
Bacillus anthracis Edema Toxin Impairs Neutrophil Actin-Based Motility
Upload
independentCategory
view
3download
0
doi: 10.1111/j.1365-2869.2010.00823.x
Sleep restriction for the duration of a work week impairs
multitasking performance
MARJA - LEENA HAAV I STO 1 , 2 , TAR JA PORKKA -HE I SKANEN 3 ,
CHR I STER HUBL IN 1 , M IKKO H ARMA 4 , P ERTT I MUTANEN 5 , K I T I
M ULLER 1 , JU S S I V IRKKALA 1 , 6 and M IKAEL SALL INEN 1 , 7
1Brain and Work Research Centre, Finnish Institute of Occupational Health, Helsinki, Finland, 2Technical Research Centre of Finland, Espoo,
Finland, 3Department of Physiology, Institute of Biomedicine, University of Helsinki, Finland, 4Centre of Expertise Human Factors at Work,
Finnish Institute of Occupational Health, Helsinki, Finland, 5Statistical Services, Finnish Institute of Occupational Health, Helsinki, Finland,6Department of Clinical Neurophysiology, Medical Imaging Centre, Pirkanmaa Hospital District, Tampere, Finland and 7Agora Center,
University of Jyvaskyla, Jyvaskyla, Finland
Accepted in revised form 2 December 2009; received 2 June 2009
SUMMARY It is important to develop shift schedules that minimise the chance for sleep-related
human error in safety-critical domains. Experimental data on the effects of sleep
restriction (SR) play a key role in this development work. In order to provide such data,
we conducted an experiment in which cognitively demanding and long-duration task
performance, simulating task performance at work, was measured under SR and
following recovery. Twenty healthy male volunteers, aged 19–29 years, participated in
the study. Thirteen of them had first two baseline days (8-h sleep opportunity per day),
then five SR days (4-h sleep) and finally two recovery days (8-h sleep). Seven controls
were allowed to sleep for 8 h each night. On each experimental day, multitask
performance was tested in 50-min sessions, physiological sleepiness was evaluated
during multitask performance using electroencephalogram (EEG) ⁄ electrooculogram(EOG) recordings, and psychomotor vigilance task performance and Karolinska
Sleepiness Scale were recorded. Sleep–wake rhythm was monitored throughout the
experiment. The multitask performance progressively deteriorated as a result of
prolongation of the SR and the time spent on the task. The effect was significant at
group level, but individual differences were large: performance was not markedly
deteriorated in all participants. Similar changes were observed also in EEG ⁄EOG-
defined sleepiness. The recovery process of performance and sleepiness from the SR
continued over the two recovery sleep opportunities. In all, our findings emphasise the
importance of shift systems that do not restrict sleep for several consecutive days.
k e y w o r d s cumulative sleep restriction, multitask performance, recovery, sleepiness
INTRODUCTION
In many safety-critical occupations, work consists of multiple
tasks that need to be performed simultaneously. To cope with
these tasks, called multitasks, the workers are required to
switch their attention between subtasks and make decisions on
priorities (Navon and Gopher, 1979; Wickens, 2002; Wickens
et al., 2003). Examples of occupations requiring multitasking
are drivers, pilots, traffic controllers and process operators,
who simultaneously search information from several sources
and have multiple parallel subtasks under processing.
Employees in safety-critical occupations often work under
acute and ⁄or cumulative sleep loss because of irregular and
long work hours (Folkard and Lombardi, 2006; Harma et al.,
2002; Sallinen et al., 2003). The combination of multitasking
and being sleep deprived is potentially hazardous: in trans-
portation and industry, where multitasking is common,
restricted sleep opportunities are a major cause of accidents
Correspondence: Marja-Leena Haavisto, Organisational Psychology,
Technical Research Centre of Finland, P.O. Box 1000, FI-02044 VTT,
Finland. Tel.: +358 40 7056443; fax: +358 20 7225888; e-mail: marja-
J. Sleep Res. (2010) 19, 444–454 Sleep restriction and multitasking
444 � 2010 European Sleep Research Society

(Caldwell, 2005; Philip and Akerstedt, 2006). For example, the
National Transportation Safety Board (NTSB) in the USA has
estimated that fatigue-related accidents involving heavy trucks
make up to 30% of fatal accidents (NTSB, 1990). Previous
research supports the view that restricted sleep markedly
degrades performance in multitasking (Caldwell and Caldwell,
1998; Caldwell and Ramspott, 1998; Elsmore, 1994; Sallinen
et al., 2008). However, from the viewpoint of work life, at least
three important questions have so far been inadequately
addressed by previous research.
The first question concerns cumulative sleep loss, that is,
partial sleep loss across several consecutive days. Until now,
studies of cumulative sleep restriction (SR) have shown that
performance on short duration, usually vigilance tasks grad-
ually degrades in the course of SR (Belenky et al., 2003; Van
Dongen et al., 2003; Webb and Agnew, 1974). Recently it has
been estimated that the extension of wakefulness to >20 h a
day (i.e. < 4 h per night) leads to an escalation of perfor-
mance impairment (McCauley et al., 2009). Although several
studies have addressed multitask performance after acute sleep
loss (Caldwell and Caldwell, 1998; Caldwell and Ramspott,
1998; Elsmore, 1994; Sallinen et al., 2008), to the best of our
knowledge performance after cumulative SR has been
addressed in a single study (Balkin et al., 2004).
Second, in many safety-critical occupations performance
needs to be maintained at a high level for long periods of time
uninterrupted. The time-on-task effect, that is, a progressive
deterioration of performance in the course of the task session,
is affected by SR (Dinges and Kribbs, 1991). Thus, it can be
argued that the duration of the task may be a critical factor in
occupational safety. Until now, the time-on-task effect on
multitasking has been studied in two acute sleep deprivation
studies. In one of these studies, the time-on-task effect was
markedly increased after the first 10 min on the task (Caldwell
and Ramspott, 1998), whereas in the other the effect just
approached significance in a 70-min test session (Sallinen
et al., 2008). Studies on the time-on-task effect on multitasking
under cumulative SR are lacking.
The third understudied question regards recovery from
partial cumulative sleep loss. This information is crucial when
planning shift work schedules; particularly when considering
the number of days off between two consecutive shift spells.
Until now, recovery of multitasking performance has been
examined in one study showing that 8-h sleep after 1 night of
partial sleep loss is not sufficient for full recovery (Sallinen
et al., 2008). Recently, Banks et al. (2007) and McCauley et al.
(2009) have shown that a single recovery night of extended
sleep after cumulative SR improves participants� performance,
but leaves their vulnerability to sleep deprivation at an
elevated level. The pace of recovery after SR is affected by at
least two determinants: the first of which is severity of SR – the
more severe the sleep loss, the longer the period of recovery
required (Lamond et al., 2007); the second element is the
nature of sleep loss – recovery from cumulative sleep loss
appears to require more time than recovery from acute sleep
loss (Axelsson et al., 2008; Belenky et al., 2003; Lamond et al.,
2007). In addition, the methods used to measure the recovery
process matter, for example, the recovery of subjective
sleepiness occurs earlier than the recovery of physiological
sleepiness (Lamond et al., 2007). With this in mind, there is a
clear need to ascertain how multitasking performance recovers
from cumulative sleep loss.
This study was aimed at elucidating the three above-
mentioned practical questions. We hypothesised that cumula-
tive sleep loss, reflecting a restriction of sleep for duration of a
work week, leads to progressive impairments in multitasking.
We predicted that the degree of impairment would be affected
by both the duration of the task (the time-on-task effect) and
the number of days of restricted sleep (extent of sleep loss). We
were also interested in the recovery process, and posed the
question whether 2 days, simulating a weekend, are sufficient
for recovery of a multitask performance of long duration. We
curtailed young male volunteers� sleep opportunity for 5 days,
and thereafter allowed them to sleep normally for 2 nights
while measuring their multitask performance in two 50-min
sessions every day. To examine the overall effect of the SR and
the recovery days, we also measured participants� subjective,behavioural and physiological sleepiness in the course of the
experiment. The results from the SR group were compared
with those from a group that was offered a sleep opportunity
of 8 h each night through the experiment.
MATERIALS AND METHODS
Participants
After signing a form for informed consent, 20 healthy men
(aged 19–29 years) with 7–9 h of habitual sleep and sleep need
voluntarily participated in the study. The measure of habitual
sleep need was based on participant�s subjective evaluation in
the questionnaire. The Ethics Committee of the Hospital
District of Helsinki and Uusimaa approved the study. Prior to
the study, participants were screened to exclude those with
extreme circadian types, sleep disorders, psychiatric illness,
chronic or recent acute medical conditions, a history of drug or
alcohol dependence, having crossed time zones during 4 weeks
preceding the beginning of the study, habitual napping, and
shift work and ⁄or night work. Positive criteria for selection
included regular lifestyles with habitual bedtime before
24:00 hours and wake-up time after 06:00 hours. The Nordic
Sleep Questionnaire (Partinen and Gislason, 1995), medical
screening questionnaires, and clinical blood and urine labora-
tory tests were used to identify and exclude individuals with
drug dependence, sleep disorders and other conditions. In the
second phase, the participants were examined by a physician to
ensure fitness for the experiment.
Procedures prior to the experiment
At least 2 weeks before the experiment, the participants
slept an adaptation night in the laboratory, and a polysomn-
ogram, including electroencephalography (EEG), bilateral
Sleep restriction and multitasking 445
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

electrooculography (EOG), submental electromyography and
electrocardiography, was recorded to exclude persons suffering
from organic sleep disorders.
The participants were instructed to maintain a regular sleep–
wake cycle for 2 weeks prior to the study, which was verified by
wrist-worn actigraphy and sleep diary recordings. In addition,
they were instructed to maintain a regular nutrition schedule,
and to refrain from caffeine, alcohol and tobacco for 2 weeks
before arriving in the sleep laboratory. Mean sleep duration
was 6 h 53 min (SD = 35.0 min) in the SR group and 7 h
(SD = 51.4 min) in the control group during the 14 days
preceding the experiment derived from actigraphy data.
Design and experimental procedures
Participants were randomly selected to the experimental or
control groups. Participants spent 10.5 consecutive days in the
laboratory (Fig. 1). The first day was an adaptation ⁄ trainingday (A) and the second served as baseline (BL). During A,
participants became familiar with the day schedule, and
practised the tasks and self-rating scales. Thirteen of the
participants underwent sleep deprivation conditions, including
the training and baseline day (8 h in bed per night, 23:00–
07:00 hours), 5 days of partial sleep loss (4 h in bed per night,
03:00–07:00 hours), and two recovery days (8 h in bed per
night, 23:00–07:00 hours). The remaining seven volunteers
were allowed to sleep for 8 h per night throughout the
experiment.
No alcohol, tobacco or caffeine was allowed during the
laboratory visit. The fixed meal hours and amount of calories
per meal were as follows: breakfast 600 kcal at 07:30 hours;
lunch 800 kcal at 12:30 hours; snack 300 kcal at 15:30 hours;
dinner 700 kcal at 18:00 hours; and snack 200 kcal at
21:30 hours. The participants were under behavioural moni-
toring, and their sleepiness was measured by EEG and EOG
24 h per day.
The study was designed to simulate a typical work week with
daily working time between 07:00 and 16:30 hours. The
participants completed a 50-min multitask session at 10:00,
11:40 and 14:00 hours on each of the 9 days. One of the two
forenoon sessions contained a 10-min rest pause, and the data
from this session were not included in this study. The
scheduling of the forenoon multitask sessions with and without
the 10-min break was counterbalanced across the days and
participants. To avoid the knowledge-of-results effect, the
participants were not provided with feedback from their
performance during the experimental days.
The psychomotor vigilance task performance (PVT) was
administrated each day at 07:10, 11:00 and 15:00 hours. The
Karolinska Sleepiness Scale (KSS) was rated at the beginning
and end of each multitask session. Each time, two participants
belonging to the same group spent the night at the laboratory
at the same time. Between the task sessions, participants were
allowed to read, watch TV or movies, and interact with each
other and the laboratory staff helped them to stay awake. In
order to avoid light exposure, going outdoors was not
permitted. In addition, physical exercise was not allowed
during the laboratory experiment. Illumination in the sleeping
room and in the test room ranged from 150 to 400 lux, and in
the living room from 350 to 600 lux. The temperature ranged
from 19 to 23 �C.
EEG recording
Electroencephalogram was recorded from 10 to 20 system
derivations Fp1-A2, Fp2-A1, C3-A2, C4-A1, O1-A2 and O2-
A1. The recordings were conducted with a digital recorder
(Embla, Flaga HF, Reykjavik, Iceland), using a sampling rate
of 200 Hz with a bandwidth of 0.5–90 Hz. Electrode imped-
ances were checked and corrected at the beginning of each
recording. The sleep periods were visually scored and classified
in 30-s epochs into sleep stages according to the criteria of
Rechtschaffen and Kales (1968).
EOG ⁄EEG-defined sleepiness
The daytime EEG and EOG data were recorded from the same
locations and at the same sampling rate as in the night
measurements. The EEG and EOG recordings during the
multitask sessions were scored into the following four catego-
ries in 20-s epochs: (1) wakefulness; (2) drowsiness indicated by
slow eye movements accompanied by theta activity of < 5 s
period in EEG; (3) microsleep indicated by theta activity for
5–10 s in EEG; and (4) stage 1 sleep indicated by theta activity
for at least a 10-s period in EEG (Sallinen et al., 2004, 2008).
All categories but category 1 were defined as increased
sleepiness during multitasking. The data of EOG ⁄EEG-
defined sleepiness and multitask performance from each
Figure 1. Study experimental design, showing nightly time in bed
across days: adaptation (A), baseline (BL), sleep restriction days (SR1–
SR5) and recovery (R1–R2), and measurements of psychomotor vig-
ilance task performance (PVT) and multitasking in the 4-h SR group.
Karolinska Sleepiness Scale (KSS) ratings were collected at the
beginning and end of each task session. The control group had a
similar schedule, with the exception that they had the opportunity to
sleep 8 h per night throughout the study. One of the two forenoon
sessions contained a 10-min break, and the data collected during this
session have been excluded from this study.
446 M.-L. Haavisto et al.
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

50-min task session were divided into five 10-min segments for
illustrations and statistical analyses.
Performance measures
Multitasking
A computerised multitask entitled Brain@Work (see Fig. 2)
consisting of four subtasks was developed at the Finnish
Institute of Occupational Health. It represents a modified
version of a multitask entitled SYNWORK that has been
previously used in sleep deprivation studies and found to be
sensitive (Caldwell and Caldwell, 1997; Caldwell and Ram-
spott, 1998; Elsmore, 1994). The four subtasks of the multitask
are introduced at scheduled intervals, and the participant must
be able to choose the optimal moment for the performance of
each subtask. The Brain@Work has been employed in our
recent acute sleep deprivation study in a similar manner as in
this study (Sallinen et al., 2008).
In the short-term memory subtask, at the beginning of each
task session for a period of 10 s the participants were shown a
string of target letters that they were instructed to learn.
During the test sessions, the participants were presented with
probe letters one at a time at 7-s intervals. The participants�task was to classify each probe letter as either a target or a
non-target by clicking on the icons �Yes� or �No�.In the arithmetic subtask, the instruction was to indicate the
sum of two numbers by clicking on digits on a digit pad on the
computer screen. The duration of each trial was 7 s. In short-
term memory and arithmetic subtasks, the participants
obtained 10 points for each correct response, )10 points for
each false response, and )20 points for not responding before
stimulus offset.
In the visual monitoring subtask, participants were
instructed to return a moving dot to the centre of the
innermost circle by clicking on the �Reset� icon. The number
of points obtained (2, 4, 6 or 10) was greater the nearer the dot
was to the outermost edge upon responding. If the participant
did not react before the dot reached the outmost circle, which
took 10 s, 10 points were deducted per each elapsed second. In
the auditory monitoring, the participants were instructed to
discriminate between a non-target tone of 1000 Hz (80%
probability) and a target tone of 1200 Hz (20% probability).
The tones (intensity 62 dB SPL, duration 50 ms) were
presented at 1.5-s intervals. The participants were instructed
to press �Enter� whenever they heard the target tone. The
points were awarded and subtracted identically for the
memory and arithmetic subtasks.
Less than 2 weeks before the experiment, each participant
practised the multitask for 30 min. The level of difficulty in the
task was defined individually for the experiment (see below).
This procedure made it possible to set task difficulty equally
according to each participant�s capacity. The participants
performed the multitask for an average duration of 75 min
(SD = 23.3 min) in the SR group and for 74 min (SD = 55.5
min) on average in the control group during the performance
level adjustment procedure. In all, the participants practised
the multitask for an average of 105 min prior to the experi-
ment to flatten the practice effect. Moreover, on the first day of
the experiment (Day A in Fig. 1), the participants practised the
multitask for 140 min.
To achieve comparable task difficulty between individuals,
difficulty level was determined in two phases. First, the
difficulty levels of the short-term memory and mental arith-
metic subtasks were defined. With this procedure, the effects of
individual differences on the performance of individual tasks
could be controlled for (Baddeley et al., 1997). In the short-
term memory subtask, the number of the target letters was set
two letters shorter than the smallest number of letters that the
participant failed twice to repeat. In the arithmetic subtask, the
number of digits was one less than the smallest number of
digits found too difficult for the participant in the additions
task (<80% of the additions correct). Second, the temporal
intensity of the arithmetic and short-term memory subtasks
was adjusted by manipulating the inter-stimulus interval (ISI,
i.e. the time between two successive items) in 5-min multitask-
ing sessions. In the first 5-min session, the ISI was 6.5 s, and it
was shortened by 0.5 s per session all the way down to 0.5 s if
needed. The adjustment of temporal intensity was finished
when the participant failed to obtain at least 70% of the total
score twice.
Vigilance
A 10-min PVT was used to evaluate behavioural alertness
(Dinges and Powell, 1985). During this task, the participants
were instructed to attend to the timer presented on the
computer screen and press the response button as quickly as
possible whenever the timer started running. The ISI varied
from 2000 to 10 000 ms. The mean number of lapses [reaction
times (RTs) longer than 500 ms] and the slowest 10% of all
responses (mean 1 ⁄RT · 1000 from slowest 10% RTs per
trial) were used as dependent measures.Figure 2. A computer screen showing the four subtasks of the
Brain@Work multitask.
Sleep restriction and multitasking 447
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454
Subjective measures
Subjective sleepiness was measured using the nine-point KSS
(Akerstedt and Gillberg, 1990). The scale varies from very alert
(1) to very sleepy ⁄fighting sleep ⁄ effort to keep awake (9).
Statistical analysis
Statistical analyses were performed using linear mixed-model
anovas, as this technique is suited for the analysis of individual
differences over time (Bliese et al., 2006; Van Dongen et al.,
2003). Traditional repeated-measures anova are inappropriate
for distinguishing stable changes from error variance across
measurement points. Moreover, repeated-measures anovas
assume equal variances within each group. Our mixed-models
included group (SR, control) and day (BL, SR1, SR2, SR3,
SR4, SR5, R1, R2) as fixed effects, and time-on-task (1st, 2nd,
3rd, 4th, 5th 10-min multitask interval) as a random effect.
Separate analyses were performed to test the effects of the SR
days (days BL, SR1, SR2, SR3, SR4 and SR5) and the
recovery days (days BL, R1, R2) on the dependent variables.
The time-of-day factor was left out of the models, as
preliminary analysis showed no difference between the fore-
noon and the afternoon sessions. A random effect on the
intercept and random slopes for centred (to the value of 3rd
10-min segment in each 50-min task session) multitask score or
percentage of EEG ⁄EOG-defined sleepiness was included in
the models to account for individual differences in the
dependent variables. For intraclass correlation (ICC) calcula-
tions, we used only random intercept models without random
slope of the time-on-task factor. In analysis we extended the
compound symmetry correlation structure for the repeated
observations using the linear covariance structure parameter
(PARM) of the SAS ⁄MIXED-procedure to account for the
different correlations during measurements in the same day
and between the different days. The data were analysed using
PROC MIXED in SAS 9.1 (Sas Institute Inc., 2004).
An ICC for multitask performance was computed from the
estimated between-subjects variance (systematic interindividu-
al variance) and the within-subjects variance (residual intra-
individual variance) separately for the experimental and the
control group. These variance components analyses were
performed by mixed-model anova, with day as a fixed linear
effect (five SR days) and participants as a random effect
(random intercept model). Confidence limits for ICC were
calculated with SAS ⁄NLMIXED procedure by the estimate
statement where the limits are based on the t-distribution
rather than on the standard normal distribution.
RESULTS
Sleep length in the experiment
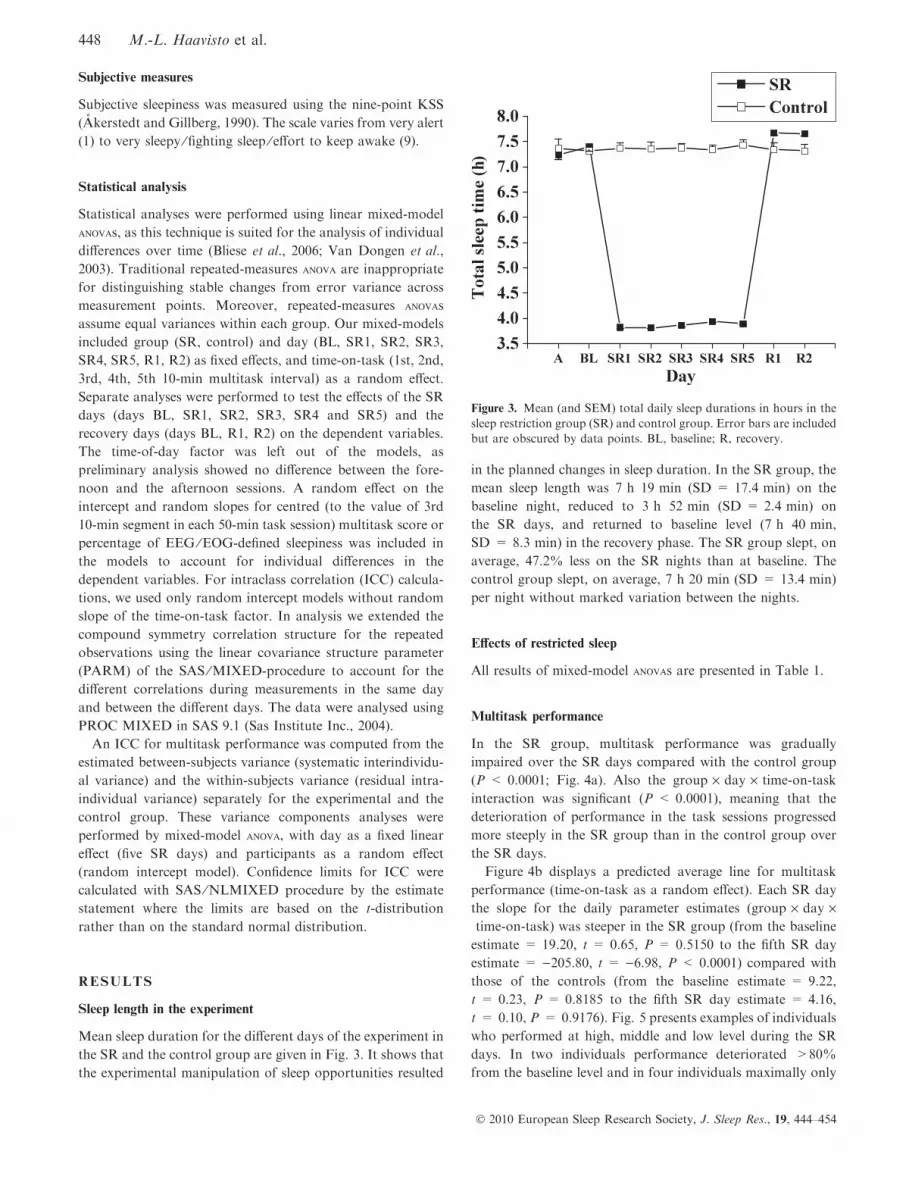
Mean sleep duration for the different days of the experiment in
the SR and the control group are given in Fig. 3. It shows that
the experimental manipulation of sleep opportunities resulted
in the planned changes in sleep duration. In the SR group, the
mean sleep length was 7 h 19 min (SD = 17.4 min) on the
baseline night, reduced to 3 h 52 min (SD = 2.4 min) on
the SR days, and returned to baseline level (7 h 40 min,
SD = 8.3 min) in the recovery phase. The SR group slept, on
average, 47.2% less on the SR nights than at baseline. The
control group slept, on average, 7 h 20 min (SD = 13.4 min)
per night without marked variation between the nights.
Effects of restricted sleep
All results of mixed-model anovas are presented in Table 1.
Multitask performance
In the SR group, multitask performance was gradually
impaired over the SR days compared with the control group
(P < 0.0001; Fig. 4a). Also the group · day · time-on-task
interaction was significant (P < 0.0001), meaning that the
deterioration of performance in the task sessions progressed
more steeply in the SR group than in the control group over
the SR days.
Figure 4b displays a predicted average line for multitask
performance (time-on-task as a random effect). Each SR day
the slope for the daily parameter estimates (group · day ·time-on-task) was steeper in the SR group (from the baseline
estimate = 19.20, t = 0.65, P = 0.5150 to the fifth SR day
estimate = )205.80, t = )6.98, P < 0.0001) compared with
those of the controls (from the baseline estimate = 9.22,
t = 0.23, P = 0.8185 to the fifth SR day estimate = 4.16,
t = 0.10, P = 0.9176). Fig. 5 presents examples of individuals
who performed at high, middle and low level during the SR
days. In two individuals performance deteriorated >80%
from the baseline level and in four individuals maximally only
Figure 3. Mean (and SEM) total daily sleep durations in hours in the
sleep restriction group (SR) and control group. Error bars are included
but are obscured by data points. BL, baseline; R, recovery.
448 M.-L. Haavisto et al.
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454
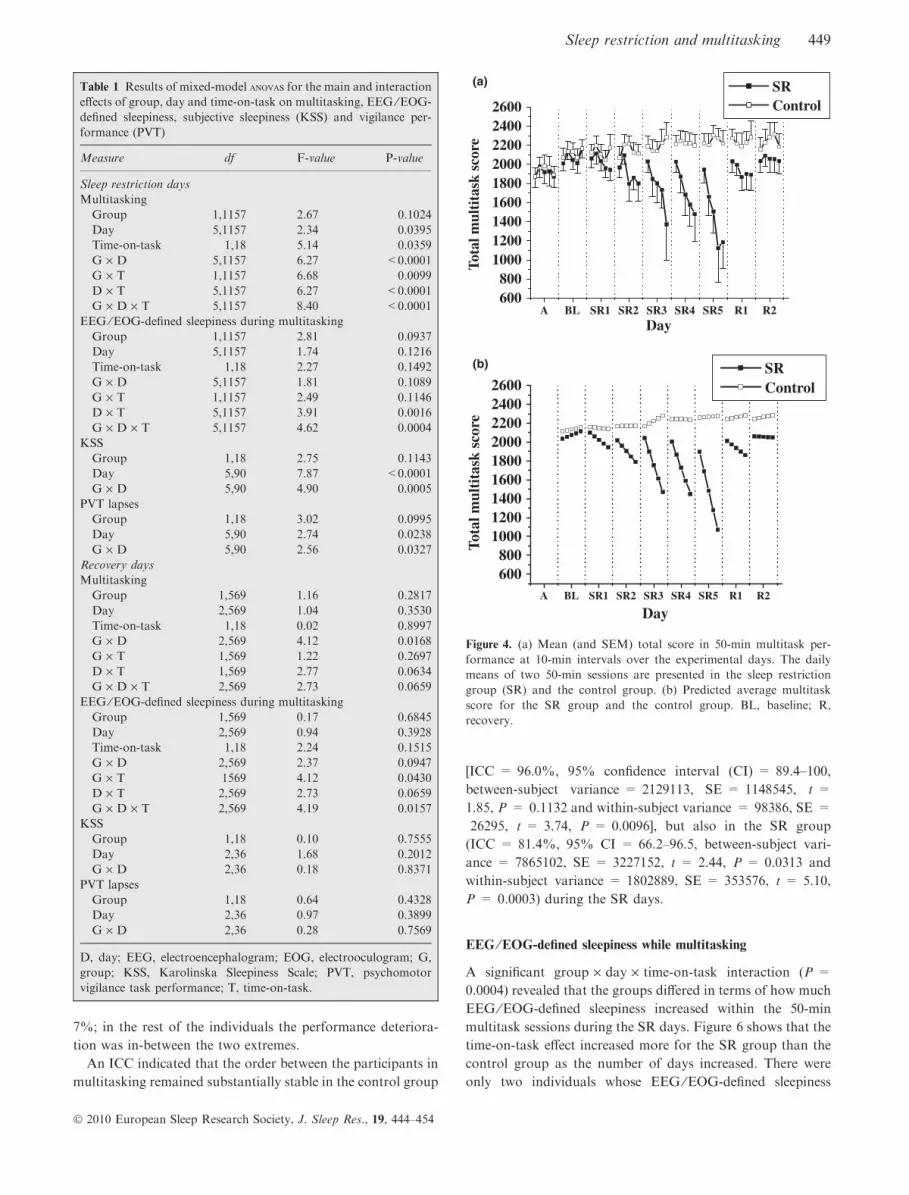
7%; in the rest of the individuals the performance deteriora-
tion was in-between the two extremes.
An ICC indicated that the order between the participants in
multitasking remained substantially stable in the control group
[ICC = 96.0%, 95% confidence interval (CI) = 89.4–100,
between-subject variance = 2129113, SE = 1148545, t =
1.85, P = 0.1132 and within-subject variance = 98386, SE =
26295, t = 3.74, P = 0.0096], but also in the SR group
(ICC = 81.4%, 95% CI = 66.2–96.5, between-subject vari-
ance = 7865102, SE = 3227152, t = 2.44, P = 0.0313 and
within-subject variance = 1802889, SE = 353576, t = 5.10,
P = 0.0003) during the SR days.
EEG ⁄EOG-defined sleepiness while multitasking
A significant group · day · time-on-task interaction (P =
0.0004) revealed that the groups differed in terms of how much
EEG ⁄EOG-defined sleepiness increased within the 50-min
multitask sessions during the SR days. Figure 6 shows that the
time-on-task effect increased more for the SR group than the
control group as the number of days increased. There were
only two individuals whose EEG ⁄EOG-defined sleepiness
Table 1 Results of mixed-model anovas for the main and interaction
effects of group, day and time-on-task on multitasking, EEG ⁄EOG-
defined sleepiness, subjective sleepiness (KSS) and vigilance per-
formance (PVT)
Measure df F-value P-value
Sleep restriction days
Multitasking
Group 1,1157 2.67 0.1024
Day 5,1157 2.34 0.0395
Time-on-task 1,18 5.14 0.0359
G · D 5,1157 6.27 <0.0001
G · T 1,1157 6.68 0.0099
D · T 5,1157 6.27 <0.0001
G · D · T 5,1157 8.40 <0.0001
EEG ⁄EOG-defined sleepiness during multitasking
Group 1,1157 2.81 0.0937
Day 5,1157 1.74 0.1216
Time-on-task 1,18 2.27 0.1492
G · D 5,1157 1.81 0.1089
G · T 1,1157 2.49 0.1146
D · T 5,1157 3.91 0.0016
G · D · T 5,1157 4.62 0.0004
KSS
Group 1,18 2.75 0.1143
Day 5,90 7.87 <0.0001
G · D 5,90 4.90 0.0005
PVT lapses
Group 1,18 3.02 0.0995
Day 5,90 2.74 0.0238
G · D 5,90 2.56 0.0327
Recovery days
Multitasking
Group 1,569 1.16 0.2817
Day 2,569 1.04 0.3530
Time-on-task 1,18 0.02 0.8997
G · D 2,569 4.12 0.0168
G · T 1,569 1.22 0.2697
D · T 1,569 2.77 0.0634
G · D · T 2,569 2.73 0.0659
EEG ⁄EOG-defined sleepiness during multitasking
Group 1,569 0.17 0.6845
Day 2,569 0.94 0.3928
Time-on-task 1,18 2.24 0.1515
G · D 2,569 2.37 0.0947
G · T 1569 4.12 0.0430
D · T 2,569 2.73 0.0659
G · D · T 2,569 4.19 0.0157
KSS
Group 1,18 0.10 0.7555
Day 2,36 1.68 0.2012
G · D 2,36 0.18 0.8371
PVT lapses
Group 1,18 0.64 0.4328
Day 2,36 0.97 0.3899
G · D 2,36 0.28 0.7569
D, day; EEG, electroencephalogram; EOG, electrooculogram; G,
group; KSS, Karolinska Sleepiness Scale; PVT, psychomotor
vigilance task performance; T, time-on-task.
A BL SR1 SR2 SR3 SR4 SR5 R1 R2600800
100012001400160018002000220024002600
SR Control
Tota
l mul
tita
sk s
core
Day
A BL SR1 SR2 SR3 SR4 SR5 R1 R2
600800
100012001400160018002000220024002600
Tota
l mul
tita
sk s
core
Day
SR Control
(a)
(b)
Figure 4. (a) Mean (and SEM) total score in 50-min multitask per-
formance at 10-min intervals over the experimental days. The daily
means of two 50-min sessions are presented in the sleep restriction
group (SR) and the control group. (b) Predicted average multitask
score for the SR group and the control group. BL, baseline; R,
recovery.
Sleep restriction and multitasking 449
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454
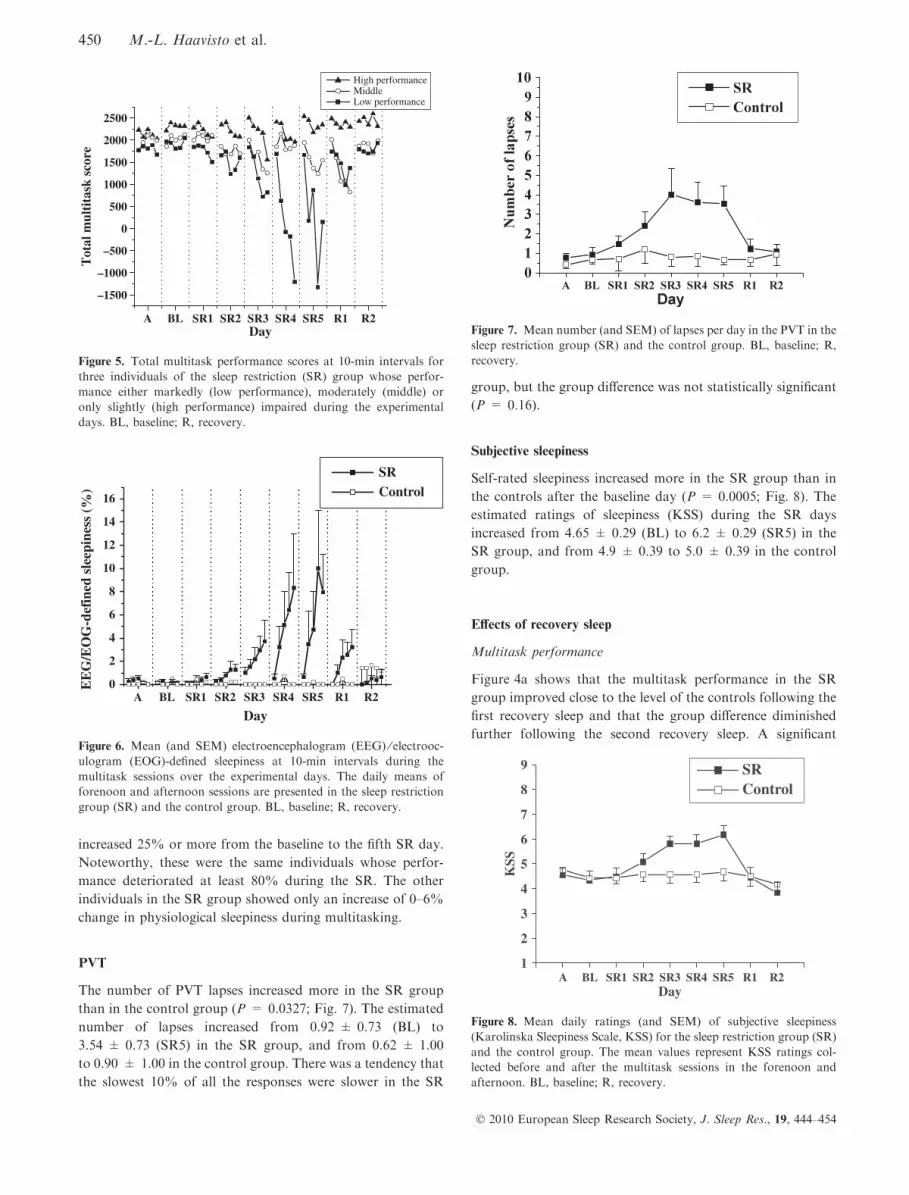
increased 25% or more from the baseline to the fifth SR day.
Noteworthy, these were the same individuals whose perfor-
mance deteriorated at least 80% during the SR. The other
individuals in the SR group showed only an increase of 0–6%
change in physiological sleepiness during multitasking.
PVT
The number of PVT lapses increased more in the SR group
than in the control group (P = 0.0327; Fig. 7). The estimated
number of lapses increased from 0.92 ± 0.73 (BL) to
3.54 ± 0.73 (SR5) in the SR group, and from 0.62 ± 1.00
to 0.90 ± 1.00 in the control group. There was a tendency that
the slowest 10% of all the responses were slower in the SR
group, but the group difference was not statistically significant
(P = 0.16).
Subjective sleepiness
Self-rated sleepiness increased more in the SR group than in
the controls after the baseline day (P = 0.0005; Fig. 8). The
estimated ratings of sleepiness (KSS) during the SR days
increased from 4.65 ± 0.29 (BL) to 6.2 ± 0.29 (SR5) in the
SR group, and from 4.9 ± 0.39 to 5.0 ± 0.39 in the control
group.
Effects of recovery sleep
Multitask performance
Figure 4a shows that the multitask performance in the SR
group improved close to the level of the controls following the
first recovery sleep and that the group difference diminished
further following the second recovery sleep. A significant
A BL SR1 SR2 SR3 SR4 SR5 R1 R20
2
4
6
8
10
12
14
16
EE
G/E
OG
-def
ined
sle
epin
ess
(
)
Day
SR Control
Figure 6. Mean (and SEM) electroencephalogram (EEG) ⁄ electrooc-ulogram (EOG)-defined sleepiness at 10-min intervals during the
multitask sessions over the experimental days. The daily means of
forenoon and afternoon sessions are presented in the sleep restriction
group (SR) and the control group. BL, baseline; R, recovery.
A BL SR1 SR2 SR3 SR4 SR5 R1 R2
–1500
–1000
–500
0
500
1000
1500
2000
2500
Tot
al m
ulti
task
sco
re
Day
High performance Middle Low performance
Figure 5. Total multitask performance scores at 10-min intervals for
three individuals of the sleep restriction (SR) group whose perfor-
mance either markedly (low performance), moderately (middle) or
only slightly (high performance) impaired during the experimental
days. BL, baseline; R, recovery.
Figure 7. Mean number (and SEM) of lapses per day in the PVT in the
sleep restriction group (SR) and the control group. BL, baseline; R,
recovery.
A BL SR1 SR2 SR3 SR4 SR5 R1 R21
2
3
4
5
6
7
8
9 SRControl
KSS
Day
Figure 8. Mean daily ratings (and SEM) of subjective sleepiness
(Karolinska Sleepiness Scale, KSS) for the sleep restriction group (SR)
and the control group. The mean values represent KSS ratings col-
lected before and after the multitask sessions in the forenoon and
afternoon. BL, baseline; R, recovery.
450 M.-L. Haavisto et al.
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

group · day interaction (P = 0.0168) indicated that the
recovery process continued still during the second night of
recovery. The daily parameter estimates for time-on-task lines
decreased from R1 (estimate = )37.63, t = )3.09, P =
0.0021) to R2 (estimate = )3.61, t = )0.30, P = 0.7673) in
the SR group, meaning that the time-on-task effect almost
disappeared during R2. The control group showed no com-
parable changes.
EEG ⁄EOG-defined sleepiness while multitasking
A significant group · time-on-task · day interaction
(P = 0.0157) demonstrated that the difference between the
groups in physiological sleepiness was dependent on both the
time spent on the task and the number of recovery days.
Figure 6 shows that the group difference in the time-on-task
effect decreased from R1 to R2.
PVT
The groups did not differ in the number of PVT lapses during
the recovery period.
Subjective sleepiness
The sleepiness ratings on the recovery days showed no group
differences.
DISCUSSION
The main findings of this study were that at a group level
multitask performance was significantly impaired by 5 days of
partial SR, and that this impairment increased as a function of
the time spent on the task. However, within the group exposed
to the SR, only few individuals showed large impairments in
their performance. Most of the sleep-deprived individuals
showed only moderate deteriorations and some individuals�performance remained virtually unchanged. In addition,
EEG ⁄EOG-defined sleepiness increased significantly at the
group level during multitasking in the course of the SR but,
actually, there were only few individuals whose sleepiness
increased markedly.
The finding that multitask performance progressively dete-
riorated with the increasing number of SR days is in line with
previous studies in which participants� performance on less
complicated and shorter tasks deteriorated in the course of SR
(Belenky et al., 2003; Dinges et al., 1997; Van Dongen et al.,
2003). From the viewpoint of work life, the strength of this
study was that the used task included two important charac-
teristics of real operational tasks that are often performed
under SR, namely high demands on cognitive processes such as
divided attention (Gopher, 1996; Wickens et al., 2003) and the
requirement of performance over long periods. In practice, our
finding can be understood that a high number of sleep-limiting
shifts in a row substantially increase the risk for human error
in operational tasks that require multitasking. However,
studies conducted in authentic work conditions are needed to
verify this conclusion, as, for example, expertise based on long
experience and awareness of the consequences of a perfor-
mance error probably also play a role in how well a person
actually performs at work while restricted of sleep.
The progressively augmenting time-on-task effect on multi-
task performance was observed in the SR but not in the
control group, indicating that it was totally dependent on the
sleep loss preceding the performance. The time-on-task effect
on multitasking or simulator performance has been also found
in previous studies on acute sleep loss (Akerstedt et al., 2005;
Caldwell and Ramspott, 1998; Sallinen et al., 2008). The
practical significance of this finding is high. In safety-critical
occupations, for example, with air-traffic controllers, most task
sessions performed under sleep loss are relatively long in
duration. Interrupting the working period with a break could
theoretically be of advantage. However, our previous work has
shown that it is unlikely that, for example, a rest pause with
light neck-and-shoulder exercise would be an effective remedy
(Sallinen et al., 2008). Cognitive performance during SR is, to
some extent, improved by napping (Mollicone et al., 2008;
Purnell et al., 2002; Sallinen et al., 1998), and thus naps during
breaks could be of advantage. Another strategy is to use
stimulants. However, continuous use often leads to tolerance
and increased dosages, which may affect sleep following the
shift.
The time-on-task effect on multitasking intensified in the
course of the SR. Previous studies have shown that 20 h of
wakefulness per day – the same amount that was used in this
study – is sufficient for escalation of performance impairments
(McCauley et al., 2009). Our new finding was that most of the
deterioration occurred during the latter part of the 50-min task
session, implying that the escalating negative effects of
extended wakefulness on multitask performance are dependent
on the time spent on the task.
There are various brain mechanisms that may explain the
observed deterioration in multitasking. The increases in
EEG ⁄EOG-defined sleepiness during the deterioration of
multitasking suggest that at least the arousal mechanisms
played a key role. The level of thalamic activation that
regulates arousal and attention has been found to decrease in
association with sleep deprivation (Chee et al., 2006, 2008;
Coull et al., 1998; Thomas et al., 2000). This deactivation
could at least to some extent explain the observed decrements
in multitask performance under SR. On the contrary, multi-
tasking places special demands on cognitive processes required
in subtasks and coordinating attention switching between the
subtasks (D�esposito et al., 1995; Dux et al., 2006; Just et al.,
2001, 2008). SR-induced changes in brain mechanisms under-
lying these cognitive processes may thus also explain the
observed deterioration in multitasking.
Large individual differences in multitasking were observed
on the SR days. Importantly, the individual differences were
substantially stable within this period: the same individuals
showed either sensitivity or tolerance to the curtailment of
sleep. Only two individuals out of 13 exhibited severe
Sleep restriction and multitasking 451
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

decrements in multitask performance and, interestingly, the
same individuals also showed the most severe increase in
EEG ⁄EOG-defined sleepiness during the task performance.
Our finding of large individual differences in multitasking
within a single SR period is in line with a study by Van Dongen
et al. (2006) in pilots. In their study, differences in flight-
simulator performance between sleep-deprived pilots were
large and stable in the course of a single experiment (Van
Dongen et al., 2006).
The individual differences observed in this study cannot be
explained by differences in cognitive aptitude. Prior to the
experiment, the cognitive demand of the task was adjusted
individually to be 70% of the maximal capacity of each
participant. The individual adjustment was carried out because
previous studies have shown that the ability to divide attention
between one or more simultaneous tasks differs greatly from
one person to another (Damos, 1993). The adjustment
protocol ensured that all individuals were able to perform
the multitask and that the starting level for each individual was
equally demanding. The observed differences suggest that there
are individual differences in tolerance for SR (Leproult et al.,
2003; Van Dongen et al., 2003, 2004). In our study, individual
differences remained stabile through the SR days, but we did
not examine whether the differences would have been replica-
ble in a repeated exposure to SR.
It is not self-evident how and to what extent the individual
differences in response to sleep loss should be accounted for in
shift work: whether they should be used as a selection criterion
when recruiting new personnel or whether they should be used
as a basis for adjusting work demands, including working
hours, on an individual basis. The seriousness of the safety
hazard associated with the task in question is one aspect to be
considered in this context. Second, it would be important to
establish the possibilities for obtaining reliable data on a
person�s sensitivity to sleep deprivation before starting shift
work. For the moment, it seems that the only reliable way of
indentifying persons with high vulnerability to sleep loss is to
subject them to such conditions, as no potential baseline
predictor has turned out to be reliable enough for this purpose
(Van Dongen and Belenky, 2009). In addition, a Bayesian
forecasting technique based on closed-loop feedback of mea-
sured performance can be used for predicting changes in a
sleep-deprived worker�s job performance (Van Dongen and
Belenky, 2009).
Electroencephalogram ⁄ electrooculogram-defined sleepiness
during multitasking responded to SR similarly to multitask
performance itself: sleepiness increased as a function of the
time spent on the task and the number of SR days, but actually
only two individuals out of 13 showed a marked increase.
Previous studies have found no clear associations between
changes in cognitive performance and concomitant physiolog-
ical sleepiness under sleep loss (Galliaud et al., 2008; Stenuit
and Kerkhofs, 2008; Wilson et al., 2007). This may be partly
due to the different durations of tasks used in the previous
studies: in this study, the task duration was much longer than
those used earlier. It can be assumed that the association is
more obvious when the effect of sleep loss on both measures
has augmented close to its maximum at the end of the task
performance. In all, our findings suggest that increased
physiological sleepiness is at least one of the factors underlying
impaired long-duration multitasking under cumulative sleep
loss.
Both standard measures of sleepiness, the KSS ratings and
number of PVT lapses, were affected by the SR. On the fifth
SR day, the mean level of the KSS ratings was close to the level
(‡7) that is known to be associated with electrophysiological
signs of extreme sleepiness and impaired driving performance
(Akerstedt and Gillberg, 1990; Ingre et al., 2006). A somewhat
surprising result was that the slowest 10% of the PVT response
times were not significantly affected by the SR. A reason for
this result may be that the PVT was always presented
immediately after a long-duration multitask session. This
protocol may have affected the level of arousal at which the
PVT task was initiated.
Recovery of long-duration multitasking from the cumulative
SR proceeded gradually. Following the first recovery sleep
period, the level of performance clearly improved as compared
with the last SR day, but still remained below that of the
control group. Performance returned to the baseline level after
the second recovery sleep period. In the course of the gradual
recovery process, the time-on-task effect and individual differ-
ences decreased. In a previous study, full recovery from a
7-day SR was not reached after three recovery nights (Belenky
et al., 2003). Interestingly, extension of sleep duration for
several days previous to SR improved the rate of performance
recovery (Rupp et al., 2009). The relationship between the
severity of the preceding SR (accumulated sleep loss) and
the pace of recovery process warrants further research. The
question is of practical significance when planning shift work
schedules: how many recovery days must be included in the
schedule after a certain number of sleep-limiting shifts?
The recovery process of EEG ⁄EOG-defined sleepiness
during multitasking resembled that of multitask performance:
the recovery process continued over the two recovery sleep
opportunities and was characterised by decreases in the time-
on-task effect and in individual differences. This finding is in
line with our recent study, where both long-duration multitask
performance and EEG ⁄EOG-defined sleepiness responded
similarly to an 8-h sleep opportunity after only 2 h of sleep
on the previous night (Sallinen et al., 2008). When considering
time, needed for recovery, it is important to notice that the
participants of this study were provided with optimal sleeping
conditions free from many sleep-disturbing factors normally
present in everyday life. Thus, it is possible that recovery takes
even longer under real working conditions than in our
laboratory environment.
There are several limitations in our study that should be
taken into account to interpret the results. First, our sample
consisted of only young healthy men, which limits the
possibilities to generalise the results to other age groups,
women, persons with health problems, and experienced shift
workers. Second, the long-term stability of the observed
452 M.-L. Haavisto et al.
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

individual differences in response to SR remains open, as the
participants were exposed to SR only once. Third, the recovery
process from 5 days of SR was followed only for 2 days.
However, shift schedules often contain repetitive spells of
sleep-limiting shifts with only a day or two off between them.
In this context, the question of how many recovery days is
needed to prevent any carry-over effect from a period of SR to
the next one is of importance, but cannot be answered on the
basis of our results. Finally, the laboratory conditions of our
study limit the possibilities to generalise the results to everyday
life and thus field studies are needed to verify our findings.
CONCLUSIONS
In conclusion, this study demonstrates that complex and long-
duration cognitive performance gradually degrades in young
healthy men when their sleep is restricted to 4 h per night for
5 days. This degradation is characterised by a strong time-on-
task effect and large individual differences, and accompanied
with an increase in physiological sleepiness. Recovery from
restricting sleep to 4 h per night for a period of a work week
takes at least two 8-h sleep opportunities.
ACKNOWLEDGEMENTS
This study was supported by the European co-funded, 6th
FW, Integrated project SENSATION (IST, 507231), Finnish
Work Environmental Found and National Technology
Agency of Finland. We would like to thank Outi Fischer,
Hannele Huhta, Seija Karas, Hannele Kataja, Nina
Lapvetelainen, Mari Marjamaki, Johanna Parikka, Teppo
Valtonen, Riitta Velin and students from the medical faculty
of Helsinki University for invaluable assistance in recruiting
and screening of the participants, and running of the
experiments. We would also like to thank Jaana Hiltunen,
Kati Hirvonen, Anu Holm and Mika Letonsaari for technical
help in this study. In addition, we received generous help and
important comments from Risto Nasanen, PhD, and Ritva
Akila, neuropsychologist.
REFERENCES
Akerstedt, T. and Gillberg, M. Subjective and objective sleepiness in
the active individual. Int. J. Neurosci., 1990, 52: 29–37.
Akerstedt, T., Peters, B., Anund, A. and Kecklund, G. Impaired
alertness and performance driving home from the night shift: a
driving simulator study. J. Sleep Res., 2005, 14: 17–20.
Axelsson, J., Kecklund, G., Akerstedt, T., Donofrio, P., Lekander, M.
and Ingre, M. Sleepiness and performance in response to repeated
sleep restriction and subsequent recovery during semi-laboratory
conditions. Chronobiol. Int., 2008, 25: 297–308.
Baddeley, A., Della Sala, S., Papagano, C. and Spinnler, H. Dual-task
performance in dysexecutive and nondysexecutive patients with a
frontal lesion. Neuropsychology, 1997, 11: 187–194.
Balkin, T. J., Bliese, P. D., Belenky, G. et al. Comparative utility of
instruments for monitoring sleepiness-related performance decre-
ments in the operational environment. J. Sleep Res., 2004, 13: 219–
227.
Banks, S., Van Dongen, H. P. A. and Dinges, D. Response to sleep
restriction depends upon pre-existing sleep dept. Sleep, 2007, 30
(Abstract Supplement), 0346.
Belenky, G., Wesensten, N. J., Thorne, D. R. et al. Patterns of
performance degradation and restoration during sleep restriction
and subsequent recovery: a sleep dose-response study. J. Sleep Res.,
2003, 12: 1–12.
Bliese, P. D., Wesensten, N. J. and Balkin, T. J. Age and individual
variability in performance during sleep restriction. J. Sleep Res.,
2006, 15: 376–385.
Caldwell, J. A. Fatigue in aviation. Travel Med. Infect. Dis., 2005, 3:
85–96.
Caldwell, J. L. and Caldwell, J. A. Recovery sleep and performance
following sleep deprivation with dextroamphetamine. J. Sleep Res.,
1997, 6: 92–101.
Caldwell, J. A. and Caldwell, J. L. Comparison of the effects of
zolpidem-induced prophylactic naps to placebo naps and forced rest
periods in prolonged work schedules. Sleep, 1998, 21: 79–90.
Caldwell, J. A. and Ramspott, S. R. Effects of task duration on
sensitivity to sleep deprivation using the multiattribute task battery.
Behav. Res. Meth. Instrum. Comput., 1998, 30: 651–660.
Chee, M. W. L., Chuah, L. Y. M., Venkatraman, V., Chan, W. Y.,
Philip, P. and Dinges, D. F. Functional imaging of working memory
following normal sleep and after 24 and 35 h of sleep deprivation:
correlations of fronto-parietal activation with performance. Neuro-
image, 2006, 31: 419–428.
Chee, M. W. L., Tan, J. C., Zheng, H. et al. Lapsing during sleep
deprivation is associated with distributed changes in brain activa-
tion. J. Neurosci., 2008, 28: 5519–5528.
Coull, J. T., Frackowiak, R. S. J. and Frith, C. D. Monitoring for
target objects: activation of right frontal and parietal cortices with
increasing time on task. Neuropsychologia, 1998, 36: 1325–1334.
Damos, D. Using meta-analysis to compare the predictive validity of
single- and multiple-task measures to flight performance. Hum.
Factors, 1993, 35: 615–628.
D�esposito, M., Detre, J. A., Alsop, D. C., Shin, R. K., Atlas, S. and
Grossman, M. The neural basis of the central executive system of
working memory. Nature, 1995, 378: 279–281.
Dinges, D. F. and Kribbs, N. B. Performing while sleepy: effects of
experimentally-induced sleepiness. In: T. H. Monk ((Ed.)) Sleep,
Sleepiness and Performance. John Wiley, New York, 1991: 97–128.
Dinges, D. F. and Powell, J. W. Microcomputer analyses of perfor-
mance on a portable, simple visual RT task during sustained
operations. Behav. Res. Meth. Instrum. Comput., 1985, 17: 652–655.
Dinges, D. F., Pack, F., Williams, K. et al. Cumulative sleepiness,
mood disturbance, and psychomotor performance decrements
during a week of sleep restricted to 4–5 hours per night. Sleep,
1997, 20: 267–277.
Dux, P. E., Ivanoff, J., Asplund, C. L. and Marois, R. Isolation of
central bottleneck of information processing with time-solved fMRI.
Neuron, 2006, 52: 1109–1120.
Elsmore, T. SYNWORK 1: a PC-based tool for assessment of
performance in a simulated work environment. Behav. Res. Meth.
Instrum. Comput., 1994, 26: 421–426.
Folkard, S. and Lombardi, D. A. Modeling the impact of the
components of long work hours on injuries and �accidents�. Am. J.
Ind. Med., 2006, 49: 953–963.
Galliaud, E., Taillard, J., Sagaspe, P., Valtat, C., Biolac, B. and Philip,
P. Sharp and sleepy: evidence for dissociation between sleep pressure
and nocturnal performance. J. Sleep Res., 2008, 17: 11–15.
Gopher, D. Attention control: explorations of the work of an executive
controller. Cogn. Brain Res., 1996, 5: 23–38.
Harma, M., Sallinen, M., Ranta, R., Mutanen, P. and Muller, K. The
effect of an irregular shift system on sleepiness at work in train
drivers and railway traffic controllers. J. Sleep Res., 2002, 11: 141–
151.
Sleep restriction and multitasking 453
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454

Ingre, M., Akerstedt, T., Peters, B., Anund, A. and Kecklund, G.
Subjective sleepiness, simulated driving performance and blink
duration: examining individual differences. J. Sleep Res., 2006, 15:
47–53.
Just, M. A., Carpenter, P. A., Keller, T. A., Emery, L., Zajac, H. and
Thulborn, K. R. Interdependence of nonoverlapping cortical
systems in dual cognitive tasks. Neuroimage, 2001, 14: 417–426.
Just, M. A., Keller, T. A. and Cynkar, J. A decrease in brain activation
associated with driving when listening to someone speak Brain Res.,
2008, 1205: 70–80.
Lamond, N., Jay, S. M., Dorrian, J., Ferguson, S. A., Jones, C. and
Dawson, D. The dynamics of neurobehavioural recovery following
sleep loss. J. Sleep Res., 2007, 16: 33–41.
Leproult, R., Colecchia, E. F., Berardi, A. M., Stickgold, R., Kosslyn,
S. M. and Van Cauter, E. Individual differences in subjective and
objective alertness during sleep deprivation are stable and unrelated.
Am. J. Physiol. Regul. Integr. Comp. Physiol., 2003, 284: R280–
R290.
McCauley, P., Kalchev, L. V., Smith, A. D., Belenky, G., Dinges, D.
F. and Van Dongen, H. P. A. A new mathematical model for the
homeostatic effects of sleep loss on neurobehavioral performance.
J. Theor. Biol., 2009, 256: 227–239.
Mollicone, D. J., Van Dongen, H. P. A., Rogers, N. J. and Dinges, D.
F. Response surface mapping of neurobehavioral performance: the
feasibility of split sleep schedules for space operations. Acta
Astronaut., 2008, 63: 833–840.
Navon, D. and Gopher, D. On the economy of the human processing
system. Psychol. Rev., 1979, 80: 214–255.
NTSB. Fatigue, alcohol, other drugs, and medical factors in fatal-to-
the-driver heavy truck crashes. National Transportation and Safety
Board, Safety Study, NTST ⁄ SS-90 ⁄ 01, 1990.Partinen, M. and Gislason, T. Basic Nordic Sleep Questionnaire
(BNSQ): a quantitated measure of subjective sleep complaints.
J. Sleep Res., 1995, 4: 150–155.
Philip, P. and Akerstedt, T. Transport and industry safety, how are
they affected by sleepiness and sleep restriction? Sleep Med. Rev.,
2006, 10: 347–356.
Purnell, M. T., Feyer, A.-M. and Herbison, G. P. The impact of a nap
opportunity during the night shift on the performance and alertness
of 12-h shift workers. J. Sleep Res., 2002, 11: 219–227.
Rechtschaffen, A. and Kales, A. A. Manual of Standardized Termi-
nology, Techniques and Scoring System for Sleep Stages of Human
Subjects. UCLA Brain Information Service, 1968.
Rupp, T. L., Wesensten, N. J., Bliese, P. D. and Balkin, T. J. Banking
sleep: realization of benefits during subsequent sleep restriction and
recovery. Sleep, 2009, 32: 311–321.
Sallinen, M., Harma, M., Akerstedt, T., Rosa, R. and Lillqvist, O.
Promoting alertness with a short nap during a night shift. J. Sleep
Res., 1998, 7: 240–247.
Sallinen, M., Harma, M., Mutanen, P., Ranta, R., Virkkala, J. and
Muller, K. Sleep-wake rhythm in an irregular shift system. J. Sleep
Res., 2003, 12: 103–112.
Sallinen, M., Harma, M., Akila, R. et al. The effects of sleep debt and
monotonous work on sleepiness and performance during a 12-h
dayshift. J. Sleep Res., 2004, 13: 285–294.
Sallinen, M., Holm, A., Hiltunen, J. et al. Recovery of cognitive
performance from sleep debt: do a short rest pause and a single
recovery night help? Chronobiol. Int., 2008, 25: 279–296.
Sas Institute Inc. Sas/Stat 9.1. user�s guide. SAS Institute, Gary, NC,
2004.
Stenuit, P. and Kerkhofs, M. Effects of sleep restriction on cognition in
women. Biol. Psychol., 2008, 77: 81–88.
Thomas, M., Sing, H., Belenky, G. et al. Neural basis of alertness and
cognitive performance impairments during sleepiness. I. Effects of
24 h of sleep deprivation on waking human regional brain activity.
J. Sleep Res., 2000, 9: 335–352.
Van Dongen, H. P. A. and Belenky, G. Individual differences in
vulnerability to sleep loss in the work environment. Ind. Health,
2009, 47: 518–526.
Van Dongen, H. P. A, Maislin, G., Mullington, J. M. and Dinges, D.
F. The cumulative cost of additional wakefulness: dose-response
effects on neurobehavioral functions and sleep physiology from
chronic sleep restriction and total sleep deprivation. Sleep, 2003, 26:
117–126.
Van Dongen, H. P. A., Baynard, M. D., Maislin, G. and Dinges, D. F.
Systematic interindividual differences in neurobehavioral impair-
ment from sleep loss: evidence of trait-like differential vulnerability.
Sleep, 2004, 27: 423–433.
Van Dongen, H. P. A., Caldwell, J. A. and Caldwell, J. L.
Investigating systematic individual differences in sleep-deprived
performance on a high-fidelity flight simulator. Behav. Res. Meth.,
2006, 38: 333–343.
Webb, W. B. and Agnew, H. W. The effects of a chronic limitation of
sleep length. Psychophysiology, 1974, 11: 265–274.
Wickens, C. Multiple resources and performance prediction. Theor.
Issues Ergon. Sci., 2002, 3: 159–177.
Wickens, C. D., Goh, J., Helleburg, J., Horrey, W. J. and Talleur, D.
A. Attentional models of multi-task pilot performance using
advanced display technology. Hum. Factors, 2003, 45: 360–380.
Wilson, G. F., Caldwell, J. A. and Russell, C. A. Performance and
psychophysiological measures of fatigue effects on aviation related
tasks of varying difficulty. Int. J. Aviat. Psychol., 2007, 17: 219–247.
454 M.-L. Haavisto et al.
� 2010 European Sleep Research Society, J. Sleep Res., 19, 444–454
Copyright © 2022 FDOKUMEN




















