
Short Title: Effects of Norepinephrine In Hepatic Stellate Cells
34
G-00537-2005.R2 1 Norepinephrine Induces Calcium Spikes And Proinflammatory Actions In Human Hepatic Stellate Cells. Short Title: Effects of Norepinephrine In Hepatic Stellate Cells Pau Sancho-Bru 1 , Ramón Bataller 1 , Jordi Colmenero 1 , Xavier Gasull 2 , Montserrat Moreno 1 , Vicente Arroyo 1 , David A. Brenner 3 , Pere Ginès 1 . 1 Liver Unit, Institut Clínic de Malalties Digestives i Metabòliques, Hospital Clínic, 2 Laboratory of Neurophysiology, University of Barcelona School of Medicine, Institut d’Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS), Barcelona, Catalonia, Spain. 3 Department of Medicine, Columbia University College of Physicians and Surgeons, New York, NY. Keywords: portal hypertension, collagen, inflammation, catecholamines, liver fibrosis. Corresponding author: Ramón Bataller, M.D. Liver Unit, Hospital Clínic Villarroel 170 08036-BARCELONA Spain Phone 34-93-227.54.10 E-mail: [email protected] Page 1 of 34 Articles in PresS. Am J Physiol Gastrointest Liver Physiol (June 15, 2006). doi:10.1152/ajpgi.00537.2005 Copyright © 2006 by the American Physiological Society.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Short Title: Effects of Norepinephrine In Hepatic Stellate Cells

G-00537-2005.R2
1
Norepinephrine Induces Calcium Spikes And Proinflammatory
Actions In Human Hepatic Stellate Cells.
Short Title: Effects of Norepinephrine In Hepatic Stellate Cells
Pau Sancho-Bru1, Ramón Bataller1, Jordi Colmenero1, Xavier Gasull2, Montserrat
Moreno1, Vicente Arroyo1, David A. Brenner3, Pere Ginès1.
1Liver Unit, Institut Clínic de Malalties Digestives i Metabòliques, Hospital Clínic,
2Laboratory of Neurophysiology, University of Barcelona School of Medicine, Institut
d’Investigacions Biomèdiques August Pi i Sunyer (IDIBAPS), Barcelona, Catalonia,
Spain. 3Department of Medicine, Columbia University College of Physicians and
Surgeons, New York, NY.
Keywords: portal hypertension, collagen, inflammation, catecholamines, liver fibrosis.
Corresponding author:
Ramón Bataller, M.D.
Liver Unit, Hospital Clínic
Villarroel 170
08036-BARCELONA
Spain
Phone 34-93-227.54.10
E-mail: [email protected]
Page 1 of 34Articles in PresS. Am J Physiol Gastrointest Liver Physiol (June 15, 2006). doi:10.1152/ajpgi.00537.2005
Copyright © 2006 by the American Physiological Society.

G-00537-2005.R2
2
ABSTRACT
Background and aims: Catecholamines participate in the pathogenesis of portal
hypertension and liver fibrosis through α1 adrenoceptors. However, the underlying
cellular and molecular mechanisms are largely unknown. Here, we investigate the
effects of norepinephrine (NE) on human hepatic stellate cells (HSC), which exert
vasoactive, inflammatory and fibrogenic actions in the injured liver.
Methods: Adrenoceptor expression was assessed in human HSC by RT-PCR and
immunocytochemistry. Intracellular calcium concentration ([Ca2+]i) was studied in fura-
2 loaded cells. Cell contraction was studied by assessing wrinkle formation and myosin
light chain II (MLC-II) phosphorylation. Cell proliferation and collagen-α1(I)
expression were assessed by 3H-thymidine incorporation and quantitative PCR,
respectively. NFκB activation was assessed by luciferase reporter gene and p65 nuclear
translocation. Chemokine secretion was assessed by ELISA.
Results: Normal human livers expressed α1A adrenoceptors, which were markedly up-
regulated in livers with advanced fibrosis. Activated human HSC expressed α1A
adrenoceptors. Norepinephrine induced multiple rapid [Ca2+]i oscillations (Ca2+ spikes).
Prazosin (α1 blocker) completely prevented NE-induced Ca2+ spikes, while propranolol
(nonspecific β blocker) partially attenuated this effect. Norepinephrine caused
phosphorylation of MLC-II, and cell contraction. In constrast, NE did not affect cell
proliferation nor collagen α1(I) expression. Importantly, NE stimulated the secretion of
inflammatory chemokines (RANTES and interleukin-8) in a dose-dependent manner.
Prazosin blocked NE-induced chemokine secretion. Norepinephrine stimulated NFκB
activation. BAY11-7082, a specific NFκB inhibitor, blocked NE-induced chemokine
secretion.
Page 2 of 34

G-00537-2005.R2
3
Conclusions: NE stimulates NFκB and induces cell contraction and proinflammatory
effects in human HSC. Catecholamines may participate in the pathogenesis of portal
hypertension and liver fibrosis by targeting HSC.
Page 3 of 34

G-00537-2005.R2
4
INTRODUCTION
Catecholamines regulate a number of physiological functions in the liver. The
normal human liver contains abundant sympathetic innervation and circulating
catecholamines regulate hepatic vascular tone as well as hepatic metabolic functions
(10; 13; 43). In patients with cirrhosis, catecholamines may participate in the
pathogenesis of portal hypertension. In these patients, elevated serum catecholamines
are believed to contribute to the increased intrahepatic vascular resistance to blood flow
(4; 16). Moreover, α and β adrenoceptor blockers markedly decrease portal pressure.
The effect of nonselective β blockers is mainly mediated by a reduction in portal venous
blood flow, while the beneficial effects of α1 receptor blockers is basically due to a
reduction in intrahepatic resistance (3; 13). In rats with experimentally-induced
cirrhosis, α1 adrenoceptor agonists increase intrahepatic vascular resistance (17). We
hypothesize that α1 adrenoceptors induce vasoactive effects in contractile -
myofibroblastic- hepatic cell types, which might mediate changes in vascular tone in
advanced liver diseases.
Besides its role in the regulation of vascular tone, catecholamines seem to be
involved in the wound healing response to injury (11; 15). Locally released
catecholamines regulate key biological features of the tissue repair process, including
myofibroblast proliferation and collagen secretion (1; 2). Therefore, catecholamines are
currently viewed as a system that participates in local response to chronic injury. Recent
evidence suggests that catecholamines may participate in the pathogenesis of liver
fibrogenesis (29). Experimentally-induced liver fibrosis is enhanced in spontaneously
hypertensive rats, which have elevated serum levels of catecholamines (21). Moreover,
both chemical sympathectomy and treatment with prazosin, a α1 adrenoceptor blocker,
attenuate experimentally-induced liver fibrosis in rats (15; 30). Finally, genetic ablation
Page 4 of 34

G-00537-2005.R2
5
of the sympathetic nervous system (SNS) reduces liver fibrosis in mice (32). Moreover,
it has been shown that rodent HSC express some adrenoceptor subtypes, and that
stimulation with NE induces proliferation and collagen gene expression (32; 33).
The cell type mediating the beneficial effect of α1 adrenoceptor blockers on
portal hypertension and liver fibrosis are unknown. Hepatic stellate cells (HSC) are
potential targets for the vasoactive actions of catecholamines in the liver. Contraction of
HSC in response to vasoconstrictors (i.e. endothelin-1 and angiotensin II) are believed
to contribute to the increased resistance to blood flow in advanced chronic liver diseases
(6; 35-37). Moreover, activated HSC secrete large amounts of collagen I as well as
inflammatory mediators in chronic liver diseases, regulating fibrogenesis and
inflammation (27). We hypothesize that NE, a major catecholamine, induces vasoactive
effects on activated HSC. Besides this action, we explore whether NE also regulates the
fibrogenic and proinflammatory actions in these cells.
Page 5 of 34

G-00537-2005.R2
6
MATERIALS AND METHODS
Human liver specimens and HSC isolation. Human cirrhotic livers were
obtained from liver explants at the time of liver transplantation from patients with end-
stage liver disease due to hepatic C virus infection (N=5). Human control livers were
obtained from resections of liver metastasis of colon cancer (N=5). Human HSC were
isolated from fragments of normal livers as described in detail previously (6).
Experiments were performed with HSC activated in culture (after the second serial
passage). A subset of experiments were performed in activated non passaged HSC. The
immunocytochemistry studies were performed also in HSC freshly isolated from normal
human livers (quiescent phenotype). In all cell cultures, no staining was found for
CD45, factor VIII related-antigens, and cam 5.2 (Dako, Glostrup, Denmark), indicating
the absence of mono/macrophagic, endothelial, and epithelial cells. Cells were cultured
in standard conditions in Iscove’s Modified Dulbecco’s Medium (IMDM,
BioWhittaker, Verviers, Belgium) containing 15% fetal bovine serum. Cells were serum
starved for at least 12 hours before the experiments. The protocol was approved by the
Investigational Review Board of the Hospital Clínic of Barcelona.
Immunocytochemistry studies. Cultured HSC were fixed in methanol at -20ºC
for 10 min, blocked in PBS containing 0.1% BSA for 30 min, and incubated with anti-
p65 or anti-α1A-adrenoceptor for 1 hour (Santa Cruz Biotechnology. Santa Cruz. CA).
Cells were incubated with Cy3 or fluorescein-labeled secondary antibody for 1 hour. An
isotype-matched antibody was used as a negative control. Cells were analyzed using a
Leica TCS SL laser scanning confocal spectral microscope (Leica Microsystems
Heidelberg GmbH, Manheim, Germany) at a wavelength of 500-535 nm.
Measurement of intracellular Ca2+ concentration ([Ca++]i). Changes in [Ca2+]i
were measured in Fura-2 (Calbiochem, San Diego, CA) loaded cells using an inverted
Page 6 of 34

G-00537-2005.R2
7
epifluorescence microscope as described in detail previously (6). A representative
experiment was performed in Fura-4 loaded cells in a confocal spectral microscope.
Cells were tested with NE, prazosin and propranolol (Sigma Chemical Co, St Luis, MI).
To study Ca2+ source, experiments were performed with and without extracellular
calcium in selected groups. To deplete intracellular Ca2+ stores, cells were incubated
with thapsigargin (Sigma) for 20 minutes before NE stimulation. Cells were considered
as responders when [Ca2+]i increased more than 50% above the resting value.
Assessment of cellular contraction: The contractile responses of cultured HSC
to NE were evaluated by wrinkle formation (19; 24). A thin layer of the polydimethyl
siloxane (12.500 cP; Sigma) was spread on microscope cover slips. The silicone fluid
layer was briefly heated on a low flame from a Bunsen burner to promote cross-linking
of the silicone fluid surface while the overlying non-cross-linked fluid served as a
lubricant and allowed the surface to move independently of the cover slip. Cells plated
on the silicone rubber membrane showed basal wrinkles after 2-3 days in culture. Cells
were photographed every 5 min on an Olympus IX70 inverted phase-contrast
microscope with a digital coupled device camera (Hamamatsu, Shizuoka, Japan) and
stimulated with the agonists to assess cellular contraction by wrinkle formation. Cell
contraction was considered when there was a significant increase in the number of
wrinkles or an increase in the length of the preexisting ones.
Gene expression assays. RNA was isolated from activated HSC using Trizol
(Life Technologies Inc., Rockville, MD). PCR primer pairs were designed to amplify
GAPDH, and adrenoceptors α1A, α1B, α2A, α2B, β1, and β2 (Table 1). Quantitative PCR
was performed with pre-designed Assays-on-Demand TaqMan probes and primer pairs
for collagen α1(I) and ribosome subunit 18s. Information on these Assay-on-Demand is
available at: http://myscience.appliedbiosystems.com/cdsEntry/Form/gene_expression
Page 7 of 34

G-00537-2005.R2
8
_keyword.jsp. TaqMan reactions were carried out in duplicate on an ABI PRISM 7900
machine (Applied Biosystems, Foster City, CA).
Cell proliferation assay. DNA synthesis was estimated by methyl-3H thymidine
(Amersham Biosciences, Buckinghamshire, UK) incorporation, as described in detail
previously (7). Cells were serum starved for 24h, stimulated for 18h with agonists and
then pulsed for 6h with 1µCi/ml methyl-3H thymidine. Results are expressed as fold-
stimulation compared to cells incubated with buffer.
Determination of chemokine secretion. HSC were cultured in 6-well plates at a
density of 4x105 cells/well for 24 h. Medium was removed and cells incubated in
serum-free medium for 24 h in the presence of agonists. Supernatants were collected
and a sandwich ELISA for human interleukin-8 (IL-8) (BLK Diagnostics, Barcelona,
Spain) and RANTES (R&D Systems, Minneapolis, MN) was performed. Cells were
preincubated with SP600125, PD231445 (Sigma), and BAY11-7082 (Calbiochem,
Darmstadt, Germany) in indicated experiments. Results are expressed as fold-increase
chemokine secretion compared to treated with buffer cells.
Recombinant Adenoviral Infection. Recombinant adenoviral vectors expressing
either a luciferase reporter gene driven by NF-κB transcriptional activation (Ad5NF-
κBLuc) (39) a dominant negative mutant form (S32A/S36A) of IκB (Ad5IκB) (23), or a
control green fluorescent protein (Ad5GFP) (28) were used. HSCs were infected with
Ad5NF-κBLuc (MOI 500) and/or Ad5IκB (MOI 1,000), Ad5GFP (MOI 1,000) for 12
hours in DMEM containing 0.5% FCS. After infection, the medium was replaced with
medium containing 0.5% FCS, and the culture was continued for additional 8 hours
before performing the individual experiments.
NFκB responsive luciferase assay. HSC were infected with Ad5NF-κBLuc for
12 hours. Medium was replaced and cells stimulated with agonists for 8 hours. NFκB-
Page 8 of 34

G-00537-2005.R2
9
mediated transcriptional induction was assessed by the luciferase assay system (BD
Pharmigen, San Diego, CA). Luciferase activity (relative light units) was normalized to
the protein concentration.
Electrophoretic mobility shift assay (EMSA). Cell nuclear proteins were
extracted as described previously (8). Eight µg of nuclear proteins were incubated with
100 pg of a 32P-labeled probe containing the activating protein-1 (AP-1) consensus site
(5’-GTAAAGCATGAGTCAGACACCTC-3’) in buffer containing 10 mM HEPES pH
7.8, 2 mM MgCl2, 50 mM KCl, 1 mM DTT, 0.1 mM EDTA and 20% glycerol in the
presence of single stranded oligonucleotide (25 µg/ml) and poly dI/dC (25 µg/ml) for 20
minutes at room temperature. For competition assay, one sample was incubated with 10
ng unlabelled probe.
Western blotting. Whole cell extracts were obtained in Triton lyses buffer
containing protease and phosphatase inhibitors. Twenty-five µg were loaded onto 10%
or 15% SDS-acrylamide gels and blotted onto nitrocellulose membranes. Membranes
were then incubated with antibodies against SAPK/JNK, phospho-SAPK/JNK, p44/42
MAP Kinase (ERK), phospho-ERK, phospho myosin light chain-II (MLC-II) (Cell
Signaling, Beverly, MA) and α1A-adrenoceptor (Santa Cruz Biotechnology). After
extensive washing, membranes were incubated with blocking buffer containing
horseradish-peroxidase conjugated secondary antibody. Proteins were detected by ECL
chemoluminescence (Amersham).
Data analysis. Data presented herein are representative of at least three
experiments. Results are expressed as mean ± SD. Statistical analysis was performed by
Student's t test and analysis of variance.
Page 9 of 34

G-00537-2005.R2
10
RESULTS
α1A adrenergic receptors are expressed in human HSC and are up-regulated
in human fibrotic livers. We first assessed the expression of adrenoceptors in human
HSC by RT-PCR. Human cultured HSC express α1A, α2B, and β2 adrenoceptors (Figure
1A). In contrast, α1B, α2A and β1 adrenoceptors were not expressed in HSC but were
detected in RNA obtained from total human liver. Because α1 receptors mediate key
biological effects of the SNS in fibrogenic cell types (15; 38), we further investigated
their expression in HSC as well as in normal and fibrotic human liver specimens. Both
quiescent and culture-activated HSC contain α1A receptors, as assessed by
immunocytochemistry (Figure 1B, 1C and 1D). Moreover, western blotting analysis of
whole liver extracts revealed that normal human livers express α1A receptors, which are
markedly up-regulated in livers with advanced fibrosis (Figure 2). These results suggest
that α1A receptors are up-regulated during liver fibrogenesis and that HSC are potential
targets for adrenergic agonists and antagonist.
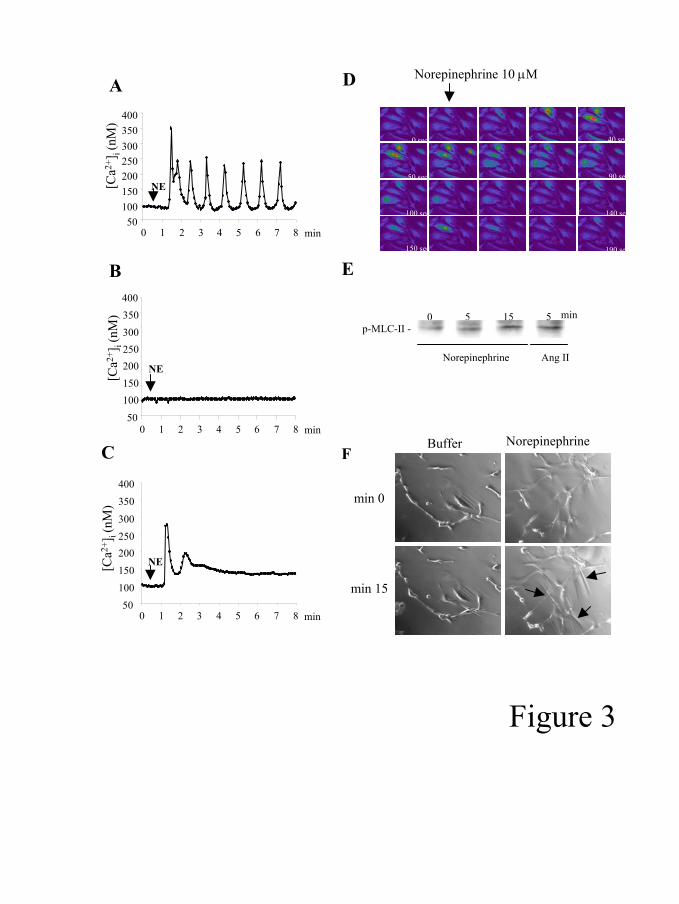
Norepinephrine induces Ca2+ spikes and cell contraction in human HSC. We
next stimulated HSC with NE, a major neurotransmitter of the SNS. Stimulation of cells
with NE (10 µM) increased [Ca2+]i in Fura-2 loaded HSC (table 2 and figure 3A and
3D). The typical pattern was characterized by an abrupt increase of [Ca2+]i followed by
an oscillatory response in 57% of the cells (Ca2+ spikes). Ten µM NE induced a
significant increase of [Ca2+]i in 84% of the cells and a mean Ca2+ peak of 317±138 nM.
Cell stimulation with 1 and 0.1 µM NE also induced a marked [Ca2+]i increase (not
shown). Removal of Ca2+ from the extracellular solution did not affect NE-induced Ca2+
response. Moreover, cell preincubation with thapsigargin, a cell-permeable SERCA
pump inhibitor that depletes Ca2+ intracellular stores, completely abolished NE-induced
Ca2+ response (data not shown). These results indicate NE mainly releases Ca2+ from
Page 10 of 34

G-00537-2005.R2
11
intracellular stores. Prazosin completely prevented NE-induced [Ca2+]i increase in all
cells (Table 2 and figure 3B). However, preincubation of cells with propranolol only
attenuated NE-induced [Ca2+]i peak and the percentage of cells showing an oscillatory
response (Figure 3C). Similar results were obtained with non-passaged cells activated in
culture (data not shown) We next explored whether NE induces cell contraction. NE
induced MLC-II phosphorylation after 5 min, which is a known prerequisite for cell
contraction (20; 44) (Figure 3E). Preincubation with 10µM of BAPTA-AM did not
significantly modify MLC-II phosphorylation, suggesting that it could be calcium-
independent (data not shown). Moreover, we found that stimulation with NE induced
cell contraction, as assessed by wrinkle formation (Figure 3F). Taken together, these
results indicate that NE induces cell contraction and may be involved in the vasoactive
effects of HSC.
Norepinephrine does not stimulate proliferation of human HSC. We
investigated whether NE regulates human HSC growth. Different concentrations of NE
(10nM - 10µM) were tested. Incubation with NE did not affect DNA synthesis, as
assessed by 3H-thymidine incorporation (Figure 4A). Stimulation with phenilephrine or
terbutaline (an α and β receptor agonist, respectively) did not affect cell proliferation
(data not shown). To elucidate whether NE regulates agonist-induced cell proliferation,
HSC were pretreated with NE (10 µM) and then challenged with platelet derived growth
factor-BB (PDGF-BB) (20 ng/ml), a potent mitogen for these cells. NE pretreatment did
not affect PDGF-induced mitogenic effect. These results indicate that NE is not
mitogenic for human HSC.
Effect of norepinephrine on gene expression of collagen α1(I) in human HSC.
We next investigated whether NE regulates the expression of collagen α1(I), the main
extracellular protein found in fibrotic human livers. Stimulation of cells with NE (10
Page 11 of 34

G-00537-2005.R2
12
µM) did not affect collagen expression, as assessed by quantitative PCR (Figure 4B).
These results suggest that NE does not directly regulate collagen synthesis in human
activated HSC.
Norepinephrine stimulates chemokine production by human HSC. We next
investigated whether NE regulates the inflammatory actions of HSC by stimulating
chemokine secretion. Norepineprhine (10 µM) induced a significant increase in
secretion of IL-8 and RANTES to the culture media, which are chemokines potentially
involved in liver fibrogenesis (34; 40) (Figure 5A). Preincubation of cells with prazosin
markedly reduced both IL-8 and RANTES secretion. Propranolol also attenuated NE-
induced chemokine production, yet only RANTES secretion was significantly reduced.
These results suggest that the inflammatory actions of NE on human HSC could be
mediated by both α1 and β adrenoceptors.
Effect of NE on intracellular signaling pathways in human HSC. We finally
explored the effect of NE on intracellular pathways known to regulate the inflammatory
actions in human HSC (8; 41). These pathways include mitogen-activated protein
kinases (MAPK) and the transcription factors NFκB and AP-1. NE (10 µM) induced a
transient increase in ERK and SAPK/JNK phosphorylation, as assessed by western
blotting (Figure 6A). NE activated NFκB, as indicated by increas NFκB-dependent
gene expression as well as nuclear translocation of the heterodimer p50/p65 (Figure 6B
and 6C). In addition, NE increased tumor necrosis factor α (TNF-α)-induced NFκB
activation, suggesting that it could amplify the inflammatory response evoked by other
cytokines (data not shown). Finally, NE stimulated AP-1 DNA binding, as assessed by
EMSA (Figure 6D). To elucidate the role of these signaling pathways on chemokine
production, HSC were preincubated with specific antagonists before NE stimulation.
Preincubation of cells with BAY11-7082 (a NFκB inhibitor), but not with SP600125 (a
Page 12 of 34

G-00537-2005.R2
13
JNK inhibitor) and PD231445 (an ERK inhibitor), attenuated IL-8 secretion (Figure.
5B). Moreover, transfection with Ad5IκB reduced IL-8 production confirming NF-κB
involvement (Figure. 5C). These data strongly indicate that NE exerts pro-inflammatory
actions in human HSC through NFκB activation.
Page 13 of 34

G-00537-2005.R2
14
DISCUSSION
In the current study, we investigate the biological effects of NE in human HSCs.
We provide evidence that HSC express some α and β receptor subtypes.
Norepinephrine stimulates key intracellular signaling pathways in these cells and
induces cell contraction and proinflammatory actions. These actions are mainly
mediated by α1 receptors, although β receptors also modulate some of the effects
evoked by NE. Moreover, α1 receptors are markedly up-regulated in human fibrotic
livers. These results suggest that HSC are potential targets for the pathogenic effect of
catecholamines in the liver and in keeping with recent data indicating that
catecholamines not only regulate vascular homeostasis, but also contribute to tissue
repair and inflammation (11; 15; 30; 31).
The hallmark biological action of NE in vascular cell types is to induce Ca2+
increase and cell contraction. This effect has been extensively studied in mesangial cells
and vascular smooth muscle cells (18; 26). Similarly, a large number of studies have
investigated the vasoactive effects of vasoconstrictors in cultured human HSC,
including angiotensin II, endothelin-1, thrombin and leukotriene D4 (36). Currently,
studies using activated HSC are widely used to investigate the cellular basis of portal
hypertension in order to identify potential targets for therapy. Surprisingly, little
information is available on the actions of adrenergic agonists in these cells. Here, we
provide evidence that NE is a potent agonist for human HSC to induce [Ca2+]i increase.
An intriguing finding of the current study is that the Ca2+ response pattern evoked by
NE markedly differ to that observed with other vasoconstrictors. Previously, we have
shown that angiotensin II induces an abrupt [Ca2+]i increase followed by a sustained
phase (6). This effect was mainly due to the entrance of Ca2+ from the extracellular
space through voltage-operated calcium channels. Interestingly, the effect of NE on
Page 14 of 34

G-00537-2005.R2
15
[Ca2+]i consists of numerous Ca2+ spikes. Baseline Ca2+ spikes are characterized by
rapidly-rising transient increases in [Ca2+]i rising from a baseline which is close to the
resting concentration (9). This pattern of response has been described in many cells in
response to hormones and neurotransmitters (18; 25). Ca2+ oscillations permit cells to
respond to agonists without being exposed to sustained levels of [Ca2+]i. It has been
shown that oscillations and specially their frequency activate cell signaling like
calmodulin-dependent protein kinase II and NFκB (14; 22). Moreover, Ca2+ oscillations
have been involved in many cellular processes like contraction, migration, cell secretion
or phagocytosis (9; 12). The biological relevance of NE-induced Ca2+ spikes in HSC is
unknown. Although NE induced MLC-II phosphorylation, a prerequisite for cell
contraction, and induced cell contraction, it failed to induce cell proliferation, which is
commonly stimulated by vasoconstrictors in HSC. It is possible that sustained [Ca2+]i
increase, rather that [Ca2+]i oscillations, are required to induce cell proliferation. Further
studies should investigate the pathophysiological relevance of agonist-induced Ca2+
spikes in HSC.
A relevant finding of the current study is that NE stimulates key intracellular
signaling pathways implicated in the pathogenesis of hepatic inflammation and
fibrogenesis. These pathways include NFκB, JNK/AP-1 and ERK. Importantly, NE
enhances chemokine secretion in a NFκB-dependent manner. Besides their role in
collagen synthesis, recent data suggest that activated HSC play a supportive role in the
inflammatory response to injury (27). These cells proliferate at the areas of liver injury
and a cross-talk between HSC and infiltrating leukocytes is likely to occur. HSC can
present antigens, express cell adhesion molecules, regulate lymphocyte proliferation and
secrete powerful chemokines such as IL-8 and RANTES (27; 42). On the other hand,
mediators derived from lymphocytes (e.g. CD40 ligand, RANTES) can regulate HSC
Page 15 of 34

G-00537-2005.R2
16
biological functions and stimulate their fibrogenic potential (40; 41). Therefore, by
stimulating chemokine secretion, NE may enhance inflammation in chronic liver
diseases. Whether NE also exerts proinflammatory actions in other non-parenchymal
hepatic cells types (i.e. Kupffer cells and sinusoidal endothelial cells) is unknown and
deserves further investigation.
In the current study, despite the possible proinflammatory effect of NE, we did
not find a direct fibrogenic effect of NE on collagen expression. In rodent early-cultured
HSC, NE stimulates collagen secretion, suggesting that it directly induces fibrogenic
effects in these cells (32; 33). These discrepant results can be explained by the fact that
we used passaged fully-activated human HSC. In this cell phenotype, collagen is highly
expressed and fibrogenic agonists commonly fail to further increase its expression.
Other biological functions, such as chemokine secretion, can be stimulated by
inflammatory mediators (40; 41). However, it is well-known that liver inflammation
precedes and promotes the progression of liver fibrosis and there is a positive
correlation between the degree of sustained hepatic inflammation and the progression of
fibrosis. Moreover, substances inhibiting the inflammatory response of the liver also
limit fibrogenesis (5). Therefore, we speculate that the proinflammatory effect of NE on
HSC may play a role in promoting hepatic inflammation in patients with chronic liver
diseases, thus favoring fibrosis development.
The biological effects of NE in HSC may have pathophysiological
consequences. First, contraction of activated HSC are believed to increase intrahepatic
vascular resistance in the cirrhotic liver and contribute to the pathogenesis of portal
hypertension (37). The finding that NE induces contraction in HSC may explain, at least
in part, the effects of catecholamines in regulating intrahepatic vascular resistance in
cirrhosis. Moreover, the fact that HSC synthesize NE suggest that not only an endocrine
Page 16 of 34

G-00537-2005.R2
17
pathways may regulate HSC contractility but also an autocrine/paracrine system may be
involved (32). Further studies should study this hypothesis. Our finding that NE
enhances chemokine secretion in HSC suggests that catecholamines may exert
proinflammatory effects in the liver. The pro inflammatory effect of NE on HSC has not
been previously described in other cell types. The finding that NE may promotes
proinflammatory actions in HSC expands the traditional view of the SNS as a system
that basically regulates systemic and hepatic hemodynamics.
In summary, the results of the current study indicate that NE mediates vasoactive
and proinflammatory actions in human HSC mainly through α1 adrenoceptors. Because
HSC are able to synthesize adrenergic agonists and the SNS is activated in patients with
advanced liver fibrosis, these findings suggest that catecholamines may increase
intrahepatic vascular resistance and promote inflammatiory actions by targeting HSC.
Our data explain, at least in part, the mechanisms underlying the beneficial effects of α1
receptor antagonists in the treatment of portal hypertension and suggest the potential use
of these drugs to modulate hepatic inflammation and subsequent fibrogenesis.
Page 17 of 34

G-00537-2005.R2
18
Table 1. PCR primer pairs for adrenoceptors and housekeeping genes. Gene Sense Antisense Product (bp)GADPH 5’-CCATGGAGAAGGCTGGGG-3’ 5’-CAAAGTTGTCATGGATGACC-3’ 198 α1A- 5’-TGGCCGACCTCCTGCTCACCTC-3’ 5’-GGCCCCGGCTTTCCCTCTTG-3’ 444 α1B- 5’-CCCCCGACGCCGTGTTCAAGGTG-3’ 5’-CTCAGGCGCGGGCAGGCTCAGGA-3’ 407 α2A- 5’-GCCGGCCTGCTCATGCTGCTCAC-3’ 5’-GCCGCCGCCGCCCTTCTTCTC-3’ 420 α2B- 5’-CAGGCCGATGGAGGTGGTTCAAAT-3’ 5’-CCGGAGGGGTGCTGGGTAAGGA-3’. 423 β1- 5’-GTGGCCCTACGCGAGCAGAA-3’ 5’-GCGGCAGTAGATGATGGGGTTGAA-3’ 195 β2- 5’-TCTGGCCGCGTTTCTGTGTTGG-3’ 5’-GGCCCCGGCTTTCCCTCTTG-3’ 295
Page 18 of 34

G-00537-2005.R2
19
Table 2. Effect of norepinephrine (NE) on [Ca2+]i in human hepatic stellate cells n Responder
cells (%)a Peak [Ca2+]i
(nM)b Oscillations (% of cells)
NE 10 µM 51 84 317±138 57
Prazosin 1 µM + NE 10 µM 27 0 - - Propranolol 1 µM + NE 10 µM 69 79 260±170* 36* aResponder cells are those showing at least a 1.5 fold increase in [Ca2+]i compared to resting values. bResponders cells only. Values are given as mean ± SEM. *p<0.05 vs cells stimulated with NE.
Page 19 of 34

G-00537-2005.R2
20
Reference List
1. Akiyama-Uchida Y, Ashizawa N, Ohtsuru A, Seto S, Tsukazaki T, Kikuchi H, Yamashita S and Yano K. Norepinephrine enhances fibrosis mediated by TGF-beta in cardiac fibroblasts. Hypertension 40: 148-154, 2002.
2. Akiyoshi H and Terada T. Mast cell, myofibroblast and nerve terminal complexes in carbon tetrachloride-induced cirrhotic rat livers. J Hepatol 29: 112-119, 1998.
3. Albillos A, Lledo JL, Rossi I, Perez-Paramo M, Tabuenca MJ, Banares R, Iborra J, Garrido A, Escartin P and Bosch J. Continuous prazosin administration in cirrhotic patients: effects on portal hemodynamics and on liver and renal function. Gastroenterology 109: 1257-1265, 1995.
4. Ballet F, Chretien Y, Rey C and Poupon R. Differential response of normal and cirrhotic liver to vasoactive agents. A study in the isolated perfused rat liver. J Pharmacol Exp Ther 244: 283-289, 1988.
5. Bataller R and Brenner DA. Hepatic stellate cells as a target for the treatment of liver fibrosis. Semin Liver Dis 21: 437-451, 2001.
6. Bataller R, Gines P, Nicolas JM, Gorbig MN, Garcia-Ramallo E, Gasull X, Bosch J, Arroyo V and Rodes J. Angiotensin II induces contraction and proliferation of human hepatic stellate cells. Gastroenterology 118: 1149-1156, 2000.
7. Bataller R, Nicolas JM, Gines P, Esteve A, Nieves GM, Garcia-Ramallo E,
Pinzani M, Ros J, Jimenez W, Thomas AP, Arroyo V and Rodes J. Arginine vasopressin induces contraction and stimulates growth of cultured human hepatic stellate cells. Gastroenterology 113: 615-624, 1997.
8. Bataller R, Schwabe RF, Choi YH, Yang L, Paik YH, Lindquist J, Qian T, Schoonhoven R, Hagedorn CH, Lemasters JJ and Brenner DA. NADPH oxidase signal transduces angiotensin II in hepatic stellate cells and is critical in hepatic fibrosis. J Clin Invest 112: 1383-1394, 2003.
9. Berridge MJ, Bootman MD and Roderick HL. Calcium signalling: dynamics,
homeostasis and remodelling. Nat Rev Mol Cell Biol 4: 517-529, 2003.
10. Bioulac-Sage P, Lafon ME, Saric J and Balabaud C. Nerves and perisinusoidal cells in human liver. J Hepatol 10: 105-112, 1990.
Page 20 of 34

G-00537-2005.R2
21
11. Bonnefont-Rousselot D, Mahmoudi A, Mougenot N, Varoquaux O, Le Nahour G, Fouret P and Lechat P. Catecholamine effects on cardiac remodelling, oxidative stress and fibrosis in experimental heart failure. Redox Rep 7: 145-151, 2002.
12. Bootman MD, Lipp P and Berridge MJ. The organisation and functions of local Ca(2+) signals. J Cell Sci 114: 2213-2222, 2001.
13. Burt AD, Tiniakos D, MacSween RN, Griffiths MR, Wisse E and Polak JM. Localization of adrenergic and neuropeptide tyrosine-containing nerves in the mammalian liver. Hepatology 9: 839-845, 1989.
14. De Koninck P and Schulman H. Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations. Science 279: 227-230, 1998.
15. Dubuisson L, Desmouliere A, Decourt B, Evade L, Bedin C, Boussarie L, Barrier L, Vidaud M and Rosenbaum J. Inhibition of rat liver fibrogenesis through noradrenergic antagonism. Hepatology 35: 325-331, 2002.
16. Esler M, Dudley F, Jennings G, Debinski H, Lambert G, Jones P, Crotty B, Colman J and Willett I. Increased sympathetic nervous activity and the effects of its inhibition with clonidine in alcoholic cirrhosis. Ann Intern Med 116: 446-455, 1992.
17. Graupera M, Garcia-Pagan JC, Abraldes JG, Peralta C, Bragulat M, Corominola H, Bosch J and Rodes J. Cyclooxygenase-derived products modulate the increased intrahepatic resistance of cirrhotic rat livers. Hepatology 37: 172-181, 2003.
18. Hajjar RJ and Bonventre JV. Oscillations of intracellular calcium induced by vasopressin in individual fura-2-loaded mesangial cells. Frequency dependence on basal calcium concentration, agonist concentration, and temperature. J Biol Chem 266: 21589-21594, 1991.
19. Harris AK, Wild P and Stopak D. Silicone rubber substrata: a new wrinkle in the study of cell locomotion. Science 208: 177-179, 1980.
20. Hirano K, Hirano M and Kanaide H. Regulation of myosin phosphorylation and myofilament Ca(2+) sensitivity in vascular smooth muscle. J Smooth Muscle Res 40: 219-236, 2004.
21. Hsu CT. The role of the sympathetic nervous system in promoting liver cirrhosis induced by carbon tetrachloride, using the essential hypertensive animal (SHR). J Auton Nerv Syst 37: 163-173, 1992.
Page 21 of 34

G-00537-2005.R2
22
22. Hu Q, Deshpande S, Irani K and Ziegelstein RC. [Ca(2+)](i) oscillation frequency regulates agonist-stimulated NF-kappaB transcriptional activity. J Biol Chem 274: 33995-33998, 1999.
23. Iimuro Y, Nishiura T, Hellerbrand C, Behrns KE, Schoonhoven R, Grisham JW and Brenner DA. NFkappaB prevents apoptosis and liver dysfunction during liver regeneration. J Clin Invest 101: 802-811, 1998.
24. Kawada N, Klein H and Decker K. Eicosanoid-mediated contractility of hepatic stellate cells. Biochem J 285: 367-371, 1992.
25. Kawanishi T, Blank LM, Harootunian AT, Smith MT and Tsien RY. Ca2+ oscillations induced by hormonal stimulation of individual fura-2-loaded hepatocytes. J Biol Chem 264: 12859-12866, 1989.
26. Koshimizu TA, Tanoue A, Hirasawa A, Yamauchi J and Tsujimoto G. Recent advances in alpha1-adrenoceptor pharmacology. Pharmacol Ther 98: 235-244, 2003.
27. Marra F. Hepatic stellate cells and the regulation of liver inflammation. J Hepatol 31: 1120-1130, 1999.
28. Moriyoshi K, Richards LJ, Akazawa C, O'Leary DD and Nakanishi S. Labeling neural cells using adenoviral gene transfer of membrane-targeted GFP. Neuron 16: 255-260, 1996.
29. Oben JA and Diehl AM. Sympathetic nervous system regulation of liver repair. Anat Rec A Discov Mol Cell Evol Biol 280: 874-883, 2004.
30. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Li Z, Torbenson M, Huang J, Guarino P, Kafrouni M and Diehl AM. Sympathetic nervous system inhibition increases hepatic progenitors and reduces liver injury. Hepatology 38: 664-673, 2003.
31. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Li Z, Torbenson M, Thomas SA and Diehl AM. Norepinephrine induces hepatic fibrogenesis in leptin deficient ob/ob mice. Biochem Biophys Res Commun 308: 284-292, 2003.
32. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Torbenson M, Smedh U, Moran TH, Li Z, Huang J, Thomas SA and Diehl AM. Hepatic fibrogenesis requires sympathetic neurotransmitters. Gut 53: 438-445, 2004.
Page 22 of 34

G-00537-2005.R2
23
33. Oben JA, Yang S, Lin H, Ono M and Diehl AM. Norepinephrine and neuropeptide Y promote proliferation and collagen gene expression of hepatic myofibroblastic stellate cells. Biochem Biophys Res Commun 302: 685-690, 2003.
34. Pinzani M and Marra F. Cytokine receptors and signaling in hepatic stellate cells. Semin Liver Dis 21: 397-416, 2001.
35. Pinzani M, Milani S, De Franco R, Grappone C, Caligiuri A, Gentilini A, Tosti-Guerra C, Maggi M, Failli P, Ruocco C and Gentilini P. Endothelin 1 is overexpressed in human cirrhotic liver and exerts multiple effects on activated hepatic stellate cells. Gastroenterology 110: 534-548, 1996.
36. Reynaert H, Thompson MG, Thomas T and Geerts A. Hepatic stellate cells: role in microcirculation and pathophysiology of portal hypertension. Gut 50: 571-581, 2002.
37. Rockey D. The cellular pathogenesis of portal hypertension: stellate cell contractility, endothelin, and nitric oxide. Hepatology 25: 2-5, 1997.
38. Saeed AE, Parmentier JH and Malik KU. Activation of alpha1A-adrenergic receptor promotes differentiation of rat-1 fibroblasts to a smooth muscle-like phenotype. BMC Cell Biol 5: 47, 2004.
39. Sanlioglu S, Williams CM, Samavati L, Butler NS, Wang G, McCray PB, Jr., Ritchie TC, Hunninghake GW, Zandi E and Engelhardt JF. Lipopolysaccharide induces Rac1-dependent reactive oxygen species formation and coordinates tumor necrosis factor-alpha secretion through IKK regulation of NF-kappa B. J Biol Chem 276: 30188-30198, 2001.
40. Schwabe RF, Bataller R and Brenner DA. Human hepatic stellate cells express CCR5 and RANTES to induce proliferation and migration. Am J Physiol Gastrointest Liver Physiol 285: G949-G958, 2003.
41. Schwabe RF, Schnabl B, Kweon YO and Brenner DA. CD40 activates NF-kappa B and c-Jun N-terminal kinase and enhances chemokine secretion on activated human hepatic stellate cells. J Immunol 166: 6812-6819, 2001.
42. Vinas O, Bataller R, Sancho-Bru P, Gines P, Berenguer C, Enrich C, Nicolas JM, Ercilla G, Gallart T, Vives J, Arroyo V and Rodes J. Human hepatic stellate cells show features of antigen-presenting cells and stimulate lymphocyte proliferation. Hepatology 38: 919-929, 2003.
Page 23 of 34

G-00537-2005.R2
24
43. vom DS, Bode JG, Reinehr RM, Monnighoff I, Kubitz R and Haussinger D. Release of osmolytes from perfused rat liver on perivascular nerve stimulation: alpha-adrenergic control of osmolyte efflux from parenchymal and nonparenchymal liver cells. Hepatology 29: 195-204, 1999.
44. Yanase M, Ikeda H, Ogata I, Matsui A, Noiri E, Tomiya T, Arai M, Inoue Y, Tejima K, Nagashima K, Nishikawa T, Shibata M, Ikebe M, Rojkind M and Fujiwara K. Functional diversity between Rho-kinase- and MLCK-mediated cytoskeletal actions in a myofibroblast-like hepatic stellate cell line. Biochem Biophys Res Commun 305: 223-228, 2003.
Page 24 of 34

G-00537-2005.R2
25
FIGURE LEGENDS
Figure 1. Expression of adrenoceptors in human hepatic stellate cells (HSC). α1A,
α2B, and β2 receptors were detected in HSC and whole liver, as assessed by RT-PCR.
α1B, α2A and β1 receptor were not detected in HSC but were amplified in mRNA
isolated from human livers (A). GAPDH was amplified as a housekeeping gene to
assess correct amplification (data not shown). Detection of α1A receptor in HSC freshly
isolated from normal human livers (quiescent) (B). Green staining shows α1A receptor
expression. Detection of α1A receptor in HSC activated after prolonged culture (C) by
immunocytochemistry. Note α1A adrenoceptor expression at cell membrane (arrow). A
negative control using a isotype-matched primary antibody was performed (C).
Figure 2. Expression of α1A adrenoceptors in normal and cirrhotic human livers.
Western blot analysis of 50 µg proteins from normal and fibrotic human livers. α1A
adrenoceptors were detected in normal livers and were overexpressed in cirrhotic livers
(51.5 KDa). Actin expression was performed by Western blotting to ensure equal
protein loading (A). Analysis of α1A receptor expression by densitometry respect to
actin (B). **p<0.01 vs normal livers.
Figure 3. Effects of norepinephrine (NE) on [Ca2+]i and cell contraction in hepatic
stellate cells (HSC). Representative graph showing the effect of NE in a single HSC.
Norepinephrine (10µM) induced an oscillatory [Ca2+]i increase (Ca2+ spikes) (A).
Preincubation with prazosin (1 µM) for 10 min blocked NE-induced [Ca2+]i increase
(B). Incubation for 10 min with propranolol (1 µM) decreased both Ca2+ peak and
percentage of cells with oscillations (C). Representative images of [Ca2+]i increase
induced by NE (10µM) in HSC (D). Images were obtained using a confocal microscope
in fluo-4 loaded HSC. Images were taken every 10 seconds. Color change into red
Page 25 of 34

G-00537-2005.R2
26
reveals [Ca2+]i increase. Phosphorylation of myosin light chain-II (MLC-II), as assessed
by western blotting (18 KDa). Stimulation with NE (10 µM) or Ang II induced MLC-II
phosphorylation at 5 min (E). Representative images of wrinkle formation (arrows)
formed by HSC stimulated with NE (10 µM) (F).
Figure 4. Effect of norepinephrine (NE) on cell growth and collagen gene
expression. Effect of NE (10µM) on cell growth, as assessed by 3H-thymidine
incorporation (A). Hepatic stellate cells were treated for 24h with NE (100µM-10nM),
platelet derived growth factor-BB (PDGF-BB, 20 ng/ml) or both. PDGF-BB induced a
marked increased in DNA synthesis (*p<.001 vs untreated cells). Norepinephrine did
not stimulate cell growth nor amplified PDGF-induced mitogenic effect. Effect of NE
(10 µM) on procollagen α1(I) gene expression, as assessed by quantitative PCR (B).
Figures are the mean of 5 independent experiments.
Figure 5. Effect of norepineprhine (NE) on chemokine secretion in human hepatic
stellate cells (HSC). (A) Norepinephrine stimulated the secretion of interleukin-8 (IL-8)
(■) and RANTES (□) to the culture media, as assessed by specific ELISA. Incubation
with prazosin prevented NE-induced chemokine secretion. Propranolol reduced NE-
induced RANTES secretion. (B) Preincubation with BAY11-7082 (a NFκB inhibitor),
but not with SP600125 (a JNK inhibitor) and PD231445 (a ERK inhibitor) prevented
NE induced IL-8 secretion. (C) Ad5IκB reduced IL-8 production induced by NE.
*p<0.05 vs untreated cells Ad5GFP was used as a control. #p<0.05 vs cells treated with
NE alone (n=5).
Figure 6. Effect of norepineprhine (NE) on intracellular signaling pathways in
human hepatic stellate cells (HSC). (A) Effect of NE on ERK and SAPK/JNK
phosphorylation, as assessed by western blotting. Stimulation with NE induced a
transient phosphorylation of ERK and SAPK/JNK. Figures are representative of three
Page 26 of 34

G-00537-2005.R2
27
independent experiments. (B) NFκB dependent gene expression, as assessed by a
luciferase assay. Norepinephrine stimulated NFκB-dependent gene expression that was
completely blunted by prazosin, but not by propranolol. *p<0.05 vs untreated cells.
#p<0.05 vs cells treated with NE alone. Results are the mean of 4 independent
experiments. (C) Immunocytochemistry analysis of the p65 subunit of NFκB in HSC.
Stimulation for 60 min with NE (10µM) resulted in the nuclear translocation of the
heterodimer p50/p65 to the nucleus, which is indicative of NFκB activation. TNFα (10
ng/ml) was used as a positive control. (D) Activation of the transcription factor AP-1, as
assessed by EMSA. Stimulation with NE (10 µM) for 60 min markedly stimulated AP-1
DNA binding. Preincubation with cold probe was performed to ensure specificity. This
figure is representative of 3 independent experiments.
Page 27 of 34

G-00537-2005.R2
28
Abbreviations.
AP-1, activating protein-1; EMSA, electrophoretic mobility shift assay; ERK,
extracellular regulated kinase; HSC, hepatic stellate cells; IL-8, interleukin-8; [Ca2+]i,
intracellular calcium concentration; JNK, Jun-N-terminal kinase; MAPK, mitogen-
activated protein kinase; MLC-II, myosin light chain-II; NFκB, nuclear factor kappa B;
NE, norepinephrine; PDGF, platelet derived growth factor-BB;SNS, sympathetic
nervous system; TNF-α, tumor necrosis factor α.
Acknowledgements.
We thank Joan Carles Garcia-Pagán for helpful discussion, Elena Juez and Cristina
Millán for her excellent technical support. We thank Maria Calvo and Anna Bosch
(Serveis Cientificotècnics, Universitat de Barcelona- IDIBAPS) for excellent technical
assistance in in vivo confocal microscopy. This work is supported by grants from the
NIH (R01 AA15055), from the Ministerio de Ciencia y Tecnología, Dirección General
de Investigación (SAF2002-03696), and from the Instituto de Salud Carlos III
(CO3/02). Pau Sancho-Bru had a grant from the Institut d’Investigacions Biomèdiques
August Pi i Sunyer (IDIBAPS). Montserrat Moreno had a grant from the Instituto Reina
Sofía de Investigación Nefrológica.
Page 28 of 34

A
B
- β1
- α2A
- α1B
α2B -
α1A -
β2 -
Whole liver
ActivatedHCS
C
D
Whole liver
ActivatedHCS
Figure 1
Page 29 of 34

Normal livers Cirrhotic livers
α1A-
A
B
Normal livers Cirrhotic livers0
25
50
75
100 **
α -1 A
rece
ptor
expr
essi
on(a
rbitr
ary
units
)
actin -
Figure 2
Page 30 of 34
A[C
a2+] i (
nM)
0 2 3 4 5 6 7 81 min50
100150200250300350400
NE
0 2 3 4 5 6 7 81 min50
100150200250300350400
NE[Ca2+
] i (nM
)
B
C
50100150200250300
350400
0 2 3 4 5 6 7 81 min
NE[Ca2+
] i (nM
)
Norepinephrine 10 µMD
0 sec 40 sec
50 sec 90 sec
100 sec 140 sec
190 sec150 sec
E
p-MLC-II -
Norepinephrine
5 150 min5
Ang II
FNorepinephrineBuffer
min 0
min 15
Figure 3
Page 31 of 34

02468
101214 *
3 H-T
hym
idin
e up
take
(F
old
incr
ease
of c
pm)
+
+
-
-
-
+
+
-
PDGF 20 ng/ml
NE 10 µM
A
Propranolol 1 µM
Prazosin 1 µMNE 10 µM
+
-+
-
--
-
-+
-
++
0.5
0.7
0.9
1.1
1.3
1.5
Nor
mal
ize d
co
llage
n m
RN
A
B
Figure 4
Page 32 of 34

0.00.51.01.52.02.53.03.54.0 * *
#
##
Cyt
okin
e s e
c re t
ed le
vels
(f
old
s tim
u lat
ion)
Propranolol 1 µM
Prazosin 1 µM
NE 10 µM
+
-
+
-
+
+
-
-
+
-
-
+
-
-
-
-
-
-
+-
-+
++
AInterleukin-8
RANTES
PD231445 20 µM
SP600125 20 µM
BAY11-7082 2 µMNE 10 µM
+
-
-+
+---
-
++
-
--
-
-+
-
-+
0.3
0.8
1.3
1.8
2.3
2.8*
#
Cyt
okin
e s e
c re t
ed le
vels
(fol
d s t
imu l
atio
n)
B
0200400
600800
10001200
Cyt
okin
e s e
c re t
edle
vels
(pg/
ml )
Ad5IκBadenovirus
Ad5GFPadenovirus
NE 10 µM
+
-
+
-
+
+
C
*
Figure 5
Page 33 of 34

A
0 min
15 m
in30
min
1h
p-ERK-
ERK-
0 min
15 m
in
30 m
in1h
P-SAPK/JNK-
SAPK/JNK-
C NECold
prob
eB
+
-
+
-
-
-
-
-
+
-
+
+
Propranolol 1 µM
Prazosin 1 µM
NE 10 µM0.6
0.8
1.01.2
1.41.6
1.82.0
#
*
NF-κB
ac t
i vit y
(luci
fer a
s e a
r bi tr
a ry
unit s
)
D
AP-1
C
Buffer NE 10 µM TNF-α
Figure 6
Page 34 of 34




















