
Simvastatin has an anti-inflammatory effect on macrophages via upregulation of an atheroprotective...
10
Simvastatin has an anti-inflammatory effect on macrophages via upregulation of an atheroprotective transcription factor, Kruppel-like factor 2 Tiina T. Tuomisto 1† , Henri Lumivuori 1† , Emilia Kansanen 1 , Sanna-Kaisa Ha ¨kkinen 1 , Mikko P. Turunen 1 , Johannes V. van Thienen 2 , Anton J. Horrevoets 2‡ , Anna-Liisa Levonen 1 , and Seppo Yla ¨-Herttuala 1,3,4 * 1 Department of Biotechnology and Molecular Medicine, A. I. Virtanen Institute, University of Kuopio, PO Box 1627, FIN-70211 Kuopio, Finland; 2 Department of Medical Biochemistry, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; 3 Department of Medicine, University of Kuopio, Kuopio, Finland; and 4 Gene Therapy Unit, Kuopio University Hospital, Kuopio, Finland Received 3 May 2007; revised 12 December 2007; accepted 4 January 2008 online publish-ahead-of-print 10 January 2008 Time for primary review: 27 days Aims Statins have beneficial vascular effects beyond their cholesterol-lowering action. Since macro- phages play a central role in atherogenesis, we characterized the effects of simvastatin on gene expression profile of human peripheral blood monocyte (HPBM)-macrophages. Methods and results Gene expression profile was studied using Affymetrix gene chip analysis. Lentiviral gene transfer of Kruppel-like factor 2 (KLF-2) was used to further study its role in macrophages. Simvastatin treatment lead to downregulation of many pro-inflammatory genes including several chemokines [e.g. monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory proteins-1a and b, interleukin-2 receptor-b], members of the tumour necrosis factor family (e.g. lymphotoxin b), vascular cell adhesion molecule-1, and tissue factor (TF). Simvastatin also modulated the expression of several transcription factors essential for inflammation: NF-kB relA/p65 subunit and ets-1 were down- regulated, and an atheroprotective transcription factor KLF-2 was upregulated. The effects of simvastatin on MCP-1 and TF could be mimicked by KLF-2 overexpression using lentiviral gene transfer. Conclusion Simvastatin has a strong anti-inflammatory effect on HPBM cells including upregulation of the atheroprotective factor KLF-2. This may partly explain the beneficial effects of statins on cardiovas- cular diseases. KEYWORDS Atherosclerosis; Gene array analysis; Inflammation; Macrophages; Statins 1. Introduction Inflammation and vessel wall macrophages play important roles in the pathogenesis of atherosclerosis. Lesion macro- phages secrete a number of growth factors, cytokines, and other molecules, such as matrix metalloproteinases (MMPs), which are involved in lesion progression: they acti- vate T-cells, enhance smooth muscle cells (SMC) prolifer- ation, and contribute to endothelial dysfunction, lesion rupture, and blood coagulation. 1 Therefore, to effectively prevent and treat cardiovascular events, treatments target- ing macrophages would be desirable. The HMG-CoA inhibitors, statins, have other beneficial effects on atherogenesis in addition to their lipid-lowering action. These pleiotropic effects include e.g. the upregulation of the production of nitric oxide in endothelial cells (ECs), decreased proliferation of vascular SMCs, inhibition of platelet activation, and increased fibrinolytic activity. 2,3 Importantly, statins have been shown to modulate the inflammatory process in the vessel wall. They reduce both the number and the activity of inflammatory cells within atherosclerotic plaques. 4 Favourable effects include the modulation of cytokine secretion and signalling, 3,5 decreased monocyte-EC adhesion, 6 decreased expression of tissue factor (TF) and MMPs 4 in macrophages, and inhi- bition of oxLDL-induced macrophage proliferation. 7 Our recent finding that human lesion macrophages overexpress HMG-CoA reductase gene may explain why statins effec- tively reduce inflammation in the vessel wall. 8 * Corresponding author. Tel: þ358 17 162075; fax: þ358 17 163751. E-mail address: seppo.ylaherttuala@uku.fi † These authors contributed equally to this work. ‡ Present address: Department of Molecular Cell Biology and Immunology, VU University Medical Center, Amsterdam, The Netherlands. Published on behalf of the European Society of Cardiology. All rights reserved. & The Author 2008. For permissions please email: [email protected] Cardiovascular Research (2008) 78, 175–184 doi:10.1093/cvr/cvn007 by guest on November 20, 2013 http://cardiovascres.oxfordjournals.org/ Downloaded from
Transcript of Simvastatin has an anti-inflammatory effect on macrophages via upregulation of an atheroprotective...

Simvastatin has an anti-inflammatory effect onmacrophages via upregulation of an atheroprotectivetranscription factor, Kruppel-like factor 2
Tiina T. Tuomisto1†, Henri Lumivuori1†, Emilia Kansanen1, Sanna-Kaisa Hakkinen1,Mikko P. Turunen1, Johannes V. van Thienen2, Anton J. Horrevoets2‡, Anna-Liisa Levonen1, andSeppo Yla-Herttuala1,3,4*
1Department of Biotechnology and Molecular Medicine, A. I. Virtanen Institute, University of Kuopio, PO Box 1627, FIN-70211Kuopio, Finland; 2Department of Medical Biochemistry, Academic Medical Center, University of Amsterdam, Amsterdam, TheNetherlands; 3Department of Medicine, University of Kuopio, Kuopio, Finland; and 4Gene Therapy Unit, Kuopio UniversityHospital, Kuopio, Finland
Received 3 May 2007; revised 12 December 2007; accepted 4 January 2008 online publish-ahead-of-print 10 January 2008
Time for primary review: 27 days
Aims Statins have beneficial vascular effects beyond their cholesterol-lowering action. Since macro-phages play a central role in atherogenesis, we characterized the effects of simvastatin on geneexpression profile of human peripheral blood monocyte (HPBM)-macrophages.Methods and results Gene expression profile was studied using Affymetrix gene chip analysis. Lentiviralgene transfer of Kruppel-like factor 2 (KLF-2) was used to further study its role in macrophages.Simvastatin treatment lead to downregulation of many pro-inflammatory genes including severalchemokines [e.g. monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory proteins-1aand b, interleukin-2 receptor-b], members of the tumour necrosis factor family (e.g. lymphotoxin b),vascular cell adhesion molecule-1, and tissue factor (TF). Simvastatin also modulated the expressionof several transcription factors essential for inflammation: NF-kB relA/p65 subunit and ets-1 were down-regulated, and an atheroprotective transcription factor KLF-2 was upregulated. The effects ofsimvastatin on MCP-1 and TF could be mimicked by KLF-2 overexpression using lentiviral gene transfer.Conclusion Simvastatin has a strong anti-inflammatory effect on HPBM cells including upregulation ofthe atheroprotective factor KLF-2. This may partly explain the beneficial effects of statins on cardiovas-cular diseases.
KEYWORDSAtherosclerosis;
Gene array analysis;
Inflammation;
Macrophages;
Statins
1. Introduction
Inflammation and vessel wall macrophages play importantroles in the pathogenesis of atherosclerosis. Lesion macro-phages secrete a number of growth factors, cytokines, andother molecules, such as matrix metalloproteinases(MMPs), which are involved in lesion progression: they acti-vate T-cells, enhance smooth muscle cells (SMC) prolifer-ation, and contribute to endothelial dysfunction, lesionrupture, and blood coagulation.1 Therefore, to effectivelyprevent and treat cardiovascular events, treatments target-ing macrophages would be desirable.
The HMG-CoA inhibitors, statins, have other beneficialeffects on atherogenesis in addition to their lipid-loweringaction. These pleiotropic effects include e.g. the upregulationof the production of nitric oxide in endothelial cells (ECs),decreased proliferation of vascular SMCs, inhibition ofplatelet activation, and increased fibrinolytic activity.2,3
Importantly, statins have been shown to modulate theinflammatory process in the vessel wall. They reduce boththe number and the activity of inflammatory cellswithin atherosclerotic plaques.4 Favourable effects includethe modulation of cytokine secretion and signalling,3,5
decreased monocyte-EC adhesion,6 decreased expressionof tissue factor (TF) and MMPs4 in macrophages, and inhi-bition of oxLDL-induced macrophage proliferation.7 Ourrecent finding that human lesion macrophages overexpressHMG-CoA reductase gene may explain why statins effec-tively reduce inflammation in the vessel wall.8
* Corresponding author. Tel: þ358 17 162075; fax: þ358 17 163751.E-mail address: [email protected]
†These authors contributed equally to this work.‡Present address: Department of Molecular Cell Biology and Immunology,VU University Medical Center, Amsterdam, The Netherlands.
Published on behalf of the European Society of Cardiology. All rights reserved. & The Author 2008.For permissions please email: [email protected]
Cardiovascular Research (2008) 78, 175–184doi:10.1093/cvr/cvn007
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

Mechanisms underlying these pleiotropic effects remainincompletely understood. Studies implicate that the inhibitionof isoprenoid synthesis [geranyl-geranyl-pyrophosphate(GGPP) and farnesyl-pyrophosphate (FPP)] mediates theeffects of statins on GTP-binding proteins (e.g. Ras andRho).2,3,9 For example, Rho regulates eNOS gene expressionand controls SMC proliferation.10 Statins have been shown toinfluence the activity of some transcription factors: theyinhibit the binding of NF-kB and activator protein AP-1 tonuclear proteins in SMCs and ECs.11 Additionally, simvastatinhas been shown to block tumour necrosis factor(TNF)-a-induced NF-kB transcriptional activity and IkB phos-phorylation/degradation. Interestingly, statins have recentlybeen shown to upregulate the expression of Kruppel-likefactor-2 (KLF-2) in ECs.12 KLF-2 is a transcription factor ident-ified from the endothelial ‘atheroprotective phenotype’: itsoverexpression inhibits pro-inflammatory and pro-thromboticgene expression in ECs, such as vascular cell adhesionmolecule-1 (VCAM-1) and plasminogen activator inhibitor-1(PAI-1) expression, and its overexpression enhances theexpression of eNOS and thrombomodulin.12–15 KLF-2expression in ECs is modulated by shear stress, and thus itmight partially mediate the atheroprotective effects ofsteady laminar flow.16,17 Less is known about the role ofKLF-2 inmonocytes, but according to recent study, KLF-2 regu-lates the pro-inflammatory activation of monocytes, whichsuggests an anti-inflammatory role for KLF-2 in monocytes.18
ETS-1 transcription factor takes part in vascular inflam-mation and remodelling. It regulates the expression of anumber of vascular-specific genes, such as adhesion mol-ecules, chemokines (e.g. MCP-1), and MMPs.19,20 Itsexpression can be induced e.g. by pro-inflammatory cytokinesand vasoactive peptides such as angiotensin II,19 but there areno studies implicating statin effects on its expression.
Although there is already evidence that statins have anti-inflammatory properties, the mechanism of action remainsincompletely understood. Especially, very little is knownabout the global effects of statins on macrophage geneexpression. In this study, we show that simvastatin has astrong anti-inflammatory effect on human peripheral bloodmonocyte (HPBM)-macrophages, including downregulationof the expression of several cytokines, members of theTNF-family, and TF. These effects are mediated by theeffects on transcription factors: simvastatin reduces NF-kBsignalling pathway and c-ets, and upregulates KLF-2, whichimplicate an atheroprotective role for KLF-2 also in macro-phages. These findings may at least partly explain the ben-eficial pleiotropic effects of statins on cardiovasculardiseases.
2. Methods
2.1 Cell culture studies
HPBMs were isolated from buffy coats from healthy blood-donor vol-unteers (Finnish Red Cross, Helsinki, Finland) using Ficoll-Paque gra-dient centrifugation.21 None of the blood donors was on statintherapy. During isolation, monocytes from three to four individualswere pooled. Adherent cells were cultured in standard medium[RPMI 1640-medium with 20% human serum (Cambrex), 1% penicillinand streptomycin, and 1% L-glutamine) for differentiation intomacrophages. The macrophage-phenotype at day 7 after isolationwas confirmed with the typical shape of macrophages and also bymacrophage-immunostaining (mAB CD68, DAKO, Denmark), where
macrophages presented .95% of the cell population (data notshown). Human monocytic THP-1 cells (ATCC TIB-202) were culturedin RPMI-1640 medium according to ATCCs instructions. Cells werestimulated by 0.1 mM phorbol 12-myristate 13-acetate (PMA)(Sigma, USA) to induce differentiation into macrophages (¼‘THP-1macrophages’). The study protocol has been accepted by theEthics Committee of the University of Kuopio. The investigation con-forms with the principles outlined in the Declaration of Helsinki foruse of human tissues.
2.2 Simvastatin treatment
Simvastatin was a gift from Merck & Co. The inactive lactone form ofsimvastatin was hydrolyzed to the corresponding b-hydroxy acid.The HPBM-macrophages were treated with statin at day 7 afterthe isolation and the THP-1 cells at day 4 after the PMA-stimulation.Twelve hours prior to statin treatment, the cell growth mediawere changed to serum-free media. The statin-treated (10 mMsimvastatin in serum-free media) cells were collected at 12 and24 h for Affymetrix analyses. Statin-stimulated THP-1 macrophageswere collected also at 48 and 72 h time-points for TaqMan analyses.The simvastatin treatment was also carried at lower concentrationsof simvastatin (0.025–1 mM) in THP-1 macrophages for TaqMan analy-sis. The toxicity of simvastatin was assessed in cell culture: 5� con-centration of simvastatin (50 mM) had no effect on cell viability.
2.3 Inhibition of protein prenylation
The THP-1 macrophages were stimulated by 20 mM concentration offarnesyl transferase inhibitor (FTI-277, Sigma) and geranyl-geranyl-transferase inhibitor (GGTI-298, Sigma). FTI and GGTI were dis-solved in DMSO. Control cells were incubated with an equivalentconcentration of DMSO.
2.4 RNA isolation
Total RNA was isolated from the cells using Trizol reagent (GibcoBRL, USA) according to the manufacturers’ instructions. Thequality of RNA was assessed by spectrophotometry (NanoDropw,USA) and by agarose gel electrophoresis.
2.5 Microarray analyses
For Affymetrix analyses, three separate HPBM cell isolation andsimvastatin experiments were performed (HPBMs from three individ-uals at each time, total n ¼ 9). Cells were collected at 12 and 24 hafter the statin treatment. Total RNA was isolated as above. cDNAswere prepared from RNAs (5 mg of RNA) with reverse transcriptase(Superscript II primed by a poly(T) oligomer/T7 promoter). cDNAswere subsequently used as a template to make biotin-labelled cRNAwith an in vitro transcription reaction. cRNAs (15 mg) were hybridizedto Affymetrix HGU133 Plus 2.0 oligonucleotide arrays, which was pro-cessed and scanned according to the manufacturer’s instructions.Each array quantifies the expression of over 47 000 transcripts(including full-length mRNA sequences and ESTs) derived from build133 of the UniGene database (www.affymetrix.com).
2.6 Microarray data analysis
Affymetrix GeneChipw Operating Software (GCOS) was used to gen-erate. CEL files which were then converted into .DCP files usingdChip (http://www.dchip.org) V1.3 software.22 The arrays werenormalized to baseline array with median probe intensity, andgene expression data were generated calculating model-basedexpression values. The t-statistic is computed as (mean1 2
mean2)/sqrt(SE(mean1)2 þ SE(mean2)2), and its P-value is com-puted based on the t-distribution and the degree of freedom is setaccording to the Welch modified two-sample t-test. In this study,genes were considered differentially expressed if they changedmore than 1.5-fold (90% confidence bound of the fold change),
T.T. Tuomisto et al.176
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

absolute difference of signals was .100, at least 40% of the sampleswere called present in both groups and False Discovery Rate was�1%. Hierarchical clustering was performed by dChip using thePearson correlation with a centroid-linkage method.23 Gene func-tion analysis was performed by using the gene ontology miningtool GoSurfer incorporated in dChip program. The.CEL files andpivot table.txt tabdelimited files (GCOS) are available at GEO repo-sitory (www.ncbi.nlm.nih.gov/GEO) with series record GSE4883.
2.7 Real-time quantitative RT-PCR (TaqMan)analyses
TaqMan analyses were performed to validate gene expression changesof the selected genes. TaqMan analyses were performed from thesame HPBM-samples as Affymetrix analyses and in THP-1 macro-phages with protein prenylation inhibitors. Total RNA was convertedto cDNA by in vitro transcription reaction (M-MuLV Reverse Transcrip-tase, Finnzymes, Finland). cDNAs (10–25 ng of cDNA depending onthe gene) were used as templates for TaqMan qRT-PCR with ABIAssays-on-Demand on ABI Prism 7900 sequence detection system.The specific assays used were Hs00360439-g1 (KLF-2),Hs00234140-m1 (MCP-1), Hs00175225-m1 (TF), Hs00234142-m1(MIP-1a), Hs00242737-m1 (LTB), and Hs00231279-m1 (p65/RelA). Allsamples were run in quarduplicate, and rRNA assay (Ribosomal RNAcontrol reagent, ABI) was used as an internal control to normalizethe RNA amount. Quarduplicates were averaged to calculate anexpression value for each sample, and data were presented asmean expression value relative to the control+ standard deviation.To evaluate statistical significances, independent samples t-testwas used for the appropriate parameters (Microsoft Excel).
2.8 Western blot analyses
HPBM and THP-1 cells treated with protein prenylation inhibitorswere lysed in lysis buffer (50 mM Tris, pH 7,5, 150 mM NaCl, 1 mMEDTA, 1% Triton X-100, 0.5% sodium deoxylacholate, 0.1% SDS, and10% glycerol). Cells were incubated on ice for 10 min and centri-fuged (10 000 g) for 10 min at þ48C. Supernatants were transferredinto new tubes and the protein concentration was determined usingthe BCA protein assay kit (Pierce, USA). For western blot, sample(20 mg of protein) was separated in 12% SDS–PAGE gel. After electro-phoresis, proteins were transferred onto nitrocellulose membranes(Trans-Blot Transfer Medium, Bio-Rad Laboratories, CA, USA). Mem-branes were blocked overnight at þ48C (5% goat serum, Vector lab-oratories) in Tris-buffered saline with 0.1% Tween 20 (TBS/T), pH7.6. Membranes were incubated with a diluted primary antibody(Anti-KLF-2, Abcam Cambridge, UK, 3 mg/mL) in 5% goat serum inTBS/T overnight at þ48C. After washing with TBS/T, membraneswere incubated with a diluted secondary antibody (Peroxidase-conjugated Affinipure Donkey Anti-Goat, Jackson Immunoresearch,USA, 8 ng/mL). Antigen–antibody complexes were detected bychemiluminecence (Supersignal West Dura Extended DurationSubstrate, Pierce) and exposed to a high-performance chemilumi-nescence film (Amersham Biosciences, UK).
2.9 Kruppel-like factor 2 overexpression studies
The lentiviral vector constructs and the preparation of the viruseshave been described elsewhere.17,24 As controls for lentivirus over-expressing KLF-2, we used respective virus without KLF-2 andno-virus-containing control. THP-1 derived macrophages were trans-duced overnight using MOI 5 and MOI 10. After 7 days, RNA was iso-lated for TaqMan analyses of KLF-2, MCP-1, and TF expression asabove. Co-stimulations of simvastatin and lentiviral overexpressionof KLF-2 were performed with THP-1 macrophages. Seven daysafter lentiviral overexpression of KLF-2 (MOI 10) and/or statinstimulation (day 6, 10 mM simvastatin), RNA was isolated and usedfor the analysis of KLF-2, MCP-1, and TF expression as above.
3. Results
3.1 Simvastatin attenuates the expression ofinflammatory genes in human peripheral bloodmonocytes
To examine the transcriptional response to simvastatin inHPBMs, we performed genome wide gene expression analysisusing Affymetrix gene chips at time-points 12 and 24 h afterthe statin treatment. From the �47 000 transcripts on theAffymetrix gene chip, a total of 589 genes showed statisti-cally significant changes in expression either at 12 or 24 has compared with the control group using a value of 1.5for the lower bound of the 90% confidence interval for thefold change as a cutoff. When analysed separately, 280genes were either up- or downregulated at 12 h and 502genes at 24 h. As a general phenomenon, we found moredownregulated genes that upregulated genes after thestatin treatment.
To assess the relationships and coordinate expression profilesbetween the regulated genes, we performed a cluster analysisof the pooled dataset. The analysis showed 10 well-definedclusters, which contained several genes that were related(e.g. the same gene family or the functional group), forexample ‘inflammatory gene cluster’ (cluster 3), ‘cell struc-ture’ (cluster 2), ‘cholesterol-metabolism-cluster’ (cluster 5),‘signal transduction with downregulation’ (cluster 4), ‘signaltransduction with upregulation’ (cluster 6), ‘KLF-cluster’(cluster 8), and ‘metallothionein cluster’ (cluster 10)(Figure 1, Table S1 in Supplementary material online).
We found that the inhibition of HMG-CoA reductase inmacrophages and the subsequent reduction in the choles-terol synthesis lead to compensatory changes at 24 h inseveral genes functioning in the cholesterol metabolism(Table 1): HMG-CoA reductase was induced 1.85-fold, aswell as enzymes participating in the HMG-CoA synthesis(Acetyl-CoA synthase 1.73-fold; HMG-CoA synthase2.82-fold). We also found induction of downstreamenzymes, such as farnesyl transferase (1.53) and squaleneepoxidase (1.96). Additionally, LDL-receptor gene wasinduced 1.79-fold. A similar pattern of changes in geneexpression (upregulation at 24 h) was also seen incholesterol-metabolism-related genes in cluster analysis:the above-mentioned genes formed a separate cluster(cluster 5) (Figure 1, Table S1 in Supplementary materialonline). Therefore, we concluded that the simvastatin treat-ment was efficient enough to affect cholesterol and prenylmetabolism, and the Affymetrix system was sensitiveenough to detect these changes in gene expression.
Simvastatin treatment led to a reduced mRNA expressionof many inflammatory genes (Table 1). These genes includeseveral chemokines, such as MCP-1 and macrophage inflam-matory protein-1a (MIP-1a) and -b, RANTES, CXC9, andCXC10; several members of TNF superfamily, such as TRAILand lymphotoxin b (LTB); interleukin receptors, adhesionmolecules, such as VCAM-1 and ICAM-3; and TF. Severalinflammatory genes and mediators of inflammatory gene sig-nalling had similar expression patterns in macrophages, asdemonstrated by cluster analysis. The cluster number 3,which includes genes with downregulated gene expression,contains almost entirely inflammatory genes, and includesthe above-mentioned members of the cytokine andTNF-families.
Simvastatin is anti-inflammatory in macrophages 177
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

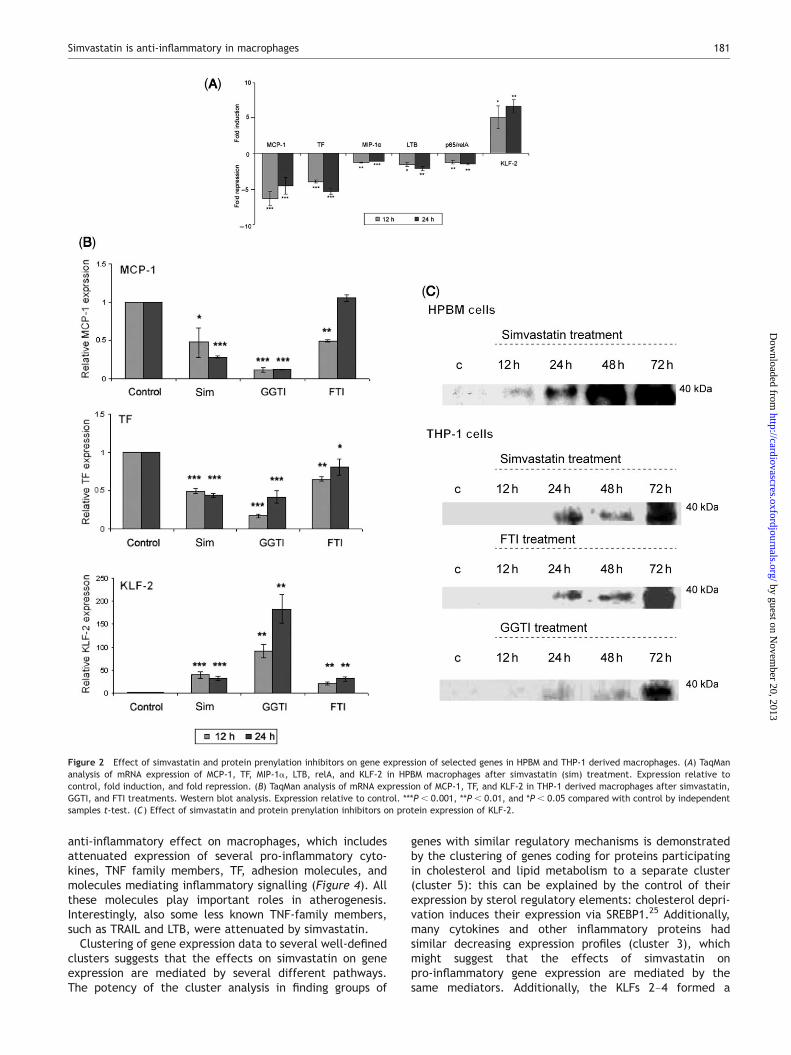
The expression changes in cytokines MCP-1 and MIP-1a,TF, and a member of TNF-family (LTB) were further con-firmed by TaqMan analyses in HPBM as well as in THP-1derived macrophages (Figure 2A and B). Downregulation ofMCP-1 and TF sustained for 48 and 72 h in THP-1 cells(data not shown).
3.2 Simvastatin attenuates inflammatory signallingin macrophages: downregulation of NF-kB relA/p65subunit mRNA expression and upregulation ofKruppel-like factor 2 expression
In Affymetrix analysis, simvastatin treatment lead to reducedexpression of several genes that mediate inflammatory sig-nalling in cells, including several guanylate-binding proteins(GBP 1–5), Ras and Rho-proteins; and several transcriptionfactors, such as NF-kB relA/p65 protein, PPARd, and v-ets.The expression of KLF-2 was strongly induced (3.9-fold upre-gulation at 12 h, 9.3-fold at 24 h). Additionally, the KLF familymembers KLF-3 and -4 had increased expression levels. Mostof the inflammatory signalling-related genes clusteredtogether with other inflammatory genes to cluster 3, which
shows decreasing expression pattern, whereas KLFs 2–4formed a separate cluster with increasing expression patterns(cluster 8) (Figure 1, Table S1 in Supplementary materialonline).
In Affymetrix analysis, several members of the NF-kBsignalling pathway showed a trend to reduced RNAexpression levels, for example NF-kB /rel proteinRelA/p65 (22.0-fold decrease at 24 h), RelB and NF-kBp50, but all the changes were not statistically signi-ficant. Additionally, several members of the signallingcascades leading to NF-kB activation had downregulatedexpression, e.g. protein kinase C (21.4) and Akt and Cot(21.4). Also JAK-STAT (STAT 21.6-fold) signalling pathwaywas donwnregulated.
The expression changes of relA/p65 and KLF-2 werefurther studied with TaqMan in HPBM and THP-1 macro-phages (Figure 2), and KLF-2 protein expression bywestern blot analysis (Figure 2C). TaqMan analysis confirmedthe downregulated levels of relA/p65 mRNA in HPBM cells(Figure 2A), as well as the induction of KLF-2 expression inHPBM and THP-1 macrophages (Figure 2A and B). Therewas also an increase in the KLF-2 protein (Figure 2C). Theeffect of simvastatin on KLF-2, TF, and MCP-1 expressionwas detected also at concentration of 1 mM simvastatin asstudied by TaqMan analysis (data not shown). However, atlower concentrations of simvastatin (0.025–0.1 mM), theeffect was not as remarkable.
3.3 The inhibition of protein prenylation has thesame effect on Kruppel-like factor 2, monocytechemotactic protein-1, and tissue factor expressionas simvastatin
THP-1 macrophages were treated with protein prenylationinhibitors GGTI (inhibitor of geranyl-geranyl transferase)and FTI (inhibitor of farnesyl transferase). Both GGTI andFTI treatments lead to the downregulation of MCP-1 andTF mRNA expression and to the upregulation of KLF-2expression (Figure 2B). Moreover, simvastatin, GGTI, andFTI treatments had similar effects on KLF-2 proteinexpression (Figure 2C).
3.4 Monocyte chemotactic protein-1 and tissuefactor are Kruppel-like factor 2 target genes inmacrophages
To test whether MCP-1 and TF are KLF-2 target genes inmacrophages, we performed lentiviral-mediated KLF-2overexpression in THP-1 macrophages. Lentiviral transduc-tion led to very strong (�6000 and �12 000-fold at 7days for MOI 5 and MOI 10, respectively) expression ofKLF-2 in macrophages (Figure 3A). In macrophages over-expressing KLF-2, the expression of MCP-1 was repressed25.9-fold (MOI 10) and TF 21.9-fold (MOI 10) relative tocontrol virus (Figure 3A). We also studied the effects of len-tiviral KLF-2 overexpression in combination with statintreatment in THP-1 macrophages (Figure 3B). However,simvastatin treatment had no additional effects over KLF-2overexpression.
4. Discussion
This gene expression array study of simvastatin effectson macrophages shows that simvastatin has a global
Figure 1 Cluster analysis of gene expression data. Clustering reveals 10well-defined clusters including ‘leukotriene signalling’ (cluster 1), ‘cell struc-ture’ (cluster 2), ‘inflammatory genes’ (cluster 3), ‘signal transduction,downregulated’ (cluster 4), ‘cholesterol metabolism’ (cluster 5), ‘signaltransduction, upregulated’ (cluster 6), ‘orosomucoid-cluster’ (cluster 7),‘KLF-cluster’ (cluster 8), ‘tiny cluster’ (cluster 9), and ‘metallothioneins’(cluster 10). Red indicates upregulated expression relative to control (base-line) and green indicates downregulated expression. The list of genes indifferent clusters is presented in Table S1 in Supplementary material online.
T.T. Tuomisto et al.178
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
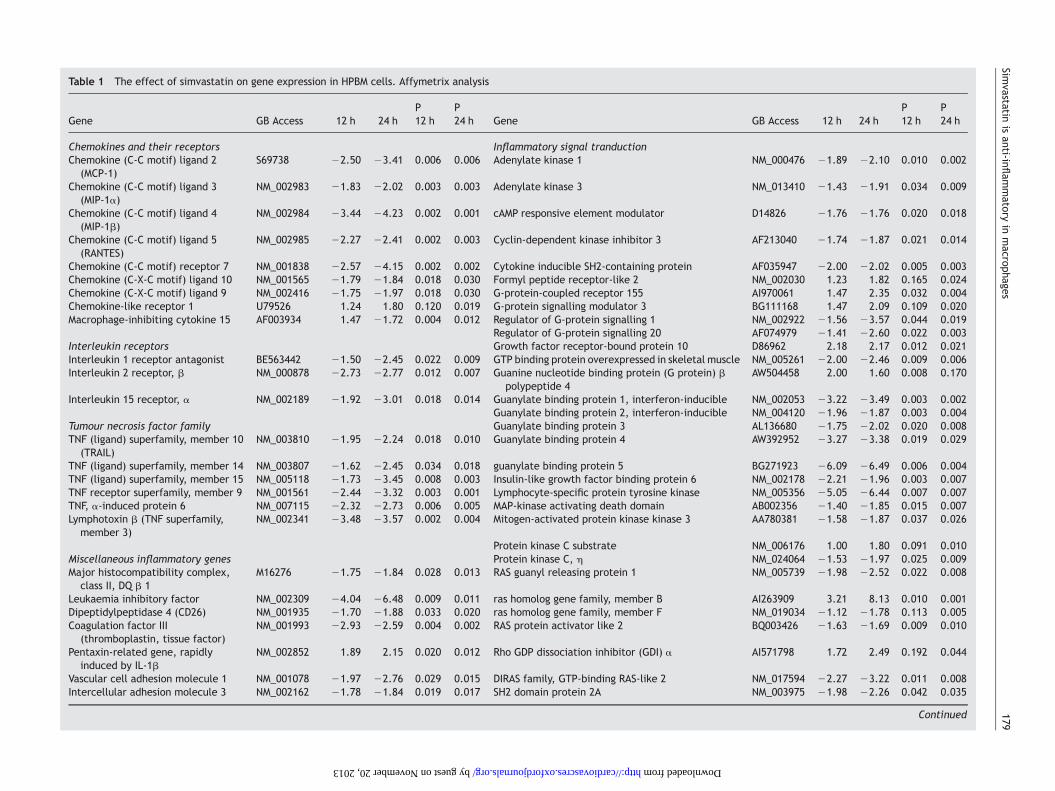
Table 1 The effect of simvastatin on gene expression in HPBM cells. Affymetrix analysis
Gene GB Access 12 h 24 hP12 h
P24 h Gene GB Access 12 h 24 h
P12 h
P24 h
Chemokines and their receptors Inflammatory signal tranductionChemokine (C-C motif) ligand 2
(MCP-1)S69738 22.50 23.41 0.006 0.006 Adenylate kinase 1 NM_000476 21.89 22.10 0.010 0.002
Chemokine (C-C motif) ligand 3(MIP-1a)
NM_002983 21.83 22.02 0.003 0.003 Adenylate kinase 3 NM_013410 21.43 21.91 0.034 0.009
Chemokine (C-C motif) ligand 4(MIP-1b)
NM_002984 23.44 24.23 0.002 0.001 cAMP responsive element modulator D14826 21.76 21.76 0.020 0.018
Chemokine (C-C motif) ligand 5(RANTES)
NM_002985 22.27 22.41 0.002 0.003 Cyclin-dependent kinase inhibitor 3 AF213040 21.74 21.87 0.021 0.014
Chemokine (C-C motif) receptor 7 NM_001838 22.57 24.15 0.002 0.002 Cytokine inducible SH2-containing protein AF035947 22.00 22.02 0.005 0.003Chemokine (C-X-C motif) ligand 10 NM_001565 21.79 21.84 0.018 0.030 Formyl peptide receptor-like 2 NM_002030 1.23 1.82 0.165 0.024Chemokine (C-X-C motif) ligand 9 NM_002416 21.75 21.97 0.018 0.030 G-protein-coupled receptor 155 AI970061 1.47 2.35 0.032 0.004Chemokine-like receptor 1 U79526 1.24 1.80 0.120 0.019 G-protein signalling modulator 3 BG111168 1.47 2.09 0.109 0.020Macrophage-inhibiting cytokine 15 AF003934 1.47 21.72 0.004 0.012 Regulator of G-protein signalling 1 NM_002922 21.56 23.57 0.044 0.019
Regulator of G-protein signalling 20 AF074979 21.41 22.60 0.022 0.003Interleukin receptors Growth factor receptor-bound protein 10 D86962 2.18 2.17 0.012 0.021Interleukin 1 receptor antagonist BE563442 21.50 22.45 0.022 0.009 GTP binding protein overexpressed in skeletal muscle NM_005261 22.00 22.46 0.009 0.006Interleukin 2 receptor, b NM_000878 22.73 22.77 0.012 0.007 Guanine nucleotide binding protein (G protein) b
polypeptide 4AW504458 2.00 1.60 0.008 0.170
Interleukin 15 receptor, a NM_002189 21.92 23.01 0.018 0.014 Guanylate binding protein 1, interferon-inducible NM_002053 23.22 23.49 0.003 0.002Guanylate binding protein 2, interferon-inducible NM_004120 21.96 21.87 0.003 0.004
Tumour necrosis factor family Guanylate binding protein 3 AL136680 21.75 22.02 0.020 0.008TNF (ligand) superfamily, member 10
(TRAIL)NM_003810 21.95 22.24 0.018 0.010 Guanylate binding protein 4 AW392952 23.27 23.38 0.019 0.029
TNF (ligand) superfamily, member 14 NM_003807 21.62 22.45 0.034 0.018 guanylate binding protein 5 BG271923 26.09 26.49 0.006 0.004TNF (ligand) superfamily, member 15 NM_005118 21.73 23.45 0.008 0.003 Insulin-like growth factor binding protein 6 NM_002178 22.21 21.96 0.003 0.007TNF receptor superfamily, member 9 NM_001561 22.44 23.32 0.003 0.001 Lymphocyte-specific protein tyrosine kinase NM_005356 25.05 26.44 0.007 0.007TNF, a-induced protein 6 NM_007115 22.32 22.73 0.006 0.005 MAP-kinase activating death domain AB002356 21.40 21.85 0.015 0.007Lymphotoxin b (TNF superfamily,
member 3)NM_002341 23.48 23.57 0.002 0.004 Mitogen-activated protein kinase kinase 3 AA780381 21.58 21.87 0.037 0.026
Protein kinase C substrate NM_006176 1.00 1.80 0.091 0.010Miscellaneous inflammatory genes Protein kinase C, h NM_024064 21.53 21.97 0.025 0.009Major histocompatibility complex,
class II, DQ b 1M16276 21.75 21.84 0.028 0.013 RAS guanyl releasing protein 1 NM_005739 21.98 22.52 0.022 0.008
Leukaemia inhibitory factor NM_002309 24.04 26.48 0.009 0.011 ras homolog gene family, member B AI263909 3.21 8.13 0.010 0.001Dipeptidylpeptidase 4 (CD26) NM_001935 21.70 21.88 0.033 0.020 ras homolog gene family, member F NM_019034 21.12 21.78 0.113 0.005Coagulation factor III
(thromboplastin, tissue factor)NM_001993 22.93 22.59 0.004 0.002 RAS protein activator like 2 BQ003426 21.63 21.69 0.009 0.010
Pentaxin-related gene, rapidlyinduced by IL-1b
NM_002852 1.89 2.15 0.020 0.012 Rho GDP dissociation inhibitor (GDI) a AI571798 1.72 2.49 0.192 0.044
Vascular cell adhesion molecule 1 NM_001078 21.97 22.76 0.029 0.015 DIRAS family, GTP-binding RAS-like 2 NM_017594 22.27 23.22 0.011 0.008Intercellular adhesion molecule 3 NM_002162 21.78 21.84 0.019 0.017 SH2 domain protein 2A NM_003975 21.98 22.26 0.042 0.035
Continued
Simvastatin
isanti-infl
ammatory
inmacrop
hages179
by guest on November 20, 2013 http://cardiovascres.oxfordjournals.org/ Downloaded from
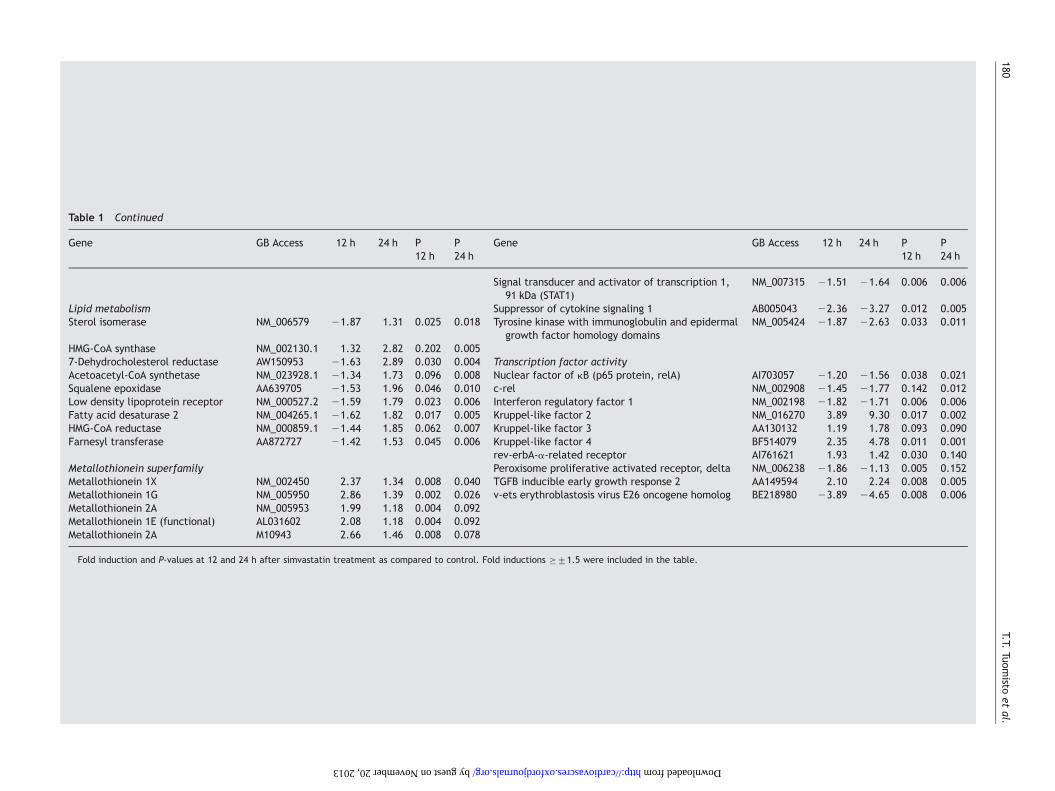
Table 1 Continued
Gene GB Access 12 h 24 h P12 h
P24 h
Gene GB Access 12 h 24 h P12 h
P24 h
Signal transducer and activator of transcription 1,91 kDa (STAT1)
NM_007315 21.51 21.64 0.006 0.006
Lipid metabolism Suppressor of cytokine signaling 1 AB005043 22.36 23.27 0.012 0.005Sterol isomerase NM_006579 21.87 1.31 0.025 0.018 Tyrosine kinase with immunoglobulin and epidermal
growth factor homology domainsNM_005424 21.87 22.63 0.033 0.011
HMG-CoA synthase NM_002130.1 1.32 2.82 0.202 0.0057-Dehydrocholesterol reductase AW150953 21.63 2.89 0.030 0.004 Transcription factor activityAcetoacetyl-CoA synthetase NM_023928.1 21.34 1.73 0.096 0.008 Nuclear factor of kB (p65 protein, relA) AI703057 21.20 21.56 0.038 0.021Squalene epoxidase AA639705 21.53 1.96 0.046 0.010 c-rel NM_002908 21.45 21.77 0.142 0.012Low density lipoprotein receptor NM_000527.2 21.59 1.79 0.023 0.006 Interferon regulatory factor 1 NM_002198 21.82 21.71 0.006 0.006Fatty acid desaturase 2 NM_004265.1 21.62 1.82 0.017 0.005 Kruppel-like factor 2 NM_016270 3.89 9.30 0.017 0.002HMG-CoA reductase NM_000859.1 21.44 1.85 0.062 0.007 Kruppel-like factor 3 AA130132 1.19 1.78 0.093 0.090Farnesyl transferase AA872727 21.42 1.53 0.045 0.006 Kruppel-like factor 4 BF514079 2.35 4.78 0.011 0.001
rev-erbA-a-related receptor AI761621 1.93 1.42 0.030 0.140Metallothionein superfamily Peroxisome proliferative activated receptor, delta NM_006238 21.86 21.13 0.005 0.152Metallothionein 1X NM_002450 2.37 1.34 0.008 0.040 TGFB inducible early growth response 2 AA149594 2.10 2.24 0.008 0.005Metallothionein 1G NM_005950 2.86 1.39 0.002 0.026 v-ets erythroblastosis virus E26 oncogene homolog BE218980 23.89 24.65 0.008 0.006Metallothionein 2A NM_005953 1.99 1.18 0.004 0.092Metallothionein 1E (functional) AL031602 2.08 1.18 0.004 0.092Metallothionein 2A M10943 2.66 1.46 0.008 0.078
Fold induction and P-values at 12 and 24 h after simvastatin treatment as compared to control. Fold inductions �+1.5 were included in the table.
T.T.Tuom
istoet
al.180
by guest on November 20, 2013 http://cardiovascres.oxfordjournals.org/ Downloaded from
anti-inflammatory effect on macrophages, which includesattenuated expression of several pro-inflammatory cyto-kines, TNF family members, TF, adhesion molecules, andmolecules mediating inflammatory signalling (Figure 4). Allthese molecules play important roles in atherogenesis.Interestingly, also some less known TNF-family members,such as TRAIL and LTB, were attenuated by simvastatin.
Clustering of gene expression data to several well-definedclusters suggests that the effects on simvastatin on geneexpression are mediated by several different pathways.The potency of the cluster analysis in finding groups of
genes with similar regulatory mechanisms is demonstratedby the clustering of genes coding for proteins participatingin cholesterol and lipid metabolism to a separate cluster(cluster 5): this can be explained by the control of theirexpression by sterol regulatory elements: cholesterol depri-vation induces their expression via SREBP1.25 Additionally,many cytokines and other inflammatory proteins hadsimilar decreasing expression profiles (cluster 3), whichmight suggest that the effects of simvastatin onpro-inflammatory gene expression are mediated by thesame mediators. Additionally, the KLFs 2–4 formed a
Figure 2 Effect of simvastatin and protein prenylation inhibitors on gene expression of selected genes in HPBM and THP-1 derived macrophages. (A) TaqMananalysis of mRNA expression of MCP-1, TF, MIP-1a, LTB, relA, and KLF-2 in HPBM macrophages after simvastatin (sim) treatment. Expression relative tocontrol, fold induction, and fold repression. (B) TaqMan analysis of mRNA expression of MCP-1, TF, and KLF-2 in THP-1 derived macrophages after simvastatin,GGTI, and FTI treatments. Western blot analysis. Expression relative to control. ***P , 0.001, **P , 0.01, and *P , 0.05 compared with control by independentsamples t-test. (C) Effect of simvastatin and protein prenylation inhibitors on protein expression of KLF-2.
Simvastatin is anti-inflammatory in macrophages 181
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
separate cluster (cluster 8), as well as metallothioneins(cluster 10). Several metallothioneins (MT 1E, X, G, and2A), which participate in the metal homeostasis as well asthe control of REDOX-balance, inflammation, and cell pro-liferation,26 are induced at 12 h by simvastatin in a similarmanner. However, their role in macrophages has not beenestablished, and there is no previous information abouttheir regulation by statins.
The anti-inflammatory effect of simvastatin can be atleast partly explained by its impact on pro-inflammatorysignal transduction. Several members of the NF-kB signallingpathway had reduced mRNA expression levels, for exampleNF-kB complex proteins p65/RelA, RelB, and p50. Addition-ally, several members of the signalling cascades leading toNF-kB activation had downregulated expression, e.g. Januskinases, protein kinase C, Akt and Cot. It has been previouslyshown that statins inhibit the binding of NF-kB to nuclearproteins11 and inhibit the phosphorylation and degradationof the NF-kB inhibitory protein IkB.27 We show here that sim-vastatin downregulates mRNA expression of the NF-kB sub-units in HPBM cells. Besides NF-kB signalling, a potent
pro-inflammatory transcription factor c-ets, which mediatese.g. MCP-1 and phospholipase expression,19,20 as well as thefamily of GBPs 1–5, and JAK-STAT signalling pathway weredownregulated by simvastatin.
We show here that KLF-2, as well as its family membersKLF-3 and KLF-4, are induced in macrophages by simvastatin.KLFs are recently characterized zinc finger transcriptionfactors that have several important functions; they modu-late cell differentiation, organ development, and prolifer-ation.16 KLF-2, -4, -5, and -6 have particular importance inthe vasculature, and they have functional as well as patho-logical roles.16 KLF-2 has been shown to be essential for thevascular development,16 and it has been identified as part ofthe ‘atheroprotective phenotype’ of ECs. It is induced bysteady laminar flow28,29 and inhibited by TNF-a28 andIL-1b14 in ECs. It has several anti-inflammatory properties,such as reduction of adhesion molecule expression.14
Additionally, KLF-2 has been shown to regulate the thrombo-tic function of ECs, e.g. the expression of thrombomodulin,eNOS, and PAI-1 as wells as cytokine-mediated production ofTF.13 Global analysis of gene expression changes after ade-noviral overexpression of KLF-2 in ECs showed that inductionof KLF-2 resulted in the regulation of endothelial transcrip-tion programmes controlling inflammation, thrombosis/hemostasis, vascular tone, and blood vessel development.12
Moreover, another microarray study showed that KLF-2 actsas a general transcriptional switch point between the quies-cent and activated status of ECs where lentiviral overex-pression of KLF-2 resulted in changes in several keyfunctional pathways such as cell migration via VEGFR2,inflammation, and hemostasis.30
Recently, it has been shown that statins upregulate KLF-2expression in ECs, which might partly explain the atheropro-tective effects of statins.12,15 The statin effect on KLF-2 wasdependent on the ability of statins to inhibit Rho-pathway,as adenoviral overexpression of Rho-protein decreasedKLF-2 expression.15
Very little is known about the role of KLF-2 in macro-phages. A recent study showed that KLF-2 regulatespro-inflammatory activation of monocytes: overexpression
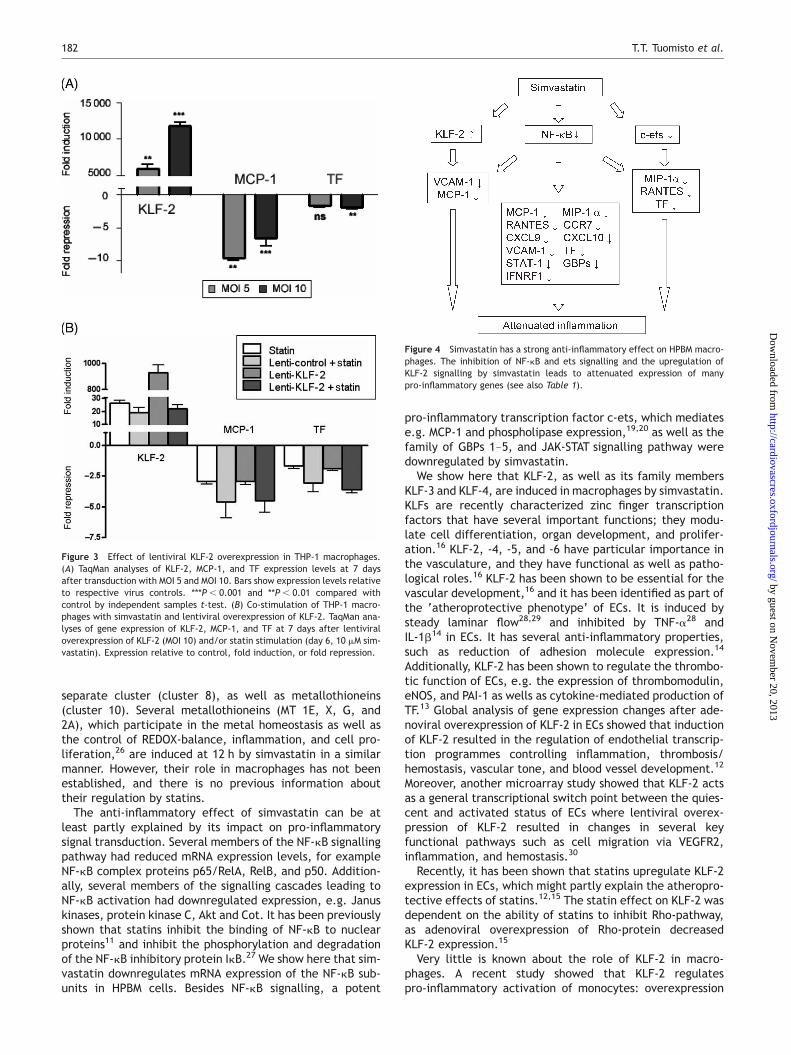
Figure 3 Effect of lentiviral KLF-2 overexpression in THP-1 macrophages.(A) TaqMan analyses of KLF-2, MCP-1, and TF expression levels at 7 daysafter transduction with MOI 5 and MOI 10. Bars show expression levels relativeto respective virus controls. ***P , 0.001 and **P , 0.01 compared withcontrol by independent samples t-test. (B) Co-stimulation of THP-1 macro-phages with simvastatin and lentiviral overexpression of KLF-2. TaqMan ana-lyses of gene expression of KLF-2, MCP-1, and TF at 7 days after lentiviraloverexpression of KLF-2 (MOI 10) and/or statin stimulation (day 6, 10 mM sim-vastatin). Expression relative to control, fold induction, or fold repression.
Figure 4 Simvastatin has a strong anti-inflammatory effect on HPBM macro-phages. The inhibition of NF-kB and ets signalling and the upregulation ofKLF-2 signalling by simvastatin leads to attenuated expression of manypro-inflammatory genes (see also Table 1).
T.T. Tuomisto et al.182
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

of KLF-2 inhibited LPS-activated production of cytokines andreduced phagocytic activity.18 Interestingly, the expressionof KLF-2 was reduced in circulating monocytes in patientswith coronary artery disease.18 In this study, we showedthat KLF-2 is induced by simvastatin in mature humanmacrophages. We also showed that the mechanism of induc-tion of KLF-2 by simvastatin in macrophages is dependent onthe inhibition of protein prenylation. Additionally, by lenti-viral overexpression of KLF-2, we showed that MCP-1 andTF are KLF-2 target genes in macrophages, and KLF-2represses their expression. Moreover, several KLF-2 ECtarget genes,12 e.g. CXCL10, CCL5, and TNF, had changedexpression levels in simvastatin-stimulated macrophages,which suggests them to be potential KLF-2 target genes inmacrophages as well. Taken together, this suggests an anti-inflammatory and vasculoprotective role for KLF-2 inmacrophages.
In addition to the induction of KLF-2, we showed that alsoits family members KLF-3 and KLF-4 are induced by simvas-tatin. However, the functions of KLF-3 and KLF-4 are poorlyknown and further studies are needed to clarify their role inmacrophages and other cell types.
The inhibition of protein prenylation by inhibitors ofgeranyl-geranylation and farnesylation had similar effectson KLF-2, MCP-1, and TF expression as simvastatin. Isopre-noid binding to Ras and Rho proteins is essential for theinitiation of G-protein signalling, which controls forexample cell proliferation and stress responses. Farnesy-lated proteins include e.g. Ras, whereas Rho and Rac acti-vations are dependent on geranyl-geranylation.9 Thesimilar effects achieved by protein prenylation inhibitorsand simvastatin suggest that the simvastatin effects mightbe dependent on the inhibition of protein prenylation. Itwould be interesting to study whether alterations in cellularcholesterol content correlate with these effects andwhether similar effects on cellular transcriptional machin-ery could be achieved via other means that can alter cellularcholesterol levels.
In conclusion, we show here that simvastatin has a stronganti-inflammatory effect on macrophages including attenu-ated expression of several cytokines, TNF family membersand some pro-inflammatory signalling molecules (Figure 4).Most interestingly, transcription factor KLF-2 is upregulatedby simvastatin in macrophages. These findings support thenotion that statins have other anti-atherogenic actionsbeyond their effects on plasma cholesterol.
Supplementary material
Supplementary material is available at CardiovascularResearch online.
Acknowledgements
We thank Ms Riina Kylatie, Ms Mervi Nieminen, Ms Anne Martikainen,and Ms Marja Poikolainen for technical assistance.
Conflict of interest: none declared.
Funding
This study has been supported by the Academy of Finland(211497), Finnish Foundation for Cardiovascular Research,
Sigrid Juselius Foundation, Finnish Cultural Foundation (toT.T.T.), Finnish Medical Foundation (to T.T.T.), the AarneKoskelo Foundation (to T.T.T.), the Paavo Nurmi Foundation(to T.T.T.), and European Vascular Genomics Network(EVGN LSHM-CT-2003-503254).
References1. Hansson GK. Inflammation, atherosclerosis, and coronary artery disease.
N Engl J Med 2005;352:1685–1695.2. Palinski W, Napoli C. Unraveling pleiotropic effects of statins on plaque
rupture. Arterioscler Thromb Vasc Biol 2002;22:1745–1750.3. Ray KK, Cannon CP. Intensive statin therapy in acute coronary syndromes:
clinical benefits and vascular biology. Curr Opin Lipidol 2004;15:637–643.
4. Aikawa M, Rabkin E, Sugiyama S, Voglic SJ, Fukumoto Y, Furukawa Yet al.An HMG-CoA reductase inhibitor, cerivastatin, suppresses growth ofmacrophages expressing matrix metalloproteinases and tissue factor invivo and in vitro. Circulation 2001;103:276–283.
5. Waehre T, Yndestad A, Smith C, Haug T, Tunheim SH, Gullestad L et al.Increased expression of interleukin-1 in coronary artery disease withdownregulatory effects of HMG-CoA reductase inhibitors. Circulation2004;109:1966–1972.
6. Yoshida M, Sawada T, Ishii H, Gerszten RE, Rosenzweig A, Gimbrone MA Jret al. Hmg-CoA reductase inhibitor modulates monocyte-EC interactionunder physiological flow conditions in vitro: involvement of Rho GTPase-dependent mechanism. Arterioscler Thromb Vasc Biol 2001;21:1165–1171.
7. Senokuchi T, Matsumura T, Sakai M, Yano M, Taguchi T, Matsuo T et al.Statins suppress oxidized low density lipoprotein-induced macrophageproliferation by inactivation of the small G protein-p38 MAPK pathway.J Biol Chem 2005;280:6627–6633.
8. Tuomisto TT, Korkeela A, Rutanen J, Viita H, Brasen JH, Riekkinen MS etal. Gene expression in macrophage-rich inflammatory cell infiltrates inhuman atherosclerotic lesions as studied by laser microdissection andDNA array: overexpression of HMG-CoA reductase, colony stimulatingfactor receptors, CD11A/CD18 integrins, and interleukin receptors.Arterioscler Thromb Vasc Biol 2003;23:2235–2240.
9. Denhardt DT. Signal-transducing protein phosphorylation cascadesmediated by Ras/Rho proteins in the mammalian cell: the potential formultiplex signalling. Biochem J 1996;318:729–747.
10. Laufs U, Liao JK. Targeting Rho in cardiovascular disease. Circ Res 2000;87:526–528.
11. Dichtl W, Dulak J, Frick M, Alber HF, Schwarzacher SP, Ares MP et al.HMG-CoA reductase inhibitors regulate inflammatory transcriptionfactors in human endothelial and vascular smooth muscle cells. Arterios-cler Thromb Vasc Biol 2003;23:58–63.
12. Parmar KM, Nambudiri V, Dai G, Larman HB, Gimbrone MA Jr,Garcia-Cardena G. Statins exert endothelial atheroprotective effectsvia the KLF2 transcription factor. J Biol Chem 2005;280:26714–26719.
13. Lin Z, Kumar A, Senbanerjee S, Staniszewski K, Parmar K, Vaughan DE etal. Kruppel-like factor 2 (KLF2) regulates endothelial thrombotic func-tion. Circ Res 2005;96:e48–e57.
14. Senbanerjee S, Lin Z, Atkins GB, Greif DM, Rao RM, Kumar A et al. KLF2 isa novel transcriptional regulator of endothelial proinflammatory acti-vation. J Exp Med 2004;199:1305–1315.
15. Sen-Banerjee S, Mir S, Lin Z, Hamik A, Atkins GB, Das H et al. Kruppel-likefactor 2 as a novel mediator of statin effects in endothelial cells. Circula-tion 2005;112:720–726.
16. Suzuki T, Aizawa K, Matsumura T, Nagai R. Vascular implications of theKruppel-like family of transcription factors. Arterioscler Thromb VascBiol 2005;25:1135–1141.
17. Dekker RJ, van Thienen JV, Rohlena J, de Jager SC, Elderkamp YW,Seppen J et al. Endothelial KLF2 links local arterial shear stress levelsto the expression of vascular tone-regulating genes. Am J Pathol 2005;167:609–618.
18. Das H, Kumar A, Lin Z, Patino WD, Hwang PM, Feinberg MW et al. Kruppel-like factor 2 (KLF2) regulates proinflammatory activation of monocytes.Proc Natl Acad Sci USA 2006;103:6653–6658.
19. Oettgen P. Regulation of vascular inflammation and remodeling by ETSfactors. Circ Res 2006;99:1159–1166.
20. Zhan Y, Brown C, Maynard E, Anshelevich A, Ni W, Ho IC et al. Ets-1is a critical regulator of Ang II-mediated vascular inflammation andremodeling. J Clin Invest 2005;115:2508–2516.
Simvastatin is anti-inflammatory in macrophages 183
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from

21. Pietarinen-Runtti P, Lakari E, Raivio KO, Kinnula VL. Expression of antiox-idant enzymes in human inflammatory cells. Am J Physiol Cell Physiol2000;278:C118–C125.
22. Li C, Wong WH. Model-based analysis of oligonucleotide arrays:expression index computation and outlier detection. Proc Natl Acad SciUSA 2001;98:31–36.
23. Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis and displayof genome-wide expression patterns. Proc Natl Acad Sci USA 1998;95:14863–14868.
24. Makinen PI, Koponen JK, Karkkainen AM, Malm TM, Pulkkinen KH,Koistinaho J et al. Stable RNA interference: comparison of U6 and H1 pro-moters in endothelial cells and in mouse brain. J Gene Med 2006;8:433–441.
25. Wang X, Sato R, Brown MS, Hua X, Goldstein JL. SREBP-1, a membrane-bound transcription factor released by sterol-regulated proteolysis.Cell 1994;77:53–62.
26. Haq F, Mahoney M, Koropatnick J. Signaling events for metallothioneininduction. Mutat Res 2003;533:211–226.
27. Hilgendorff A, Muth H, Parviz B, Staubitz A, Haberbosch W, Tillmanns H etal. Statins differ in their ability to block NF-kappaB activation in humanblood monocytes. Int J Clin Pharmacol Ther 2003;41:397–401.
28. Dekker RJ, van Soest S, Fontijn RD, Salamanca S, de Groot PG, VanBavel Eet al. Prolonged fluid shear stress induces a distinct set of endothelial cellgenes, most specifically lung Kruppel-like factor (KLF2). Blood 2002;100:1689–1698.
29. van Thienen JV, Fledderus JO, Dekker RJ, Rohlena J, van Ijzendoorn GA,Kootstra NA et al. Shear stress sustains atheroprotective endothelialKLF2 expression more potently than statins through mRNA stabilization.Cardiovasc Res 2006;72:231–240.
30. Dekker RJ, Boon RA, Rondaij MG, Kragt A, Volger OL, Elderkamp YW et al.KLF2 provokes a gene expression pattern that establishes functionalquiescent differentiation of the endothelium. Blood 2006;107:4354–4363.
T.T. Tuomisto et al.184
by guest on Novem
ber 20, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from




















