fanden dort Besucher aus
aller Welt ein enges idyllisches, von Bäumen bewachsenes Tal vor, das tief in
den Kalkfels geschnitten war. Die Felsklippen ragten bis zu sechzig Meter
gegen den Himmel und waren mit einem umfangreichen Höhlensystem
versehen, das aus mindestens neun Höhlen und Felsdächern bestand
(Bolus/Schmitz 2006). Zur gleichen Zeit wurden diese Höhlen –insbesondere
die größte des Tals (Neanderhöhle auf dem nördlichen Ufer) sowie die größte
der beiden Feldhofer Grotten (Feldhofer Kirche auf dem südlichen Ufer)– oft
von den Künstlern der Düsseldorfer Malerschule besucht. Diese feierten dort
ausgiebige Feste, wobei viele Abbildungen des Tals gefertigt wurden. Jenen
Zeichnungen ist es zu verdanken, dass wir heute noch eine Vorstellung des
Neandertals vor Mitte des 19. Jahrhunderts haben (Bolus/Schmitz 2006).
Nachdem es in England zur industriellen Revolution kam und diese sich
ihren Weg nach Deutschland bahnte, veränderte sich die Situation im
Neandertal drastisch. Der Kalkstein wurde in der Industrie als Zuschlagstoff für
Hochöfen gebraucht. So kam es, dass ab ca. 1850 das Tal als Steinbruch für
Kalk verwendet wurde. Schicht für Schicht wurde der Kalkstein abgebaut bis
dieser einst idyllische Ort nicht mehr wiederzuerkennen war (Bolus/Schmitz
2006). Bereits zu diesem Zeitpunkt wurden Stimmen laut, die sich kritisch über
das Vorhaben im „Gesteins“ äußerten. Trotzdem erfolgte weiterhin der Abbau
des Kalkfels. Dem Tal wurde erst zu einem späteren Zeitpunkt die Ehre zuteil,
die es verdient hatte. 1921 wurde es, mit Hilfe des im Jahr davor gegründeten 1 Eigentlich hieß der in Bremen geborene Joachim Neander mit dem Nachnamen Neumann. Er ließ seinen Namen, wie es zu dieser Zeit Mode war, ins griechische umwandeln, was dann Neander ergab (Ackermann 1980). 2 Abb.1: Geographische Lage des Neandertals. (J. Kraft, Rheinisches Landesmuseum Bonn)

4
Naturschutzvereins Neandertal, durch die preußische Regierung unter
Naturschutz gestellt (http://www.neanderthal.de/museum-tal/im-
tal/wildgehege/naturschutzverein/print.html). Heute erinnert, nicht weit von der
Fundstätte des Typus Exemplars (Neandertaler) entfernt, ein im Oktober 1996
errichtetes modernes Museum an das einstige Tal und seinen bedeutsamen
Fund.
2.2 Eine unverhoffte Entdeckung und dessen Auswirkung auf die Nachwelt
Der Abbau des Felsens wurde systematisch vorgenommen. Zuerst wurden
die Dächer der Höhlen entfernt und danach die Verfüllung jener
hinausbefördert, um nur den reinen Kalkfels zurückzubehalten. Während dieser
Abbauarbeiten waren im Jahr 1856 zwei Steinbrucharbeiter damit beschäftigt
das Erdreich, welches sich über mehrere Jahrtausende darin abgelagert hatte,
aus einer der Höhlen auszuräumen (Bolus/Schmitz 2006). In einer Tiefe von
etwa 60 cm wurden fossile Knochen, in Ablagerungen von mehr als 1,50 m
Dicke, der sogenannten Feldhofer Grotte gefunden. Zunächst wurden sie von
den Arbeitern unbeachtet in das knapp zwanzig Meter tiefer gelegene Tal
geschaufelt, in dem sie einer der Steinbruchbesitzer, Wilhelm Beckershoff,
entdeckte (Kuckenburg 2005). Auf dringliches Bitten Beckershoffs stiegen die
beiden Steinbrucharbeiter ins Tal und bargen einen Teil der Knochen. Nur
einem Zufall ist es zu verdanken, dass diese geborgen wurden, denn Wilhelm
Beckershoff nahm an, dass es sich um Höhlenbärenknochen handele. Friedrich
Wilhelm Pieper, Mitbesitzer des Steinbruchs, übergab sie einige Wochen später
an einen Eberfelder Lehrer Namens Johann Carl Fuhlrott34, der an solchen
Funden Interesse zeigte (Kuckenburg 2005; Bolus/Schmitz 2006).
Leider wurden 1856 die Knochen so unsachgemäß geborgen, dass damals
nur insgesamt 16 Knochen eingesammelt wurden. Darunter befanden sich
neben der Schädelkalotte, fünf Rippen, ein Schlüsselbein, ein
Schulterblattfragment, eine fast vollständige Beckenhälfte als auch mehrere
Arm- (Humerus, Ulna und Radius) und Beinknochen (Femur) (Kuckenburg 3 Fuhlrott hatte einen Naturwissenschaftlichen Verein für Elberfeld und Barmen gegründet, in dem auch beide Steinbruchbesitzer, Wilhelm Beckershoff und Friedrich Wilhelm Pieper, waren (Bosinski 2013). Es lag also nahe, dass die fossilen Überreste an ihn überreicht wurden. 4 Abb.2: Johann Carl Fuhlrott

5
2005; Bolus/Schmitz 2006; s.a.: Fuhlrott 1859: 137)56. Da die fossilen Knochen
mit Spitzhacken aus der Höhle befördert wurden, wiesen die meisten Brüche
und Absplitterungen auf (Bolus/Schmitz 2006).
Es ist bekannt, dass Fuhlrott damals im Neandertal war, um sich den Fund
anzusehen. Da die Grotte bereits ausgeräumt war, hätten im Schutt weitere
Knochenteile gefunden werden können. Fuhlrott unterließ es jedoch diesen zu
untersuchen, weswegen es bei der geringen Anzahl an Knochen blieb. Als er
die Gebeine sah erkannte er auf den ersten Blick (Fuhlrott 1859: 137), dass es
sich nicht wie von Beckershoff angenommen um Höhlenbärenknochen handelt,
sondern um die eines Menschen mit auffallend abnormen Merkmalen (Bosinski
2013). Vor allem am Schädel fiel ihm die Geschwulstähnliche Ausformung der
Augenbrauenbogen auf, die in der Mitte miteinander verschmolzen. Die
Schädeldecke hat eine länglich-ovale Form und ist ungewöhnlich groß. Die
Stirn hingegen ist schmal und flach, fast fliehend (Kuckenburg 2005; Fuhlrott
1859). Fuhlrott befragte auch die Steinbrucharbeiter, welche ihm erklärten, dass
sie die Knochen in einer Tiefe von etwa 60 cm unter der Oberfläche gefunden
hätten. Ebenfalls sagten sie aus, das Skelett hätte im korrekten anatomischen
Verbund mit dem Kopf in Richtung des Höhleneingangs gelegen (Bosinski
2103; Kuckenburg 2005). In einer ersten Äußerung –wegen der Aussagen der
Arbeiter– ging Fuhlrott davon aus, dass in der Höhle ein komplettes Skelett
gelegen haben könnte (Kuckenburg 2005). Aufgrund damaliger Diskussionen
widerrief er jedoch jene Ausführung wieder (Bosinski 2013). Er wies darauf hin,
dass die Höhlenöffnungen viel zu weit oberhalb des Tales am Berghang lägen
und so zu beschwerlich zu erreichen gewesen wären. Nach seinem Widerruf
bekräftigte er allerdings, dass jenes Skelett zufällig in die Höhle hineingelangt
wäre (Kuckenburg 2005). Nach weiteren Untersuchungen bei denen auch ein
Arzt (Dr. Meisenberg) aus Elberfeld hinzugezogen wurde, stand für Fuhlrott
dann schlussendlich fest, dass es sich bei den Überresten um eine
altertümliche Menschenform handeln muss (Bolus/Schmitz 2006; Fuhlrott
1859).
5 Abb.3: Neandertaler Knochen aus dem Neandertal. Abgebildet sind die Knochen von 1856 sowie weitere Fragmente aus den Grabungskampagnen von 1997 und dem Jahr 2000. Foto Universität Bonn/ G. Oleschinski 6 Abb.4: Zeichnung von Gerhard Wandel mit den 1856 gefunden Knochen des Neandertalers.

6
2.2.1 Befürworter und Gegner einer umstrittenen Behauptung
Nachdem in mehreren Zeitungen über den Fund aus dem Neandertal
berichtet wurde, erhielt Fuhlrott ein Gesuch von zwei Professoren der
Universität Bonn, in dem er gebeten wurde, die Gebeine an die Universität zu
schicken, damit diese genauer untersucht werden können. Hierbei handelte es
sich zum einen um Geh. Rath Dr. Mayer und zum anderen um den 40-jährigen
Dr. Hermann Schaaffhausen 7 , der schon damals für sein fortschrittliches
Denken bekannt war (Bolus/Schmitz 2006; Fuhlrott 1859). Da Herr Geh. Rath
Dr. Mayer zu jenem Zeitpunkt als Fuhlrott nach Bonn kam bettlägerig war,
untersuchte Schaaffhausen das Skelett im Winter 1856/57 und ließ Fuhlrott
daraufhin einen genauen Bericht8 –welcher 1858 veröffentlicht wurde– über
seine Erkenntnisse zukommen (Fuhlrott 1859). Dank der Unterstützung
Schaaffhausens ging Fuhlrott im Jahr 1857 zur Generalversammlung des
Naturhistorischen Vereins der Preußischen Rheinlande und Westphalens, um
vom Neandertalerfund zu berichten (Bosinski 2013). Doch wie anzunehmen
war, schenkten ihm die meisten Gelehrten keinen Glauben, vor allem weil die
Gebeine nicht in Verbindung mit Knochen von Tieren sowie Steinwerkzeugen
aus vordiluvialer Zeit (Eiszeit) gefunden worden waren. Auch seine
Argumentation, basierend auf den Flecken aus einer Mangan- und
Eisenverbindung, die auf den Gebeinen durch Geh. Rath Dr. Mayer entdeckt
wurden, stieß auf taube Ohren. Gleichwohl bei anderen Fundkontexten mit
tierischen Gebeinen aus der Diluvialzeit gleiche Beobachtungen gemacht
worden waren (Fuhlrott 1859).
Wenngleich viele Gelehrte jener Zeit dem Evolutionsgedanken nicht folgten
und Theorien wie die Fuhlrotts beiseite schoben, kam es zu grundlegenden
Veränderungen. Im gleichen Jahr in dem Fuhlrott seinen Bericht 9
veröffentlichte, erschien die Evolutionstheorie von Charles Darwin, die neue
und bedeutende Wege in der Forschung ebenen sollte. In den darauf folgenden
Jahren wurden immer wieder neue Berichte in Bezug auf den Neandertaler
veröffentlicht, die seine Altertümlichkeit in Frage stellten. Während einige davon
ausgingen, dass es sich bei den Überresten um die eines Schwachsinnigen 7 Abb.5: Hermann Schaaffhausen 8 „Zur Kenntnis der ältesten Rassenschädel“ 9 „Menschliche Überreste aus einer Felsgrotte des Düsselthals. Ein Beitrag zur Frage über die Existenz fossiler Menschen“ (1859).

7
handelte, gingen andere davon aus, dass es sich um einen Kelten, einen alten
Holländer oder sogar um einen Kosaken aus den napoleonischen Kriegen
handeln müsse (Bolus/Schmitz 2006).
Die Forschung am Neandertaler wurde, vor allem durch die Aussage eines
der größten Gelehrten aus dem 19. Jahrhundert, in Deutschland und seinen
Nachbarländern für Jahrzehnte in falsche Bahnen gelenkt. Rudolf Virchow, ein
deutscher Arzt, erhielt 1872 während der Abwesenheit Fuhlrotts durch dessen
Frau Zugang zum Originalskelett. Bei seiner Untersuchung stellte er an den
Gebeinen pathologische Veränderungen fest –die auf Verletzungen
zurückzuführen waren– die schon Schaaffhausen erkannt hatte und auch heute
von Wissenschaftlern belegt werden konnten. Ihm unterlief allerdings der Fehler
auch natürliche Abweichungen im Knochenbau als krankhaft anzusehen. Nach
seiner Auffassung hatte dieses Individuum in seiner Kindheit wahrscheinlich an
Rachitis gelitten, sich davon aber wieder erholt und ging zuweilen als
Erwachsener einer kräftigen Tätigkeit nach. Weiter vermutete er, diese Person
habe sich im Erwachsenenalter schwere Kopfverletzungen zugezogen, die aber
glimpflich verliefen, sodass sie sich davon ebenfalls erholte. Zu einem späteren
Zeitpunkt in seinem Leben litt jener zudem an Arthritis deformans und anderen
Krankheiten, die im hohen Alter auftreten, wodurch der linke Arm fast ganz
unbeweglich war (Virchow 1872; Kuckenburg 2005). Seine Schlussfolgerung
nach der Untersuchung am Skelett war: So wenig ich mich berechtigt fühlen würde, heutigen Tages die Natur einer Race nach
einem Schädel zu beurtheilen, welcher grosse und wesentliche Spuren krankhafter
Störungen an sich trägt, (…), so meine ich auch, dass man es wird aufgeben müssen,
den Neanderthal-Schädel als hinreichendes Zeugniss einer Race anzusehen, welche
den gleichen Typus der Schädelbildung gehabt haben. Denn dann müsste man
glauben, dass es ganz und gar eine pathologische Race gewesen sei (Virchow 1872:
163). Weiter schreibt er:
Ich meine aber, der Neanderthal-Schädel wird vorläufig nur als eine merkwürdige
Einzelerscheinung gelten dürfen, und ehe wir nicht durch parallele Funde weitere
Aufklärung erlangt haben, müssen wir daran festhalten, dass eine durchaus individuelle
Bildung vorliegt (Virchow 1872: 163-164).
Bis zu seinem Lebensende wich er von seiner Sicht der Dinge auf das
Neandertalerskelett nicht mehr ab. Als einer der erfolgreichsten Gelehrten
seiner Zeit prägte Virchow andere Anatomen mit seinen Aussagen. Das führte

8
zu vermehrten Anfeindungen, denen sich Fuhlrott und Schaaffhausen
ausgesetzt sahen. 1886 kam es in Spy (Belgien) zu zwei weiteren
Neandertalerfunden mit identischen Merkmalen. Dies hielt Virchow aber nicht
davon ab weiterhin auf seiner Meinung zu beharren (Bolus/Schmitz 2006).
In den 1860e Jahren, noch vor den Geschehnissen mit Rudolf Virchow, gab
es auf den britischen Inseln Kunde über den Neandertalerfund. 1860 reiste
Charles Lyell, ein ebenso fortschrittlich denkender Wissenschaftler seiner Zeit,
nach Deutschland und ließ sich die Fundstelle im Neandertal sowie die
Schädelkalotte persönlich von Fuhlrott zeigen. Ihm wurde ein Abguss der
Schädeldecke überreicht, den er nach seiner Rückkehr nach England an den
Zoologen Thomas Henry Huxley übergab. Dieser war ein großer Verfechter der
Evolutionstheorie und beschäftigte sich nach Erhalt dieses Abgusses fortan
auch mit dem Neandertaler. Anders als in Deutschland scheint es, dass der
Neandertaler in Großbritannien eine wahre Chance bekommen hat. Denn
wenngleich sich Lyell Anfangs skeptisch zeigte, vor allem aufgrund fehlender
Schichtzusammenhänge, ging er äußerst objektiv an den Fund aus dem
Neandertal in seinem Werk „The Geological Evidence of Antiquity of Man“ von
1863 heran (Bolus/Schmitz 2006). Ebenso widmete Huxley im gleichen Jahr in
seinem Werk „Evidence as to Man’s Place in Nature“ seine Aufmerksamkeit
dem Neandertalerschädel und versuchte ihn nach bestmöglicher Objektivität zu
beschreiben und einzugliedern.
Im darauf folgenden Jahr 1864 wurde ein bereits 16 Jahre alter Fund von
einem anderen Zoologen Namens George Busk der Öffentlichkeit vorgestellt.
Er stellte fest, dass der Schädel morphologische Gemeinsamkeiten zum Typus-
Exemplar aufwies, jedoch graziler als jener war und somit zu einem Skelett
einer Frau gehört, die der gleichen Gattung wie der Neandertaler angehört
(Kuckenburg 2005)10. Hugh Falconer ein schottischer Paläontologe stellte fest,
dass der Schädel abweichende Proportionen zum anatomisch modernen
Menschen aufwies und ein beträchtliches Alter haben musste. Er sah in den 10 George Busk ging sogar einen Schritt weiter, nachdem er das Potential der Funde aus dem Neandertal sowie von Gibraltar erkannte, schien er mit einigen seiner Aussagen seine Kollegen auf dem Festland verspotten zu wollen. Vor allem war seine Kritik an Franz Joseph Carl Mayer gerichtet, der davon ausging, dass es sich beim Neandertaler (Typus-Exemplar) um einen Kosaken aus dem napoleonischen Krieg handeln musste, der sich in der Höhle verkrochen hatte (s.d.: Kuckenburg 2005: 45 ff.). Zum Fund aus Gibraltar schrieb er: „Selbst Professor Mayer wird kaum behaupten, dass sich ein rachitischer Kosak des Feldzuges von 1814 in die versiegelten Spalten des Felsens von Gibraltar verkrochen habe“ (zit. bei Kuckenburg 2005: 53).

9
Überresten eine andere Spezies und schlug den Namen „Homo var. calpius“ –
benannt nach Mons Calpe in Gibraltar– vor (Barton et al.: 1999). Er erkannte
jedoch nicht, dass es sich bei dem Fund von Gibraltar um den Schädel eines
Neandertalers handelte. Es war schlussendlich William King, der 1864 dem
Neandertaler seinem Namen „Homo neanderthalensis“ gab (Barton et al.:
1999).
2.3 Homo sapiens, eine Menschenform mit einer vorsintflutlichen Vergangenheit?
Ab der Mitte des 19. Jahrhunderts beschäftigten sich die Gelehrten vor allem
mit einem Thema, dem Menschen und der damit verbundenen
Evolutionstheorie, sei es nun um jenes Thema zu bekräftigen oder zu
diskreditieren. Die Frage, ob der Mensch eine „vorsintflutliche Vergangenheit“
hat oder nicht, wurde über mehrere Jahrzehnte kontrovers diskutiert. Die
Generation an Wissenschaftlern aus dem 19. Jahrhundert wuchs, wie auch
schon viele vor ihnen, in dem Glauben auf, dass die Welt nur wenige Tausend
Jahre alt ist. 1658 legte der irische Erzbischof James Ussher die Entstehung
der Erde und der Menschen auf das Jahr 4004 v. Chr. Ein britischer Priester
Namens John Lightfoot wurde diesbezüglich noch genauer und legte den Tag
der Erschaffung auf den 23. Oktober 4004 v. Chr. und da der Mensch zur
dritten Stunde des Tages erschaffen wurde, sprach er sich für neun Uhr
morgens aus (Kuckenburg 2005; http://www.ccel.org/ccel/lightfoot).
Mutige Gelehrte wie John Frere, der Naturforscher Paul Tournal oder Jaques
Boucher de Perthes wagten schon zu früheren Zeitpunkten erste Aussagen –
wenn auch nicht alle immer richtig waren– zur Dauer der menschlichen
Existenz auf dieser Welt, da bereits im 18. sowie frühen 19. Jahrhundert an
vielen Orten Überreste von Menschen, Tieren und deren Artefakte aus der
sogenannten „vorsintflutlichen Periode“ gefunden wurden (Kuckenburg 2005).
Allerdings sollte es –wie schon weiter oben ausführlich beschrieben– noch bis
zur Mitte des 19. Jahrhunderts dauern bis sich der Gedanke diesbezüglich
allmählich änderte und anerkannt wurde, dass der Mensch wesentlich älter ist
als angenommen und es auch altertümliche Formen aus längst vergangener
Zeit gab. Aussagen wie die von Paul Tournal aus dem Jahr 1829 sollten sich
nach nur wenigen Jahrzehnten bewahrheiten:

10
„Die Geologie wird unsere kurzen Geschichtsannalen ergänzen und den Menschen stolz
auf das hohe Alter seiner Rasse machen, denn sie allein kann künftig uns bestimmte
Angaben über Zeit des ersten Erscheinens des Menschen auf unserer Erde geben“ (zit.
bei Kuckenburg 2005: 37).
3 Bearbeiteter Zeitraum und geographische Ausdehnung
Die vorliegende Arbeit konzentriert sich auf eine begrenzte Zeitspanne von
ungefähr 15.000 Jahren, wobei eine Periode aus der Altsteinzeit bearbeitet
wird, die den Zeitraum zwischen ca. 50.000 bis 35.000 v.Chr. umspannt. Diese
ist von großer Bedeutung, da es eine Zeit –klimatisch wie auch kulturell– der
großen Umbrüche war. Zum Einen befinden wir uns am Ende des
Mittelpaläolithikums, das über Jahrtausende durch archäologische Kulturen wie
das Moustérien und das Micoquien geprägt wurde, zum Anderen an einem
wichtigen Wendepunkt der Geschichte, an dem das Mittelpaläolithikum in das
Jungpaläolithikum übergeht. Neben den genannten lithischen Kulturen
entwickeln sich nun Übergangsindustrien mit Elementen aus Mittel- und
Jungpaläolithikum. Die Blattspitzengruppen gehören zu den prägnantesten
jener Zeit. Nachdem der Neandertaler nahezu 300.000 Jahre der einzige
Vertreter der Gattung Homo in Europa war, trat um ca. 45.000 v. Chr. ein
weiterer Vertreter dieser Gattung auf den Plan. Der anatomisch moderne
Mensch (Homo sapiens sapiens) war über die letzten Jahrtausende auf seinen
Wanderwegen nach Europa gelangt.
Da das Subsistenzverhalten beider Gattungen miteinander verglichen
werden soll, sowie Einflüsse vom Neandertaler auf den anatomisch modernen
Menschen und/oder umgedreht in Betracht gezogen werden sollen, ist es nötig
einen Raum zu definieren in dem beide gegebenenfalls für eine gewisse
Zeitspanne koexistiert haben. Von Interesse sind daher Regionen wie die
iberische Halbinsel (Spanien und Portugal) und Frankreich (Südwest Europa),
Großbritannien, Belgien, Deutschland und Österreich (Nordwest sowie
Mitteleuropa), Italien und Griechenland (Südost Europa) sowie Polen, die

11
Tschechische Republik (Mähren/Morawien) und die Donau Region (östliches
Mitteleuropa)11.
4 Klima
4.1 Einblicke zum jüngsten Eiszeitalter und der dazu gehörigen Bestimmungsmethoden
Gegenwärtig befinden wir uns im jüngsten Eiszeitalter unserer Erde, dem
Quartär, das vor ca. 2.6 Ma begonnen hat (Gibbard/Head 2010)12. In den
letzten Jahrmillionen dieses Zeitabschnitts wechselten sich mehrere Kalt- und
Warmzeitzyklen ab, zu denen unter anderem die folgenden sechs jüngeren
Abschnitte gehören: die Cromer-Warmzeit, die Elster-Kaltzeit, die Holstein-
Warmzeit, die Saale-Kaltzeit, die Eem-Warmzeit, die Weichsel-Kaltzeit sowie
das Holozän, der gegenwärtigen Warmzeit in der wir leben. Jene Namen der
aufgezählten Zyklen werden in Norddeutschland und seinen angrenzenden
Gebieten verwendet. Für das Alpenland und den süddeutschen Raum gelten
andere Begriffe, die von dem Geographen Albrecht Penck und seinem Schüler
Eduard Brückner Anfang des 20. Jahrhunderts geprägt wurden (Ehlers 2011).
Die vier letzten Kaltzeiten im Alpenraum wurden dementsprechend nach den
Nebenflüssen der Donau als auch nach einem Voralpenfluss benannt: Günz-
Eiszeit, Mindel-Eiszeit, Riß-Eiszeit und die Würm-Eiszeit (Bolus/Schmitz 2006;
Ehlers 2011). Und die sich dazwischen befindlichen Warmzeiten lediglich als
Günz/Mindel-, Mindel/Riß- und Riß/Würm-Warmzeit. Es sollte erwähnt werden,
dass die genannten Gliederungen nur regional aussagekräftig sind und es
überregional zu Unterschieden in der Abfolge der Gletschervorstöße kommen
kann. Alleine die Würm- und die Weichseleiszeit stimmen in ihrem Verlauf
überein (Bolus/Schmitz 2006). Um etwaige Verwirrungen zu vermeiden, werden
ausschließlich die Bezeichnungen für das nördliche Mitteleuropa verwendet.
Im Laufe des 20. Jahrhunderts entwickelten sich die Methoden, um
Klimaabfolgen genauer zu definieren, stetig weiter. Mitte der 1950e wurde eine
innovative Verfahrensweise –basierend auf der Analyse von Sauerstoffisotopen
11 Abb.6: Geographische Übersichtskarte 12 In älteren Quellen wurden andere Daten diskutiert wobei 2.4 Ma und 1.8 Ma die beiden meistgenannten waren. In jüngster Zeit wurde sich jedoch auf den oben genannten ungefähren Zeitpunkt von circa 2.6 (2.588) Ma geeinigt (Gibbard/Head 2010).

12
(O16 & O18) aus den Ozeansedimenten– entwickelt, die bis heute in der
Forschung für die Rekonstruktion von klimatischen Bedingungen verwendet
wird (Bolus/Schmitz 2006). Die Sauerstoff-Isotopenstufe 13 –abgekürzt MIS
(Marine Isotopic Stage) oder OIS (Oxygen Isotopic Stage)– wird in
verschiedene Stadien untergliedert, wobei für das Quartär 103 bekannt sind,
(http://quaternary.stratigraphy.org/images/TablaCuaternarioIbéricov_209.pdf).
Während gerade Zahlen für Glaziale (Kaltzeiten) stehen, werden ungerade
Zahlen für Interglaziale (Warmzeiten) angewandt.
In den 1990e wurde das grönländische Eisbohrprojekt ins Leben gerufen, das
nach einem ähnlichen Prinzip wie die Sauerstoff-Isotopenstufe funktioniert. Da
die Eisschicht in Grönland mehr als 3 km dick ist, befindet sich dort ein in der
Regel gut datierbares Klimaarchiv. Wie bei OIS wird auf die im Eis
eingeschlossenen Sauerstoffisotopen zurückgegriffen. 1992 wurde in einer
ersten Bohrung (GRIP) ein 3029 m langer Bohrkern zu Tage gebracht und im
darauffolgenden Jahr, knapp 30 km von der ersten Bohrstelle entfernt, ein 3053
m langer Kern (GRIP 2), von einem amerikanischen Team14. Aus den Analysen
konnten die Klimaschwankungen aus den letzten 105.000 Jahren ermittelt
werden. Alle Daten darüber hinaus sind unpräzise, da die letzten ∼300 m der
beiden Bohrkerne –zurückzuführen auf Bodenwellen, die zu Stauchungen im
Eis führen– Störungen in der Beschaffenheit aufwiesen
(http://www.iceandclimate.nbi.ku.dk/research/drill_analysing/history_drilling/cent
ral_ice_cores/; Bolus/Schmitz 2006). Interessant an diesen Verfahren ist, dass
sie im Gegensatz zu anderen Methoden nicht auf eine Region begrenzt sind,
allerdings sollte auch bei diesen beachtet werden, dass sie nicht in allen
Arbeitsbereichen anwendbar sind. Sie haben gegenüber anderer Verfahren
Vorteile, da sich pro Jahr eine Schicht bildet –sei es Sediment oder
Schnee/Eis–, die wie bei Jahresringen von Bäumen einfach nur zurückgezählt
werden müssen, um einen genauen Einblick in einen Zeitabschnitt zu
bekommen (Bolus/Schmitz 2006). Es sei noch erwähnt, dass jede Methode ihre
Vor- und Nachteile mit sich bringt und gestellte Fragen nicht immer eindeutig
beantwortet werden können. Indem mehrere Verfahren miteinander verglichen
13 Abb.7: Sauerstoff- Isotopenstufen der letzten 110 ka 14 Abb.8: GRIP & GRIP 2 grönländisches Eisbohrprojekt

13
oder zusammen verwendet werden, besteht die Möglichkeit einen detaillierteren
Einblick über den Klimaverlauf aus vergangener Zeit zu bekommen.
4.2 Klimatische Bedingungen während der letzten Kaltzeit und des Übergangs vom Mittel-zum Jungpaläolithikum
Ein Teil des Mittelpaläolithikums und die Transition vom Mittel- zum
Jungpaläolithikum fanden während der Weichsel-Kaltzeit statt. Diese begann
nach der Eem-Warmzeit mit einer ersten Kaltphase, dem Herning-Stadial (OIS
5d) (Litt et al. 2007), um 115.000 BP und erstreckte sich über einen Zeitraum
von ungefähr 100.000 Jahren, bis zum Anfang des Holozäns vor ca. 11.700 b2k
(Walker et al. 2009). Wie auch während der Eiszeitalter wechselten sich immer
wieder Wärme- und Kältephasen ab, die während einer Kaltzeit aber als Stadial
(Kältephase) und Interstadial (Wärmephase) bezeichnet werden. Zu Beginn der
Weichsel-Eiszeit wurde es langsam aber stetig kälter, erste Anzeichen für einen
Vorstoß des skandinavischen Inlandgletschers gibt es ab OIS 4 um 65 ka BP.
Diese erste Abkühlung hielt nur ein paar tausend Jahre an bis um 59 ka BP das
OIS 3 begann und das Klima wieder deutlich milder wurde. Jenes wärmere
Klima wurde zwischenzeitlich von kürzeren Kälteperioden (s. Kapitel 4.3)
unterbrochen, war jedoch bis 43 ka BP relativ stabil. Danach begann das Klima
anhaltend schlechter zu werden, wobei kältere Phasen dichter aufeinander
folgten. Die letzte Wärmephase endete 37 ka BP und war deutlich kürzer und
kälter als die vorherigen. Um 30 BP wurde ein Kältemaximum erreicht, das dem
von OIS 2 (21 ka BP) gleichzustellen ist (vgl.: Davies/Gollop 2003: 131). Die
Temperaturen während OIS 3 waren denen heutiger Zeit sehr ähnlich,
allgemein war es aber 4°-6°C kühler. Sommertemperaturen über 18°C wurden
nur in Spanien und dem westlichen Mittelmeerraum erreicht. Die Temperaturen
fielen im Winter nahe des Inlandgletschers weit unter -8° bis -20°C und bis -
30°C auf den Eisflächen. Im Gegensatz zu heute waren die Winter eher trocken
(Vgl.: Davies/Gollop 2003: 132 ff.). Abrupte Klimaschwankungen, von kalten zu
deutlich milderen Wetterkonditionen, zeichnen die gesamte Weichselkaltzeit
aus (Kjellström et al. 2010; Rahmstarf 2002). Untersuchungen von NGRIP
ergaben, dass es zu insgesamt 25 Klimawechsel kam (s. Literaturverzeichnis:
andere Medien {N°6}). Diese Schwankungen sind uns heute als Dansgaard-

14
Oeschger Zyklen15 –benannt nach ihren Entdeckern– bekannt (Kjellström et al.
2010). Aufgrund des letzten Stadials (OIS 2), das vor 25 ka BP begann und vor
allem wegen der Ausbreitung des fennoskandischen Eisschildes während des
letzten Kältemaximums (21 ka BP), ist es schwierig genaue Aussagen über das
OIS 3 zu fällen, da Paläodaten sehr spärlich sind (Kjellström et al. 2010). Um
sich dennoch ein genaueres Bild über das Klima während des OIS 3 in Europa
machen zu können, wurde in jüngster Zeit unter anderem ein Projekt von
Kjellström et al. (2010) gestartet.
4.3 Heinrich Ereignisse (Heinrich-Events)
Heinrich Ereignisse wurden in den 1980e von Hartmut Heinrich als dünne
Sedimentschichten (IRD) in drei Bohrkernen aus dem Nordatlantik entdeckt
(Broecker et al. 1992). 1988 wurden diese Ablagerungen und eine erste
Definition jener in einem Paper in der Quartenary Research 29 von ihm
vorgestellt. Er beschreibt sie als Phasen oder Ereignisse periodisch massiver
Eisvorstöße –während des späten Pleistozäns– vom laurentidischen Eisschild
in die Hudsonstraße einer Meeresenge in Kanada (Heinrich 1988; Marcott et al.
2011). Durch den Abrieb der Gletschermassen wurden Sedimente mit Hilfe der
Eisschollen auf das offene Meer getragen, wo sie nach Abschmelzen jener auf
den Meeresgrund sanken (Heinrich 1988). Heinrich Ereignisse sind Stadiale,
die Weltweit durch extreme Konditionen gekennzeichnet sind (Stieglitz et al.
2014).
Bis Dato wurden sechs Lagen eindeutig belegt, davon sind alle in die letzte
Kaltzeit einzuordnen. Gekennzeichnet sind sie mit den Bezeichnungen H6-H1
(oder HE6-HE1; ∼60-15 ka BP), wobei H1 (∼18-15 ka BP) das jüngste Ereignis
darstellt (Álvarez-Solas et al. 2011). Nach Ansicht einiger Autoren gibt es noch
ein siebtes HE, jenes wäre demnach mit der Bezeichnung H0 (jüngere
Dryaszeit; ∼12 ka BP) versehen (Broecker 1994). Vier der sechs Ereignisse
entspringen ihrer Zusammensetzung nach der gleichen Quelle. Hierbei handelt
es sich um H1, H2, H4 und H5, die ihren Ursprung in einer Region nahe der
Hudsonstraße haben. H3 und H6 ähneln dem Sediment aus dem Bohrkern
15 Abb.9: NGRIP: Dansgaard-Oeschger Zyklen

15
V28-82 (Vgl.: Hemming 2004, S. 17) und haben wahrscheinlich ihren Ursprung
in der europäischen Arktis (Grousset 2001).
HE treten in einem Intervall von 7.000 bis 14.000 Jahre auf (Heinrich 1988;
Broecker et al. 1992; Broecker 1994; Bond/Lotti 1995; Grousset 2001;
Hemming 2004). Neben den Intervallen und dem Zeitpunkt16 in denen sie
auftreten, wurde versucht eine Aussage über deren Dauer zu fällen. Aufgrund
der vielen Faktoren (verschiedene Bohrkerne, Abweichungen und
Ungenauigkeiten in der Messung mit 14C, Bioturbation nach den Ereignissen
usw.) stellt es sich als schwierig heraus eine genaue Dauer zu errechnen. Da
es sich jedoch um kurzweilige Ereignisse handelt, kann davon ausgegangen
werden, dass ein Event weniger als ein Jahrtausend gedauert hat (Dowdeswell
et al. 1995; Veiga-Pires/Hillaire-Marcel 1999; Roche et al. 2004; Hemming
2004). Wobei aufgrund der unterschiedlichen Messtechniken Datierungen
zwischen 250 und 2000 Jahre rauskommen (Roche et al. 2004; Hemming
2004; Veiga-Pires/ Hillaire-Marcel 1999), die für jedes HE unterschiedlich
ausfallen. Im Durchschnitt hat ein Event –wenn man sich an H1 und H2
orientiert– knapp ein halbes Jahrtausend (495 +/- 255 Jahre) gedauert
(Hemming 2004).
5 Kampanischer Ignimbrit, (KI)/Y5 Eruption
Während der Transition vom Mittel- zum Jungpaläolithikum kam es neben
umfangreichen kulturellen und klimatischen Änderungen zu einem
geologischen Großereignis. Zu jener Zeit ereignete sich in der neapolitanisch-
kampanischen Region (im Süden Italiens) der größte Vulkanausbrauch der
letzten 100.000 Jahre im mediterranen Gebiet (Pyle et al. 2006; Sparks et al.
2005). Dasjenige Ereignis überschneidet sich mit dem Beginn des HE 4’s, dem
kältesten und trockensten Zeitabschnitt während der letzten Kaltzeit (Vgl.:
Costa et al. 2012, S.1; Fitzsimmons et al. 2013, S.1). Die Verbindung beider
Begebenheiten hatte mit großer Wahrscheinlichkeit einen erheblichen Impact
auf das Ökosystem, wovon sowohl Fauna, Flora als auch der Mensch und der
Neandertaler in direkter und entfernter Umgebung, über Jahrhunderte eventuell
sogar Jahrtausende, betroffen waren (Fedele et al. 2002, 2003, 2007; Pyle et
16 Tab.1: Datierung von Heinrich Ereignissen

16
al. 2006; Fitzsimmons et al. 2013). Diese Super-Eruption, allgemein bekannt als
kampanischer Ignimbrit (KI), hatte ihren Ursprung in den Phlegräischen Feldern
im Westen von Neapel. Die daraus resultierende Asche –(2.5-5) x 1014 kg–
bedeckte ein Gebiet von ungefähr 5.000.000 km2 (Pyle et al. 2006; Giaccio et
al. 2006), das sich in Teilen von Süd- bis weit nach Osteuropa erstreckt (Italien,
Balkan Region, südöstlich mediterraner Raum, Nord Afrika und die russische
Ebene; Fitzsimmons et al. 2013) und reichte bis in eine Höhe von 40-45 km
(Fedele et al. 2003; Fitzsimmons et al. 2013). In der direkten Umgebung kam in
einem Umkreis von ca. 30.000 km2 ein pyroklastischer Strom aus Schutt und
Geröll runter (der Schutt lagerte sich in einer Distanz von bis zu 80 km vom
Ausbruchort ab; Vigliotti 2014; Hoffecker et al. 2008). Der Ausbruch selbst
hinterließ eine Caldera von geschätzt 230 km2 (Hoffecker et al. 2008). Das
kampanische Ignimbrit Tephra wurde in der Stratigraphie vieler archäologischer
Fundkomplexe identifiziert, die von Italien, Griechenland, Bulgarien bis nach
Südwest Russland in die Kostenki Region am Fluss Don reichen (Pyle et al.
2006)17.
Die Ascheschicht aus der KI Eruption wurde des Weiteren in marinen
Sedimenten und Sedimenten aus Seen (z.B.: Lago Grande di Monticchio;
Engwell et al. 2014) sowie im grönländischen Eis –im GISP2 Bohrkern
zwischen GI und GS 9 und dem darauffolgenden HE 4 zu finden (Hoffecker et
al. 2008)– ausgemacht. Die marinen Ablagerungen 18 sind uns als C-13
(tyrrhenisches Meer) und Y5 Tephra (adriatisches Meer) bekannt (Fedele et al.
2003), wobei letzter Begriff allgemein populärer ist und zudem als jenes
(Tephra) gilt, welches sich am weitesten ausgebreitet hat. Aufgrund der
Ausmaße der Eruption und seiner guten Datierbarkeit gilt das KI als
bedeutender chronologischer Marker während der Transition vom Mittel- zum
Jungpaläolithikum im südöstlich gelegenen mediterranen Raum. Mittels 40Ar/39Ar Datierung –stammend von 18 Proben– wurde der Ausbruch auf ein
Alter von 39.280 +/- 110 BP berechnet. Weitere Datierungen –mit Hilfe der
gleichen Methode– mit Proben aus dem tyrrhenischen Meer ergeben ähnlichen
17 Abb.10: Geographische Verbreitung des KI Tephras 18 Zwei Phasen der Eruption konnten ausgemacht werden: zum einen die plinianische Phase, bei der große Mengen an Material in die Atmosphäre geschleudert werden und in der direkten Umgebung wieder herunterregnen und zum anderen, in der darüber liegenden Schicht, die ignimbrit/co-ignimbrit Phase, in der sich Material über große Distanzen verbreitet aufgrund seiner geringen Masse (Engwell et al. 2014).

17
Daten um 41.000 +/- 400 BP (Fedele et al. 2003). Die Datierung des Y5
Tephras im Grönland Eis ergibt einen Zeitraum von 43.000-40.000 BPGISP2
(Hoffecker et al. 2008). Nach Ansicht von Fedele et al. (2008) liegen die am
Besten bestimmten Daten bei 40.012 ka BPGISP2 und 39.395 +/- ka BP40Ar/39Ar.
Vergleicht man die 14C Datierungen fällt auf, dass sie im Gegensatz zu den
obigen Ergebnissen, im Durchschnitt bis zu 8.000 Jahre jünger ausfallen (s.d.:
32.000-29.000 14C BP: Giaccio et al. 2006; 32.420 +/- 400/420 14C BP:
Sinitsyn/Hoffecker 2006; 32.300 +/- 600 und 34.600 +/- 150 14C BP: Fedele et
al. 2002; Deino et al. 1994; 36.33,5-26.9 ka 14C BP: Scandone et al. 1991). Sie
ergeben Zahlen zwischen c. 27.000 und c. 42.000 14C BP (Fedele et al. 2003),
die Größtenteils zu jung für die KI Eruption angesetzt sind.
6 Der Neandertaler
Die klassische Form des Neandertalers macht nur einen Bruchteil seiner
Entwicklungsgeschichte aus. Seine Geschichte lässt sich mittels seiner
Vorfahren über Jahrtausende zurückverfolgen. Als Schlüsselgebiet seiner
Entwicklung gilt die Sierra de Atapuerca19 bei Burgos in Spanien20. Zwar waren
die Neandertaler nicht die Ersten, die in Europa gelebt haben, dennoch gelten
sie bis heute als die ersten echten Europäer. Sie gingen aus der Gattung Homo
heidelbergensis hervor, die als deren direkter Vorfahr gilt. Vor rund 200.000
Jahren entwickelten sich die ersten markanten Merkmale, die den Neandertaler
als jenen erkennen ließen. Die klassische Form, die uns von Plastiken und
Zeichnungen aus Museen oder Zeitschriften bekannt ist, trat zum ersten Mal
vor ∼115.000 Jahren, mit dem Beginn der letzten Kaltzeit (Weichsel-Kaltzeit), in
Erscheinung. Aus dieser Zeit des Mittelpaläolithikums ist eine relativ hohe
Anzahl an Knochen bekannt. Jene, die dem klassischen Neandertaler genau
19 Abb.11: Sierra de Atapuerca 20 Zahlreiche Funde aus der Gran Dolina Grotte sowie ein Schädel aus Crepano in Italien konnten auf ein Alter von ca. 800.000 BP datiert werden. Sie galten lange als die ältesten Funde in Europa, bis im Jahr 2007 in der Sima del Elefante Grotte ein Unterkiefer (ATE9-1) geborgen wurde, der auf ein Alter von 1.3 Ma eingeordnet werden konnte (Carbonell et al. 2008; Bermúdez de Castro et. al: 2011). Die Sima de los Huesos gilt als Schlüsselplatz für die Entwicklung der Neandertaler. Die dort entdeckten bis zu 600.000 Jahre alten Überreste (Homo heidelbergensis) weisen erste Zeichen einer sogenannten „Neandertalisation“ auf (Bischoff et al. 2007; Serangeli/Bolus 2008: 84).

18
zugeordnet werden konnten, geben Auskunft über deren
Mindestausbreitungsgebiet.
6.1 Verbreitungsraum
An 174 Orten wurden Knochen vom klassischen Neandertaler entdeckt,
deren Fundorte sich über ganz Europa und sogar nach Vorder- und
Zentralasien erstrecken (Serangeli/Bolus 2008). Wichtige Fundplätze aus dem
Südwesten Mitteleuropas liegen auf der iberischen Halbinsel (Portugal/Spanien
und Gibraltar) und Frankreich (Bolus 2009). Besonders in Frankreich ist die
Konzentration an Funden sehr hoch. Das lässt darauf schließen, dass dieses
Areal als Kerngebiet fungiert haben könnte. An vielen Plätzen wurden großen
Mengen an Material, seien es Knochen –von menschlicher oder tierischer
(Schlachtabfälle) Herkunft– oder Werkzeuge gefunden. Der bekannteste
Fundort dürfte le Moustier bei Peysac-le-Moustier in der Dordogne (Frankreich)
sein. Von dieser Fundstelle wurde der Name das Moustérien abgeleitet der für
die lithischen Kulturen, die im Mittelpaläolithikum existiert haben, steht. Weiter
sind zu erwähnen la Ferrassie und la Micoque, die ebenfalls in der Dordogne
liegen als auch la Quina und Chapelle-aux-Saints im Département Charente
(Bolus/Schmitz 2006; Serangeli/Bolus 2008; Bolus 2009). In la Quina war die
Funddichte an Neandertalerresten besonders groß. Hier wurden von mehr als
20 Individuen Überreste ausgegraben (Martin 1913). Im Nordosten Europas
wurden in Großbritannien (Robin Hood Grotte), Belgien (Grotte de Spy),
Deutschland (Ranis) und Polen (Nietoperzowa Grotte in Jermanowice) unter
anderem Artefakte (Blattspitzen, Kratzer usw.) und fast zwei vollständige
Skelette (Spy) ausgegraben (Flas 2011). Weitere wichtige Fundplätze befinden
sich im Südosten Europas in Italien und Griechenland, wobei erwähnt werden
sollte, dass aus Griechenland bisher nur wenig Fundmaterial zur Verfügung
steht –bis 2007 waren es gerade einmal fünf mittelpaläolithische Fundplätze–
(Darlas 2007). Erst in den letzten Jahren wurden weiter Neandertalerüberreste
in der Kalamakia Grotte gefunden (Harvati et al. 2013). In Nord- (Peresani
2011; Longo et al. 2012), Zentral- und Süditalien (Boscato/Crezzeni 2012) sind
unzählige Plätze aufgetaucht, die Fundschichten von mehreren Tausend
Jahren aufweisen. Viele davon können bis zum OIS 3 datiert werden
(Grimaldi/Santaniello 2014). Im Osten Mitteleuropas, in Polen und der

19
Tschechischen Republik in der Donau Region, wurden ebenfalls Fundplätze
(Bohunice, Stránská skála) aus dem späten Mittel- und dem Anfang des
Jungpaläolithikums ausgegraben. Aus diesen Grabungen gingen Funde hervor,
die auf Industrien des Übergangs hindeuten (Škrdla 2013).
Das Ausbreitungsgebiet der Neandertaler limitierte sich aber nicht nur auf
Mitteleuropa, es erstreckte sich wie schon kurz erwähnt, bis nach Vorder- und
Zentralasien. In Vorderasien sind Fundplätze wie Amud, Kebara und Tabun in
Israel von Bedeutung, in Syrien Dederiyeh und Shanidar im Gebiet wo sich
heute der Irak befindet. In Zentralasien gilt Teshik-Tash als bekannteste
Fundstelle für klassische Neandertalerüberreste (Serangeli/Bolus 2008)21. Auch
wenn die Gebiete außerhalb Europas nicht in den bearbeiteten Zeitraum von
ca. 50.000 bis 35.000 v. Chr. passen, gehören sie zu den wichtigsten
Fundplätzen von klassischen Neandertalerüberresten. Darüber hinaus gehören
sie zu den Schlüsselgebieten in denen es bereits vor der Transition vom Mittel-
zum Jungpaläolithikum (TMJ) zu Kontakten mit anatomisch modernen
Menschen gekommen ist.
6.2 Welche archäologischen Kulturen sind von Bedeutung?
Während der TMJ hat es zahlreiche Kulturen gegeben. Zu diesen zählten
sowohl der Neandertaler als auch von anatomisch moderne Mensch. Wenn von
Kulturen gesprochen wird, handelt es sich in diesem Fall nicht um Kulturkreise
wie wir sie im heutigen Sinne verstehen, vielmehr geht es um die lithischen und
knöchernen Gegenstände, die von Gruppen hergestellt und im Nachhinein –sei
es durch Verlust, Aussonderung oder Defekt– in Höhlen, Felsüberhängen sowie
auf freien Flächen zurückgelassen wurden. Es sind jene Hinterlassenschaften,
die uns einen Einblick in den Alltag (s.d.: Kapitel 6.2) und die verwendeten
Techniken aus jener Zeit geben. Für den Zeitraum der TMJ wurden mehrere
Kulturen mit den Neandertalern in Verbindung gebracht. Zu jenen gehören das
Moustérien, das Châtelperronien, das Lincombien-Ranisien-Jerzmanowicien,
das Szeletien und das Bohunicien. Allerdings wird bei einigen immer wieder die
Frage aufgeworfen, ob es sich nun wirklich um Artefakte handelt, die von
Neandertaler Hand gefertigt wurden, oder ob nicht vielleicht doch der
21 Abb.12: Verbreitungsgebiet des klassischen Neandertalers

20
anatomisch moderne Mensch der Schöpfer dieser Gegenstände ist. Auf den
darauf folgenden Seiten wird jede archäologische Kultur mit ihren
dazugehörigen Leitformen einzeln vorgestellt. Hierbei werden sie in Zeit und
Raum platziert und kurz auf die Problematik der Einordnung eingegangen. In
einem weiteren Kapitel (8 Beispielplätze) werden dann noch ausgewählte
Beispiele vorgestellt, um ein genaueres Bild zu erstellen.
6.2.1 Das Moustérien in Europa
Das Moustérien ist eine der uns am längsten bekanntesten
Werkzeugkulturen deren Fundstätten zum Teil bereits im 19. Jahrhundert
entdeckt und ausgegraben wurden. Es war Gabriel de Mortillet, der 1872 dieser
Kultur ihren Namen –benannt nach dem Typus Fundplatz „Le Moustier“– gab.
Diese lithische Industrie war vorwiegend in Frankreich verbreitet, dennoch sind
in ganz Westeuropa, in Teilen von Westasien und sogar im Nahen Osten bis in
die Sahelzone in Afrika Fundstellen bekannt (Bordes 1981; Fiedler et al. 2011).
Seine Wurzeln hat das Moustérien –dessen Ursprung im Jungacheuléen liegt–
in Europa um circa 300 ka BP und prägte das gesamte Mittelpaläolithikum
sowie einen Teil der TMJ bis ungefähr 41.030-39.260 cal. BP (Higham et al.
2014). Stratigraphisch unterliegt es je nach Region Technokomplexen wie dem
Châtelperronien und dem teilweise parallel verlaufendem Uluzzien als auch
dem Szeletien. Zu jenem Zeitpunkt als sich diese lithische Industrie entwickelte,
war der Neandertaler der einzige Vertreter der Gattung Homo auf dem
eurasischen Kontinent. Dieser kann aufgrund von zahlreichen Knochenfunden
in Verbindung mit Artefakten eindeutig mit dem Moustérien assoziiert werden
(Bsp.: La Quina, Le Moustier usw.).
Das Moustérien ist eine polymorphe Industrie (Pradel 1954), welche mehrere
Technokomplexe in sich vereint. Überregional als auch regional unterscheiden
sich die angewandten Methoden deutlich voneinander. Es gibt eine Vielzahl an
Werkzeugen, die je nach Typ mehr oder weniger vorkommen oder gänzlich
fehlen. Zu jenen Varianten gehören unter anderem das typische Moustérien
(erste Erscheinung während der Kaltzeit Würm I bis Würm II; Bordes 1981), das
Moustérien des Charentetyps, das aus zwei Fazies besteht (Moustérien vom
Typ Quina: verwendet seit OIS 5 aber am meisten während OIS 4/3;
Bourguinon 1997; Moustérien vom Typ Ferrassie: erste Erscheinung während

21
Würm I, existierte auch während Würm II; Bordes 1981), das Moustérien „à
denticulés“ (erste Erscheinung am Anfang von Würm I und kommt relativ häufig
vor während Würm II vor; Bordes 1981), der Abbau diskoider Kerne (untypisch
für Moustérien Industrien: ab OIS 6; Soressi 2004), das Moustérien „de tradition
acheuléenne“ (MTA: eine genaue Dauer dieser Phase und der
Klingenproduktion ist aufgrund der hohen Standardabweichungen von
Datierungsmethoden nicht bekannt; Soressi 2004: 13) sowie die am meisten
und längsten verwendete Levallois-Technik (auch Moustérien à lames genannt:
verwendet ab mindestens OIS 8 bis OIS 3; Soressi 2004; Bataille 2013; laut
anderen Quellen: seit OIS 6 bis OIS 3 ❲zwischen 175.000 und 40.000 BP❳
Fiedler et al. 2011).
Das späte Moustérien
Am Ende des Mittelpaläolithikums haben zwei nahezu parallel verlaufende
Formen des Moustérien existiert. Zum einen das Micoquien, eine
Keilmessergruppe, welche von West- bis Osteuropa verbreitet war (Fiedler et
al. 2011)22. Eingegliedert wird sie in die Zeitspanne zwischen 85.000 und
45.000 BC (Jöris 2002). Ihren Namen hat diese Form vom Fundplatz la
Micoque bei Les Eyzies in Frankreich (Fiedler et al. 2011). Und zum anderen
das Moustérien „de tradition acheuléenne“, das in Westeuropa verbreitet war.
Sein Ausbreitungsgebiet reicht von den Pyrenäen bis zu den Tälern Ariège,
Tarn und Quercy (Frankreich) mit einer Großzahl an Fundplätzen im Gebiet des
Périgord (Soressi 2002) 23 . Das MTA setzt sich aus zwei Hauptgruppen
zusammen: das MTA (A) und MTA (B) (Fiedler et al. 2011) und wird
chronologisch ca. zwischen 60.000 bis 40.000 BP eingegliedert (Soressi
2002)24.
Leitformen, Technologie und Typologie
Die zwei Formen des späten Moustériens –dessen beider Ursprung das
Jungacheuléen ist– verfügen über Gemeinsamkeiten bei ihren Leitformen und
weichen bei den restlichen Gerätespektren nur graduell voneinander ab.
22 Abb.13: Geographische Verbreitung des Micoquiens 23 Abb.14: Geographische Verbreitung der Fundplätze des MTA 24 Tab.2: Radiometrische Datierungen für das MTA

22
Sowohl das MTA als auch das Micoquien verfügen über Faustkeile, dennoch
werden sie voneinander getrennt und als eigenständige Gruppen innerhalb des
Moustérien behandelt. Problematisch ist jedoch, dass die für das Micoquien
typischen Formen in vielen anderen Inventaren des Mittelpaläolithikums
vorkommen und somit immer wieder die Frage aufkommt (Fiedler et al. 2011),
ob es sich wirklich um eine autonome Gruppe handelt (Ruebens 2007).
Ruebens beantworte diese Frage folgendermaßen: (…) based on the presented evidence, the Mousterian and Micoquian are in my opinion two closely interlinked but different behavioural entities. Despite their similar basic knapping and retouching techniques, clear differences occur (Ruebens 2007: 71).
Das Micoquien gehört zu den Keilmessergruppen. Zu seinen Leitformen
gehören spitze Faustkeile, Halbkeile, kleinere ovale oder herzförmige
Faustkeile, Keilmesser, Grochaki (kleine rundliche Kratzer), blattspitzenartige
Geräte, flächig retuschierte Schaber sowie steil retuschierte einfache Schaber
(Ruebens 2007; Fiedler et al. 2011; http://altsteinzeit-hessen.de/?p=269)25.
Beim MTA kommen Faustkeile auch relativ häufig vor. Sie sind besonders
sorgfältig gearbeitet und haben vorwiegend eine dreieckige oder herzförmige
Form. Wie oben bereits kurz erwähnt, gibt es zwei Hauptgruppen, wobei jede
für sich ein individuelles Gerätespektrum umfasst. Überdies gibt es noch
weitere Zwischenformen, bei denen es zu weiteren Unterschieden der
einzelnen Geräteformen kommen kann (z.B. bei Faustkeilformen, Kostenki-
Technik, bifaciell retuschierte Schaber und Messer usw.). Das MTA (A) hat in
seinem Inventar herzförmige und flache große dreieckige Faustkeile, breite
rückenretuschierte Messer, diverse Bohrer, Stichel und Schaber und sogar
Jerzmanovice-Spitzen. Das MTA (B) hat entgegensetzt zu (A) weniger
Faustkeile, besitzt dafür aber viele rückenretuschierte Messer (Couteau à dos
Moustérien und Châtelperron-Messer). Die Faustkeile sind kleiner und von
ovaler oder herzförmiger Form. Außerdem gehören zu diesem Inventar,
gezähnte Geräte, Kratzer und grobe Stichel sowie eine gute Klingentechnik
(Soressi 2002; Fiedler et al. 2011)26.
25 Taf.1: Leitformen des Micoquiens in Mittel- und Osteruopa 26 Taf.2: Leitformen des MTA in Westeuropa

23
6.2.2 Das Châtelperronien im südwestlichen Europa (Frankreich und Spanien)
Das Châtelperronien, früher auch als Castelperronien bekannt, ist eine
typische Kultur des Übergangs vom Mittel- zum Jungpaläolithikum. Benannt
wurde sie nach der Kommune Châtelperron im Département de l’Allier
(Frankreich), wo zwischen den 1850e und 1870e in einer Grotte namens Grotte
des/aux Fées eine umfangreiche Industrie ausgegraben wurde (Gravina et al.
2005; Pelegrin/Soressi 2007). Eingegliedert wurde sie erstmalig durch den
Geistlichen Henri Breuil, einem passionierten Prähistoriker (Breuil 1912). So
fand das Châtelperronien seinen Platz zwischen dem Moustérien und dem
darauffolgenden frühen Aurignacien und wurde von ihm als erste lithische
Industrie des Jungpaläolithikums eingestuft. Trotz dieser genauen
Eingliederung und den umfangreichen Stratigraphien scheint bisher eine
genaue Datierung schwierig zu sein. Dies wird durch zwei Faktoren beeinflusst:
zum Einen stößt die 14C Methode an ihre Grenzen und zum Anderen waren
viele der anfänglichen Schichten durch Erosionsprozesse (Pelegrin/Soressi
2007; Bordes/Teyssandier 2012), die während des Würm Interstadials
stattfanden, zerstört worden (Pelegrin/Soressi 2007). Dies führte zu
Interstratifikationen, die ungenaue Datierungen zu Folge hatten. Pelegrin und
Soressi (2007) schlagen eine Datierung zwischen 40 ka bis 36 ka 14C BP vor.
Bordes (2002), Lucas et al. (2003) und Higham et al. (2010) tendieren aufgrund
der erhaltenen Resultate (2/3 von 21 14C Neudatierungen fallen in den
gezeigten Zeitraum) jedoch zu einer kürzeren Zeitspanne zwischen 41,5/40 ka
bis 38 ka 14C BP (Bordes/Teyssandier 2012). Seine Anfänge hat es
wahrscheinlich zwischen 45.000 und 40.500 BP (s.d.: Higham et al. 2010;
Talamo et al. 2012; Hublin et al. 2012 (Roussel/Soressi 2014)). Zur
Datierungsproblematik kommt hinzu, dass nicht ersichtlich ist, welchen
Ursprung das Châtelperronien hat. Hinsichtlich der Rückenretuschierten
Artefakte im Inventar schlussfolgerten Peyrony (1948) und Bordes (1954/55),
dass sich die Industrie aus dem Moustérien de „tradition acheuléene“ B heraus
entwickelt hat. Stratigraphische Superposition (Châtelperronien/Moustérien)
wurde auch gerne angeführt, um die Idee von Kontinuität im technischen als
auch im kulturellen Sinne zu bestätigen (Vgl.: Lucas et al. 2003: 290). Diese
Theorien wurde seitdem mehrfach mit Hilfe weiterer Argumente versucht zu

24
bekräftigen (s.d.: Pelegrin 1995; d’Errico et al. 1998; Soressi 2002). Neben dem
MTA (B) wurde von Leroi-Gourhan das Moustérien „à denticulé“ als möglicher
Ursprung vorgeschlagen (Lucas et al. 2003). Da beim Châtelperronien weder
eine konkrete Weiterentwicklung aus dem MTA (B) noch aus dem Moustérien
„à denticulé“ zu erkennen ist (Bordes/Teyssandier 2012), ist bis Dato nicht
vollends geklärt aus welcher Kultur sie sich entwickelt hat. Die Interpretationen
(das Moustérien de „tradition acheuléene“ B als Ursprung) von Peyrony und
Bordes sind dennoch bis heute allgemein gültig (Lucas et al. 2003;
Pelegrin/Soressi 2007). Heimisch ist das Châtelperronien im gesamten
Südwesten Frankreichs und Teilen Spaniens, wobei Teile Osteuropas auch
beeinflusst wurden (Fiedler et al. 2011). Die größte Konzentration an Funden
befindet sich jedoch im Département der Dordogne in Zentralfrankreich (Bsp.:
La Ferrassie, Le Moustier, Roc-de-Combe-Capelle usw.)27.
Leitformen, Technologie und Typologie
Das Châtelperronien ist vor allem für seine Klingenproduktion mit seinen
Spitzen und Messern bekannt. Besonders charakterisierend ist die
Châtelperron-Spitze (Châtelperron-Messer), dessen Rücken steil retuschiert ist
und leicht bogenförmig verläuft (Fiedler et al. 2011). Die bearbeitete Basis weist
darauf hin, dass einige dieser Spitzen geschäftet waren. Es wurden vor allem
zwei Typen an Gebrauchsspuren identifiziert: Abgebrochene Spitzen verweisen
auf einen Gebrauch an einer Lanze zu Jagdzwecken hin, Abnutzungsspuren
entlang der Klinge hingegen auf einen Gebrauch als Messer (Pelegrin/Soressi
2007). Die Klingen sind in der Regel zwischen 3 und 7 cm lang. Neben der
klassischen Variante existiert noch die „Pointe des Cottés“ (Fundstelle Saint-
Pierre-de-Maillé), eine schlankere Form des Châtelperron-Messers (Pradel
1963; Fiedler et al. 2011) sowie weitere Übergangsformen.
Des Weiteren gehören zum charakteristischen Inventar Klingen mit
unregelmäßiger Retusche, Messer mit Rücken, einige Bohrer (Becs),
Klingenkratzer und etwas dickere Rundkratzer. Seltener findet man auch
Schaber, einige gezahnte und zersplitterte Stücke als auch Bohrer. Aufgrund
ihres schlechten Erhalts kommen Knochengeräte nur sehr selten vor –mit dem
27 Abb.15: Geographische Verteilung der Hauptfundplätze des Châtelperroniens (Abb. nach Pelegrin 1995)

25
Fundplatz d’Arcy-sur-Cure als Ausnahme– und sind darüber hinaus nicht sehr
vielfältig vertreten. Aus der entwickelten Phase sind dennoch Schmuckstücke
aus Zähnen oder Knochen und schlanke Knochenspitzen bekannt
(Pelegrin/Soressi 2007)28.
Die Klingen des Châtelperronien wurden vor allem aus Kernen mit
gegenüberliegenden Schlagflächen gewonnen. Wahrscheinlich handelt es sich
bei dieser Klingentechnik um eine Weiterentwicklung aus Levallois-
Klingenkernen (Fiedler et al. 2011).
6.2.3 Das Lincombien-Ranisien-Jerzmanowicien im Nordosten Europas
(Großbritannien, Belgien, Deutschland und Polen)
Das Lincombien-Ranisien-Jerzmanowicien (LRJ) gehört zu den weniger
bekannten und weniger gut erforschten Industrien der TMJ. Da dieser
Technokomplex im Grunde genommen lediglich aus einem Faziestyp
(Blattspitzen) besteht (Flas 2011a), wurde das LRJ lange nicht als eigene
lithische Kultur anerkannt und dementsprechend behandelt. Die ersten
Blattspitzen wurden in Grabungen diverser Länder (Großbritannien,
Deutschland Belgien und Polen) im 19e und Anfang des 20e Jahrhunderts
entdeckt. Aufgrund eines kleinen Fundus (dadurch bedingt, dass es sich bei
den meisten Fundplätzen um Jagdplätze handelt) und schlecht ausgeführter
Grabungen kam es vermehrt zu Vermischungen der Spitzen mit anderen
Inventaren (Flas 2011b). Über die Jahre wurde von diversen Kollegen
deswegen des Öfteren versucht diesen Faziestyp in bereits bestehende und
anerkannte Technokomplexe aus Frankreich zu integrieren (Flas 2002). So
ordnete Henri Breuil ihn zum Beispiel dem Protosolutréen zu (Flas 2006). Seine
Aussage hatte soviel Gewicht, dass vor allem die Funde aus England und
Belgien für längere Zeit diesem Komplex zugewiesen wurden. Nach weiteren
Grabungen in der Nietoperzowa Grotte in Jerzmanowice und Vergleichen mit
anderen Funden (Kostenki Tel’manskaya und Ranis Schicht X ❲Ranisien❳) war
es W. Chmielewski (1961), der die Eigenständigkeit der Blattspitzengruppe
erkannte und den Begriff „Jerzmanowicien“ für die polnische Variante prägte
(Flas 2006, 2008). 1978 erkannte M. Otte einen Zusammenhang zwischen den
Faziestypen aus Belgien und Deutschland und 1980 wurden schlussendlich die 28 Taf.3: Leitformen des Châtelperroniens

26
Blattspitzen aus Großbritannien durch J.B. Campbell unter dem Namen
„Lincombien“ vereint. Aufgrund der signifikanten Gemeinsamkeiten waren es
Desbrosse und Kozłowski, die 1988 den endgültigen Sammelbegriff „Komplex
des Lincombien-Ranisien-Jerzmanowicien“ festlegten (Flas 2006). 2008
bekräftigte Flas in seiner Dissertation auf ein Neues, dass es sich beim LRJ um
einen eigenständigen Technokomplex handelt. Den Ursprung dieser lithischen
Kultur zu ermitteln, stellt sich als genauso schwierig heraus und bleibt bis Dato
unklar. Als mögliche Ursprungsindustrie wurde des Öfteren eine
mittelpaläolithische Industrie das „Altmühlien“ aus Mauern vorgestellt (z.B.:
Kozłowski 1990; Bosinski 2001), mit einem hypothetischen Werdegang von
Mauern F, über Ranis 2 zu Nietoperzowa Schicht 6 (Flas 2008, 2011a). Flas
bemerkt jedoch, dass Ranis 2 das älteste Ensemble des LRJ ist und aus der
Anfangsphase des OIS 3 stammt und somit besser als Wegweiser für den
Ursprung des LRJ geeignet sei (Flas 2006: 595).
Das LRJ ist über weite Teile Nordost-Europas 29 verteilt, wobei in
Großbritannien (29 Fundkomplexe) die größte Konzentration an Funden
herrscht. In den Niederlanden, im Norden Deutschlands und dem Krakau-
Tschenstochauer Jura fällt die Anzahl wesentlich geringer aus (Vgl.: Flas
2011a: 261). Bisher konnten 40 Fundplätze dem LRJ sicher zugewiesen
werden. Den Komplex chronologisch einzugliedern gestaltet sich als schwierig,
da es während früher Grabungskampagnen häufig zu Vermischungen mit dem
darunter oder darüber liegenden Moustérien und/oder Aurignacien/Gravettien
kam. Dennoch gibt es einige Daten die mit Hilfe der 14C Methode ermittelt
werden konnten. 1961 ließ Chmielewski eine Kohleprobe aus Schicht 6 der
Nietoperzowa Grotte per 14C analysieren und erhielt eine Datierung von 38.160
+/- 1.250 BP. Anhand einer weiteren Probe der gleichen Schicht –dieses Mal
aus einem Stück Tierknochen gewonnen– erhielt Kosłowski (2002) eine
Datierung von 37.600 +/- 1.300 BP und bestätigte somit den ersten
Datierungsversuch. Weiter wurde in England (Pin Hole, Badger Hole und Bench
Tunnel Cavern) mit Hilfe von Ultrafiltration eine Datierung zwischen 37.760 +/-
340 und 36.000 +/- 450 BP ermittelt. Hier gilt es jedoch zu erwähnen, dass die
Proben aus einem stratigraphisch problematischen Kontext stammen. Eine
Knochenprobe aus Glaston ergibt eine Datierung um 38.000 BP (Flas 2011a).
29 Abb.16: Geographische Ausbreitung des Lincombien-Ranisien-Jerzmanowicien

27
Als jüngste Datierung ging die verbrannte Blattspitze aus Beedings mit einer
Datierung von 31.100 +/- 5.700 BP (TL) hervor, welche als eventuelles
Mindestalter angesehen werden sollte (Jacobi 2007). Laut Flas (2011a) gibt es
jedoch keine zuverlässigen Angaben dafür, dass das LRJ länger als 35.000 BP
existiert hat. Eine Datierung sei demnach älter als das Resultat aus Beedings
anzusetzen. Er schlägt daher eine Datierung zwischen 39.000 uncal. BP und
35.000 uncal. BP vor (Flas 2011a).
Da das LRJ genau in die Zeitspanne fällt während der sich der anatomisch
moderne Mensch in Europa ausbreitet und der Neandertaler kurz vor dem
Aussterben steht, stellt sich die Frage wer eigentlich die Macher dieser
Industrie sind. Trotz Funden von Knochen in einigen Fundplätzen (Kent’s
Cavern, Grotte de Spy, trou de l’Âbime à Couvin, trou de Walou) herrscht
Uneinigkeit. Die eine Fraktion (S. Swainston 1999; van Andel et al. 2003) sieht
den anatomisch modernen Menschen als alleinigen Macher, aufgrund des
„modernen“ Charakters der Industrie. Die andere und auch größere Fraktion
(s.d: Otte 1990; Kozłowski 1995; Jacobi 1999; Pettitt 1999, Flas 2008) jedoch
den Neandertaler, da sie eine Kontinuität von lokalen Gruppen aus dem
Mittelpaläolithikum darin sehen (Flas 2008: 130). Es ergibt sich bei den
gefunden Knochen das Problem, dass sie sich nicht in direkter Verbindung mit
LRJ Blattspitzen befanden und gelegentlich aus unsicheren Kontexten (Kent’s
Cavern) stammen. Aufgrund des geringfügigen Knochenmaterials (Kent’s
Cavern) konnten einige Überreste nicht eindeutig einer Gattung zugeordnet
werden. Andere Knochenreste wiederum konnten als die von Neandertalern
identifiziert werden und passen chronologisch in den Zeitraum des LRJ (Spy:
36.000 BP), oder werden als möglichen Ursprung angesehen (trou d’Âbime de
Couvin; trou de Walou: ca. 40.000 BP; Flas 2008). Dennoch bleiben eindeutige
Beweise für eine genaue Zuordnung aus. Aus mehreren Gründen wird trotzdem
eher dazu tendiert das LRJ dem Neandertaler zuzuschreiben (Flas 2008):
1. Am Ende des Mittelpaläolithikums war der Neandertaler der einzige
Vertreter der Gattung „Homo“ im Norden Europas. Eine
Weiterentwicklung zu Übergangsindustrien ist somit gar nicht so
unwahrscheinlich.
2. Die Datierungen der Knochen von Spy passen in den Zeitraum des
LRJ und fallen zu früh aus, um dem Moustérien anzugehören.

28
3. Die ersten anatomisch modernen Menschen treten in geographischer
Entfernung zum LRJ auf und fallen jünger aus.
Flas (2008: 130) bemerkt jedoch, dass das LRJ ein technokultureller
Komplex ist, welcher chronologisch wie auch geographisch weit verbreitet ist
und somit die Möglichkeit besteht, dass diese Technik der Blattspitzen-
Herstellung, hypothetisch gesehen, von zwei verschiedenen Gattungen hätte
verwendet werden können.
Leitformen, Technologie und Typologie
Wie bereits weiter Oben erklärt, besteht die Industrie bis Dato aus einem
einzigen Faziestyp. Hierbei handelt es sich um die sogenannten Blattspitzen,
wovon es zwei Varianten gibt. Zum Einen, eine partiell retuschierte Blattspitze
aus einer Klinge und zum Anderen eine bifacielle Blattspitze aus einem
Abschlag, wobei erstere vom Umfang her dominiert30. Die Umrissform ist nicht
standardisiert sondern fällt sehr variabel aus. So können Jerzmanowice-Spitzen
zum Beispiel lanzettförmig und gestreckt-oval sein. Seltener wurde auch eine
schlanke Herzform festgestellt. Des Weiteren sind sie proximal bearbeitet und
somit für eine Schäftung geeignet. Die Symmetrie der Stücke deutet auch
darauf hin (Flas 2002). Die Spitzen haben im Durschnitt eine Dicke von 1 cm,
sind ca. 3 cm breit und zwischen 9 und 10 cm lang (Flas 2006).
In einigen Fundinventaren wurden neben Blattspitzen andere
Steinwerkzeuge festgestellt. Flas (2002) weist darauf hin, dass diese aufgrund
ihrer Bearbeitungsweise dem LRJ zugewiesen werden sollten. Ein Teil dieser
Artefakte stammt aus stratigraphisch relativ intakten Kontexten (Ranis 2),
andere wiederum aus stratigraphisch gestörten Kontexten. Darunter befinden
sich unter anderem angespitzte Klingen, welche morphologisch den
Blattspitzen ähneln, retuschierte dickere Klingen, bifacielle Kratzer (Racloir) und
Schaber, blattförmige Abschlagkratzer und retuschierte Klingenkratzer (Ranis
2); Stichel und Kratzer (Grattoir) (Nietoperzowa Grotte); Kratzer, Doppelkratzer,
Bohrer, gezähnte Klingen, retuschierte Klingen und mit Kerben versehene
Artefakte (Badger Hole, stratigraphisch unsicherer Kontext) (Flas 2002).
Des Weiteren gesellen sich zu den Steinwerkzeugen des LRJ auch
Knochengeräte und Geräte aus Bein bzw. Fragmente (Lanzenspitze). Sie 30 Taf.4: Blattspitzen aus dem LRJ. Verschiedene Beispiele.

29
wurden vorwiegend in britischen Inventaren (Hyaena Den, Efynnon Beuno
Cave, Uphil Soldier’s Hole) entdeckt. Allerdings werden diese Funde vielseitig
angezweifelt, da sie aus stratigraphisch unsicheren Kontexten stammen und
teilweise sogar in andere Perioden (Aurignacien) eingegliedert wurden 31 .
Angesichts der unsicheren Herkunft der Artefakte, die neben den Blattspitzen
ausgegraben wurden, ist es schwierig jene eindeutig dem LRJ zuzuordnen.
Lediglich zwei aus Ranis 2 stammende Artefakte (Stichel aus Knochen und ein
Fragment eines kreisförmigen durchlöcherten Gerätes aus Elfenbein) könnten
dem LRJ zugeordnet werden (Flas 2014).
6.2.4 Das Szeletien im östlichen Mitteleuropa
Das Szeletien ist eine der Schlüsselkulturen des Übergangs vom Mittel- zum
Jungpaläolithikum im östlichen Mitteleuropa (Mester 2014). Es gehört wie das
LRJ zu den Blattspitzengruppen und lässt sich als einzige Kultur eindeutig den
Neandertalern zuordnen (Allsworth-Jones 1986). Das führte des Öfteren dazu,
dass versucht wurde Blattspitzen aus anderen Inventaren, wie z.B. aus dem
LRJ oder dem Bohunicien, in das Szeletien einzugliedern (Flas 2002). Der
Name „Szeletien“ wird als Sammelbegriff verwendet und vereint
unterschiedliche Ensembles, die an sich nur Blattspitzen als gemeinsamen
Faktor haben (Mester 2014). Diese lithische Kultur ist in seiner Ausbreitung auf
einen kleinen Verbreitungsraum begrenzt, der sich von Ungarn, der
Tschechischen Republik (Mähren), der Slowakischen Republik bis nach Polen
als nördlichster Punkt erstreckt (Weißmüller 1995; Martin 1995; Mester 2014)32.
Seit seiner Entdeckung in Ungarn wurde darüber gerätselt welchen Ursprung
das Szeletien haben könnte. Seit den Anfängen des 20e Jahrhunderts wurden
die unterschiedlichsten Theorien vorgelegt (Entwicklung des Protosolutréen aus
dem lokalen Moustérien: Breuil 1923; Er widersprach Breuil und sah eine
Vermischung von Neandertalern und Cro-Magnon als Ursprung: Hillebrand
1926; Übernahm die Idee von Breuil, sah aber noch einen Einfluss des
Aurignaciens in den Funden: Freund 1952; Ursprung im lokalen Moustérien aus
dem Karpatenbecken mit Einfluss des Aurignaciens während Würm I und
Anfang von Interstadial Würm I/II: Prošek 1953; Vertés 1964). Die Idee, dass 31 Taf.5: Leitformen des LRJ 32 Abb.17: Verbreitungsgebiet des Szeletiens

30
das Szeletien sich aus dem lokalen Moustérien entwickelt hat, wurde
mittlerweile komplett verworfen. 1967 schlug Bosinski das Micoquien aus
Zentraleuropa als möglichen Ursprung vor. Funde aus verschiedenen Regionen
aus den darauffolgenden Jahren bekräftigen diese Idee (s.d.: Gábori 1976; Otte
1996, Kozłowski/Kozłowski 1996; Valoch 1996) Mester 2014). Nachdem Ringer
(1983) in der Umgebung von Kiskolc lithische Artefakte mit einem Micoquien
Charakter aufgelesen hatte und jene als Bábonyien beschrieb, schlug er diese
als weitere Möglichkeit vor. Seitdem wurde versucht einen Zusammenhang
zwischen dem Szeletien und dem Bábonyien herzustellen (s.d.: Ringer 2008-
2009 ❲Mester 2014❳). Da es aber keine genaueren Informationen über
spätpaläolithische Kulturen in der Region gibt, kann nur angenommen werden,
dass das Szeletien seine Wurzeln in Transdanubien, in der ungarischen
Version des Micoquiens (Bábonyien) oder evtl. sogar dem Jankovichien hat
(Vgl.: Neruda/Nerudová 2013, 4).
Eine genaue Datierung des Szeletien erweist sich als schwierig. In den
letzten 20 Jahren wurde eine Serie von Datierungen an verschiedenen
Fundplätzen (Vedrovice V: Valoch et al. 1993; Moravský Krumlov IV, vr. 0:
Neruda et al. 2004 und Nerudová/Davies 2009; Moravany-Dlhá (SK): Nemergut
2010; Brno-Bohunice II: unpubliziert; Pod Hradem: unpubliziert 33 )
vorgenommen. Die daraus resultierenden Datierungen weisen auf einen
Zeitraum von ca. 43.900 +/- 520 cal. BP bis 38.600 +/- 1.590 cal. BP hin,
dennoch sollte aufgrund unterschiedlicher Probleme –sei es durch Verlust von
Horizonten, um Datierungen zu verifizieren, Knochenproben ohne
anthropogene Spuren, Interstratifikationen usw.– jener Zeitraum nicht als
absolute Chronologie für diese lithische Kultur verwendet werden
(Neruda/Nerudová 2013). Grob wird das Szeletien zwischen 45.000 und 35.000
BP eingegliedert (Fiedler et al 2011).
33 Tab.3: Kalibrierte Daten in Bezug auf die Chronologie des Szeletiens (Tabellenausschnitt von Tab.3 bei Neruda/Nerudová (2013).

31
Leitformen, Technologie und Typologie
Da viele Funde aus alten Grabungen (Interstratifikation, Verluste usw.)
stammen, stellt sich auch beim Szeletien die Frage welche lithischen Artefakte
wo einzuordnen sind. Die folgende Aufzählung an Artefakten wird daher als
problematisch angesehen. 1968 stellte Vértes ein Modell der Leitformen aus
Schichten der Szeleta und Jankovich Grotte vor. Blattspitzen kommen mit 36%
am häufigsten vor, zu denen gesellen sich Schaber, die mit 23% die
zweithäufigsten Artefakte darstellen, danach folgen Klingen mit 17%, mit
Kerben versehene Artefakte (Encoche) 10% und dann Kratzer und Stichel mit
7%.
In den 1990e untersuchte Karel Valoch Ensembles aus der Gegend von
Brno, von deren Integrität er überzeugt war. Aus seinen Recherchen ging
hervor, dass das Szeletien einen transitorischen Charakter hat (Mester 2014).
Eine weitere Grabung (Vedrovice V) belegte die vorher erhaltenen Resultate. In
der Grabung von Vedrovice V wurden ca. 17.000 Artefakte in situ gefunden.
5.000 davon flossen in die Wertung ein. 24% der Werkzeuge haben einen
jungpaläolithischen Charakter und 35% einen mittelpaläolithischen. Ein Drittel
des Ensembles setzen sich aus mit Kerben versehenen Artefakten (27%) und
gezähnten Stücken (14%) zusammen. Sie können weder dem Jung- noch dem
Mittelpaläolithikum zugeordnet werden. 20% bestehen aus mittelpaläolithischen
Schabern, die sehr unterschiedlich ausfallen, dennoch einfach gearbeitet sind.
Blattspitzen (ovaler, dreieckiger Form oder als Doppelspitze) sind dagegen nur
mit 9,5% vertreten. Faustkeile und Messer repräsentieren 1,5% und
Moustérien-Spitzen nur 0,5%. Im jungpaläolithischen Teil sind die aus
Abschlägen gefertigte Kratzer mit 8% am häufigsten vertreten, darauf folgen
retuschierte Abschläge und Klingen mit 7%. Meist abgewinkelte Stichel sind zu
3,5% vorhanden, Bohrer zu 2,5% und abgesplitterte Stücke zu 1,8% (Valoch et
al. 1993; Mester 2014)34.
An einem weiteren Beispiel von Neruda und Nerudová aus dem Jahr 2005
wurde gezeigt, dass im Szeletien drei Methoden der Chaîne opératoire
verwendet wurden. Für Abschläge und Klingen wurde die diskoide Kerntechnik
und die Klingentechnik verwendet. Für beide Abläufe benötigt man einen Kern
34 Taf.6: Leitformen des Szeletiens

32
(entweder vielseitig, segmentförmig, dreieckig, gestreckt barrenförmig) von dem
das Material abgearbeitet wird. Die bifacielle Bearbeitung wurde für das
Herstellen von blattförmigen Werkzeugen verwendet. In den seltensten Fällen
wurde neben den drei Methoden die Levallois Methode hinzugezogen (Mester
2014).
6.2.5 Das Bohunicien im östlichen Mitteleuropa
Im mittleren Donaugebiet waren im Laufe der TMJ zwei Übergangsindustrien
beheimatet. Einerseits das zuvor vorgestellte Szeletien und andererseits das
Bohunicien (Škrdla 2013). Seit dem Ende des 19. Jahrhunderts tauchten immer
wieder Oberflächenfunde mit Levallois ähnlichem Charakter und mit
Übergangscharakter im Brno Bassin auf (Škrdla 2014). Der wichtigste
Fundplatz befindet sich in Bohunice-Kejbaly einem Vorort im Süden der Stadt
Brno (Tostevin/ Škrdla 2006; Richter et al. 2008). Er wurde erstmalig von
Valoch ausgegraben und fungierte als Namensgeber der dort entdeckten
Industrie (Bohunicien) (Valoch 1976). Stránská skálá (s. Kapitel 8.14) ist eine
weitere wichtige Fundstelle des Bohunicien (Valoch et al. 2000, 2009), als auch
des Aurignacien in diesem Gebiet. Beide Stätten besitzen eine umfangreiche
Stratigraphie. Die größte Konzentration an Fundstellen befindet sich im Bassin
von Brno (Mähren) bis auf eine Distanz von 40 km außerhalb der Stadt (Škrdla
2014). Drei weitere Ansammlungen an Funden befinden sich im Mohelno
Gebiet, im Prostějov Gebiet und im Bobrava Gebiet (Škrdla 2013). Des
weiteren existieren Fundplätze des Bohuniciens in Polen, in der Tschechischen
Republik (Dzierzyslaw I/Třebom), im nördlichen Zentralböhmen
(Hradsko/Sedlec), in der Ost-Slovakei, die Ukraine (Kulychivka) und in
Bulgarien (Meignen, Temnata Grotte; Škrdla 2013, 2014)35.
Um das Bohunicien in Mähren in einen Zeitraum einzugliedern, wurden drei
verschiedene Methoden verwendet. Zu jenen gehören die 14C Methode, die TL
und die OSL Datierung (Škrdla 2014). Mit der 14C Methode wurden drei Serien
berechnet (Valoch 1976; Svoboda 1991, 2003; Škrdla/Tostevin 2005). Die
erhaltenen Resultate besitzen eine weite Bandbreite und reichen von 48 bis 40
ka BP. Die TL Daten stammen aus den untersten Schichten des Fundplatzes
35 Abb.17: Verbreitungsgebiet des Bohuniciens

33
von Brno-Bohunice Grabung aus dem Jahr 2002. Es wurden 11 Proben
untersucht. Die erhaltenen Datierungen ergaben einen Durchschnittswert von
48,2 +/- 1,8 ka BPTL (Richter et al. 2008)36. Mit der OSL Methode wurden in
Bohunice 2002 und Stránská skálá Sedimentproben verschiedenster Level
(Boh_Area A, Boh-TP1&2; SS- IIIc1&c2) analysiert. Es gingen Resultate hervor,
die den untersten Schichten ein Alter von 104,3 +/- 10 ka (Boh_Area A) bzw.
141,90 +/- 11 ka (SS-IIIc1) zuweisen. Die jüngsten Schichten wurden auf ein
Alter von 30,9 +/- 3,1 ka (Boh_Area A) bzw. 26,60 +/- 2,30 ka (SS-IIIc1)
datiert 37 . Radiokarbon und OSL Daten haben eher die Tendenz durch
geologische Prozesse beeinflusst zu werden und können daher ungenauere
Resultate liefern. In diesem Fall lagen die erhaltenen TL Daten näher an der
Chronostratigraphie der besagten Fundplätze (Škrdla 2014). Das Bohunicien
gilt als älteste Kultur der TMJ in dieser Region und wird in einen Zeitraum von
ca. 50 bis 40 ka BP eingegliedert. Neben ihr existieren parallel noch zwei
weitere Kulturen wie das Szeletien und das frühe Aurignacien (z.B.: Willendorf
II; Nigst et al. 2008). Das Bohunicien, als auch das Szeletien verschwanden
kurz nach 40 ka BP. Die KI Eruption (ca. 40/39 ka cal. BP) und das
darauffolgende H4 können für das Ende beider Kulturen als
chronostratigraphische Marker gesetzt werden (Hoffecker et al. 2008). Nach
ihrem Verschwinden breitet sich das mittlere Aurignacien aus (Škrdla 2014) und
leitet somit endgültig das Jungpaläolithikum in Mähren und seiner Umgebung
ein.
Leitformen, Technologie und Typologie
Als typische Industrie der TMJ besitzt das Bohunicien in seinem
Gerätespektrum, Artefakte mit einem mittelpaläolithischen, als auch
jungpaläolithischen Charakter. Zu Artefakten mit mittelpaläolithischem
Charakter zählen eingekerbte und gezähnte Geräte, Schaber, meist
unretuschierte längliche Levallois Spitzen sowie retuschierte Levallois Spitzen,
die aber weniger häufig vorkommen. Dazu gesellen sich Moustérien-, Quina-
und Châtelperron-Spitzen. Im Grabungsinventar von Bohunice findet man
zudem Blattspitzen, als auch bifaciell retuschierte Spitzen. Das

34
jungpaläolithische Ensemble setzt sich aus anderen Artefakten zusammen.
Kratzer mit Aurignacien ähnlichen Formen treten am häufigsten auf. Sie wurden
aus Abschlägen oder Klingen (mit oder ohne oder nur teilweise erhaltenem
Kortex) hergestellt, die mittels Levallois Technik, aber auch anderen Methoden
gefertigt wurden. Dazu kommen noch einige Stichel (Valoch et al. 2009;
Neruda/Nerudová 2013; Škrdla 2014)38.
Die Technologie des Bohuniciens basiert auf einer konzeptuellen Fusion der
Levallois Technik und jungpaläolithischen Technologien (Škrdla 2003). Das
bedeutet soviel, dass zwei Methoden der Reduktion auf einem Kern ausgeführt
wurden. Zu dieser Erkenntnis gelangte man nachdem aus Stránská skálá 14
Kerne wieder zusammengesetzt und analysiert wurden (Škrdla 2003). Das
Material stammt zu 90% aus der Region (Stránská skálá Felswand, Krumlovský
und Ondratice/Želeč), nur ein geringer Teil wurde importiert. Feuerstein- und
Ortho-Quarzit-Artefakte entspringen der lokalen Produktion, während Radiolarit
(Karpaten) und Flint (Mähren) importiert wurden (Neruda/Nerudová 2013;
Škrdla 2014).
6.3 Subsistenzverhalten
In jedem Zeitalter entwickeln sich Kulturen mit für ihre Periode spezifischen
Fertigkeiten. Diese erlernten Fähigkeiten, die Neugierde und der Drang neue
Landstriche zu erobern und zu besiedeln sowie deren Anpassungsfähigkeit,
tragen zu einem erfolgreichen Überleben bei. Das Subsistenzverhalten von
prähistorischen Gruppen wird seit der Hälfte des 20e Jahrhunderts vermehrt
diskutiert. Es war Bordes, der (1953) in seinem «Essai de Classification des
industries „moustériennes“» erstmalig auf die Verhaltensunterschiede von
Neandertalern aufmerksam machte (Delagnes/Rendu 2011). Mitte der 1960e
wurde das Thema der Großwildjagd und seine Konsequenzen auf soziale
Strukturen sowie das Verhalten unserer Urahnen mit auf die Agenda des
Symposiums „Man the Hunter“ gesetzt (Vgl.: Gaudzinski-Windheuser/Kindler
2012: 59). Fortan erhielten Subsistenzstrategien Einzug in die Wissenschaft
und wurden fester Bestand unterschiedlicher Untersuchungen. Das
Subsistenzverhalten –Umgangssprachlich auch: sämtliches Eingreifen von
Menschen in die Natur oder seine Umwelt, sei es durch Jagd oder andere 38 Taf.7: Leitformen des Bohuniciens

35
Tätigkeiten, um seinen Lebensunterhalt zu gewährleisten– umfasst viele
verschiedene Punkte, wie Technologie, Mobilität, Jagdstrategien, Beuteschema
usw., die gleichermaßen behandelt werden sollten. In den letzten Jahrzehnten
standen vor allem die Steinkulturen mit ihren dazu gehörigen Chaînes
Opératoires im Fokus. So wurde zwar der Weg vom Rohmaterial zum fertigen
Artefakt verstanden, der kulturelle Hintergrund blieb allerdings im Dunkeln.
Binford (1978, 1980) gilt auf dem Gebiet der Ethnoarchäologie als Pionier
(Delagnes/Rendu 2011) und ebnete den Weg weitere Bereiche zu erforschen
und ein besseres Verständnis für paläolithische Lebensweisen zu entwickeln. In
den letzten fünf Jahren beschäftigte sich die Forschung wieder vermehrt mit
Subsistenzstrategien. Unter anderem standen die Mobilität und Technologie
(Daujeard/Moncel 2010; Delagnes/Rendu 2011), die Landnutzung
(Daujeard/Moncel 2010; Djindjian 2011), das Jagdverhalten/Jagdbeuteschema
(Gaudzinski-Windheuser/Kindler 2012) mit dem gesamten Nahrungsspektrum
(Richards/Trinkaus 2009; Ready 2013; Fiorenza et al. 2015) im Mittelpunkt von
Untersuchungen.
6.3.1 Lebensweise
Die Lebensweise des Neandertalers lässt sich am besten als die eines
Jägers und Sammlers bezeichnen, der ein nomadisches, eventuell auch ein
seminomadisches Leben führte. Je nach Region (steil abfallende bewaldete
oder felsige Plateaus, Täler sowie kalte und offene Steppen) und Saison
standen ihm unterschiedliche Ressourcen zur Verfügung, die er durch
vorausschauendes Planen verstand zu verwerten. Zu den üblichen Habitaten
gehörten Höhleneingänge mit großen Vorhallen, Felsnischen, aber auch
kleinere Höhlenkammern an steilen Felsvorhängen sowie selbst gebaute
Hütten aus Knochen und Tierhäuten. Letztere waren an Freilandplätzen zu
finden und blieben nur selten erhalten, sodass kaum etwas über diese Art der
Behausung bekannt ist (Daujaerd/Moncel 2010). Dennoch gibt es einige
Bespiele aus unterschiedlichen Perioden, wie zum Beispiel ein Fundplatz mit
einer umfangreichen Stratigraphie aus der Ukraine (Moldovo I, Schicht 4,
∼44.000 BP), der zeigt, dass Mammuts nicht nur als Nahrungsmittel fungierten,

36
sondern deren Knochen auch zum Bau von Hütten verwendet wurden (Demay
et al. 2012).
Um einen gewissen Überblick über Subsistenzstrategien und die
Organisation von Territorien zu bekommen, wurden in den letzten 15 Jahren
lithische Artefakte und faunische Überreste analysiert. Der Typ und die Anzahl
geben Auskunft über die Mobilität von Gruppen innerhalb eines Territoriums.
Außerdem können sie dazu beitragen den Umfang (Dauer und Größe) eines
ausgebeuteten Gebietes sowie die Art der Behausung, um welche es sich
handelt zu ermitteln (Daujeard/Moncel 2010). Zwei Mobilitätsmodelle können
anhand der Relation zu den Steinartefakten identifiziert werden:
1. Zirkulationsmodell: Dieses Modell besagt, dass kleine Gruppen
Saisonal viel unterwegs sind. Plätze an denen sie verweilen sind von
kurzer Dauer und sind für spezifische Aufgaben gedacht. Dazu
gehören unter anderem Jagd- und Schlachtplätze. Die Steingeräte
sind in der Regel spezialisiert oder multifunktional und kommen aus
weiter Entfernung. Diese gehörten zur persönlichen Ausrüstung einer
Person. Da Rohmaterial nicht immer greifbar war, wurde
dementsprechend auf diese Geräte aufgepasst.
2. Satellitenmodell (radiating model) oder Zentralort Modell (central place
foraging model): Dieses Modell besteht darin, dass Gruppen (evtl.
größere Familiengruppen) Langzeitcamps mit in der Umgebung
verteilten Satellitencamps haben von denen aus die benötigten
Ressourcen in Exkursionen herangeschafft werden. Viele der
Artefakte werden erst im Basiscamp hergestellt und kommen in Form
von Knollen ins Camp (Daujeard/Moncel 2010; Morin 2012).
Die räumliche Organisation limitiert sich jedoch nicht nur auf große
Territorien, wie das Beispiel von Riparo Bombrini (Nordwest Italien) zeigt. Aus
Untersuchungen der Schichten des späten Moustériens ging hervor, dass der
Neandertaler seine Camps je nach Mobilitätstrategie nach einem gewissen
Muster ordnete und Schlaf-, Arbeits- (z.B.: Steinbearbeitung) und Kochbereich
voneinander zu trennen wusste. Dies ist eine Eigenschaft, die bisher nur dem
anatomisch modernen Menschen zugeschrieben wurde (Vaquero/Pastó 2001;
Riel-Salvatore et al. 2013).

37
6.3.2 Jagdwaffenspektrum
Steinzeitliche Jagdwaffen haben eine lange Geschichte der Entwicklung
hinter sich, die vom normalen Stein bis hin zu Pfeil und Bogen reicht. Viele
faunische Überreste weisen darauf hin, dass der Neandertaler ein
Großwildjäger war und große Mengen an Fleisch konsumiert hat, dennoch sind
Jagdwaffen aus vergänglichem Material in archäologischen Inventaren rar
gesät. Nur in Ausnahmefällen, wenn die richtigen Konservierungsbedingungen
gegeben sind, bleiben leicht vergängliche Materialien aus Holz, Geweih,
Knochen, Federn, Leder, Fell, Binde- oder Klebemittel erhalten
(Stodiek/Paulsen 1996). Nur knapp 20 % des Jagdwaffenspektrums bleiben
bestehen. Steine (10%) haben die besten Erhaltungschancen, da sie jeglicher
Witterung gut standhalten. Knochen und Geweih (10%) platzieren sich an
zweiter Stelle, denn auch sie haben je nach Lagerung ausreichend bis gute
Chancen die Zeit zu überdauern39.
Zu den häufigsten Relikten steinzeitlicher Jagdwaffen gehören Projektile.
Man vermutete, dass sie ab dem späten Mittelpaläolithikum (vor etwa 60.000
bis 40.000 Jahren) verwendet wurden (Stodiek/Paulsen 1996). Einige Spitzen
weisen jedoch darauf hin, dass sie viel früher in Verwendung waren als
angenommen und das lange vor der Ankunft des anatomisch modernen
Menschen in Europa (z.B.: Projektil in 3. Halswirbel von einem Esel (Equus
africanus): Boëda et al. 1999; „Pointes d’Angé“ von vor ca. 80.000 Jahren:
Soressi/Locht 2010). Sie wurden vorne an Speer- und Lanzenenden geschäftet,
welche zuvor lediglich angespitzt und in Feuer gehärtet wurden. Viele der
Projektile wurden aus Silex (Feuerstein) hergestellt, da das Material hart ist,
sich aber auch gut spalten lässt. Es wurden aber auch welche aus Knochen
oder Geweih gefertigt. Jedes Material hat seine Vor- und Nachteile. Während
Knochen und Geweih sich als besonders resistent bei Impacten auf Knochen
herausstellen, haben sie im Vergleich zu Steinspitzen weniger Schneidkraft
beim Eindringen in den Körper. Bei Steinspitzen verhält es sich genau
umgekehrt (Stodiek/Paulsen 1996). Neben Speerspitzen werden, wie Beispiele
aus Südwest Frankreich zeigen, auch andere Werkzeuge verwendet. In MTA
Ensembles wurden verschiedene Schlacht- bzw. Zerlegungsgeräte erkannt. Zu
39 Tab.4: Materialien (1-8) gruppiert nach Erhaltungschancen

38
jenen gehören bifacielle MTA Faustkeile, bifacielle Abschläge, die entweder als
Abschlag verwendet oder zu leichten Schabern umfunktioniert wurden
(Delagnes/Renu 2011).
1994 wurden im Braunkohlewerk in Schöningen sieben im Durchschnitt über
zwei Meter lange Speere ausgegraben. Sie sind eine wahre Sensation, denn
sie stammen aus der Zeit des Homo heidelbergensis vor 300.000 Jahren und
sind somit die ältesten erhaltenen Holzwaffen. Auf diesem Jagdlagerplatz
wurden noch andere interessante Holzobjekte ausgegraben –unter den Funden
befanden sich weitere Speerfragmente, ein Wurfholz und eine Lanze–
(http://www.palaeon.de/schoeninger-speere.html). Die Speere sind sehr
sorgfältig gearbeitet, haben eine ergonomische Form und sind für eine optimale
Flugbahn austariert (Soressi/Locht 2010). Diese Funde lassen erkennen, dass
Homo heidelbergensis vorausschauend arbeiten konnte sowie diverse
technische Fähigkeiten beherrschte. Zudem zeigt es, dass er in einem mehr
oder weniger komplexen sozialen Gefüge lebte und unterschiedlichen
Jagdstrategien mächtig war. Bis Dato wurden keine vergleichbaren Funde
entdeckt. Vermutungen zum Jagdwaffenspektrum bei Neandertalern müssen
dementsprechend von diesen Funden abgeleitet werden.
6.3.3 Nahrungsspektrum
Aus zahlreichen Studien ging immer wieder hervor, dass Homo
neanderthalensis zu den Topprädatoren ihrer Zeit gehörten und ihre Ernährung
vorwiegend aus Fleisch bestand. Daher tendierte man lange dazu, das
Verhalten von Neandertalern zu homogenisieren. Mittlerweile ist allgemein
anerkannt, dass dessen Subsistenzverhalten variabler und komplexer war als
zuvor angenommen (Burke 2000). Die Ernährung der verschiedenen Gruppen
war wahrscheinlich durch eine Vielzahl an unterschiedlichen Diäten gegliedert,
die dennoch Regional abhängig waren. So stehen neben mittelgroßen (Ibex,
Chamois, Gämse, Rentier) und großen Säugetieren (Pferde, Rinderartige,
Wollnashorn, Wollhaarmammut) auch kleine Tiere wie Hasen und Schildkröten
sowie Meerestiere im Fokus (Stringer et al. 2008; Hardy/Moncel 2011;
Yravedra/Cobo-Sánches 2015). Pflanzen spielen ebenfalls eine wichtige Rolle
in der Ernährung. Seit den letzten 20 Jahren beschäftigen sich vier

39
unterschiedliche Disziplinen mit dem Subsistenzverhalten der Neandertaler. Die
Zooarchäologie, die Paläobotanik, die Osteologie und die Isotopenanalyse.
Jede für sich hat Resultate hervorgebracht, die mit anderen übereinstimmen,
andere ergänzen, oder sogar das Gegenteil von dem ergeben, was als
rekonstruiert galt (Ready 2010).
Die Zooarchäologie wird seit der Mitte des 20e Jahrhunderts verwendet. Sie
beschäftigt sich mit den faunischen Überresten (Knochen und Zähne) aus
Fundkomplexen und ermöglicht somit Einblicke in soziales Verhalten
(Gaudzinski-Windheuser/Kindler 2012). Aus diesen Resten lässt sich ablesen,
ob Spuren vom Menschen oder Karnivoren vorhanden sind. Je nach Größe der
Knochen oder Zähne können Genus, Geschlecht und Alter ermittelt werden.
Quantifizierungsanalysen geben Auskunft über die Anzahl von eventuell
gejagten Tieren sowie über die bevorzugt mitgebrachten bzw. verwerteten
Körperteile. Je nach Periode, Saison und Gebiet variierte das Jagbeuteschema
deutlich (Bar-Yosef 2004).
Da Pflanzen die Zeit nur sehr schlecht überdauern, wurden sie als
Ernährungsbestandteil der Neandertalerdiät allgemein unterschätzt. Hinweise
gibt es dennoch aus dem Nahen Osten. Aus Moustérien Schichten der
Gorham’s Höhle (Gibraltar) und der Kebara Höhle (Israel) sind verkohlte Samen
bekannt (s.d: Barton et al. 1999; Lev et al. 2005). Weitere Spuren wurden auf
Steinartefakten entdeckt, allerdings konnte selten bis gar nicht herausgefunden
werden um welchen Pflanzentyp es sich handelt.
Osteologische Analysen an Knochen und Zähnen fördern interessante
Ergebnisse zu Tage und geben Anlass zu weiteren Interpretationen.
Untersuchungen an Neandertalerskeletten belegen, dass diese, im Gegensatz
zu anderen archaischen Menschen oder heute lebenden Kulturgruppen, eine
ungewöhnlich hohe Rate an Verletzungen vorweisen. Besonders der Kopf und
die Nackenpartie waren betroffen. Vergleichbare Verletzungsmuster wurden
lediglich bei Rodeoreitern identifiziert. Man folgerte aus diesen Erkenntnissen,
dass diese Verletzungen das Resultat von Jagdaktivitäten sein müssen, bei
denen die Neandertaler nahen Kontakt mit mittelgroßen bis großen Säugetieren
hatten (Berger/Trinkaus 1995; Ready 2010). Weitere Untersuchungen
konzentrierten sich auf die Zähne von Neandertalern, welche mit Inuit Gebissen
verglichen wurden. Jene Gegenüberstellung ergab, dass die Ernährung im

40
Gegensatz zu der der Inuit, deren Ernährung fast ausschließlich auf Fleisch
basierte und die Zähne des Neandertalers wesentlich größere
Abnutzungsspuren aufweisen. Dies könnte darauf hindeuten, dass jene
ebenfalls als Werkzeuge genutzt wurden, aber auch, dass der Anteil an
abrasiven Nahrungsmittel wesentlich höher war als angenommen
(Spencer/Demes 1993; Ready 2010). Die vorgenommenen Analysen
begrenzen sich jedoch nicht nur auf Abnutzungspuren der Zähne. Henry et al.
(2014) stellten in einem Artikel im Journal of Human Evolution die Resultate
ihrer Untersuchungen des Zahnsteins vor. Sie entnahmen 209 Proben von 30
verschiedenen Völkergruppen (Neandertaler und anatomisch moderne
Menschen inbegriffen) aus unterschiedlichen Regionen der Welt (Naher Osten,
Europa und Afrika aus einem Zeitraum von ca. 130-10 ka). Untersucht wurden
im Zahnstein eingelagerte Mikroreste, um ein Ernährungsschema zu erstellen.
Die Spy-Proben wiesen eine große Menge an stärkehaltigen Speiseresten auf,
die der Wasserlilien Familie (Nympheae) und Gräsern zugeordnet werden
konnten. In Zahnsteinproben von Neandertalern aus anderen europäischen
Fundplätzen wurde zwar eine geringere Menge an Pflanzenresten
(Speisestärke von Gräsern Triticeae) festgestellt, dennoch schien der Konsum
hoch genug gewesen zu sein, um sich abzulagern. Proben aus dem Nahen
Osten beinhalteten ebenfalls Reste von Gräsern. Die meisten Proben aus
Europa verfügen im Durchschnitt fünf verschiedene Sorten an Mikroresten von
Pflanzen. Die Proben aus Spy und Shanidar tragen hingegen fast 50 (Henry et
al. 2014; Fiorenza et al. 2015). Dies lässt darauf schließen, dass pflanzliche
Nahrungsmittel wahrscheinlich eine weitaus größere Rolle spielten als bisher
angenommen und Neandertaler vielleicht im gleichen Umfang pflanzliche
Fasern zu sich nahmen wie der anatomisch moderne Mensch.
Seit Anfang der 1990e werden Stabilisotopenanalysen an Neandertalern aus
dem Zeitraum zwischen 120 und 30 ka durchgeführt. Diese Methode ist eine
weitere Möglichkeit einen besseren Einblick in das Nahrungsspektrum zu
bekommen, allerdings hat sie wie jede andere Methode auch ihre Nachteile.
Kollagen ist ein tierisches Protein, welches am Aufbau von Knochen und
Zähnen beteiligt ist. Stirbt ein Individuum, baut sich dieses Protein stetig ab.
Möchte man Analysen durchführen, empfiehlt es sich daher Proben von
Knochen zu entnehmen, die kälteren Konditionen ausgesetzt waren. Somit

41
erhöht sich die Chance genügend Kollagen zu extrahieren. Bei bisher
durchgeführten Untersuchungen (s.d: Bocherens et al. 1999, 2001, 2005; Smith
et al. 1999; Richards et al. 2000, 2008; Higham et al. 2006; Richards/Schmitz
2008) lag die Konzentration vor allem auf Neandertalern aus kalten
klimatischen Zyklen (Fiorenza et al. 2015). Die Resultate ergaben generell
ergeben, dass tierische Proteine –meistens von mittelgroßen bis großen
Landsäugetieren– verspeist wurden. Informationen über Meerestiere oder
Pflanzen gingen aus diesen Untersuchungen nicht hervor (Richards/Trinkaus
2009; Fiorenza et al. 2015). Es sollte dennoch nicht von einer einseitigen
Ernährung ausgegangen werden. Vielmehr dient diese Methode als ein
weiteres Hilfsmittel und zur Ergänzung anderer Methoden. Eine
Weiterentwicklung der Technik kann in Zukunft evtl. neue Informationen über
das Nahrungsspektrum geben.
7 Der anatomisch moderne Mensch
Der anatomisch moderne Mensch entwickelte sich, anders als der
Neandertaler, nicht auf dem europäischen Kontinent. In den letzten
Jahrzehnten wurde heftig über seine Herkunft debattiert, sodass verschiedene
mehr oder weniger plausible Modelle vorgestellt wurden. Die beiden
bekanntesten Thesen sind die Out-of-Africa Theorie und die multiregional
Theorie (Multiregionaler Ursprung). Bei der „multiregional“ Theorie wird davon
ausgegangen, dass sich vor 1.7 Ma Gruppen der Gattung Homo erectus in
verschiedenen Regionen der Welt ansiedelten und sich der Homo sapiens aus
jenen parallel entwickelte. Dies begründet sich auf dem Austausch von
genetischem Material trotz großer Distanzen. Somit sei verhindert worden, dass
sich neben dem Homo sapiens noch weitere Gattungen entwickelt hätten
(Thorne/Wolpoff 2003). Die Unterschiede zwischen den einzelnen heute
lebenden Menschengruppen basieren dementsprechend auf der
multiregionalen Entwicklung (Raghavan et al. 2009). Die „Out-of-Africa“ Theorie
basiert hingegen darauf, dass sich der anatomisch moderne Mensch aus den in
Afrika gebliebenen Homo erectus weiterentwickelte und es somit einen
gemeinsamen Ursprung in Afrika gibt (Stringer 2003). Sie gilt als die
plausibelste Theorie. Fossile Funde sowie DNA Analysen weisen auf einen

42
Ursprung vor 150.000 und 200.000 Jahren in Afrika (Äthiopien) hin (Mellars
2004; Raghavan et al. 2009). Der bisher älteste Schädel eines anatomisch
modernen Menschen (Omo I aus der Kibish Formation in Süd Äthiopien) wurde
auf ein Alter von 196 +/- 2 ka datiert (Mcdougall et al. 2005; Trinkau 2005;
Fleagle et al. 2008). Auf anderen Kontinenten wurden aus diesem Zeitraum
keine vergleichbaren Funde gemacht. Aus den meisten Teilen Asiens gibt es
zum Bespiel keine Überreste moderner Menschen, die älter als 40.000 bis
45.000 Jahre BP sind (Mellars 2004b; Trinkaus 2005). Vor 60.000 bis 50.000
Jahren begann der anatomisch moderne Mensch sich von Afrika aus in die
gesamte Welt auszubreiten. Vor ∼45.000-43.000 cal. BP Jahren erreichte er
Europa und breitete sich bis 35.000 cal. BP aus (Mellars 2006), um den dort
lebenden Neandertaler –der neusten Informationen nach bereits um ∼41.000-
39.000 cal. BP europaweit ausgestorben war (Süden der iberischen Halbinsel
bis auf weiteres ausgenommen)– zu ersetzen (Higham et al. 2014). Die
möglichen Ausbreitungsrouten werden in Kapitel 9.3 vorgestellt.
7.1 Verbreitungsraum
Aufgrund der Gegenüberstellung mit dem Neandertaler wird der gleiche
Verbreitungsraum behandelt und wenn möglich sogar der gleiche Fundplatz.
Nachdem der anatomisch moderne Mensch Europa erreichte, breitete er sich
weiter aus bis er jede Nische des Kontinents besiedelte. Diese Ausbreitung
geschah jedoch nicht kontinuierlich und nicht von Anfang an flächendeckend.
Es kommt also vor, dass in Regionen in denen während der TMJ
Übergansindustrien vorhanden waren, keine Gruppen anatomisch moderner
Menschen lebten. Umgedreht konnte dies auch der Fall sein. Des Weiteren ist
zu beobachten, dass im Falle des Aurignaciens nicht allen Fundplätzen jede
Phase (Aurignacien 0,I,II oder III) vertreten war und je nach Region zu
unterschiedlichen Zeitpunkten dort eintraf. In einer Tabelle wurden nach
Angaben von Djindjian (Vgl. 2010: 21-25) die wichtigsten Regionen mit den
aussagekräftigsten Fundplätzen zusammengetragen, um eine
Chronostratigraphie zusammenzustellen40.
40 Tab.5: Übersicht der wichtigsten stratigraphischen Sequenzen des Aurignacien, aufgeteilt in 13 Regionen.

43
7.2 Welche archäologischen Kulturen sind von Bedeutung?
Während der TMJ kommt es im Zeitraum zwischen ∼45.000 bis 35.000 BP
zur Einwanderung von anatomisch modernen Menschen nach Europa. Anders
als bei den Neandertalern, bei denen mehrere Technokomplexe für den
Übergang vom Jung- zum Mittelpaläolithikum bekannt und erforscht sind, fällt
die Auswahl an lithischen Kulturen beim anatomisch modernen Menschen
wesentlich geringfügiger aus. Bis Dato wird eine Übergangsindustrie dem
anatomisch modernen Menschen zugeschrieben. Hierbei handelt es sich um
das Uluzzien, welches in Italien und Griechenland verbreitet ist und bis vor ein
paar Jahren noch den Neandertalern zugewiesen wurde. Des Weiteren wird
das Aurignacien vorgestellt. Diese Kultur gilt als die erste Kultur des
Jungpaläolithikums, welche nach Meinung vieler Forscher ein Paket an
Merkmalen kultureller Modernität mit sich brachte und somit eine neue Ära
einleitete. Wie schon im Kapitel über die Neandertaler (Kapitel 6) werden beide
Kulturen mit ihren Leitformen kurz vorgestellt, indem sie in Raum und Zeit
platziert werden.
7.2.1 Das Uluzzien im südöstlichen Europa (Italien und Griechenland)
Das Uluzzien, als lithische Industrie, wurde in den 1960e von Arturo Palma di
Cesnola entdeckt und das erste Mal 1965/66 vorgestellt (Palma di Cesnola
1965/66). Benannt wurde sie nach der Uluzzo Bucht, in der sich auch der
Typus-Fundplatz –die Grotta del Cavallo– befindet (Palma di Cesnola 1965/66;
d’Errico et al. 2012; Boscato/ Crezzini 2012). Der Technokomplex ist
insbesondere im Süden Italiens, in Kampanien, Apulien und in der Toskana
verbreitet. Im Norden ist er mit einem Fundplatz –der Grotta die Fumane–
vertreten und außerhalb Italiens in Griechenland mit der Klissoura Grotte
(d’Errico et al. 2012; Boscato/ Crezzini 2012)41. Da alle drei Fundplätze in
Kapitel 8 vorgestellt werden, wird hier nicht weiter auf sie eingegangen. In
Zentraleuropa sowie der franko-kantabrischen Region zählt das Uluzzien –
neben dem Châtelperronien und dem nahezu zeitgleichen Szeletien– zu den
drei wichtigsten Übergangsindustrien. Stratigraphisch ist das Uluzzien zwischen
41 Abb.18: Verbreitungsgebiet des Uluzziens

44
dem Ende des Moustériens –von dem es in der Regel (Grotta di Fumane
ausgenommen) durch eine sterile Schicht getrennt ist– und dem beginnenden
Aurignacien einzuordnen (Benazzi et al. 2011; Boscato/ Crezzini 2012).
Nachdem 1964 zwei Molare (Cavallo B und Cavallo C) in der Grotta del
Cavallo ausgegraben wurden, assoziierte man das Uluzzien zunächst mit dem
Neandertaler (Benazzi et al. 2011), wobei Palma di Cesnola davon ausgeht,
dass sich das Uluzzien aus dem Moustérien denticulé der Toskana entwickelt
hat (d’Errico et al. 2012). Benazzi et al. (2011) belegten mit neu durchgeführten
Untersuchungen der Zähne und der Datierung von Muschelresten aus der
Grotta del Cavallo, dass es sich anders als lange angenommen und von vielen
wissenschaftlichen Kollegen (u.a. Palma di Cesnola und Messeri 1967,
Churchill und Smith 2000) akzeptiert nicht um die lithische Industrie von
Neandertalern handelt, sondern um die des anatomisch modernen Menschen.
Mit einem angenommenen Alter von 45.000 bis 43.000 BP gelten die Zähne als
ältester Beweis für die Ausbreitung des Menschen über den europäischen
Kontinent (Benazzi et al. 2011). Diskussionen über eine mögliche Ausdehnung
über eine südliche und somit mediterrane Route wurden mit diesem Beweis
neu entfacht (Moroni et al. 2013). Ebenso deutet, neben den absoluten Daten in
Süditalien, die Art und Weise der Herstellung der Artefakte darauf hin, dass es
sich bei den Schöpfern des Uluzziens um anatomisch moderne Menschen
handelt. Die Beschaffenheit der Artefakte weist größere Gemeinsamkeiten mit
dem darauffolgenden Aurignacien auf als mit dem späten Moustérien (Vgl.:
Benazzi et al. 2011, 527). Aufgrund der Ähnlichkeiten des Uluzzien und des
Châtelperronien wurde wieder die Frage laut, zu welcher Gattung letztere
gehört (Bar-Yosef/Bordes 2010; Higham et al. 2010).
Nachdem das Uluzzien durch Benazzi et al. (2011) wieder mehr in den
Mittelpunkt von Untersuchungen gerückt wurde, starteten Douka et al. (2014)
ein neues Projekt, um diese Kultur chronostratigraphisch genauer
einzugliedern. Dazu wurden 18 Proben (Knochen aus der Fumane Grotte,
Kollagen aus Knochen, Kohle und Muschelproben aus den drei anderen
Grotten) mit der 14C Methode aus Schichten der Grotta del Cavallo, Fumane,
Castelcivita und Klissoura untersucht. Die Datierungen aus der Fumane Grotte
fielen relativ kurz aus. Sie reichten lediglich von 39 ka bis 42 ka BP. In der
Grotta del Cavallo stimmen die ermittelten Daten mit denen aus der

45
Stratigraphie der Grotte überein. Die Proben ergaben Datierungen für einen
Zeitraum zwischen 35 ka bis 40 ka BP. Lediglich für die unterste Schicht der
archaischen Periode enthielt man keine Ergebnisse. Aus der Castelcivita Grotte
wurde ein Stück Kohle doppelt per 14C datiert, welches 50 bis 55 cm unter dem
CI-Tephra in einer Schicht mit identifizierten Feuerstellen lag. Aufgrund der
unterschiedlichen Vorbehandlungsmethoden (ABA und ABOx-SC Methode)
kam es zu einer Diskrepanz in den resultierenden Enddaten. Das Ergebnis mit
der ABA Methode fiel deutlich jünger aus als erwartet, denn dabei wurde ein
Alter von 33 ka BP (∼37,0-38,8 ka cal. BP) festgestellt. Problematisch ist, dass
dieses Datum wesentlich jünger ausfällt als jenes, das im darüber liegenden Kl-
Tephra berechnet wurde, und somit nicht in Frage kommen kann. Die zweite
Messung ergab ein realistischeres Alter von 36.1 ka BP (∼40,5-42,0 ka cal.
BP). Auch hier liegen keine Daten für den Beginn des Uluzziens vor. Die in
Klissoura untersuchte Schicht ist mit darunter und darüber liegenden Schichten
kontaminiert. An der Grenzfläche zwischen den Schichten IV und V wurde eine
hohe Konzentration an KI-Tephra festgestellt. Sie wird als terminus ante quem
für Schicht V angesehen. Untersucht wurden zwei Muschelschalen, wobei eine
auf ein Alter von 34,6 +/- 0,2 ka BP (∼39,9-38,5 ka cal. BP) datiert wird, die
zweite fällt mit einem Alter von ∼28 ka BP wesentlich jünger aus. Die
erhaltenen Datierungen42 lassen darauf schließen, dass das Uluzzien bereits
kurz vor 45 ka cal. BP begann und bis ca. 39 ka cal. BP andauerte (Douka et al.
2014).
Leitformen, Technologie und Typologie
Das Uluzzien ist eine Industrie in der Abschläge dominieren. Hergestellt
wurden vorwiegend Abschläge mit durchschnittlicher Dicke sowie
Klingenabschläge, die zeitweise eine natürliche Rückenretusche aufweisen.
Zusätzlich wurden Klingen und auch eine kleine Anzahl an Lamellen gefertigt.
Zum Inventar gehören aber auch Kratzer, Schaber, gezähnte Stücke und
Stichel sowie andere typische Formen des Uluzziens:
abgesplitterte/zersplitterte Stücke (pièces esquillées) und semilunar 42 Tab.6: Neue Datierungen für den Anfang und das Ende des Uluzziens. Resultate sind in cal. BP widergegeben.

46
rückenretuschierte Artefakte (Douka et al. 2014; Peresani 2014) 43 . Zur
Produktion von Klingen und Lamellen wurden zwei verschiedene
Herstellungstechniken verwendet. Zum einen die bipolare Ambosstechnik (eine
Form der bipolaren Kerntechnik) und zum anderen eine unipolare Bearbeitung
des Nukleus. Trotz dieser unterschiedlichen Methoden sind die hergestellten
Objekte miteinander vergleichbar (Peresani 2014).
Während des Uluzziens gehörten Artefakte aus Knochen und Bein zum
normalen Gerätespektrum. Vorwiegend wurden Stichel mit einer abgeflachten
Basis produziert. Daneben entstanden auch zylindrisch geformte Spitzen mit
konisch abgeschrägten Kanten und Fragmente zylindrischer Objekte wurden
ebenso gefunden (d’Errico et al. 2012). In Castelcivita wurde neben den
üblichen Modellen eine Doppelspitze zu Tage gebracht. Peresani (2014) nimmt
an, es könne sich dabei aus funktioneller Sicht um einen Angelhaken handeln.
An einigen Stücken konnte man noch erkennen von welchen Tieren sie
stammten. Pferde- und Hirschknochen wurden bevorzugt verarbeitet. Zudem
wurde festgestellt, dass mindestens drei verschiedene Techniken Anwendung
fanden, um die Diaphysen von langen Knochen zu bearbeiten (d’Errico et al.
2012):
1. Die Enden von natürlich spitzen Knochen werden mittels schaben
geringfügig modifiziert.
2. Die Epiphysen von langen Knochen werden mittels schaben
modifiziert.
3. Die dünnen Fragmente von langen Knochen werden mittels schaben
in Form gebracht bzw. modelliert.
7.2.2 Das Proto-Aurignacien/Aurignacien in West-. Mittel- Ost- und
Südeuropa
Das Aurignacien gilt innerhalb der Wissenschaft als die erste Kultur des
Jungpaläolithikums, welche in Verbindung mit der Ausbreitung des anatomisch
modernen Menschen steht und somit wesentliche Neuerungen in sozialen,
ökologischen und demographischen Verhaltensmustern mit sich brachte
(Mellars 1973, 1996, 2004a; Gamble 1999; Bar-Yosef 2002). Diese
Innovationen verursachten einen Bruch mit dem Mittelpaläolithikum und 43 Taf.8: Leitformen des Uluzziens

47
läuteten eine neue Periode ein. Erste Artefakte dieser lithischen Kultur wurden
1860 von Edouard Lartet in der Grotte von Aurignac entdeckt (Bouyssonie
1954). Seit ihrem Fund wurden viele Fragen aufgeworfen, sodass das
Aurignacien über das letzte Jahrhundert Mittelpunkt vieler Debatten war. In den
insgesamt vier großen Diskussionen wurden unterschiedliche Aspekte des
Themas allumfassend betrachtet.
Die erste Debatte fand zwischen Gabriel de Mortillet, dessen Sohn Adrien de
Mortillet (nach dem Tod seines Vaters) und Henri Breuil statt. Es ging um die
Frage der Eigenständigkeit bzw. der Existenz des Aurignaciens und seiner
stratigraphischen Eingliederung nach dem Moustérien. Breuil sah in dieser
Kultur die Sukzession zum Mittelpaläolithikum und gliederte sie in das
Jungpaläolithikum ein. 1912 erreichte diese Debatte ihren Höhepunkt indem
Breuil eine eindrucksvolle Rede auf dem C.I.A.A.P. (Congès International
d’Anthropologie et d’Archéologie Préhistoriques) in Genf über
Chronostratigraphie und die Sukzession von Kulturen hielt (Davies 2009;
Djindjian 2010; s. zusätzliche Medien N°10 ❲Pigeaud 2006❳).
Die zweite erstreckte sich über einen wesentlich längeren Zeitraum von 1950
bis 1994. 1933 stellte Denis Peyrony das Modell des Perigordien in Aquitanien
vor. Dieses beinhaltete, dass an Stelle von nur einer Kultur, sich in Aquitanien
zwei parallel voneinander entwickelten, das Aurignacien (I-V) und das
Perigordien (I-V). 1994 leitet das Ende dieser Debatte ein, indem das
Perigordien und das Aurignacien V als nicht existent eingestuft werden
(Djindjian 2010).
1966 stellte George Laplace das Modell „le synthétotype“ vor und löste
prompt eine neue Debatte aus. Er erkannte europaweit ein weiteres Stadium
des Aurignaciens, nämlich das Proto-Aurignacien, mit Dufour Lamellen in
unterschiedlichsten Inventaren. Lange nicht anerkannt, ist heute gewusst, dass
das Proto-Aurignacien –auch archaisches Aurignacien genannt– in Teilen
Europas existiert hat, wenngleich nicht alle von Laplace vorgeschlagenen
Fundplätze berücksichtigt werden können (Laplace 1966; Djindjian 2010).
Die vierte Debatte hat ihren Anfang in den 1980e. Die französische Schule
der „Châine opératoire“ widersetzte sich dem gängigen Bild der Typologie
sowie der Kulturen des Jungpaläolithikums und zweifelte das Aurignacien in
Frankreich an. Stattdessen schlugen sie zwei regionale parallel verlaufende

48
Fazies, ohne Evolution in der Machart der Artefakte, für Aquitanien und von der
Languedoc Küste bis zum Fundplatz d’Arcy-sur-Cure im Département Yonne,
vor (Djindjian 2010).
Mittlerweile sind für das Aurignacien mehrere Phasen44 anerkannt, die mittels
Chronostratigraphie und anderen Datierungsmethoden (14C Methode) ermittelt
wurden. Da der Technokomplex über ganz Europa verteilt ist45, gibt es eine
Bandbreite an Datierungen (38.000 BP bis 28.000 BP: Djindjian 2010; 40.000
bis 27 uncal. BP : Haesaerts/Pirson 2010). Die Fundplätze aus den Gebieten
Österreich, Mähren und Westkarpaten werden in einen Zeitraum zwischen +/-
39.000 bis 27.500 BP platziert. In Zentraleuropa kommen in diesem besagten
Zeitraum zwei verschiedene Aurignacien Gruppen vor. Zum einen das
Aurignacien ancien (39.000 bis 36.000 BP, Schwallenbach I Interstadial;
Willendorf III, Keilberg-Kirche und Senftenberg) und zum anderen das
klassische Aurignacien (+/- 33.000 bis 27.500 BP, Schwallenbach II Interstatial;
Mitoc-Malus Galben (32.700-27.500 BP), Stratzing (31.700-29.200 BP);
Haesaerts/Pirson 2010).
Das Proto-Aurignacien das Laplace (1966) erstmalig vorstellte, wurde von
Bon et al. (2006) neu untersucht und aufgearbeitet, wobei es in Zeit und Raum
gesetzt wurde. Das archaische Aurignacien –datiert in den Zeitraum von 42,0-
41,2 ka cal. BP (Banks et al. 2013)– ist vor allem im mediterranen Raum
vertreten (Spanien, französische Pyrenäen, im Baskenland, südöstliches
Frankreich und Norditalien). Dennoch wurden in jüngster Zeit einige
Fundstellen in Aquitanien, welches sich im Südwesten Frankreichs befindet, als
auch im südöstlichen Pariser Becken (Grotte du Renne, Schichte VII), im
Donaugebiet (Krems Hundssteig in Österreich), im Banat (Tincova in
Rumänien) und im Balkangebiet (Kozarnika) entdeckt (s.d.: Bar-Yosef 2003;
Otte/Kozłowski 2004; Tsanova; 2006; Teyssandier 2007 ❲Bon et al. 2010❳).
Das klassische Aurignacien (Aurignacien I oder ancien typique) ist die
bekannteste Stufe dieses Technokomplexes. Irrtümlicherweise wurde früher
angenommen, diese Phase sei über ganz Europa verbreitet gewesen.
Mittlerweile weiß man jedoch, dass sie wie das Proto-Aurignacien auf gewisse
Provinzen begrenzt war. Zum Kerngebiert mit den wichtigsten Fundplätzen
gehört der Südwesten Frankreichs (La Ferrassie, La Quina, Le Piage, usw.) mit 44 Tab.6: Phasen des Aurignaciens in Europa 45 Abb.19: Verbreitungsgebiet des Aurignaciens

49
dem Baskenland (Labeko Koba und Gatzzaria) und Kantabrien sowie die
schwäbische Alb (Vogelherd, Geißenklösterle und Hohle Fels). Außerhalb von
diesem Radius ist das Aurignacien ancien typique nur durch einzelne Funde
(Aurignac-Spitze) nachgewiesen und dementsprechend nur spärlich bekannt.
Zeitlich wird diese Phase um 40,0 ka cal. BP bis 39,2 ka cal. BP eingegliedert
(Banks et al. 2013).
Auf das Aurignacien I folgen weitere Stufen, das Aurignacien récent (II) und
das Ende des Aurignaciens (III). Da beide Phasen allerdings jünger als der
vorher festgelegte Zeitraum von ca. 50 ka bis 35 ka BP ausfallen, werden sie in
diesem Rahmen nur kurz vorgestellt. Der Technokomplex ist erstmals
flächendeckend in Europa bis in den Balkan anzutreffen (Djindjian 2010; Bon et
al. 2010). Das Aurignacien récent beginnt ca. 32.000 BP und geht 31.000 BP
(nach Banks et al. 2013: 37.0-36.5 ka cal. BP) in die Endphase über. Ab 29.500
bis 29.000 BP sind immer weniger Fundplätze vorzufinden. Das Aurignacien
befindet sich zu diesem Zeitpunkt im Übergang zum Gravettien –welches seine
Anfänge je nach Region bereits um 29.300 +/- 750 BP hat (Kozłowski 2015)–
und verschwindet schlussendlich vor +/- 27.000 uncal. BP (Bon et al. 2010).
Die Frage zum Ursprung des Aurignaciens wird seit Jahren kontrovers
diskutiert. Dabei wurden verschiedene Modelle vorgestellt. In einem Modell wird
von manchen Forschern die Meinung vertreten, dass das Aurignacien eine
homogene europaweite Kultur mit einem gemeinsamen Ursprung außerhalb
Europas (Balkan) ist, dennoch herrscht teilweise Uneinigkeit in Bezug auf die
Chronologie der Anfänge, wodurch weitere Fragen aufgeworfen werden (s.d.:
Mellars 1989, 1999, 2004b; Kozłowski 1993; Otte 1996; Zilhão und d’Errico
1999; Kozłowski und Otte 2000 ❲Teyssandier 2005❳). In einem anderen Modell
wird das Aurignacien als eine polyzentrische Kultur mit unterschiedlichen
Ursprungsherden angesehen (s.d.: Clark und Lindly 1989; Valoch 1990; Straus
1996; Karavanić und Smith 2000; Cabrera et al. 2001). Die verschiedenen
Phasen und Regionen (Proto-Aurignacien, Aurignacien I-III ❲ancien typique,
récent, final❳) werden als Beweis angeführt. In einem weiteren Modell
wiederum wird die Meinung vertreten, dass zwar ein gemeinsamer Ursprung
außerhalb von Europa liegt, der Balkan wird allerdings als Region abgelehnt
(s.d.: Conard/Bolus 2003; Bolus 2004). Weiter wird davon ausgegangen, dass
das Aurignacien „nicht von Anfang an das komplette Paket an Merkmalen

50
kultureller Modernität“ (Teyssandier 2005: 12) mit sich führte. Diese
entwickelten sich lediglich im Laufe der Zeit bis zum klassischen Aurignacien
(Teyssandier 2005).
Leitformen, Technologie und Typologie
Mit der neuen Gattung des Menschen, die sich ihren Weg nach Europa
gebahnt hat, kommt auch eine neuartige Steinkultur. Es werden ganz neue
Techniken der Verarbeitung angewandt, sodass innovative und zugleich
grazilere Artefakte entstehen. Die Pluralität des Mittelpaläolithikums weicht
einem homogenen Konzept, das sich auf ein paar wenige, aber
hochspezialisierte Steinartefaktformen reduziert. Klingen und Lamellen gehören
zu den Grundprodukten und werden systematisch hergestellt. Geräte aus
Geweih, Knochen und Elfenbein (Aurignacien-Spitze) erleben einen
Aufschwung und sind in vielfältigen sowie komplexen Abwandlungen vertreten
(Mellars 2005). Sie sind oftmals mit verschiedenen Motiven verziert.
Kunstäußerungen treten ab diesem Zeitpunkt in verschiedensten Formen auf,
dazu gehören Schmuckstücke aus Muscheln und durchlöcherten Zähnen
(Ketten), Flöten aus Knochen oder Elfenbein, als auch Höhlenmalereien (Liolios
2010; Floss/Conard 2010; Delluc 2010; Vanhaeren 2010).
Im Proto-Aurignacien dominieren große Lamellen (z.B.: Font-Yves-Lamelle),
welche systematisch hergestellt werden. Gradlinige oder leicht bogenförmige,
schlanke und wechselseitig retuschierte Formen werden bevorzugt (Dufour
Lamelle) (Le Brun-Ricalens 2013). Diese sind in der Regel zwischen 20 und 70
mm lang und 12 mm breit (Fiedler et al. 2011). Im Osten findet man auch eine
noch schlankere Version, die Krems-Spitze. Sie ist beidkantig oder partiell
beidkantig, marginal zur Spitze hin retuschiert. Die meisten sind beidkantig
dorsal zugerichtet, einige davon wurden wechselseitig gearbeitet und in
einzelnen Fällen findet man auch ventral bearbeitete Stücke
(Teyssandier/Bordes 2010; Fiedler et al. 2011). Zusätzlich wurden leichte
Klingen für die Produktion von Stichel, Schaber und retuschierten Klingen
hergestellt. Größtenteils wurden unipolare, gerne aber auch pyramidenförmige
Kerne verwendet. Die Lamellen wurden mittels Schlaggeräten aus tierischen
Materialien mit einer direkten Schlagtechnik vom Kern abgetragen. In manchen

51
Inventaren wurden Lamellen entdeckt, die aus kernförmigen Sticheln oder
Kielkratzern gefertigt wurden (Teyssandier/Bordes 2010)46.
Im Aurignacien ancien typique wurden vor allem kleine axiale Dufour
Lamellen –nach Demars und Laurent (1989) ein „sous-type Dufour“– mit einem
gradlinigen oder bogenförmigen Profil geschaffen. Sie wurden aus Kern-
Kielkratzern bevorzugt mit breiter Stirn oder Kern-Nasenkratzern abgebaut (Le
Brun-Ricalens 2013). Es wird zu einer Microlithisation tendiert. Die Lamellen
sind zwischen 12 und 25 mm lang, 3 bis 6 mm breit und 1 bis 2 mm dick47.
Das Aurignacien récent wird wieder durch einen anderen Typ Lamellen
gekennzeichnet. Aus Kern-Kielstichel, Kern-Bogenstichel oder Kern-
Nasenkratzer mit meistens einer schmalen Kratzerstirn wurden Dufour
Lamellen vom Typus sous-type Roc-de-Combe hergestellt (Le Brun-Ricalens
2013)48. Sie sind durchschnittlich zwischen 8 und 20 mm lang (Fiedler et al.
2011). Im Aurignacien final waren es große, axiale und leicht tordierte Lamellen
mit direkter Retusche49. Seltener sind sie ventral oder alternierend. Abgebaut
wurden sie von Kern-Flachstichel oder Vachons-Stichel (Le Brun-Ricalens
2013).
7.3 Subsistenzverhalten
Das Subsistenzverhalten des anatomisch modernen Menschen spielt eine
ebenso wichtige Rolle wie bei Neandertalern und wird seit Jahrzehnten intensiv
untersucht. Vor allem Disziplinen wie die Evolutionäre Anthropologie,
Archäologie und in den letzten Jahren sogar vermehrt die Humanmedizin
beschäftigen sich mit den unterschiedlichen Aspekten. Subsistenzstrategien
haben sich im Laufe der Entwicklungsgeschichte des Menschen stetig
geändert, sei es wegen den Unterschieden zwischen den Gruppen oder den
klimatischen Begebenheiten (Discamps et al. 2011). Auch die Umwelt ist
zeitlichen Veränderungen unterworfen (Kuhn/Stiner 2001), sodass Strategien
dementsprechend angepasst werden müssen.
46 Taf.9: Leitformen des Proto-Aurignaciens

52
7.3.1 Lebensweise
Trotz der teils großen Unterschiede, die zwischen und innerhalb von
Menscharten im Jungpleistozän herrschten, lassen sich dennoch
Gemeinsamkeiten in der Lebensweise finden. Wie schon der Neandertaler
waren die anatomisch modernen Menschen Jäger sowie Sammler und führten
einen nomadischen, eventuell auch seminomadischen Lebensstil. An den
zahlreichen Stratigraphien der europäischen Fundplätze lässt sich ablesen,
dass die gleichen Stellen über mehrere Jahrtausende aufgesucht und bewohnt
wurden. Runde Behausungen aus vergänglichen Materialen (Holz, Knochen
und Tierhäuten) erhielten sich vor allem aus dem Gravettien des mittleren
Jungpaläolithikums erhalten. Zooarchäologische Analysen von Vercoutère und
Patou-Mathis (2010) aus Frankreich, dem Mediterranen Raum, Nord-, Zentral-
und Osteuropa zeigen, dass im Aurignacien Tiere nicht nur zum Verzehr gejagt
wurden. Vor allem Wild (Ren, Pferd und Hirsch) wurde generell
ausgeschlachtet und zu unterschiedlichen Utensilien weiter verarbeitet.
Besonders beliebt waren dabei Elemente wie Jagdwaffen, oder aber auch
Schmuck. Weiter ging aus diesen Analysen hervor, dass primär in der
Mediterranen Region, Nordfrankeich und Europa Langzeitcamps aufgebaut
wurden. In Kroatien, Slowenien, der Krim und Russland (Kostenki I) treten
hingegen vorwiegend Kurzzeitcamps auf, die als saisonale Behausungen
angesehen werden können, oder als Jagdstationen (Vercoutère/Patou-Mathis
2010). Es also kann davon ausgegangen werden, dass bereits frühere
anatomisch moderne Menschen ihre Behausungen, je nach Mobilitätsmodell
(s.: Kapitel 6.3.1), mit sich führten und an strategisch ausgewählten Punkten
wieder aufbauten. Grotten sind im Gegensatz zu Freilandbehausungen (z.B.:
Kostenki I, Barca II, Stratzing) eher als Langzeitcamps (z.B.: Fumane,
Vogelherd, Geißenklösterle, l’Abri Pataud Schicht 7) geläufig, von denen aus
die Gruppenmitglieder ausscherten, um Ressourcen heranzuschaffen. In der
Regel wurden relativ kleine Territorien ausgebeutet, die in einen Umkreis von
∼20 km lagen. Nichtsdestotrotz konnten sich diese auch auf über 70 km
ausdehnen. Einige der Behausungen dienten entweder als Sommer- oder
Winterquartier (z.B: Vogelherd und die Grotte du Renne d’Arcy-sur-Cure
wurden als Winterquartier genutzt; Fumane eher im Frühling und Spätsommer;

53
Geißenklösterle im Herbst) genutzt. Auch der anatomisch moderne Mensch
organisierte die Fläche in seinen Behausungen nach einem bestimmten Muster
(Noiret 2010).
Die Kunst spielte auch eine wichtige Rolle im Leben der frühen Menschen.
Zahlreiche Höhlenmalereien, Schnitzereien und Gravierungen zeugen von den
Eindrücken, die von der Umgebung gewonnen wurden. Kleidung und Haare
wurden mit teilweise zeitaufwändigen Schmuckstücken aus Muscheln, Bein
oder Knochen verziert. Lange wurde diese Eigenart nur dem Menschen
zugeschrieben. In der Gorham Grotte auf Gibraltar wurde kürzlich eine
geometrische Gravur eingeritzt in den Felsen aus ungestörten
Moustérienschichten, die älter als 39.000 cal. BP ausfallen, entdeckt
(Rodríguez-Vidal et al. 2014) und in Krapina zu Schmuck umfunktionierte
130.000 Jahre Seeadlerkrallen (Radovčić et al. 2015). Die Gravur und die
Kralen geben einmal mehr Anlass dazu die kognitiven Fähigkeiten (Gunz et al.
2010) des Neandertalers zu überdenken und in ein neues Licht zu rücken.
Zumal dieser seine Toten beerdigte und somit eine Form des rituellen Denkens
beherrschte, scheint es nicht abwegig zu sein, dass bis zu einem gewissen
Niveau auch Kunst eine Rolle spielte.
7.3.2 Jagdwaffenspektrum
Das Jagdwaffenspektrum unterschied sich im Wesentlichen nur in der
Hinsicht des Typs zum Mittelpaläolithikum. Die Gerätschaften wurden aus den
gleichen Materialien gefertigt und blieben demnach nur in Ausnahmenfällen,
wenn es die Beschaffenheit des Bodens (Sumpfgebiet, sehr trockener Boden,
Permafrost) zuließ, in Form von Fragmenten oder sehr vereinzelt sogar
komplett erhalten. (s.: Kapitel 6.3.3). Zu 90% bestanden die Waffen aus Holz
(Cattelain 2010), die mit anderen Elementen (die restlichen 10%) –sei es aus
Knochen, Geweih oder Elfenbein– kombiniert wurden, um eine besser
Durchschlagskraft oder Reichweite zu erlangen. Während des Übergangs vom
Mittel- zu Jungpaläolithikum entwickelten sich in unterschiedlichen Regionen
neue Kulturen. Zum mittelpaläolithischen Inventar treten neue Formen auf, die
einerseits noch vor dem Erscheinen des anatomisch modernen Mensch
erfunden und verwendet wurden, und andererseits von dieser „neuen“ Gattung
mit nach Europa gebracht wurden. Ab dem Jungpaläolithikum wird ein ganz

54
neuer Typ an Geräten und Waffen hergestellt. Lamellen und Klingen gehören
zu den Grundprodukten aus denen andere Produkte (Spitzen, Messer,
Kratzer/Schaber usw.) konzipiert werden. Diese werden für die Bewaffnung, für
Schlacht- und Zerlegungsvorgänge sowie den häuslichen Gebrauch (Beispiel
Grotte von Isturitz) verwendet (Normand et al. 2008). Waffenspitzen aus
tierischen Materialien sind aus der Phase des Proto-Aurignaciens nur spärlich
vorhanden, dafür sind sie aus früheren Perioden wesentlich bekannter und
gelten als Leitformen des klassischen und späten Aurignaciens (Cattelain
2010). Die unterschiedlichen Formen an Spitzen Knochen und Geweih haben
dazu veranlasst Hypothesen zur Schäftungsweise zu erstellen. Nach
umfassenden Experimenten in den 1990e stellte Knecht (2000) einige
Methoden vor50.
Welche Waffen wurden in der ersten Hälfte des Jungpaläolithikums
verwendet? Der Stand der Forschung erlaubt es nur Hypothesen zu erstellen,
da es bis Dato keine Funde gibt, die erahnen lassen, wie breit gefächert das
Spektrum war. In Isturitz wurde zum Beispiel das Fragment von einer
Hirschgeweih-Spitze gefunden. Dieses Stück zeigt, dass die Schäfte der
Waffen aus einem vergänglichen Material (Holz) gefertigt wurden. Ob es nun
ursprünglich Lanzen oder Speere waren, lässt sich nur vermuten. Mit Hilfe der
Experimentellen Archäologie wurde das Flugverhalten von Spitzen aus Stein
und aus anderen Materialien aus der ersten Hälfte des Jungpaläolithikums an
Pfeilen getestet. Das Resultat hat gezeigt, dass bei Vielen ein gutes
Flugverhalten erzielt werden kann. Aufgrund dieser Ergebnisse kam der
Vorschlag, das Erscheinen von Pfeil und Bogen evtl. auch der Speerschleuder
bereits an den Anfang des Jungpaläolithikums zu verlegen (Cattelain 2006).
7.3.3 Nahrungsspektrum
Die Ernährungsweise des Homo sapiens wurde im gleichen Umfang, wie die
des Neandertalers, erforscht. Jedoch ergaben anfängliche Resultate ein
gänzlich anderes Bild. Aus den Analysen ging hervor, dass sich das
Ernährungsbild seit der mittleren Steinzeit ständig veränderte und immer mehr
unterschiedliche Ressourcen herangezogen wurden. Weiterentwickelte
50 Abb.20: Potenzielle Schäftungsweisen im Jungpaläolithikum

55
Jagdstrategien und Waffen haben dabei weitergeholfen, sodass nach Ansicht
einiger Wissenschaftler ein Wettbewerb um die Ressourcen ausgelöst wurde,
dem der Neandertaler unterlegen war und auf lange Dauer zu seinem
Aussterben führte (Flores 1998; Banks et al. 2008; Fa et al. 2013; Henry et al.
2014; Benazzi et al. 2015). Isotopenanalysen zufolge wurde dem anatomisch
modernen Mensch ein wesentlich größeres Nahrungsspektrum zugesagt als
dem Neandertaler (Drucker/Bocherens 2004; Richards/Trinkaus 2009). Diese
Methode zeigt aber lediglich den Anteil an Proteinen die konsumiert wurden,
Pflanzen bleiben außen vor. Marine Tiere (Fisch, Molluske, Schildkröten;
Marean et al. 2007; O’Connor et al. 2011) und kleine Tiere (Hasen, Vögel,
Fuchs usw.; Conard et al. 2013) standen genauso auf dem Speiseplan wie
mittelgroße und große Landsäugetiere (Pferde, Hirsch und Reh, Ren, Gämse,
Rhino, Mammut usw.; Conard et al. 2013). Je nach Region und Zeitraum gab
es unterschiedliche Präferenzen. Da Pflanzen sich über die Zeit nur schlecht
bis gar nicht erhalten, konnte lange nicht nachvollzogen werden welche Arten
konsumiert wurden und in welchem Umfang. Henry et al. (2014) analysierten,
wie schon weiter oben vorgestellt, 209 Zahnsteinproben von Neandertalern und
anatomisch modernen Menschen. In diesen Proben, wie auch auf Werkzeugen,
waren unter anderem stärkehaltige Mikroreste von Gräsern (Triticeae),
Schwertliliengewächse (Iridaceae) und Seerosengewächse (Nymphaceae) zu
finden. Aus den Ergebnissen unterschiedlicher Projekte geht die Kenntnis
hervor, dass das Nahrungsspektrum beider Arten sich wesentlich ähnlicher war
als bisher angenommen, auf geographischer Ebene jedoch sehr unterschiedlich
ausfallen konnte (Fiorenza et al. 2011).
8 Beispielplätze
Folgende Fallbeispiele spiegeln die oben vorgestellten Elemente wieder.
Einige Fundplätze wurden umfassend erforscht und beschrieben, sodass ein
einigermaßen homogenes Bild der Fauna, der Flora sowie der Geräte bekannt
ist. Chronostratigraphische Marker wie die KI Eruption fördern eine leichtere
Einordung einzelner Level. Die Wahl der Jagdbeute, die Raumorganisation des
Fundplatzes sowie der Stil der Werkzeuge –sofern eine Beschreibung
vorhanden ist– geben Auskunft über den klimatischen Zyklus, die Saison, die

56
Art der Behausung sowie über die persönlichen Präferenzen und angewandten
Technologien innerhalb einer Region und eines Zeitabschnitts. Bei manchen
Fallbeispielen fehlt ein Teil der Beschreibung. Dies ist darauf zurückzuführen,
dass viele Fundstellen zwar ausgegraben, bisher aber nicht vollständig
ausgewertet und publiziert wurden. Dennoch werden sie hier vorgestellt, da sie
wichtige Belege der Forschung innerhalb der TMJ darstellen.
8.1 Abric Romaní de Capellades, Level J (ca. 50 ka)
1909 entdeckte Amador Romaní nahe der Stadt Capellades, die ca. 42 km
Luftlinie von Barcelona entfernt liegt51, einen Fundplatz mit umfangreichen
archäologischen Resten (http://www.iphes.cat/print/node/32). Dabei handelt es
sich um einen Abri (Felsnische/-überhang) auf der Westbank des Anoia
Flusses. Die Felsnische, deren Öffnung in Richtung Nord-Osten zeigt, liegt auf
einer Höhe von ungefähr 265 m.ü.M. in einer karstigen Landschaft, bestehend
aus Travertine Gestein, welches in der Umgebung Cinglera del Capelló
genannt wird (Rosell et al. 2012; Vallverdú-Poch et al. 2012).
Bis zum heutigen Tag wurden im Abri vier Kampagnen realisiert. Die erste
(1909-1930) führte Amador Romani, der Entdecker der Felsnische, durch, zwei
weitere standen in den 1950e und 1960e unter der Leitung von Dr. Eduard Ripoll
und die letzte Kampagne erstreckte sich über die Jahre 1983-2009. (Vallverdú-
Poch et al. 2012; http://www.iphes.cat/print/node/32). Bei diesen Ausgrabungen
kam eine Stratigraphie von mehr als 20 m zum Vorschein, die mittels 14C und
Uran-Thorium Datierung in einen Zeitraum von 70 bis 40 ka BP (OIS 4 und 3)
datiert wurde. Mittlerweile wurden weitere Meter der Grotte an der Felswand
des Abric Romaní entdeckt, wodurch die Gesamtstratigraphie auf fast 40 m
Dicke anwächst (http://www.iphes.cat/print/node/32). Insgesamt wurden 16
Schichten freigelegt, die von Level (A) bis (P) reichen (Rosell et al. 2012). Die
einzelnen Ablagerungen der Stratigraphie, die alle bis auf Level (A) zum
Mittelpaläolithikum gehören, sind durch eine sterile Kalktuffschicht voneinander
getrennt (http://www.iphes.cat/print/node/32). Aufgrund der umfangreichen
sowie gut erhaltenen Funde können zahlreiche Rückschlüsse auf das
Subsistenzverhalten der Neandertaler, die über Jahrtausende den Ort
aufsuchten, gezogen werden. Da die ersten fünf Level (A-E) allerdings 51 Abb.21: Geographische Lage des Abric Romaní de Capellades

57
größtenteils in früheren Kampagnen ausgegraben wurden, ist es schwierig jene
genauer zu interpretieren. Speziell die räumliche Verteilung des Fundinventars.
Alle tiefer liegenden Schichten (ab H) wurden zu einem rezenteren Zeitpunkt
ausgehoben und sind somit aussagekräftiger
(http://www.iphes.cat/print/node/32). Aus eben diesem genannten Grund wird
hauptsächlich Level J präsentiert52, welches zentral in der Gesamtstratigraphie
liegt (Rosell et al. 2012). Zum einen wurde diese Fundschicht gründlich
untersucht sowie dokumentiert und zum anderen kann sie an den Anfang des
zu bearbeitenden Zeitraums gesetzt werden. Zum Gesamtinventar des Abris
gehören Kohle von mehr als 200 Feuerstellen, Holzstein (wood
pseudomorphs), faunische Überreste als auch lithische Artefakte.
Innerhalb der Gesamtstratigraphie wurden faunische Überreste von 13
verschiedenen Spezies identifiziert, sechs Herbivoren und sieben Karnivoren.
Pferd (Equss ferus) und Rothirsch (Cervus elaphus) dominieren in allen
Sequenzen. Des Weiteren wurden in einigen Schichten Überreste von Tieren
entdeckt, die in bewaldeten Ökosystemen oder im Flachland leben, darunter
befanden sich Auerochse (Bos primigenius), Nashorn (Stephanorhinus
hemitoechus) und Rüsseltiere. Eine geringe Anzahl an Gämsen aus felsigen
Regionen zählen ebenfalls. Die Überreste der gefundenen Herbivoren wiesen
sowohl anthropogene Spuren als auch Spuren von Karnivoren auf. Die meisten
Karnivoren fanden sich außerhalb des archäologischen Kontextes auf einer
Travertinplattform. Sie deuten darauf hin, dass die Felsniche zumindest
zeitweise als Refugium für diese Spezies diente. Vielleicht wurde diese
Saisonal abwechselnd von Mensch und Tier genutzt. Die Karnivorenknochen,
die im archäologischen Kontext auftauchen, weisen keine anthropogene
Anzeichen auf. Sie gelangten wahrscheinlich ohne menschliches Zutun in den
Abri. Ferner befinden sich in der Grotte Reste von einer Höhlenhyäne (Crocuta
crocuta), Wildkatze (Felis silvestris), Höhlen-Luchs (Lynx sp.), Leopard
(Panthera pardus), Höhlen-Löwe (Panthera leo spelaea), Wolf (Canis lupus)
und Höhlen-Bär (Ursus sp.) (Rosell et al. 2012).
Level J, welches sich in Ja und Jb aufgliedert (beide Sublevel sind durch
eine Schicht Travertine voneinander getrennt; Sañudo et al. 2012), wurde
zwischen 1993 und 1998 ausgegraben und mit Hilfe von zwei
52 Abb.22: Position von Level J innerhalb der Gesamtstratigraphie

58
Datierungsmethoden auf ein Alter zwischen 49.3 +/- 1.6 ka und 50.4 +/- 1.6 ka
(Uranium-Thorium Datierung), beziehungsweise auf 47 ka 14C (AMS) datiert,
womit es sich in die kalte Phase des OIS 3 einfügt. Es wurden 67 Feuerstellen
identifiziert, die sich auf die beiden Sublevel aufteilen –42 auf Level Ja und 25
auf Level Jb–. Die zooarchäologischen Reste lassen sich in zwei Arten von
Kategorien einordnen. Die eine bezieht sich auf die Gewichtsklasse (fünf
Kategorien) des Tieres und die andere auf das Alter in dem es starb. Kategorie
1 entspricht sehr kleine Tieren mit einem Gewicht <20 kg; Kategorie 2
entspricht kleinen Tieren zwischen 20 und 100 kg; Kategorie 3 entspricht
mittelgroßen Tieren zwischen 100 und 300 kg; Kategorie 4 entspricht großen
Tieren zwischen 300 und 1000 kg und Kategorie 5 entspricht sehr großen
Tieren mit einem Gewicht >1000 kg. Vier Kategorien beschreiben hingegen
unterschiedliche Altersstufen der Tiere. Kategorie 1 entspricht der
Säuglingsphase (Milchzähne); Kategorie 2 entspricht der Jungendphase in der
M3 durchbricht; Kategorie 3 entspricht der adulten Phase in der das gesamte
Gebiss ausgeprägt ist und Kategorie 4 entspricht alten Tieren mit stark
abgenutzten Zähnen. In Level J lagen Überreste von 8460 Säugetieren, davon
wurden 6738 aus Level Ja geborgen und nur 1722 aus Level Jb. Da fast alle
Epiphysen fehlten und ein Großteil der Knochen zerbrochen war, konnten nur
knapp 30% identifiziert werden –14,6% (984) in Level Ja und 16,3% (281) in
Level Jb–. Überwiegend befanden sich unter den Überresten Schädelknochen
und Gliedmaßen von mittelgroßen sowie von großen Tieren. Die Knochen
weisen vorwiegend Einschnitte auf, die von einer Häutung oder von der
Entnahme der Eingeweide herrühren (Schädel- und Rippenknochen). Weiter
wurden einige Schnitt- und Kratzspuren mit länglichem Muster identifiziert, die
auf entfleischen hindeuten. Hack- und Zerlegungsspuren tauchen hingegen nur
sehr selten auf. Anhand der Beschaffenheit der Frakturen lässt sich erkennen,
dass die Knochen im frischen Zustand bearbeitet wurden. Die meisten dieser
Knochen lagen im Zentrum der Felsnische oder nahe der Wände bei den
Feuerstellen. Spuren, die durch Karnivoren verursacht wurden, sind in Level J
nur zu knapp 1% enthalten. Hauptvertreter der Knochenfunde ist das Pferd und
der Rothirsch, gefolgt von einigen Resten Auerochse, Nashorn und Gämse. Die
meisten dieser Tiere waren im adulten Alter als sie erlegt wurden, dennoch
lassen sich auch Jungtiere und ältere Tiere finden. Pferde und Auerochse

59
wurden bevorzugt im Frühling und Sommer gejagt, Rothirsch dagegen im
Herbst. Karnivoren lagen nur sehr selten vor (Rosell et al. 2012).
Neben den zooarchäologischen Funden wurden fast 7.000 Steinartefakte
geborgen. Diese verteilten zum größten Teil um die Feuerstellen. Das
Rohmaterial für die Artefakte stammt vorwiegend aus zwei Zonen, die in einem
Umkreis bis 5 km (Quarz und Kalkstein) bzw. in einem Umkreis von 5 bis 25 km
(Feuerstein aus dem Ebro Bassin) lagen. Nur selten machte man sich die Mühe
Material aus mehr als 30 km Entfernung zusammen zutragen. Feuerstein aus
der zweiten Zone wurde hauptsächlich verarbeitet, so bestehen 75% der
Artefakte in Level J aus ihm. Kalkstein und Quarz bestreiten dagegen nur 10%
des Bestandes. Weniger als 3% der Gegenstände/Gerätschaften bestehen aus
zwei weiteren Sorten Stein - Gedankenstrich Porphyr und Quarzit (Vaquero et
al. 2012a). Gestein wurde in fünf verschieden Stadien in den Abri gebracht.
Entweder als ganze oder teile einer Knolle; teilweise bearbeitete Kerne; als
eckige Fragmente; als Abschläge oder retuschierte Artefakte (Vaquero et al.
2012b). Gezähnte Geräte und Geräte mit Kerben fanden sich am häufigsten,
retuschierte Artefakte verkörpern lediglich 3% des Inventars. Spezialisierte
Artefakte wie Schaber sind praktisch gar nicht vorhanden. Zwei
unterschiedliche Strategien lassen sich am Befund ablesen: entweder wurden
aus ganzen oder teilweise bearbeiteten Kernen/Knollen in situ Geräte
hergestellt, oder häufiger vorkommend fertige retuschierte Geräte
(wahrscheinlich zugehörend zur persönlichen Ausrüstung) wurden in die
Felsnische mitgenommen, was häufiger der Fall war. Auch in der Größe
variierten die Artefakte deutlich. Man erkannte durch das Zusammensetzten
gebrochener Kerne, dass jene, die in situ gefertigt wurden wesentlich kleiner
ausfielen als diejenigen, die mitgebracht wurden (Vequero et al. 2012a).
Allgemein waren die Artefakte nur wenig abgenutzt. Dies begründet sich in den
großen Mengen an Rohmaterial innerhalb eines Umkreises von 30km, wodurch
nicht an Ressourcen gespart werden musste.

60
8.2 Cueva del Esquilleu in Kantabrien (spätes Moustérien)
Die Grotte von Esquilleu gehört zu den wichtigsten Fundplätzen für das
späte Moustérien in Kantabrien. Im Gegensatz zu anderen in der Region
(Cillorigo de Liébana) besitzt diese Fundstätte eine nahezu vollständige
chronologische Abfolge (53-30 ka BP) für fast das gesamte MIS 3, als auch die
regionalen H5 bis H3 Ereignisse (Baena et al. 2012). Die Grotte liegt im Westen
Kantabriens, 26 km von der Küste entfernt auf einer Höhe von 350 m ü.d.M. Sie
befindet sich in der „La Hermida“ Schlucht, dessen Eingang nach Süd-Osten
zeigt.
Die Gesamtstratigraphie weist eine Dicke von 4,20 m auf, ist in vier Einheiten
(A,B,C und D) aufgeteilt und besitzt 41 Fundschichten (Uzquiano et al. 2012),
von denen bis Dato 27 Schichten datiert53, wobei lediglich bei zweien (Schicht
III und VI) Ungereimtheiten auftauchten (Baena et al. 2012). Die umfangreiche
lithische Kultur, die generell dem Moustérien zugewiesen werden kann, deutet
auf eine Belegung der Grotte in drei Phasen hin. Die Schichten drei bis fünf
dienten als kurzfristige Besiedlung für Gruppen mit einer diskoiden Kerntechnik;
die Schichten sechs bis vierzehn wurden von Gruppen, die mit der Quina-
Technik vertraut waren belegt, und waren somit dichter als die vorherigen
besiedelt; die dritte und letzte Phase reicht von Schicht fünfzehn bis dreißig. In
jener Phase ist eine Mischung aus mehreren Techniken vertreten, die Levallois-
, die diskoide- als auch sporadisch die Quina-Technik (Baena et al. 2005).
99,2 % des Rohmaterials für die Artefakte entstammt aus der Region. Primär
wurde Quarzit verwendet, sekundär auch Quarz und andere Stoffe von
schlechterer Qualität. Das Flussbett des Devas aus der Gegend liefert Material
für die oberen Schichten (3-6). Die Masse der tiefer liegenden Schichten (7-20)
entnahm man nicht nur aus dem Fluss, sondern sie wurde auch von der 26 km
entfernt liegenden Küste zum Siedlungsplatz transportiert (Baena et al. 2012;
Uzquiano et al. 2012).
Neben den Artefakten befand sich in der Grotte von Esquilleu eine große
Ansammlung an Knochen gefunden. Die Identifikation der einzelnen Taxa
gestaltete sich allerdings als schwierig, da sich das Ensemble aus 71,814 stark
fragmentierten Knochen zusammensetzt. Mehr als 90 % der Gebeine sind
53 Tab.7: Datierung der Fundschichten in der Grotte von Esquilleu

61
kleiner als 2 cm (Baena et al. 2005; Yravedra et al. 2009). Unter den
Fragmenten aus den Taxa, die identifiziert werden konnten, sticht besonders
der Iberiensteinbock (Capra pyrenaica) als dominante Art hervor, gefolgt von
der Pyrenäen-Gämse (Rupicapra pyrenaica) und gelegentlich dem Rothirsch
(Cervus elaphus). In allen Schichten sind komplette Skelette von Steinböcken
vorhanden, in Schicht (11f) auch vom Rothirsch. Von kleinen Huftieren
(Steinbock und Gämse) fand man meist nur Teile des Kopfes, andere Teile sind
dagegen weniger repräsentativ. Der Hirsch ist hauptsächlich durch Hinterläufe
seine Hinterläufe vertreten, gefolgt von seinen Vorderläufen (Yravedra/ Gómez-
Castanedo 2014). Es ist ersichtlich, dass Tiere aus der felsigen Umgebung,
welche um den Abri herum lebten, dominieren (Uzquiano et al 2012). Neben
jenen wurden in einigen Fundschichten Überreste von Karnivoren und anderen
Huftieren wie dem Rind gefunden (Baena et al. 2005). Zwischen den einzelnen
Schichten sind klare Unterschiede zu erkennen. In den oberen Ablagerungen
(3-5) sind kaum Hinweise auf die Präsenz von Neandertaler rar gesät. Knapp
2% der dort gefundenen Knochen weisen Schnitt- und Zerschlagungsspuren
auf, wobei 42 % bis 77 % über Bissspuren von Tieren verfügen. In den darunter
liegenden Schichten (6-13) bergen wesentlich mehr Knochenspuren, die von
Neandertalern herrühren. Bisspuren von Karnivoren treten vereinzelter auf. In
den untersten Schichten (21, 23, 25, 28, 29 und 31) wurden hochfragmentierte,
verbrannte Knochen gefunden (Yravedra/ Gómez-Castenado 2014; Uzquiano
et al. 2012). Da in diesen Ablagerungen keine Kohlereste existieren, wurde die
Vermutung geäußert, dass Knochen als Brennstoff dienten (Baena et al. 2005).
In erste Linie wurde während fast der gesamten Belegung der Grotte Holz –
Kiefer (Pinus), Birke (Betula) und Vogelbeeren (Sorbus)– aus der Umgebung
für Feuer verwendet, wie Funde aus den darüber liegenden Schichten belegen.
Verbrannte Knochen tauchen in diesen hingegen kaum auf. Pollenanalysen aus
der gegebenen Zeit zeigen, dass es kein Feuerholzmangel gab, Uzquiano et al.
(Vgl.: 2012, S. 88) vermuten daher eine systematische Verbrennung von
organischem Material nach dessen Verzehr. In Bezug auf das Alter der Beute
kann gesagt werden, dass bevorzugt adulte Tiere gejagt wurden, dennoch
befanden sich unter der Jagdbeute auch einige Jungtiere und adoleszente
Tiere sowie ein Fuchs (Baena et al. 2005).

62
8.3 Labeko Koba (Châtelperronien und Proto-Aurignacien)
Die Grotte von Labeko Koba –bekannt seit 1971– war eine Fundstelle im
spanischen Baskenland, die sich im Südwesten der Provinz Guipúzcoa im
oberen Teil des Tals vom Fluss Deva befand. Sie lag 28 km von der heutigen
Küste entfernt, auf einer Höhe von 246 m ü.d.M. (Arrizabalaga 2000). Um einer
Zerstörung durch den Bau einer Straße zu entgehen, musste Labeko Koba die
Grotte in einer programmierten Bergungsgrabung, unter der Leitung von
Arrizabalaga A. (Arrizabalaga 2000; Rios-Garaizar et al. 2012), zwischen
September 1987 und Dezember im darauf folgenden Jahr komplett
ausgegraben werden (Arrizabalaga et al. 2002).
Die Morphologie der Grotte war sehr speziell54. Die Öffnung befand sich an
der höchsten Stelle des Hohlraums und war nur dann sichtbar, wenn man sich
ihr auf wenige Meter näherte. Durch eben diese Gegebenheit wurde die Höhle
insbesonders für Herbivoren zur Todesfalle. Im inneren verlief sie stufenartig
bis zum Grund, während die gegenüberliegende Wand einen Überhang bildete
(Rios-Garaizar et al. 2012). Die Gesamtstratigraphie weist mit seinen zehn
Levels eine Dicke von mehr als 4,50 m auf, die in das Jungpaläolithikum datiert
wurden. In den Ablagerungen ist zu erkennen, dass die Grotte von mehreren
Spezies genutzt wurde, wobei die Hyäne und der Mensch eine prädominante
Rolle spielten. Der untere Teil der Höhle besaß die Form eines konusförmigen
Schachtes, in dem sich die ersten Schichten gebildet haben. Die unterste
Schicht 10 ist archäologisch steril. Trotzdem wurden im obersten Teil dieser
Schicht archäozoologische Reste gefunden sowie ein abgebrochenes Stück
von einem Stichel, das mit einem anderen aus Schicht 9, zusammengefügt
werden konnte. Es wurde angenommen, dass Level 10 mit der darüber
liegenden Schicht einher geht und aus der Zeit der Anfänge der Besuche
stammen könnte und archäologisches und so archäologische Material dahin
gelangt sein könnte (Abfallgrube?; Arrizabalaga/Altuna 2000). Schicht 9 welche
in verschiedenen Abschnitten eine Dicke von 2 m aufweist, dominiert fast die
Hälfte des Ensembles. Aufgrund ihres Ausmaßes wurde sie in einen unteren
(inf.) und einen oberen (sup.) Teil gegliedert. Für diese Schicht existieren bis
dato drei Datierungen, wobei der untere Teil auf 34,215 +/- 1,265 BP datiert
54 Abb.23: Gesamtstratigraphie von Labeko Koba

63
wurde und der obere auf 29,750 +/- 740 BP. Neuere Analysen eines
Geweihfragments eines Riesenhirschs (Megaloceros giganteus) ergaben
mittels Ultrafitration jedoch 42,200 +/- 2,400 BP (Rios-Garaizar et al. 2012).
Datierungen mit einer solch extremen Zeitspanne erschweren eine genaue
Prognose über absolute Zeitabläufe von Fundschichten. Wobei in diesem
speziellen Fall eine Datierung um 42, 000 BP plausibel erscheint, da mittels
Ultrafiltration eine Kontamination von Proben minimiert wird (Minami et al.
2012). Die lithische Industrie aus der 9. Schicht setzt sich aus 81 Teilen
zusammen, die sich im unteren Bereich konzentrieren. Ein kleiner Teil dieses
Bereichs wurde in einer früheren Kampagne (1973) aus dem Konus geholt.
Lediglich 12 der 81 Stücke besitzen Spuren von Retusche und gerade einmal
11 sind noch intakt, alle anderen sind fragmentiert. Ein Großteil des Ensembles
zeichnet sich durch 15 mm breite Lamellen mit flachen trapezförmigen
Abschnitten (Bipolarität) aus. Jener Lamellen Typus ist charakteristisch für die
lithische Kultur des Châtelperronien. Somit kann Schicht 9 in die Zeitspanne
des Châtelperronien eingegliedert werden. 85 % der Artefakte sind aus zwei
Sorten Feuerstein gefertigt, die in einem Umkreis von 30 km an verschiedenen
Stellen an der Küste von Bilbao und Bayonne vorkommen sowie in Richtung
Süden an der Wasserscheide (Rios-Garaizar et al. 2012).
Die Fauna der Schicht 10 und 9 setzt sich aus zwei verschiedenen Typen
zusammen. Im konusförmigen Schacht (inf. und sup.) und der Schicht 9 inf.
wurden maßgeblich Tierknochen, wie Rothirsch (Cervus elaphus), Wildpferd
(Equus ferus) und Bos/Bison (Bovini) gefunden, die gegenüber großen
Temperaturschwankungen unempfindlich sind. Vereinzelt sind auch Ren
(Rangifer tarandus) und Wollnashorn (Coelodonta antiquitatis) vorhanden. In
Schicht 9 sup. herrschen vier Taxa vor, welche in Kantabrien nicht gängig
waren. Zu jenen gehören Ren, Riesenhirsch (Megaloceros giganteus), Mammut
(Mammuthus primigenius) und das Wollnashorn aus der kaltzeitlichen Fauna.
Letzteres repräsentiert mit sieben Individuen 8% des gesamten Ensembles,
wobei fast komplette Hirschskelette (>60%), Analysen zufolge, den Großteil der
Fauna in den untersuchten Fundschichten ausmachen. Viele der Knochen sind
wenig fragmentiert und verfügen über Bissspuren von verschiedensten
Karnivoren (11,8%), unter anderem von Höhlen-Hyänen (Crocuta crocuta
spelaeus), Wölfen (Canis lupus), Bären (Ursus spelaeus) und Füchsen

64
(Vulpes). Überdies wurden Überreste von Hyänen geborgen, die ebenfalls zu
den repräsentativen Arten des Fundplatzes gehören, wurden geborgen. Das
Skelett des jüngsten Exemplars wurde auf ein Alter von mehr als neun Monaten
geschätzt und war somit ein subadultes Tier. Jungtierknochen sind nicht
vorhanden. Dies lässt darauf schließen, dass jene Tiere, die in der Höhle
verendeten, nach dem Verzehr ihrer Beute nicht mehr heraus kamen und dort
verendeten.
Die Zusammensetzung des lithischen Ensembles, der Taxa und der Zustand
der Knochen deuten eher auf sporadische und kurze Aufenthalte der
Neandertaler in der Grotte hin, sei es zur Verwertung von Kadavern –nur drei
Arten verfügen über Zerschlagungs-, Schnitt- und Brandspuren: Wildpferd,
Rothirsch und Bos/Bison–, oder um Unterschlupf zu suchen.
8.4 Saint-Césaire (Moustérien denticulé,
Moustérien/Châtelperronien/Aurignacien)
Der Fundplatz von Saint-Césaire55, der auch als la Roche-à-Pierrot bekannt
ist, liegt im Süd-Westen Frankreichs im Landkreis Charente-Maritimes (Morin et
al. 2005; Morin 2012). Die Stelle, die 1970 während dem Bau einer
Zufahrtsstraße –zu dem sich damals dort befindlichen Steinbruch, der als
Pilzfarm genutzt wurde– wahrgenommen wurde, befindet sich in einem Tal auf
einer fünf bis sechs Meter hohen, gelben Kalksteinklippe (Morin 2012). Nach
ihrer Entdeckung wurde die Fundstelle in zwölf aufeinander folgenden
Kampagnen bis ins Jahr 1987 freigelegt (Morin et al. 2005). Während
Ausgrabungen wurden Fundschichten zu Tage gebracht, die sich über mehrere
Jahrtausende erstreckten. Es wird angenommen, dass es bei dem Fundplatz
ehemals um eine Grotte oder einen Felsüberhang gehandelt haben muss.
Einige Indizien, wie die Lage der Artefakte, deuten auf diesen Umstand. Aus
der Sagittalansicht ist ersichtlich, dass die einzelnen Lagen die Tendenz
besitzen sich zur Klippe hin abzusenken 56 . Ähnliches konnte bereits bei
anderen Ausgrabungen observiert werden (Morin 2012).
Zwei große stratigraphische Gruppen stachen besonders hervor (Morin et al.
2005; Morin 2012) –zum einen die obere gelbe Schicht und die tiefer liegende
55 Abb.24: Geographische Lokalisierung des Fundplatzes von Saint-Césaire 56 Abb.25: Sagittalansicht der einzelnen Schichten

65
graue Schicht. Der gelbe Abschnitt weist eine Dicke von circa 1,60 Metern auf,
Komma gehört zum Jungpaläolithikum und beinhaltet Artefakte sowie
Faunenreste aus dem Moustérien/Châtelperronien und dem Aurignacien (Morin
2012). Die tiefer liegende graue Schicht besitzt eine Dicke von circa 1,20
Metern und beinhaltet Artefakte sowie Faunenreste aus dem Moustérien de
tradition acheuléene und dem Moustérien denticulé 57 . Für den kulturellen
Vergleich sind die Schichten EGF 12 bis EJO sup. 65859 von Bedeutung, da sie
in den bearbeiteten Zeitraum eingegliedert werden können. Die Schichten des
Moustérien denticulé60 (EGF 12 bis EGPF 10) fügen sich mit Hilfe der TL
Datierung in einen Zeitraum von im Durchschnitt 40.000 BP ein. Die Daten
variieren zwischen 36,8 +/- 3,7 und 42,4 +/- 4,3. Das jüngere
Moustérien/Châtelperronien (EJOP inf. 9 bis EJOP sup. 8) in einen ungefähren
Zeitraum von 36,300 BP (TL) verankern und das Aurignacien (EJOP inf. 7 bis
EJOP 6) in einen Zeitraum von circa 34,000 BP (TL) +/- 3,900 (s. d. Tab.:10 )
(Morin 2012).
Im Juli 1979 wurden in der Schicht EJOP sup. 8 Überreste von einem
Neandertaler gefunden. Das Skelett war unvollständig und in einen Umkreis
von 70 cm verstreut. Es fehlten die Füße, die linke Hälfte vom Schädel, ein paar
Zähne als auch einige andere Teile. 1993 warf Vandermeersch in seinem
Artikel die Frage auf, ob das Skelett intentionell dort abgelegt wurde, da er ein
solches Szenario für möglich hielt (Vandermeersch 1993; Morin 2012).
Unterstützung für seine These erhielt er von Morin et al. (2005). In der gleichen
Schicht wurden Zähne von mindestens zwei weiteren Individuen entdeckt sowie
ein Knochen, der an einer Schnittstelle zu einer höher liegenden Ebene lag. Es
stellte sich heraus, dass er (proximal Phalanx) zu einem weiteren Individuum
gehört. Untersuchungen zufolge wurde er der Mittel-Aurigancien Ebene (EJF)
zugwiesen. Das Neandertalerskelett wurde in vieler Hinsicht untersucht, wobei
sich zum einen mit der Anatomie befasst wurde (Trinkaus et al. 1998;
Crèvecoeur 2002) und zum anderen mit seinem Nahrungsspektrum. Die 57 Tab.8: Darstellung der Stratigraphie von Saint-Césaire mit TL Datierung der verschiedenen Level 58 Tab.9: Stratigraphische Zuordnung der kulturellen Abfolge 59 Die Fundschichten von Saint-Césaire wurden 2011 von Bordes neu eingestuft, somit gibt es zu früheren Publikationen Unterschiede in der Abfolge (s. Tab.9: Stratigraphische Zuordnung der kulturellen Abfolge). Für diese Arbeit wurde sich an der neuen Abfolge orientiert. 60 Ältere Fundschichten des Moustérien wurden in Saint-Césaire noch nicht vollends untersucht, daher werden diese hier außer Acht gelassen. Morin weist darauf hin, dass jene in einer noch kommenden Publikation vorgestellt werden sollen (Morin 2012, 57).

66
Untersuchung der Stabilen Isotopen Komposition im Skelett ergab, dass sich
dieses Individuum größtenteils von Huftieren ernährte (Morin 2012).
Womit wir bei der Frage wären, wie sich die Fauna in den verschiedenen
Fundschichten zusammensetze und welche Tiere bevorzugt gejagt wurden? In
Saint-Césaire wurden in den verschiedenen Schichten mehrere Tausend
intakte Konchen sowie Knochenfragmente ausgegraben. Um die großen
Massen an Knochen irgendeiner Tierart zuweisen zu können, startete Anfang
der 2000e Jahre eine umfangreiche Studie, die nicht identifizierte Knochen
einem Taxon zuordnen sollte. Des Weiteren wurden alle Fundschichten einzeln
untersucht, um Unterschiede in den kulturellen Sequenzen feststellen zu
können. Daraus ging hervor, dass es große Unterschiede im Vorkommen der
Knochen zwischen den einzelnen Level gab. Die größten Differenzen konnte
man zwischen Level 8 und 6 feststellen (Morin et al. 2005). Rinderartige
(Bison/Auerochs), Ren und Pferd gehören zu den meist vertretenen Taxa in
Saint-Césaire. Sie bestreiten unter den identifizierten Knochen mit 91,7 % bis
99,5 % den Großteil des Ensembles. Im Moustérien denticulé dominieren das
Pferd und Rinderartige (Bison) das Spektrum, währenddessen im
Moustérien/Châtelperronien Ren und Horntiere als Hauptvertreter agieren und
Pferde in den Hintergrund treten. Das Aurignacien wird deutlich von Ren
bestimmt. Oftmals fand man von diesen Gattungen nur noch Schädelknochen
(Bison und Pferd), Humerus-, Tibia-, Metakarpalknochen (Pferd) und
Hinterläufe (Ren). Andere Arten wie das Mammut, der Rothirsch und das
Rhinozeros sind dagegen nur selten anzutreffen. Von ihnen tauchten vermehrt
Rippen, Langknochen und Zähnen auf. Ebenso rar sind Karnivoren, wie der
Bär, der Wolf und die Hyäne. Je nach Periode verzeichnen sie gerade einmal
0,5% bis 6,7% der Funde.
Paläoökologische Untersuchungen von Saint-Césaire gewähren einen
Einblick in das Klima der Region jener Zeit. Lavaud-Girard beschäftigte sich in
den 1980e und 1990e mit dem Zeitabschnitt des Aurignaciens in Saint-Césaire.
Ihrer Meinung nach deutet der hohe Anteil an Ren während dieser Periode
darauf hin, dass das Klima eher trocken, aber auch milder als in der Umgebung
war. Sie verbindet diesen Zustand mit der Nähe des Fundplatzes zum Atlantik.
Untersuchungen der Mikrofauna durch Jean-Claude Marquet (1993) bestätigten
ihre Aussagen.

67
8.5 Grotte von Spy (Moustérien/Lincombian-Ranisian-Jerzmanowice/Aurignacien)
Die Grotte von Spy wurde Mitte des 19. Jahrhunderts von Marcel De Puydt
et Maximin Lohest entdeckt61. Sie ist neben ihrem umfassenden Inventar auch
für die beiden 1886 in einem stratigraphisch intakten Kontext gefundenen
Neandertalerskelette bekannt. Während der zahlreichen Grabungskampagnen
stellte man eine Gesamtstratigraphie fest, welche vom Mittelpaläolithikum bis in
das Holozän reichte. Diese Grotte umfasst fast das gesamte paläolithische
Wissen aus Belgien.
Unterhalb des LRJ befand sich eine Industrie, die dem Moustérien
zugeschrieben wurde. Ulrix-Closset platzierte die Objekte, welche sich aus
Moustérien-Spitzen und Blattspitzen zusammensetzen in das Moustérien
„évolué“. Flas teilte die Blattspitzen in seinen späteren Recherchen dem LRJ
zu. Des Weiteren zählen bifacielle Stücke zu den Funden dieses Levels, die
Kleimessern oder Blattspitzen gleichgestellt wurden. Aufgrund ihrer Typologie
wird ihnen ein orientalischer Einfluss zugesprochen. Die säuberliche bifacielle
Bearbeitung war ausschlaggebend sie dem Moustérien „évolué“ zuzuordnen.
Da jener Begriff in diesem Rahmen aber kritisch gesehen wird –wir befinden
uns am Ende des Mittelpaläolithikums–, wurde er in Moustérien récent à
pointes foliacées umgetauft (Semal et al. 2011).
Im darüber liegenden LRJ wurden invers retuschierte Blattspitzen freigelegt,
von denen ein Teil in stratigraphisch gestörten Schichten lagen. Sie wurden
Anfangs als „Pointes de Spy“ bezeichnet, sind mittlerweile aber unter dem
Namen Jerzmanowice Spitzen bekannt, die über 30 Fundplätzen in
Nordosteuropa verteilen (Semal et al. 2011).
Die umfangreichste Schicht wird durch das Aurignacien mit seinen Stein- und
Knochenartefakten verkörpert. Da allerdings keine stratigraphisch sicher
Kontext vorhanden ist, lassen sich die Stücke nur schwer bestimmen. Aus
diesem Grund existiert aus der Anfangszeit der Grabungen lediglich eine kurze
Beschreibung der Fragmente. Nach Flas Aufarbeitung des Ensembles konnten
die Geräte in unterschiedliche Phasen gegliedert werden. Ein Teil des Inventars
wurde in das Aurignacien ancien eingruppiert. Unter den Artefakten fanden sich
61Abb.26: Geographische Lokalisierung der Spy Grotte

68
kielförmige Kratzer, mittelgroße Dufour Lamellen sowie Spitzen mit einer
gespaltenen Basis. Zum Inventar des Aurignacien récent gehören viele Kratzer
mit einer Nase sowie Vachon-Stichel, hakenförmige Stichel und eine Reihe an
Geräten aus Elfenbein (Semal et al. 2011).
8.6 Ranis (Lincombian-Ranisian-Jerzmanowice)
Die Ilsenhöhle wurde bereits Anfang und Mitte des 20e Jahrhunderts
ausgegraben. Sie liegt in Thüringen nahe der Stadt Ranis unterhalb der
Hauptburg. Ausgegraben und beschrieben wurde sie erstmals von Werner
Hülle (1977). Dieser Fundplatz ist insofern interessant, da er eine gut
dokumentierte Gesamtstratigraphie von 12 Level vorweisen kann 62 . Eine
besondere Stellung bezieht Ranis 2 mit seinen Steinartefakten und
„Knochengeräten“, in die Zeit des LRJ eingegliedert werden. Die
Knochengeräte, die heute verloren sind, umfassten 25 Exemplare. Zwei zogen
insbesondere die Aufmerksamkeit aus sich. Bei dem einen Stück handelte es
sich um eine Knochenspitze, die als Vogelpfeil interpretiert wurde. Das andere
Exemplar wurde als Elfenbeinscheibe beschrieben, welche mittig mit einem
Loch versehen war (Hülle 1977)63. Aufgrund ihrer Position und Form, wurden
sie von Flas als mögliche Geräte des LRJ angesehen.
8.7 Vedrovice V (Szeletien)
Vedrovice ist eine Ortschaft, die auf den östlichen Hängen des südlichen
Teils des Kromauer Waldes circa 40 südwestlich von Brno liegt. Die Fundstelle
Vedrovice V wurde Mitte der 1950e von Effenberger entdeckt und infolge des
Fundes eines Blattspitzenfragmentes dem Szeletien zugeordnet. Die
Besiedlung wurde in mehreren Kampagnen ausgegraben, während denen eine
umfangreiche Spaltindustrie mit einigen aus zähem Material bestehenden
Geröllen zu Tage gebracht wurde. Im Gegensatz zu anderen Gebieten
befanden sich im Inventar auch kleine Klümpchen bzw. amorphe Stückchen
Farbstoffarten. Zu erkennen waren Limonit, Hämatit und Graphit. Bis auf einige
Fragmente wurden keine Knochen entdeckt, die aufgrund von
Erosionsprozessen fast vollkommen vernichtet wurden. Der Fundplatz 62 Abb.27: Gesamtstratigraphie der Ilsenhöhle 63 Abb.28: Mögliche Knochenspitze und Elfenbeinscheibe des LRJ

69
beherbergte mehrere Feuerstellen, die einmalig oder mehrmals benutzt wurden
und zu deren Inhalt verkohlte Knochen gehörten. Die Artefakte konzentrierten
sich in bestimmten Bereichen des Fundplatzes. Das Rohmaterial wurde aus
einer Distanz von 200-300 m herangebracht und bestand zu mehr als 99% aus
Silizite. Die Grabungen erbrachten 17.000 Artefakte, von denen zwei Drittel aus
Absplissen (kleiner als 2 cm) bestand. Das Inventar setzte sich aus zwei
Typengemeinden zusammen. Auf der einen Seite mittelpaläolithische (u.a.:
Fäustel und Keilmesser, Moustier- und Pseudolevallois-Spitzen, einen
Chopper und Blattspitzen) und auf der anderen Seite jungpaläolithische
Artefakte (u.a.: Endretuschen, Kratzer, Stichel, Längsretuschen, Klingen und
Abschläge, aufgesplitterte Stücke an Kostienki-Messer erringende Geräte:
(Karel Valoch 19(?)).
8.8 Brno-Bohunice (Bohunicien)
Brno-Bohunice gilt als Typus-Fundplatz für das Bohunicien, Komma stammt
aber zugleich aus einem schwierigen Kontext. Die Stelle wurde von einem
Amateur Archäologe ausgegraben, der die Artefakte aus Bulldozergraben
auflas. Er versuchte sie mit Hilfe von anerkannten Protokollen zu identifizieren,
konnte aber keine Übereinstimmungen finden. Da die Funde nicht
Ordnungsgemäß geborgen wurden und ist es trotz Rettungsversuchen seitens
Valoch schwierig diese Artefakte in einen Kontext zu integrieren. Im Gegensatz
zu anderen Fundplätzen des Bohuniciens ist die Vielfalt an Artefakte größer. Zu
den Funden zählt eine Varietät an Rohmaterialien, aber auch retuschierte
Geräte. Besonders stechen bifaciell gearbeitete Blattspitzen hervor, da sie
einzigartig für das Bohunicien sind. Durch ihre Existenz wird die Frage laut, ob
sie einen Hinweis auf die Vermischung mit dem Szeletien darstellen. Um dieser
Frage eine Antwort zu liefern, wurde der Fundplatz 2002 erneut ausgegraben.
So ließ sich das Durchmischen von Horizonten ausschließen und das
Ensemble konnte als homogene Unität anerkannt werden. Weiterhin wurde ein
neues Inventar aus dem späten Mittel- und dem frühen Jungpaläolithikum
freigelegt. Auch in diesem befanden sich bifacielle Spitzen. Aufgrund dieses
Gerätetyps, der in anderen Fundplätzen des Bohuniciens nicht vorkommt, gilt
es zu verstehen wie innerhalb eines Technokomplexes eine solche Diskrepanz
entstehen kann.

70
8.9 Stránská skála (Bohunicien)
Stránská skála gehört zu den wichtigsten Fundplätzen des Bohuniciens. Es
liegt westlich von Brno 310 m.ü.M64. Es handelt sich um eine Freilandfundstelle,
der mehrere teils vermischte Horizonte unterliegen. In den untersten Schichten
fand man paläolithische Artefakte. Aufgrund seiner Dimensionen wurde der
Platz in mehrere Sektoren aufgeteilt. Es wurden überwiegend Steinartefakte
freigelegt sowie eine kleine Anzahl an Knochen und Kohlefragmente. Eisenerz
wurde auch identifiziert. Bis auf zwei Stellen hielt sich die Menge der Objekte in
Grenzen. Gewöhnlich wurden weniger als 50 Geräte bzw. Abschlagabfall pro
Sektor geborgen. In den Bereichen, in denen die Konzentration an
Schlagabfällen am höchsten war, wurde die Vermutung aufgestellt, dass jene
Stelle als Werkzeugherstellungsplatz fungierte. Trotz Kohlefunde wurden
innerhalb der Stratigraphien keine Feuerstellen festgestellt. Eisenerz kam
vermehrt vor. Welchen Zweck es jedoch hatte, wurde nicht herausgefunden, da
weder Brand- noch andere Arten an Spuren daran erkannt wurden. Aufgrund
der hohen Anzahl, schätzte man jedoch, dass sie für die Gruppe eine
Bedeutung haben mussten. Da die faunische Überreste für die Forschung
verloren gingen, ist es nicht möglich eine Aussage zu treffen. Die Konstellation
des Fundplatzes verweist auf einen reinen Arbeitsplatz, der für die Herstellung
von Werkzeugen vorgesehen war. Es ist nicht gewusst, ob sich eine
Behausung in der Nähe dieses Camps befunden hat (Valoch et al. 2009).
8.10 Grotta del Cavallo (Moustérien/Uluzzien)
Die Grotta del Cavallo liegt in Apulien im Süden Italiens in der Nähe von
Santa Caterina in einer Bucht Namens Uluzzo. Sie befindet sich nur einige
Meter über dem Meeresspiegel an deren felsigen Küste im ionischen Meer. Sie
wurde 1960 entdeckt. In den folgenden Jahren nahm Arturo Palma di Cesnola
stratigraphisch Stichproben. Die Grotte besitzt eine Gesamtstratigraphie von 31
Schichten mit drei verschieden Technokomplexen65. Das Moustérien mit seinen
16 Schichten (FI-N) übernimmt den Großteil der Stratigraphie gefolgt vom
Uluzzien mit 6 Schichten (DIa-EIII). Im obersten Bereich ruht eine lithische
64 Abb.29: Geographische Lokalisierung des Fundplatzes Stránská skála 65 Abb.30: Stratigraphie aus der Grotta del Cavallo

71
Industrie aus dem späten Epigravettien (BIa-BII), welche im Umfang dieser
Arbeit nicht weiter beachtet wird. Die Kulturen sind jeweils mit einer Schicht
Höhlensinter (DIa) (Hublin 2014) und Tephra (CII und Fa) voneinander getrennt
(Boscato/ Crezzini 2012).
Bis heute wurden nur wenige Schichten der Grotta del Cavallo komplett
aufgearbeitet, daher ist Gesamtvergleich zwischen dem Moustérien und dem
Uluzzien nicht möglich. Bisher erfuhren die Fundschichten EIII5, F, H, I und L
eine nähere Untersuchung. Das homogene Ensemble setzt sich vorwiegend
aus Huftieren zusammen, jedoch herrscht in jeder Schicht ein anderes Taxon
vor. In den Ablagerungen EIII5, FI und FII konnten insgesamt 393 Knochen
identifiziert werden (Boscato/Crezzini 2012). Von jenen sind 204 Stück in
Schicht EIII5 zu lokalisieren, wovon acht zu Karnivoren gehören und die
restlichen zu Huftieren. Von diesen identifizierten Knochen wurden etwa 17,5 %
bearbeitet. Punkt Glieder sind vornehmlich –was auf eine Zerlegung der Tiere
hinweist– mit Schnittspuren versehen. In FI wurden 76 Knochen identifiziert,
von denen 66 Überreste von Huftiere sind, während FII 113 aufweist, wovon
104 Huftieren zugeordnet werden. Im Uluzzien sind im Gegensatz zum
Moustérien viele Handwurzel- und Fußwurzelknochen, Finger-/Zehenknochen
und Sesambeine nicht unüblich. Jene markieren den größten Unterschied
zwischen den Ensembles des Mittel- und Jungpaläolithikums der Grotta del
Cavallo. Die drei Fundschichten wurde bezeugen, dass neben Rothirsch und
Pferd überwiegend Auerochse (Bos primigenius) gejagt wurde. In Schicht F
gesellen sich zum Auerochsen geringe Mengen an Damhirsch (Dama dama).
Letzterer ist in Schicht EIII5 nicht mehr vorzufinden. In Schicht tauchen
überwiegend Pferdknochen auf und in den beiden Schichten H und L Rothirsch.
Des Weiteren wurden Wildschweinreste (Sus scrofa), europäischer Wildesel
(Equus hydruntinus) als auch eine heute ausgestorbene Nashorn Gattung
(Stephanorhinus) gefunden. Bis auf zwei Überreste Alpensteinbock (Ibex) in
Schicht I, ist die Unterfamilie der Caprinae nicht vertreten. Karnivoren sind
ebenfalls Teil der Funde, sind jedoch weitaus weniger präsent als Herbivoren.
Vier Taxa überwiegen in allen Schichten: der Wolf (Canis lupus), der Rotfuchs
(Vulpes vulpes), der eurasische Luchs (Lynx lynx) und die Wildkatze (Felis
silvestris) (Boscato/Crezzini 2012).

72
8.11 Grotta di Fumane (Moustérien/Uluzzien)
Fumane ist eine kleine Gemeinde im Nordöstlichen Italien in der Provinz
Verona. Sie liegt in den Vizentiner Voralpen und ist Namensgeber für die
gleichnamige Grotta di Fumane, welche auf eine Höhe von 350 m ü.d.M. am
Randgebiet der Lessinischen Bergen liegt (Tagliacozzo et al. 2013; Benazzi et
al. 2014).
Die Grotte ist seit dem 19. Jahrhundert bekannt und wurde das erste Mal im
Jahr 1964 vom Historischen Museum aus Verona untersucht. Unter dem
Patronage des UNESCO Weltkulturerbe wird die Höhle seit 1988 in
regelmäßigen Abständen ausgegraben. Dabei wurde eine 12 m dicke
Stratigraphie aus dem Mittel- und Jungpaläolithikum zutage gebracht (Peresani
2012)66. Diese umfasst 50.000 Jahre, die sich vom Moustérien bis in das
Uluzzien ausdehnen. Vier Hauptsequenzen (S, BR, A und D) sind während der
Grabungen erstellt worden. Faunische Überreste (Tagliacozzo et al. 2011),
lithischen Artefakte (Peresani 2012), Feuerstellen (Peresani et al. 2011) sowie
menschliche Zähne (Benazzi et al. 2014) aus dem Moustérien (Fumane 1, 4, 5)
und dem Uluzzien (Fumane 6) zeugen von wiederholten und auch
umfassenden Besuchen (Peresani 2012). Die Sequenzen B11, A11, A8-A9 und
A5-A6 waren Phasen, welche besonders von Menschen besucht wurden. Die
älteren Schichten bis A6-A5 waren vom Moustérien belegt. Die Schichten A4
und A3 vom Uluzzien, A2 und A1 vom Proto-Aurignacien und die darüber
liegenden Schichten A3 und A1 vom Aurignacien. Um eine einigermaßen
genaue Vorstellung über diese Zeitspanne zu erhalten, wurden aus
unterschiedlichen Level Proben mittels 14C Methode (ABA und ABOx-SC), U-
Th/ ESR Datierung sowie TL Datierung untersucht. Das späte Moustérien
umspannte das OIS 3 und das Uluzzien einen Zeitraum von 44.400 +/- 4.000
(U-Th/ ESR) bis 37.828 +/- 430 (ABA 14C) (43,6-43,0 und 41,2-40,4 cal. BP,
nach Higham et al. 2009; Peresani 2012).
Die Sequenzen A4 und A3 des Uluzzien enthielten 36.944 Knochen. Diese
beliefen sich auf 3% des gesamten Ensembles. Gerade einmal 1.188 konnten
zugewiesen werden. Insgesamt identifizierte man 17 Genera und 19
Gattungen, die zu fünf verschiedenen Säugetierordnungen gehören. Reh,
66 Abb.31: Gesamtstratigraphie der Fumane Grotte

73
Rothirsch, Alpensteinbock, und Füchse waren zahlreich vertreten. Marder,
Höhlenbär, Höhlenhyäne, Lux, Wollnashorn und Leopard waren dagegen
lediglich durch wenige Exemplare repräsentiert. Beide Schichten bargen 575
Vogelüberreste und obschon die meisten Knochen stark fragmentiert und zu
95% kleiner als 3 cm waren, gelang es 47,1% (Genus und Gattung) zu
identifizieren. Alpendohle, Birkhuhn und Wachtelkönig herrschten vor.
8.12 Klissoura Grotte (Uluzzien/Aurignacien)
Die Klissoura Grotte 1 liegt nordwestlich des Peleponnes in einer Schlucht
mit dem gleichen Namen (Starkovich 2014). Sie wurde ende der 1980e während
einer Feldbegehung entdeckt. Nachdem Anfang der 1990e erste
Probegrabungen stattfanden, stieß man auf eine Gesamtstratigraphie, die vom
späten Pleistozän bis in das Holozän hineinreicht67. In einer der untersten
Schichten V wurden Artefakte aus einer frühen Phase des Jungpaläolithikums
identifiziert (Uluzzien), in zwei darüber liegenden Schichten mehrere Phasen
des Aurignacien und die Schichten II bis I stammen aus dem Epigravettien,
dem Mesolithikum und der klassischen Bronzezeit (Koumouzelis et al. 2001).
Neusten Datierungen zufolge, wird Schicht V auf ein Alter von 39.280 +/- cal.
BP geschätzt. Die darüber liegenden Schichten des Aurignaciens umspannen
den Zeitraum von 37.500-35.000 cal. BP (Kuhn et al. 2010; Lowe et al. 2012).
In Schicht V wurde ein Inventar von 2.800 Artefakten ausgegraben, welches
sich aus 1.500 kleinen Abschlägen, ca. 130 Klingen und 370 Abschlägen
zusammensetzt. 132 Geräte wurden aus zersplitterten Stücken, oder aus
bogenförmigen rückenretuschierten Klingen hergestellt. Kleine Kratzer und in
seltenen Fällen Stichel und Bohrer. Des Weiteren wurde ein Mikrostichel sowie
eine mikrolithische Endretusche, die aussieht wie eine Zonhoven Spitze
geborgen. Beide gelten als besonders interessant. 58% der Artefakte wurden
aus R1 Radiolarit hergestellt. Feuersteine wurden am zweithäufigsten
verwendet. Der erste Rohstoff wurde nicht in jüngeren Fundschichten
festgestellt. In Schicht III ist primär das frühe Aurignacien von Bedeutung, da
jenes an das Uluzzien grenzt. In seiner frühen Phase sind Endschaber,
67 Abb.32: Gesamtstratigraphie der Klissoura Grotte

74
zweiflächige Stichel, zersplitterte Stücke sowie retuschierte Abschläge
vorhanden (Koumouzelis et al. 2001).
Aus den Schichten wurde ein umfangreiches faunisches Ensemble
herausgeholt. Dieses stellte sich unteranderem aus Säugetieren, Vögel aber
auch Mollusken zusammen. Bei den Säugetieren wurden 18 unterschiedliche
Arten bestimmt, die auf 1774 partielle oder ganze Knochen aufgeteilt sind.
Damwild und Hase dominieren mit 56% und 23%. Karnivore und andere
Kleintiere sind seltener vertreten. Die Knochen weisen keine Bisspuren auf,
dafür sind 25% mit Brandspuren versehen. Die Tierknochen verfügen über
anthropogene Spuren, die von Jagd- und Schlachtaktivitäten herrühren. Die
121 Vogelknochen repräsentieren mindestens fünf Taxa. Die meisten Gebeine
gehören zum Steinhuhn (Alectoris graeca), der Großtrappe (Otis tarda), Eule
(Asio), Dohle (Corvus monedula) und Rabe (Corvus corone). Die beiden ersten
Exemplare wurden wahrscheinlich aufgrund ihrer Größe gejagt. Das Inventar
der Mollusken enthält 1239 Schalen von Gastropoden. Vier bis fünf Arten
gehören den Landschnecken an, andere dagegen den Meeresschnecken.
Zweischalige Exemplare waren auch vertreten, konnten jedoch nicht identifiziert
werden. Die meisten dieser Schalen (Helix figulina) –919 an der Zahl– wurden
aus den Schichten des Aurignaciens geborgen. Schicht V schloss Fragmente
zweier Spezies Dentalium und Helix figulina ein. Recherchen von Starkovich
(2014) zeigen ein bestimmtes Bild für die Wahl der Jagdbeute. In den meisten
Sequenzen dominieren mittelgroße Huftiere. Im Aurignacien gewinnen große
Huftiere aus einer milderen Phase an Bedeutung. Hasen wurden in allen
Phasen gerne gejagt, kommen aber erst nach dem Uluzzien häufiger vor.
Steinhuhn und Großtrappe wurden in späteren Phasen des Aurignaciens
bevorzugt.
8.13 Roc-de-Combe (Châtelperronien, Proto-
Aurignacien/Aurignacien)
Roc-de-Combe ist eine kleine Grotte im Südwesten Frankreichs. Sie ist für
die TMJ von großer Bedeutung da sie eine der wenigen Fundplätze in der
Region ist, die ein gutes faunisches Ensemble für das Châtelperronien und
Aurignacien sowie mehrere Aurignacien Ensembles vorweisen kann. Sie liegt
im Gebiet Bouriane etwa 9,5 km südlich des Dordogne Flusses. 1950 wurde

75
der Fundplatz von einem Mann namens Jean Labrot entdeckt und erstmals
1959 ausgegraben. Bei dieser Grabung wurden archäologische Sequenzen aus
dem Moustérien, dem Châtelperronien und dem Aurignacien identifiziert. 1966
wurde eine weitere Kampagne von François Bordes und Jacques Labrot
durchgeführt, die in den Schichten Perigordien (Châtelperronien), Aurignacien,
Perigordien und wieder Aurignacien sahen. Sie nahmen eine wechselnde
Besiedlung beider Gruppen in der Grotte an (Grayson/Delpech 2008). Ein Teil
der Stratigraphie wurde in den 1990e und 2000e mithilfe der 14C Methode
datiert. Schicht 8 (Châtelperronien) wurde auf ein ungefähres Alter von 40.000
BP geschätzt und die Schicht 7-5 zwischen 34.800 +/- 1200 BP und 28.500 +/-
700 BP (Grayson/Delpech 2008). Zur richtigen Identifikation der Level hat
allerdings erst viel später Bordes J.-G. dazu beigetragen. In seiner Dissertation
(2002) arbeitete er den gesamten Fundkomplex noch einmal vollständig auf.
Im Roc- de-Combe stellte man große Unterschiede in der Anzahl sowie der
Taxa der faunischen Reste fest, welche auf die klimatischen Bedingungen und
andere Faktoren zurückzuführen sind. In einer neu durchgeführten Analyse,
wurden die Levels 5-8 (Châtelperronien und Aurignacien) aufgearbeitet. Alles
umfassend wurden 12 verschiedene Taxa innerhalb der Huftiere68 identifiziert
und 10 innerhalb der Karnivoren69. Zu jenen gehören Bos/Bison, Capra sp.,
Capreolus capreolus, Cervus elaphus, Equus sp., Equss caballus, Equus
hydruntinus, Rangifer tarandus, Rhino, Rupicapra sp., Sus sp. und Mammut. Es
wurden 3506 Knochen bestimmt. Rentier dominiert mit 3070 Knochen
(Grayson/Delpech 2008). Vollständige Knochen sind in beiden Ensembles
(Châtelperronien und Aurignacien) nur selten vertreten (Soulier/Mallye 2011).
Da die Anzahl an Knochen lediglich beim letzten genannten Taxon hoch war,
bestand die Möglichkeit bei dieser Art eine Körperteilanalyse durchzuführen.
Aufgrund der geringen Anzahl bei den anderen Individuen, war es nicht möglich
eine Gesamtübersicht der Präferenzen zu erstellen. Aus den Resultaten der
Körperteilanalyse beim Rentier geht hervor, welche Körperteile bevorzugt mit in
die Höhle gebracht und weiterverarbeitet wurden. In allen Schichten überwiegt
Ren gefolgt von Rind, Pferd und Hirsch. Andere Arten umfassen weniger als
3%. Die größte Vielfallt lässt sich in Schicht 8 und die geringste in Schicht 5
68 Tab.11: Anzahl der identifizierten Taxa bei den Huftieren. Mammut nicht mit in der Berechnung inbegriffen. 69 Tab.12: Anzahl der identifizierten Taxa bei den Karnivoren

76
erkennen. Die beiden anderen Level liegen im Durschnitt. Die Karnivoren
setzten sich aus Meles meles, Mustela nivalis, Vulpes/Alopex, Ursus sp.,
Mustela erminea, Crocuta crocuta, Canis lupus, Lynx sp., Marte sp. und
Panthera spelaea zusammen. 147 Knochen konnten bestimmt werden, wobei
Vulpes/Alopex mit 88 Knochen in allen Level der häufigste Vertreter ist. Die
Tierkombination verweist auf ein kaltes Milieu. Knapp 306 Herbivorenknochen
sind mit Schnittspuren versehen, der Rest blieb unberührt. Jene Spuren sind
diejenigen von Schlachtaktivitäten (Häutung). Anhand der Beschaffenheit der
Frakturen lässt sich erkennen, dass die Knochen im frischen Zustand bearbeitet
wurden. 33 wurden durch Beißspuren beschädigt und nur 49 verfügen über
Verbrennungsspuren, wobei der Anteile im Ensemble des Aurignaciens bei
50% liegt und im Châtelperronien nur bei 10% (Grayson/Delpech 2008;
Soulier/Mallye 2011). Karnivorenknochen der Schicht 7 sind mit Schnittspuren
versehen, welche nicht unbedingt als Schlachtaktivität interpretiert wurden. In
Schicht 8 konnten keine Spuren festgestellt werden. Anhand der
Rentierknochen wurde ein Altersprofil erstellt. Daraus ging hervor, dass primär
Tiere unter einem Alter von 5 Jahren erlegt wurden. In den Schichten des
Aurignaciens identifizierte man Knochen ältere Tiere. Unter den Pferden
(Châtelperronien) wurden drei trächtige Tiere beobachtet. Nähere Angaben
zum Alter konnten jedoch nicht gemacht werden. Aufgrund fehlender Substanz,
war ein Erstellen weiterer Profile nicht möglich. Die Größen der Föten ließen
erahnen, dass im Châtelperronien im Spätsommer bis Herbst und im
Aurignacien im Spätsommer gejagt wurde (Soulier/Mallye 2011).
9 Problematik des vorgestellten Zeitraums und der dazugehörigen archäologischen Kulturen
Trotz intensiver Erforschung bringt der Zeitraum zwischen 50.000 und
35.000 v.Chr. eine Reihe an Problemen mit sich, angefangen mit der Datierung.
Wie an den vorgestellten Beispielen zu erkennen ist, fällt es schwer viele der
einzelnen Technokomplexe einzugliedern. Das liegt zum einen an den häufigen
Interstratifikationen bei den Schnittstellen der einzelnen Schichten (bei
Datierungsversuchen kann es zu Verunreinigungen durch die jüngeren
Gegenstände kommen) und zum anderen kommen Datierungsmethoden wie

77
die Radiokarbonmethode an ihre Grenzen. Proben können nur bis zu einem
maximalen Alter von 50.000 bis 60.000 Jahren datiert werden. Je älter die
Proben sind desto weniger 14C enthalten sie. Durch modernen Kohlestoff kann
es zu Verunreinigungen kommen, wodurch das Alter der Probe wesentlich
jünger ausfällt als angedacht (http://www.14c.uni-erlangen.de/grundlagen.html).
Methoden wie die Lumineszenz kommen dann zum Einsatz wenn 14C an seine
Grenzen stößt. Angewandt wird sie für den Zeitraum von 50.000 bis 100.000
Jahren (in manchen Fällen auch älter), um verbrannte Sedimente sowie Flint
(Quarz, Quarzite, Feuerstein etc.) zu analysieren. Problematisch ist jedoch,
dass auch diese Methode begrenzte Fähigkeiten hat. Besonders bei jüngeren
und sehr alten Proben kann es zu Ungenauigkeiten kommen. Des Weiteren
können Umwelteinflüsse diese Methode verfälschen, sodass falsche
Altersangaben als Resultat angezeigt werden. Doch richtig angewandt und
ausgewertet, kann jene eine Bereicherung und Ergänzung zu anderen
darstellen, da zum Beispiel der Zeitpunkt des letzten Erhitzens einer Feuerstelle
ermittelt werden kann (Richter 2007).
9.1 Neandertaler oder anatomisch moderner Mensch?
Da wir uns an einem Wendepunkt befinden, der sich auf kultureller sowie
biologischer Basis abspielt, haben sich viele mit der Frage beschäftigt, wer die
eigentlichen Initiatoren der Übergangskulturen70 innerhalb der TMJ waren (u.a.:
Flas 2008; Bar-Yosef-Bordes 2010; Benazzi et al. 2011; Morin 2012). Viele
sehen in diesen großen Veränderungen den anatomisch modernen Mensch,
der ein gereiftes Paket an modernen Eigenschaften und Technologien mit sich
führte (Hublin 2013). Andere favorisieren wiederum den Neandertaler, da er zu
bereits früheren Zeitpunkten selbst eine „Art der Verhaltensbezogenen
Modernisierung“ durchlief. McBrearty und Brooks (2000) argumentieren, dass
ein solcher Prozess sich über Raum und Zeit unabhängig von Arten allmählich
entwickelte und nicht urplötzlich irgendwo zwischen 40-50 ka auftauchte, wie es
das „human revolution“ Modell suggeriert (s.a.: „Multi Species Model“ in d’Errico
2003). Im Zentrum der Debatten stehen die kognitiven Fähigkeiten des
Neandertalers sich neuen Gegebenheiten anzupassen und „moderne“
70 Abb.33: Übersichtsplan der besprochenen Übergangsindustrien, gegliedert nach deren chronologischen Erscheinen in 14C cal. Daten und NGRIP.

78
Merkmale zu entwickeln (Mellars 1989, 1996, 2005; Klein 1995; Conard 2010).
Dies führt zu einer weiteren Frage, die sich darauf bezieht, ob diese Kulturen
eventuell Produkt einer Akkulturation beider Arten waren und ob der
Neandertaler lediglich die Technologien des anatomisch modernen Menschen
kopierte (s.: Kapitel 9.2). Das Szeletien (z.B: Allsworth-Jones P. 1986), das
Uluzzien (z.B.: Riel-Salvatore 2009) und allen voran gestellt das
Châtelperronien (z.B.: Roussel/Soressi 2014) wurden lange als die
Schlüsselkulturen für die Weiterentwicklung des Neandertalers hervorgehoben.
Argumente diesbezüglich werden primär in geographischen, aber auch in
technotypologischen Merkmalen gesucht, wobei das erstgenannte kritisch
betrachtet wird, da einige der Kulturen geographisch und chronologisch sehr
begrenzt sind, sodass eine Zuordnung zu einer homininen Gruppe zusätzlich
erschwert wird (Hublin 2015: 198, 205). Dieses Bild kam jedoch ins Wanken
nachdem Benazzi et al. (2011) die in der Grotte von Cavallo gefundenen Zähne
(Schichten EIII & EII-I) dem anatomisch modernen Menschen zuordneten.
Vergleiche, wie zwischen dem Châtelperronien und dem Uluzzien –welches
aufgrund seiner ähnlichen typologischen Merkmale an das Châtelperronien
herangerückt wurde– (Schintler 2011) müssen somit in ein neues Licht gerückt
werden (Akkulturation, Koexistenz/Interaktion?). Angesichts von eindeutig
identifizierten Knochen anderer Fundstellen (Spy in Belgien und Saint-Césaire
in Frankreich) kann zumindest ein Teil der Übergangsindustrien dem
Neandertaler als Schöpfer zugeschrieben werden (Hublin 2015). Sich diese
Situation als Gesamtbild vor Augen zu führen bleibt schwierig, da viele der
Funde früherer Grabungen in inadäquater Form geborgen wurden. Dadurch
gingen viele Informationen verloren. Das größte Problem besteht im Mangel an
menschlichen fossilen Resten und solange keine weiteren zutage gebracht
werden, bleibt nur die Möglichkeit in den anderen Fällen Hypothesen zu
erstellen, welche sich auf unterschiedliche Indizien (Ursprung einer Industrie:
Moustérien, Micoquien oder anderen eindringenden Kulturen) stützen (Djindjian
2011).

79
9.2 Akkulturation, Wunschdenken oder Realität? Und wie wahrscheinlich ist eine Koexistenz verschiedener Kulturgruppen, mit sporadischem oder regelmäßigem Austausch kultureller Aspekte sowie „Tauschgut“?
In Verbindung mit der Ausbreitung des Homo sapiens innerhalb Eurasiens ist
die Frage der Akkulturation eine der meistdiskutierten (d’Errico et al. 1998;
Tostevin 2007). Innerhalb Westeuropas wird der Fortschritt archaischer
Kulturen oftmals damit begründet, dass sie sich ihre technologischen
Fähigkeiten bei eingewanderten Kulturen angeeignet haben. Mit einer
außerordentlichen Reihe von Kritiken und Gegenargumenten wurde versucht
jene Theorie zu widerlegen. Infolge dessen wurden unterschiedliche Modelle
der Koexistenz und Akkulturation vorgestellt (Tostevin 2007: 341). Das Problem
der Thematik liegt in der vielfachen Verwendung der Stratigraphien (Gravina et
al. 2005), um Pros oder Contras für die eigene Theorie zu finden. Heute
oberhalb der Sahara lebende Menschen (Eurasien) weisen zwischen 1-3%
Neandertaler-Gene auf. Als möglicher Zeitpunkt wurde ein Zeitraum zwischen
47-65 ka vorgeschlagen (Sankararaman et al. 2012). Aktuelle Genetische
Analysen an einem aus Peştera cu Oase in Rumänien stammenden
Individuum, dessen Alter auf ∼37.000-42.000 14C BP geschätzt wird, ergaben
einen 6-9% Wert der Neandertaler-Gene. Daraus folgt, dass einer seiner
Vorfahren, vier bis sechs Generationen zuvor, ein Neandertaler war (Fu et al.
2015). Bei einem weiteren ∼45.000 Jahren alten Mann aus Sibirien wurde ein
möglicher Zeitpunkt für den Austausch genetischen Materials zwischen 42.000
und 45.000 Jahren festgelegt (Fu et al. 2014). Mit dem Eintreffen des
anatomisch modernen Menschen in Europa vor ∼45.000-43.000 Jahren
koexistierten beide Arten zwischen 2.600 und 5.400 Jahren (95,4%
Wahrscheinlichkeit: Higham et al. 43.500 cal. BP: 2014; Nigst 2014). Da es zu
genetischem Austausch gekommen ist, gab es zumindest bei einigen Gruppen
irgendeine Form von Interaktionen. Bei der Annahme, dass beide
Kulturgruppen ihrerseits eine „Modernisierung“ durchlaufen haben und es bei
Begegnungen mit der anderen Art zur Akkulturation gekommen ist, stellt sich
nun die Frage, ob durch die bisher nicht eindeutig zugeordneten
Übergangsindustrien erkannt werden kann, wer die Initiatoren sind. Ebenso gut

80
könnten einige der Technokomplexe, wie bereits von Flas (2008) erwähnt, von
zwei unterschiedlichen Kulturgruppe kreiert, wobei jede seine eigenen
Technologien mit einbrachte (Beispiel: Châtelperronien, Uluzzien). Da bisher
von beiden Hominini keine fossilen Reste gemeinsam in Fundschichten
entdeckt und auf ein gleiches Alter datiert wurden, können dazu lediglich
Vermutungen aufgestellt werden.
9.3 Auf welchem Weg kam der anatomisch moderne Mensch
nach Westeuropa?
Zwischen 60/50-50/40 ka fing die Expansion des anatomisch modernen
Menschen nach Eurasien an (Seguin-Orlando et al. 2014; Nigst et al. 2014). Da
aus diesem Zeitraum fossile Reste allerdings nur spärlich vorhanden sind, wird
viel über mögliche Routen spekuliert. Von der Levante sind erste
jungpaläolithische Industrien von vor ∼48.000 Jahren bekannt, deswegen
wurde schon früh angenommen, dass es sich hierbei um eine mögliche Route
handeln könnte (Seguin-Orlando et al. 2014). Ein Multinationales Forscherteam
unter der Leitung des Max-Planck-Instituts untersuchte kürzlich eine der
wenigen jungpaläolithischen Fundstätten im Nahen Osten (Ksâr `Akil). Mittels 14C Methode wurden die Daten von einem Skelett, einem Unterkiefer („Egbert“
und „Ethelruda“) sowie wie von Schneckenhäusern (Phorcus turbinatus)
ermittelt. Daraus resultierten Datierungen zwischen 43.000-42.900 cal. BP für
„Egbert“ und ∼45.900 cal. BP. Vor allem die Zweite Datierung gilt als wichtig,
da sie als „Marker“ für die Ausbreitung des Jungpaläolithikums nach Europa
gesehen werden kann (Bosch et al. 2015). Weitere Indizien finden sich nördlich
der Alpen am Fundplatz Willendorf II, der zu den ältesten in Europa gehört. Der
archäologische Horizont 3 (AH3) wird um 43.500 ca. BP eingestuft, an den
Anfang des Grönland Interstadial 11 und fällt somit älter als bisher datierte
Aurignacien Ensembles aus (Nigst et al. 2014: 14394). Eine südliche Route
könnte somit unterstützt werden (Beyin 2011).
Aus Radiokarbon und Lumineszenz Datierungen von Fundplätzen am Don
Fluss geht hervor, dass dort bereits vor 45.000 bis 42.000 Jahren (≤ 48.000 cal.
BP; Hoffecker 2009) Industrien siedelten. Angesichts der Artefakte aus
Knochen und Bein, dem Schmuck aus Muscheln, der aus einer Distanz von 500

81
km heran geschafft wurde und der figurativen Kunst ist es wahrscheinlich, dass
sie vom Homo sapiens stammen. Sie wären dementsprechend der jüngste
Beweis für anatomisch moderne Menschen in Zentral- und Osteuropa. Die
hohe Anzahl an Fundkomplexen aus der Periode vor der KI-Eruption zeigt die
Wichtigkeit dieses Gebietes für die weitere Ausbreitung und könnte als
mögliche alternative parallel verlaufende Route angesehen werden (Anikovich
et al. 2007). Ein weiterer wichtiger Fund stammt vom Kostenki 14 Fundplatz
(Russland). Ein 1954 ausgegrabenes Skelett (K14) wurde kürzlich auf ein Alter
von 38.700 bis 36.200 cal BP datiert. Es grenzt an die KI-Eruption (∼39.3 ka
cal. BP) und unterliegt einer frühen jungpaläolithischen Industrie. Dieses
Individuum ist in sofern interessant da Untersuchungen des Genoms zeigen,
dass jenes aus einer Zeit nach der Trennung der Erblinien (westeurasische
„europäische„ und nordeurasische „asiatische“ Merkmale) stammt. Es gehört zu
der Abstammungslinie aus der sich die heutigen Europäer entwickelt haben und
zeigt am meisten Ähnlichkeit mit den heutigen Nordeuropäern (Litauer,
Skandinavier etc.). Gemeinsame Allele zu europäischen Farmern aus dem
Neolithikum und heute im mittleren Osten und im Kaukasus lebenden
Menschen, sowie Sibirien lebenden Menschen (aus dem Gebiet des Yenisei
Flusses) wurden auch festgestellt. Wie die weiter oben vorgestellten Individuen
aus Rumänien und Sibirien enthält K14 Spuren von Neandertaler-Genen, die in
einen ungefähren Zeitraum vor 54.000 Jahren zurückverfolgt werden können.
Das Beispiel von diesem Subjekt zeigt wie komplex die Abläufe zu jener Zeit
waren und welchen Beitrag es zu späteren Generationen vom
Jungpaläolithikum bis in die heutige Zeit beigetragen hat (Segeuin-Orlando
2014).
9.4 Was führte zum Aussterben der Neandertaler?
Der Neandertaler entwickelte sich vor etwa 200.000 Jahren auf dem
eurasischen Kontinent und reifte zu einer der vorherrschen Spezies heran. Er
passte sich unterschiedlichen klimatischen sowie geographischen Bedingungen
an und verstand es seine Umwelt zu seinen Gunsten zu gebrauchen, bis in
einen Zeitraum herein in dem eine weitere Art der Gattung Homo auf den
Kontinent einwanderte und ihn allmählich ersetzte. Das Aussterben des

82
Neandertalers warf viele Fragen auf und wurde vielseitig besprochen. Da es
keine ersichtlichen Gründe für das Verschwinden dieser Art zu geben scheint,
wurden in den letzten Jahrzehnten Hypothesen entwickelt, die deren ableben
erklären sollen. So besagen einige Thesen, dass Krankheiten (Epidemien,
genetisch bedingte Krankheiten; Riel-Salvatore 2008; ����� ����
������������ � ������� Underknown 2008), Gifte (Størmer/Mysterud 2007),
Klimaveränderungen (d’Errico/ Sánchez Goñi 2004; Jiménez-Espejo et al.
2007), Naturkatastrophen, Hungersnöte, Probleme bei der Geburtenrate, sowie
ein ausgedehnter Konkurrenzkampf um die Ressourcen zwischen den beiden
Arten (Banks et al. 2008; Shipman 2012) schwerwiegende Auslöser sein
können. Szenarien, zu denen auch Underknowns (2008) Spongiform
Enzephalopathie (TSE) zählt, werden allgemein nicht unterstützt. Ferner
scheint das Argument der Hungersnöte ebenfalls fragwürdig zu sein, da sich
Nahrungsengpässe bei einer angenommen Population von ca. 10.000
Individuen, die sich auf der gesamten Fläche Europas verteilten, nur kurzfristig
auf der regionalen Ebene auswirkt hätten (Sørensen 2011). Ein Modell zur
Klimaveränderung und den damit verbunden Konsequenzen (die letzte
Rückzugsnische des Neandertalers im Süden von Gibraltar zwischen 30.000
und ∼28.000 BP) stellten unter anderem Jiménez-Espejo et al. (2007) vor.
Diese Idee basiert auf der These, dass fallende und stark schwankende
Temperaturen während des OIS 3 eine tragende Rolle spielten. Da sich Fauna
und Flora infolge jenes klimatischen Umschwungs drastisch veränderten,
gelang es der restlichen Population nicht sich an diese neuen Konditionen
anzupassen. Besonders Kinder und ältere Personen wären die Leidtragenden
gewesen, sodass es zu einem Ungleichgewicht entstanden wäre. Sørensen
(2011) benannte in seinem Klimamodell die Neandertaler aus dem Saale
Kältemaximum als eine Referenzgruppe und beschrieb drei mögliche Szenarien
für die Entwicklung der Population. Im ersten Szenario wurde die klimatisch
bedingte Todesrate um 50% zur Referenzkurbe (C-1) erhöht, wodurch die
südeuropäische Population ausstarb. Die klimatisch bedingte Todesrate
verringert sich im zweiten Szenario um 30% zur Referenzkurbe (C-1). Dies
führte zu einem Anstieg der Population, sodass Hungersnöte als Ursache für
das Aussterben nicht mehr so unrealistisch erscheinen. Im dritten Szenario
wurde anhand der Referenzkurbe eine „normale“ klimatisch bedingte Todesrate

83
errechnet, die eine wesentlich geringere Population im Gegensatz zur
Wechselkaltzeit während des Saale Kältemaximums hervor gerufen hätte. Ein
Überleben der Art am Kälteminimum der letzten Kaltzeit müsste demnach
möglich gewesen sein. Sørensen bemerkt jedoch, dass in seinem Modell die
Gesamtheit der Klimaschwankungen beider Kaltzeiten nicht berücksichtigt
wurden und das Klima wohl auch kein bedeutungsvolles Kriterium sei.
Naturkatastrophen könnten hingegen substanziell sein, denn innerhalb eines
kurzen Zeitraums ereigneten sich zwei große Vulkanausbrüche. Vor 75.000
Jahren brach der Pina Toba (die größte bisher dokumentierte Eruption) aus und
um ∼39.000 BP wurde die KI Eruption registriert. Sekundäre Effekte –
Abwandern der Jagdbeute oder Veränderung der Flora– könnten zu einer
Umsiedelung in den Süden geführt haben. Da im Zeitraum nach der KI Eruption
in Mittel- und Nordeuropa aber immer noch die bevorzugte Jagdbeute zugegen
war, wurde auch diese Theorie verworfen (Sørensen 2011). Ein Aussterben
aufgrund Probleme bei der Geburtenrate (Kindbetttot, hohe Rate an Säuglings-
und Kindersterblichkeit) wurde zwar auch in Betracht gezogen, schnell aber
wieder beiseite gelegt. Die Todesrate bei Geburten war beim anatomisch
modernen Menschen nicht geringer als bei seinem ausgestorbenen
Verwandten. Dies hätte sich dementsprechend auf beide Arten auswirken
müssen (Sørensen 2011). Die Gewalteinwirkung durch Dritte kann auch
vernachlässigt werden. Skelette aus dem Zeitraum der Koexistenz weisen
keine merklichen Verletzungsmuster auf, die auf gängige Gewalteinwirkungen
hindeuten könnten. Eine gezielte Ausrottung kann somit ausgeschlossen
werden. Eine weitere Theorie bezieht sich auf Krankheiten, die Epidemien unter
den Neandertalern verursachten. Aus der jüngeren Geschichte ist ein
besonderer Fall bekannt, in dem Populationen durch das Einschleppen von
Viren und Bakterien fast vollständig ausgerottet wurden. Die Rede ist von den
Pockendecken, die an die indianische Bevölkerung ausgeteilt wurden. Da ihr
Immunsystem solche Erreger nicht kannte, entflammte eine tödliche Seuche in
Mitten der Bevölkerung. Auch im Bezug auf das Aussterben der Neandertaler
wird die Argumentation laut, dass der anatomisch moderne Mensch einen
unbekannten Erreger einschleppte, der sich ähnlich auf die Neandertaler
auswirkte (Wolff/Greenwood 2010). Unterstützend wird angeführt, dass die
Neandertaler von Ost nach Südwest verschwanden und dies genau dem Weg

84
des Menschen entspräche (Sørensen 2011). Das „Competitive exclusion“
Modell wird in der Literatur vielfältig besprochen (Stewart 2004; Banks et al.
2008; Shipman 2012; Fa et al. 2013; Hortalà/Martínez-Navarro 2012 usw.).
Aufgrund der Ausbreitung des anatomisch modernen Menschen soll der
Neandertaler einer ungewohnten Stresssituation ausgesetzt gewesen sein.
Verbesserte Technologien, eine bessere Jagdstrategie sowie eine Konkurrenz
in Jagdbeuteschema –mit dem Unterschied, dass der Speiseplan des
Menschen variabler war–, seien signifikante Faktoren. Zurückdrängung und die
Unfähigkeit sich an die neuen Konditionen anzupassen, indem zum Beispiel
nicht auf andere Ressourcen zurück gegriffen oder innovative Technologien
entworfen wurden, führten zu einem unausweichlichen Aussterben unseres
nahen Verwandten. Angesicht der In der herrschenden Uneinigkeit spiegelt sich
die Komplexität dieser Thematik wieder und es scheint, als würde jede
mutmaßliche Antwort neue Fragen aufwerfen.
10 Zusammenfassung und Fazit
Die Transition vom Mittel- zum Jungpaläolithikum überbrückt einen relativ
kurzen Zeitraum im Jungpleistozän. Während dieser Zeitspanne treten auf der
klimatischen, geologischen, kulturellen und biologischen Ebene drastische
Veränderungen auf, welche die Zukunft des Menschen und seiner nahen
Verwandten besiegelt. Klimatisch bestimmt das OIS 3 die Verhältnisse, das für
seine kurzen sowie drastischen Klimaschwankungen (H-Events) und für ein
geologisches Großereignis (KI Eruption) bekannt ist. Vor 50.000 Jahren
herrscht die Spätphase des Moustériens, welches kurz vor seinem Ende steht.
Parallel und kurz danach entwickeln sich lithische Kulturen, die im Gegensatz
zu Mittelpaläolithikum Neuerungen in der angewandten Technik, aber auch im
Gerätespektrum aufweisen. Sie gelten als Übergangsindustrien zum
Jungpaläolithikum. In jener Phase zwischen 50.000 und 35.000 v. Chr.
wanderte indes auch der anatomisch moderne Mensch auf den eurasischen
Kontinent (vor ∼45.000 bis 43.000 Jahren) ein, sodass fossile Funden
innerhalb von Stratigraphien, die in den Industrien rar gesät sind, nur mit
Schwierigkeit einer besagten Kultur einer Art zugeordnet werden können.
Lediglich die in stratigraphisch intakten Kontexten gefundenen Skelette konnten

85
Aufschluss über einige dieser Übergangsindustrien geben. Diese Zeitspanne ist
insofern interessant, da sie neue Denkanstöße zu alteingesessenen Meinungen
liefert. Bemühungen diese Kulturen einer gewissen Menschengruppe
zuzuschreiben, führen unweigerlich zu Vergleichen beider Arten, bei denen in
der Regel der Homo sapiens als überlegene Spezies angepriesen wird. Dies
lässt sich besonders bei dem Subsistenzverhalten beobachten. Die These, der
anatomisch moderne Mensch sei mit einem Gesamtpaket an modernen
Verhaltensmustern und Technologien nach Eurasien eingewandert, ist
besonders bekannt. Der Neandertaler sei hingegen anlässlich seiner kognitiven
Kapazitäten in seiner Evolution stagniert.
Neuste Funde (geometrische Gravur, umfunktionierte Seeadlerkallen)
bezeugen jedoch, dass dieses Monopol der „Modernisierung“ nicht länger
ausschließlich dem Homo sapiens gebührt. Sie vollzog sich vielmehr an
unterschiedlichen Orten zu unterschiedlichen Zeiten unabhängig der Art. Bei
der Betrachtung der kognitiven Fähigkeiten könnte die Schnelligkeit in der eine
solche Entwicklung erfolgte als ein Argument für eine Überlegenheit
herangezogen werden. Im direkten Vergleich der beiden Arten ließen sich im
Umfang dieser Arbeit zugleich große Unterschiede neben einigen
Gemeinsamkeiten erkennen. Die lithischen Kulturen weisen definitiv eine
fortschreitende Evolution ausgehend von eher großen Allzweckgeräten
(Faustkeile, Keilmesser usw.) zu feineren spezialisierten Artefakten auf, die je
nach Region bei den Übergangsindustrien anders ausfallen. Während bei
einigen Technokomplexen Blattspitzen eine tragende Rolle spielen, wurden bei
anderen eher Abschläge oder Klingenabschläge produziert. Zu diesen
Artefakten gesellten sich zusätzlich verschiedene Sorten an Schabern, Bohrern,
Sticheln, Pièces esquillées usw. Mit dem Anfang des Jungpaläolithikums ändert
sich die Produktionsweise vollkommen. Zur Basisproduktion gehören ab jenem
Zeitpunkt Klingen und Lamellen unterschiedlicher Größen, die größtenteils zu
Spitzen weiterverarbeitet wurden. Stichel und Schaber zählen ebenfalls zum
Inventar und Geräte aus Geweih, Knochen und Bein erleben einen
Aufschwung, da sie zu Geschossspitzen ummodelliert wurden. Die signifikanten
Unterschiede zwischen den Waffen beider Arten lassen sich nur erahnen,
Komma da lediglich witterungsbeständige Fragmente erhalten bleiben. Es steht
außer Frage, dass sowohl Homo neanderthalensis als auch Homo sapiens

86
Komposit -lanzen und -speere verwendeten. Pfeil und Bogen sowie
Speerschleudern könnten potentiell schon seit Beginn des Jungpaläolithikums
gefertigt worden sein. Einschlägige Beweise fehlen für diese Annahme
allerdings. Über den gesamten Umfang der angewandten Jagdstrategien kann
auch nur spekuliert werden. Aufgrund der zahlreich entdeckten Verletzungen an
den Neandertalerknochen, ist es wahrscheinlich, dass unsere nahen
Verwandten auf eine Jagd von kurzer Distanz spezialisiert waren. Die Funde
von Lanzen und Speere deuten ebenso in diese Richtung. Während der TMJ
fehlen Knochen des anatomisch modernen Menschen vollständig, sodass in
ihrem Fall ausschließlich Vermutungen geäußert werden können.
Die vorgestellten Beispiele bezeugen eine Tendenz des Jagbeuteschemas.
Generell wurden innerhalb beider Menschengruppen mittelgroße bis große
Säugetiere gejagt, mit dem Unterschied, dass man bei jungpaläolithischen
Fundplätzen vermehrt auf kleinere Tiere trifft. Je nach Saison und Gebiet fielen
die Präferenzen unterschiedlich aus, in der Frage des Alters war man sich
jedoch Größtenteils einig. Bevorzugt wurden adulte Tiere zur Strecke gebracht,
Jungtiere und ältere Tiere dagegen selten. Die Stratigraphien bieten keine
Hilfestellung bei der Frage, ob sich ein Austausch zwischen beiden
Menschengruppen vollzog. In einigen Fundplätzen wie zum Beispiel Saint
Césaire wurden zwar Interstratifikationen einer Lage mit der darüber liegenden
Schicht aus dem Aurignacien festgestellt, dennoch werden diese oftmals nicht
als Beweis anerkannt und werden eher Witterungsbedingungen zugeschrieben.
Obwohl keine offensichtlichen Anzeichnen zu sehen sind, ist eine
Kontaktaufnahme der Arten nicht völlig unmöglich, zumal sich diverse dieser
Übergangskulturen mit anderen zeitlich überschnitten. Des Weiteren sollte man
nicht der Annahme erlegen sein, dass es sich um ein statisches System handle.
Der Homo neanderthalensis und der Homo sapiens führten einen mobilen
Lebensstil und konnten große Distanzen zurücklegen. Außerdem koexistierten
sie zwischen 2.600 und 5.400 Jahren, so muss es trotz kleinerer
Gruppierungen zu Kontakten gekommen sein. Die Stratigraphien
dokumentieren teilweise eine Abfolge von über 50.000 Jahren, die vom
Mittelpaläolithikum bis in das Jungpaläolithikum reicht. So lässt sich ablesen,
dass über Generationen hinweg die gleichen Plätze aufgesucht wurden.
Demnach kann nicht ausgeschlossen werden, dass Gruppen unterschiedlicher

87
Menschen Arten das Gebiet eines anderen durchstreiften. Eine Vermischung
beider Hominini innerhalb der TMJ lässt sich bis Dato nicht bestätigen.
Die 1-3% Neandertaler-DNA, die in der heutigen eurasischen Population
auftaucht, verweist auf einen weiter zurückliegenden Zeitpunkt.
Fand innerhalb der Zeitspanne der TMJ eine erneute Vermischung statt,
würden die Nachkommen aus einer Erblinie stammen, die nicht mit dem
heutigen Stammbaum synchron verläuft. Der prägnanteste Unterschied beider
Spezies beläuft sich auf den Fakt, dass der Neandertaler vor ∼41.000-39.000
cal. BP ausstarb und der anatomisch moderne Mensch sich zu der
dominierenden Art auf der Erde weiterentwickelte.

Anhang

2
Literaturverzeichnis
1. Ackermann H.: Joachim Neander- Sein Leben, seine Lieder, sein Tal. Presseverband der evangelischen Kirche im Rheinland, Düsseldorf, 1980.
2. Allsworth-Jones P.: The Szeletian and the Transition from Middle to Upper Palaeolithic in Central Europe. Oxford, Clarendon Press (1986), 412 S.
3. Álvarez-Solas J./ Montoya M./ Ritz C./ Ramstein G./ Charbit S./ Dumas C./ Nisancioglu K./ Dokken T./ Ganoploski A.: Heinrich Event 1: an example of dynamical ice-sheet reaction to oceanic changes, –in: Climate of the Past 7 (2011), S. 1297-1306.
4. Arrizabalga A.: El yacimiento arqueológico de Labeko Koba (Arrasate, País Vasco). Entorno. Crónica de las investigaciones. Estratigrafía y estructuras. Cronología absoluta, –in: Munibe (Antropologia-Arkeologia) N° 52 (2000), S. 15-72.
5. Arrizabalaga A./ Altuna J.: Consideraciones generales sobre el depósito arqueológico de Labeko Koba (2000a), S. 385-395.
6. Arrizabalaga A./ Altuna J.: Labeko Koba (País Vasco). Hienas y Humanos en los albores del Paleolítico superior, –in: Sociedad de Ciencias Aranzadi-Aranzadi Zientzia Elkartea, Donostia-San Sebastián (2000b).
7. Bachellerie F./ Bon F./ Deschamps M./ Eizenberg L./ Henry-Gambier D./ Mourre V./ Normand C./ Pelegrin J./ Primault J./ Scandiuzzi R./ Thiébaut C.: Archaeological Signatures of Hunting Activities Applied to Comparisons of Mousterian, Chatelperronian and Aurignacian Industries in the Pyrenees: the Nature of Hunting Tools and Site Functions, –in: Bon F., Costamagno S., Valdeyron N. (eds.), Hunting Camps in Prehistory. Current Archaeological Approaches, Proceedings of the International Symposium, May 13- 15 2009, University Toulouse II - Le Mirail, P@lethnology, 3 (2011), S. 131-167.
8. Baena J./ Carrión E./ Ruiz B./ Ellwood B./ Sesé C./ Yravedra J./ Jordá J./ Uzquiano P./ Velásquez R./ Manzano I./ Sánchez-Marco A./ Hernández F.: Paleoecología y comportamiento humano durante el Pleistoceno Superior en la comarca de Liébana: La secuencia del al Cueva de El Esquilleu (Occidente de Cantabria, Espagña), –in: Museo de Altamira. Monografías N° 20 (2005), S. 461-487.
9. Baena J./ Carrión E./ Cuartero F./ Fluck H.: A chronicle of crisis: The late Mousterian in north Iberia (Cueva del Esuilleu, Cantabria, Spain), –in: Quaternary International 247 (2012), S. 199-211.
10. Banks W.E./ d'Errico F./ Peterson A.T./ Kageyama M./ Sima A./ Sánchez-Goñi M-F.: Neanderthal Extinction by Competitive Exclusion, –in: PLoS ONE 3(12) (2008): e3972.
11. Banks W.E./ d’Errico F./ Zilhão J.: Revisiting the chronology of the Proto-Aurignacian and the Early Aurignacian in Europe: A reply to Higham et al.’s comments on Banks et al. 2013, –in: Journal of human evolution (2013), S. 1-8.
12. Barton R.N.E/ Currant A.P./ Fernandez-Alvo Y./ Finlayson J.C./ Goldberg P./ Macphail R./ Pettiitt P.B./ Stringer C.B.: Gibraltar Neanderthals and results of recent excavations in Gorham’s, Vanguard and Ibex Caves, –in: Antiquity 73 (1999), S. 13-23.
13. Bar-Yosef O.: The Upper Palaeolithic revolution. Ann. Rev. Anthropol. 31 (2002), S. 363–393. 14. Bar-Yosef O./ Bordes J.G.: Who were the makers of the Châtelperronian culture? Journal of
Human Evolution 59 (2010), S. 586-593. 15. Benazzi S./ Bailey S.E./ Peresani M./ Mannino M.A./ Romandini M./ M.P./ Hublin J.-J.: Middle
Paleolithic and Uluzzian human remains from Fumane Cave, Italy, –in: Journal of Human Evolution 70 (2014), S. 61-68.
16. Benazzi S./ Douka K./ Fornai C./ Bauer C.C./ Kullmer O./ Svoboda J./ Pap I./ Mallegni F./ Bayle P./ Coquerelle M./ Condemi S./ Ronchitelli A./ Harvati K./ Weber G.W.: Early dispersal of modern humans in Europe and implications for Neanderthal behaviour, –in: Nature 479 (2011), S. 525–528.
17. Benazzi S./ Slon V./ Talamo S./ Negrino F./ Peresani M./ Bailey S.E./ Sawyer S./ Panetta D./ Vicino G./ Starnini E./ Mannino M.A./ Salvadori P.A./ Meyer M./ Pääbo S./ Hublin J.-J.: The makers of the Protoaurignacian and implications for Neandertal extinction, –in: Science Vol. 348, Issue 6236 (2015), S. 793-796.
18. Berger T.D./ Trinkaus E.: Patterns of Trauma Among the Neandertals. Journal of Archaeological Science 22 (1995) :841–852.
19. Bischoff J. L./ Williams R. W./ Rosenbauer R. J./ Aramburu A./ Arsuaga J. L./ García N./ Cuenca-Bescós G.: High- resolution U-series dates from the Sima de los Huesos hominids yields 600 +∞/-66 kyrs: implications for the evolution of the early Neanderthal lineage, –in: Journal of Archaeological Science 34 (2007): S. 763-770.

3
20. Beyin A.: Upper Pleistocene Human Dispersals out of Africa: A Review of the Current State of the Debate, 2011.
21. Bocherens H, Billiou D, Mariotti A.: Palaeoenvironmental and palaeodietary implications of isotopic biogeochemistry of last interglacial Neanderthal and mammal bones in Scladina Cave (Belgium), –in: J. Archaeol. Sci. 26 (1999), S. 599–607.
22. Bocherens H, Billiou D, Mariotti A, Toussaint M, Patou-Mathis M, Bonjean D, Otte M.: New isotopic evidence for dietary habits of Neandertals from Belgium, –in: J. Hum. Evol. 40 (2001) S. 497-505.
23. Bocherens H, Drucker D, Billiou D, Patou-Mathis M, Vandermeersch B.: Isotopic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal: review and use of a multi-source mixing model, –in: J. Hum. Evol. 49 (2005) S. 71-87.
24. Bolus M.: Der Übergang vom Mittel- zum Jungpaläolithikum in Europa. Eine Bestandsaufnahme unter besonderer Berücksichtigung Mitteleuropas. Germania 82 (2004), S. 1–54.
25. Bolus M.: Die Neandertaler- Verbreitung und Expansion einer europäischen Menschenform, –in: Ditte Bandini und Ulrich Kronauer: 100 Jahre Heidelberger Akademie der Wissenschaften, Früchte vom Baum des Wissens, Eine Festschrift der wissenschaftlichen Mitarbeiter (2009), S. 21-33.
26. Bolus M./ Schmitz R.W.: Der Neandertaler. Ostfildern, Thorbecke, 2006. 27. Bon F./ Maíllo Fernández José M./ Ortega Cobos D.: En torno a los conceptos de
Protoauriñaciense, de auriñaciense arcaico, inicial y antiguo (Unidad y variabilidad de los componentes tecnológicos de los primeros humanos modernos en el sur de Francia y norte de España), Madrid, Espacio, Tiempo y Forma. Prehistoria y Arqueología, t. 15, 2002, S. 13-15 (2006), Actes de la table ronde de Toulouse (2003), 262 S.
28. Bon F./ Teyssandier N./ Bordes J.-G.: Les significations culturelle des équipements lithiques, S. 49-72, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
29. Bond G.C./ Lotti R.: Iceberg Discharges into the North Atlantic on Millennial Time Scales During the Last Glaciation, –in: Science Vol. 267 (1995), S. 1005-1010.
30. Bordes F.: Essai de Classification des industries „moustériennes“, –in: Bulletin de la Société préhistorique française, tome 50, N°7-8 (1953), S. 457-466.
31. Bordes F.: avec une note paléontologique de J. Bouchud. Les gisements du Pech de l’Azé (Dordogne), I : Le Moustérien de tradition acheuléenne. L’Anthropologie, 58 (1954), S. 401-432 et 59 (1955), S. 1-38.
32. Bordes F.: Vingt-cinq ans après: le compexe moustérien revisité, –in: Bulletin de la Société préhistorique française, tome 78, N°3 (1981), S. 77-87.
33. Bordes J.-G.: Les interstratifications Châtelperronien/Aurignacien du Roc-de-Combe et du Piage (Lot, France). Analyse taphonomique des industries lithiques; implication archéologiques. Ph.D dissertation, University Bordeaux I (2002).
34. Bordes J.-G./Teyssandier N.: Reprint of: Upper Paleolithic nature of the Châtelperronian in south-western France: Archeostratigraphic and lithic evidence, –in: Quaternary International 259 (2012), S. 95-101.
35. Bosinski G.: Die mittelpaläolithischen Funde im westlichen Mitteleuropa, –in: Fundamenta A4, Köln-Graz, Böhlau Verlag (1967).
36. Bosinski G.: El Paleolítico medio en Europa central, Zephyrus, t. 53-54 (2000-2001), S. 79-142. 37. Bosinski G.: Urgeschichte am Rhein. Tübingen, Kerns Verlag, 2008. 38. Boscato P./ Crezzini J.: Middle–Upper Palaeolithic transition in Southern Italy: Uluzzian
macromammals from Grotta del Cavallo (Apulia), –in: Quaternary International 252 (2012), S. 90-98.
39. Breuil E.P.: Les subdivisions du paléolithique supérieur et leur signification : Congrès international d'anthropologie et d'archéologie préhistoriques, compte rendu de la XIV session, Genève (1912), Lagny: publ. 2e Ed. 1937, 78 S.
40. Breuil H.: Notes de voyage paléolithique en Europe centrale. I. Les industries paléolithiques en Hongrie, –in: L’Antrophologie, 33 (1923), S. 323-346.
41. Broecker W.: Massive iceberg discharges as triggers for global climate change, –in: Nature Vol. 372 (1994), S. 421-424.
42. Broecker W./ Bond G./ Klas M./ Clark E./ McManus J: Origin of the nothern Atlantic’s Heinrich events, –in: Climate Dynamics 6 (1992), S. 265-273.
43. Budek A./ Kalicki T./ Karminská L./ Kozlowski J.K./ Mester Z.: Interpleniglacial profiles on open-air sites in Hungary and Slovakia, –in: Quaternary Int. 294 (2013), S. 82-98.
44. Burke A.: HUnting in the Middle Palaeolithic. International Journal of Osteoarchaeology 10 (2000), S. 281-285.
45. Cabrera, V./ Maillo J. M./ Lloret M./ Bernaldo de Quiros F.: La transition vers le Paléolithique

4
supérieur dans la grotte du Castillo (Cantabrie, Espagne): la couche 18. L’Anthropologie 105 (2001), S. 505–532.
46. Campbell J.B.: Les problèmes des subdivisions du Paléolithique supérieur britannique dans son cadre européen, Bulletin de la Société royale belge d’Anthropologie et de Préhistoire, t. 91 (1980), S. 39-77.
47. Carbonell E./ Bermúdez de Castro J.M./ Parés J.M./ Pérez-González A./ Cuenca-Bescós G./ Ollé A./ Mosquera M./ Huguet R./ van der Made J./ Rosas A./ Sala R./ Vallverdú J./ García N./ Granger D. E./ Martinón-Torres M./ Rodríguez X. P./ Stock G.M./ Vergès J.M./ Allué E./ Burjachs F./ Cáceres I./ Canals A./ Benito A./ Díez C./ Lozano M./ Mateos A./ Navazo M./ Rodríguez J./ Rosell J. & Arsuaga R.L.: The first hominin in Europe, –in: Nature Vol. 452 (2008), S. 465-469.
48. Carbonell i Roura E.: High Resolution Archaeology and Neanderthal Behavior – Time and Space in Level J of Abric Romaní (Capellades, Spain), Springer, Dordrecht Heidelberg New York London, 2012.
49. Cattelain P.: Apparition et évolution de l’arc et des pointes de flèches dans la Préhistoire européenne (Paléo-, Méso-, Néolithique), –in: Catene operative dell'arco preistorico, Bellintani, P. et Cavulli, F., (Éds.), Incontro di archeologia sperimentale, San Lorenzo in Banale-Fiavè, 30 août - 2 septembre 2002, Giunta della Provincia Autonoma di Trento, Soprintendenza per i Beni Archeologici (2006), S. 45-66.
50. Cattelain P.: Les armes, S. 113-128, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris (2010), 300 S.
51. Chmielewski W.: La civilisation de Jerzmanowice, Instytut Historii Kultury Materialnej Polskej Akademii Nauk, Wrocław- Warszawa-Kraków (1961), 92 S.
52. Clark G. A./ Lindly J. M.: The Case for Continuity: Observations on the Biocultural Transition in Europe and Western Asia. –in: P. Mellars und C. Stringer (Hrsg.), The Human Revolution. Behavioural and Biological Perspectives on the Origins of Modern Humans. Edinburgh: Edinburgh University Press (1989), S. 626–676.
53. Conard N.J./ Bolus M.: Radiocarbon dating the appearance of modern humans and timing of cul- tural innovations in Europe: new results and new challenges. Journal of Human Evolution 44 (2003), S. 331–371.
54. Conard N.J.: Cultural modernity: Consensus or conundrum?, –in: PNAS 107 (17) (2010), S. 7621–7622.
55. Conard N.J./ Kitagawa K./ Krönneck P./ Böhme M./ Münzel S.C.: The Importance of Fish, Fowl and Small Mammals in the Paleolithic Diet of the Swabian Jura, Southwestern Germany. In Zooarchaeology and Modern Human Origins: Human Hunting Behavior During the Later Pleistocene, edited by J. L. Clark and J. D. Speth, Vertebrate Paleobiology and Paleoanthropology. Springer, New York, NY (2013), S. 173-190.
56. Cordain L.: Implications of Plio-Pleistocene Hominin Diets for Modern Humans. –in: Early Hominin Diets: The Known, the Unknown, and the Unknowable. Ungar, P (Ed.), Oxford University Press, Oxford (2006), S. 363-383.
57. Costa A./ Folch A./ Macedonio G./ Giaccio B./ Isaia R./ Smith V.C.: Quantifiying volcanic ash dispersal and impact of Campanian Ignimbrite super-eruption, –in: Geophysical Research Letters, Vol. 39, L10310, 2012.
58. Daujeard C./ Moncel M.-H.: On Neanderthal subsistence strategies and land use: A regional focus on the Rhone Valley area in southeastern France, –in: Journal of Anthropological Archaeology 29 (2010), S. 368–391.
59. Daujeard C./ Fernandes P./ Guadelli J.-L./ Moncel M.-H./ Santagata C./ Raynal J.-P.: Neanderthal subsistence strategies in Southeastern France between the plains of the Rhone Valley and the mid-mountains of the Massif Central (MIS 7 to MIS 3), -in: Quaternary International 252 (2012), S. 32-47.
60. Davies W./ Gallop P.: The Human Presence in Europe during the Last Glacial Period II: Climate Tolerance and Climate Preferences of Mid- and Late Glacial Hominids, Chapter 8, –in: van Andel T.H. & Davies W.: Neanderthals and modern humans in the European landscape during the last glaciation: Archaeological Results of the Stage 3 Project, McDonald Institute for Archaeological Research, 2003.
61. Davies W.: The Abbé Henri Breuil (1877–1961), –in: R. Hosfield, F. Wenban-Smith & M. Pope (eds.) Great Prehistorians: 150 Years of Palaeolithic Research, 1859–2009 (Special Volume 30 of Lithics: The Journal of the Lithic Studies Society): 127Ŕ 141. Lithic Studies Society, London, 2009.
62. Deino A.L./ Southon J./ Terrasi F./ Campajola L./ Orsi G.: 14C and 40Ar/39Ar dating of the Campanian Ignimbrite, Phlegrean Fields, Italy, –in: „Abstracts, ICOG 1994, Berkerley, CA“, 1994.
63. Delagnes A./ Rendu W.: Shifts in Neandertal mobility, technology and subsistence strategies in western France, –in: Journal of Archaeological Science 38 (2011), S. 1771-1783.

5
64. Delluc B..: L’art pariétal, S.215-236, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris (2010), 300 S.
65. Demay L./ Péan S./ Patou-Mathis M.: Mammoths used as food and building resources by Neanderthals: Zooarchaeological study applied to layer 4, Molodova I (Ukraine), –in: Quarernary International 276/77 (2012), S. 212-226.
66. Djindjian F.: La Chronologie, S. 17-32, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
67. Douka K./ Higham T.F.G./ Wood R./ Boscato P./ Gambassini P./ Karkanas P./ Peresani M. Ronchitelli A.M.: On the chronology of the Uluzzian, –in: Journal of Human Evolution 68 (2014), S. 1-13.
68. Drucker D.G./ Bocherens H.: Carbon and nitrogen stable isotopes as tracers of change in diet breadth during the Middle and Upper Paleolithic in Europe. Int. J. Osteoarchaeol 14 (2004), S. 162-177.
69. d'Errico F.: "The Invisible Frontier A Multiple Species Model for the Origin of Behavioral Modernity", S. 188-202, –in: Evolutionary Anthropology 12 (2003).
70. d’Errico F./ Borgia V./ Ronchitelli A.: Uluzzian bone technology and its implications for the origin of behavioural modernity, –in: Quaternary International 259 (2012), S. 59-71.
71. d'Errico F./, Joao Zilhão, Michèle Julien, Dominique Baffier and Jacques Pelegrin: Neanderthal Acculturation in Western Europe? A Critical Review of the Evidence and Its Interpretation, –in: Current Anthropology Vol. 39, No. S1, Special Issue The Neanderthal Problem and the Evolution of Human Behavior (1998), S. 1-44.
72. Ehlers J.: Das Eiszeitalter, Spektrum Akademischer Verlag, Heidelberg, 2011. 73. Engwell S.L./ Sparks R.S.J./ Carey S.: Physical characetristics of tephra layers in the deep sea
realm: the Campanian Ignimbrite eruption, –in: Marine tephrochronologie 398, Geological Society, London, Special Publications, 2014, S. 47-64.
74. Fa J.E./ Stewart J.R./ Lloveras L./ Vargas J.M.: Rabbits and hominin survival in Iberia. Journal of Human Evolution 64 (4) (2013), S. 233-241.
75. Fedele F.G./ Giaccio B./ Isaia R./ Orsi G.: Ecosystem Impact of the Campanian Ignimbrite Eruption in late Pleistocene Europe, –in: Quaternary Research 57 (2002), S. 420-424.
76. Fedele F.G./ Giaccio B./ Isaia R./ Orsi G.: The Campanian Ignimbrite Eruption, Heinrich 4, and Palaolithic Change in Europe: a High-Resolution Investigation, –in: Volcanism and the Earth’s Atmosphere, Geophysical Monogrpaph 139 (2003), S.301-325.
77. Fedele F.G./ Giaccio B./ Isaia R./ Orsi G./ Carroll M./ Scaillet B.: The Campanian Ignimbrite Factor: Towards a reapraisal of the Middle to Upper Paleolithic ‚Transition’, –in: Grattan J./ Torrence R., ed.: Living in the shadow: the cultural Impacts of volcanic eruptions, Left Coast Press: Walnut Creek (CA) (2007), S. 19-41.
78. Fedele F.G./ Giaccio B./ Hajdas I.: Timescale and cultural process at 40,000 BP in the light of the Campanian Ignimbrite eruption, Western Eurasia, –in: Journal of Human Evolution 55 (2008), S. 834-857.
79. Fiedler L./ Rosendahl G./ Rosendahl W.: Altsteinzeit von A bis Z. WGB, Wissen verbindet, Publikationen der Reiss-Engelhorn-Museen, Band 44, 2011, 415 S.
80. Fiorenza L./ Benazzi S./ Tausch J./ Kullmer O./ Bromage T.G./ Schrenk F.: Molar Macrowear Reveals Neanderthal Eco-Geographic Dietary Variation. Rosenberg K, ed. PLoS ONE 6(3) (2011), e14769.
81. Fiorenza L./ Benazzi S./ Henry A.G./ Salazar-Garc D.C./ Blasco R./ Picin A./ Wroe S./ Kullmer O.: To Meat or Not to Meat? New Perspectives on Neanderthal Ecology, –in: Yearbook of Physical Anthropology 156 (2015), S. 43-71.
82. Fitzsimmons K.E./ Hambach U./ Veres D./ Iovita R.: The Campanian Ignimbrite Eruption: New Data on Volcanic Ash Dispersal and Its Potential Impact on Human Evolution, PLoS ONE 8(6): e65839, 2013.
83. Flas D.: Les débuts du paléolithique supérieur dans la Nord-Ouest de L’Europe: le Lincombien-Ranisien-Jerzmanowicien. État de la question, –in: Anthropologica et Praehistoria 113 (2002), S. 25-49.
84. Flas D.: La transition du Paléolithique moyen au supérieur dans la plaine septentrionale de l’Europe. Les problématiques due Lincombien-Ranisien-Jerzmanowicien (2006), –in: Bulletin de la Société préhistorique française, tome 104 N°3, 2007, S. 589-609.
85. Flas D.: Les pointes foliaciées et les changements techniques autour de la transition du Paléolithique moyen au supérieur dans le Nord-Ouest de l’Europe, –in: Toussaint M./ Di Modica K./ Pison St.: Le Paléolithique moyen en Belgique. Bulletin de Chercheurs de la Wallonie, Hors-Série N°4 (2011a), S. 261-276.

6
86. Flas D.: The Middle to Upper Paleolithic transition in Northern Europe: the Lincombian-Ranisian-Jerzmanowician and the issue of acculturation of the last Neanderthals, –in: World Archaeology, Vol. 43, Iss. 4 (2011b), S. 605-627.
87. Flas D.: The Lincombian-Ranisian-Jerzmanowician and the Limits of Aurignacien expansion on the Northern European Plain, –in: (ed.) François Djindjian, Janusz Kozlowski, Nuno Bicho: Le concept de territoires dans le Paléolithique supérieur européen, BAR International Series 1938, 2009, S. 135-142.
88. Flas D.: Les industries à pointes foliacées dans les régions septentrionales de l’Europe, S. 97-122, –in: Otte M. (Ed.): Néandertal/Cromagnon. La Rencontre, Édition errance, Arles (2014), 301 S.
89. Flores J.C.: A mathematical model for Neanderthal extinction, –in: Journal of theoretical Biology (1998), S. 1-9.
90. Floss H.: Steinartefakte - vom Altpaläoloithikum bis in die Neuzeit. Tübingen Publications in Prehistory, Tübingen, Kerns Verlag, 2013.
91. Floss H./ Conard N.J.: L’art mobilier du Jura souabe, S. 201-214, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
92. Freund G.: Die Blattspitzen de Paläolithikums in Europa. Bonn, Ludwig Röhrscheid Verlag, Quartär-Bibliothek 1 (1952).
93. Fu Q./ Li H./ Moorjani P./ Jay F./ Slepchenko S.M./ Bondarev A. A./ Johnson Ph.L.F./ Aximu-Petri
A./ Prüfer K./ Cesare de Filippo,/ Meyer M./ Zwyns N./ Salazar-García D.C./ Kuzmin Y.V./ Keates S.G./ Kosintsev P.A./ Razhev D.I./ Richards M.P./ Peristov N.V./ Lachmann M./ Douka K./ Thomas F./ Higham G./ Slatkin M./ Hublin J.-J./ Reich D./ Kelso J./ Viola T. B./ Pääbo S.: Genome sequence of a 45,000-year-old modern human from western Siberia, –in Nature 2014.
94. Fuhlrott J.C.: Menschliche Überreste aus einer Felsgrotte des Düsselthals. Ein Beitrag zur Frage über die Existenz fossiler Menschen. Verhandlungen des naturhistorischen Veriens der preussischen Rheinlande und Westphalens 16, S. 131-153, Tafel I; s.a.: zusätzliche Medien (N°2).
95. Gábori M.: Les civilisations du Paléolithique moyen entre les Alpes et l’Oural. Esquisse historique. Budapest, Akadémiai Kiadó, (1976).
96. Gamble C.: The Palaeolithic societies of Europe. Cambridge: Cambridge University Press (1999). 97. Gaudzinski-Windheuser S./ Kindler L.: Research perspectives for the study of Neandertal
subsistence strategies based on the analysis of archaeozoological assemblages,–in: Quaternary International 247 (2012), S. 59-68.
98. Giaccio B./ Hajdas I./ Peresani M./ Fedele F.G./ Isaia R.: The Campanian Ignimbrite an its Relevance for the timing of the Middle to Upper Paleolithic shift: timescales and regional correlations, –in: Conard N.: When Neanderthals and Modern Humans met, Kerns Verlag, Tübingen (2006), S. 343-375.
99. Gibbard Ph.L./Head M.J.: The newly-ratified definition of the Quaternary System/Period and redefinition of the Pleistocene Series/Epoch, and comparison of proposals advanced prior to formal ratification, –in: Episodes N°33, Vol. 3 (2010), S. 152-158.
100. Gravina B./ Mellars P./ Ramsey C.B.: Radiocarbon dating of interstratified Neanderthal and early modern Human occupations at the Chatelperronien type-site, –in: Nature, Vol. 438/3 (2005), S. 51-56.
101. Grayson D.K./ Delpech F.: The large mammals of Roc de Combe (Lot, France): The Châtelperronian and Aurignacien assemblages, –in: Journal of Anthropological Archaeology 27 (2008), S. 338-362.
102. Grimaldi S./ Santaniello F.: New insights into Final Mousterian lithic production in western Italy, –in: Quaternary International (2014), S. 116-129.
103. Grousset F.: Les changements abrupts du climat depuis 60.000 ans, –in: Quartenaire, Volume 12, Numéro 4 (2001), S. 203-211.
104. Gunz P./ Simon Neubauer, Bruno Maureille, and Jean-Jacques Hublin: Brain development after birth differs between Neanderthals and modern humans, –in: Current Biology 2010.
105. Hardy BL./ Moncel M-H.: Neanderthal Use of Fish, Mammals, Birds, Starchy Plants and Wood 125-250,000 Years Ago, –in: PLoS ONE 6(8 (2011): e23768.
106. Haesaerts P./ Pirson S.: Paléoenvironnement et chronologie, S. 33-48 –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
107. Heinrich H.: Origin and Consequences of Cyclic Ice Rafting in the Northeast Atlantic Ocean during the Past 130.000 Years, –in: Quaternary Research 29 (1988), S. 142-152.
108. Henry A.G./ Brooks A./ Piperno D.R.: Plant foods an the dietary ecology of Neanderthals and early modern humans, –in: Journal of human evolution 69 (2014), S. 44-54.

7
109. Hemming S.R.: Heinrich Events: Massive late Pleistocene Detritus Layers of the North Atlantic and their Global Climate Imprint, –in: Review of Geophisics 42 (2004), S.1-43.
110. Higham, T./ Brock F./ Peresani M./ Broglio A./ Wood R./ Douka K.: Problems with radiocarbon dating the Middle and Upper Palaeolithic transition in Italy, –in: Quaternary Science Reviews, 28(13–14) (2009), S. 1257–1267.
111. Higham T./Jacobi R./ Julien M./ David F./ Basell L./ Wood R./ Davies W./ Ramsey C.B.: Chronology of the Grotte du Renne (France) and implications for the context of ornaments and human remains within the Châtelperronian, –in: Proceedings, National Academy of Science USA (PNAS), Vol. 107, N° 47 (2010), S. 20234-20239.
112. Higham T./ Compton T./ Stringer Ch./ Jacobi R./ Shapiro B./ Trinkaus E./ Chandler B./ Gröning F./ Collins Ch./ Hillson S./ O’Higgins P./ FitzGerald Ch./ Fagan M.: The earliest evidence for anatomically modern humans in northwestern Europe, –in: Nature 479 (2011), S. 521–524.
113. Higham T./ Douka K./ Wood R. R./ Christopher B./ Brock F./ Basell L./ Camps M./ Arrizabalaga A./ Baena J./ Barroso-Ruiz C./ Bergman C./ Boitard C./ Boscato P./ Caparros M./ Conard N. J./ Draily C./ Froment A./ Galvan B./ Gambassini P./ Garcia-Moreno A./ Grimaldi S./ Haesaerts P./ Holt B./ Iriarte-Chiapusso M.-J./ Jelinek A./ Jorda Pardo J. F./ Maillo-Fernandez J.-M./ Marom A./ Maroto J./ Menendez M./ Metz L./ Morin E./ Moroni, A./ Negrino F./ Panagopoulou E./ Peresani M./ Pirson S./ de la Rasilla M./ Riel-Salvatore J./ Ronchitelli A./ Santamaria D./ Semal P./ Slimak L./ Soler J./ Soler N./ Villaluenga A./ Pinhasi R./ Jacobi R.: The timing and the spatiotemporal patterning of Neanderthal disappearance, –in: Nature Letter, Vol. 512 (2014), S.306-309.
114. Hillebrand J.: Über neuere Funde aus dem ungaländlischen Paläolithikum, –in: Die Eiszeit, 3 (1926), S. 3-5.
115. Hocket B./ Haws J.A: Nutritional ecology and the human demography of Neandertal extinction, –in: Quaternary International 137 (2005), S. 21-34.
116. Hoffecker J.F./ Holliday V.T./ Anikovich M.V./ Sinitsyn A.A./ Popov V.V./ Lisistsyn S.N./ Levkovskaya G.M./ Pospelova G.A./ Forman S.L./ Giaccio B.: From the Bay of Naples to the River Don: the Campanian Ignimbrite eruption and the Middle to Upper Palaeolithic transition in Eastern Europe, –in: Journal of Human Evolution 55 (2008), S. 858-870.
117. Hortalà/Martínez-Navarro: The Quaternary megafaunal extinction and the fate of Neanderthals: An integrative working hypothesis, –in: Quat. Int. 295 (2013), S. 69-72.
118. Hublin J.-J.: The Makers of the Early Upper Paleolithic in Western Eurasia (Chapter 6), S.223-252, –in: Fred H. Smith and James C. M. Ahern. (Eds.), The Origins of Modern Humans: Biology Reconsidered, Second Edition, 2013.
119. Hublin J.-J.: The modern human colonization of western Eurasia: when and where? –in: Quaternary Science Reviews (2014/2015), S. 1-17/ S. 194-210.
120. Hülle W.M.: Die Ilsenburg unter Berg Ranis/Thürngen, 1977, 203 S. 121. Jacobi R.M.: Some Observations on the British Earlier Palaeolithic, –in: DAVIES W. &
CHARLES R. (éd.), Dorothy Garrod and the Progress of the Palaeolithic: Studies in the Prehistoric Archaeology of the Near East and Europe. Oxbow Books. Oxford 1999 : S. 35-40.
122. Jacobi R.M.: A collection of early Upper Palaeolithic artefacts from Beedings, near Pulborough, West Sussex and the context of similar finds from British Iles, –in: Proceedings of the Prehistoric Society, 73 (2007), S. 229-325.
123. Jéquier C.A./ Romandini M./ Peresani M.: Les retouchoirs en matières dures animales : une comparaison entre Moustérien final et Uluzzien, –in: Comptes Rendus Palevol, Volume 11, Issue 4, (2012), S. 283-292.
124. Jiménez-Espejo F.J./ Martínez-Ruiz F./ Finlayson C./ Paytan A./ Sakamoto T./ Ortega-Huertas M./ Finlayson G./ Iijima K./ Gallego-Torres D./ Fa D.: Climate forcing and Neanderthal extinction in Southern Iberia: insights from a multiproxy marine record, –in: Quaternary Science Reviews, Volume 26, Issues 7–8, April 2007, Pages 836-852.
125. Karavanić I./ Smith, F. H.: More on the Neanderthal problem: the Vindija Case. Current Anthro- pology 41 (2000), S. 838–840.
126. Klein R.: "Anatomy, behavior, and modern human origins". Journal of World Prehistory 9 (1995): S. 167–198.
127. Knecht H.: Design Strategiesof Early Upper Paleolithic Bone and Antler ProjectileTechnologie, –in: Anthropologie et Préhistoire 111 (2000), S. 28-36.
128. Krause J./, Ludovic Orlando, David Serre, Bence Viola, Kay Pru fer, Michael P. Richards, Jean-Jacques Hublin, Catherine Ha nni, Anatoly P. Derevianko & Svante Pa a bo: Neanderthals in central Asia and Siberia, –in: Nature (2007), S.902-904.
129. Kuckenburg M.: Der Neandertaler – auf den Spuren des ersten Europäers'. Stuttgart, Klett-Cotta, 2005, 339 S.

8
130. Koumouzelis M./ Ginter B./ Kozłowski J.K./ Pawlikowski M./ Bar-Yosef O./ Albert R.M./ Litynska-Zajac M./ Stworzewicz E./ Wojtal P./ Lipecki G./ Tomek T./ Bochen_ski Z.M./ Pazdur A.: The Early Upper Palaeolithic in Greece: the excavations in Klissoura cave. J. Archaeol. Sci. 28 (2001), S. 515-539.
131. Kuhn S.L./ Pigati, J./ Karkanas P./ Koumouzelis M./ Kozłowski J.K./ Ntinou M./ Stiner M.C.: Radiocarbon dating results for the Early Upper Paleolithic of Klissoura Cave 1. Eurasian Prehist. 7 (2010), S. 37-46.
132. Kaminská/ L'ubomíra/ Kozlowski J.K./ Škrdla P.: New Approach to the Szeletian - Chronology and Cultural Variability, –in: Eurasian Prehistory 8 1/2 (2011).
133. Kozłowski J.: Les origines, S. 73-94, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
134. Kozłowski J.K.: La fin du Paléolithique moyen en Pologne, Anthropologie, t. 27, n° 2-3 (1989), S. 133-142.
135. Kozłowski J.K.: Certains aspects techno-morphologiques des pointes foliacées de la fin du Paléolithique moyen et du début du Paléolithique supérieur en Europe centrale, –in: C. Farizy (dir.): Paléolithique moyen récent et Paléolithique supérieur ancien en Europe, Mémoires du Musée de Préhistoire d’Île-de-France, 3, APRAIF, Nemours (1990), S. 125-133.
136. Kozłowski J.K.: L’Aurignacien en Europe et au Proche-Orient, –in: Bánesz L. et Kozłowski J.K. (Eds.), Aurignacien en Europe et au Proche-Orient. Bratislava, Institut Archéologique de l’Académie Slovaque des Sciences (1993), S. 283-291.
137. Kozłowski J.K.: La signification des "outils foliacés". Paléo, supplément 1 (1995), S. 91-99. 138. Kozłowski J.K./Kozłowski S.K.: Le Paléolithique en Pologne, –in: Préhistoire d’Europe 2,
Grenoble, Jérôme Millon, (1996). 139. Kozłowski J. K./ Otte M.: The formation of the Aurignacian in Europe, –in: Journal of
Anthropological Research 56 (2000), S. 513–534. 140. Kuhn S.L./Stiner M.C.: The antiquity of hunter-gatherers, –in: Panter-Brick C, Layton R H,
Rowley-Conwy P A, eds. Another Day, Another Camp: An Interdisciplinary View of Hunter-gatherers. Cambridge: Cambridge University Press, 2001, S. 99-142.
141. Le Brun-Ricalens F.: Retuschierte Lamellen im Aurignacien: Dufour et alii, S. 357-366, –in: Floss H. (Ed.): Steinartefakte vom Altpaläolithikum bis in die Neuzeit, Tübingen Publications in Prehistory, Kerns Verlag Tübingen, 2. Edition, 2013, 986 S.
142. Lev E./ Kislev M.E./ Bar-Yosef O.: Mousterian vegetal food in Kebara Cave, Mt. Carmel. J.: –in: Archaeological Science 32 (2005), S.475–484.
143. Liolios D.: Les instruments osseux, S. 137-152, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
144. Litt Th./ Behre K.-E./ Meyer K.-D./ Stephan H.-J. & Wansa St.: Stratigraphische Begriffe für das Quartär des norddeutschen Vereisungsgebietes, –in: Eiszeitalter und Gegenwart Quaternary Science Journal 56/1-2 (2007), S. 7-65.
145. Longo L./ Boaretto E./ Caramelli D./ Giunti P./ Lari M./ Milani L./ Mannino M. A./ Sala B./ Thun Hohenstein U./ Condemi S.: Did Neandertals and anatomically modern humans coexist in northern Italy during the late MIS 3? –in: Quaternary International 259 (2012), S. 102-112.
146. Lowe, J./ Barton N./ Blockley S./ Ramsey C.B./ Cullen V.L./ Davies W./ Gamble C.S./ Grant K./ Hardiman M./ Housley R./ Lane C.S./ Lee S./ Lewis M./ Macleod A./ Menzies/ M. Müller W./ Pollard M./ Price C./ Roberts A.P./ Rohling E.J./ Satow C./ Smith V.C./ Stringer C.B./ Tomlinson E.L./ White D./ Albert P./ Arienzo I./ Barker G./ Boric D./ Carandente A./ Civetta L./ Ferrier C./ Guadelli J.- L./ Karkanas P./ Koumouzelis M./ Müller U.C./ Orsi G./ Pross J./ Rosi M./ Sha- lamanov-Korobar/ L./ Sirakov N./ Tzedakis P.C.: Volcanic ash layers illuminate the resilience of Neanderthals and early modern humans to natural hazards. Proc. Natl. Acad. Sci. U. S. A. 109 (2012), 1-6.
147. Lucas G./ Rigaud J.-P. Simek/ Soressi J.F.: The Châtelperronian of Grotte XVI, Cénac-et-Saints-Julien (Dordogne, France), –in: Zilhão J./ d’Errico F. (Eds.), The chronology of the Aurignacien and of the Transitional technocomplexes. Dating, stratigraphies, cultural implications. Instituto Português de Arquelogia, Lisboa, 2003, S.289-298.
148. Marean C.W./ Bar-Matthews M./ Bernatchez J./ Fisher E./ Goldberg P./ Herries A.I.R./ Jacobs Z./ Jerardino A./ Karkanas P./ Minichillo T./ Nilssen P.J./ Thompson E./ Watts I./ Williams H.M.: Early human use of marine resources and pigment in South Africa during the Middle Pleistocene, –in: Nature 449 (2007), S. 905-908.
149. Martin H.: Nouvelle série de débris humains disséminés, trouvés en 1913, dans la Gisement Moustérien de La Quina, –in: Bulletin de la Société préhistorique de France, tome 10, N°10 (1913), S. 540-543.
150. Marcott S.A./ Clark P.U./ Padman L./ Klinkhammer G.P./ Springer S.R./ Liu Z./ Otto-Bliesner B.L./ Carlson A.E./ Ungerer A./ Padman J./ He F./ Cheng J./ Schmittner A.: Ice-shelf collapse from

9
subsurface warming as a trigger for Heinrich events, –in: PNAS Vol.108, N°33 (2011), S. 13415-13419.
151. McBrearty S./ Brooks A.: "The revolution that wasn’t: a new interpretation of the origin of modern human behavior". Journal of Human Evolution 39 (2000): S. 453–563.
152. Mester Z.: Le Szélétien, S. 149-188, –in: Otte M. (Ed.): Néandertal/Cromagnon. La Rencontre, Édition errance, Arles (2014), 301 S.
153. Mellars P.: The character of the Middle-Upper Palaeolithic transition in south-west France., –in: Renfrew AC, editor. The explanation of culture change: models in prehistory. London: Duckworth (1973), S. 255–276.
154. Mellars P.: Major Issues in the emergence of modern humans, –in: Current Anthropology 30 (1989), S. 349-385.
155. Mellars P.: The Neanderthal legacy: an archaeological perspective from Western Europe. Princeton: Princeton University Press (1996).
156. Mellars, P.: The Neanderthal Problem Continued (mit Kommentaren). Current Anthropology 40 (1999), S. 341–364.
157. Mellars P.: Reindeer specialization in the early Upper Palaeolithic: the evidence from southwest France, –in: J. Archaeol. Sci. 31 (2004a), S.613–617.
158. Mellars P.: Neanderthals and modern human colonization of Europe, –in: Nature review article (2004b), S. 461-465.
159. Mellars P.:The impossible coincidence: A single species model for the origins of modern human behavior in Europe, –in: Evol. Anthropol. 14 (2005), S. 12–27.
160. Mellars P.: A new radiocarbon revolution and the dispersal of modern humans in Eurasia, –Nature Reviews (2006), S. 931-935.
161. Minami M./ Yamazaki K./ Omori T./ Nakumara T.: Radiocarbon dating of VIRI bone samples using ultrafiltration, –in: Nuclear Instruments and Methods in Physics Research B 294 (2013), S. 240–245.
162. Morin E.: Reassessing Paleolithic Subsistence - The Neandertal and Modern Human Foragers of Saint-Césaire. New York Cambridge University Press (2012).
163. Morin E./ Tsanova T./ Sirakov N./ Rendu W./ Mallye J.-B./ Lévêque F.: Bone refits in stratified deposits: testing the chronological grain at Saint-Césaire, –in: Journal of Archaological Science 32 (2005), S. 1083-1098.
164. Moroni A./ Boscato P./ Ronchitelli A.: What roots for the Uluzzian? Modern behaviour in Central-Southern Italy and hypotheses on AMH dispersal routes, –in: Quaternary International 316 (2013), S. 27-44.
165. Neruda P./ Nerudová Z.: The development of the production of lithic industry in the Early Upper Palaeolithic of Moravia, –in: Archeologické rozhledy LVII (2005), S. 263-292.
166. Neruda P./ Nerudová Z.: The Middle-Upper Paleolithic transition in Moravia in the context of the Middle Danube region, –in: Quaternary International 294 (2013), S. 3-19.
167. Nigst Ph.R./ Bence Viola Th.B./ Haesaerts P./ Blockley S./ Damblon F./ Frank Ch./ Fuchs M./ Götzinger M./ Hambach U./ Mallol C./ Moreau L./ Niven L./ Richards M./ Richter D./ Zöller L./ Trnka G./Hublin J.-J.: New Research on the Aurignacian of Central Europe: A first Note on the 2006 Fieldwork at Willendorf II, –in: Quartär 55 (2008), S. 9-15.
168. Nigst Ph.R.: First Modern Human Occupation of Europe: The Middle Danube Region as a Case Study, 2014, S. 35-47.
169. Noiret P.: Habitations et territoires, S. 181-200, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
170. Normand Ch./ O’Farrell M./ Rios Garaisar J.: Quelles(s) utilisations(s) pour les productions lamellaires de l'Aurignacien archaïque? Quelques données et réflexions à partir des exemplaires se la grotte d'Isturitz ( Pyrénées Atlantiques ; France). Paléthnologie, 1 (2008), S. 7-46.
171. O’Connor S./ Ono R./ Clarkson C.: Pelagic fishing at 42,000 years before the present and the maritime skills of modern humans, –in: Science 334 (2011), S. 1117-1121.
172. Otte M.: Le Paléolithique supérieur ancien en Belgique, Monographies d’archéologie nationale, 5, Musée royaux d’Art et d’Histoire, Bruxelles (1979), 684 S.
173. Otte M.: Les industries aux pointes foliacées du Nord-Ouest européen (1990), –in: Kozłowski J.K. (éd.), Feuilles de pierre. Les industries à pointes foliacées du Paléolithique supérieur européen. Actes du collo- que de Cracovie (1989), ERAUL 42, Liège, S. 247- 269.
174. Otte M.: Le Paléolithique inférieur et moyen en Europe, Paris, Amand Colin, (1996). 175. Otte M.: Le bouleversement de l’humanité en Eurasie vers 40.000 ans. In: E. Carbonell und
M. Vaquero (Hrsg.), The Last Neandertals, the First Anatomically Modern Humans: A Tale About the Human Diver- sity. Cultural Change and Human Evolution: the Crisis at 40 ka BP. Tarragona: Universitat Rovira i Virgili (1996), S. 95–106.
176. Pelegrin J.: Technologie lithique : le Châtelperronien de Roc de Combe (Lot) et de La Côte

10
(Dordogne). Paris, CNRS (1995), 297 S. (Cahiers du Quaternaire ; 20). 177. Pelegrin J./ Soressi M.: Le Châtelperronien et ses rapports avec le Moustérien, –in: Les
Néandertaliens. Biologie et cultures, Paris, Éditions du CTHS (2007), Documents préhistoriques; 23), S. 283-296.
178. Peresani M.: Fifty thousand years of flint knapping and tool shaping across the Mousterian and Uluzzian sequence of Fumane cave: –in: Quaternary International 247 (2012), S. 125-150.
179. Peresani M.: L’Uluzzien en Italie, S. 61-80, –in: Otte M. (Ed.): Néandertal/Cromagnon. La Rencontre, Édition errance, Arles (2014), 301 S.
180. Pettitt P.B.: Disappearing from the world: an archaeological perspective on Neanderthal extinction, –in: Oxford Journal of Archaeology, 18 (3) (1999), S. 217-240.
181. Peyrony D.: Une mise au point au sujet de l’industrie de l’abri Audit et de celle de Châtelperron. Bulletin de la Société préhistorique française, 45 (1948), S. 34-35.
182. Radovčić D./ Sršen A.O./ Radovčić J./ Frayer D.W.: Evidence for Neandertal Jewelry: Modified White-Tailed Eagle Claws at Krapina, –in: PLoS ONE 10(3) (2015): e0119802.
183. Richter, D. :Advantages and limitations of thermoluminescence dating of heated flint from Paleolithic sites, –in: Geoarchaeology, 22 (2007), S. 671–683.
184. Riel-Salvatore J.: What Is a ‘Transitional’ Industry? The Uluzzian of Southern Italy as a Case Study, S. 377-396, –in: M. Camps, P. Chauhan (eds.), Sourcebook of Paleolithic Transitions, 2009.
185. Riel-Salvatore J.: Mad Neanderthal disease? Some comments on “A potential role for Transmissible Spongiform Encephalopathies in Neanderthal extinction”, –in: Medical Hypotheses, Volume 71, Issue 3, September (2008), S. 473-474.
186. Riel-Salvatore J./ Ludeke I./ Negrino F./ Holt B.M.: A spatial analyses of the late Mousterian levels of Riparo Bombrini (Balzi Rossi, Italy), –in: Canadian Journal of Archaeology /Journal Canadien d’Archéologie 37 (2013): S. 70-92.
187. Pradel L. (Dr.): Le Moustérien, –in: Bulletin de la Société préhistorique française, tome 51, N° 8 (1954), S. 35-43.
188. Pradel L. (Dr.): La pointe des Cottés, –in: Bulletin de la Société préhistorique de France. 1963, tome 60, N. 9-10 (1963), S. 582-590.
189. Pradel L.: La pointe de Font-Yves. In: Bulletin de la Société préhistorique française. 1978, tome 75, N. 8, S. 233-236.
190. Prošek F.: Szeletien na Slovensku (Le Szélétien en Slovaquie), –in: Slovenská Archeologia, 1 (1953), S. 133-194.
191. Pyle D.M./ Ricketts G.D./ Margari V./ van Andel T.H./ Sinnitsyn A.A./ Praslov N.D./ Lisitsyn S.: Wide dispersal and deposition of the distal tephra during the Pleistocene ‘Campanian Ignimbrite/Y5‘ eruption, Italy, –in: Quaternary Science Reviews 25 (2006), S. 2713-2728.
192. Rahmstarf S.: Ocean circulation and climate during the past 120,000 years, –in: Nature, insight review Articles (2002), S. 207.214.
193. Ready E.: Neandertal man the hunter: a history of Nean- dertal subsistence. Explor Anthropol 10 (2010):58–80.
194. Ready E.: Neandertal foraging during the late Mousterian in the Pyrenees: new insights based on faunal remains from Gatzarria Cave; –in: Journal of Archaeological Science 40 (2013), S. 1568-1578.
195. Richards MP, Schmitz R.: Isotope evidence for the diet of the Neanderthal type specimen. Antiquity 82 (2008), S. 553–559.
196. Richards M.P/ Erik Trinkaus: Isotopic evidence for the diets of European Neanderthals and early modern humans, –in: PNAS (2009), S. 16034-16039.
197. Richter D./ Tostevin G./ Škrdla P.: Bohunician technology and thermoluminiscence dating the type locality of Brno-Bohunice (Czech Republic): comparision of TL, OSL, IRSL and 14C dating results, –in: Journal of Archeological Science Reviews, 36 (2009), S. 708-720.
198. Ringer Á.: Bábinonien. Eine mittelpaläolithische Blattwerkzeugindustrie in Nordostungarn. Dissertation Archaeologicae Ser. II. N° 11, Budapest, Eötvös Loránd Tudománygetem Régészeti Intézete, (1983).
199. Ringer Á.: Nouvells données sur le Szélétien de Bükk, –in: Praehistoria, 9-10 (2008-2009), S. 21-34.
200. Rios-Garaizar J./ Arrizabalaga A./ Villaluenga A.: Haltes de chasse du Châtelperronien de la Péninsule Ibérique : Labeko Koba et Ekain (Pays Basque Péninsulaire), –in: L’anthropologie 116 (2012), S. 532-549.
201. Roche D./ Paillard D./ Cortijo E.: Constraints on the duration and freshwater release of Heinrich event 4 trough isotope modelling, –in: Nature, letters to Nature (2004), S. 379-382.
202. Rodríguez-Vidal J./ d’Errico F./ Pacheco F.G./ Blasco R./ Rosell J./ Jennings R.P./ Queffelec A./ Finlayson G./ Fa D.A./ Gutiérrez López J.M./ Carrión J.S./ Negro J.J./ Finlayson S./ Cáceres

11
L.M./ Bernal M.A./ Fernández Jiménez S./ Finlayson C.: A rock engraving made by Neanderthals in Gibraltar, –in: Proc. Natl. Acad. Sci. 111 (2014), S. 13301–13306.
203. Roebroeks W.: Time for the middle to upper Palaeolithic Transition, –in: Journal of Human Evolution 55 (2008), S. 918-926.
204. Romagnoli F.: A second life: Recycling production waste during the Middle Palaeolithic in layer L at Grotta del Cavallo (Lecce, Southeast Italy), –in: Quat. Int. 361 (2015), S. 200-211.
205. Rosell J./ Cáceras I./ Blasco R./ Bennàsar M./ Bravo P./ Campeny G./ Esteban-Nadal M./ Fernández-Laso M.C./ Gabucio M.J. Gabucio/ Huguet R./ Ibáñez N./ Martín P./ Rivals F./ Rodríguez-Hidalgo A./ Saladié P.: A zooachaeological contribution to establish occupational patterns in Level J of Abric Romaní (Barcelona, Spain), –in: Quaternary International 247 (2012), S. 69-84.
206. Roussel M./Soressi M.: Le Châtelperronien, S.31-60, –in: Otte M. (Ed.): Néandertal/Cromagnon. La Rencontre, Édition errance, Arles (2014), 301 S.
207. Sañudo P./ Vallverdú-Poch J./ Canals A.: Spatial Patterns in Level J, –in: Ed. Carbonell i Roura E.: High Resolution Archaeology and Neanderthal Behavior, Time and Space in Level J of Abric Romaní (Capellades, Spain), VertebratePaleobiology and Paleoanthropology Series, Springer, Dordrecht, Heidelberg, New York, London (2012), S. 47-76.
208. Scandone R.F./ Bellucci L./ Rolandi G.: The structure of the campanian Plain and the activity of the neapolitan volcanos (Italy), Journal of Volcanology and Geothermal Research 48 (1991), S. 1-31.
209. Seguin-Orlando A., Thorfinn S. Korneliussen, Martin Sikora, Anna-Sapfo Malaspinas,
Andrea Manica, Ida Moltke, Anders Albrechtsen, Amy Ko, Ashot Margaryan, Vyacheslav Moiseyev, Ted Goebel, Michael Westaway, David Lambert, Valeri Khartanovich, Jeffrey D. Wall,
Philip R. Nigst, Robert A. Foley, Marta Mirazon Lahr, Rasmus Nielsen, Ludovic Orlando, Eske Willerslev: Genomic structure in Europeans dating back at least 36,200 years, 2014.
210. Schintler A.: La frontière entre le Châtelperronien et l’Uluzzien : analyse comparative des typologies lithiques sur la base des dernières découvertes, –in: L'Anthropologie, Volume 115, Issue 5 (2011), S. 585-599.
211. Schmitz R.W.: Die Entwicklungsgeschichte des Menschen im Weltbild vergangener Jahrhunderte, S. 111-116. –in: Roots – Wurzeln der Menschheit. Mainz, Rheinisches Landesmuseum Bonn, 2006.
212. Schmitz R.W.: Interdisziplinäre Untersuchungen an den Neufunden aus dem Neandertal – Johann Carl Fuhlrott (1803 - 1877) gewidmet, –in: Mitteilungen der Gesellschaft für Urgeschichte 12 (2003), S. 25-45.
213. Semal P./ Jungels C./ Di Modica K./ Flas D./ Hauzeur A./ Toussaint M./ Pirson S./ Khlopachev G./ Pesesse D./ Tartar E./ Crevecoeur I./ Rougier H./ Maureille B.: La grotte de Spy (Jemeppe-sur-Sambre ; prov. Namur), –in: Bulletin des Chercheurs de la Wallonie, hors-série no 4 (2011), S. 305-321
214. Shipman P.: Do the eyes have it?, –in: American scientist 100 (3) (2012): 198. 215. Sinitsyn A.A./Hoffecker J.F.: Radiocarbon dating and chronology of the Early Upper
Paleolithic at Kostenki, –in: Quaternary International, Vol. 152 & 153 (2006), S. 164-174. 216. Škrdla P.: The Bohunician in Moravia and Adjoining Regions, –in: Archaeology, Ethnology
and Anthropology of Eurasia, Volume 41, Issue 3 (2013), S. 2-13. 217. Škrdla P.: Moravian Bohunician, S. 123-148, –in: Otte M. (Ed.): Néandertal/Cromagnon. La
Rencontre, Édition errance, Arles (2014), 301 S. 218. Škrdla P.: Bohunician Technology: A Refitting Approach. In Stránská skála. Origins of the
Upper Paleolithic in the Brno Basin, Moravia, Czech Republic, –in: (ed.) J. A. Svoboda and O. Bar-Yosef, (2003), S. 119–51, American School of Prehistoric Research Bulletin 47. Cambridge: Peabody Museum of Anthropology and Ethnology, Havard University.
219. Škrdla P./ Tostevin G.: Brno-Bohunice. Analýza materiálu z výzkumu v roce 2002, Přehled výzkumů, 50 (2005), S. 35-61.
220. Smith FH, Trinkaus E, Pettitt PB, Karavanic I, Paunovic M.: Direct radiocarbon dates for Vindija G1 and Velika Pecina Late Pleistocene hominid remains. Proc Natl Acad Sci USA 96 (1999), S. 12281–12286.
221. Soressi M.: Le Moustérien de tradition acheuléenne du sud-ouest de la France. Discussion sur la signification du faciès à partir de l’étude comparée de quatre sites : Pech de l’Azé I, Le Moustier, La Rochette et la Grotte XVI. Thèse, Université Bordeaux I (2002), 399 S.
222. Soressi M.: Die Steintechnologie des Spätmoustérien. Ihre Bedeutung für die Entstehungsgeschwindigkeit modernen Verhaltens und die Beziehung zwischen modernem Verhalten und biologischer Modernität, –in: Mitteilungen der Gesellschaft für Urgeschichte, 13 (2004), S. 9-28.

12
223. Soressi M. & Locht J.-L.: Les armes de chasse de neandertal. Première analyse des pointes moustériennes d'Angé. Archéopages, 27 (2010), S. 6-11.
224. Soressi M./ Roussel M. (2014). European Middle to Upper Paleolithic Transitional Industries: Châtelperronian, –in Encyclopedia of Global Archaeology, Springer New York (2014), S.2679-2693.
225. Soulier M.-C./ Mallye J.-B..: Hominid subsistence strategies in the South-West of France: A new look at the early Upper Palaeolithic faunal material from Roc-de-Combe (Lot, France), –in: Quaternary International, Volume 252, 27 February 2012, S. 99-108.
226. Sørensen B.: Demography and the extinction of European Neanderthals, –in: Journal of Anthropological Archaeology, Volume 30, Issue 1 (2011), S. 17-29.
227. Sparks S./ Self S./ Grattan J./ Oppenheimer C./ Pyle D./ Rymer H.: Super-eruptions: Global effects and future threats, report of a Geological Society of London Working Group, The Geol. Soc., London, 2005. [Verfügbar unter: http://www.geolsoc.org.uk/gsl/education/page2965.html.]
228. Spencer M.A./ Demes B.: Biomechanical Analysis of Masticatory System Configuration in Neandertals and Inuits, –in: American Journal of Physical Anthropology 91 (1993): S. 1-20.
229. Starkovich B.M: Optimal foraging, dietary change, and site use during the Paleolithic at Klissoura Cave 1 (southern Greece), –in: Journal of Arch. Sc. 52 (2014), S. 39-55.
230. Stieglitz J.L./ Schmidt M.W./ Henry L.G./ Curry W.B./ Skinner L.C./ Mulitza S./ Zhang R./ Chang P.: Muted change in Atlantic overturning circulation over some glacial-aged Heinrich events, –in: Nature Geoscience Vol. 7 (2014), S. 144-150.
231. Stodiek U./ Paulsen H.: „Mit Pfeil und Bogen…“ Technik der steinzeitlichen Jagd, Isensee Verlag, Oldenburg, 1996.
232. Størmer F.C./ Mysterud I.: Cave smoke: Air pollution poisoning involved in Neanderthal extinction?, –in: Medical Hypotheses, Volume 68, Issue 3, 2007, S. 723-724.
233. Straus L. G.: Continuity or rupture; convergence or invasion; adaptation or catastrophe; mosaic or monolith: views on the Middle to Upper Paleolithic transition in Iberia, –in: E. Carbonell und M. Vaquero (Hrsg.), The Last Neandertals, the First Anatomically Modern Humans: A Tale About the Human Diver- sity. Cultural Change and Human Evolution: the Crisis at 40 ka BP. Tarragona: Universitat Rovira i Virgili, 1996, S. 203–218.
234. Stringer Ch.: Out of Ethiopia, –in: Nature News and views, Vol. 423 (2003), S. 692-695. 235. Stringer Ch./ Pälike H./ van Andel T.H./ Huntley B./ Valdes P./ Allen J.R.M.: Climatic Stress
and the Extinction of the Neanderthals, –in: Neanderthals and Modern Humans in the European Landscape during the Last Deglaciation (eds van Andel, T. H. & Davies, W.), McDonald Institute Monographs, Cambridge (2003), S. 233-240
236. Stringer C.B./ Finlayson J.C./ Barton R.N.E./ Fernández-Jalvo Y./ Cáceres I./ Sabinh R.C./ Rhodes E.J./ Currant A.P./ Rodríguez- Vidal J./ Giles-Pacheco F./ Riquelme-Cantal J.A.: Neanderthal exploitation of marine mammals in Gibraltar. Proc Natl. Acad. Sci. USA 105 (2008) S.14319-14324.
237. Svoboda J.: Stánská skála. Výsledky výzkumu v letech 1985-87, –in: Památky archeologické, 82 (1991), S. 5-47.
238. Svoboda J.: The Neandertal extinction in eastern Central Europe, –in: Quaternary International 137 (2005): S. 69-75.
239. Tagliacozzo A./ Romandini M./ Fiore I./ Gala M./ Peresani M.: Animal Exploitation Strategies during the Uluzzian at Grotta di Fumane (Verona, Italy), 2011.
240. Teyssandier N.: Revolution or evolution: the emergence of the Upper Paleolithic in Europe. World Archaeol. 40 (2008), S. 493-519.
241. Totevin G.B.: Social Intimacy, Artefact Visibility and Acculturation Models of Neanderthal–Modern Human Interaction, –in: Rethinking the Human Revolution: New Behavioural and Biological Perspectives on the Origins and Dispersal of Modern Humans. P. Mellars, K. Boyle, O. Bar-Yosef, & C. Stringer (eds.). MacDonald Institute for Archaeological Research Monographs, Cambridge University, UK 2007, S. 341-357.
242. Tostevin G.B./ Škrdla P.: New excavations at Bohunice and the question of Uniqueness of the Type-site for the Bohunicien Industrial Type, –in: Anthropologie, 44 (2006), Brno, S. 31-48.
243. Trinkaus E.: Modern human versus Neandertal evolutionary distinctiveness, –in: Curr. Anthropol. 47 (2006), 597-620.
244. Underdown S.: A potential role for Transmissible Spongiform Encephalopathies in Neanderthal extinction, –in: Medical Hypotheses, Volume 71, Issue 1 (2008), S. 4-7.
245. Uzquiano P./ Yravedra J./ Zapata B.R./ Garcia M.J.G./ Sesé C./ Baena J.: Human bihaviour and adaptations to MIS 3 environmental trends (>53-30 ka BP) at Esquilleu cave (Cantabria, northern Spain), –in: Quaternary International 252 (2012), S. 82-89.
246. Valoch K.: Verdovice, eine Siedlung des Szeletien in Südmähren (19(?)).

13
247. Valoch K.: Die altsteinzeitliche Fundstelle in Brno-Bohunice, –in: Studie Archeologického Ústavu Československé Akademie Věd v Brně ; roč. 4, sv. 1, Praha: Academia (1976), 120 S.
248. Valoch K.: La Moravie il y a 40 000 ans. In: C. Farizy (Hrsg.), Paléolithique moyen récent et Paléoli- thique supérieur ancien en Europe. Ruptures et transitions: examen critique des documents archéologi- ques. Mémoires du Musée de Préhistoire d’Ile-de-France 3. Nemours: Ed. A.P.R.A.I.F., 1990, S. 115–124.
249. Valoch K.: Le Paléolithique en Tchéquie et en Slovaquie, –in: Préhistoire de l’Europe 3, Grenoble, Jérôme Millon, (1996).
250. Valoch K./ Kočí A./ Mook W.G./ Opravile E./ Van der Plicht J./ Smolíková L./ Weber Z.: Vedrovive V, eine Siedlung des Szeletien in Südmähren, –in: Quartär, 43/44 (1993), S. 7-93.
251. Valoch K./ Nerudová Z./ Neruda P.: Stránská skála III-1. Ateliers des Bohunicien, –in: Památky archeologické, 41 (2000), S. 5-13.
252. Valoch K./ Neruda P./ Nerudová Z.: The Bohunician Technology from Stránská skála Open-air Site (the Czech Republic), –in: Jan Michał Burdukiewicz, Krysztof Cyrek, Piotr Dyczek, Karol Szymczak (Eds.): Understanding the past. Papers offered to Stefan K. Kozłowski, 2009.
253. Vallverdú-Poch J./ Gómez de Soler B./ Vaquero M./ Bischoff J.L.: The Abric Romaní Site and the Capellades Region (Chapter 2), –in: Ed. Carbonell i Roura E.: High Resolution Archaeology and Neanderthal Behavior, Time and Space in Level J of Abric Romaní (Capellades, Spain), VertebratePaleobiology and Paleoanthropology Series, Springer Dordrecht, Heidelberg, New York, London, 2012, S. 19-46.
254. van Andel T.H./ Davies W. (eds.): Neanderthals and modern humans in the European landscape during the last glaciation: archaeological results of the Stage 3 Project, –in: Cambridge, UK, McDonald Institute for Archaeological Research monographs (McDonald Institute monographs) (2003), 265 S.
255. Vanhaeren M.: La parure, S. 253-270, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
256. Vaquero M., María Gema Chacón, María Dolores García-Antón, Bruno Gómez de Soler, Kenneth Martínez, Felipe Cuartero: Time and space in the formation of lithic assemblages: The example of Abric Romaní Level J, –in: Quart. Int. 247 (2012), S. 162-181.
257. Vaquero M., Amelia Bargallo, María Gema Chacon , Francesca Romagnoli, Pablo Sanudo: Lithic recycling in a Middle Paleolithic expedient context: Evidence from the Abric Romaní (Capellades, Spain), –in: Quaternary Int. 361 (2015), 212-228.
258. Vercoutère C./Patou-Mathis M.: L’animal comme une ressource alimentaire…pas seulement S. 153-180, –in: Otte M. (Ed.): Civilisations et Cultures, Les Aurignaciens. Édition errance, Paris, 2010, 300 S.
259. Vertés L.: Tata, Eine mittelpaläolithische Travertin-Siedlung in Ungarn. Budapest, Akadémiai Kiadó, A Magyar Régézet Kéziköyve 1 (1964), 284 S.
260. Vigliotti L.: Magnetic properties of the Campanian Ignimbrite and the marine Y5 tephra layer, –in: Geological Society 396, London, Special Publications, 2014.
261. Veiga-Pires C.C./ Hillaire-Marcel C.: U and Th isotope constraints on the duration of Heinrich events H0-H4 in the southern Labrador Sea, –in: Paleoceanography, Volume 14, N°2, 1999, S. 187-199.
262. Weißmüller W.: Drei Fundstellen mit Blattformen aus dem südostbayerischen Donauraum. Ein Beitrag zur Westausbreitung des Szeletien, –in: Quartär 45/46 (1995), S. 99-134.
263. Wood R.E./ Barroso-Ruíz C./ Caparrós M./ Pardo J.F.J./ Santos B.G./ Higham T.F.G.: Radiocarbon dating casts doubt on the late chronology of the Middle to Upper Palaeolithic transition in southern Iberia, –in: PNAS 110 (8) (2013), S. 2781-2786.
264. Walker M./ Johnsen S./ Rasmussen S. O./ Popp T./ Steffensen J.-P./ Gibbard P./ Hoek W./ Lowe J./ Andrews J./ Björck S./ Cwynar L. C./ Hughen K./ Kershaw P./ Kromer B./ Litt T./ Lowe D. J./ Nakagawa T./ Newnham R. and Schwander J.: Formal definition and dating of the GSSP (Global Stratotype Section and Point) for the base of the Holocene using the Greenland NGRIP ice core, and selected auxiliary records, –in: Journal of Quaternary Science 24 (2009), S. 3-17.
265. Yravedra J/ Cobo-Sánchez L.: Neanderthal exploitation of ibex and chamois in southwestern Europe, –in: J. Hum. Evol. 78 (2015), S. 12-32.
266. Yravedra J./ Domínguez-Rodrigo M.: The shaft-based methodological approach of the quantification of long limb bones and its relevance to understanding hominid subsistence in the Pleistocene: application to four Palaeolithic sites, –in: Journal of Quaternary Science 24 (2009), S. 85-96.
267. Yravedra J./ Gómez-Castenado A.: Taphonomic implications for the late Mousterian of South-West Europe at Esquilleu Cave (Spain), –in: Quaternary International 337 (2014), S. 225-236.
268. Zilhão J./ d’Errico F.: The Chronology and Taphonomy of the Earliest Aurignacian and Its Implica- tions for the Understanding of Neandertal Extinction. Journal of World Prehistory 13

14
(1999), S. 1-68. 269. Zilhão, J./ D’Errico, F.: The Chronology of the Aurignacian and of the Transitional
Technocomplexes. Dating, Stratigraphies, Cultural Implications. Instituto Portugues de Arqueologia, Lisbon (2003).
270. Zilhão, J./ Trinkaus, E./ Constantin S./ Milota, S./ Gherase M./ Saecina L./ Rougier H./ Quilès, J./ Rodrigo R.: The Pestera cu Oase people, Europe’s earliest modern humans, –in: Mellars, P., Boyle, K., Bar-Yosef, O., Stringer, C. (Eds.), Rethinking the Human Revolution, McDonald Institute for Archaeological Research, Cambridge University Press, Cambridge (2007), S. 249-262.
Zusätzliche Medien:
1. http://www.neanderthal.de/museum-tal/im-tal/wildgehege/naturschutzverein/print.html 2. http://www.naturhistorischerverein.de/neandertaler_ebook.pdf (Menschliche Überreste aus einer
Felsgrotte des Düsselthals. Ein Beitrag zur Frage übder die Existenz fossiler Menschen. Von Dr. C. Fuhlrott)
3. http://www.ccel.org/ccel/lightfoot 4. http://quaternary.stratigraphy.org/images/TablaCuaternarioIbéricov_209.pdf 5. http://www.iceandclimate.nbi.ku.dk/research/drill_analysing/history_drilling/central_ice_cores/ 6. http://www.google.de/url?sa=t&rct=j&q=&esrc=s&source=web&cd=3&ved=0CC4QFjAC&url=http%
3A%2F%2Fwww.uni-graz.at%2F~foelsche%2FVO%2FKlima%2FKlima_09_Abrupte_Klimaaenderungen.ppt&ei=3zHSVKPnEoa7ygOl9oHIAQ&usg=AFQjCNHfJwqC4JeBPEL7N8VirV7OcyzCSA&sig2=cZPk3Kw0WtFybR425ZG74A (Dansgaard-Oeschger Zyklen)
7. http://www.iphes.cat/abric-roman%C3%AD oder http://www.iphes.cat/print/node/32 8. http://altsteinzeit-hessen.de/?p=269 9. http://www.palaeon.de/schoeninger-speere.html 10. http://www.larecherche.fr/savoirs/histoire-sciences/bataille-aurignacienne-abbe-breuil-01-11-2006-
75209 11. http://www.14c.uni-erlangen.de/grundlagen.html

15
Abbildungsverzeichnis Abb. 1: Geographische Lage des Neandertals. (J. Kraft, Rheinisches Landesmuseum Bonn) ......................................................................................................................... 16!Abb. 2: Johann Carl Fuhlrott ...................................................................................... 16!Abb. 3: Neandertaler Knochen aus dem Neandertal. Abgebildet sind die Knochen von 1856 sowie weitere Fragmente aus den Grabungskampagnen von 1997 und dem Jahr 2000. Foto Universität Bonn/ G. Oleschinski ............................................. 17!Abb. 4: Zeichnung von Gerhard Wandel mit den 1856 gefunden Knochen des Neandertalers ............................................................................................................ 17!Abb. 5: Hermann Schaaffhausen ............................................................................... 18!Abb. 6: Geographische Übersichtskarte .................................................................... 18!Abb. 7: Sauerstoff-Isotopenstufen der letzten 110 ka. ............................................... 19!Abb. 8: GRIP & GRIP 2 grönländisches Eisbohrprojekt ............................................. 20!Abb. 9: NGRIP: Dansgaard-Oeschger Zyklen ........................................................... 20!Abb. 10: Geographische Verbreitung des KI Tephras ............................................... 21!Abb. 11: Sierra de Atapuerca ..................................................................................... 21!Abb. 12: Verbreitungsgebiet des klassischen Neandertalers .................................... 22!Abb. 13: Geographische Ausbreitung des Micoquiens .............................................. 23!Abb. 14: Geographische Ausbreitung der Fundplätze des MTA ................................ 23!Abb. 15: Geographischer Raum und Verteilung der Hauptfundplätze des Châtelperroniens (Abb. (unten) nach Pelegrin 1995) ................................................ 24!Abb. 16: Geographische Ausbreitung des Lincombien-Ranisien-Jerzmanowicien .... 25!Abb. 17: Verbreitungsgebiet des Bohuniciens und des Szeletiens ............................ 26!Abb. 18: Verbreitungsgebiet des Uluzziens ............................................................... 27!Abb. 19: Verbreitungsgebiet des Aurignaciens .......................................................... 27!Abb. 20: Potenzielle Schäftungsweisen im Jungpaläolithikum .................................. 28!Abb. 21: Geographische Lage des Abric Romaní de Capellades .............................. 29!Abb. 22.: Position von Level J innerhalb der Gesamtstratigraphie ............................ 30!Abb. 23: Gesamtstratigraphie von Labeko Koba ....................................................... 31!Abb. 24: Geographische Lokalisierung des Fundplatzes von Saint-Césaire ............. 31!Abb. 25: Sagittalansicht der einzelnen Schichten ...................................................... 32!Abb. 26: Geographische Lokalisierung der Spy Grotte .............................................. 32!Abb. 27: Gesamtstratigraphie der Ilsenhöhle ............................................................. 33!Abb. 28: Mögliche Knochenspitze und Elfenbeinscheibe des LRJ ............................ 34!Abb. 29: Geographische Lokalisierung des Fundplatzes Stránská skála .................. 35!Abb. 30: Stratigraphie aus der Grotta del Cavallo ...................................................... 36!Abb. 31: Gesamtstratigraphie der Fumane Grotte ..................................................... 37!Abb.: 32: Gesamtstratigraphie der Klissoura Grotte .................................................. 37!Abb. 33: Übersichtsplan der besprochenen Übergangsindustrien, gegliedert nach deren chronologischen Erscheinen in 14C cal. Daten und NGRIP. ............................ 38!

16
I Abbildungen
Abb. 1: Geographische Lage des Neandertals. (J. Kraft, Rheinisches Landesmuseum Bonn)
Abb. 2: Johann Carl Fuhlrott
Mitteilungen der Gesellschaft für Urgeschichte — 12 (2003) 25
Interdisziplinäre Untersuchungen an den Neufunden aus dem Neandertal
Johann Carl Fuhlrott (1803 - 1877) gewidmet
Ralf W. SchmitzInstitut für Ur- und Frühgeschichte und Archäologie des Mittelalters
Abteilung Ältere Urgeschichte und QuartärökologieSchloss Hohentübingen, 72070 Tübingen
Zusammenfassung: Nachgrabungen an der Fundstelle des Neandertaler-Typusexemplares erbrachten in den Jahren 1997 und 2000 erstmals kulturelle Hinterlassenschaften des pleistozänen Menschen aus dem Bereich der Höhlen „Feldhofer Kirche” und „Kleine Feldhofer Grotte”. Bei den Artefakten handelt es sich um Typen des Gravettien und der Keilmessergruppen. Weiterhin konnten neben ergänzenden Skelettelementen des namengebenden Neandertalers auch Knochenreste eines bis dahin unbekannten zweiten Neandertaler-Individuums geborgen werden. Die Ergebnisse der Analysen von mitochondrialer DNA aus der Knochensubstanz beider Neandertaler stützen das Out of Afrika-Modell und sprechen gegen einen Beitrag der Neandertaler zum Genpool der heutigen Menschen. 14C-Daten von ~ 40 000 Jahren BP für die Fossilien Neandertal 1 und 2 betten nicht nur diese beiden Neandertaler, sondern auch die aus ihnen gewonnenen DNA-Sequenzen in das Sauerstoff-Isotopenstadium 3 ein.
Abstract: Renewed excavations of the sediments from the caves “Feldhofer Kirche” and “Kleine Feldhofer Grotte” in the Neander Valley during 1997 and 2000 yielded the first cultural remains from the Neanderthal type site. The artefacts found are Gravettian and Micoquian forms. In addition, bones belonging to the Neanderthal type specimen and bones which could be assigned to a second Neanderthal individual were recovered. The results of mitochondrial DNA analyses using mtDNA from both specimens support the Out of Africa-model and do not indicate a contribution of Neanderthal DNA to the modern human gene pool. Radiocarbon measurements date Neanderthal 1 and Neanderthal 2, and the corresponding mtDNA sequences, to ca. 40 000 years BP and consequently to Oxygene Isotope Stage 3.
!
!
"
Neandertal
Düsseldorf
Köln
Abb. 1: Geographische Lage des Neandertals. (J. Kraft, Rheinisches Landesmuseum Bonn).
Mittelungen12.indd 25 23.01.04, 12:57:49
10.01.15 11:56Fuhlrott.jpg 262×400 Pixel
Seite 1 von 1http://upload.wikimedia.org/wikipedia/commons/e/ed/Fuhlrott.jpg

17
Abb. 3: Neandertaler Knochen aus dem Neandertal. Abgebildet sind die Knochen von 1856 sowie weitere Fragmente aus den Grabungskampagnen von 1997 und dem Jahr 2000. Foto Universität Bonn/ G. Oleschinski
Abb. 4: Zeichnung von Gerhard Wandel mit den 1856 gefunden Knochen des Neandertalers
08.01.15 20:5920244.jpg 1.798×3.008 Pixel
Seite 1 von 1http://www.lwl.org/pressemitteilungen/daten/bilder/20244.jpg

18
Abb. 5: Hermann Schaaffhausen
Abb. 6: Geographische Übersichtskarte
10.01.15 11:56Hermann_Schaafhausen.jpg 972×1.314 Pixel
Seite 1 von 1http://upload.wikimedia.org/wikipedia/commons/7/75/Hermann_Schaafhausen.jpg

19
Abb. 7: Sauerstoff-Isotopenstufen der letzten 110 ka.
132
Chapter 8
OIS-2 OIS-3 OIS-4 OIS-5a OIS-5b OIS-5c
LGM cold E.cold Trans. Stable warm EGM
Figure 8.1. Second- (labelled) and third-order (Dansgaard/Oeschger) climate
changes for the period 110–10 ka BP from the GISP2 ice core, Greenland (Meese
et al. 1997; Johnsen et al. 2001; Stuiver & Grootes 2000). The broken heavy
line depicts the general trend of climate changes for the interval (from Chapter
4: van Andel et al. 2003b, Fig. 4.1). Climate Phases: Last Glacial Maximum
Cold Phase (LGM Cold); Early Cold Phase (E. Cold); Transitional Phase
(Trans.); Stable Warm Phase (Stable warm); Early Glacial Maximum (EGM).
Fennoscandian ice sheet began. For details of the
climate history itself, see Chapter 2 (van Andel 2003).
Simulating the late Quaternary climate of Europe
Even supercomputers lack the capacity to display at
high resolution the millennial-scale climate changes
typical of OIS-3. Therefore, the Stage 3 models were
limited to ‘snapshots’ intended as typical for the end
members of the glacial climate range. An extensive
set of modern climate data was used to evaluate the
modelling programme itself and to provide a com-
parison with the rigours of the climates of the last
glacial period (Chapter 5: Barron & Pollard 2002;
Pollard & Barron 2003). The results give us reason-
able confidence in the use of high-resolution, nested
global and regional circulation models.
When evaluating hypotheses that changing mid-
and late glacial human settlement patterns in Eu-
rope were partly or mainly responses to climatic
forcing, contrasts between ranges of climate vari-
ables are often more informative than absolute values.
Among the about 50 variables of the Stage 3 simula-
tion set (Chapter 5: Barron et al. 2003, Appendix) that
may be directly or indirectly relevant to the human
case, we believe the following to have had the larg-
est impact: summer and winter monthly mean tem-
peratures, day/night temperature contrasts, wind-chill,
precipitation, including thickness of snow-cover and
annual snow days, as well as the seasonal ranges of
those variables.
The European climate today
The European climate is domi-
nated by two major gradients, a
strong N–S gradient from arctic
Scandinavia to the subtropical
Mediterranean (Fig. 8.2: top),
and an equally strong transition
from the maritime Atlantic
coasts to the continental condi-
tions in the east. The N–S gradi-
ent is disrupted by the high W–E
mountain barrier of the Pyr-
enees, Alps and Carpathians,
which separates the temperate
European zone from the hot,
summer-dry Mediterranean zone.
Farther east, beyond the 25th
meridian, the subarctic climate
zone passes more gradually into
the subtropical Mediterranean.
The transition from the
maritime Atlantic region to con-
tinental eastern Europe has temperature contrasts as
stark as those between the Mediterranean and Arctic
regions; note, for instance the 15°C northward shift
of the summer-warm zone east of the 15°E meridian
(see also Chapter 5: Barron et al. 2003, Figs. 5.11–
5.16). The eastward increasing continentality trans-
forms the mild, moist winters and cool summers of
the west coast into the harsh winters and hot sum-
mers of Russia. Striking regional contrasts in the
amounts of precipitation add further complexity to
the European climate (Chapter 5: Barron et al. 2003).
The climate of the last glaciation
The climate pattern of the Stage 3 Stable Warm Phase,
about 59–42 ka BP (Fig. 8.2: centre), resembles that of
the present, except for being 4°–6°C cooler than the
modern state at each latitude. Mean summer tem-
peratures above 18°C occurred only in southern Spain
and the western Mediterranean, but in the east, well
beyond 20°E, they reached as far north as the 50°–
55°N parallel in Scandinavia (only in western Russia
did they extend below the 50°N one).
During the Last Glacial Maximum, conditions
were more extreme (Fig. 8.2: bottom). Summer days
above 18°C existed only in the southern Mediterra-
nean and the extreme southeast, south of 45°N. In
western Europe, the summer rarely reached 10°C,
while in winter only the Atlantic shores of southern
France, Iberia and the Mediterranean coasts had
‘mild’ average temperatures of 4°–6°C. Elsewhere,

20
Abb. 8: GRIP & GRIP 2 grönländisches Eisbohrprojekt
Abb. 9: NGRIP: Dansgaard-Oeschger Zyklen
1. Because glacier ice is a sediment, defining the Holoceneboundary stratotype in an ice core is as justified as basing astratotype on hard or soft rock sequences. Moreover, while itmight be argued that there is a potential impermanenceabout the Greenland ice sheet (i.e. it could melt withaccelerated global warming), it could equally be arguedthat no conventional geological section or exposure has anassured permanence. Quarrying, flooding, erosion orinstability/collapse are just four of the potential threats tothe preservation of a conventional GSSP.
2. Ice sheets form through the annual incremental accumu-lation of snow. This means that there is a continuity ofaccumulation (sedimentation) across the Pleistocene–Holocene boundary. Moreover, the relatively heavy snow-fall at the NGRIP site at the end of the last cold stage alsomeans this key part of the record is very highly resolved –far more so than is the case with many terrestrial sedimen-tary contexts.
3. Because of its geographical location in the high-latitudeNorth Atlantic, Greenland is a sensitive indicator ofhemispherical-scale climate change. This is likely to havebeen even more the case at the Pleistocene–Holocene
transition when the Greenland ice sheet lay mid-waybetween the wasting Eurasian and Laurentide ice masses.As noted above, a range of climate proxies is preservedwithin the Greenland ice, a number of which are immedi-ately responsive to climate shifts. Accordingly, early signalsof the marked climatic change that defines the onset of theHolocene Epochwill be registered very clearly in the Green-land ice-core record.
4. If defined on the basis of theNGRIP climatic record, the baseof the Holocene can be very precisely dated by annual ice-layer counting (see below), and can be replicated in otherGreenland ice-core records (see Greenland Ice Core Chron-ology 2005/GICC05, above). The boundary stratotype forthe Holocene (GSSP) is therefore at a level of chronologicalprecision that is unlikely to be attainable in any otherterrestrial stratigraphic context.
5. The Greenland ice-core record has been proposed by anINQUA project group (INTIMATE) as the stratotype for theLate Pleistocene in the North Atlantic region (Walker et al.,1999) and an ’event stratigraphy’ was initially developed forthe Last Termination based on the oxygen isotope record inthe GRIP ice core (Bjorck et al., 1998). More recently, the
Figure 1 The locations of five deep drilling sites on the Greenland ice sheet: NGRIP (75.18N, 42.38W), GRIP (72.58N, 37.38W), GISP2 (72.58N,38.38W), Dye-3 (65.28N, 43.88W) and Camp Century (77.28N, 61.18W). Also shown is the shallower site (324m) of Renland (71.38N, 26.78W).At all of these sites, the ice-core record extends back to the last (Eemian) interglacial. Map by S. Ekholm, Danish Cadastre
Copyright ! 2008 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 24(1) 3–17 (2009)DOI: 10.1002/jqs
GSSP FOR THE BASE OF THE HOLOCENE IN THE NGRIP ICE CORE 7
Dansgaard-Oeschger-Zyklen
Am Ende der Eem-Warmzeit begannen die Temperaturen zunächst langsam zu sinken.
Der letzte Kaltzeit-Zyklus
Klima 74
Schon bald darauf setzten im Bereich um den Nordatlantik Klimaschwankungen zwischen kalten Stadialen und deutlich wärmeren Interstadialen (insgesamt 25) ein, bei denen vor allem die Erwärmungen abrupt erfolgten. Sie prägten die gesamte letzte Kaltzeit. Nach ihren Entdeckern werden die Klimaschwankungen als Dansgaard-Oeschger-Zyklen bezeichnet.
Willi Dansgaard (links), Hans Oeschger (rechts), dazwischen Chester C. Langway (nach dem nichts benannt ist).
Temperatur-rekonstruktion basierend auf dem NGRIP Eisbohr-kern aus Grönland (oben) und auf einem Sediment-bohrkern vor der spanischen Küste (Quelle: Nature):

21
Abb. 10: Geographische Verbreitung des KI Tephras
Abb. 11: Sierra de Atapuerca
neutrally termed the European Late Pleistocene shift[Fedele et al., 2002]. In the cultural sphere, the Mousteriantool-making traditions of the so-called Middle Palaeolithiceventually changed into the industries or cultures of theUpper Palaeolithic, which many authors regard as charac-terized by an unprecedented explosion of artifacts reflectingideological purposes. In broad temporal concordance, awidely accepted shift has been noted in biological popula-tions, where the Neandertal anatomical type appears to have
been substituted by fully “modern” humans, a supposedlyunrelated morphology (formal taxonomic names are avoid-ed). The interplay of similarities and differences across theshift is a subject of intense and continuing controversy [e.g.Carbonell and Vaquero, 1996; Clark and Willermet, 1997;Fox, 1998; Akazawa et al., 1998; Klein, 1999; Zilhão andD’Errico, 1999; Bar-Yosef and Pilbeam, 2000; Straus andBar-Yosef, 2001]. No consensus yet exists as to whether theobserved changes in both cultural and biological domains
FEDELE ET AL. 303
Figure 1. Geographic distribution of the Campanian Ignimbrite deposits, including archaeological and sampling sitesmentioned in the paper. Solid squares: CI tephra occurrences and related thickness in cm (in italics if reworked) [mod-ified from Cornell et al., 1983; Amirkhanov et al., 1993; Cini Castagnoli et al., 1995; Narcisi and Vezzoli, 1999; Fedeleet al., 2002; Upton et al., 2002]. Dotted area in central Italy: distribution of the CI-derived paleosol (CI-PS [Frezzottiand Narcisi, 1996]). Solid triangles: archaeological sites with CI ash layer. Blank triangles: archaeological sites with ashlayer attributed to the CI on the basis of cultural-stratigraphic position and 14C dating (Fedele et al., 2002; Giaccio andIsaia, on file). Solid circles: selected European Palaeolithic sites within the area potentially affected by the CI air-fall.
75963_301-328.qxd 10/7/2003 2:37 PM Page 303
06.02.15 16:22ScienceDirect - Full Size Image
Seite 1 von 1http://www.sciencedirect.com/science?_ob=MiamiCaptionURL&_met…&_pii=S0305440306001580&md5=efcbf552cdc0076f972a37698accd542
�

22
Abb. 12: Verbreitungsgebiet des klassischen Neandertalers
199
4;H
olli
day
,199
7).
Yet
,a
nu
mbe
rof
trait
s,su
chas
the
hyp
er-b
arr
el-s
hape
ofth
eth
orax,
the
thic
ken
edco
rtic
al
bon
ean
dp
ron
oun
ced
are
as
ofm
usc
lea
ttach
men
t,th
ele
ngth
ofth
eca
lcan
eus
an
dth
ela
rge
nasa
lp
irif
orm
ap
ertu
re,
are
thou
gh
tto
refl
ect
extr
emel
yh
igh
level
sof
ph
ysi
cal
act
ivit
y(e
.g.,
Tri
nk
au
s,198
6;
Fra
nci
scu
san
dC
hu
rch
ill,
20
02;
Wei
nst
ein
,2008;
Ra
ich
len
etal.
,2011
).S
uch
act
ivit
yp
att
ern
s,to
get
her
wit
hth
ep
hysi
olog
ica
lst
ress
ind
uce
dby
cold
clim
ate
s,w
ould
ha
ve
imp
osed
sign
ifica
nt
hig
h-e
ner
gy
dem
an
ds
onN
ean
der
tha
lbod
ies
(Sor
ense
nan
dL
eon
ard
,2001;
Ste
egm
an
net
al.
,20
02;
Ch
urc
hil
l,2
006):
met
abol
icre
qu
irem
ents
that
wou
ldh
ave
forc
edN
ean
der
tha
lsto
con
sum
ean
ener
gy-r
ich
die
t.S
up
por
tfo
rth
isarg
um
ent
mos
tly
der
ived
from
sta
ble
isot
opic
an
aly
ses
(e.g
.,R
ich
ard
sa
nd
Tri
nk
au
s,20
09),
wh
ich
sugges
ted
that
Nea
nd
erth
als
obta
ined
nea
rly
all
ofth
eir
die
tary
pro
tein
sfr
oman
imal
sou
rces
,an
doc
cup
ied
the
hig
her
level
ofth
efo
odtr
oph
icw
eb,
sim
ila
rto
the
top
-lev
elca
rniv
ores
ofth
at
tim
e.T
he
tra
-d
itio
na
lin
terp
reta
tion
ofN
ean
der
tha
lsa
sex
clu
sivel
ym
eat-
eate
rs,
has,
how
ever
,bee
nch
all
enged
onse
ver
al
fron
tsov
erre
cen
td
eca
des
.M
oun
tin
gev
iden
cesu
gges
tsth
at
the
nit
rogen
isot
ope
reco
rdm
ay
un
der
esti
ma
teth
eco
nsu
mp
tion
ofst
arc
h/c
arb
ohyd
rate
rich
pla
nt
food
sb
yh
um
an
s(A
mbro
seet
al.
,2003;
Hed
ges
an
dR
eyn
ard
,20
07;
Sp
onh
eim
eran
dL
ee-T
hor
p,
2007
).M
ore
imp
or-
tan
tly,
an
aly
ses
ofn
ewa
rch
aeo
logic
al
evid
ence
from
the
Med
iter
ran
ean
coa
stli
nes
ofS
outh
ern
Eu
rop
ean
dth
eN
ear
East
,d
enta
lw
ear
stu
die
s,an
dm
icro
foss
ils
inca
lcu
lus,
ha
ve
dem
onst
rate
dth
at
Nea
nd
erth
als
wer
eex
plo
itin
ga
wid
eva
riet
yof
food
sou
rces
(e.g
.S
trin
ger
etal.
,2008;
El
Zaa
tari
etal.
,2011
;F
iore
nza
eta
l.,
20
11a;
Hen
ryet
al.
,2
011
;S
ala
zar-
Garc
! ıaet
al.
,2
013;
Sis
tiaga
etal.
,2014
).T
his
un
exp
ecte
dd
ieta
ryco
mp
lex-
ity,
cou
ple
dw
ith
aver
yw
ide
tem
por
al
an
dgeo
gra
ph
icra
nge,
ha
sle
dm
an
yto
reco
nsi
der
the
ecol
ogy
an
dfo
r-agin
gst
rate
gie
sof
Nea
nd
erth
als
.T
oacc
omp
lish
this
task
,h
ere
we
revie
wso
me
trad
i-ti
onal,
as
wel
las
inn
ovati
ve
ap
pro
ach
esu
sed
inth
ere
con
stru
ctio
nof
Nea
nd
erth
al
die
ts,
such
as
lith
ican
dzo
oarc
haeo
logic
al
an
aly
ses,
stable
isot
opic
met
hod
s,p
ale
-
opath
olog
y,to
oth
wea
rst
ud
ies,
an
du
seof
den
tal
calc
u-
lus.
Fir
st,
how
ever
,it
isim
por
tan
tto
giv
ea
gen
eral
intr
odu
ctio
non
Nea
nd
erth
al
geo
gra
ph
icd
istr
ibu
tion
,w
ith
afo
cus
oncl
imate
an
dveg
etati
onin
Eu
rasi
ad
uri
ng
the
Mid
dle
an
dL
ate
Ple
isto
cen
e.It
wil
lbe
foll
owed
by
ad
iscu
ssio
nw
her
ein
we
sum
mari
zeth
isin
form
ati
on,
tak
-in
gin
toacc
oun
tec
olog
ical
vari
ati
onam
ong
Nea
nd
er-
thals
an
din
clu
de
nu
trit
ion
an
dp
ath
olog
y-b
ase
darg
um
ents
for
ad
iet
rich
inan
imal
pro
tein
s.
NE
AN
DE
RT
HA
LW
OR
LD
:G
EO
GR
AP
HIC
DIS
TR
IBU
TIO
N,
CL
IMA
TE
AN
DV
EG
ETA
TIO
N
Ow
ing
toth
ein
exor
able
lin
ks
bet
wee
na
habit
at
an
dit
sor
gan
ism
s(e
.g.,
spec
ies
ofan
imals
an
dp
lan
ts),
an
ass
essm
ent
ofN
ean
der
thal
subsi
sten
cep
att
ern
sre
qu
ires
an
over
vie
wof
the
envir
onm
ents
inw
hic
hth
eyli
ved
.N
ean
der
thals
occu
pie
dw
este
rnE
ura
sia,
aw
ide
geo
-gra
ph
icare
ara
ngin
gW
est
from
the
Iber
ian
Pen
insu
laE
ast
toS
iber
ia(K
rau
seet
al.
,2007),
an
dN
orth
from
Ger
man
yS
outh
toth
eL
evan
t(e
.g.
McD
erm
ott
etal.
,1993;
Wol
pof
f,1996)
(Fig
.1).
Th
isre
gio
nen
com
pas
sed
con
sider
abl
eto
pog
rap
hic
alh
eter
ogen
eity
(e.g
.,in
teri
oran
dco
asta
lar
eas,
mou
nta
in-
ous
orre
lati
vely
flat
terr
ain
s),
soil
com
pos
itio
n,
and
vari
a-
tion
inte
mper
atu
reh
um
idit
y,la
rgel
yin
flu
ence
dby
lati
tud
e.B
esid
esth
at,
Nea
nd
erth
als
are
mos
tly
doc
u-
men
ted
from
mar
ine
isot
ope
stag
es(M
IS)
6th
rou
gh
3,
alo
ng
per
iod
ofti
me
char
acte
rize
dby
sever
ecl
imati
cco
nd
i-ti
ons,
wit
hal
tern
ati
ng
gla
cial
/in
terg
laci
al
cycl
es.
Th
isw
ide
geog
raph
icd
istr
ibu
tion
and
clim
ati
cfl
uct
uati
ons
du
rin
gth
eM
iddle
an
dL
ate
Ple
isto
cen
edro
veth
efo
rma-
tion
ofd
iver
sean
dd
isco
nti
nu
ous
envir
onm
ents
,sp
ann
ing
from
war
man
dfo
rest
edto
cold
an
dop
enh
abit
ats
.C
han
g-
ing
clim
ate
also
stro
ngly
infl
uen
ced
the
dis
trib
uti
onp
at-
tern
sof
mam
mal
ian
her
biv
ores
and
oth
erfo
odso
urc
esfo
rcin
gN
ean
der
thals
toad
apt
toth
en
ewco
nd
itio
ns.
Th
eM
id-L
ate
Ple
isto
cen
ew
as
mark
edby
agen
eral
incr
ease
inari
difi
cati
on,
bu
tals
ogre
at
inst
abil
ity,
wit
hep
isod
esof
max
imu
mic
eex
pan
sion
char
act
eriz
edby
cold
,d
ryco
ndit
ion
san
dse
ale
vel
sd
rops
ofh
un
dre
ds
met
ers
Fig
.1.
Nea
nd
erth
al
geo
gra
ph
icd
istr
ibu
tion
from
hu
man
foss
ilre
main
s(R
epro
du
ced
wit
hp
erm
issi
onfr
omK
rau
seet
al.
,N
atu
re,
2007,
449,
902-9
04)
.
44
L.
FIO
RE
NZ
AE
TA
L.
Yea
rboo
kof
Ph
ysi
cal
An
thro
pol
ogy

23
Abb. 13: Geographische Ausbreitung des Micoquiens
Abb. 14: Geographische Ausbreitung der Fundplätze des MTA
'SPN�5PPMT�UP�4ZNCPMT
���
#PSEFT �������.FMMBST �������BOE�TFF�B�SFDFOU�TZOUIFTJT�JO�4PSFTTJ ��������o� �BOE�B�QSFDJTF�UJNF�EVSBUJPO �FOEVSJOH�CSJFnZ�FJUIFS�TJEF�PG����,Z�#1�'JHT�� ����.5"�JT�DIBSBDUFSJTFE�CZ�TNBMM�BOE�mOFMZ�SFUPVDIFE�DPSEJGPSN�CJGBDFT�BOE�CZ�CBDLFE�FMPOHBUFE�nBLFT� BT� XFMM� BT� XFMM� GPSNFE� FOE�TDSBQFST� BOE� CPSFST� #PSEFT � ������ 'JH�� ��� 5IF�BTTFNCMBHFT�VTFE�GPS�UIJT�BOBMZTJT�XFSF�UIF�FQPOZNPVT�BTTFNCMBHFT��-F�.PVTUJFS�MBZFST�(�BOE�) �1FDI�EF�M�"[�MBZFST���UP�� �BT�XFMM�BT�UXP�PUIFST�o�-B�3PDIFUUF�MBZFS���BOE�-B�(SPUUF�97*�MBZFS�$��5IFTF�XFSF�BMM�EBUFE�CZ�SBEJPNFUSJD�NFUIPET�TFF�SFGFSFODFT�GPS�UIF�EBUJOH�JO�UIF�MFHFOE�PG�'JH�����
4UBCJMJUZ�PG�.PVTUFSJBO�UFDIOPMPHZ "DDPSEJOH�UP�TPNF�BVUIPST �BNPOH�UIFN�TFWFSBM�XIP�EFGFOE�UIF�3FWPMVUJPO�NPEFM�
PG�UIF�EFWFMPQNFOU�PG�CFIBWJPVSBM�NPEFSOJUZ �.PVTUFSJBO�MJUIJD�UFDIOPMPHZ�XPVME�CF�JOUFSOBMMZ�TUBUJD�BOE�DPOTJTUFOU�UISPVHI�UIF�.JEEMF�BOE�UIF�-BUF�1MFJTUPDFOF�DPNQBSFE�
'JHVSF��� �(FPHSBQIJD�TQSFBE�PG�UIF�.PVTUFSJBO�PG�"DIFVMJBO�USBEJUJPO�XJUI�DPSEJGPSN�CJGBDFT�PG�UIF�MBUF�.JDPRVJBO�PG�
$FOUSBM�&VSPQF �BOE�MPDBUJPO�PG�TJUFT�NFOUJPOFE�JO�UIF�UFYU�
7
arm oricain à pièces bifaciales accom pagnées de hachereaux sur éclats, in itialem ent rap-portés à un MTA breton (parm i ces sites, celui de Bois-du-Rocher dans les Côtes-d’Arm or oucelui de Kervouster dans le sud du Fin istère ; Monnier, 1980 : 265-290 et 299-338, 1986 ;figure I 1) son t actuellem ent rapprochés du Micoquien d’Europe cen trale, en s’appuyant enparticulier dans ces séries sur la présence de Prondniks, couteaux à dos bifaciaux (Molines etal., 2001).
Enfin , un ensem ble de séries m oustériennes «à bifaces triangulaires plats» du Nord de laFrance** est égalem ent rapporté au MTA (Bordes, 1954 ; Som m é et Tuffreau, 1971 ; Tuffreau,1976, 1977 ; Farizy et Tuffreau, 1986 ; Tuffreau, 1990 : Cliquet, 2001 ; figure I 1 et figure I 2).Ces séries ainsi que les ensem bles à «bout-coupés» anglais (Roe, 1981 : 240-252) seraien tplus ancien que le MTA à cordiform es (voir p. 10).
LE MOUSTÉRIEN DE TRADITION ACHEULÉENNE : RAPPEL DE L’ÉTAT DES CONNAISSANCES ET DES INTERPRÉTATIONS
Figure I 1. Extension géographique des sites moustériens de tradition acheuléenne(d’après en particulier Boëda et Mazière, 1989 ; Cliquet, 2001 ; Delporte, 1976 ; Farizy etTuffreau, 1986 ; Janot, 1981 ; Jaubert et Rouzaud, 1985 ; Jaubert, 1994, 2000, 2001 ;Gruet, 1976 ; Mazières et Raynal, 1976 ; Molines et al., 20001 ; Roe, 1981 ; Tuffreau, 1990)
* : Des gisem en ts MTA pourraien t égalem en t se trouver au sud du Luxem bourg, en particulier dans lesvallées de la Moselle et de l’Our (Le Brun-Ricalens, 1994). **: Notons que des pièces bifaciales de m orpholo-gies sim ilaires, découvertes dans des gisem en ts de surface de la Vienne et de l’Indre-et-Loire, on t été publiéespar F. Bordes (1961 : planches 56-59).

24
Abb. 15: Geographischer Raum und Verteilung der Hauptfundplätze des Châtelperroniens (Abb. (unten) nach Pelegrin 1995)
Yet, arguments for a local Mousterian of Acheu-
lean Tradition origin for the Chatelperronian are
as follows:
1. The Mousterian of Acheulean Tradition and
the Chatelperronian share a unique interest for
backing tools and for unretouched backed
blanks,
2. They also share a unique combination of elon-
gated and backed blanks or retouched tools,
which does not exist in any other contempora-
neous industry,
3. The necessity to obtain backed artifacts
(retouched and unretouched) is actually guid-
ing the production of Mousterian of Acheu-
lean Tradition elongated flakes (Pelegrin &
Soressi 2007; Soressi 2005) as well as
the production of Chatelperronian blades
(Roussel 2011 & in press). Backed elongated
flakes and backed blades (i.e. with an asym-
metrical transversal section) are obtained
directly during the production. The method
used to produce blanks within the two indus-
tries is relying on the obtainment of a high
quantity of backed blanks, some of which
will be retouched. Some extra backed arti-
facts, retouched backed knifes and
chatelperron points, are obtained through a-
posteriori retouch of blanks symmetric in
section.
4. Geographic distribution of the
Chatelperronian matches outstandingly that
of the Mousterian of Acheulean Tradition
(Fig. 2), and they are chronologically compat-
ible (Soressi 2005).
European Middle to Upper Paleolithic TransitionalIndustries: Chatelperronian, Fig. 2 Map of the distri-bution of the Mousterian of Acheulean Tradition (MTA;
bolded line), the Mousterian of Acheulean Tradition typeb (shaded line) and the Chatelperronian (dotted line)
European Middle to Upper Paleolithic Transitional Industries: Chatelperronian 2683 E
E
285
J. Pelegrin et M. Soressi — Le Châtelperronien et ses rapports avec le Moustérien
Figure 2. Localisation géographique des principaux sites châtelperroniens (d’après Pelegrin, 1995)
En revanche, les débuts du Châtelperronien sont très mal connus et de datation très incertaine. D’une part, très peu de ses dépôts ont résisté à l’érosion de l’interstade würmien (une période ou oscillation climatique plus tempérée et humide entre le Würm ancien et le Würm récent) ; d’autre part, la méthode du carbone 14 trouve ici sa limite de viabi-lité : les tentatives de datation d’échantillons plus vieux que 38 000 BP se heurtent à la quasi-disparition du 14C qu’ils contiennent, au prix d’une imprécision grandissante. Nous verrons que cette méconnaissance des débuts du phé-nomène châtelperronien et de leur durée, au cours de l’obs-cur interstade würmien, complique l’appréciation de son ori-gine, même si une hypothèse peut être avancée.
Cultures synchrones
Par sa position chronologique, comme par ses particula-rités industrielles et son ambiguïté anthropologique, le
Châtelperronien peut être considéré comme l’un des tech-nocomplexes « transitionnels » entre la très longue pério-de des industries moustériennes et les sociétés beaucoup mieux connues du Paléolithique supérieur (Aurignacien, Gravettien, etc.). Dans cette même tranche d’âge, on reconnaît en Italie le technocomplexe uluzzien, caractérisé par des petites pièces à dos arqué en demi-lune. Dans une large portion sud de la péninsule Ibérique (Espagne et Portugal), perdureraient des industries moustériennes, comme dans le centre-sud de la France. En Europe centrale, des industries à débitage Levallois (Bohunicien) et/ou à pièces foliacées (Széléttien, Jankovichien) en sont contemporaines (fig. 4).
Les sites du Châtelperronien
Les sites sous abri rocheux (en pied de falaise, avec un auvent plus ou moins avancé) sont les plus fréquents.

25
Abb. 16: Geographische Ausbreitung des Lincombien-Ranisien-Jerzmanowicien
D. FLAS
Jerzmanowice point fragments (two burins on break, a splintered piece, a Jerzmanowice point with rounded base).
JERZMANOWICE POINTS: AURIGNA-
CIAN ARTEFACTS?
May the Jerzmanowice points coming from the second level of Spy be considered as part of the Aurignacian assemblage yielded from the same deposit? For several reasons, the an-swer is negative.
This “second fauna-bearing level” in-cluded an obviously mixed assemblage with clear Aurignacian and Mousterian elements (and also
some pottery), unsurprising considering the ex-cavation techniques used in the late 19th cen-tury7. While Jerzmanowice points were long considered as part of the Aurignacian as-semblage, this was mainly because they are made on blades and classified as “Protosolutrean”. As D. de Sonnevilles-Bordes (1961) already under-scored, in such fuzzy stratigraphic context, noth-ing really suggests that these points are Aurig-nacian. It is thus not possible to group the Jerz-manowice points with other artefacts or to give them a precise chronological position.
222
7 Because of sediment thickness, M. De Puydt and M. Lohest used a gallery system of excavation (for a detailed analysis of the problems linked with inaccuracy of the old excavations at Spy, cf. Dewez, 1980: 37-41; Semal et al., this volume: chapter II).
1: Paviland 11: Pin Hole 21: Moordown 31: Goldcliff 2: Ffynnon Beuno cave 12: Robin Hood Cave 22: Sutton Courtenay 32: Temple Mills 3: Bench Tunnel Cavern 13: Glaston Grange Farm 23: Osney Lock 33: Aardjesberg 4: Windmill Hill cave 14: Beedings 24: Creffield Road 34: Spy 5: Kent’s Cavern 15: Earl of Dysart’s Pit 25: Wallow Camp 35: Goyet 6: Uphill Quarry cave 8 16: Drayton 26: Brandon 36: Zwergloch 7: Soldier’s Hole 17: Warren Hill 27: Town Pit 37: Ranis 2 8: Hyaena Den 18: White Colne Pit I 28: Bapchild 38: Nietoperzowa cave 9: Badger Hole 19: Eastall’s Pit 29: Conningbrook Manor Pit 39: Koziarnia cave 10: King Arthur’s Cave 20: Bramford Road Pit 30: Hainey Hill 40: Puchacza Skała cave
Figure 3. Geographic distribution of LRJ sites (modified after Flas, 2006 and Semal et al., 2009).Triangles = cave sites; circles = open air sites. Upper right: Jerzmanowice point from Spy cave.

26
Modell für die Entwicklung vom Siedlungsverhalten während der Transition vom Mittel- zum Jungpaläolithikum. A – ∼50-40 ka uncal BP; B – ∼40-35 ka uncal BP; C – ∼35-30 ka uncal BP. Abb. 17: Verbreitungsgebiet des Bohuniciens und des Szeletiens

27
Abb. 18: Verbreitungsgebiet des Uluzziens
Abb. 19: Verbreitungsgebiet des Aurignaciens
15.06.15 19:08ScienceDirect - Full Size Image
Seite 1 von 1http://www.sciencedirect.com/science?_ob=MiamiCaptionURL&_me…&_pii=S0047248414000074&md5=aec411e4fb4905e1de22f3deb0a9d046
�

28
Oben links: Schäftung einer rautenförmigen Spitze. Unten links: Schäftung einer spindelförmigen Spitze. Unten rechts: Schäftung einer Spitze mit abgeschrägter Basis.
Schäftung einer Spitze mit gespaltener Basis Abb. 20: Potenzielle Schäftungsweisen im Jungpaläolithikum
34 Heidi KNEcHT
Thickness in mmT2
experimentatiory the spears were found to easilyenter and even pass through the animal.
7. Mammoth-rib pointsProjectile points manufactured from mam-
moth ribs are known from several Gravettiansites in Germany. Pieces representative of eachstage of the manufacfuring process of mammothrib points were studied. The mammoth rib wasfirst divided into flat segments. The segmentwas next sectioned longitudinally. The fragmentwas then reduced by longitudinal scraping toform a gently tapering point that was oval orcircular in section. During shaping, the compactbone was completely removed from the piece.The resultant point was formed entirely ofcancellous bone.
Many of the mammoth rib points are scoredwith fine incisions which run both perpendic-ular and oblique to the longitudinal axis ofthe piece. Some of the mammoth rib pointshave flattened portions along their otherwiserounded shafts. Several features of mammothrib points are analogous to attributes of Perigor-dian single-beveled points: the use of cancellousbone; scoring or incising along lateral edges;and flattened surfaces. The system for haftinomammoth rib points remains to be explored.
8. Conclusion: Early Upper Paleolithicprojectile technologiesThere are distinct differences among the
various Early Upper Paleolithic bone and antlerprojectile technologies. These technologies aredistinguishable from the outset of the manu-facturing process on the basis of the selectionof bone or antler as the raw material forproduction. Given the selected raw materiaftechniques suitable for working of that materialwere used to produce different types of pro-jectile points. Particular attention was givento the manufacture of various forms of haftingmechanisms. Attached to shafts, the differenttypes of projectile points would have allowedfor the construction of several different butequally effective hunting weapons.
Both the manufacture and hafting of EarlyAurignacian split based points were based onthe principles of splitting and wedging. Giventhe mechanism used for attachment of the pointto the spear shaft, it would not have beennecessary to use resins to hold the point in place.
8
Width in mm7 6
Fig. 14 - Points with single-beveled bases: width x thickness.
down to form a bevel at an angle which matchedthat of the beveled base of the points (fig. 15).During experimentation, a step-like structurewas left at the end of the bevel on the handle toimpede the dislocation of the point upon impact.Also, resin was applied to the beveled surfacesto increase adhesion. The irregular surface of thebevel created by the natural openings of the can-cellous bone increased the tenacitv between thebase and the end of the handle; smooth surfaceswould have allowed for slippage. The shafts ofGravettian projectile points with single-beveledbases were sometimes scored with incisions
Fig. 15 - Hafting a point with single-beveled base.
perpendicular to the axis at the distal end of thebevel. It is likely that these incisions reducedslippage of the ligature with which the point wasbound to the shaft.
6.4. PefiormanceArmed with these weapons, Paleolithic
hunters would have been capable of maiming orkilling both large and small animals. During
30 Heidi KNpcnr
3.3. HaftingAn association between tongued pieces
(piices d languette) and split based points haslong been recognized. Since Peyrony's (1928)suggestion that tongued pieces are by-productsgenerated during the fabrication of split basedpoints, this explanation of their association hasbeen accepted (see fig. 1).
While studying the split based points andtongued pieces from Abri Castanet in thecollections of the Musde Nationnl de ln Prdhistoireat Les Eyzies, 27 pieces which appeared to benegative removals from tongued pieces werefound. Observations of these small rectangularpieces and numerous tongued pieces fromsites throughout France has allowed for thedelineation of the sequence of manufacture oftongued pieces. Examples of each stage of thissequence of production have been observedamong archaeological assemblages (fig. 5).1. A parallel-sided blank was shaped from
a semi-cylindrical segment of antler. Theproximal end was left unworked.
2. Perpendicular incisions were made acrossboth the inferior and superior faces of theblank.
3. Flat rectangular pieces were removed fromthe distal end of the blank by splitting orcleavage.
C I!,IJ J
l, n,[ ,.n.tN,[ -EJ
Fig. 5 - Manufacture of tongued piece and wedges.
TWo small wedges and one tongued pieceresulted from this procedure. The tonguedpiece was then used as the source of additionalwedges:4. First, the tongue was snapped off of the
tongued piece.5. Then perpendicular incisions were again
made across both faces of the tongued piece.
6. TWo small wedges were once again split offproducing a new tongue.The entire process could be repeated until
all that remained of the tongued piece wasthe unworked proximal portion and a smallremnant of the worked blank.
Fig. 6 - Hafting a split based point.
Experimentation supports the probabilitythat these small wedges were used in haftingsplit based points (fig. 6). It is likely that splitbased points were attached to shafts by useof a wedging mechanism. When antler is splitby cleavage, the split remains closed. For theexperimental reconstructions, the distal end ofa wood spear shaft was whittled out to forma U-shaped housing for the proximal end ofthe split based point. The split base point wasinserted into the handle and held in place bya ligature. When the ligature was wrappedaround the point, an opening was left so that thesplit was accessible. Once the point was boundto the shaft a small wedge like those removedfrom tongued pieces was inserted into the splitby gentle tapping. The wedge forced open thesplit until the wings of the base pressed firmlyagainst the inside of the housing in the woodshaft.
3.4. Performance
During experimentatiort spears armed withsplit based points penetrated the hide and softtissue of the animal with no damage to the pointor the haft regardless of the size or form of thepoint. When a point was projected into bone, itspenetration was halted, but the point and haftusually remained intact.

29
Abb. 21: Geographische Lage des Abric Romaní de Capellades

30
Abb. 22.: Position von Level J innerhalb der Gesamtstratigraphie
(Table 3). Examining the % Skeletal Survival Rate by skeletal seg-ments, the proximal appendicular skeleton is the most complete(83.33%Surv.), whereas the axial skeleton is lacking most elements(23.19% Surv.) (Fig. 2). The non-coordinated remains from the bagsof general !nds and wet sieving were checked for another study
(Gabucio et al., submitted for publication), and the few remains of F.silvestris recovered have also been considered in this study.Therefore, the anatomical bias is not due to the excavation process.This anatomical bias, moreover, does not correlate with the bonedensity values provided by Pavao and Stahl (1999) (Table 3).
Fig. 1. Location of Abric Romaní site (Capellades, Barcelona) with respect to the Iberian Peninsula (top left) and the region near the Capellades town (middle left). General stra-tigraphy and levels dating from the Abric Romaní (top right). Level O in the excavation of 2005 (bottom left) and 2010 (bottom right).
M.J. Gabucio et al. / Quaternary International 326-327 (2014) 307e318310

31
Abb. 23: Gesamtstratigraphie von Labeko Koba
Abb. 24: Geographische Lokalisierung des Fundplatzes von Saint-Césaire
Aurignacian occupations. In July 1979, a Neandertalskeleton associated with a Chatelperronian industry wasuncovered at Saint-Cesaire, suggesting that this archaicform was the author of the assemblage [24,35]. This wasa revolutionary statement at that time, given thatmodern humans were then considered the only makersof Upper Paleolithic industries in Western Europe (e.g.,[10,16]).
The association uncovered at Saint-Cesaire wasimmediately denied by F. Bordes [2] who was moreinclined to attribute the Chatelperronian to modernhumans, in agreement with his bi-lineal scheme ofbiocultural evolution for the Middle to Upper Paleo-lithic transition (for a fuller discussion see [28]). Amongother things, Bordes [2:644] expressed his skepticismconcerning the stratigraphic position of the Neandertalskeleton within the Saint-Cesaire sequence, arguingthat the skeleton was exhumed en petit comite (bya small party of people). The homogeneity of theChatelperronian assemblage was also questioned by deSonneville-Bordes [33] who stressed that the presence ofMousterian-looking artifacts in the Chatelperronianassemblage is a possible indication of contamination.These allegations, however, were disputed by Levequeand Vandermeersch [19 (p. 103), 25].
Stimulated by these discussions, Backer [1] analyzedthe artifact distribution in the Chatelperronian occupa-tion. She observed that bovine remains tend to clusterspatially with tools more typical of the MiddlePaleolithic, whereas reindeer remains tend to followthe distribution of the Chatelperron points. Elaborating
on these results, Guilbaud, followed by Rigaud [29,30],brought up the possibility of mixing of occupations:‘‘none of the results of this analysis prove that levelEJOP superior [the Chatelperronian level], stratigraph-ically very distinct, does not contain the results ofsuccessive, di!erent activities that spanned severalcenturies, even millennia’’ [11:49]. In support of thishypothesis, Guilbaud [11,12] stressed similarities in lithicreduction strategies between the ‘‘Mousterian’’ compo-nent of the Chatelperronian assemblage and materialsfrom the Denticulate Mousterian EGPF level. There-fore, is the ‘‘Mousterian’’ component of the Chatelper-ronian the result of mixing with the DenticulateMousterian? This question raises a broader issue: isthe cultural stratigraphy of Saint-Cesaire reliable? Inorder to answer these questions, the vertical and spatialdistribution of bone refits within and between eightfaunal assemblages from this site are presented here, aswell as data on changes in species composition, spatialdistribution, and body part representation. These resultsfocus on bones only as stone tool counts are notavailable for all the assemblages.
2. The Saint-Cesaire site
The Saint-Cesaire site takes its name from a villagenear the city of Saintes in Charente-Maritimes, France(Fig. 1). The site is found at the base of a 5e6 mlimestone cli! that had been previously exploited asa quarry [20]. The deposits were excavated during 12
Fig. 1. Geographical location of the Saint-Cesaire site.
1084 E. Morin et al. / Journal of Archaeological Science 32 (2005) 1083e1098

32
Abb. 25: Sagittalansicht der einzelnen Schichten
Abb. 26: Geographische Lokalisierung der Spy Grotte
the EJO inf layer likely rules out the possibility ofextensive contamination between the Chatelperronianand the Aurignacian bone samples. However, contam-ination between the Mousterian and Chatelperronianoccupations cannot be ruled out based on this data.
3.2. Changes in taxonomic composition
All else being equal, stratified assemblages that di!erdrastically in faunal composition are likely to indicatea low level of mixing. At Saint-Cesaire, the level-by-levelfaunal composition is examined using ratios of un-identified long bones of size 2 ungulates (most probablyreindeer, see [28] for a description of the classification)versus unidentified long bones of size 3e4 ungulates(most probably bison and horse) for a total sample of7283 long bones (Table 3). In spite of a loss of resolution
in taxonomic identification, this approach has theadvantage of increasing sample size in most of the smallassemblages. In order to address the possibility ofchange due to di!erences in horizontal sampling, onlythose squares (n=11) that are found in at least sevenof the eight levels studied here were included in theanalysis. Ratios obtained using NISP counts are alsopresented in Table 3. In this case, the ratios compare theabundance of reindeer versus horse and bison elementsfor the total excavated area. These three taxa wereselected because they represent the vast majority (95%)of the specimens identified at Saint-Cesaire.
As seen in Fig. 4, size 3e4 ungulates are far morecommon in the Mousterian and Chatelperronian levelsthan in the reindeer-dominated Aurignacian occupa-tions. Results obtained with NISP counts confirm thatthese two sets of occupations di!er significantly from
Table 2Saint-Cesaire (abundance of faunal remains by level, excluding birds and microfauna)
Layer Cultural attribution Surfacestudied (m2)
Volumestudied (m3)
Total bonecounts
Density
EJJ (3) Evolved Aurignacian 21.50 5.1 4231 830EJM (4) Evolved Aurignacian 17.25 3.1 8033 2591EJF (5) Aurignacian I 31.00 9.3 40,075 4309EJO sup (6) Aurignacian 0? 17.50 2.9 5610 1934EJO inf (7) very low density 17.50 2.2 801 (147) 364 (67)EJOP SUP (8) Chatelperronian 28.75 7.1 28,872 4066EJOP inf (9) Chatelperronian? 29.50 5.7 9510 1668EGPF-sample (10) Denticulate Mousterian 16.75 2.2 21,084 9584
Total/average 179.75 37.6 1,18,216 3144
Density is calculated by dividing the number of bones by the volume of sediments studied. Italic numbers are counts after excluding the threeunusually ‘‘rich’’ squares I4, I5, and J5. Density is probably over-estimated in the Denticulate Mousterian, as the sample included in this study favorrich decapages.
Fig. 2. Reconstruction of the Saint-Cesaire sagittal stratigraphy based on a three-dimensional analysis of piece-plotted artifacts. The dashed sectionto the right indicates the area of the deposit in which layers tend to merge.
1087E. Morin et al. / Journal of Archaeological Science 32 (2005) 1083e1098
!. Introduction
La grotte est connue pour ses richesses préhistoriquesdepuis la seconde moitié du XIX! siècle. La découverteen 1886, par Marcel De Puydt et Maximin Lohest, desrestes de deux individus adultes néandertaliens, et cedans un contexte stratigraphique authentifié par unprocès-verbal, lui confèrera une renommée internatio-nale. En e"et, l’observation stratigraphique de la positionde ces squelettes dans un « niveau ossifère » contenantune faune typique de « l’Âge du Mammouth » (L#$%&%,1861) et des artefacts de l’époque moustérienne('& M($%)**&%, 1883) est e"ectuée dans des conditionsscientifiquement acceptables. Elle démontre l’âge quater-naire de cet Homme plus archaïque que l’actuel, auquelWilliam King avait attribué le nom spécifique d’Homoneanderthalensis (K)+,, 1864).
". Localisation et description du site
La grotte de la Betche aux Rotches (P)$-(+, à paraître),plus communément appelée grotte de Spy, se trouve àl’ouest du village de Spy sur le territoire de la commune deJemeppe-sur-Sambre (prov. de Namur ; .),. /). La cavitéest creusée dans un massif calcaire du Viséen(Carbonifère), en contrebas d’une saillie rocheuse(P)$-(+ et al., à paraître). Elle est surplombée par unvaste plateau qui o"re un beau point de vue sur la valléede l’Orneau, a0uent de la Sambre, et sur les terres agri-coles contenant de nombreux rognons de silex résiduels,à l’emplacement d’une ancienne terrasse alluviale. Lescoordonnées géographiques du site sont de 50° 28’ 49”nord et 41 40’ 29” est.
Le porche de la grotte s’ouvre vers le sud sur une largeterrasse qui est située, à l’époque de la découverte, àenviron 18 m au-dessus du niveau de la rive gauche del’Orneau et dont la surface faisait 11m sur 6m (D& P23'%& L(4&-%, 1886, 1887).
A l’avant, une pente abrupte débouche sur une « basseterrasse » ou « terrasse inférieure » qui borde l’Orneau(D&5&6, 1980 et 1981). Son profil est aujourd’hui le
FIG. !Situation de la grotte de Spy. A. : localisation sur la carte deBelgique ; B. : carte topographique ; C. : vue S.-E. depuis l’Orneau,cliché de Rahir 1904, IRPA n 3353b.
A
B
C
2011 : 305-321
M7*#+,&- M#$,2&$)%& U*$)8-C*(--&%L& P#*7(*)%4)92& :(3&+ &+ B&*,)92&
La grotte de Spy (Jemeppe-sur-Sambre ; prov. Namur)PATRICK SEMAL, CÉCILE JUNGELS, KÉVIN DI MODICA, DAMIEN FLAS, ANNE HAUZEUR,
MICHEL TOUSSAINT, STÉPHANE PIRSON, GENNADY KHLOPACHEV, DAMIEN PESESSE,ÉLISE TARTAR, ISABELLE CREVECOEUR, HÉLÈNE ROUGIER & BRUNO MAUREILLE
ERAUL/;<
B2**&%)+ '&- C4&$=4&2$- '&*# W#**(+)&, 4($---7$)& +( >

33
Abb. 27: Gesamtstratigraphie der Ilsenhöhle

34
Abb. 28: Mögliche Knochenspitze und Elfenbeinscheibe des LRJ

35
Abb. 29: Geographische Lokalisierung des Fundplatzes Stránská skála
388 KAREL VALOCH, PETR NERUDA and ZDENKA NERUDOVÁ
F!"#$% &. Position and structure of Stránská Skála open-air site. A – position of the site on the map of Europe,B – position of the site on the SE border of the Brno City, C – distribution of the site on Stránská Skála hill,D – the shape of the eneolithic pit with the position of sectors A, B and C.

36
Abb. 30: Stratigraphie aus der Grotta del Cavallo
Mousterian layer F. These layers are separated by a thin volcanicstratum (Table 1). Auroch is the most common ungulate in FII, FIand EIII5, followed by red deer and horse. Fallow deer, decreasing inthe F layer, is absent in EIII5. The sporadic presence of Equushydruntinus and wild boar, the latter present as only one specimenin FII, is registered in this layer.
Species composition in these three layers indicates thata homogeneous climate existed in this part of Apulia from the LateMousterian to the Early Uluzzian, with a tendency towardsa decrease in temperature and precipitation (Nocentini, 2007).However, the ungulate skeletal elements indicate a signi!cantdifference between the Uluzzian layer EIII5 and the Mousterianseries below. In the Uluzzian layer EIII5, the percentages of theskeletal elements, including sesamoids, phalanges, carpals andtarsals, arewell above the averageof theentireMousterian sequence
of Grotta del Cavallo (Table 2). Sesamoids and phalanges, almostalways broken, are most abundant (30.1% vs. 7.9% average of theMousterian layer F). Tarsals and carpals are frequent (15.3% vs.1.4%).
3.1. Unidenti!able material
In the unidenti!able material from layer EIII5, excluding thecategory “others”, the spongy bone fragments represent the highestpercentage (34.8%). Pieces of diaphysis comprise a lowerpercentage (17.5%), followed by fragments of ribs (12.3%) and ofepiphysis (11.9%). A signi!cant number of skull elements, composedlargely of small teeth fragments (7.8%) is also present (Table 3).
Metric analysis of the unidenti!able material recovered in layerEIII5 shows dominance of the smaller fragments (1e3 and 3e6 cm),but signi!cant quantities of elements larger than 6 cm are alsopresent (Table 3). Table 4 shows that the fragmentation in thissample (1e3 cm) is lower than the one recorded for the similarsamples of the layers FI and FII and for the Mousterian layer 4/1 ofRiparo l’Oscurusciuto.
3.2. Signs of human and animal activity
Of the identi!ed bones in layer EIII5, 17.5% show anthropogenictraces. In the unidenti!able group, this percentage drops to 2.4%(Table 5). Cut marks are concentrated mainly on the limb bones ofungulates. Their position on the bones indicates that these tracesare attributable largely to the disarticulation of carcasses. Cutsobserved near a fracture on a horse scapula also suggest disartic-ulation. In addition, there are cut marks on the left and rightcondyles of an atlas of Equus ferus, indicating beheading of the prey.
Marks related to the use of cutting tools are the most frequentanthropic traces in the unidenti!able sample (Table 5). Excludingthe “others” these traces mainly affect the fragments of the shaftsand ribs, and are probably related to activities aimed at acquiringmeat. As with the cut marks, impact traces and fractures are mainlydistributed on the limb elements of the identi!able sample and onthe diaphysis fractions of the unidenti!able sample. In both cases,these traces are related to breakage of long bones for the extractionof marrow. Fractures have curved pro!les (Villa and Mahieu, 1991,Fig. 7b) with smooth edges (Morlan, 1984; Villa and Mahieu, 1991),and the angle formed by the cortical bone surface is oblique(Morlan, 1984; Johnson, 1985; Villa and Mahieu, 1991). Theseobservations suggest that bones were broken while still fresh. Two
Fig. 2. Schematic stratigraphic sequence of Grotta del Cavallo.
Table 1Grotta del Cavallo: number of identi!ed remains of the Uluzzian and Final Mous-terian. The percentages refer only to ungulate NISP. MNI ! Minimum number ofindividuals.
Uluzzian Final Mousterian
EIII5 FI FII
NISP-% MNI NISP-% MNI NISP-% MNI
Lepus sp. 1 1 4 1 2 1Crocuta crocuta 1 1 / /Canis lupus 2 1 / /Vulpes vulpes 4 1 6 1 7 2Equus ferus 53 (27.0) 3 14 (21.2) 2 21 (20.2) 2Equus hydruntinus 1 (0.5) 1 / /Equus sp. 1(0.5) 1 / /Sus scrofa 3 (1.5) 1 / 1 (0.9) 1Cervus elaphus 52 (26.5) 2 22 (33.3) 2 26 (25.0) 3Dama dama / 3 (4.5) 1 7 (6.7) 1Capreolus capreolus / / 1 (0.9) 1Cervidae sp. / 2 (3.0) 2 (1.9)Bos primigenius 86 (43.9) 3 25 (37.9) 3 46 (44.2) 3Total Ungulates 196 66 104Total NISP 204 76 113
P. Boscato, J. Crezzini / Quaternary International 252 (2012) 90e9892

37
Abb. 31: Gesamtstratigraphie der Fumane Grotte
Abb.: 32: Gesamtstratigraphie der Klissoura Grotte
almost exclusive. The overall sedimentary sequence displays vary-ing amounts of these basic lithological components as well as in thedensity of cultural remains. More specifically, layers A13 and A12are composed of loose breccia with prevalent sandy matrix, mostlyvertically arranged stones and affected by densely spaceddeformations incorporating the overlying unit A11. This latter unitand the overlying unit A10 mark the units with the highest contentof cultural and organic remains; they are thin levels and somestone-lines are interbedded and sediment includes loose, partiallyopen-worked breccia, with slabs often vertically arranged asa consequence of freeze–thaw action. A11 was excavated during the1988 and 1989 field seasons. Various levels were recognized withinthis unit, such as A11a and A11c (organic levels with dwellingstructures), A11d (ephemeral occupation level) and A11base (thinlayer overlying unit A12). Similarly, unit A10, excavated in the 1990trench, was subdivided into five fine levels as a function of inter-bedding and limited prevalence of gravel, aeolian loam, charcoal,ash and organic matter.
Unit A9 and sub-unit A8, the dispersed portion of A9 upper part,are stony layers with various amounts of aeolian dust. Unit A7 iscompletely lacking in primary cultural traces and thus marks aninterruption in human occupation. The latter unit clearly contrastswith unit A6 with its dense amount of organic matter, charcoal,faunal and lithic remains and some remains of combustion struc-tures such as the one we have labelled A6base SI, discovered in
squares 95 and 105. Regarding A5!A6, during excavation a thindiscontinuous anthropic horizon gradually vanishing in the upperportion of A6 emerged, which we labelled A5, but given that therewas no clear boundary between the two levels, we have labelled itas A5!A6. The uppermost units A4 and A3 are stony or sandylayers with few cultural remains and with slabs (A4I–A4III)progressively substituted by aeolian dust outside the present-daydrip-line (i.e. A4IV).
2.2. Palaeoecological setting and archaeological content
The palaeoecological setting related to the sequence describedabove is provided by palaeontological and charcoal diagrams.Pollen is rare and data on micromammals and other palae-oecological proxies are not yet available macromammals, andspecifically hunted ungulates found from units A12 to A4, revealmoist-cool climatic phases that favoured the expansion of forests infavour of alpine grasslands. Nevertheless, the presence of Capraibex, Rupicapra rupicapra and Marmota marmota in addition tosome birds – Pyrrhocorax graculus and Lagopus mutus – indicatesthat open alpine environments still characterized the cavesurroundings. In units A11–A10 cervids progressively and clearlyincrease relative to caprids revealing a moist–temperate climaticshift in parallel with forest expansion. Aside from a certaintendency towards cooler conditions observable towards the top of
Fig. 2. Stratigraphic sequence of Fumane Cave reported for the main sagittal section running from outside the cave (right). Lithological features of the most significant units withinthe S, BR, A and D complexes are reported. Key: 1 – rendzina, upper soil; 2 – slope deposits with boulders; 3 – living floors, with high concentration of organic matter or charcoal; 4 –loess and sandy loess; 5 – CaCO3 cemented layers; 6 – sandy deposits; 7 – unweathered and weathered bedrock. TL dates (after Martini et al., 2001) are reported for units S7, BR12and BR11.
M. Peresani et al. / Journal of Archaeological Science 35 (2008) 2986–29962988

38
Abb. 33: Übersichtsplan der besprochenen Übergangsindustrien, gegliedert nach deren chronologischen Erscheinen in 14C cal. Daten und NGRIP.
tilts toward the Neandertal side. Another aspect to take into
consideration relates to the fact that, to some degree, contrary towhat is observed with the IUP further east, some arguments sup-port local continuity for some of these western European assem-blages. The case seems stronger for the LRJ, but it should also beconsidered for the Uluzzian and for the Chatelperronian. If some orall of these assemblages were really produced by Neandertals, andtaking into account that they display clear UP features, it istempting to consider that the cultural changes at work in theNeandertal world of western Europe since the !nal Mousterianphases (Soressi et al., 2013) might result from cultural stimulusdiffusion (sensu Kroeber, 1940) from adjacent regions wheremodern humans had already been settled. The source of thisstimulus diffusionwould be represented !rst by the IUP, possibly asearly as 48 ka cal BP and later by the !rst phases of the Aurignaciancomplex. Although there is no evidence for prolonged local co-existence of Neandertals and modern humans in any region ofEurope, the ages of the latest directly dated Neandertals of WesternEurope (Spy and Saint-C!esaire) suggest that the chronologicaloverlap between the two groups might have been quite long on acontinental scale.
One can likely assign most, if not all, of the early phases of theAurignacian to modern humans, although the biological nature ofthe makers of these earliest phases is poorly documented inwestern Europe. In the Mediterranean world, the Early Ahmarian/Kozarnikian/Protoaurignacian seems to represent a rather consis-tent techno-typological ensemble, spreading rather quickly, mostlikely out of the Levant. Its occurrence in northern Italy and thePyrenees region predates the Campanian Ignimbrite and after 40 kaBP it further expanded in both central France and central Italy.Current stratigraphical and radiocarbon evidence suggest a partialchronological overlap with the Chatelperronian and Uluzzian. Thelimited development of body ornaments in both assemblages aswell as the production of bladelets in the Chatelperronian with a
chaîne op!eratoire different from that of the Protoaurignacian hasbeen suggested to result from direct cultural diffusion betweenthese groups (e.g. Mellars, 2005; Roussel, 2011; Hublin, 2012;Hublin et al., 2012a). The earliest stages of the Early Aurignacianare documented north of the Alps at Willendorf II and Gei-ßenkl"osterle c. 43e42 ka cal BP and may derive from the samesource as the Protoaurignacian encountered south of the Alps. Itsextension further west as well as further east, and ultimately intothe Levant, occurred later. Although the fossil record of the EarlyAurignacian is limited and has been yielded by the later phases ofthis industry, to date its modern nature is quite clear, which isconsistent with the later Aurignacian fossil evidence.
In addition to a clari!cation of the chronology, a better under-standing of the evolution of the various assemblages present inEurope during this crucial period of recent human evolution isessential to elucidate of the dynamic of the modern humanmigration into western Eurasia, including the possible cultural in-teractions with and eventual replacement of the local Neandertalpopulations. Palaeogenetics are also expected to provide newinsight into the biological aspects of these processes.
Acknowledgements
I am grateful to Nina D"orschner, Kathryn Fitzsimmons, DamienFlas, Marion Hernandez, Marcello Mannino, Shannon McPherron,Philip Nigst, Alyson Reid, Karen Ruebens, Marike Schreiber, PatrickSemal, Sahra Talamo, Christine Verna, Bence Viola, Nicolas Zwynsand two anonymous referees for their assistance in the preparationof this article.
References
Adams, B., Ringer, !A., 2004. New C14 dates for the Hungarian early Upper Palae-olithic. Curr. Anthropol. 45, 541e551.
Fig. 7. Synthetic table of the assemblages discussed in the text with their estimated chronological span. The colour code matches that of Fig. 1 and 3. The calibrated timeline on theright side and the 14C timeline on the left are scaled using Bronk Ramsey (2009) and Reimer et al. (2009). The d18O variations established by the North Greenland Ice-Core Project(NGRIP) are taken from http://www.gfy.ku.dk/~www-glac/data/gripdelta.dat. Cold stadials (GS) are indicated by grey horizontal bands and interstadial periods (GIS) by whitehorizontal bands. The red line indicates the time of the Campanian Ignimbrite (CI) and the blue bands the Heinrich Events 4 (H4) and 5 (H5).
J.-J. Hublin / Quaternary Science Reviews 118 (2015) 194e210206

39
Tafelverzeichnis
Taf. 1: Leitformen des Micoquiens in Mittel- und Osteruopa ...................................... 40!Taf. 2: Leitformen des MTA in Westeuropa ............................................................... 41!Taf. 3: Leitformen des Châtelperroniens .................................................................... 44!Taf. 4: Blattspitzen aus dem LRJ. Verschiedene Beisiele. ........................................ 46!Taf. 5: Leitformen des LRJ ......................................................................................... 51!Taf. 6: Leitformen des Szeletiens ............................................................................... 54!Taf. 7: Leitformen des Bohuniciens ........................................................................... 59!Taf. 8: Leitformen des Uluzziens ............................................................................... 62!Taf. 9: Leitformen des Proto-Aurignaciens ................................................................ 64!
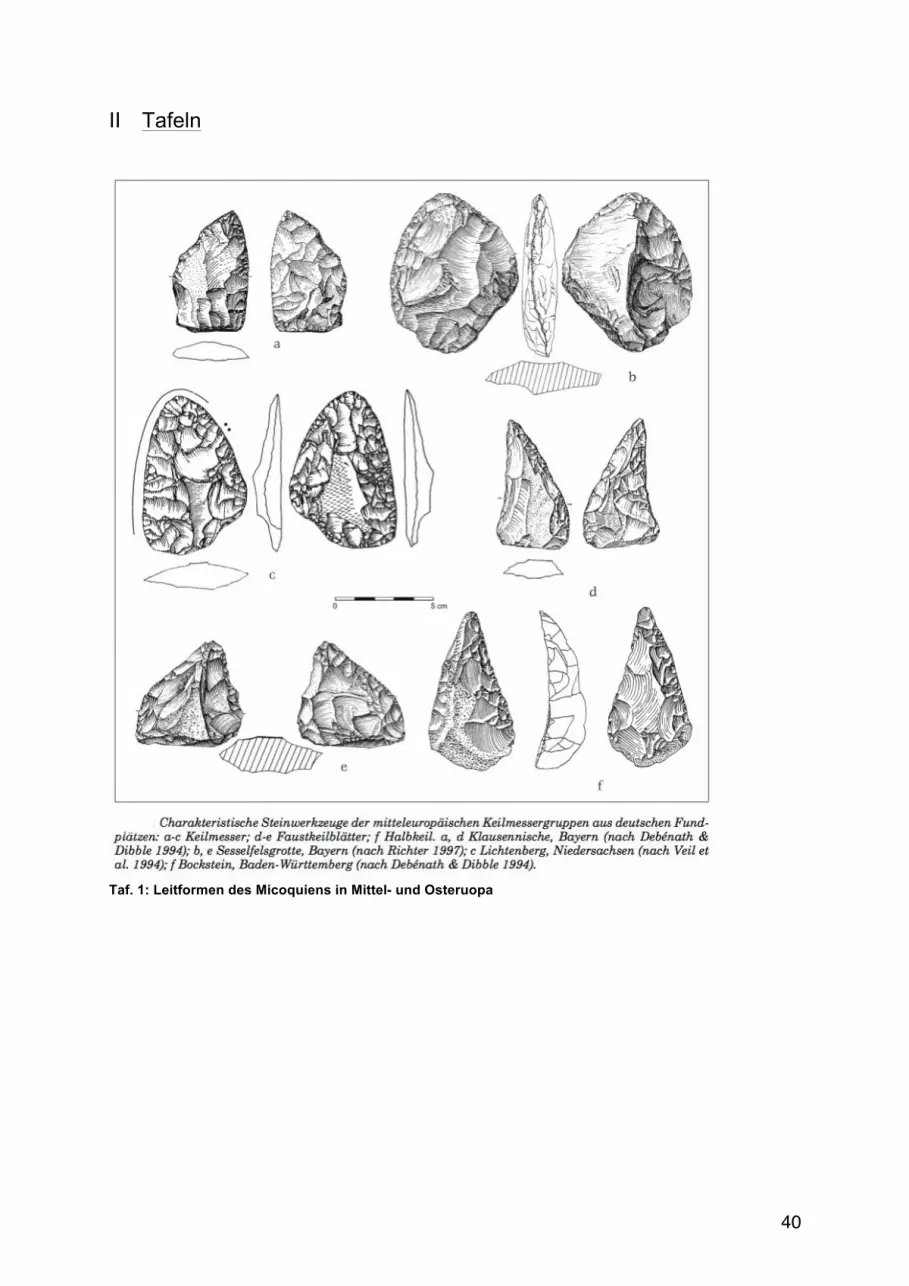
40
II Tafeln
Taf. 1: Leitformen des Micoquiens in Mittel- und Osteruopa

41
Taf. 2: Leitformen des MTA in Westeuropa

42
Inventar des Châtelperroniens aus Quinçay; Châtelperron-Spitzen Klingenschaber, Stichel und Rundkratzer.
290
Les Néandertaliens. Biologie et cultures
Figure 7. Outillage châtelperronien de Quinçay (d’après Airvaux et al., 2005)

43
Châtelperron-Spitzen, Messer mit natürlichem Rücken, Klingen und Rundkratzer aus der Grotte des Fées in Châtelperron.
284
Les Néandertaliens. Biologie et cultures
niveaux aurignaciens anciens. Seuls deux abris-sous-roche du nord-ouest du Lot faisaient exception à cette généralité : Roc de Combe et Le Piage, où F. Bordes et collaborateurs avaient cru distinguer une « interstratification » de Châ-telperronien au-dessus de témoins aurignaciens (F. Bordes et Labrot, 1967 ; F. Bordes, 1968 ; Champagne et Espita-lié, 1981). Cette occurrence tardive plaidait ainsi pour une coexistence d’une durée significative entre groupes châtel-perroniens et aurignaciens dans une même région. Mais, déjà perçues avec suspicion, ces interstratifications ont été récem-ment expliquées comme le résultat de perturbations des cou-ches archéologiques (J.-G. Bordes, 2002). Le Châtelperronien est donc toujours antérieur à l’Aurignacien, et la notion d’une longue coexistence a perdu son meilleur argument. Cette position chronoculturelle confirmée, entre Mousté-rien et Aurignacien ancien, renvoie à un âge mieux cerné
du Châtelperronien, tout au moins quant à sa terminaison (fig. 3). Ce progrès est dû à de nouvelles datations 14C, mieux étalonnées, de plusieurs niveaux d’Aurignacien ancien d’Aquitaine, qui confirment la riche et cohérente séquence de l’abri Pataud. Cet Aurignacien ancien, très cohérent dans sa culture matérielle et sa parure, s’étend de 35-34 000 à 32 000 BP : sous-jacent, le Châtelperronien ne peut qu’être plus ancien, ce que quelques récentes mesures 14C viennent d’ailleurs confirmer. Une tranche d’ancienneté 14C BP de 40 à 36 000 ans (soit 45-44 à 40 000 ans avant J.-C.) est donc acceptable pour le Châtelperronien. Dans l’ensemble, cette tranche de temps correspond aux prémisses du Würm récent, peu avant la phase très froide et sèche qui voit dans tout le sud-ouest de la France la puissante occupation de l’Aurignacien ancien, avec une faune à renne très dominant.
Figure 1. Outillage de la grotte des Fées à Châtelperron, dans l’Allier (d’après Delporte, 1999)

44
Eine vermutlich durch Impact abgebrochene Châtelperron-Spitze
Châtelperron Knochengeräte und Schmuck aus der Grotte du Renne à Arcy-sur-Cure Taf. 3: Leitformen des Châtelperroniens
289
J. Pelegrin et M. Soressi — Le Châtelperronien et ses rapports avec le Moustérien
lames à dos et des lames à retouche irrégulière, quelques burins simples (fig. 7). S’y ajoutent, moins caractéristiques car présents à d’autres périodes, des denticulés, des becs, quelques racloirs et pièces esquillées.Dans deux sites de Dordogne, un outil rare, le micro-denti-culé, a été identifié. Ces micro-denticulés sont formés, sur le bord d’un éclat ou d’une lame, d’une succession serrée de petites coches obtenues par pression à l’aide d’un autre tranchant de silex. Plutôt que d’une « scie », il pourrait s’agir d’un outil utilisé pour nettoyer, en les peignant, des fibres végétales.
Techniques et méthodes de taille des outils de silex
Le débitage châtelperronien est, dans la plupart des sites, orienté vers la production d’éclats allongés détachés parallèlement l’un après l’autre, c’est-à-dire de véritables « lames ». Ce débitage est souvent mené sur la tranche et la face inférieure d’un gros éclat, après aménagement d’une crête simple et d’un plan de frappe principal, complété d’un plan de frappe opposé. La technique de débitage recourt à des percuteurs de pierre tendre soigneusement sélectionnés, comme des galets de grès ou de calcaire tendre.Les petites lames les plus régulières sont ensuite retouchées en couteaux et en pointes d’armatures.Les lames plus larges ou plus épaisses servent pour quelques grattoirs, burins, bords retouchés ou simplement utilisés. Sur les éclats de mise en forme, épais ou minces, sont retouchés d’autres grattoirs (circulaires à front épais, minces à front plus étroit), des denticulés, racloirs, etc.
Outils en os et parure
Dans la plupart des sites où les vestiges osseux sont conser-vés, les outils en os du Châtelperronien sont plutôt rares et peu diversifiés. Il s’agit surtout de poinçons sur os ou esquilles appointés par raclage, qui existent déjà dans le Moustérien. Le site d’Arcy-sur-Cure (Yonne) fait exception, avec une riche industrie osseuse, diversifiée et élaborée : poinçons souvent entièrement façonnés, longues épingles en os, sagaies obtenues par rainurage, dont une probable en ivoire de mammouth, fins bâtonnets d’ivoire et d’os, tubes en os d’oiseau sciés au silex, etc. (fig. 8).
À La Grande Roche, à Quinçay (Vienne, fouilles F. Lévêque), 6 dents percées de renard, loup et cervidé proviennent de niveaux Châtelperroniens (Granger et Lévêque, 1997), comme il en existe à Arcy-sur-Cure en plus de dents rai-nurées, et à l’unité (et peut-être intrusives), dans quelques autres sites.
L’origine du Châtelperronien
On a vu plus haut que les débuts du Châtelperronien étaient à la fois très mal connus (dépôts archéologiques très rares ou érodés) et mal datés. Son origine reste donc incertaine, sur la base de comparaisons industrielles sujettes à caution. Pourtant, déjà en 1948, D. Peyrony voyait les racines du Châ-telperronien dans les industries moustériennes qui le précè-dent, et en particulier dans le faciès moustérien dit « de tra-dition acheuléenne » sous sa forme récente dite « B » ; non pas pour les bifaces de ce dernier, mais pour d’autres outils
Figure 6. Probable trace d’impact liée à l’utilisation en pro-jectile d’une pointe de Châtelperron d’Arcy-sur-Cure (d’après Plisson et Schmider, 1990)
291
J. Pelegrin et M. Soressi — Le Châtelperronien et ses rapports avec le Moustérien
similaires (« prémonitoires » pourrait-on dire) qu’il contient : pièces à dos rares et de forme assez variable, quelques grat-toirs, abondance de denticulés (fig. 9). Cette proposition, confirmée par les fouilles de F. Bordes au Pech de l’Azé, en Dordogne (Bordes, 1954-55), reste valable, d’autant qu’elle a été renforcée par l’analyse des intentions du débitage de certains ensembles MTA, cette analyse mon-trant que ce sont les seuls ensembles moustériens connus qui témoignent de l’intérêt pour des produits allongés et à dos (fig. 10). Nous trouvons donc des niveaux moustériens récents (de la fin du Würm ancien, vers 45 000 ans BP) dans lesquels des pièces à dos sont déjà présentes, mais avec un débitage laminaire encore inabouti, qualitativement et quan-titativement, et avec des panoplies d’outils variables dont certains vont être privilégiés et adaptés au Châtelperronien.
Figure 8. Outillage en os et parure châtelperroniens de la grotte du Renne à Arcy-sur-Cure (d’après Ba!er, 1999)
De plus, les répartitions géographiques du MTA et du Châtelperronien se superposent presque parfaitement. Le sud-est, l’est et le nord de la France ne sont pas occupés (alors que d’autres Moustériens, comme le Moustérien de type Quina ou le denticulé, y sont présents), tandis que le Centre-Ouest et le Sud-Ouest sont au cœur de la réparti-tion spatiale de ces deux industries. Par ailleurs, de récentes études technologiques ont sug-géré que le succès d’une invention et sa généralisation par adoption (devenant alors une innovation) pourraient expliquer l’ensemble de la formation de l’industrie lithi-que du Châtelperronien depuis un « fonds » moustérien. Cette invention consisterait à fixer, selon un même prin-cipe, une petite lame pointue à dos dans un manche pour s’en servir de couteau, et au bout d’une hampe pour armer la pointe de sagaies ou de piques. L’adoption d’une telle innovation rendrait alors bien compte des particularités du débitage laminaire châtel-perronien, dont l’étude technologique a justement mon-tré qu’elles cherchaient en priorité à satisfaire la produc-tion de petites lames transformables en pointes de Châ-telperron. Elle rendrait aussi bien compte, en cascade, de la transformation secondaire d’autres types d’outils du Châtelperronien, confectionnés non plus sur des éclats produits à partir de nucléus à éclats comme dans le Moustérien, mais sur des produits de second choix et des sous-produits du débitage des petites lames à pointes de Châtelperron. Le mécanisme ici évoqué n’est qu’une hypothèse. Nous ne connaissons ou ne percevons que les termes d’un changement, et pas ses « raisons » intimes qui ne sont pas directement démontrables. Mais du moins cette hypothèse est-elle parcimonieuse — elle propose de ren-dre compte de l’évolution de plusieurs caractères typolo-giques et technologiques à partir d’une seule cause, d’un même primum movens — et elle permet également de poser quelques implications prédictives que de nouveaux faits pourront satisfaire ou contredire.L’une de ces implications est le développement de l’em-manchement précédant la généralisation des pointes de projectiles. Il s’agit de multiplier les études fonctionnel-les sur la fin du Moustérien et sur le Châtelperronien pour tester cette hypothèse par la recherche d’emman-chements et de pointes de projectiles en silex. Une autre serait que cette innovation aurait pu aussi intéresser d’autres groupes moustériens que ceux du MTA B dans laquelle Peyrony voyait sa racine, en témoin de leur

45
Mauern F (nach Bosinski, 1967). 1&2 bifacielle Blattspitzen; 3 patriell bifaciell retuschierte Spitze; 4&5 Schaber.
FIG. !Mauern F (d’après BOSINSKI, 1967). 1 et 2 : pointes foliacées bifaciales ; 3 : pointe à retouche bifaciale partielle ; 4 et 5 : racloirs.
0 2 cm2
3
4
5
1
D!"#$% F&!'268

46
1&2 Jerzmanovice-Spitzen (nach Jacobi, 2007); 3 bifacielle Blattspitze aus Kent’s Cavern.
Taf. 4: Blattspitzen aus dem LRJ. Verschiedene Beisiele.
204
Damien FLAS
Fig. 17. — Kent’s Cavern. 1 et 2 : pointes de Jerzmanowice (d’après Jacobi, 2007) ; 3 : pointe foliacée bifaciale.

47
1. Retuschierte Klinge; 2. Kratzer- Stichel mit Endretusche; 3. Jerzmanovice-Spitze aus Beedings (nach Jacobi, 2007)
193
La transition du Paléolithique moyen au supérieur dans la plaine septentrionale de l’Europe
Fig. 6. — Beedings. 1 : lame retouchée ; 2 : grattoir – burin sur troncature ; 3 : pointes de Jerzmanowice, avec « flûtage » (d’après Jacobi, 2007).

48
1. Jerzmanovice-Spitze aus Balding Hills; 2. Jerzmanovice-Spitze aus Bapchild; 3&4 Klingen aus Beedings (nach Jacobi, 2007)
189
La transition du Paléolithique moyen au supérieur dans la plaine septentrionale de l’Europe
Fig. 2. — 1. Balding Hills, pointe de Jerzmanowice ; 2. Bapchild, pointe de Jerzmanowice ; 3 et 4. Beedings, lames (d’après Jacobi, 2007).

49
1&2 Stichel; abgewinkelter Stichel/Bohrer; kernförmiger Stichel aus Beedings (nach Jacobi, 2007)
195
La transition du Paléolithique moyen au supérieur dans la plaine septentrionale de l’Europe
Fig. 8. — Beedings. 1 et 2 : burins multiples ; 3 : burin dièdre : 4 : burin nucléiforme (d’après Jacobi, 2007).

50
1&2 Kratzer aus Beedings (nach Jacobi, 2007)
194
Damien FLAS
Fig. 7. — Beedings. 1 et 2 : grattoirs (d’après Jacobi, 2007).

51
1-3 Kratzer aus Ranis 2 (nach Hülle, 1977); Doppelstichel mit abgebrochener Spitze aus der Robin Hood’s Cave (nach Campbell, 1977); 4 Stichel aus der Nietoperzowa Grotte (nach Kozłowski, 1996)
Taf. 5: Leitformen des LRJ
34 Damien Fras
Elle comporte principalement trois types d'ou-tils : des pointes foliacées laminaires à retouchesplates bifaciales partielles (surtout ventrales),des burins et des couteaux de Kostenki. Or,
, 1f l
certaines pièces présentent la combinaison deces différents aménagements. On rencontre desburins sur lame à retouches ventrales plates,des burins avec troncature de type Kostenki
0 3 c mc m - l
F ig.6 - 1et3:raclo i rs,Ranis2(d 'aprèsHûIIe,1977);2:bur indoublesurcassure,RobinHood'sCave(d'après Campbell, 1977);4:buriry grotte Nietoperzowa (dâprès Kozlowski & Koztowski, 1996).

52
TrenèianskéTeplice-Plieöky, ausgewählte Spitzen aus der Prospetion von Bárta’s
Szeletian – chro nol ogy and cul tural variability 43
Fig. 10. Trenèianské Teplice-Plieöky, se lected points from Bárta’s sur face sur vey

53
TrenèianskéTeplice-Plieöky, ausgesuchtes Material aus der Kaminská’s Grabung
44 L. Kaminská et al.
Fig. 11. Trenèianské Teplice-Plieöky, se lected ma te rial from Kaminská’s ex ca va tion

54
Éeleöice-Hoynerhüge, ausgesuchtes Material. 1-15 Material aus der Grabung von 2010; 16-31 Flächenfunde.
Taf. 6: Leitformen des Szeletiens
34 L. Kaminská et al.
Fig. 3. éeleöice-Hoynerhügel, se lected ma te rial. 1-15: ma te rial from 2010 ex ca va tion; 16-31: sur face col lec tion

55
Ausgesuchte lithische Artefakte aus dem unteren Paleosol aus Brno-Bohunice 2002 Grabung. 1-3 Blattspitzen; 4-14 Levallois Produkte; 15-19 Kratzer; 20, 24: Stichel; 21 zusammenlaufender Schaber; 22 bifaciell retuschierter Schaber; 23, 25, 26, 28 Kerne; 27 Kernkantenklinge.
stratigraphic section of the 2002 excavation also corroborates thestratigraphic picture seen at the Stranska skala localities. In general,the geology of the site is well-studied as it is directly adjacent toa classic Pleistocene geological profile (Damblon et al., 1996): themodern soil overlies a later Upper Paleolithic loess stratum, which
overlies two paleosols, the Upper and Lower Last Interpleniglacialsoils (indicated by lines in Fig. 3), about 30 cm and 30–50 cm thick,respectively, above a thick loess deposit.
Five hypotheses of site formation processes are systematicallyevaluated and might explain the apparent differences between the
Fig. 2. Selected lithic artifacts from the Lower Paleosol assemblage from the Brno-Bohunice 2002 excavation: 1–3) leaf points; 4–14) Levallois products; 15–19) endscrapers; 20, 24)burins; 21) convergent sidescraper; 22) bifacially-retouched scraper with refitted flake; 23, 25, 26, 28) cores; and 27) crested blade.
D. Richter et al. / Journal of Human Evolution 55 (2008) 871–885876

56
Ausgesuchte lithische Artefakte aus dem unteren Paleosol aus Brno-Bohunice 2002 Grabung. Rohmaterial Bestimmung: Stránska skála Feuerstein: 1-2, 7-13, 15-16, 18-19. Krumlovský les Feuerstein: 3-4, 14, 17. Radiolarit: 5-6.
36
Gilbert B. Tostevin, Petr !krdla
FIGURE 2. Selected artifacts from the 2002 assemblage from Brno-Bohunice Lower Paleosol. Raw material determinations: Stránská skála chert: 1–2, 7–13, 15–16, 18–19. Krumlovsk! les chert: 3–4, 14, 17. Radiolarite: 5–6.

57
Jungpaläolithische Blattspitzen und Rohlinge von Blattspitzen aus Eger-Kőpros.
4.2. Tren!cianska Turná-Vrla!cka 2
At this site, four trenches were dug: trench S2 in the saddlebetween sites 1 and 2, and three trenches (from east to west: S3, S1and S4) on the slope, above the saddle, transversally to the axis ofthe hill where site 2 is situated. The pro!les show grey-reddish-brown loess (designated as layers 4 and 4a in trenches S3, S1 andS4), with well developed oxidationereduction processes. The meansize (Mz) of the sediments is about 64, badly andmoderately sorted(Fig. 16).
Thin section sample M5 shows subangular and channel micro-structures. Quartz grains in coarse material are weathered andcracked. The groundmass is mainly brown-gray but in zones withiron oxide, the groundmass is reddish. Within some !ssuressecondary calcium carbonates appear, in the form of sparite ormicrite, connected with freeze and thaw cycles (Fig. 17). These
forms of the calcium carbonate !ll all channels and encrust walls ofvoids (Van Vliet-Lanoë, 1985; Haesaerts and Mestdagh, 2000;Mroczek, 2008). In the thin section, round micromasses representtraces of mass movement in this horizon. Various-sized iron andiron-manganese nodules commonly occur. Fragments of very welldecomposed organic matter appear in the groundmass, withpreserved structure of tissues.
This formation shows the occurrence of angular and roundedquartz grains and exhibits micromorphological features indicatingfrost sorting. In all likelihood, this is a Bt horizon of the Eemian soiltransformed by frost processes in the Vistulian, and corresponds tolayers 4 and 4a at site 1 (trenches S1, S2, S4 and S5).
As at site 1 at site 2, the Bt horizon is cut by erosion andmantledby a layer of typical loess (layer 2). The typical loess layer !lls thelarge ice-wedge casts, up to 0.7 m wide and up to 1.2 m deep, thatcut the Bt horizon (layer 2b in trenches S4 and S1, possibly also S3).
Fig. 13. Upper Palaeolithic leaf points and preforms of leaf points from Eger-K}oporos.
A. Budek et al. / Quaternary International 294 (2013) 82e9892

58
Mittelpaläolithische Artefakte aus der Eger Region: 1, 5, 7 Egerszalók-Kővágó; 2, 3, 4, 6 Eger-Kőpros; 1, 3 typisches Moustérien, 4, 5 Moustérien vom Quina Typus, 6, 7 Micoquien (1 Moustérien Spitze; 2 transversaler Schaber; 3, 5 convexe Schaber; 4 bifacielle retuschierte Schaber; 6 Keilmesser; 7 bifacielles Gerät.
the Late Pleistocene were covered by loesses and decalci!ed loamyloesses (Lukni!s a kol., 1972). The Tren!cín microregionwas attractivefor settlement for several reasons. The terraces were an excellentstrategic position for Palaeolithic hunters, and the river bedprovided good quality raw materials, especially radiolarites whoseprimary deposits are situated at a small distance from Tren!cín(Cheben et al., 1995). The Vlára mountain pass enabled easy contactwith the territory of Moravia (!Skrdla, 2005), and the Moravian Gateopened the route to Poland. In the eastern direction, the Jastrebskésedlo mountain pass enabled contact with the valleys of theBobrava and the Nitra rivers.
In the Tren!cín region, Palaeolithic sites, some of them multi-phase settlements, have been known for a long time. The earliestinformation about Palaeolithic sites can be found in the works byBabor (1927) and Skutil (1938). The Middle Palaeolithic Micoquian(previously belived to be the Szeletian e Pro!sek, 1953) wasdiscovered at Zamarovce. The industries from the transition
between the Middle and the Upper Palaeolithic were identi!ed atIvanovce-Skala (Pro!sek, 1953) and at Tren!cín IV (Bárta, 1965a). In2009, a Szeletian site in Tren!cianske Teplice (Kaminská, in press),earlier designated as Vel’k! Kola!cín (Bárta, 1974, p. 16) wasinvestigated.
Most Palaeolithic traces are related to the Gravettian. SporadicGravettian !nds come from Nem!sová (Bárta, 1961, p. 16; Chebenand Kaminská, 2002), and provided a radiocarbon date of28,570 ! 1,345 BP (GrN-2470 e Bárta, 1965b, p. 127). The richestEarly Gravettian site is Tren!cianske Bohuslavice, excavated by Bárta(1988) in the years 1981e1986. Radiocarbon dates for this site arewithin the time span between 25,650 ! 160 BP (GrA-16162) and20,300 ! 500 BP (Gd-4011) (Verpoorte, 2002). A speci!c feature ofthe Late Gravettian from Tren!cianske Bohuslavice is the occurenceof leaf points with an absence of shouldered points (Koz1owski,1996).
In recent years, a number of new sites have been discovered inthe southern part of the Tren!cín microregion: Tren!cianske Stan-kovce, Tren!cianska Turná, Mníchova Lehota, mainly thanks to thecollections of Michalík (2003, 2004, 2006, 2007). The newly-discovered sites yielded mainly Gravettian and Epigravettian arte-facts, and individual items with bifacial retouch. In the excavationsconducted in 2007 at the sites of Tren!cianska Turná-Vrla!cka 1 and2, Tren!cianska Turná-Hámre, Tren!cianske Stankovce III and V, andMníchova Lehota, stratigraphic data were obtained and occupa-tional episodes were distinguished (Kaminská et al., 2008). Some ofthe results from these excavations are presented in this paper.
4.1. Tren!cianska Turná-Vrla!cka 1
In trench S3, situated on an elevation, the lowermost layerrepresents a Bt horizon of fossil soil. In thin sections M1 and M2,channel and subangular blocky structures occur (Table 1). Thegroundmass contains mainly rough and cracked quartz grains, butrare grains are fresh and rounded. Thin plates of muscovite andbiotite are rare. Wetting and draining of sediments producedmosaic and granostriated b-fabric. Clay and silt coatings in channelsare very well developed. The coatings are small microlayers indi-cating polycyclic development of the clay illuviation (Kemp, 1985,1998; Haesaerts and Mestdagh, 2000; Budek et al., 2004; Kühnet al., 2006; Mroczek, 2008) (Fig. 15). These pedofeatures arecharacteristic of the Bt horizon of luvisols.
Disrupted clay coatings in channels and development of papulesare connected with the destruction of the Bt horizon (Kemp, 1985;Mroczek, 2008). In some cases, voids are !lled with grey, coarsermaterial. This in!lling comes probably from the destroyed eluvial(Eet) horizon. The groundmass has typical iron nodules with quartzgrains inside. Ferruginous nodules commonly have iron and claycoatings and hypocoatings. In channels, moderately decomposedorganic matter has been occurred. This fossil soil is truncated byerosion and overlain by a laminated slope deposit.
Within the Bt horizon, in the cold climate of the Vistulian, frostsorting of different sized material took place (unstrati!ed brownloess e layer 4a from trench S4). On the southern slope (sample M3from trench S2) all pedofeatures indicate the existence of Bt horizonof the luvisol. In channels and voids, well developed traces of claymovement have been preserved. Diagenetic (oxidation–reduction) isre"ected as ring-shape and diffuse ferrous-manganese nodules. Thislayer was cut by a large ice-wedge cast observable in the pro!le oftrench S4. The formation of this ice wedge can be placed at themaximum of the Last Glaciation.
Level Bt, transformed by cryogenic processes, is overlain bya layer of typical loamy loess, yellow, up to 40 cm thick, in trenchesS3, S4 and S5. This formation disappears in the eastern direction.Loess !lls the large icewedge cast in pro!le S4 (layer 2b) and covers
Fig. 11. Middle Palaeolithic from the Eger region: 1,5,7 e Egerszalók-K}ovágó; 2,3,4,6 e
Eger-K}oporos; 1e3 e Typical Mousterian, 4,5 e Quina type Mousterian, 6,7 e Mico-quian (1 e Mousterian point, 2 e transversal side-scraper, 3,5 e convex side-scrapers,4 e bifacially retouched side-scraper, 6 e Keilmesser, 7 e bifacial tool.
A. Budek et al. / Quaternary International 294 (2013) 82e9890

59
Stránská skála III. The sub-prismatiche Methode. 1 Zusammensetzten eines Kerns (A) und Abschläge/Klingen (a-n); 2 Zusammensetzten eines Kerns (A1+A2) und teilweise fertiggestellte Klinge (B).
Taf. 7: Leitformen des Bohuniciens
nounced exploitability of the cores, approaches to the preparation of the raw material arenot well documented. Their character is best attested by a prepared, non-exploited corewith a worked crest blade on the narrow edge of a raw material frost fragment (fig. 3: 3).
In the refittings this approach is best shown by the reffiting of a uni-directional core witha final blade and secondary crest blade that, within the framework of the core reduction, isevidently secondary (reparatory; fig. 3: 2). The original core (A) was larger, and primarilybi-directional. The removal of a series of flakes, however, changed the morphology of the
NERUDA – NERUDOVÁ: The development of the production …270
Fig. 2. Stránská skála III. The sub-prismatic method. 1 – refitting of a core (A) and flakes/blades (a-n);2 – refitting of a core (A1+A2) and step terminated blade (B). – Obr. 2. Stránská skála III. Subprizmatickámetoda. 1 – skládanka jádra (A) a ú!t"p#/$epelí (a-n); 2 – skládanka jádra (A1+A2) a $epele zab"hlév t"%ní plo!e (B).

60
Knochengeräte aus der Grotta del Cavallo (1-8), Grotta della Cala (9) und Grotta di Castelcivita (10-14)
Fig. 2. Uluzzian bone tools from Grotta del Cavallo (n. 1e8), Grotta della Cala (n. 9), and Grotta di Castelcivita (n. 10e14).
F. d’Errico et al. / Quaternary International 259 (2012) 59e7162

61
Eine Auswahl an lithischen Artefakten aus der Grotta del Cavallo

62
Eine Auswahl an lithischen Artefakten aus der Grotta Fabbricia: 1-4, 7-8: Kratzen; 5,6: Pièces esquillées; 11: Unipolarer Nukleus.
Taf. 8: Leitformen des Uluzziens

63
Font-Yves-Lamelle
����
��
ϩ�
ϙϪϽ�
ϒ� 0�
)LJ�� �� �� ᕤV� �� ¢� ��� �� SRLQWHV� GH� )RQW��<YHV� GH� OD� JURWWH� «SRQ\PH�� /HV� VHFWLRQV� GHV� SLªFHV� VRQW� SULVHV� ¢� PL�KDXWHXU� �� HOOHV�QH� UHSU«VHQWHQW� SDV� IRUF«PHQW� O«SDLVVHXU� PD[LPDOH� GRQQ«H� SDU� OH� WDEOHDX� ,��
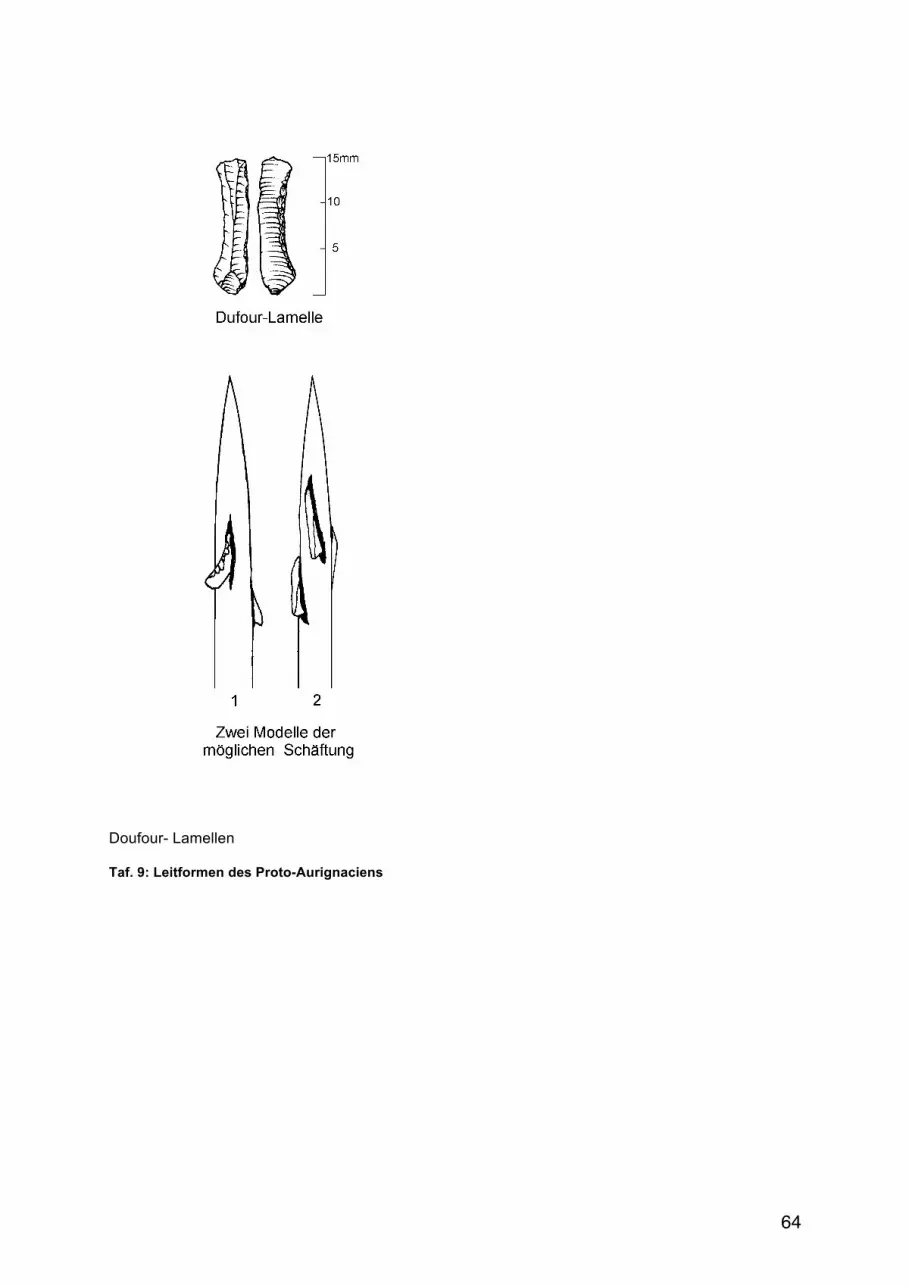
64
Doufour- Lamellen Taf. 9: Leitformen des Proto-Aurignaciens
15.07.15 04:43Modell-Dufor-Lam.JPEG.jpg 2.373×3.086 Pixel
Seite 1 von 1http://altsteinzeit-hessen.de/wp-content/uploads/2013/12/Modell-Dufor-Lam.JPEG.jpg

65
Tabellenverzeichnis Tab. 1: Datierung von Heinrich Ereignissen .............................................................. 65!Tab. 2: Radiometrische Datierungen für das MTA ..................................................... 66!Tab. 3: Kalibrierte Daten in Bezug auf die Chronologie des Szeletiens. Tab.3 bei Neruda/Nerudová (2013). .......................................................................................... 66!Tab. 4: Materialen (1-8) gruppiert nach Erhaltungschancen ...................................... 67!Tab. 5: Übersicht der wichtigsten stratigraphischen Sequenzen des Aurignacien, aufgeteilt in 13 Regionen. Die Tabelle zeigt in welcher Region das Aurignacien in welchem Stadion seiner Entwicklung vorzufinden ist und ermöglich somit eine Chronostratigraphie dieser Industrie. Diese Tabelle wurde nach den Angaben von Djindjian (Vgl. 2010: 21-25) zusammengestellt. ........................................................ 69!Tab. 6: Neue Datierungen für den Anfang und das Ende des Uluzziens. Resultate sind in cal. BP widergegen. ........................................................................................ 70!Tab. 7: Datierung der Fundschichten in der Grotte von Esquilleu ............................. 70!Tab. 8: Darstellung der Stratigraphie von Saint-Césaire mit TL Datierung der verschiedenen Level .................................................................................................. 71!Tab. 9: Stratigraphische Zuordnung der kulturellen Abfolge ...................................... 72!Tab. 10: Eine Übersicht verschiedener Datierungen für das Moustérien und das Uluzzien in der Grotta di Fumane .............................................................................. 72!Tab. 11: Anzahl der identifizierten Taxa bei den Huftieren. Mammut nicht mit in der Berechnung inbegriffen. ............................................................................................. 72!Tab. 12: Anzahl der identifizierten Taxa bei den Karnivoren ..................................... 72!
III Tabellen
Tab. 1: Datierung von Heinrich Ereignissen
estimates of IRD concentration in the ice and glacialogicalprocesses, and using their mapped volume of H1 and H2,Dowdeswell et al. [1995] suggested 1.4 ! 105 to 1.4 !106 km3 of water released over 250–1250 years (0.39–3.9 m sea level rise).
5.2. What Caused the Heinrich Layers?
[57] A successful Heinrich layer model, that is, specifi-cally for the Hudson Strait Heinrich layers, must producesufficient IRD and distribute it across the North Atlantic[Hulbe, 1997]. It must explain the limited source region forthe terrigenous sediments and the presence of abundantdetrital carbonate only within the Heinrich layers. It mustaccount for the apparent association with the cold phases ofthe D-O cycles, with the dramatic ocean circulationchanges, and with the rapid warming that directly followsthem. Finally, if Hudson Strait Heinrich events accompa-nied global mountain glacier advances [e.g., Denton et al.,1999], then they must be interpreted in the context of globalclimate change. Three proposed mechanisms to model theIRD and freshwater delivery of Hudson Strait Heinrichlayers are reviewed here: catastrophic ice sheet purging,jokulhlaup activity, and ice shelf buildup/collapse. Anadditional model that involves a sea ice switch to triggerthe abrupt warming (Y. Kaspi et al., A ‘triple sea-ice state’mechanism for the abrupt warming and synchronous icesheet collapses during Heinrich events, submitted to Nature,2003, hereinafter referred to as Kaspi et al., submittedmanuscript, 2003) is described briefly after the discussionof glacialogical mechanisms.[58] MacAyeal [1993] formulated an internally driven
‘‘binge-purge’’ model. The premise of a binge-purge modelis that a large ice sheet will build up gradually during thebinge stage, dependent on air temperature and moisturesupply. Some combination of geothermal heat, advection ofheat from the upper surface, and internal friction in thebottom boundary zone at the base of the ice sheet will act toeventually destabilize the ice sheet, resulting in a rapidpurge phase, also known as Heinrich events. MacAyeal[1993] assumed geothermal heating is the driving forcefor purging the interior of the ice sheet, while Verbitsky andSaltzman [1995] found friction and heat advection from thesurface to be more important parameters. Marshall andClarke [1997] provided a continuum mixture model be-
tween sheet ice, which deforms by viscous creep, andstream ice with fluxes dictated by basal sliding and sedi-ment deformation. They used this model to test the sensi-tivity of a Hudson Strait ice stream and concluded that it isunlikely for an ice stream in Hudson Strait to drain theinterior portions of the ice sheet. Clarke et al. [1999]concluded that it is likely that the onset of a Heinrich eventoccurs when glacier flow instability is triggered, and theyfavor episodic surging of an ice stream in Hudson Strait. Analternative purge mechanism was proposed by Hunt andMalin [1998], that is, that ice-load-induced earthquakes mayhave destabilized the Laurentide ice sheet (although theycall for successively shorter intervals between the Heinrichevents, whereas my reassessment of the timing (Table 4)suggests a nonchanging, 7 kyr spacing).[59] The volume MacAyeal [1993] estimated is 1.25 !
106 km3 of fresh water (he estimated an area of 1 ! 106 km2
with an average thickness of 1250 m), introduced into theNorth Atlantic over about 250–500 years, implying a 0.16–0.08 Sv flux. Dividing the volume by the area of the worldocean, 3.61 ! 108 km2, MacAyeal estimated a sea level riseof about 3.5 m. If the Heinrich events represent a purging ofa very large part of the Laurentide ice sheet, then it wouldtake an interval considerably longer than a single D-O cycleto rebuild the ice sheet and create the conditions necessaryto trigger a purge. The binge interval estimated fromMacAyeal’s model was about 7 kyr (remarkably close tothe recurrence time of Heinrich layers, see Table 4). Thewater volumes and event durations calculated by MacAyeal[1993] abide by the maximum constraints discussed above.Marshall and Clarke [1997] modeled the behavior of an icestream in Hudson Strait using mixed bed conditions, andthey were able to produce a maximum flux of 0.03 Sv oficebergs, which is about 10 times lower than that implied bythe d18O-based calculations given above. One possibleexplanation they proposed is that their ice streams do nottap deep enough into the core of the ice sheet. However,because the maximum ice stream velocity in their simula-tions is 900 m yr"1, ice originating at the head (750 km in)takes 833 years to get to the outlet. Accordingly, Marshalland Clarke [1997] concluded that it is unlikely for an icestream in Hudson Strait to drain the interior portions of theice sheet. They left open the possibility that their omissionof hydrological conditions under the ice sheet may have led
TABLE 4. Ages of Heinrich Layersa
Heinrich Layer Calendar Age, years Basis Source
H1 16800 14C#14200$ 2600% Bond et al: &1992; 1993'H2 24000 14C (20500 + 3500) Bond et al. [1992, 1993]H3 (31000 correlation to Greenlanda GISP2 [Meese et al., 1997]H4 38000 correlation to Greenlanda GISP2 [Meese et al., 1997]H5 45000 correlation to Greenlanda GISP2 [Meese et al., 1997]H6 (60000 correlation to Greenlanda GISP2 [Meese et al., 1997]
aBond et al. [1993] set the basis for making correlations between North Atlantic records and Greenland ice core d18O. The ages from H3 through H5 arebased on this strategy, but the ages are different and are based on current age estimates. The ages for H1 through H4 are compatible with those derived fromU-Th of dated carbonate correlated to GISP2 based on d18O pattern matching the Hulu (China) speleothem record [Wang et al., 2001]. That for H5 wouldbe 47,000 with the correlation to the Hulu speleothem record [Wang et al., 2001]. The position of H3 is more difficult to discern, and thus the age isuncertain but probably is within ±1000 years. The age of H6 is very poorly constrained and could be different by at least ±5000 years.
RG1005 Hemming: HEINRICH EVENTS
29 of 43
RG1005

66
Tab. 2: Radiometrische Datierungen für das MTA
Tab. 3: Kalibrierte Daten in Bezug auf die Chronologie des Szeletiens. Tab.3 bei Neruda/Nerudová (2013).
9
La lim itation de l’extension de sites MTA à la partie occiden tale de la France est difficile-m ent explicable. Dans la Vallée du Tarn par exem ple où les industries MTA sont fréquentesen haute et m oyenne vallée m ais absen tes dans la basse vallée, cette distribution géogra-phique ne sem ble pas pouvoir être attribuée n i à la disponibilité des m atières prem ières n ià la géom orphologie régionale (Tavoso, 1978 : 360, 1984). A. Turq propose d’expliquer l’ab-sence de sites MTA dans le sud-est de la France, du m oins pour partie, par la nécessitéd’avoir des blocs de dim ensions raisonnables pour produire les pièces bifaciales MTA (Turq,2000 : 389).
Position chronologique
La chronologie relative perm ettait de penser que le MTA du sud-ouest de la France sepositionnait principalem ent à la fin de la prem ière période du Würm ancien ; le MTA detype B se rencontran t jusque dans l’in terstade würm ancien / würm récen t (Bordes, 1984 :137). Les résultats des datations radiom étriques on t perm is de préciser la chronologie rela-tive dans le sud-ouest de la France ; leurs résultats son t indiqués dans le tableau I 1. Elles sesituen t toutes dans le stade isotopique trois ou à la lim ite du stade isotopique quatre et troispour le n iveau C de la Grotte XVI. Les dates effectuées dans le cadre de ce travail von t dansle m êm e sens ; elles seron t présen tées en détail dans le chapitre suivan t.
LE MOUSTÉRIEN DE TRADITION ACHEULÉENNE : RAPPEL DE L’ÉTAT DES CONNAISSANCES ET DES INTERPRÉTATIONS
!"#$%"&' (' )*' +",",-./0' 1"2-.34,1-5&%0' 2-06./-#$%0' 6.&1' 2%0' /-7%"&8' 9!:' ;$%0' 2",%0' 14"$-04%0' 2"/0' $%' <"21%' 2%' <%
,1"7"-$=' -/2-5&4%0' 2"/0' <%' ,"#$%"&' 0.&0' $"' >.13%' 2?@A%0' 3.B%/0=' 0%1./,' 24,"-$$4%0' 2"/0' $%' <C"6-,1%' 0&-7"/,D
!"#$%&'()('$*+$,% -(#./0$% 12#$%
32%45"*2%&6.2'$*#$7%-$'+"$'%$#%8299202:7%;<<=,%
*">$25%?2%&25@0$::5:%05%*">$25%-AB,%
*">$25%=%&25@0$::/5:%05%*">$25%-AB,%
A3%:5'%:"9$C% *D%?2%E%FG%FGG%!H?GG7%FI%IGG%!F?GG%
*D%=%E%FF%IGG%!FJGG7%IH%GGG%!IGGG%
K$+.@0$@9LBM(%N8%&1/'0/O*$7%P/QR2*7%;<=J,% A3%:5'%:"9$C% FH%HIG%!H<GG%&SO$%R/T$*,%
P2'U2:%NNN7%6F%&1/'0/O*$7%P/V02%!"#$%D7%;<<?,% 6;F%B-!% H=%HGG%!IGG7%FH%IGG%!JJGG%
W/*:$"O*$'7%1%:5X%&1/'0/O*$7%8299202:%!"#$%D7%;<=Y,% A3%:5'%:"9$C% IG%IGG!IHGG%&SO$%R/T$*,%
Z'/##$%[8N7%6%&1/'0/O*$7%Z5"U$'#%!"#$%D7%;<<<,% A3%:5'%:(0"R$*#% ?F%?GG%!H;GG%&SO$%R/T$*,%%
3$%-/5:#"$'7%Z%&1/'0/O*$7%8299202:%!"#$%D7%;<=Y,% A3%:5'%:"9$C% IG%HGG%!IIGG7%II%=GG%!IGGG%&SO$%R/T$*,%
3$%-/5:#"$'7%Z%&1/'0/O*$7%-$992':%$#%Z'\*7%;<<;,% ]!^% FH%GGG%!JHGG7%FY%GGG%!JIGG%&SO$%R/T$*,%
3$%-/5:#"$'7%_%&1/'0/O*$7%8299202:%$#%29D7%;<=Y,% A3%:5'%:"9$C% FJ%IGG%!JGGG7%F?%HGG%!HGGG%&SO$%R/T$*,%
3$%-/5:#"$'7%_%&1/'0/O*$7%-$992':%$#%Z'\*7%;<<;,% ]!^% H<%YGG%!JFGG7%F;%GGG%!J?GG%&SO$%R/T$*,%
K$+.@0$@9LBM(%N7%Y%&1/'0/O*$7%`/*$:%!"#$%D7%>/"'%+.2X"#'$%NN,% ]!^% FJ%GGG%!=GGG%&]a,7%F<%GGG%!YGGG%&3a,%
K$+.@0$@9LBM(%N7%?%&1/'0/O*$7%`/*$:%!"#$%D7%>/"'%+.2X"#'$%NN,% ]!^% H<%GGG%!JGGG%&]a,7%FY%GGG%!FGGG%&3a,%
K$+.@0$@9LBM(%N7%I%&1/'0/O*$7%`/*$:%!"#$%D7%>/"'%+.2X"#'$%NN,% ]!^% F<%GGG%!?GGG%&]a,7%I;%GGG%!YGGG%&3a,%
32%^/+.$##$7%Y%&1/'0/O*$7%1$%8'"$:%$#%b2#$'U/9c7%;<I=,% 6;F% H?%GGG%!IGG7%HG%YGG%!FGG%
32%^/+.$##$7%Y%&1/'0/O*$7%8299202:%!"#$%D7%>/"'%+.2X"#'$%NN,% 6;F%B-!% IJ%IGG%!HFGG%
!

67
Tab. 4: Materialen (1-8) gruppiert nach Erhaltungschancen

68
Regionen
Chronologie Bemerkung
Region 1: Nordwest-Europa • Großbritannien: Kent’s Cavern;
Paviland Cave (Wales) • Belgien: Trou Walou, Trou
Margeritte • Frankreich: Grotte du Renne, Les
Cottés, Quincay
∼31.000-29.000 BP Zwischen 38.000 und 33.000 BP sind in dieser Region mehrere Übergangsindustrien vorhanden. Das Lincombien in Großbritannien, das Ranisien in Belgien und Deutschland sowie das Châtelperronien in Frankreich.
Region 2: Westeuropa (Frankreich, Spanien) • Charente: St.-Césaire, La Quina • PErigord/ Quercy: La Ferrassie,
Pataud, Le Facteur, Roc-de-Combe, Caminade, Trou de la Chèvre, Le Flageolet, Le Piage, La Rochette, Grotte 16, Combe-Saunière, Castanet
• Pyrenäen und Kantabrien: Brassembouy, Gatzarria, Isturitz, Labeko Koba, Cueva del Pendo, Cueva Morin, Cueva del Castillo, La Vina
• Châtelperronien ab 38.000-35.000 BP
• Kein Aurignacien 0 vorhanden • Aurignacien I ab ∼ 35.000-31.000
BP • Aurignacien très ancien ab
∼36.500 BP (Gatzzaria, Isturitz, Cueva Morin 8, Castillo 16, Vina XII)
• Aurignacien I ab 35.000-32.000 BP
In dieser Region existiert vor dem Aurignacien I eine Übergangsindustrie, das Châtelperronien. Die Datierungen und Stratigraphien der Fundplätze in Spanien müssen überarbeitet werden. In Castillo soll es eine Evolution aus dem lokalen Moustérien ab 40.000 BP geben.
Region 3: Westlicher Mittelmeerraum • Frankreich: Abri Mochi (Ligurie),
Grotte Rainaude, Grotte Mandrin (Provence), l’Esquicho-Grapaou, la Laouza (Languedoc Ost), Grotte de Bize (Langedoc West).
• Spanien: l’Arbedra, Abri Romani, Reclau-Viver (Katalanien)
Wahrscheinlich ab 36.500 BP Bisher sind keine zuverlässigen 14C Datierungen vorhanden. Muss überprüft werden.
Region 4: Nordeuropa (Niederlande, Dänemark, Nordost Deutschland, Polen)
• Jerzmanowicien (Übergangsindustrie) vorhanden
• Kein Aurignacien ancien oder très ancien vorhanden
• Aurignacien évolué ab 31.000 BP (Breitenbach in Türingen, Krakow-Spadzista C, Oblazowa in Polen)
Region 5: Zentraleuropa • Deutschland: Geißenklösterle
(Schwäbische Alb), Wildscheuer (Rheinland)
• Österreich: Willendrof II, Seftenberg, Krems
• Frankreich: Solutre (Saône Becken)
• keine Datierung vorhanden für das Aurignacien
• Gravettien beginnt schon kurz nach 30.000 BP
In dieser Region gab es keine Übergangsindustrie zwischen dem Moustérien (Micoquien) und dem Aurignacien. Es wurden drei Sequenzen ausgemacht, das Aurignacien trèse ancien, ancien (I) und récent (II)/l’Olchévien. Ausbreitung des Aurignacien ancien in Richtung Westen über das Rhone Tal, danach nach Norden in das Rheinland (Wildheuer, Lommersum). Als A. évolué (II) geht es dann in Richtung Belgien und England. Des Weiteren breitet sich das A. évolué nach Westen in das Saône Becken aus, wo es auf das Châtelperronien folgt (Germolles, Arcy-sur-Cure, Châtelperron).
Region 5 bis: Zentraleuropa • Ungarn: Szeleta, Tata, Istallosko • Mähren: Stránská skálá, Milovice,
Pod Hradem, Verdrovice, Mladec • Slowenien: Certova Pec
Keine aktuellen 14C Datierungen vorhanden für das auf die Übergangsindustrien folgende Aurignacien. Alte 14C Datierungen von Istallöskö und Peskö verworfen
In dieser Region waren zwei Übergangsindustrien vorhanden, das Bohunicien in Bohunice und Stránská skálá sowie das Szeletien in Szeleta, Tata, Certova Pec, Vedrovice V.
Region 6: Adriatisches Becken • Italien: Fumane (Venetien),
Paglici (Apulien)
• Aurignacien ancien ab 34.000 BP (Paglici)
• Gravettien ancien ab 28.300 BP
In Fumane sind zwei Phasen vorzufinden, ein Aurignacien très ancien und ein Aurignacien ancien.

69
Tab. 5: Übersicht der wichtigsten stratigraphischen Sequenzen des Aurignacien, aufgeteilt in 13 Regionen. Die Tabelle zeigt in welcher Region das Aurignacien in welchem Stadion seiner Entwicklung vorzufinden ist und ermöglich somit eine Chronostratigraphie dieser Industrie. Diese Tabelle wurde nach den Angaben von Djindjian (Vgl. 2010: 21-25) zusammengestellt.
Region 7: Nordosteuropa Kein Aurignacien vorhanden In dieser Region sind weitere Übergangsindustrien vorhanden. Das Streletien, welches sich zum Streletien récent mit Aurignacien ähnlichen Zügen weiterentwickelt, und im Ural ist eine Kultur namens Uralien/Ouralien vorhanden. Datierungen müssen bestätigt werden.
Region 8: Osteuropa • Don Becken: Kostenki (Russland) • Dniepr Becken: Mira,
Kulychchivka (Ukraine) • Becken des Dniepr und des Prut:
Mitoc Malu Galben, Ripiceni Ivzor (Rumänien)
Aurignacien erst ab 32.600 BP In dieser Region sind jungpaläolithische Industrien vorhanden, die in anderen Gebieten nicht vorzufinden sind. Des Weiteren findet man dort mehrere Übergangsindustrien. Im Nordosten Europas findet man das Streletien (Kostienki, Soungir) und im Südosten das Gorotsovien (Kostienki, Mira). Weitere zum Bohunicien vergleichbare Industrien (Levallois Klingen Technik) befinden sich in Bulgarien, Moldavien und Rumänien (Ripiceni-Izvor) sowie der Ostukraine (Kulychivka, Korolevo).
Region 9: Südost-Europa • Bulgarien: Bacho-Kiro, Temnata
• Moustérien • Bacho-Kirien (Übergangsindustrie)
38.500-34.800 BP • Aurignacien 33.300-29.150 BP • Gravettien
Das Bacho-Kirien ist eine Übergangindustrie mit einer Levallois-Klingen-Technik. Sie ähnelt den Industrien aus Mähren und der Ukraine.
Region 10: Iberische Halbinsel Gibraltar: Gorham Cave
Aurignacien évolué ab 30.250 BP In dieser Region waren weder Übergangsindustrien noch das Châtelperronien vorhanden, dafür überdauerte das Moustérien länger als in allen anderen Gebieten. In Andalusien und Portugal war es bis 28.000 BP präsent. In der Gorham Cave auf Gibraltar gibt es die einzige zuverlässige Sequenz für das Aurignacien auf der iberischen Insel.
Region 11: Tyrrhenische Küste in Italien • Kampanien: La Cala, Castelcivita
• Uluzzien bis ∼ 34/33.000 BP • Ankunft des Aurignacien ab
∼34.000 BP
In dieser Region war eine Übergangsindustrie (Uluzzien) vorhanden, welche vom Aurignacien abgelöst wurde. Das Aurignacien breitete sich an der Küste nach Süden aus.
Region 12: Balkan Region Griechenland: Klissoura Grotte 1
Aurignacien ancien ab 34.700 BP-22.000 BP
In dieser Region war vor dem Aurignacien das Uluzzien vorhanden. Das jung ausfallende Datum ist eine Folge von Kontaminierung der Proben.
Region 13: Krim • Siuren 1, Buran-Kaya III
• Moustérien wahrscheinlich bis 28.000 BP
• Jungpaläolithische Industrie mit Aurignacien Zügen ab 29.550 BP (?)
Auf der Krim war das Moustérien weit verbreitet und hat bis 28.000 BP angedauert. Die junge Datierung ist dennoch evtl. die Folge von Kontaminierung. Zwischen den Schichten in Siuren 1 kam es zu Vermischungen, somit fallen die Daten evtl. zu jung aus. In Buran-Kaya werfen die vermischten Schichten zwischen dem Micoquien, dem Streletien und der jungpaläolithischen Industrie Fragen auf.

70
Tab. 6: Neue Datierungen für den Anfang und das Ende des Uluzziens. Resultate sind in cal. BP widergegen.
Tab. 7: Datierung der Fundschichten in der Grotte von Esquilleu
15.06.15 19:06ScienceDirect - Full Size Image
Seite 1 von 1http://www.sciencedirect.com/science?_ob=MiamiCaptionURL&_me…&_pii=S0047248414000074&md5=ebba6d25fae00d652a368e13e67f9f46
�
ecological and geographic conditions of this region make it aninteresting area to model the changes in !ora and fauna pop-ulations at this time. Almost all the Cantabrian Late Mousteriansites are dated to the latter part of the “Würm II” glacial, as is seenelsewhere in Europe (Moncel, 1997; Otte, 2007), although there area few exceptions. At Cueva del Castillo, evidence of earlier Mous-terian occupation was found in only a few horizons of this longsequence: Level 24 (>89 ka, U-Series dating from Castillo 23;Bishoff et al., 1992) and Level 22 (70.1 ka ! 9400; Rink et al., 1996;Soto Barreiro, 2003). A few other early dates have been recorded: atPendo XVII (83.079 ka ! 8291, dated by TL), but from a mixedsequence (Montes Barquín and Sanguino González, 2001); atLezetxiki V (dated between 186 and 57 ka; Arrizabalaga et al.,2005). However, other dated Mousterian assemblages from Can-tabria offer chronologies occasionally in the 60e50 ka range (e.g. ElEsquilleu)but most usually in the 50e40 ka (El Esquilleu, La Viña,Arrillor, Castillo, Axlor, Covalejos, El Mirón) and, increasingly, in the40e30 ka interval (e.g. El Esquilleu, Morín, Castillo, El Sidrón,Covalejos, Arrillor; Baena et al., 2005; Maíllo et al., 2001; CabreraValdés et al., 2005; Fortea, 1998; Fortea et al., 2006; Sanguino andMontes, 2005; Hoyos et al. 1999; González Urquijo et al. 2005;Ríos Garaizar, 2006).
In general, the Cantabrian dates are all dated comfortably within“Würm II”, predating the H4 event (dated to 40e38.5 cal ka;D’Errico and Sánchez Goñi, 2003) and persisting at some sites intothe classic warm episode of Hengelo-Les Cottés (c. 36 ka). At thispoint elsewhere in Europe, and particularly across the Pyrenees, theAurignacian was emerging (Maroto et al., 2005).
In Cantabria, there are also classic sequences which presentmore conventional chronologies for the transition to the UpperPalaeolithic. For example: Mirón 130 at 41.2 ka (Straus andGonzález Morales 2001); the "nal levels of Covalejos (D and J)which mark the end of a Mousterian sequence with classic chro-nologies at 40.6 ka and 38.3 ka (Sanguino and Montes, 2005);Morín (Aurignacian) level 8 at 36.6 ka (Maíllo et al., 2001).
The presence of these sites reinforces the uniqueness of thefollowing two sites which are located in environmentally similarcontexts, but which have Mousterian assemblages dated beyond35 ka. One is Arrillor Cave (Zigoitía, Álava), with level SMK-I datedto 43.1 ka ! 1700, has four Mousterian levels within the early partof “Würm III” (Lmc, dated to 37.1 ka ! 1000, Hoyos et al. 1999). Thesecond is Esquilleu Cave (Castro-Cillórigo, Cantabria) which hasa sequence that has classical Mousterian dates in the lower levelsand continues into “Würm III”, with an unequivocal recentsequence between levels VI and IV (Level VI dates to34.38 ka ! 670; Level IV dates to 22.84 ka ! 280/250). The datingsobtained for Esquilleu, although they could provide some shades inthe overall coherence of the sequence, maintain, even strengthen,the uniqueness of the site (vid infra.) in terms of continuation.
However, there are two additional factors which contribute tothe complexity of the situation in Cantabria. The attribution ofmany Cantabrian sites to the Châtelperronian is frequently de"nedon a purely statistical basis. For example: Morín 10 at36,950 ! 6580 and 28,515 ! 840 (González Echegaray andFreeman, 1978; Butzer, 1981; Laville and Hoyos, 1994); Ekain at30 ka (Straus and Heller, 1988); Labeko Koba at 34.1 ka(Arrizabalaga, 2000); La Güelga at 32.4 ka (Menéndez et al., 2005);Gatzarria (Sáenz De Buruaga, 1991) and Santimamiñe (RuízIdarraga, 1990). Above all, Castillo (Cabrera and Bischoff, 1989;Cabrera et al., 1996, 2005), with its 40 ka Aurignacian, althoughquestioned (see Zilhão and D’èrrico, 2000; Maroto et al., 2005;Zilhão, 2006), highlights the presence of a particularly complexmosaic of situations in the Cantabria region. On closer inspection,the later Middle Palaeolithic and Early Upper Palaeolithic sites ofCantabria do not "t comfortably with the traditional patterns of
interpretation for the region but present a much more complexpattern of behavior and technological repertoire.
4.2. El Esquilleu
The cave site of Esquilleu has one of the most complete chro-nological sequences in the Cantabrian Mousterian (Fig. 2) with animportant battery of dates (e.g. Baena et al., 2004, 2005) which arealmost continuous from the beginning of MIS3, including MIS3cand MIS3b, and the environmental events H5 and H4, until the H3event in the middle part of MIS3a.
The dates obtained so far are as follows:
So far, only the date obtained for Level VI (with discrepancies indates of level XIX) was inconsistent: 40,110 ! 500/420 BP; GrA33,816; AMS on charcoal. The date for Level III (12,050! 130 BP; AA29664; AMS bone) is also considered anomalous as it does not "twith the observed archaeological sequence (Jordá Pardo et al.2009).Nevertheless it is possible that the sequence may stretchinto markedly colder stages at the beginning of Level VI whichcould allow the occupation to include the H3 event at c.30 ka.
All of this underlines the unique nature of the Cantabrianassemblages. It is possible that the earliest Aurignacian in south-east Europe (Maroto et al., 2005) is coincident with one of themost signi"cant Mousterian occupations at El Esquilleu. This siteis also exceptional, since the most recent dates of other Cantabrian(e.g. Arrillor) or northern Spanish sites (e.g. Ermitons, Fuentes deSan Cristóbal, Roca dels Bous), although well recorded, do notexceed 30 ka (Vaquero et al., 2006; Maroto et al., 2005). Twopoints indicate the overall uniqueness of the sequence: from LevelVI downwards (in a range nearly typical in a peninsular context)and the uppermost levels, for which radiometric reviews areproviding extremely surprising but very consistent dating.Certainly, some differences were detected in the dates subse-quently obtained for Level VI (between 44.1 and 30.1 ka 14C BP)and Level XIX (30 ka 14C BP) with respect to those previouslyobtained, but thus, the new dates offered from Level III(19,300 ! 100 and 19,310 ! 80 14C ka BP) and Level IIIB
Level Cuture Sample Reference Lab. Date
III Mousterian charcoal GrA-33829 3640 ! 90 BPIII Mousterian Bone AA-29664 12,050 ! 130 BPIII Mousterian bone OxA-19967 19,300 ! 100 BPIII Mousterian bone OxA-19968 19,310 ! 80 BPIII B Mousterian bone OxA-19246 20,810 ! 110 BPIV Mousterian charcoal GrA-35064 22,840 " 280e250 BPIV Mousterian charcoal GrA-35064 23,560 ! 120 BPV Mousterian charcoal GrA-35065 30,250 " 500e430 BPVIF Mousterian charcoal AA37883 34,380 ! 670 BPVI Mousterian charcoal GrA-33816 40,110 " 500e420 BPVI-1 Mousterian bone OxA-19965 43,700 ! 1400 BPVI-2 Mousterian bone OxA-19966 44,100 ! 1300 BPXIF Mousterian charcoal AA37882 36,500 ! 830 BPXIII Mousterian charcoal Beta149320 39,000 ! 300 BPXVII-1 Mousterian charcoal OxA-X-2297-31 49,400 ! 1300 BPXVII-2 Mousterian charcoal OxA-20320 52,600 ! 1200 BPXVII Mousterian charcoal OxA-20318 53,400 ! 1300 BPXVII Mousterian charcoal OxA-19993 >54,000 BPXVII Mousterian charcoal OxA-20319 >58,500 BPXVIII Mousterian charcoal OxA-19993 49,700 ! 1600 BPXIX-1 Mousterian charcoal OxA-19085 39,280 ! 340 BPXIX-2 Mousterian charcoal OxA-19086 >54,600 BPXIX-3 Mousterian charcoal OxA-V-2284-29 39,600 ! 400 BPXIX-4 Mousterian charcoal OxA-V-2284-30 39,650 ! 450 BPXXI-I Mousterian charcoal OxA-20321 >59,600 BPXXId Mousterian burnt clay Mad3299 (TL) 51,034 ! 5114 BPXXIb Mousterian burnt clay Mad3300 (TL) 53,491 ! 5114 BP
Baena et al. / Quaternary International 247 (2012) 199e211202

71
Tab. 8: Darstellung der Stratigraphie von Saint-Césaire mit TL Datierung der verschiedenen Level

72
Tab. 9: Stratigraphische Zuordnung der kulturellen Abfolge
Tab. 10: Eine Übersicht verschiedener Datierungen für das Moustérien und das Uluzzien in der Grotta di Fumane
Tab. 11: Anzahl der identifizierten Taxa bei den Huftieren. Mammut nicht mit in der Berechnung inbegriffen.
Tab. 12: Anzahl der identifizierten Taxa bei den Karnivoren
3. Lithic assemblages and the cultural sequence
3.1. Lithic sources
The macroscopic features (texture, structure, colour and mor-phoscopy of cortical surfaces) of the !int exploited in Mousterianand Uluzzian industries vary according to the carbonatic forma-tions, which, in the western Monti Lessini, are dated from theUpper Jurassic to the middle Eocene. The most widespread areMaiolica (Biancone, Bi) and its varieties, and Scaglia Rossa (SR)formations, as are Eocene calcarenites (Eoc) and Scaglia Variegata(SV). Conversely, !int from other formations and oolitic limestones(Ool) is rare. The associations re!ect the lithological variability ofValpolicella, an area rich in easily provisionable lithic sourceswithin a range of 5e10 km from the site, where the !int is alsoavailable loose in !uvial and stream deposits and in deposits on theslopes and in the soils. Most of the raw material introduced ontothe site outcrops in the immediate surroundings of the cave (Bi andSR !ints). Also to be noted is the use of older patinated artefacts,recovered elsewhere and exploited as cores.
The oolitic !ints originate from marly limestones, marls andcalcarenites in the Jurassic Tenno formation (S. Vigilio Group,Toarcian-Aalenian), which crops out 5 km away in the upper Val-pantena and Adige valleys. Several varieties are to be found in theMaiolica micritic limestone formation (Titonian-Aptian), whichthickens out from the western (25e30) to the eastern fringe of theMonti Lessini (70e75 in Valpantena). Yellowish-dark !int (Bi-y)outcrops extensively in the western series and in the Fumanevalley, and is highly valued because of its excellent quality. Thesame applies to the grey variety (Bi), which outcrops a few kmsnorth-west of the site and is abundant in stream gravels. Thecolours often vary from light to dark grey, with the possibility ofshading within the same nodule. There are various types of !intdistributed randomly in Scaglia Variegata marly limestones(Aptian-Cenomanian). The grey-green (SV-gb) type has excellent!aking properties but nodules are rare. The grey-blackish (SV-gb)type is concentrated at the top of the formation in dark grey thinlayers, its use hampered by its poor quality (intense "ssuration) andsmall plate size. The dark-reddish !ints from the Scaglia Rossamarly limestones (Turonian-Maastrichtian) are excellent qualityand, although rarely found in the immediate site surroundings, arepresent abundantly in the form of nodules along the westernwatershed of the Fumane basin. The Paleocene and lower Eocenemarls and marly micritic limestones (Peoc) of the southern slopesof western Lessini contain scarce amounts of "nely textured !int,coloured grey, dark green or olive green. The succession ends withthe middle Eocene calcarenites (Eoc). Again, the !int nodules are
scarse. They are grey or brownwith Nummulites and Discocyclines,with the coarse texture of the parent rock.
3.2. Previous studies on the Mousterian lithic industries of Fumane
In the reporting of the "rst excavations carried out by theNatural History Museum of Verona, the lithic industries of theMousterian sequence were merely mentioned and divided up intolayers and hearths, and this remained the case until the "rsttechno-typological analyses were carried out by A. Sartorelli afterthe re-examination of the stratigraphy by Cremaschi in 1985(Cremaschi et al., 1986). Sartorelli used the Bordes (1961) protocoland examined the techno-typological features of 682 artefacts in"ve groups, created either according to stratigraphic provenance orin order to make up an adequate number for statistical analysis.Starting from the top, the “strati alti” (layers 12e9) and “focolareIV” group has been attributed to the Typical Mousterian of Levalloistechnology and Levalloisian facies, whereas the underlying layer 8,with a high incidence of denticulates, Levallois technology andnon-Levalloisian facies, has been labelled as Eastern FerrassieMousterian, although some doubts have emerged over the homo-geneity of the industry due to the Quina features observed in the3rd arbitrary cut and the recovery of a fragmentary acheuleanbiface. The following group (“focolare V”) is characterised by theLevallois technique, with a large number of scrapers, and has beenascribed to the Ferrassie Mousterian. Finally, various indexes andfeatures have located the industry of the lowermost layers (layer 7and “focolare IV”) in the Charentian of the Levalloisian technique.The very few artefacts recovered at the bottom of the sequence areLevallois !akes and scrapers, similar to those found in the overlyingindustries.
With the resumption of the research, the "rst preliminaryobservations on the Mousterian cultural sequence have, ata general level, con"rmed the key features of the Levallois assem-blages, i.e. the variability due to the presence of bifaces and Quinaretouched tools (Peresani and Sartorelli, 1998; Cremaschi et al.,2005). However, they have also revealed new details acrossmacro-unit A. The technological replacement observed from A10 toA9 led to the setting up of an integrated analysis project, focusingon the techno-functional signi"cance of the discoid production inA8-A9 (Peresani, 1998; Lemorini et al., 2003) and its abruptreplacement as from the Levallois in the overlying A5-A6 complex(Peresani et al., in press). Moreover, recent con"rmation of Uluzzianlevels embedded between the Mousterian and the Proto-aurignacian has led to new investigations into the lithic industryand the dynamics of the site occupation (Peresani, 2008).
Table 1A summary of numerical ages from the Mousterian and Uluzzian levels of Grotta di Fumane. Details on the TL and the combined ESR/U-series data sets are given in Peresaniet al. (2008). The Radiocarbon dates are on single charcoal fragments and are here reported at 1s con"dence. From the large 14C dataset available for this sequence, only ABOX-Sc 14C and one single ABA 14C measurements have been here reported (see Higham et al., 2009 for discussion).
Context Technocomplex Nature Lab. Ref. Method Date
A3SIV-425 Uluzzian Charcoal LTL1795A ABA14C 37,828 ! 430A4I Uluzzian Tooth FU0003 U/Th e ESR 44,000 ! 4000A5 Moust. Levallois Charcoal OxA-17980 ABOx-SC 14C 40,150 ! 350A5 Charcoal OxA-X-2275-45 ABOx-SC 14C 41,650 ! 650A5 " A6 Moust. Levallois Tooth FU9606 U/Th e ESR 38,000 ! 4000A5 " A6 Charcoal OxA-17566 ABOx-SC 14C 40,460 ! 360A6 Moust. Levallois Burnt !int 4 TL 50,000 ! 8000A6 Tooth FU9607 U/Th e ESR 38,000 ! 4000A9 Moust. Discoid Tooth FU0004 U/Th e ESR 46,000 ! 5000A11 a Moust. Levallois Tooth FU0005 U/Th e ESR 49,000 ! 5000BR11 Moust. Levallois Burnt !int MA TL 55,000 ! 7000BR12 Moust. Levallois Burnt !int 6 TL 57,000 ! 8000S7 Moust. Levallois Burnt !int 7 TL 79,000 ! 11,000
M. Peresani / Quaternary International 247 (2012) 125e150128
Subsequently, Morin (2004) provided a detailed analysis of theMousterian, Châtelperronian, and Aurignacian assemblages at St.Césaire, located some 180 km northeast of Grotte XVI. His excellentwork detected ‘‘no significant change in subsistence strategies”across the Middle-to-Upper Paleolithic transition (2004: 366). Thislack of difference includes little, if any, significant change in skele-tal part representation, seasonal use of the site, the use of low-ranked (i.e., small) vertebrate taxa, and even in such things as mar-row extraction and the possible use of specific skeletal parts asfuel.
Although these two sets of analyses are distinguished by thefact that they include detailed examinations of Châtelperronianassemblages, they are not alone in looking for, but failing to find,significant differences between Middle and early Upper Paleolithiccentral and western Eurasian archaeological vertebrate faunalassemblages (e.g., Villa et al., 2004; Bar-Oz and Adler, 2005; Adleret al., 2006; Grayson and Delpech, 2002, 2006 and references there-in), other than differences attributable to climate change.
The accomplishments of Bordes (2002) now make it possible toadd a third Châtelperronian–Aurignacian comparison to the list.That comparison is the major focus of the work reported here.Unfortunately, it did not seem appropriate to us to include theRoc-de-Combe Mousterian ungulate assemblage in this analysis.
We excluded this material for two reasons. First, there is verylittle information available on its stratigraphic context. Indeed, F.Bordes himself paid little attention to the Roc de Combe Mouste-rian during excavation. After the excavations were completed, heexpressed no interest in it whatsoever: he had come to the siteto investigate the relationship between the Châtelperronian andthe Aurignacian. The Mousterian materials remained in the bagsinto which they were put in the field, and were neither washednor numbered (J.-Ph. Rigaud, pers. comm.). Laville (1973, 1975),who provided the only detailed discussion of the Roc de Combestratigraphy and sediments prior to Bordes (2002), noted simplythat the Mousterian deposits had been subjected to significantfreeze–thaw episodes, had been heavily cemented, and that theirsurface had been heavily eroded. While Bordes (2002) discussedthe Mousterian materials from the site, his focus was on the poten-tially interstratified Châtelperronian and Aurignacian deposits. As aresult, any evaluation of the scientific importance of the Mouste-rian faunal assemblages from Roc de Combe must await an assess-ment of the stratigraphic contexts that provided thoseassemblages. However, J.-Ph. Rigaud (pers. comm.), who partici-pated in the excavations, observes that the Mousterian depositsin front of the cave were steeply sloped and lacked clear stratigra-phy. This, he suggests, might provide another reason why F. Bordesshowed little interest in them.
Second, F. Bordes’ approach to the Roc de Combe excavationswas similar to that he applied to other Paleolithic sites. Hescreened all sediments from Upper Paleolithic deposits, using aca. 2 mm mesh screen at Roc de Combe, although, as we discussbelow, he did not keep all the items caught by those screens (Bor-des, 2002; see Figs. 5 and 6). However, he did not screen depositsfrom Middle Paleolithic units, either at Roc de Combe or elsewhere.He made this distinction because of the importance he placed onsmall blade fragments, objects he expected to find in the Upper,but not the Middle, Paleolithic (J.-Ph. Rigaud, pers. comm.).
We return to this issue below, here noting only that because theMousterian deposits at Roc de Combe were not screened, there isno reason to think that the quantitative structure of this assem-blage would be comparable to that of the assemblages that werescreened.
In short, because the stratigraphic context of the Mousterianfaunal materials from Roc de Combe is not well understood and be-cause the Mousterian sediments at the site were not screened, wedo not examine that material here, observing only that it is rich inthe remains of large ungulates, particularly bovids. Instead, we fo-cus entirely on the assemblages from strata 5 through 8, the stratathat contained Aurignacian and Châtelperronian assemblages.
Delpech (1972, 1983) identified and analyzed the larger mam-mals from Roc de Combe strata 1–10. These analyses includedmaterial that Bordes (2002) showed to have been derived fromuntrustworthy stratigraphic contexts. For the work reported here,we reidentified all of the Roc de Combe larger mammals from stra-ta 5–8 and included all of this material in our subsequent tapho-nomic analyses (these analyses followed the protocol discussedin Grayson et al. (2001) and Grayson and Delpech (2003)). We theneliminated from further study all faunal materials from excavationrow G and all Aurignacian (strata 5, 6, and 7) materials from exca-vation row H, following Bordes (2002, p. 94). Differences betweenthe numbers reported here and in Delpech (1983) stem in partfrom this vetting procedure. However, we were also able to iden-tify a larger fraction of the ungulate collection than was done inthe earlier attempt (3506 ungulate specimens from strata 5–8,excluding material from rejected excavation units, compared with2466 specimens from all units). In addition, a very small number ofspecimens were assigned to different taxa during the reidentifica-tion process.
Tables 2–7 provide the raw data for many of our ensuing discus-sions. Tables 2 and 3 provide the number of identified ungulate andcarnivore specimens per stratum, respectively. Tables 4–7 provideNISP, MNE, and MAU values and associated information by stratumand skeletal part for the Roc de Combe reindeer, the only taxonpresent in sufficient abundance to merit a body part analysis.
Table 2Numbers of identified specimens per ungulate taxon by Roc de Combe stratum; mammoth tusk specimens excluded from percentage calculations
Taxon Stratum 5 Stratum 6 Stratum 7 Stratum 8 R
NISP % NISP % NISP % NISP %
Bos/Bison 14 0.92 31 10.44 24 1.78 102 32.48 171Capra sp. 10 0.66 4 1.35 4 0.30 7 2.23 25Capreolus capreolus 3 0.20 1 0.34 0 0.00 1 0.32 5Cervus elaphus 6 0.39 8 2.69 9 0.67 11 3.50 34Equus sp. 0 0.00 0 0.00 2 0.15 0 0.00 2Equus caballus 10 0.66 10 3.37 53 3.93 76 24.20 149Equus hydruntinus 0 0.00 0 0.00 0 0.00 1 0.32 1Rangifer tarandus 1470 96.71 236 79.46 1252 92.74 112 35.67 3070Rhinoceros 1 0.07 1 0.34 1 0.07 0 0.00 3Rupricapra sp. 6 0.39 1 0.34 2 0.15 2 0.64 11Sus sp. 0 0.00 5 1.68 3 0.22 2 0.64 10R 1520 297 1350 314 3481Mammoth 19 6 0 0 25
D.K. Grayson, F. Delpech / Journal of Anthropological Archaeology 27 (2008) 338–362 343
Collection bias
Although all of the excavated Upper Paleolithic sediments atRoc de Combe were screened, not all of the materials caught inthese screens were retrieved. As Bordes (2002, pp. 105,191) hasnoted, an unknown number of smaller lithic objects were dis-carded at the site. It is also possible that F. Bordes himself dis-carded further objects after they were collected, includingunretouched microdebitage (J.-Ph. Rigaud, pers. comm.; R.G. Klein,pers. comm.).
The situation might be somewhat worse for the faunal material.Although he required that screens be used, F. Bordes also askedthat only ‘‘identifiable” bones be kept, leaving it up to the excava-tors to define what was ‘‘identifiable” (pers. comm. from Ph. Delp-ech, J.-Ph. Rigaud, and J.-P. Texier, all of whom worked at the site).This process, of course, opened the possibility of introducing signif-icant bias into the retrieved Châtelperronian and Aurignacian fau-nal assemblages. Two of the excavators, J.-Ph. Rigaud and F.Delpech, recall that teeth, mandibles, vertebrae, phalanges, andthe articular ends of long bones were generally kept, but that shaftfragments were not.
Without excavating the Roc de Combe backdirt, there is no wayof quantifying the severity of such bias securely. Even excavatingthe backdirt might be problematic. This material was depositedwell beneath the site, where it has been subjected both to erosionand to the activities of collectors (J.-Ph. Rigaud, pers. comm.). Thereare, however, ways of determining if the impacts of the faunal tri-age conducted at the site are likely to have been severe. For in-stance, it is certainly appropriate to assume that long bone shaftfragments were among the least likely specimens to have been re-tained (e.g., Pickering et al., 2003) and teeth among the most likely.Indeed, this is precisely what the Roc de Combe excavators recallwas the case. Given this, a general bias estimate can be obtainedby comparing the ratios of long bone shaft fragments to teeth be-tween the identified ungulates from Roc de Combe and those froma comparable site excavated using more modern techniques.
We have chosen Grotte XVI (Rigaud et al., 1995; Grayson et al.,2001; Karkanas et al., 2002; Grayson and Delpech, 2003) for thiscomparison both because the data are readily available to us andbecause of the collection procedures that were in use at this site.Here, all faunal specimens greater than 1 cm in any dimensionwere collected in situ and given point provenience. In addition,all excavated sediments were passed through 2 mm mesh screensand all vertebrate faunal specimens from those screens saved (withthe exception of ‘‘microvertebrates”, which were sampled in otherways).
Table 8 provides data on tooth and long bone shaft abundancesin the Châtelperronian and Aurignacian assemblages at both GrotteXVI and Roc de Combe. At Grotte XVI, the ratio of tooth to shaftspecimens varies from 0.63 to 1.06. At Roc de Combe, this ratio var-
ies from 2.48 to 5.63. As a whole, shafts are more abundant thanteeth in the Grotte XVI units (teeth/long bone shaft ratio = 0.74),but the reverse is true at Roc de Combe (ratio = 3.53). This differ-ence is dramatically significant (v2 = 409.54, p < .001), with teethfar overrepresented in the Roc de Combe assemblages.
A similar measure can be built by comparing the abundances ofrib heads to rib shafts in the assemblages (Table 8). Although thedifferences here are not as pronounced as they are for teeth andlong bone shafts—and the values for Roc de Combe stratum 6and Grotte XVI stratum B are virtually identical—the compositevalues once again differ significantly (v2 = 46.52, p < .001), withrib heads greatly overrepresented in the Roc de Combe assem-blages as a whole.
Since we identified and analyzed the faunas from both of thesesites, the differences cannot reflect different approaches and skillsbrought to the identification process. It is possible, of course, thatRoc de Combe simply had more teeth and rib heads than long boneand rib shafts than did Grotte XVI. However, it seems far morelikely that these differences reflect differences in collectionprocedures.
Thus, even though the Upper Paleolithic deposits at Roc deCombe were screened, there are good reasons to think that thepost-screening triage that took place significantly impacted certainaspects of the ungulate assemblages retrieved. Unfortunately, thelikely under-representation of long bone shaft fragments at Rocde Combe has the potential of causing interpretive problems. Eventhough, we place heavy emphasis on the identification of long boneshaft fragments to avoid the introduction of observer bias into ourunderstanding of skeletal element patterning (e.g., Marean andFrey, 1997; Marean and Kim, 1998; Marean, 1998; Pickering etal., 2003), we cannot identify what may once have been presentbut was not collected. While we see great value in attempting toextract information relevant to current archaeological problemswith data collected using now-outmoded excavation procedures,we fully recognize the validity of the issues raised by Mareanand his colleagues.
However, the approach we have taken in examining subsis-tence-related aspects of the Middle-to-Upper Paleolithic transitiondoes not require that we determine whether skeletal element pat-terning within a given assemblage is more consistent with one thatmight have resulted from, for instance, hunting or scavenging.Rather than attempting to reconstruct the behavior that produceda given pattern or set of patterns, we ask whether a given patterndiffers significantly from those shown by other assemblages,regardless of the specific nature of that pattern. Thus, in our anal-yses of the Grotte XVI ungulate assemblages, we asked whether thepatterning of relevant skeletal variables in the Mousterian, Châ-telperronian, and Aurignacian assemblages differed from one an-other significantly (Grayson and Delpech, 2003). We showedthat, for the variables we measured, they did not. In addition, weasked whether such differences were greater than those betweenthe Grotte XVI Gravettian, Solutrean, and Magdalenian assem-blages and showed that they were not either. At Roc de Combe,we ask whether the magnitude of differences between Châtelper-ronian stratum 8 and Aurignacian stratum 7 is significant, andwhether those differences are greater than those between the laterAurignacian assemblages here. That is, rather than trying to recon-struct details of past human behaviors, we ask whether the resultsof those behaviors differed significantly through time.
Since this is our approach, the fact that the faunal assemblagesfrom Roc de Combe underwent post-screening triage may be lessharmful than it would otherwise be. Although we have no way ofknowing whether the distribution of mid-shaft fragments acrossskeletal elements at Roc de Combe reflects the nature of the assem-blages as they were encountered on excavation or, as seems farmore likely, reflects the discard decisions that were made after
Table 3Numbers of identified specimens per carnivore taxon by Roc de Combe stratum
Taxon Stratum
5 6 7a 7b 7c 7a–c R7 8 R
Meles meles 0 0 1 0 0 0 1 0 1Mustela nivalis 2 9 9 1 0 5 15 7 33Vulpes/Alopex 18 15 28 1 0 10 39 16 88Ursus sp. 0 0 3 0 0 0 3 1 4Mustela erminea 1 2 1 0 0 0 1 0 4Crocuta crocuta 0 1 0 0 0 0 0 0 1Canis lupus 0 2 1 0 1 2 4 2 8Lynx sp. 3 0 0 0 0 0 0 0 3Martes sp. 3 1 0 0 0 0 0 0 4Panthera spelaea 0 0 0 0 0 0 0 1 1R 27 30 43 2 1 17 63 27 147
344 D.K. Grayson, F. Delpech / Journal of Anthropological Archaeology 27 (2008) 338–362

73
IV Abbildungsnachweise • Abb.1: Schmitz 2003, S. 25. • Abb.2: http://upload.wikimedia.org/wikipedia/commons/e/ed/Fuhlrott.jpg • Abb.3: http://www.lwl.org/pressemitteilungen/daten/bilder/20244.jpg • Abb.4: Bosinski 2013, S. 20; Bolus/Schmitz 2006, S. 23. • Abb.5: http://upload.wikimedia.org/wikipedia/commons/7/75/Hermann_Schaafhausen.jpg • Abb.6: Karte bei Google Earth selbst erstellt • Abb.7:http://www.google.de/url?sa=t&rct=j&q=&esrc=s&source=web&cd=3&ved=0CC4QFjAC&url
=http%3A%2F%2Fwww.uni-graz.at%2F~foelsche%2FVO%2FKlima%2FKlima_09_Abrupte_Klimaaenderungen.ppt&ei=3zHSVKPnEoa7ygOl9oHIAQ&usg=AFQjCNHfJwqC4JeBPEL7N8VirV7OcyzCSA&sig2=cZPk3Kw0WtFybR425ZG74A (Dansgaard-Oeschger)
• Abb.8: Walker et al. 2009, S. 7. • Abb.9: (?) • Abb.10: Fidele et al. 2003, S. 303. • Abb.11: Bischoff et al. 2007, S. 764. • Abb.12: Grimaldi/Santaniello 2014, S. 127; Krause et al. 2007, S. 309. • Abb.13: Soressi 2004: 392. • Abb.14: Soressi 2004: 11. • Abb.15: Pelegrin/Soressi 2007: 285. Soressie/Roussel 2014: 2683. • Abb.16: Flas 2011: 607. • Abb.17: Nerudo/Nerudova 2013: 15. • Abb.18: Douka et al. 2014: 13. • Abb.19: https://upload.wikimedia.org/wikipedia/commons/thumb/9/9e/Aurignacian_culture_map-
de.svg/1280px-Aurignacian_culture_map-de.svg.png • Abb.20: Knecht 2000: 29, 30, 32, 34. • Abb.21: http://www.iphes.cat/abric-roman%C3%AD oder http://www.iphes.cat/print/node/32 • Abb.22: Rosell et al. 2012: 71. • Abb.23: Rios-Garaizar J. et. al. 2012: 535. • Abb.24: Morin 2012: 55. • Abb.25: Morin 2012: 62. • Abb.26: Semal et al. 2011: 305. • Abb.27: Hülle 1977: 58 • Abb.28: Hülle 1977: 101 • Abb.29: Valoch 2009: 388. • Abb.30: Romagnoli 2014: 3. • Abb.31: Peresani 2012: 127. • Abb.32: Koumouzelis et al. 2001: 519. • Abb.33: Hublin 2015: 206.
V Tabellennachweise • Tab.1: Hemming 2003: 29. • Tab.2: Soressie 2002: 9. • Tab.3: Nerudo/Nerudova 2013: 12. • Tab.4: Stodiek/Paulsen, 1996: 16. • Tab.5: Djindjian 2010: 21-25. • Tab.6: Douka et al. 2014: 8. • Tab.7: Baena et al. 2012: 202. • Tab.8:Morin 2012: 59. • Tab.9: Morin 2012: 58. • Tab.10: Peresani et al. 2008: 2991. • Tab.11: Grayson/Delpech 2008: 343. • Tab.12: Grayson/Delpech 2008: 344.

74
VI Tafelnachweise • Taf. 1: Soressi 2004: 15. • Taf. 2: Soressi 2004: 12. • Taf. 3: Pelegrin/Soressi 2007: 284, 289, 290, 291. • Taf. 4: Flas 2008: 189, 193. • Taf. 5: Flas 2002 34; 2008: 194, 195; 2011: 268. • Taf. 6: Kaminská et al. 2011: 34, 44. • Taf. 7: Neruda/Nerudova 2000: 270; Richert et al. 2008: 876; Hublin: 1347; Budek et al. 2013: 90,
92. • Taf. 8: Peresani 2010: 69, 70, 73. • Taf. 9: http://altsteinzeit-hessen.de/wp-content/uploads/2013/12/Modell-Dufor-Lam.JPEG.jpg;
Pradel 1978: 234

75
VII Eidesstaatliche Erklärung
„Ich versichere hiermit, dass die Masterarbeit mit dem Titel „Späte Neandertaler und
frühe anatomisch moderne Menschen in Europa. Zur Frage der Begegnung und des
kulturellen Vergleichs.“ von mir selbst und ohne jede unerlaubte Hilfe angefertigt
wurde, dass sie noch an keiner anderen Hochschule zur Prüfung vorgelegen hat und
dass sie weder ganz noch in Auszügen veröffentlicht worden ist. Die Stellen der
Arbeit – einschließlich Tabellen, Karten, Abbildungen usw. –, die anderen Werken
dem Wortlaut oder dem Sinn nach entnommen sind, habe ich in jedem einzelnen Fall
kenntlich gemacht.“
Jennifer Ligbado
(Datum)………………………………(Unterschrift)………………………………
Copyright © 2022 FDOKUMEN




















