
Solution-Gated Graphene Transistors for Chemical and Biological Sensors
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 354 (2007) 668–675
Sp-tetraKCNG: A novel cyclic nucleotide gated K+ channel
Blanca Estela Galindo a,b,c,*, Jose Luis de la Vega-Beltran a, Pedro Labarca d,Victor D. Vacquier c, Alberto Darszon a
a Depto. de Genetica del Desarrollo y Fisiologıa Molecular del Instituto de Biotecnologıa, Universidad Nacional Autonoma de Mexico,
Apdo Postal 510-3, Cuernavaca, Morelos 62271, Mexicob CINVESTAV, Unidad Monterrey, Av. Cerro de las Mitras 2565, Col. Obispado, C.P. 64060, Monterrey, Nuevo Leon, Mexico
c Scripps Institution of Oceanography, University of California San Diego, La Jolla, CA 92093-0202, USAd Depto. de Biofısica y Fisiologıa Molecular, Centro de Estudios Cientıficos, Arturo Prat 514, Valdivia, Chile
Received 24 December 2006Available online 17 January 2007
Abstract
The sequence of a novel cGMP-regulated, tetrameric, K+ selective channel (Sp-tetraKCNG) was discovered in the sea urchin Strong-
ylocentrotus purpuratus. The Sp-tetraKCNG is a single polypeptide made of four KCNG domains similar to voltage-dependent Na+ andCa2+ channels. Each KCNG domain has six transmembrane segments (S1–S6), the ion pore having the K+ selectivity signature GYGDand a cyclic nucleotide-binding domain (CNBD). This novel channel is evolutionary located between K+-selective and voltage-dependentEAG channels and voltage-independent cationic CNG channels. Bilayer reconstitutions demonstrate such a cGMP-regulated K+ selec-tive channel in sea urchin spermatozoa.� 2007 Elsevier Inc. All rights reserved.
Keywords: Sperm; Cyclic nucleotides; Flagellar motility; Ion channels; Fertilization; Plasma membrane
Small peptides from sea urchin eggs regulate spermmotility by elevating cyclic nucleotides and changing mem-brane ion permeability. These responses are essential forchemotaxis to the egg [1,2]. Speract, a decapeptide isolatedfrom eggs of Strongylocentrotus purpuratus [3,4], transient-ly activates a guanylyl cyclase, increasing cGMP [5,6] andhyperpolarizing sperm [7]. Evidence indicates that acGMP-gated, K+-selective channel mediates hyperpolar-ization [8–13]. Opening this channel modulates severalvoltage-dependent ion transporters, such as the voltage-de-pendent Ca2+ channels (Cav), which regulate the trajectoryof sperm in response to cGMP and speract [14–16]. Thisalso occurs in Arbacia punctulata urchin sperm, the onlysea urchin species demonstrating chemotaxis in responseto the egg peptide, resact [1,17,18].
0006-291X/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2007.01.035
* Corresponding author. Fax: +52 81 82201741.E-mail address: [email protected] (B.E. Galindo).
Two families of channels are activated by cGMP orcAMP: the cyclic nucleotide-gated channels (CNG) andthe hyperpolarization-activated and cyclic nucleotide-gatedchannels (HCN) [19]. Both families belong to the super-family of voltage-dependent K+ channels, which containsix transmembrane segments (S1–S6) and possess a posi-tively charged S4 segment and an ion pore between S5and S6. None of these channels are selective for K+ ions.
Evidence for K+ selective, cGMP-gated channels insperm was acquired through two efforts: searching the S.
purpuratus genome and planar bilayer reconstitution. Wedescribe the sequence of a novel, single polypeptide, tetra-meric, cGMP-regulated, K+-selective channel, named Sp-tetraKCNG. Bilayer reconstitution experiments show thepresence of such a channel in S. purpuratus sperm flagella.
Materials and methods
Sea urchins (S. purpuratus) were from Marinus (Long Beach, CA), orPamanes, (Ensenada, Baja California, Mexico). Salts, buffers, cAMP and

B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675 669
cGMP were from Sigma (St. Louis, MO). Diphitanoylphosphatidylcholine(DPPC) was from Avanti Polar Lipids (Alabaster, AL). 3 0,4 0-Dicholo-robenzamil (DCB) was synthesized as described [20] and dissolved indimethylformamide at 20 mM. ZD7288 was from Tocris Cookson (UK)and dissolved in distilled water at 100 mM.
Head and flagellar S. purpuratus sperm membranes for planar bilayerswere isolated as described [21] or by the pH 9.1 method for tandem massspectroscopy (MS/MS) [22].
Identification of sea urchin sperm flagellar proteins by MS/MS [23,24]yielded a sequence WASATGASVGYGDIHANSVSE [25] (see Fig. 1, P1)that was found in a translated 77 residue open reading frame (ORF) of thesea urchin genome project from the Human Genome Sequencing Center atBaylor College of Medicine (http://www.hgsc.bcm.tmc.edu/projects/seaurchin/). This ORF was then used in a BLASTp search against the non-redundant protein database at NCBI (http://www.ncbi.nlm.nih.gov/BLAST/). A KAT-like channel from carrot (CAG27094, Daucus carota)was the highest scoring match. Specific primers KIR-IF forward (5 0-AGTGTTCCCGAGGACCTTCAG-3 0) and reverse KIR-1R (5 0-ACCCGTCTTCAGAGACCAC-3 0) were designed from the sea urchin ORF from thegenome. The cDNA sequence of this putative K+ channel was thenobtained by PCR using a Lambda Zap II (Stratagene, La Jolla, CA) testiscDNA library as template. Testis total RNA was isolated by Trizol(Invitrogen, Carlsbad, CA) extraction to synthesize cDNA. Using thesecond assembly of the sea urchin genome, a scaffold was found corre-sponding to the partial cDNA sequence. This scaffold was expanded 5 0
using genome sequence. Thereafter, more primers were designed for the
Fig. 1. Amino acid sequence of the Sp-tetraKCNG channel. The full-length prosix putative transmembrane segments, which are boxed and labeled S1–S6. Thenucleotide-binding domains (CNBD) are double underlined. Peptide sequencesrich region and an Asn-rich domain are shown by a single, thin underline in theby a downward arrow.
5 0-region. 3 0-RACE was performed to obtain the 3 0-end, and the 5 0-end(684 nucleotides) was predicted by FGENESH-M, a program at theSoftberry website (http://www.softberry.com/berry.phtml). A 6717 bpfull-length cDNA sequence, encoding 2238 amino acids was obtained. Ofthis, 6033 bp were sequenced from cDNA and the remaining 684 bp of the5 0-end were predicted from the genome.
Transmembrane segments (TM) and domains were predicted with theSMART program (http://smart.embl-heidelberg.de). The BioEdit pro-gram was used for alignments (CLUSTALW) and for hydropathy plots.Alignments were done using a BLOSUM62 matrix. In addition to thechannel sequences used in the phylogenetic analysis, representativesequences from the human Kv channel pore were used for alignment of thepore region. GenBank numbers for the human Kv channels were:NP_000208 for Kv1.1 NP_004966 for Kv2.1, NP_004967 for Kv3.1 andNP_004970 for Kv4.1. The GenBank accession number for the Sp-tetra-KCNG is EF151800.
Ion channel ligand selectivity was explored by examining the primarystructure of its CNBDs. An amino acid alignment was done with differentCNBDs of representative molecules: mouse cAMP-dependent proteinkinase (PKA) I (AAH03461), sea urchin PKAII (Q26619), human cGMP-dependent protein kinase (PKG) (NP_006249), human PKGII(NP_006250) and: cyclic nucleotide gated (CNG), ether-a-go-go (EAG)-K+-like, hyperpolarization and cyclic nucleotide-gated (HCN) and plantK+ (KAT).
A neighbor-joining tree was made with complete sequences of channelsthat have an alpha subunit composed of 6 TM domains and a CNBD.
tein is 2238 amino acids. Each of the four KCNG domains (I–IV) containspore region (P) and selectivity filter (F) are underlined and the four cyclicobtained by MS/MS are overlined and labeled P1, PA, PB and PC. A Ser-third KCNG domain. The putative start of each KCNG domain is marked

670 B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675
Protein channels from various organisms were used. The fruit fly Shakerchannel (Kv1.1), was used as an outgroup. The alignment and tree wereconstructed using the program MEGA3.1 [26] with 4000 replications andthe Poisson correction. GenBank accession numbers were: P29973,CNG1; NP_005131, CNG2 and Q16281 for CNG3 for human. Othersequences were: NP_001038211, zebrafish CNG5; Q9M671, wheat AKT1;NP_999729, SpHCN1 and AAZ03555 for SpHCN2 from sea urchin;AAQ16311, lobster; O60741, HCN1; NP_001185 HCN2 and O95259,Kv10.1 from human; XP_001190426 predicted CNG2 and XP_787483predicted EAG from sea urchin; AAY84142, zebrafish Kv11.3; Q96L42,human Kv12.1; Q6ZXS4, carrot KAT-like; NP_199436, KAT1 A. thali-
ana; AAL24466, KAT-like wine grape; CAA56175, KST1 potato andBAD81033, tobacco NKT2; sequences NP_476713, ERG and CAA29917,Shaker were from fruit fly.
Planar bilayers were formed with DPPC and flagellar membranesisolated and preloaded with 0.5 M sucrose were fused after addition tothe cis compartment (1–25 lg protein), as described [21]. Single channelcurrents were acquired and analyzed as in [24]. Figure legends indicatespecific bilayer conditions. Relative ion permeability (PK+/PNa+) wascalculated with the Goldman–Hodgkin–Katz equation [21]. Average I–V curves from single channel transitions of four experiments were usedto estimate Vrev.
Results and discussion
Lee and Garbers [8] showed that speract rapidlyincreased cGMP in sperm [5], and induced a K+-dependent
Fig. 2. Amino acid alignment of the four CNG domains of the Sp-tetraKCNcontained on each CNG domain (I–IV), as well as the pore region (P) and the swith downward arrowheads. Black-boxed residues indicate identities and grey
hyperpolarization in isolated flagella, which stimulated anincrease in intracellular pH. Babcock et al. [9], demonstrat-ed that speract induced a fast K+-selective hyperpolariza-tion in swollen sperm that preceded the [Ca2+]i increasealso triggered by speract, which also induced K+-selectivesingle channel activity [10]. Galindo et al. [10], then showedthat a K+-selective channel was activated directly bycGMP in sperm flagellar plasma membrane vesicles. Asexpected, speract caused a K+-dependent transient hyper-polarization in sperm [11].
Recently, fast membrane potential (Em) recordings in A.
punctulata sperm revealed that the first ionic permeabilitychange triggered by egg peptide is a cGMP-gated, K+
channel that is transiently activated. As was suggested ear-lier [16], the resulting hyperpolarization is necessary toremove inactivation from Cav channels, possibly of theT-type [18]. This paper also reported the sequence of amonomeric K+-selective CNG channel and indicated thatit could be a tetramer. Here we report the sequence of anovel cyclic nucleotide-gated channel that is K+ selective(Sp-tetraKCNG) and is constructed as four repeat modulessimilar to a Nav or Cav channels.
Peptide sequences identified using MS/MS from spermflagellar membranes and the sequence of the CNBD were
G channel. The alignment shows the six transmembrane domains (S1–S6)electivity filter (F). The charged amino acids in the putative S4 are marked-boxed amino acids are similarities according to the BLOSUM62 matrix.

B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675 671
used to explore the sea urchin genome database and obtainthe full-length DNA sequence of Sp-tetraKCNG. Threeadditional peptide sequences of this novel channel werereported as a protein associated with the soluble adenylylcyclase (sAC) from sea urchin sperm [27]. These peptideswere from a protein ‘‘similar to a cyclic nucleotide-gatedion channel’’ (LCNG1, GenBank Accession No.XP_780236). This predicted channel represented a partialsequence of the Sp-tetraKCNG channel and the peptides.Peptide A (Fig. 1, PA) (G413EIEVLSADDDDTPIAR429)is localized in the CNBD from the KCNG I domain. Pep-tide B (Fig. 1, PB; H857NLTPDLQQR866) is found downstream of S6 in the KCNGII domain. Peptide C (Fig. 1,PC S1091EDFQEPFQEISLLSR1106), reported by Nomuraand Vacquier as belonging to a ‘‘predicted olfactoryCNG channel’’, is located upstream of the IIIS1 of KCNGIII. The predicted molecular mass of the 2238 amino acidsequence of Sp-tetraKCNG is 253 kDa.
The Sp-tetraKCNG sequence is not an artefact causedby ligation of cDNAs during library construction, sincesequences between the four KCNG domains that connect
CKarteT
CKarteT
CKarteT
CKarteT
2GNCnamuH
CKarteTIII
aa 8322 = htgneL
A
B C
PMNc
P
NPMNc
P
noiger eroP
F
GNCKarteT I Y V I S L Y W S AV T A T S T G Y G D I VAS KEN E
GNCKarteT II Y I Q S L Y W A AA T M T S T G Y G D I HAS GITS
GNCKarteT III Y V T S M Y W A AS T G A S V G Y G D I NAH SVS E
GNCKarteT VI Y C TD I Y W A AV T L T S T G Y G D I YAH PVS E
notcnalpooZ vK I QP S M Y W A VI T M T T V G Y G D V V P H GITT
ylF tiurF rekahS-1.1vK DPI FA W W A VV T M T T V G Y G D M T P V GWVG
namuH bahS-1.2vK API FS W W A IT T M T T V G Y G D I Y P K GLLT
namuH wahS-1.3vK FGIPI W W A VV T M T T L G Y G D M Y P Q GSWT
namuH lahS-1.4vK API FA W Y T VI T M T T L G Y G D M V P S GAIT
torraC TAK Y V T S I Y W S TI T L T T V G Y G D L AH Q GTN E
taehW 1TKA Y V T S V Y W S TI T L T T V G Y G D Y EAH RIN E
occaboT 2TKN Y I A A L Y W S TI T M T T V G Y G D L VAH LTN E
anailaht.A 1TAK Y V T A L Y W S TI T L T T T G Y G D F EAH RPN E
namuH 1GNC Y V Y S L Y W S LT T L T T I G - E T PP P V SDR E
namuH 2GNC Y I CY L Y W S LT T L T T I G - PPTE P V EDK E
hsifarbeZ 5GNC Y V CY L Y W S LT T L T T I G - APME P V EDQ E
ylf tiurF GRE Y I T A L Y F TFT S L T S V G F G N AV P N ADT E
namuH 1GAE 1.01vK Y I S S L Y F T TM S L T S V G F G N AI P S IDT E
hsifarbeZ 3.11vK Y V T A L Y F SFT S L T S V G F G N SV P N SNT E
namuH 1.21vK Y I A A L Y F T SL S L T S V G F G N ASV N ADT E
1NCHpS Y T W A L F K A HSL LM C I G Y G K PF P Q TIS D
2NCHpS Y S W FS KF A HCI IM S I G F G R PF P M TVN E
retsboL NCH Y S W FA KF A HSM CLM I G Y G S PF P Q TLN D
namuH 1NCH Y S Y A L F K A HSM CLM I G Y G A AQ P V SMS D
namuH 2NCH Y S F A L F K A HSM CLM I G Y G R AQ P E TMS D
Fig. 3. (A) Schematic representation of the Sp-tetraKCNG channel. The four KS6) and containing a pore loop (P) and a cyclic nucleotide-binding domain (CNof representative CNG, HCN, EAG-like, KAT and Kv channels. Note that thBlack-boxed residues indicate identity in at least 12 sequences and grey-boxeHydropathy plots of the four KCNG domains and the orthologous domain ofloop as P.
each domain could be amplified from testis cDNA. There-fore, the Sp-tetraKCN is constructed of four KCNGdomains with 24 TM helices (Figs. 2 and 3A). Hydropathyplots of KCNG domains are compared to human CNG2 inFig. 3C. Each domain contains an ion pore having thesequence GYGD, the characteristic selectivity filter forK+ channels (Figs. 2 and 3B). The sequence of eachdomain in the Sp-tetraKCNG channel is similar to thoseof CNG, KAT and EAG channels, all possessing a CNBD.Voltage-gated K+ channels have only 6 TM helices andrequire tetramerization to constitute a functional channel.However, as in Cav or Nav channels, Sp-tetraKCNG hasthe four domains in one protein chain.
The primary structures of the four CNBDs, specificallythe residues needed for cyclic nucleotide binding, werecompared with those of PKA, PKG, and CNG, EAG-like,HCN and KAT family channels (Fig. 4). We searchedwithin these domains for a Thr residue that forms a hydro-gen bond with the guanine 2-amino group of cGMP, butnot with cAMP [28,29]. All four KCNG domains havethe Thr residue at the same position as CNG channels
1S 2S 3S4S
5S 6S
P
1S2S 3S
4S5S 6S
P
1S 2S 3S4S
5S 6S
P
1S 2S 3S4S
5S 6SP
1S 2S3S
4S5S 6S
P
GN I
GN II
GN III
GN VI
DBNC
GNVIIII
losotyc CPMNc
P
PMNc
P
CNG domains (I–IV) each composed of six transmembrane segments (S1–BD) are shown. (B) Alignment of the Sp-tetraKCNG pore filters with thosee GYGD signature is shared by Sp-tetraKCNG, Kv and KAT channels.d amino acids are similarities according to the BLOSUM62 matrix. (C)human CNG2. Transmembrane segments are labeled S1–S6 and the pore

Fig. 4. Amino acid alignment of the four CNBD from the Sp-tetraKCNG channel and CNBDs from other representative channels. Key residues for cyclicnucleotide binding are denoted by an asterisk. Green-boxed residues show those amino acids necessary for binding of both cAMP and cGMP. Blue-boxedresidues denote amino acids that could participate only in the binding of cAMP. Red-boxed residues participate exclusively in cGMP binding. Black-boxed residues indicate identity in at least 12 sequences and grey-boxed amino acids are similarities according to the BLOSUM62 matrix.
672 B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675
and PKGs (Fig. 4, red asterisk, Thr at positions 448, 992,1541 and 2178). In addition, Fig. 4 shows these domainshave an Arg (green asterisk) known to interact with theribose group of cGMP or cAMP. Blue-boxed amino acidsare involved in cAMP binding [29–31]. This indicates thatthe four tetraKCNG CNBDs have higher similarity tothe cGMP-binding region of CNG channels, and of PKGs,than to the cAMP-binding domain of PKAs (Fig. 4).
Comparing the four KCNG domains to each other(Fig. 2), shows percent identities of amino acids are: I–II29%, I–III 33%, I–IV 28.5%, II–III 29%, II–IV 30% andIII–IV 29.5%. This high amount of divergence suggestsan ancient duplication of these four KCNG domains.
Database searches using the full-length Sp-tetraKCNGsequence, or any of its four repetitive domains, yieldedalignments with all known CNG channels, such as theKAT-like, HCN and EAG-like channels. Therefore, tosearch for orthologues of Sp-tetraKCNG, the interdomainregions were used as queries for BLASTP or TBLASTXsearches of GenBank. With the exception of the partial pre-dicted sequence for this channel from S. purpuratus
(XP_780236) and one partially sequenced orthologue fromA. punctulata sperm (ABH10136 [18], no significant match-es were found.
A neighbor-joining tree (Fig. 5) was made using thefour KCNG domains separately and including 22 otherchannels that contain a CNBD from the EAG, CNG,HCN and KAT channel families. Kv1.1 (Shaker-like)channel from Drosophila was used as an outgroup.KCNG domains from the Sp-tetraKCNG channel consti-tuted a new branch located between the HCN channelsand the EAG channel family (Fig. 5). Because the greatestdifferences in alignments of these channels are upstreamof S1 and downstream of the CNBD, the tree was remadewithout these two end regions. The ‘‘core KCNGdomain’’, from S1 through the CNBD, yielded the exactbranching order, as does the tree made with the full-length sequences.
Three separated alignments were made between the fourdomains of a human Cav2.1, a human Cav3.2 and a humanNav1.1 and Nav1.3 (Accession Nos.: NP_000059, O95180,P35498 and NP_008853). Each alignment was used to con-struct its respective phylogenetic tree. All trees showed thesame branching topology as was shown with the tree com-posed of the four KCNG domains (Fig. 5). For the Sp-tet-raKCNG channel, domains I and III group together andare separated from domains II and IV that also grouptogether (Fig. 5).

SpHCN1 Sea urchin
HCN Lobster
HCN1-Human
HCN2 Human
SpHCN2 Sea urchin
PRED:CNG2 Sea urchinCNG1 HumanCNG3 Human
CNG2 Human
CNG5 Zebrafish
TetraKCNG IVTetraKCNG II
TetraKCNG IIITetraKCNG I
PRED:EAG Sea urchin
Kv10.1 EAG1 Human
Kv12.1 Human
Kv11.3 Zebrafish
ERG Fruit fly
NKT2 Tobacco
AKT1 Wheat
KAT Carrot
KST1 Potato
KAT-like Wine grape
KAT1 A. thalianaShaker Fruit Fly
100
99
100
97
99
100
96
100
100
100
100
83
86
100
100
78
100
85
84
74 90
0.2
HCN Family
CNG Family
TetraKCNG
ERG Family
KAT-like Family
Plants
Fig. 5. Phylogenetic analysis of channel alpha subunits composed of six transmembrane segments and a CNBD. A neighbor-joining tree made with thealignment of 24 full-length sequences from different species. The Drosophila shaker channel was used as an outgroup. The four KCNG domains from seaurchin Sp-tetraKCNG are in bold. Numbers at nodes are bootstrap percentages from 4000 replications.
B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675 673
The Sp-tetraKCNG channel, the first of its class, is evo-lutionary located between K+-selective and voltage-depen-dent EAG channels, and the voltage-independent cationicCNG channels, both families possessing a CNBD. The for-mation of a separated branch in the phylogenetic tree indi-cates that this is a new channel family whose componentsshare characteristics with K+ selective channels and CNGchannels.
We searched for the activity of such a channel by incor-porating sperm flagellar membranes into planar bilayers[21]. Planar lipid bilayer experiments have permitted docu-mentation of several ion channels, both in sea urchin andmouse sperm membranes [12,13]. This experimentalapproach provided the first recordings of cAMP-gated, cat-ion-selective channels from purified sea urchin sperm fla-gellar membranes [21]. Direct patch clamping of swollensea urchin sperm revealed a channel with similar character-istics [32,33].
Though previous functional studies had indicated theexistence of cGMP-activated K+-selective channels in seaurchin sperm that are involved in motility and chemotaxis[1,13], no direct recording of such channels had been doc-umented. To investigate cGMP-gated K+-selective chan-nels in sperm membranes, we monitored the activity ofsuch a channel by fusing purified flagellar plasma mem-branes with planar lipid bilayers [34]. We found that a highproportion of bilayer fusions (300/350) resulted in insertion
of cation-selective cAMP-activated channels that we havepreviously described [21]. Channels were activated by add-ing 200 lM cAMP to the cis bilayer chamber (Fig. 6A).These channels are refractory to 3 0,4 0-dicholorobenzamil(DCB), a blocker of cGMP-activated K+ channels(Fig. 6A) [35]. However ZD7288, which blocks HCN chan-nels [36], also blocks this cAMP-gated cation channel(Fig. 6A, fourth trace).
cAMP-gated cation channel fluctuations in the recordindicated ion channel insertion to the planar lipid bilayer.Thereafter, the presence of cGMP-gated channels was test-ed by monitoring ion channel currents that were refractoryto ZD7288, activated by cGMP, and blocked by DCB.Fig. 6B shows ion channel activity in planar lipid bilayerswith ZD7288 (control, upper record), which were activatedby adding cGMP to the cis bilayer chamber (middlerecord). These cGMP-activated channels were blocked by25 lM DCB to the cis bilayer chamber (lower record). Only30 out of 350 assays yielded insertion of cGMP-activated,DCB-blocked ion channels. Fig. 6C illustrates the subcon-ductance states exhibited by the cGMP-activated channels(49 (O1), 92 (O2) and 139 pS (O3)). Substate reversal poten-tials showed that cGMP-activated channels were perfectlycation selective. Fig. 6C compares the probability thatthe cGMP-activated channel opens to the different subcon-ductance states under control conditions and followingcGMP addition to the cis chamber, at different applied
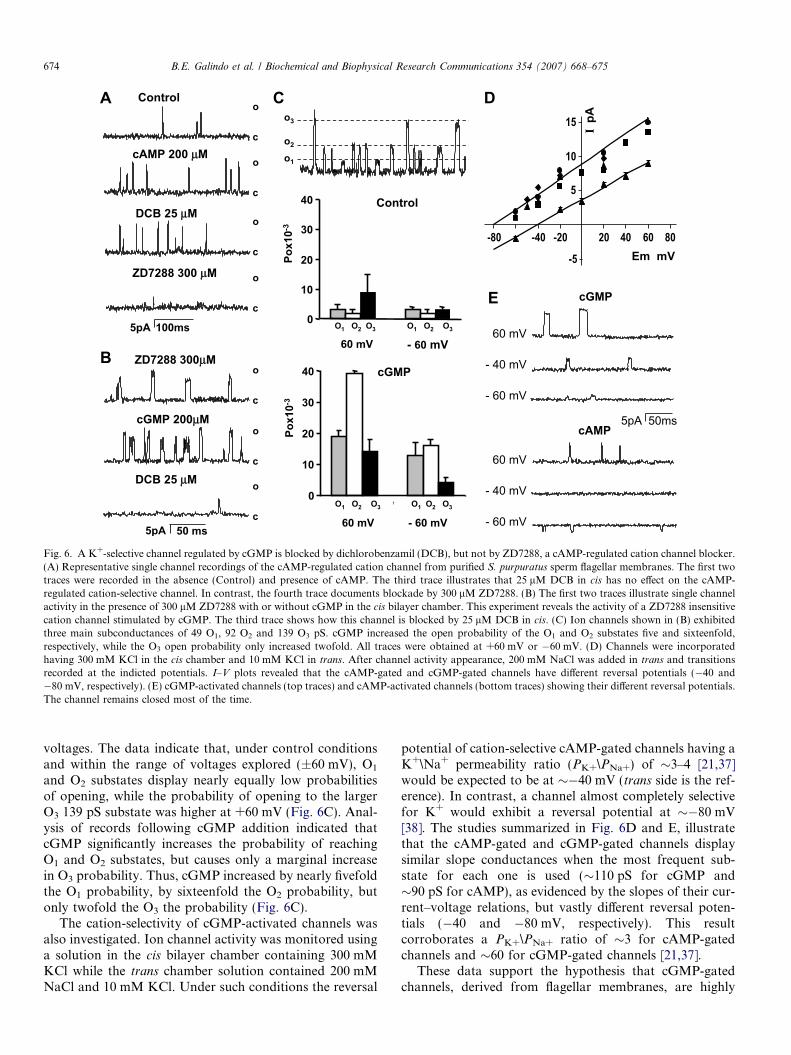
Fig. 6. A K+-selective channel regulated by cGMP is blocked by dichlorobenzamil (DCB), but not by ZD7288, a cAMP-regulated cation channel blocker.(A) Representative single channel recordings of the cAMP-regulated cation channel from purified S. purpuratus sperm flagellar membranes. The first twotraces were recorded in the absence (Control) and presence of cAMP. The third trace illustrates that 25 lM DCB in cis has no effect on the cAMP-regulated cation-selective channel. In contrast, the fourth trace documents blockade by 300 lM ZD7288. (B) The first two traces illustrate single channelactivity in the presence of 300 lM ZD7288 with or without cGMP in the cis bilayer chamber. This experiment reveals the activity of a ZD7288 insensitivecation channel stimulated by cGMP. The third trace shows how this channel is blocked by 25 lM DCB in cis. (C) Ion channels shown in (B) exhibitedthree main subconductances of 49 O1, 92 O2 and 139 O3 pS. cGMP increased the open probability of the O1 and O2 substates five and sixteenfold,respectively, while the O3 open probability only increased twofold. All traces were obtained at +60 mV or �60 mV. (D) Channels were incorporatedhaving 300 mM KCl in the cis chamber and 10 mM KCl in trans. After channel activity appearance, 200 mM NaCl was added in trans and transitionsrecorded at the indicted potentials. I–V plots revealed that the cAMP-gated and cGMP-gated channels have different reversal potentials (�40 and�80 mV, respectively). (E) cGMP-activated channels (top traces) and cAMP-activated channels (bottom traces) showing their different reversal potentials.The channel remains closed most of the time.
674 B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675
voltages. The data indicate that, under control conditionsand within the range of voltages explored (±60 mV), O1
and O2 substates display nearly equally low probabilitiesof opening, while the probability of opening to the largerO3 139 pS substate was higher at +60 mV (Fig. 6C). Anal-ysis of records following cGMP addition indicated thatcGMP significantly increases the probability of reachingO1 and O2 substates, but causes only a marginal increasein O3 probability. Thus, cGMP increased by nearly fivefoldthe O1 probability, by sixteenfold the O2 probability, butonly twofold the O3 the probability (Fig. 6C).
The cation-selectivity of cGMP-activated channels wasalso investigated. Ion channel activity was monitored usinga solution in the cis bilayer chamber containing 300 mMKCl while the trans chamber solution contained 200 mMNaCl and 10 mM KCl. Under such conditions the reversal
potential of cation-selective cAMP-gated channels having aK+\Na+ permeability ratio (PK+\PNa+) of �3–4 [21,37]would be expected to be at ��40 mV (trans side is the ref-erence). In contrast, a channel almost completely selectivefor K+ would exhibit a reversal potential at ��80 mV[38]. The studies summarized in Fig. 6D and E, illustratethat the cAMP-gated and cGMP-gated channels displaysimilar slope conductances when the most frequent sub-state for each one is used (�110 pS for cGMP and�90 pS for cAMP), as evidenced by the slopes of their cur-rent–voltage relations, but vastly different reversal poten-tials (�40 and �80 mV, respectively). This resultcorroborates a PK+\PNa+ ratio of �3 for cAMP-gatedchannels and �60 for cGMP-gated channels [21,37].
These data support the hypothesis that cGMP-gatedchannels, derived from flagellar membranes, are highly

B.E. Galindo et al. / Biochemical and Biophysical Research Communications 354 (2007) 668–675 675
K+ selective. Opening these cGMP-gated channels wouldhyperpolarize sea urchin sperm, consistent with the ideathat cGMP-gated channels participate in the speract-in-duced sperm hyperpolarization. Because channel openingcan occur in the absence of cGMP, this channel could affectthe sperm resting potential.
Acknowledgments
This work was supported by NIH Grant HD12986 toV.D.V. and by Grants DGAPA IN225406-3, CONACyT,the Wellcome Trust and FIRCA RO3 TW 006121 to A.D.
References
[1] U.B. Kaupp, E. Hildebrand, I. Weyand, Sperm chemotaxis in marineinvertebrates—molecules and mechanisms, J. Cell Physiol. 208 (2006)487–494.
[2] A. Darszon, J.J. Acevedo, B.E. Galindo, E.O. Hernandez-Gonzalez, T.Nishigaki, C.L. Trevino, C. Wood, C. Beltran, Sperm channel diversityand functional multiplicity, Reproduction 131 (2006) 977–988.
[3] J.R. Hansbrough, D.L. Garbers, Speract. Purification and character-ization of a peptide associated with eggs that activates spermatozoa,J. Biol. Chem. 256 (1981) 1447–1452.
[4] N. Suzuki, K. Nomura, H. Ohtake, S. Isaka, Purification and theprimary structure of sperm-activity peptides from the jelly coat of seaurchin eggs, Biochem. Biophys. Res. Commun. 99 (1981) 1238–1244.
[5] D.L. Garbers, Molecular basis of fertilization, Annu. Rev. Biochem.58 (1989) 719–742.
[6] P.S. Yuen, D.L. Garbers, Guanylyl cyclase-linked receptors, Annu.Rev. Neurosci. 15 (1992) 193–225.
[7] A. Darszon, T. Nishigaki, C. Wood, C.L. Trevino, R. Felix, C.Beltran, Calcium channels and Ca2+ fluctuations in sperm physiology,Int. Rev. Cytol. 243 (2005) 79–172.
[8] H.C. Lee, D.L. Garbers, Modulation of the voltage-sensitive Na+/H+
exchange in sea urchin spermatozoa through membrane potentialchanges induced by the egg peptide speract, J. Biol. Chem. 261 (1986)16026–16032.
[9] D.F. Babcock, M.M. Bosma, D.E. Battaglia, A. Darszon, Earlypersistent activation of sperm K+ channels by the egg peptide speract,Proc. Natl. Acad. Sci. USA 89 (1992) 6001–6005.
[10] B.E. Galindo, C. Beltran, E.J. Cragoe Jr., A. Darszon, Participationof a K(+) channel modulated directly by cGMP in the speract-induced signaling cascade of strongylocentrotus purpuratus seaurchin sperm, Dev. Biol. 221 (2000) 285–294.
[11] C. Beltran, O. Zapata, A. Darszon, Membrane potential regulates seaurchin sperm adenylylcyclase, Biochemistry 35 (1996) 7591–7598.
[12] A. Darszon, P. Labarca, T. Nishigaki, F. Espinosa, Ion channels insperm physiology, Physiol. Rev. 79 (1999) 481–510.
[13] A. Darszon, J.J. Acevedo, B.E. Galindo, E.O. Hernandez-Gonzalez, T.Nishigaki, C.L. Trevino, C. Wood, C. Beltran, Sperm channel diversityand functional multiplicity, Reproduction 131 (2006) 977–988.
[14] C.D. Wood, A. Darszon, M. Whitaker, Speract induces calciumoscillations in the sperm tail, J. Cell Biol. 161 (2003) 89–101.
[15] C.D. Wood, T. Nishigaki, T. Furuta, S.A. Baba, A. Darszon, Real-time analysis of the role of Ca(2+) in flagellar movement and motilityin single sea urchin sperm, J. Cell Biol. 169 (2005) 725–731.
[16] G. Granados-Gonzalez, I. Mendoza-Lujambio, E. Rodriguez, B.E.Galindo, C. Beltran, A. Darszon, Identification of voltage-dependentCa2+ channels in sea urchin sperm, FEBS Lett. 579 (2005) 6667–6672.
[17] G. E. Ward, C. J. Brokaw, D. L. Garbers, and V. D. Vacquier,Chemotaxis of Arbacia punctulata spermatozoa to resact, a peptidefrom the egg jelly layer, in J. Cell Biol., 1985, pp. 2324–2329.
[18] T. Strunker, I. Weyand, W. Bonigk, Q. Van, A. Loogen, J.E. Brown,N. Kashikar, V. Hagen, E. Krause, U.B. Kaupp, A K+selectivecGMP-gated ion channel controls chemosensation of sperm, Nat.Cell Biol. 8 (2006) 1149–1154.
[19] F. Hofmann, M. Biel, U.B. Kaupp, International Union of Pharma-cology. LI. Nomenclature and structure-function relationships ofcyclic nucleotide-regulated channels, Pharmacol. Rev. 57 (2005) 455–462.
[20] E.J. Cragoe Jr., O.W. Woltersdorf Jr., J.B. Bicking, S.F. Kwong, J.H.Jones, Pyrazine diuretics. II. N-amidino-3-amino-5-substituted 6-halopyrazinecarboxamides, J. Med. Chem. 10 (1967) 66–75.
[21] P. Labarca, C. Santi, O. Zapata, E. Morales, C. Beltr’an, A. Li’evano,A. Darszon, A cAMP regulated K+-selective channel from the seaurchin sperm plasma membrane, Dev. Biol. 174 (1996) 271–280.
[22] V.D. Vacquier, N. Hirohashi, Sea urchin spermatozoa, Methods CellBiol. 74 (2004) 523–544.
[23] M.P. Washburn, D. Wolters, J.R. Yates 3rd, Large-scale analysis ofthe yeast proteome by multidimensional protein identification tech-nology, Nat. Biotechnol. 19 (2001) 242–247.
[24] C.C. Wu, M.J. MacCoss, K.E. Howell, J.R. Yates 3rd, A method forthe comprehensive proteomic analysis of membrane proteins, Nat.Biotechnol. 21 (2003) 532–538.
[25] B.E. Galindo, A.T. Neill, V.D. Vacquier, A new hyperpolarization-activated, cyclic nucleotide-gated channel from sea urchin spermflagella, Biochem. Biophys. Res. Commun. 334 (2005) 96–101.
[26] S. Kumar, K. Tamura, M. Nei, MEGA3: Integrated software forMolecular Evolutionary Genetics Analysis and sequence alignment,Brief Bioinform 5 (2004) 150–163.
[27] M. Nomura, V.D. Vacquier, Proteins associated with soluble adenylylcyclase in sea urchin sperm flagella, Cell Motil. Cytoskeleton 63(2006) 582–590.
[28] W. Altenhofen, J. Ludwig, E. Eismann, W. Kraus, W. Bonigk, U.B.Kaupp, Control of ligand specificity in cyclic nucleotide-gatedchannels from rod photoreceptors and olfactory epithelium, Proc.Natl. Acad. Sci. USA 88 (1991) 9868–9872.
[29] W.N. Zagotta, N.B. Olivier, K.D. Black, E.C. Young, R. Olson, E.Gouaux, Structural basis for modulation and agonist specificity ofHCN pacemaker channels, Nature 425 (2003) 200–205.
[30] G.R. Tibbs, D.T. Liu, B.G. Leypold, S.A. Siegelbaum, A state-independent interaction between ligand and a conserved arginineresidue in cyclic nucleotide-gated channels reveals a functionalpolarity of the cyclic nucleotide binding site, J. Biol. Chem. 273(1998) 4497–4505.
[31] K.B. Craven, W.N. Zagotta, CNG and HCN channels: two peas, onepod, Annu. Rev. Physiol. 68 (2006) 375–401.
[32] J. Gorelik, Y. Gu, H.A. Spohr, A.I. Shevchuk, M.J. Lab, S.E.Harding, C.R. Edwards, M. Whitaker, G.W. Moss, D.C. Benton, D.Sanchez, A. Darszon, I. Vodyanoy, D. Klenerman, Y.E. Korchev,Ion channels in small cells and subcellular structures can be studiedwith a smart patch-clamp system, Biophys. J. 83 (2002) 3296–3303.
[33] D. Sanchez, P. Labarca, A. Darszon, Sea urchin sperm cation-selective channels directly modulated by cAMP, FEBS Lett. 503(2001) 111–115.
[34] A. Darszon, P. Labarca, C. Beltran, J. Garcia-Soto, A. Lievano, SeaUrchin Sperm: an ion channel reconstitution study case, Methods 6(1994) 37–50.
[35] W.N. Zagotta, S.A. Siegelbaum, Structure and function of cyclicnucleotide-gated channels, Annu. Rev. Neurosci. 19 (1996) 235–263.
[36] K.S. Shin, B.S. Rothberg, G. Yellen, Blocker state dependence andtrapping in hyperpolarization-activated cation channels: evidence foran intracellular activation gate, J. Gen. Physiol. 117 (2001) 91–101.
[37] R. Gauss, R. Seifert, U.B. Kaupp, Molecular identification of ahyperpolarization-activated channel in sea urchin sperm, Nature 393(1998) 583–587.
[38] B. Hille, Ion Channels of Excitable Membranes, third Ed., SinauerAssociates Inc., 2001.
Copyright © 2022 FDOKUMEN




















