
Solubilization and characterization of a cell wall-bound trehalase from ascospores of the fission...
8
Microbiological Research 164 (2009) 304—311 Solubilization and characterization of a cell wall-bound trehalase from ascospores of the fission yeast Schizosaccharomyces pombe J. Vicente-Soler, T. Soto, M. Madrid, A. Nu `n ˜ez, J. Cansado, M. Gacto Department of Genetics and Microbiology, Facultad de Biologı ´a, University of Murcia, 30071 Murcia, Spain Received 13 June 2006; received in revised form 3 October 2006; accepted 8 December 2006 KEYWORDS Trehalase; Fission yeast; Trehalose mobilization; Ascospores Summary The genome of the fission yeast Schizosaccharomyces pombe lacks sequence homologs to ath1 genes coding for acid trehalases in other yeasts or filamentous fungi. However, acid trehalase activity is present at the spore stage in the life cycle of the fission yeast. The enzyme responsible for this activity behaves as a surface enzyme covalently linked to the spore cell walls in both wild-type and ntp1 mutant strains devoid of neutral trehalase. Lytic treatment of particulated cell wall fractions allowed the solubilization of the enzyme into an active form. We have characterized this soluble enzyme and found that its kinetic parameters, optimum pH and temperature, thermal denaturation and salt responses are closely similar to other conventional acid trehalases. Hence, this rather unusual enzyme can be recognized as acid trehalase by its biochemical properties although it does not share genetic homology with other known acid trehalases. The potential role of such acid trehalase in the mobilization of trehalose is discussed. & 2007 Elsevier GmbH. All rights reserved. Introduction Trehalose plays a key physiological role in yeast cells as reserve and stress metabolite (van Laere, 1989). Enzymes that breakdown this storage dis- accharide are specific hydrolases known as treha- lases. Yeast trehalases are classified in two classes, neutral (regulatory) and acid (non regulatory) treha- lases, on the bases of their optimum pH for activity, ability to become activated by covalent modification and localization into the cells (Thevelein, 1984). Neutral trehalases coexist with trehalose within the cytosol as cryptic enzymes whose activity can be triggered by cAMP-dependent phosphorylation of the enzyme protein. Acid trehalases are compart- mentalized into vacuoles or at the cell surface and their activity is not regulated by post-traslational ARTICLE IN PRESS www.elsevier.de/micres 0944-5013/$ - see front matter & 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.micres.2006.12.005 Corresponding author. Fax: +34968 363963. E-mail address: [email protected] (M. Gacto).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Solubilization and characterization of a cell wall-bound trehalase from ascospores of the fission...

ARTICLE IN PRESS
Microbiological Research 164 (2009) 304—311
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/micres
Solubilization and characterization of acell wall-bound trehalase from ascospores of thefission yeast Schizosaccharomyces pombe
J. Vicente-Soler, T. Soto, M. Madrid, A. Nunez, J. Cansado, M. Gacto�
Department of Genetics and Microbiology, Facultad de Biologıa, University of Murcia, 30071 Murcia, Spain
Received 13 June 2006; received in revised form 3 October 2006; accepted 8 December 2006
KEYWORDSTrehalase;Fission yeast;Trehalosemobilization;Ascospores
ee front matter & 2007micres.2006.12.005
ing author. Fax: +34 968ess: [email protected] (M. G
SummaryThe genome of the fission yeast Schizosaccharomyces pombe lacks sequencehomologs to ath1 genes coding for acid trehalases in other yeasts or filamentousfungi. However, acid trehalase activity is present at the spore stage in the life cycleof the fission yeast. The enzyme responsible for this activity behaves as a surfaceenzyme covalently linked to the spore cell walls in both wild-type and ntp1 mutantstrains devoid of neutral trehalase. Lytic treatment of particulated cell wallfractions allowed the solubilization of the enzyme into an active form. We havecharacterized this soluble enzyme and found that its kinetic parameters, optimumpH and temperature, thermal denaturation and salt responses are closely similar toother conventional acid trehalases. Hence, this rather unusual enzyme can berecognized as acid trehalase by its biochemical properties although it does not sharegenetic homology with other known acid trehalases. The potential role of such acidtrehalase in the mobilization of trehalose is discussed.& 2007 Elsevier GmbH. All rights reserved.
Introduction
Trehalose plays a key physiological role in yeastcells as reserve and stress metabolite (van Laere,1989). Enzymes that breakdown this storage dis-accharide are specific hydrolases known as treha-lases. Yeast trehalases are classified in two classes,
Elsevier GmbH. All rights rese
363963.acto).
neutral (regulatory) and acid (non regulatory) treha-lases, on the bases of their optimum pH for activity,ability to become activated by covalent modificationand localization into the cells (Thevelein, 1984).Neutral trehalases coexist with trehalose within thecytosol as cryptic enzymes whose activity can betriggered by cAMP-dependent phosphorylation ofthe enzyme protein. Acid trehalases are compart-mentalized into vacuoles or at the cell surface andtheir activity is not regulated by post-traslational
rved.

ARTICLE IN PRESS
Solubilization and characterization of a cell wall-bound trehalase 305
mechanisms (Thevelein, 1984). Recently, it hasbeen suggested to rename neutral and acidtrehalases as cytosolic and extracellular trehalases,respectively, to describe more adequately theirlocalization and function in the yeast cells (Parrouet al., 2005). Trehalases with mixed biochemicalproperties have been described in thermophilicfungi (Lucio-Eterovic et al., 2005).
The implication of neutral trehalases in thehydrolysis of endogenous trehalose is well docu-mented (Cansado et al., 1998). Besides, it isgenerally accepted that the main function of acidtrehalases is related to the utilization of exogenoustrehalose as carbon source (d’Enfert and Fontaine,1997; Parrou et al., 2005). However, the involve-ment of acid trehalases in the catabolism ofintracellular trehalose has been suggested inseveral reports. In particular, Inoue and Shimoda(1981) proposed that the mobilization of endogen-ous trehalose in the fission yeast Schizosacchar-omyces pombe during spore germination might becarried out by a cell wall-bound acid trehalase.They also suggested that the germination stimuliwould alter plasma membrane permeability toallow the cytosolic trehalose to reach the trehalaseenzyme located outside the spore. Later workdemonstrated that in addition to the sporulation-specific acid trehalase, a cytosolic neutral treha-lase exists in the vegetative cells of the fissionyeast, leading to the assumption that such enzymewould be responsible for the mobilization in sporesof the stored trehalose (De Virgilio et al., 1991).However, subsequent studies described that treha-lose was still mobilized during germination inspores from Sch. pombe mutants defective inneutral trehalase, although at a much lower ratethan in wild type spores (Beltran et al., 2000).Moreover, inhibition of the acid trehalase in thesemutants blocked trehalose catabolism and greatlyretarded spore germination, suggesting an impor-tant role in the breakdown of this reserve carbohy-drate during the germination process (Beltranet al., 2000). The decrease in trehalose contentobserved in the absence of neutral enzyme wasattributed to the acid trehalase, which was thusconsidered to participate in the hydrolysis of theendogenous trehalose as an ancillary enzyme. Incontrast to all biochemical data supporting thepresence of an acid trehalase activity duringsporulation, the complete sequencing of theSch. pombe genome revealed the absence ofsequence homologs to known acid trehalases fromother yeasts and fungi (Parrou et al., 2005).This unexpected finding has hampered directgenetic approaches in studies on the nature andphysiological significance of the acid trehalase of
Sch. pombe. In view of these conflicting results, wereinvestigated the trehalase enzyme of the fissionyeast spores to present additional data related toits localization, solubilization and biochemicalcharacterization.
Materials and methods
Cell strains, culture and ascospore isolation
Sch. pombe strain 968 (h90 ura4-D18) and strainMMT48 (h90 ura4-D18 ntp1:: ura4+), which lacksneutral trehalase activity, were used in this study(Beltran et al., 2000). To obtain spores, cells werepre-grown until mid-exponential phase in YES liquidmedium containing 2% glucose plus 0.6% yeastextract, washed in sterile distilled water andfurther incubated for 6 days in liquid MEL sporula-tion medium, which contained 3% malt extract in50mM sodium phosphate buffer pH 5.9 (Gutz et al.,1974). For ascospore isolation, samples of sporu-lated cultures were subjected to isopycnic centri-fugation on linear density gradients of urografin(25–55%, v/v) in a swinging-bucket rotor at 25,000gfor 30min and 4 1C (Nishi et al., 1978). The samplesresulted partitioned after centrifugation in threedistinct isolated bands that were analyzed byphase-contrast microscopy. The upper fractioncontained cellular debris and unbroken asci whilethe intermediate fraction was composed of remain-ing whole vegetative cells. Isolated intact ascos-pores were located purified in the band of moredensity (45–50% urografin), extensively washedwith distilled water at low-speed centrifugation,resuspended in assay buffer and directly used asenzyme source in most experiments.
Treatment of spores, preparation of cellsextracts and cell walls
Mild acid treatments of spores were performedessentially as indicated by Arnold (1972). Briefly,spores were subjected to 1-h treatments at roomtemperature in 0.1 N HCl or 3 N H3PO4 and thenexhaustively washed on the centrifuge with 50mMacetate buffer, pH 4.6. The percentage of live cellswas ascertained by the methylene blue test inconjunction with a hemocytometer (Arnold, 1972).In parallel, spore viability was determined by theplate forming units method. Cell-free extracts fromtreated spores were prepared by mechanicaldisruption with glass beads as described previously(Carrillo et al., 1992) and tested for trehalaseand marker enzymes against cell extracts from

ARTICLE IN PRESS
Table 1. Distribution of trehalase activity after iso-picnic centrifugation of cell cultures in linear gradients ofurografin (25–55%)
Fraction Trehalase(units/assay)
Specificactivity(units/109
cells)
Content
Upper 0 0 Cell debrisIntermediate 0 0 Vegetative
cellsBottom 2.7 28.4 Spores
J. Vicente-Soler et al.306
untreated spores. Isolated cell walls were obtainedaccording to Perez and Ribas (2004). The lysingenzyme complex from Trichoderma harzianum(Glucanex; Novozyme Corporation, Sigma), whichwas found free of trehalase activity, was used totreat spore cell walls. The cell walls were washedin 50mM acetate buffer pH 4.6, resuspended to anOD600 of 1.0 in the same buffer containing 20 ml/mlGlucanex and the suspension was incubated at 30 1Cwith shaking. The resulting supernatants weremonitored every hour for trehalase after elimina-tion of the glucose background by gel filtration asindicated below.
Enzyme assays
In trehalase assays, 0.25ml of either sporesuspension (109 spores/ml) or the correspondingcell extract was added to 0.25ml containing200moles trehalose in 50mM acetate buffer pH4.6. Sodium azide (1mM) was routinely included inthese assays to reduce potential uptake of theresulting glucose. After incubation at 30 1C withshaking the reactions were stopped by heating. Theassay mixtures were centrifuged at low-speedcentrifugation and the glucose originated in thereactions was measured in the supernatants bythe glucose oxidase–peroxidase method (Carrilloet al., 1994). One enzyme unit released 1 nmolof glucose per min under the stated conditions.The intracellular enzymes alkaline phosphatase anda-glucosidase were used as reference markers inexperiments for enzyme location and their activitydetermined according to Fernandez et al. (1995).Maltase activity was measured as for a-glucosidasebut with maltose as substrate. Assays for trehalaseenzyme activation by phosphorylation were per-formed as described previously (Arguelles et al.,1986).
Gel filtration and non-denaturingelectrophoresis
Elimination of glucose from lysate samples wasperformed by gel filtration using small SephadexG-50 packed columns and recovering the enzyme inthe elution void volume. The relative molecularweight of trehalase solubilized from the cell wallswas estimated by gel filtration through a SephadexG-200 column (60 cm� 2.5 cm) equilibrated withassay buffer. The elution pattern was calibratedusing the following markers: dextran blue(2000 kDa), ferritin (450 kDa), catalase (240 kDa)and aldolase (158 kDa). The trehalase was resolvedunder native conditions by electrophoresis through
20 cm long, 5% polyacrylamide gels. The gels wererun for 24 h at 250 V and trehalase activity wasdetected in situ by staining as described by Morenoet al. (1990) except that sucrose was changed fortrehalose.
Results
Trehalase activity in ascospores
Intact ascospores of the wild type Sch. pombestrain 968 showed enzyme activity in standard acidtrehalase assays. Table 1 shows a typical result onthe distribution of the enzyme activity among thedifferent bands obtained after fractionation ofsporulated cultures by isopycnic centrifugation(see Materials and Methods). When cell samples ofthe resulting bands were washed and used asenzyme source without previous breakage, treha-lase activity correlated exclusively to the presenceof the undisrupted ascospores. No trehalase wasdetected associated to intact vegetative cells.
Since Sch. pombe cells of the homothallic strain968 also contain a neutral trehalase it could beargued that the potential contribution of suchenzyme to the trehalose breakdown was notcompletely dismissed in the above assays. To avoidmiss-interpretation of the results we used insubsequent experiments ascospores from the ntp1-disrupted strain MMT48, which is isogenic to strain968 but devoid of neutral trehalase activity due todisruption of the ntp1 gene (Beltran et al., 2000).The results showed that whole ascospores of Sch.pombe lacking neutral trehalase also hydrolyzedtrehalose, pointing to the existence of an indepen-dent enzyme able to cleave this disaccharide.
Localization of acid trehalase
Exposure of intact ascospores from the MMT48strain to mild acid treatments completely inactivated

ARTICLE IN PRESS
Solubilization and characterization of a cell wall-bound trehalase 307
the acid trehalase activity originally present inuntreated samples under conditions in which morethan 90% of the spores remained viable. In contrast,as summarized in Table 2, the acid treatments did notsignificantly affect the activity of intracellularenzyme markers. These results are congruent withthe interpretation that the trehalase enzyme islocated at the cell surface. The fact that acid-treated ntp1-disrupted spores contain almost a fullcomplement of intracellular marker enzymes whilethey are no longer potential sources for trehalaseactivity, together with the observation of a rapidinactivation of the solubilized enzyme at pH valuesbelow 2.0 (see later), supports that this trehalasemust be located external to the cell membrane.
To gain more insight into the localization of theenzyme we devised an assay method for trehalasedetermination alternative to the original glucose-oxidase/peroxidase procedure. The assay wasbased on the use of eugenol (2-metoxi-4-alil-fenol)instead of the chromogenic oxidized substrateo-dianisidine. The complete procedure involved acoupled reaction between the oxidation of theglucose generated from trehalose by trehalase andthe oxidation of eugenol by peroxidase, whichresults in polymerized eugenol as a white insolubleprecipitate where glucose is being released. Therationale behind this approach was that if glucosewas being produced outside the cells, the forma-tion of a white precipitate in the enzyme assayswould then be observed. The data supported thatglucose originates extracellularly in assays withintact ascospores and exogenously added treha-lose. These findings suggested that the enzyme was
Table 2. Relative location of acid trehalase in ascos-pores from ntp1-disrupted Sch. pombe
Treatment of spores
None 0.1N HClfor 10min
3N H3PO4
for 10min
Viability(methylene bluetest)a
99.1% 96.2% 88.7%
Viability (platecounts)b
100% 83.2% 76.4%
Alkalinephosphatasec
515.1 502.6 461.4
a-glucosidasec 283.4 266.1 230.7Acid trehalased 27.9 o0.1 o0.1
aPercentage of live cells as judged by methylene blue test.bColony formation ability after 6-day incubation.cEnzyme activities are given as units per 109 cells determined inspore extracts.dEnzyme activity given as units per 109 cells determined in wholespores.
either periplasmic or cell wall-located in vivo.Additional evidence for the external location of thistrehalase derived from fractionation of spore cellextracts. After low speed centrifugation (3000g for5min), the bulk of the enzyme activity present inthe extracts (490%) was in the particulatedfraction and recoverable in the cell wall-rich pelletobtained after exhaustive washings.
Fractions enriched in cell walls were also assayedfor maltase and a-glucosidase and no activity wasfound. The absence of activity in these fractionscontaining acid trehalase using p-nitrophenyl-a-glucoside as substrate is congruent with theobservation that trehalase is acid-sensitive in vivowhereas a-glucosidase is not (Table 2).
Solubilization of the cell wall-boundtrehalase
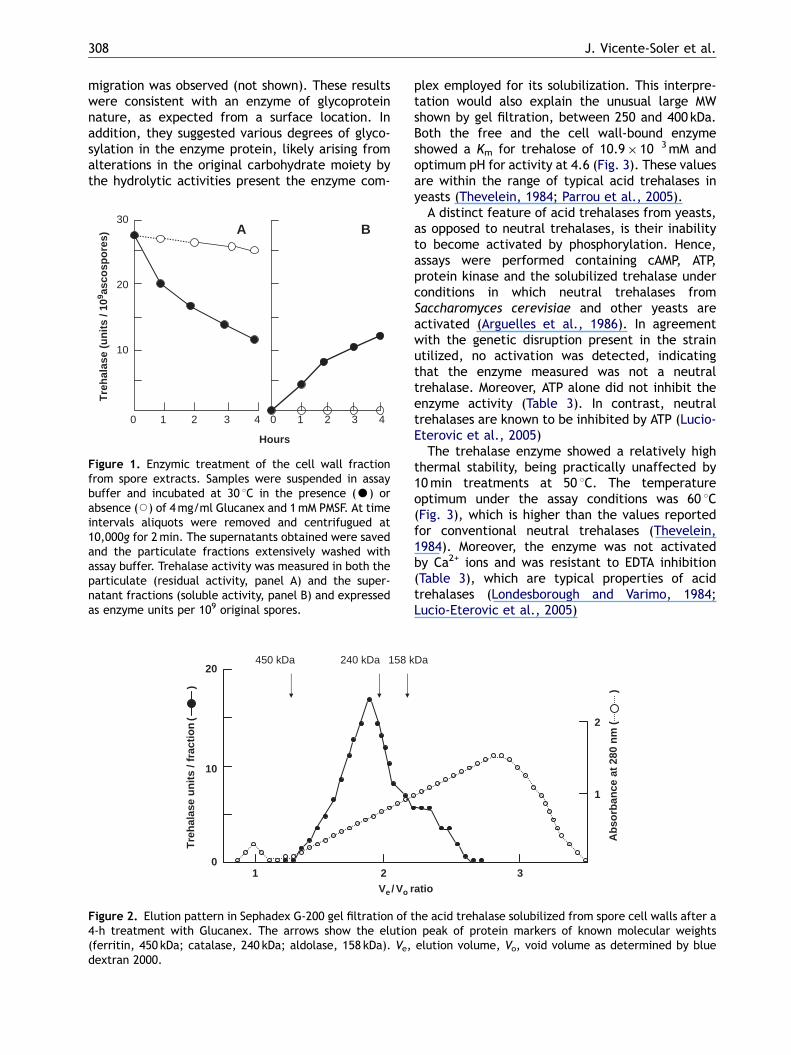
Attempts to release the acid trehalase bytreating the particulated fractions with solutionsof increased NaCl concentrations or 2% SDS failed torender soluble the acid trehalase activity in thecorresponding supernatants, suggesting a covalentlinkage (Cassone et al., 1979; Perez and Ribas,2004). The cell wall fractions were then treated at30 1C with the lytic complex Glucanex, which isable to partially degrade the ascospore envelope.The resulting enzyme lysates showed a high glucosecontent, which seriously hindered trehalase deter-minations, likely arising from partial hydrolysis ofthe cell wall structural glucans. Hence, to performin the absence of glucose a follow-up of thepotential cell wall-bound trehalase being released,the supernatants of the lysates were analyzed fortrehalase determination after passing throughsmall Sephadex G-50 columns and employing theobtained void volume fractions as enzyme source.Fig. 1 shows data on the decrease of trehalase inthe particulate fraction and its concomitant ap-pearance as soluble form upon treatment with thelysing complex. Typical results indicated that about60% of the original enzyme activity was recoveredas free active form after 4 h of incubation withGlucanex (Fig. 1).
Properties of the solubilized enzyme
The trehalase solubilized from the ascosporecell walls showed a broad elution pattern bygel filtration chromatography in Sephadex G-200(Fig. 2). Moreover, when the activity was detectedon polyacrylamide gels by several in situ stainingprocedures after non-denaturing electrophoresis(Moreno et al., 1990; Sanchez et al., 2003) a fuzzy
ARTICLE IN PRESS
J. Vicente-Soler et al.308
migration was observed (not shown). These resultswere consistent with an enzyme of glycoproteinnature, as expected from a surface location. Inaddition, they suggested various degrees of glyco-sylation in the enzyme protein, likely arising fromalterations in the original carbohydrate moiety bythe hydrolytic activities present the enzyme com-
1
Ve/ Vo r
0
10
20450 kDa 240 kDa 158 k
Tre
hala
se u
nit
s / f
racti
on
( )
2
Figure 2. Elution pattern in Sephadex G-200 gel filtration of t4-h treatment with Glucanex. The arrows show the elution(ferritin, 450 kDa; catalase, 240 kDa; aldolase, 158 kDa). Ve,dextran 2000.
1 2 3 40 1 3
10
20
30A
Tre
hala
se (
un
its / 1
09asco
sp
ore
s)
Hours
B
420
Figure 1. Enzymic treatment of the cell wall fractionfrom spore extracts. Samples were suspended in assaybuffer and incubated at 30 1C in the presence (K) orabsence (J) of 4mg/ml Glucanex and 1mM PMSF. At timeintervals aliquots were removed and centrifugued at10,000g for 2min. The supernatants obtained were savedand the particulate fractions extensively washed withassay buffer. Trehalase activity was measured in both theparticulate (residual activity, panel A) and the super-natant fractions (soluble activity, panel B) and expressedas enzyme units per 109 original spores.
plex employed for its solubilization. This interpre-tation would also explain the unusual large MWshown by gel filtration, between 250 and 400 kDa.Both the free and the cell wall-bound enzymeshowed a Km for trehalose of 10.9� 10�3mM andoptimum pH for activity at 4.6 (Fig. 3). These valuesare within the range of typical acid trehalases inyeasts (Thevelein, 1984; Parrou et al., 2005).
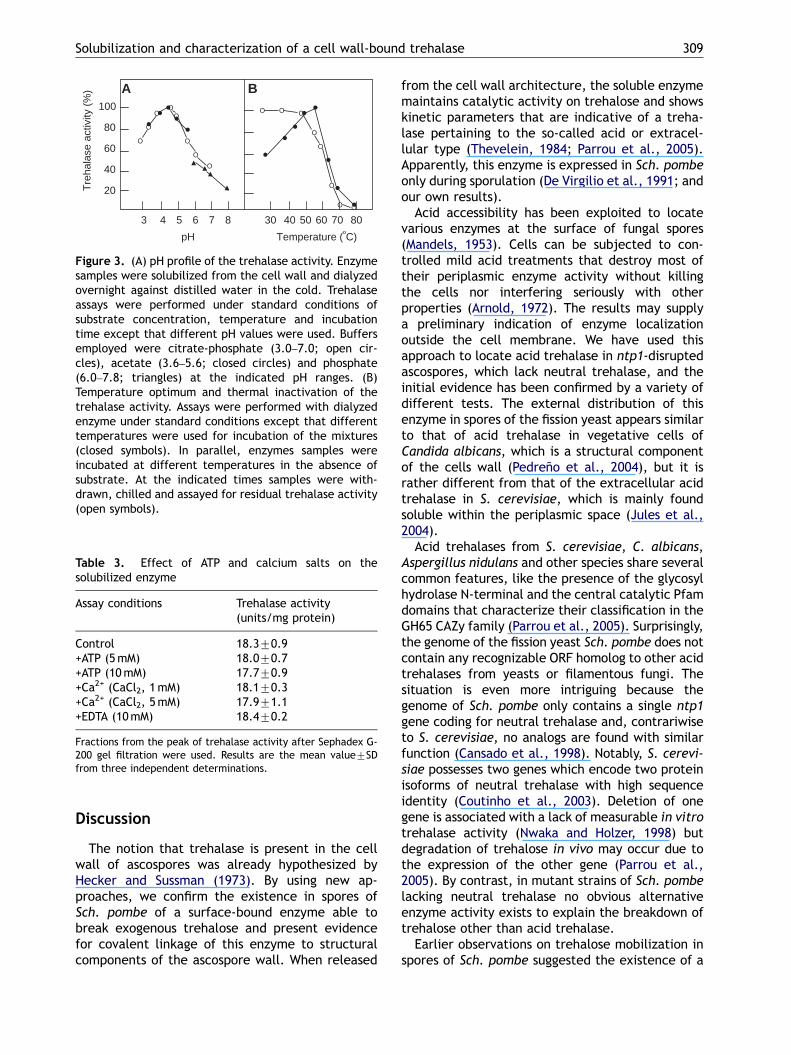
A distinct feature of acid trehalases from yeasts,as opposed to neutral trehalases, is their inabilityto become activated by phosphorylation. Hence,assays were performed containing cAMP, ATP,protein kinase and the solubilized trehalase underconditions in which neutral trehalases fromSaccharomyces cerevisiae and other yeasts areactivated (Arguelles et al., 1986). In agreementwith the genetic disruption present in the strainutilized, no activation was detected, indicatingthat the enzyme measured was not a neutraltrehalase. Moreover, ATP alone did not inhibit theenzyme activity (Table 3). In contrast, neutraltrehalases are known to be inhibited by ATP (Lucio-Eterovic et al., 2005)
The trehalase enzyme showed a relatively highthermal stability, being practically unaffected by10min treatments at 50 1C. The temperatureoptimum under the assay conditions was 60 1C(Fig. 3), which is higher than the values reportedfor conventional neutral trehalases (Thevelein,1984). Moreover, the enzyme was not activatedby Ca2+ ions and was resistant to EDTA inhibition(Table 3), which are typical properties of acidtrehalases (Londesborough and Varimo, 1984;Lucio-Eterovic et al., 2005)
3
atio
1
2
Da
Ab
so
rban
ce a
t 280 n
m (
)
he acid trehalase solubilized from spore cell walls after apeak of protein markers of known molecular weights
elution volume, Vo, void volume as determined by blue
ARTICLE IN PRESS
Tre
hala
se a
ctivity (
%)
3 4 5 6 7 8
20
40
60
80
100
30 40 50 60 70 80
pH Temperature (ºC)
A B
Figure 3. (A) pH profile of the trehalase activity. Enzymesamples were solubilized from the cell wall and dialyzedovernight against distilled water in the cold. Trehalaseassays were performed under standard conditions ofsubstrate concentration, temperature and incubationtime except that different pH values were used. Buffersemployed were citrate-phosphate (3.0–7.0; open cir-cles), acetate (3.6–5.6; closed circles) and phosphate(6.0–7.8; triangles) at the indicated pH ranges. (B)Temperature optimum and thermal inactivation of thetrehalase activity. Assays were performed with dialyzedenzyme under standard conditions except that differenttemperatures were used for incubation of the mixtures(closed symbols). In parallel, enzymes samples wereincubated at different temperatures in the absence ofsubstrate. At the indicated times samples were with-drawn, chilled and assayed for residual trehalase activity(open symbols).
Table 3. Effect of ATP and calcium salts on thesolubilized enzyme
Assay conditions Trehalase activity(units/mg protein)
Control 18.370.9+ATP (5mM) 18.070.7+ATP (10mM) 17.770.9+Ca2+ (CaCl2, 1mM) 18.170.3+Ca2+ (CaCl2, 5mM) 17.971.1+EDTA (10mM) 18.470.2
Fractions from the peak of trehalase activity after Sephadex G-200 gel filtration were used. Results are the mean value7SDfrom three independent determinations.
Solubilization and characterization of a cell wall-bound trehalase 309
Discussion
The notion that trehalase is present in the cellwall of ascospores was already hypothesized byHecker and Sussman (1973). By using new ap-proaches, we confirm the existence in spores ofSch. pombe of a surface-bound enzyme able tobreak exogenous trehalose and present evidencefor covalent linkage of this enzyme to structuralcomponents of the ascospore wall. When released
from the cell wall architecture, the soluble enzymemaintains catalytic activity on trehalose and showskinetic parameters that are indicative of a treha-lase pertaining to the so-called acid or extracel-lular type (Thevelein, 1984; Parrou et al., 2005).Apparently, this enzyme is expressed in Sch. pombeonly during sporulation (De Virgilio et al., 1991; andour own results).
Acid accessibility has been exploited to locatevarious enzymes at the surface of fungal spores(Mandels, 1953). Cells can be subjected to con-trolled mild acid treatments that destroy most oftheir periplasmic enzyme activity without killingthe cells nor interfering seriously with otherproperties (Arnold, 1972). The results may supplya preliminary indication of enzyme localizationoutside the cell membrane. We have used thisapproach to locate acid trehalase in ntp1-disruptedascospores, which lack neutral trehalase, and theinitial evidence has been confirmed by a variety ofdifferent tests. The external distribution of thisenzyme in spores of the fission yeast appears similarto that of acid trehalase in vegetative cells ofCandida albicans, which is a structural componentof the cells wall (Pedreno et al., 2004), but it israther different from that of the extracellular acidtrehalase in S. cerevisiae, which is mainly foundsoluble within the periplasmic space (Jules et al.,2004).
Acid trehalases from S. cerevisiae, C. albicans,Aspergillus nidulans and other species share severalcommon features, like the presence of the glycosylhydrolase N-terminal and the central catalytic Pfamdomains that characterize their classification in theGH65 CAZy family (Parrou et al., 2005). Surprisingly,the genome of the fission yeast Sch. pombe does notcontain any recognizable ORF homolog to other acidtrehalases from yeasts or filamentous fungi. Thesituation is even more intriguing because thegenome of Sch. pombe only contains a single ntp1gene coding for neutral trehalase and, contrariwiseto S. cerevisiae, no analogs are found with similarfunction (Cansado et al., 1998). Notably, S. cerevi-siae possesses two genes which encode two proteinisoforms of neutral trehalase with high sequenceidentity (Coutinho et al., 2003). Deletion of onegene is associated with a lack of measurable in vitrotrehalase activity (Nwaka and Holzer, 1998) butdegradation of trehalose in vivo may occur due tothe expression of the other gene (Parrou et al.,2005). By contrast, in mutant strains of Sch. pombelacking neutral trehalase no obvious alternativeenzyme activity exists to explain the breakdown oftrehalose other than acid trehalase.
Earlier observations on trehalose mobilization inspores of Sch. pombe suggested the existence of a

ARTICLE IN PRESS
J. Vicente-Soler et al.310
mechanism during germination that would requirethe export of the disaccharide, its extracellularhydrolysis, and the subsequent uptake of theglucose released (Inoue and Shimoda, 1981; Beltranet al., 2000). This hypothesis has been in partsupported by the observation that the deletion ofath1+ (encoding acid trehalase) in nth1�nth2�
double mutant cells of S. cerevisiae (which lackthe two genes coding for neutral trehalases) resultsin the accumulation of extracellular trehalose(Parrou et al., 2005). Data presented here con-cerning the enzyme location are thus compatiblewith the above interpretation.
Several lines of evidence support that in spite oflacking common sequences, the cell wall-locatedenzyme of Sch. pombe shares properties similar toother acid trehalases from different yeasts species.First, not only its optimum pH is within the acidrange, but it exhibits both a temperature optimumand a thermal resistance higher than neutraltrehalases (Thevelein, 1984). Second, it is insensi-tive to calcium salts and lacks inhibition bynucleotides such as ATP. Finally, like many otheracid trehalases, it is located outside the cell and,hence, far from the post-translational controlwhich regulate the activity of neutral trehalases.However, it does not appear to be inhibited byacetate, unlike other reported acid trehalases(Fig. 3) (Londesborough and Varimo, 1984; Ar-guelles and Gacto, 1985).
Taken as a whole, the results presented in thisstudy add insight into the nature and physiologicalsignificance of the acid trehalase in the fission yeast,which was biochemically recognized as such byseveral workers before fully sequencing of the fissionyeast genome. The solubilization of this enzymefavors its purification and may eventually facilitatethe design of molecular probes for the identificationof the corresponding gene within the Sch. pombegenome. The enzyme might represent a novel classof trehalases not fitting among the types so farestablished on the bases of sequence homology.Further characterization of enzymes similar to thisparticular hydrolase of the spore coat of Sch. pombemay help to broaden the knowledge on trehalasesand their role in yeast biology.
References
Arguelles JC, Gacto M. Evidence for regulatory trehalaseactivity in Candida utilis. Can J Microbiol 1985;31:529–37.
Arguelles JC, Vicente-Soler J, Gacto M. Protein phosphor-ylation and trehalase activation in Candida utilis.FEMS Microbiol Lett 1986;34:361–5.
Arnold WN. The structure of yeast cell wall: solubilizationof a marker enzyme, b-fructofuranosidase, by theautolytic enzyme system. J Biol Chem 1972;247:1161–9.
Beltran FF, Castillo R, Vicente-Soler J, Cansado J, GactoM. Role for trehalase during germination of spores inthe fission yeast Schizosaccharomyces pombe. FEMSMicrobiol Lett 2000;193:117–21.
Cansado J, Soto T, Fernandez J, Vicente-Soler J, Gacto M.Characterization of mutants devoid of neutral treha-lase activity in the fission yeast Schizosaccharomycespombe: partial protection from heat shock and high-salt stress. J Bacteriol 1998;180:1342–5.
Carrillo D, Vicente-Soler J, Gacto M. Activation of neutraltrehalase by fermentable sugars and cAMP in thefission yeast Schizosaccharomyces pombe. FEMS Micro-biol Lett 1992;98:61–6.
Carrillo D, Vicente-Soler J, Gacto M. Cyclic AMP signallingpathway and trehalase activation in the fission yeastSchizosaccharomyces pombe. Microbiology 1994;140:1467–72.
Cassone A, Kerridge D, Gale EF. Ultrastructural changes inthe cell wall of Candida albicans following cessation ofgrowth and their possible relationship to the devel-opment of polyene resistance. J Gen Microbiol 1979;110:339–49.
Coutinho PM, Deleury E, Davies GJ, Henrissat B. Anevolving hierarchical family classification for glycosyl-transferases. J Mol Biol 2003;328:307–17.
De Virgilio C, Muller J, Boller T, Wiemken A. Aconstitutive, heat-shock activated neutral trehalaseoccurs in Schizosaccharomyces pombe in addition tothe sporulation-specific acid trehalase. FEMS MicrobiolLett 1991;84:85–90.
d’Enfert C, Fontaine T. Molecular characterization of theAspergillus nidulans treA gene encoding an acidtrehalase required for growth on trehalose. MolMicrobiol 1997;24:203–16.
Fernandez J, Soto T, Vicente-Soler J, Cansado J, Gacto M.Increased thermal stability of the enzyme content inpermeabilized whole cells from the fission yeastSchizosaccharomyces pombe by exogenous trehaloseand other compounds. Can J Microbiol 1995;41:936–41.
Gutz H, Heslot H, Leupold U, Loprieno N. Schizosacchar-omyces pombe. In: King RC, editor. Handbook ofgenetics, vol. I. New York, London: Plenum Press;1974. p. 395–446.
Hecker LI, Sussman AS. Localization of trehalase in theascospores of Neurospora: relation to ascosporedormancy and germination. J Bacteriol 1973;115:592–9.
Inoue H, Shimoda C. Changes in trehalose content andtrehalase activity during spore germination in fissionyeast, Schizosaccharomyces pombe. Arch Microbiol1981;129:19–22.
Jules M, Guillou V, Franc-ois J, Parrou JL. Two distinctpathways for trehalose assimilation in the yeastSaccharomyces cerevisiae. Appl Environ Microbiol2004;70:2771–8.

ARTICLE IN PRESS
Solubilization and characterization of a cell wall-bound trehalase 311
Londesborough J, Varimo K. Characterization of twotrehalases in baker’s yeast. Biochem J 1984;219:511–8.
Lucio-Eterovic AK, Jorge JA, Polizeli ML, Terenzi HF.Biochemical characterisation of the trehalase ofthermophilic fungi: an enzyme with mixed propertiesof neutral and acid trehalase. Biochim Biophys Acta2005;1723:201–7.
Mandels GR. Localization of carbohydrases at the surfaceof fungus spores by acid treatment. Exp Cell Res1953;5:48–55.
Moreno S, Sanchez Y, Rodriguez L. Purification andcharacterization of the invertase from Schizosacchar-omyces pombe. A comparative analysis with theinvertase from Saccharomyces cerevisiae. Biochem J1990;267:697–702.
Nishi K, Shimoda C, Hayashibe M. Germination andoutgrout of Schizosaccharomyces pombe ascosporesisolated by urografin density gradient centrifugation.Can J Microbiol 1978;24:893–7.
Nwaka S, Holzer H. Molecular biology of trehaloseand the trehalases in the yeast Saccharomyces
cerevisiae. Prog Nucleic Acid Res Mol Biol 1998;58:197–237.
Parrou JL, Jules M, Beltran G, Franc-ois J. Acid trehalasein yeasts and filamentous fungi: localization, regula-tion and physiological function. FEMS Yeast Res2005;5:503–11.
Pedreno Y, Maicas S, Arguelles JC, Sentandreu R,Valentin E. The ATC1 gene encodes a cell wall-linked acid trehalase required for growth ontrehalose in Candida albicans. J Biol Chem 2004;279:40852–60.
Perez P, Ribas JC. Cell wall analysis. Methods 2004;33:245–51.
Sanchez R, Franco A, Gacto M, Notario V, Cansado J.Characterization of gdp1+ as encoding a GDPase in thefission yeast Schizosaccharomyces pombe. FEMS Micro-biol Lett 2003;228:33–8.
Thevelein JM. Regulation of trehalose mobilization infungi. Microbiol Rev 1984;48:42–59.
Van Laere A. Trehalose, reserve and/or stress metabolite?FEMS Microbiol Rev 1989;63:201–10.




















