
Sensory map transfer to the neocortex relies on pretarget ordering of thalamic axons
7
Current Biology 23, 810–816, May 6, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.03.062 Report Sensory Map Transfer to the Neocortex Relies on Pretarget Ordering of Thalamic Axons Ludmilla Lokmane, 1,2,3 Re ´ mi Proville, 1,2,3 Nicolas Narboux-Ne ˆ me, 4,5,6 Ildiko Gyo ¨ ry, 7 Maryama Keita, 1,2,3 Caroline Mailhes, 1,2,3 Cle ´ment Le ´na, 1,2,3 Patricia Gaspar, 4,5,6 Rudolf Grosschedl, 7 and Sonia Garel 1,2,3, * 1 Ecole Normale Supe ´ rieure, Institut de Biologie de l’ENS (IBENS), 46 Rue d’Ulm, 75005 Paris, France 2 INSERM, U1024, 75005 Paris, France 3 CNRS, UMR 8197, 75005 Paris, France 4 INSERM, UMR-S 839, 17 Rue du Fer a ` Moulin, 75005 Paris, France 5 Universite ´ Pierre et Marie Curie, 75005 Paris, France 6 Institut du Fer a ` Moulin, 75005 Paris, France 7 Department of Cellular and Molecular Immunology, Max Planck Institute of Immunobiology, Stuebeweg 51, 79108 Freiburg, Germany Summary Sensory maps, such as the representation of mouse facial whiskers, are conveyed throughout the nervous system by topographic axonal projections that preserve neighboring relationships between adjacent neurons [1]. In particular, the map transfer to the neocortex is ensured by thalamocort- ical axons (TCAs), whose terminals are topographically organized in response to intrinsic cortical signals [2–5]. However, TCAs already show a topographic order early in development, as they navigate toward their target [6, 7]. Here, we show that this preordering of TCAs is required for the transfer of the whisker map to the neocortex. Using Ebf1 conditional inactivation that specifically perturbs the development of an intermediate target, the basal ganglia, we scrambled TCA topography en route to the neocortex without affecting the thalamus or neocortex. Notably, embry- onic somatosensory TCAs were shifted toward the visual cortex and showed a substantial intermixing along their trajectory. Somatosensory TCAs rewired postnatally to reach the somatosensory cortex but failed to form a topo- graphic anatomical or functional map. Our study reveals that sensory map transfer relies not only on positional infor- mation in the projecting and target structures but also on preordering of axons along their trajectory, thereby opening novel perspectives on brain wiring. Results and Discussion Topographic axonal connections, in which neighboring neu- rons project to adjacent targets, are essential for the transfer of sensory maps from the periphery to brain centers. Thalamo- cortical axons (TCAs), which convey sensory information to the neocortex, constitute a unique system to understand how such projections are formed in that they exhibit two levels of topography. First, distinct thalamic nuclei relay specific information to different neocortical areas. For instance, dorsal lateral geniculate nucleus (dLGN) axons relay visual informa- tion, ventroposterior (VP) axons relay somatosensory inputs, and ventrolateral (VL) axons convey motor information. Second, each nucleus shows a precise internal topographic organization, which is transferred in a point-to-point manner onto the neocortex. This feature is well illustrated in the rodent somatosensory map, in which the large facial whiskers form a barreloid map in the thalamus (VP), which is transferred as a barrel map in the primary somatosensory (S1) neocortex [1, 8, 9]. Although activity has been shown to refine TCA target- ing, the global topography of these projections is established during embryonic and early postnatal development by region- alized cortical guidance cues [4, 10]. Modifications in cortical regionalization shift somatosensory TCA targeting to the displaced cortical area, where they form a topographically arranged barrel map [2, 4, 11–13] via a spectacular rerouting within the deep neocortical subplate [14]. Thus, the topog- raphy of TCAs is controlled by positional information inside the neocortical target, similarly to what has been described in other axonal projections like the retinotectal system [15, 16]. However, TCAs en route to the neocortex already exhibit a precise topographic organization before reaching the cortex: axons originating from distinct thalamic nuclei fan out in the subpallium (SubP) or basal ganglia primordium, following distinct rostrocaudal trajectories [6]. This topo- graphic arrangement of TCAs is tightly controlled by subpallial guidance cues, thereby raising the question of its function [6, 17–29]. Ebf1 Inactivation in the Subpallium Scrambles TCAs En Route to the Neocortex To test whether TCA ordering in the SubP participates in their final addressing in the neocortex, we specifically perturbed SubP development, thereby affecting TCA positioning en passant, without affecting the thalamus or neocortex. It had been shown that full inactivation of the transcription factor Ebf1 [30–34], which is expressed in differentiating subpallial neurons [33], perturbs SubP development, leading to a mis- routing of the TCAs formed by dLGN axons and to a caudal shift of other TCAs [32, 35]. Because these defects could be due to Ebf1 inactivation in the thalamus where it is also ex- pressed, we performed a conditional inactivation of Ebf1 in the SubP, by crossing transgenic Dlx5/6::cre mice expressing the Cre recombinase in the SubP and prethalamus [36, 37] with Ebf1 flox/flox mice [38]. This procedure led to a specific inactiva- tion of Ebf1 in the SubP (see Figure S1 available online) [39] with no apparent defect in thalamus patterning and cortical regionalization (Figure S2). To assess whether conditional Ebf1 mutants recapitulated the defect in TCA ordering observed in null mutants, we performed axonal tracing using carbocyanine dye injections at embryonic day (E) 16.5 and E18.5 in the presumptive somatosensory (S1) (n control = 13 and n mutant = 8) or visual (V1) neocortex (n control = 5 and n mutant = 8) (Figures 1A and 1B; data not shown). These procedures specifically back-label thalamic neurons that have projected to distinct cortical regions at E16.5, and label both thalamic neurons and reciprocal cortical axons at E18.5 [7]. As ex- pected, in E18.5 control embryos, DiI injections in presumptive *Correspondence: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Sensory map transfer to the neocortex relies on pretarget ordering of thalamic axons

Current Biology 23, 810–816, May 6, 2013 ª2013 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2013.03.062
ReportSensory Map Transfer to the NeocortexRelies on Pretarget Orderingof Thalamic Axons
Ludmilla Lokmane,1,2,3 Remi Proville,1,2,3
Nicolas Narboux-Neme,4,5,6 Ildiko Gyory,7
Maryama Keita,1,2,3 Caroline Mailhes,1,2,3 Clement Lena,1,2,3
Patricia Gaspar,4,5,6 Rudolf Grosschedl,7
and Sonia Garel1,2,3,*1Ecole Normale Superieure, Institut de Biologie de l’ENS(IBENS), 46 Rue d’Ulm, 75005 Paris, France2INSERM, U1024, 75005 Paris, France3CNRS, UMR 8197, 75005 Paris, France4INSERM, UMR-S 839, 17 Rue du Fer a Moulin,75005 Paris, France5Universite Pierre et Marie Curie, 75005 Paris, France6Institut du Fer a Moulin, 75005 Paris, France7Department of Cellular and Molecular Immunology,Max Planck Institute of Immunobiology, Stuebeweg 51,79108 Freiburg, Germany
Summary
Sensory maps, such as the representation of mouse facialwhiskers, are conveyed throughout the nervous system bytopographic axonal projections that preserve neighboringrelationships between adjacent neurons [1]. In particular,themap transfer to the neocortex is ensured by thalamocort-ical axons (TCAs), whose terminals are topographicallyorganized in response to intrinsic cortical signals [2–5].However, TCAs already show a topographic order early indevelopment, as they navigate toward their target [6, 7].Here, we show that this preordering of TCAs is required forthe transfer of the whisker map to the neocortex. UsingEbf1 conditional inactivation that specifically perturbs thedevelopment of an intermediate target, the basal ganglia,we scrambled TCA topography en route to the neocortexwithout affecting the thalamus or neocortex. Notably, embry-onic somatosensory TCAs were shifted toward the visualcortex and showed a substantial intermixing along theirtrajectory. Somatosensory TCAs rewired postnatally toreach the somatosensory cortex but failed to form a topo-graphic anatomical or functional map. Our study revealsthat sensory map transfer relies not only on positional infor-mation in the projecting and target structures but also onpreordering of axons along their trajectory, thereby openingnovel perspectives on brain wiring.
Results and Discussion
Topographic axonal connections, in which neighboring neu-rons project to adjacent targets, are essential for the transferof sensory maps from the periphery to brain centers. Thalamo-cortical axons (TCAs), which convey sensory information tothe neocortex, constitute a unique system to understandhow such projections are formed in that they exhibit two levelsof topography. First, distinct thalamic nuclei relay specificinformation to different neocortical areas. For instance, dorsal
lateral geniculate nucleus (dLGN) axons relay visual informa-tion, ventroposterior (VP) axons relay somatosensory inputs,and ventrolateral (VL) axons convey motor information.Second, each nucleus shows a precise internal topographicorganization, which is transferred in a point-to-point manneronto the neocortex. This feature is well illustrated in the rodentsomatosensory map, in which the large facial whiskers forma barreloid map in the thalamus (VP), which is transferred asa barrel map in the primary somatosensory (S1) neocortex[1, 8, 9]. Although activity has been shown to refine TCA target-ing, the global topography of these projections is establishedduring embryonic and early postnatal development by region-alized cortical guidance cues [4, 10]. Modifications in corticalregionalization shift somatosensory TCA targeting to thedisplaced cortical area, where they form a topographicallyarranged barrel map [2, 4, 11–13] via a spectacular reroutingwithin the deep neocortical subplate [14]. Thus, the topog-raphy of TCAs is controlled by positional information insidethe neocortical target, similarly to what has been describedin other axonal projections like the retinotectal system[15, 16]. However, TCAs en route to the neocortex alreadyexhibit a precise topographic organization before reachingthe cortex: axons originating from distinct thalamic nuclei fanout in the subpallium (SubP) or basal ganglia primordium,following distinct rostrocaudal trajectories [6]. This topo-graphic arrangement of TCAs is tightly controlled by subpallialguidance cues, thereby raising the question of its function[6, 17–29].
Ebf1 Inactivation in the Subpallium Scrambles TCAs EnRoute to the NeocortexTo test whether TCA ordering in the SubP participates in theirfinal addressing in the neocortex, we specifically perturbedSubP development, thereby affecting TCA positioning enpassant, without affecting the thalamus or neocortex. It hadbeen shown that full inactivation of the transcription factorEbf1 [30–34], which is expressed in differentiating subpallialneurons [33], perturbs SubP development, leading to a mis-routing of the TCAs formed by dLGN axons and to a caudalshift of other TCAs [32, 35]. Because these defects could bedue to Ebf1 inactivation in the thalamus where it is also ex-pressed, we performed a conditional inactivation of Ebf1 inthe SubP, by crossing transgenic Dlx5/6::cre mice expressingthe Cre recombinase in the SubP and prethalamus [36, 37] withEbf1flox/flox mice [38]. This procedure led to a specific inactiva-tion of Ebf1 in the SubP (see Figure S1 available online) [39]with no apparent defect in thalamus patterning and corticalregionalization (Figure S2). To assess whether conditionalEbf1 mutants recapitulated the defect in TCA orderingobserved in null mutants, we performed axonal tracing usingcarbocyanine dye injections at embryonic day (E) 16.5 andE18.5 in the presumptive somatosensory (S1) (ncontrol = 13and nmutant = 8) or visual (V1) neocortex (ncontrol = 5 and nmutant =8) (Figures 1A and 1B; data not shown). These proceduresspecifically back-label thalamic neurons that have projectedto distinct cortical regions at E16.5, and label both thalamicneurons and reciprocal cortical axons at E18.5 [7]. As ex-pected, in E18.5 control embryos, DiI injections in presumptive*Correspondence: [email protected]

V1 neocortex back-labeled dLGN neurons (Figure 1C), andinjections in the presumptive S1 neocortex back-labeled neu-rons in the VP (Figure 1D). In contrast, in Dlx5/6::cre; Ebf1fl/2
embryos, DiI injections in the presumptive V1 neocortexabnormally back-labeled neurons in the VP (Figure 1E),whereas injections in the presumptive S1 neocortex aberrantlyback-labeled neurons in the VL (Figure 1F). Furthermore, wefound that dLGN TCAs were misrouted within the embryonicSubP of Dlx5/6::cre; Ebf1fl/2 embryos as in null mutants ([32];Figure S1). Consistent with defects in TCA navigation andtopography, the morphology of SubP corridor guideposts,which provide cues for navigating thalamic axons [6, 7, 23,26], was impaired in conditional Ebf1 embryos as in null mu-tants ([32]; Figure S1). These data show that defects in interme-diate target development in Dlx5/6::cre; Ebf1fl/2 embryos leadto a phenotype similar to the one observed in null mutants [32]:(1) VP TCAs navigate to the presumptive V1 neocortex, and (2)VL TCAs navigate to the presumptive S1 neocortex (Figure 1G).
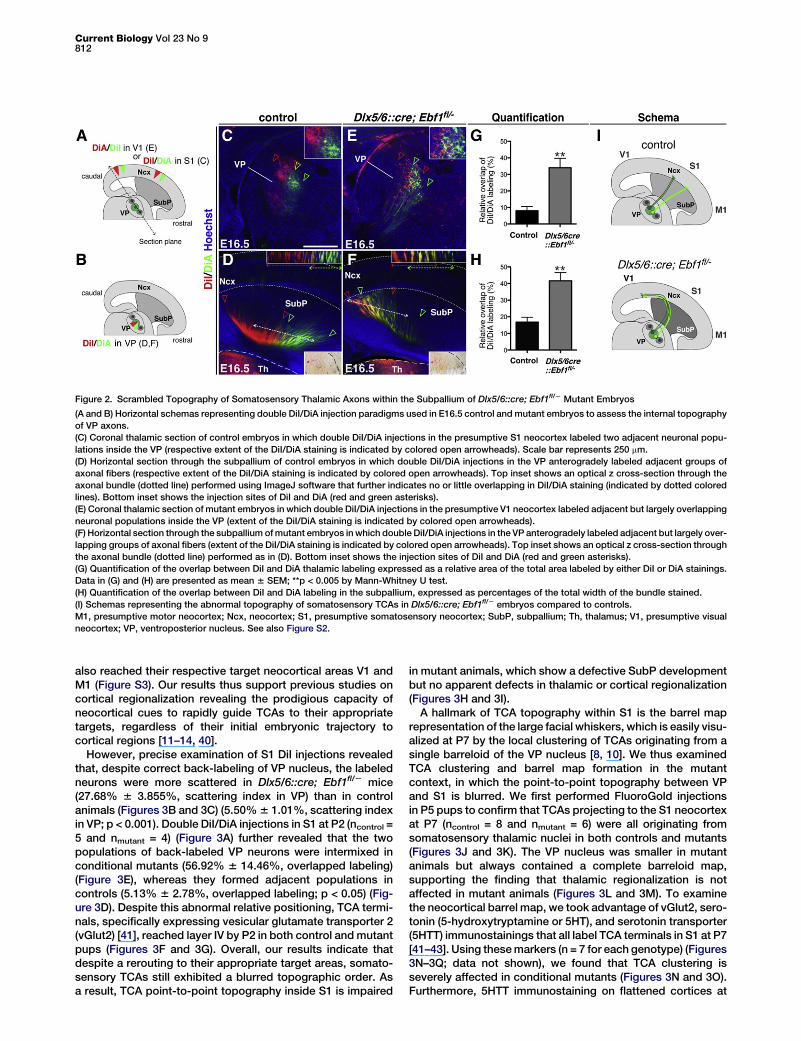
To further assess the defect of TCA topography in condi-tional mutant embryos, we examined somatosensory TCAsby performing double DiI/DiA injections at E16.5 (Figures 2Aand 2B). Double injections in presumptive S1 neocortex ofcontrol embryos (n = 6) back-labeled adjacent subpopulationsof thalamic VP neurons that showed little to no intermixing(8.01% 6 2.51% of overlap; Figures 2C and 2G). Similarly,double injections in control VP nuclei (n = 8), which specificallylabel TCAs at this stage [7, 8], anterogradely labeled adjacentbut largely nonoverlapping axonal bundles in the SubP(16.82% 6 2.88% of overlap; Figures 2D and 2H), reflecting asharp positioning of VP axons en route to the neocortex. Incontrast, in Dlx5/6::cre; Ebf1fl/2 embryos, double injections
in the presumptive V1 neocortex (n = 6), which back-labeledsomatosensory TCAs, stained neurons in adjacent but largelyoverlapping domains of the VP nucleus (34.03% 6 5.67% ofoverlap; **p < 0.005; Figures 2E and 2G). Similarly, anterogradedouble injections in mutant VP nuclei (n = 7) labeled largelyoverlapping axonal bundles in the SubP (41.62% 6 4.94% ofoverlap; **p < 0.005; Figures 2F–2H), indicating a substantialintermixing of somatosensory TCAs en route to the neocortex.Taken together, our results reveal that defective SubP devel-opment in Dlx5/6::cre; Ebf1fl/2 embryos leads to (1) caudalnavigation of VP somatosensory TCAs to the presumptive V1neocortex instead of S1, and (2) blurring in the pretargetordering of somatosensory TCAs en route to the neocortex(Figure 2I).
Scrambled Somatosensory TCAs Do Not Form aTopographic Barrel MapTCAs initially reach the embryonic neocortical subplate andonly invade their target cortical layers postnatally [8, 10]. Wethus investigated how the observed embryonic defects insomatosensory TCA ordering affect their final topographyand sensory map transfer, by analyzing map formation in thebarrel cortex. In contrast with embryonic analyses, DiI injec-tions in S1 neocortical area at postnatal day 2 (P2) (n = 12 foreach genotype) (Figure 3A) back-labeled neurons in the VPnucleus of both control and Dlx5/6::cre; Ebf1fl/2 animals (Fig-ures 3B and 3C). Following axonal trajectories in mutantanimals, we found that a majority of back-labeled VP axonstraveled first to V1 and then rerouted to S1 through the sub-plate region (Figure S3). Similarly, dLGN visual TCAs and VLmotor TCAs, which were misrouted during embryogenesis,
Figure 1. Abnormal Topography of TCAs in Dlx5/6::cre; Ebf1fl/2 Mutant Embryos
(A and B) Horizontal schemas of DiI injection paradigms used in control and mutant embryos to analyze TCA topography. Neurons back-labeled by DiIinjections within the presumptive V1 or S1 neocortex were analyzed on sections through the thalamus (dotted lines).(C and D) Coronal sections through control thalami showed retrolabeled neurons in the dLGN (dotted lines in C) or in the VP nucleus (indicated by the openarrowheads in D) after DiI injections in the presumptive V1 (C) and S1 neocortex (D), respectively. Scale bar represents 250 mm.(E and F) Coronal sections through Dlx5/6::cre; Ebf1fl/2 mutant thalami showed retrolabeled neurons in the VP nucleus (indicated by open arrowheads in E)or in the VL nucleus (indicated by solid arrowheads in F) after DiI injections in presumptive V1 (E) and S1 neocortex (F), respectively.(G) Schemas representing the shift observed in the embryonic topography of TCAs in Dlx5/6::cre; Ebf1fl/2 mice compared to controls.dLGN, dorsal lateral geniculate nucleus; M1, presumptive motor neocortex; Ncx, neocortex; S1, presumptive somatosensory neocortex; SubP, subpallium;Th, thalamus; V1, presumptive visual neocortex; VL, ventrolateral nucleus; VP, ventroposterior nucleus. See also Figure S1.
Pretarget Axonal Ordering in Sensory Map Transfer811
also reached their respective target neocortical areas V1 andM1 (Figure S3). Our results thus support previous studies oncortical regionalization revealing the prodigious capacity ofneocortical cues to rapidly guide TCAs to their appropriatetargets, regardless of their initial embryonic trajectory tocortical regions [11–14, 40].
However, precise examination of S1 DiI injections revealedthat, despite correct back-labeling of VP nucleus, the labeledneurons were more scattered in Dlx5/6::cre; Ebf1fl/2 mice(27.68% 6 3.855%, scattering index in VP) than in controlanimals (Figures 3B and 3C) (5.50%6 1.01%, scattering indexin VP; p < 0.001). Double DiI/DiA injections in S1 at P2 (ncontrol =5 and nmutant = 4) (Figure 3A) further revealed that the twopopulations of back-labeled VP neurons were intermixed inconditional mutants (56.92% 6 14.46%, overlapped labeling)(Figure 3E), whereas they formed adjacent populations incontrols (5.13% 6 2.78%, overlapped labeling; p < 0.05) (Fig-ure 3D). Despite this abnormal relative positioning, TCA termi-nals, specifically expressing vesicular glutamate transporter 2(vGlut2) [41], reached layer IV by P2 in both control and mutantpups (Figures 3F and 3G). Overall, our results indicate thatdespite a rerouting to their appropriate target areas, somato-sensory TCAs still exhibited a blurred topographic order. Asa result, TCA point-to-point topography inside S1 is impaired
in mutant animals, which show a defective SubP developmentbut no apparent defects in thalamic or cortical regionalization(Figures 3H and 3I).A hallmark of TCA topography within S1 is the barrel map
representation of the large facial whiskers, which is easily visu-alized at P7 by the local clustering of TCAs originating from asingle barreloid of the VP nucleus [8, 10]. We thus examinedTCA clustering and barrel map formation in the mutantcontext, in which the point-to-point topography between VPand S1 is blurred. We first performed FluoroGold injectionsin P5 pups to confirm that TCAs projecting to the S1 neocortexat P7 (ncontrol = 8 and nmutant = 6) were all originating fromsomatosensory thalamic nuclei in both controls and mutants(Figures 3J and 3K). The VP nucleus was smaller in mutantanimals but always contained a complete barreloid map,supporting the finding that thalamic regionalization is notaffected in mutant animals (Figures 3L and 3M). To examinethe neocortical barrel map, we took advantage of vGlut2, sero-tonin (5-hydroxytryptamine or 5HT), and serotonin transporter(5HTT) immunostainings that all label TCA terminals in S1 at P7[41–43]. Using thesemarkers (n = 7 for each genotype) (Figures3N–3Q; data not shown), we found that TCA clustering isseverely affected in conditional mutants (Figures 3N and 3O).Furthermore, 5HTT immunostaining on flattened cortices at
Figure 2. Scrambled Topography of Somatosensory Thalamic Axons within the Subpallium of Dlx5/6::cre; Ebf1fl/2 Mutant Embryos
(A and B) Horizontal schemas representing double DiI/DiA injection paradigms used in E16.5 control andmutant embryos to assess the internal topographyof VP axons.(C) Coronal thalamic section of control embryos in which double DiI/DiA injections in the presumptive S1 neocortex labeled two adjacent neuronal popu-lations inside the VP (respective extent of the DiI/DiA staining is indicated by colored open arrowheads). Scale bar represents 250 mm.(D) Horizontal section through the subpallium of control embryos in which double DiI/DiA injections in the VP anterogradely labeled adjacent groups ofaxonal fibers (respective extent of the DiI/DiA staining is indicated by colored open arrowheads). Top inset shows an optical z cross-section through theaxonal bundle (dotted line) performed using ImageJ software that further indicates no or little overlapping in DiI/DiA staining (indicated by dotted coloredlines). Bottom inset shows the injection sites of DiI and DiA (red and green asterisks).(E) Coronal thalamic section of mutant embryos in which double DiI/DiA injections in the presumptive V1 neocortex labeled adjacent but largely overlappingneuronal populations inside the VP (extent of the DiI/DiA staining is indicated by colored open arrowheads).(F) Horizontal section through the subpallium ofmutant embryos inwhich double DiI/DiA injections in the VP anterogradely labeled adjacent but largely over-lapping groups of axonal fibers (extent of the DiI/DiA staining is indicated by colored open arrowheads). Top inset shows an optical z cross-section throughthe axonal bundle (dotted line) performed as in (D). Bottom inset shows the injection sites of DiI and DiA (red and green asterisks).(G) Quantification of the overlap between DiI and DiA thalamic labeling expressed as a relative area of the total area labeled by either DiI or DiA stainings.Data in (G) and (H) are presented as mean 6 SEM; **p < 0.005 by Mann-Whitney U test.(H) Quantification of the overlap between DiI and DiA labeling in the subpallium, expressed as percentages of the total width of the bundle stained.(I) Schemas representing the abnormal topography of somatosensory TCAs in Dlx5/6::cre; Ebf1fl/2 embryos compared to controls.M1, presumptive motor neocortex; Ncx, neocortex; S1, presumptive somatosensory neocortex; SubP, subpallium; Th, thalamus; V1, presumptive visualneocortex; VP, ventroposterior nucleus. See also Figure S2.
Current Biology Vol 23 No 9812

P7 (ncontrol = 6 and nmutant = 5) confirmed the impaired anatom-ical TCA-barrel like clustering in a normally shaped S1 (Fig-ure 3Q). This finding cannot be explained by a delayed arrivalof TCAs in layer IV, because barrel formation can occur up toP7 once the pattern is imprinted in the thalamus [8, 44]. Collec-tively, our results thus indicate that defective SubP develop-ment leads to a caudal shift of somatosensory TCAs and ascrambling of their relative positioning during embryogenesis.Although targeting to S1 is rapidly rescued postnatally, TCAscrambling persists after birth, leading to a lack of point-to-point topography between VP and S1 and an impairment of
TCA clustering (Figures 3R and 3S), which is required for theemergence of barrels [8].
TCA Scrambling Impairs the Emergence of a FunctionalMap in Barrel CortexBecause sensory map topography has been shown to existin mice that lack anatomically visible TCA clusters or barrels[45–47], we examined the functional topography in thesomatosensory barrel map of adult mutant mice. To this end,we performed in vivo autofluorescence imaging of corticalactivity. This technique reveals local increases in metabolism
Figure 3. Impairment of Axonal Topographic Map Formation in the Barrel Cortex of Dlx5/6::cre; Ebf1fl/2 Pups
(A) Schema of DiI/DiA injection paradigms used in P2 control and Dlx5/6::cre; Ebf1fl/2 mice.(B andC) Thalamus coronal sections at P2 after DiI injections in S1 area show back-labeled neurons in the VP (indicated by open arrowheads) of both control(B) and Dlx5/6::cre; Ebf1fl/2 mice (C), positioned as either a dense group of neurons in controls (inset in B) or scattered cells in mutant mice (inset in C).(D and E) Thalamus coronal sections at P2 after double DiI/DiA injections in S1 show back-labeled neurons in the VP (indicated by open arrowheads) thatform either adjacent nonoverlapping neuronal groups in controls (D and inset) or overlapping neuronal groups in mutants (E and inset).(F and G) CTIP2 and vGlut2 coimmunostaining in S1 similarly marks layer V neurons and TCA terminals in layer IV, respectively, in both P2 control (F) andmutant pups (G).(H and I) Schemas recapitulating the topography defect of somatosensory TCAs at P2 in Dlx5/6::cre; Ebf1fl/2 pups (I) compared to controls (H).(J and K) Thalamus coronal sections at P7 after FluoroGold injection in S1 show back-labeled neurons in the VP nucleus and other somatosensory nuclei(indicated by open arrowheads) in both controls (J) and Dlx5/6::cre; Ebf1fl/2 mice (K).(L and M) Autofluorescence in the green channel of the sections presented in (J) and (K) outlines the barreloid map in the VP nuclei of controls (L) andDlx5/6::cre; Ebf1fl/2 mice (M).(N andO) Double immunostaining on coronal sections through S1 at P7 showsCTIP2-positive neurons in layer V and vGlut2-positive TCA terminal clusters inlayer IV of controls (N), whereas in mutant mice, vGlut2-positive TCA terminals do not cluster in layer IV (O).(P and Q) 5HTT immunostaining on P7 flattened cortex shows TCA clustering in the barrel field of control mice (P), whereas individual clusters are not clearlydetectable in mutants (Q).(R and S) Schemas representing the topography of somatosensory TCAs and the formation of the barrel clusters in controls (R), which are both impaired inDlx5/6::cre; Ebf1fl/2 mice (S).Ncx, neocortex; S1, somatosensory neocortex; SP, subplate; SubP, subpallium; VP, ventroposterior nucleus. Scale bars represent 250 mm (B, D, J, L, N, andP) and 100 mm (F and G). See also Figure S3.
Pretarget Axonal Ordering in Sensory Map Transfer813
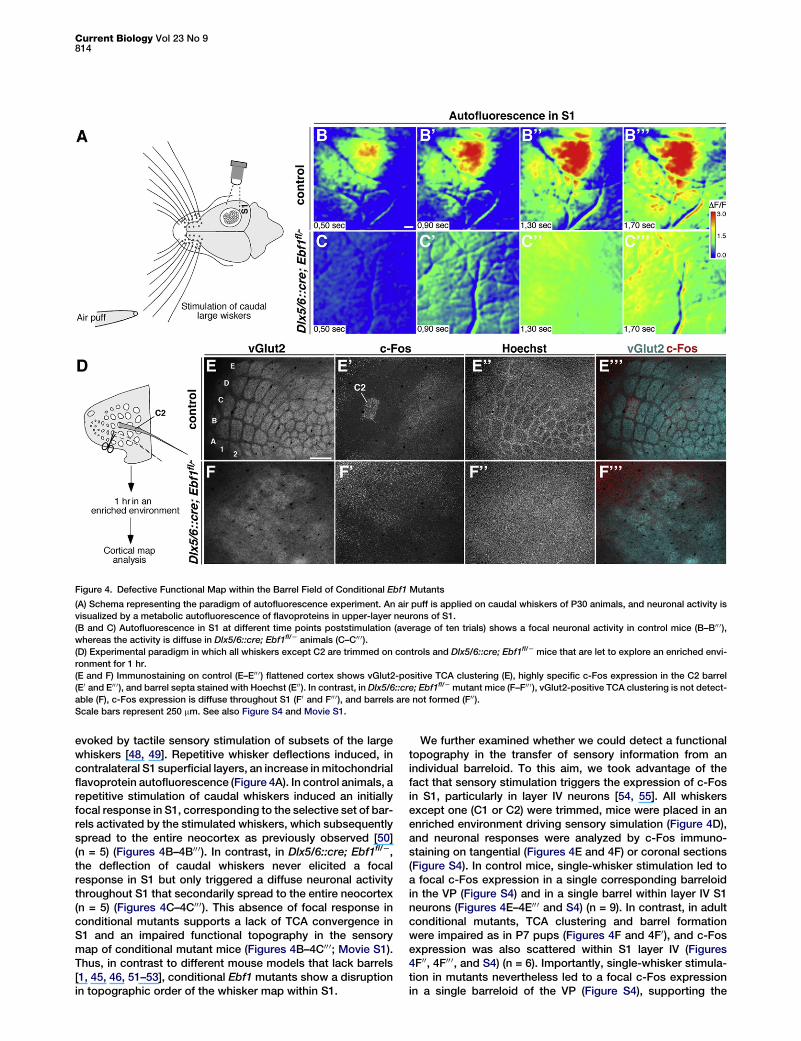
evoked by tactile sensory stimulation of subsets of the largewhiskers [48, 49]. Repetitive whisker deflections induced, incontralateral S1 superficial layers, an increase inmitochondrialflavoprotein autofluorescence (Figure 4A). In control animals, arepetitive stimulation of caudal whiskers induced an initiallyfocal response in S1, corresponding to the selective set of bar-rels activated by the stimulated whiskers, which subsequentlyspread to the entire neocortex as previously observed [50](n = 5) (Figures 4B–4B00 0). In contrast, in Dlx5/6::cre; Ebf1fl/2,the deflection of caudal whiskers never elicited a focalresponse in S1 but only triggered a diffuse neuronal activitythroughout S1 that secondarily spread to the entire neocortex(n = 5) (Figures 4C–4C00 0). This absence of focal response inconditional mutants supports a lack of TCA convergence inS1 and an impaired functional topography in the sensorymap of conditional mutant mice (Figures 4B–4C00 0; Movie S1).Thus, in contrast to different mouse models that lack barrels[1, 45, 46, 51–53], conditional Ebf1 mutants show a disruptionin topographic order of the whisker map within S1.
We further examined whether we could detect a functionaltopography in the transfer of sensory information from anindividual barreloid. To this aim, we took advantage of thefact that sensory stimulation triggers the expression of c-Fosin S1, particularly in layer IV neurons [54, 55]. All whiskersexcept one (C1 or C2) were trimmed, mice were placed in anenriched environment driving sensory simulation (Figure 4D),and neuronal responses were analyzed by c-Fos immuno-staining on tangential (Figures 4E and 4F) or coronal sections(Figure S4). In control mice, single-whisker stimulation led toa focal c-Fos expression in a single corresponding barreloidin the VP (Figure S4) and in a single barrel within layer IV S1neurons (Figures 4E–4E00 0 and S4) (n = 9). In contrast, in adultconditional mutants, TCA clustering and barrel formationwere impaired as in P7 pups (Figures 4F and 4F0), and c-Fosexpression was also scattered within S1 layer IV (Figures4F00, 4F00 0, and S4) (n = 6). Importantly, single-whisker stimula-tion in mutants nevertheless led to a focal c-Fos expressionin a single barreloid of the VP (Figure S4), supporting the
Figure 4. Defective Functional Map within the Barrel Field of Conditional Ebf1 Mutants
(A) Schema representing the paradigm of autofluorescence experiment. An air puff is applied on caudal whiskers of P30 animals, and neuronal activity isvisualized by a metabolic autofluorescence of flavoproteins in upper-layer neurons of S1.(B and C) Autofluorescence in S1 at different time points poststimulation (average of ten trials) shows a focal neuronal activity in control mice (B–B00 0),whereas the activity is diffuse in Dlx5/6::cre; Ebf1fl/2 animals (C–C00 0).(D) Experimental paradigm in which all whiskers except C2 are trimmed on controls and Dlx5/6::cre; Ebf1fl/2 mice that are let to explore an enriched envi-ronment for 1 hr.(E and F) Immunostaining on control (E–E00 0) flattened cortex shows vGlut2-positive TCA clustering (E), highly specific c-Fos expression in the C2 barrel(E0 and E00 0), and barrel septa stained with Hoechst (E00). In contrast, in Dlx5/6::cre; Ebf1fl/2 mutant mice (F–F00 0), vGlut2-positive TCA clustering is not detect-able (F), c-Fos expression is diffuse throughout S1 (F0 and F00 0), and barrels are not formed (F00).Scale bars represent 250 mm. See also Figure S4 and Movie S1.
Current Biology Vol 23 No 9814

finding that the whisker map is properly relayed up to thethalamus. Thus, the stimulation of a single whisker does notdrive focal neuronal activity in S1, thereby revealing a lack offunctional topography in the barrel cortex. Collectively, theseresults indicate that perturbing the developmental preorderingof TCAs, without affecting thalamus and neocortex, compro-mises the establishment of the topographic sensory whiskermap in the adult neocortex.
ConclusionsWe have shown that precise TCA topography at the level of anintermediate target, the subpallium, is required for the correcttransfer of a sensory map to the neocortex. Indeed, usinga novel viable mouse model, we have demonstrated thatalthough global developmental TCA topography defects canbe postnatally rescued by positional information in theneocortex, the loss of fine axonal neighboring relationshipsalong their trajectory leads to a defective transfer of a somato-sensory map. Our study reveals that, in addition to topograph-ically organized cues in the projecting and target structures[15], preordering of axons along their trajectory is required forthe transfer of an anatomical and functional sensory mapto the neocortex. This work shows that defects in subpallialdevelopment may unexpectedly impact on cortical mapformation, thereby opening novel perspectives on brainphysiopathology. Furthermore, it reveals that the global topog-raphy of TCAs within the neocortex and the fine topography ofsomatosensory TCAswithin their target areas are controlled bystrikingly different processes. Finally, our study raises theintriguing possibility that the fine axonal ordering within tracts,which has been described in various systems [16, 56–59], mayhave a general role in organizing neural circuit wiring.
Supplemental Information
Supplemental Information includes four figures, Supplemental Experi-mental Procedures, and one movie and can be found with this article onlineat http://dx.doi.org/10.1016/j.cub.2013.03.062.
Acknowledgments
We are grateful to Christo Goridis and members of the Garel lab for stimu-lating discussions and critical comments on the manuscript, and toBenjamin Mathieu, Christophe Auger, and Deborah Souchet for excellenttechnical assistance. This work was supported by grants from the INSERM‘‘Avenir’’ program to S.G., the city of Paris to S.G., the EURYI program toS.G., and the ANR project Sensocode (ANR-11-BSV4-028) to C.L. S.G. isan EMBO YIP awardee, and both S.G. and P.G. are part of the Ecole desNeurosciences de Paris Ile-de-France network. L.L. was supported by apostdoctoral fellowship from the Fondation ARC pour la Recherche sur leCancer, and R.P. was supported by doctoral fellowships from the Ministerede l’Enseignement Superieur et de la Recherche and the FRM. All animalhousing and experimental procedures were performed following French/EU regulations and in compliance with the local animal research committee.
Received: December 11, 2012Revised: February 21, 2013Accepted: March 22, 2013Published: April 25, 2013
References
1. Wu, C.S., Ballester Rosado, C.J., and Lu, H.C. (2011). What can we getfrom ‘barrels’: the rodent barrel cortex as a model for studying theestablishment of neural circuits. Eur. J. Neurosci. 34, 1663–1676.
2. Assimacopoulos, S., Kao, T., Issa, N.P., and Grove, E.A. (2012).Fibroblast growth factor 8 organizes the neocortical area map and reg-ulates sensory map topography. J. Neurosci. 32, 7191–7201.
3. Fukuchi-Shimogori, T., and Grove, E.A. (2001). Neocortex patterning bythe secreted signaling molecule FGF8. Science 294, 1071–1074.
4. O’Leary, D.D., Chou, S.J., and Sahara, S. (2007). Area patterning of themammalian cortex. Neuron 56, 252–269.
5. O’Leary, D.D., and Sahara, S. (2008). Genetic regulation of arealizationof the neocortex. Curr. Opin. Neurobiol. 18, 90–100.
6. Bielle, F., Marcos-Mondejar, P., Leyva-Dıaz, E., Lokmane, L., Mire, E.,Mailhes, C., Keita, M., Garcıa, N., Tessier-Lavigne, M., Garel, S., andLopez-Bendito, G. (2011). Emergent growth cone responses to combi-nations of Slit1 and Netrin 1 in thalamocortical axon topography. Curr.Biol. 21, 1748–1755.
7. Molnar, Z., Garel, S., Lopez-Bendito, G., Maness, P., and Price, D.J.(2012). Mechanisms controlling the guidance of thalamocortical axonsthrough the embryonic forebrain. Eur. J. Neurosci. 35, 1573–1585.
8. Erzurumlu, R.S., and Gaspar, P. (2012). Development and critical periodplasticity of the barrel cortex. Eur. J. Neurosci. 35, 1540–1553.
9. Erzurumlu, R.S., Murakami, Y., and Rijli, F.M. (2010). Mapping the face inthe somatosensory brainstem. Nat. Rev. Neurosci. 11, 252–263.
10. Agmon, A., Yang, L.T., Jones, E.G., and O’Dowd, D.K. (1995).Topological precision in the thalamic projection to neonatal mouse bar-rel cortex. J. Neurosci. 15, 549–561.
11. Hamasaki, T., Leingartner, A., Ringstedt, T., and O’Leary, D.D. (2004).EMX2 regulates sizes and positioning of the primary sensory and motorareas in neocortex by direct specification of cortical progenitors.Neuron 43, 359–372.
12. Pinon, M.C., Tuoc, T.C., Ashery-Padan, R., Molnar, Z., and Stoykova, A.(2008). Altered molecular regionalization and normal thalamocorticalconnections in cortex-specific Pax6 knock-out mice. J. Neurosci. 28,8724–8734.
13. Armentano, M., Chou, S.J., Tomassy, G.S., Leingartner, A., O’Leary,D.D., and Studer, M. (2007). COUP-TFI regulates the balance of corticalpatterning between frontal/motor and sensory areas. Nat. Neurosci. 10,1277–1286.
14. Shimogori, T., and Grove, E.A. (2005). Fibroblast growth factor 8 regu-lates neocortical guidance of area-specific thalamic innervation.J. Neurosci. 25, 6550–6560.
15. Sperry, R.W. (1943). Effect of 180 degree rotation of the retinal field onvisuomotor coordination. J. Exp. Zool. 92, 263–279.
16. Luo, L., and Flanagan, J.G. (2007). Development of continuous anddiscrete neural maps. Neuron 56, 284–300.
17. Lokmane, L., and Garel, S. (2011). Screening for genes that wire the ce-rebral cortex. BMC Biol. 9, 1.
18. Braisted, J.E., Tuttle, R., and O’leary, D.D. (1999). Thalamocorticalaxons are influenced by chemorepellent and chemoattractant activitieslocalized to decision points along their path. Dev. Biol. 208, 430–440.
19. Bonnin, A., Torii, M., Wang, L., Rakic, P., and Levitt, P. (2007). Serotoninmodulates the response of embryonic thalamocortical axons to netrin-1. Nat. Neurosci. 10, 588–597.
20. Powell, A.W., Sassa, T., Wu, Y., Tessier-Lavigne, M., and Polleux, F.(2008). Topography of thalamic projections requires attractive andrepulsive functions of Netrin-1 in the ventral telencephalon. PLoS Biol.6, e116.
21. Wright, A.G., Demyanenko, G.P., Powell, A., Schachner, M., Enriquez-Barreto, L., Tran, T.S., Polleux, F., andManess, P.F. (2007). Close homo-log of L1 and neuropilin 1 mediate guidance of thalamocortical axons atthe ventral telencephalon. J. Neurosci. 27, 13667–13679.
22. Dufour, A., Seibt, J., Passante, L., Depaepe, V., Ciossek, T., Frisen, J.,Kullander, K., Flanagan, J.G., Polleux, F., and Vanderhaeghen, P.(2003). Area specificity and topography of thalamocortical projectionsare controlled by ephrin/Eph genes. Neuron 39, 453–465.
23. Lopez-Bendito, G., Cautinat, A., Sanchez, J.A., Bielle, F., Flames, N.,Garratt, A.N., Talmage, D.A., Role, L.W., Charnay, P., Marın, O., andGarel, S. (2006). Tangential neuronal migration controls axon guidance:a role for neuregulin-1 in thalamocortical axon navigation. Cell 125,127–142.
24. Metin, C., and Godement, P. (1996). The ganglionic eminencemay be anintermediate target for corticofugal and thalamocortical axons.J. Neurosci. 16, 3219–3235.
25. Zhou, L., Bar, I., Achouri, Y., Campbell, K., De Backer, O., Hebert, J.M.,Jones, K., Kessaris, N., de Rouvroit, C.L., O’Leary, D., et al. (2008). Earlyforebrain wiring: genetic dissection using conditional Celsr3 mutantmice. Science 320, 946–949.
26. Bielle, F., Marcos-Mondejar, P., Keita, M., Mailhes, C., Verney, C.,Nguyen Ba-Charvet, K., Tessier-Lavigne, M., Lopez-Bendito, G., and
Pretarget Axonal Ordering in Sensory Map Transfer815

Garel, S. (2011). Slit2 activity in the migration of guidepost neuronsshapes thalamic projections during development and evolution.Neuron 69, 1085–1098.
27. Bagri, A., Marın, O., Plump, A.S., Mak, J., Pleasure, S.J., Rubenstein,J.L., and Tessier-Lavigne, M. (2002). Slit proteins prevent midlinecrossing and determine the dorsoventral position of major axonal path-ways in the mammalian forebrain. Neuron 33, 233–248.
28. Braisted, J.E., Ringstedt, T., and O’Leary, D.D. (2009). Slits are chemo-repellents endogenous to hypothalamus and steer thalamocorticalaxons into ventral telencephalon. Cereb. Cortex 19(Suppl 1 ), i144–i151.
29. Lopez-Bendito, G., and Molnar, Z. (2003). Thalamocortical develop-ment: how are we going to get there? Nat. Rev. Neurosci. 4, 276–289.
30. Hesslein, D.G., Fretz, J.A., Xi, Y., Nelson, T., Zhou, S., Lorenzo, J.A.,Schatz, D.G., and Horowitz, M.C. (2009). Ebf1-dependent control ofthe osteoblast and adipocyte lineages. Bone 44, 537–546.
31. Jin, K., and Xiang, M. (2011). Ebf1 deficiency causes increase of Mullercells in the retina and abnormal topographic projection at the opticchiasm. Biochem. Biophys. Res. Commun. 414, 539–544.
32. Garel, S., Yun, K., Grosschedl, R., and Rubenstein, J.L. (2002). The earlytopography of thalamocortical projections is shifted in Ebf1 and Dlx1/2mutant mice. Development 129, 5621–5634.
33. Garel, S., Marın, F., Grosschedl, R., and Charnay, P. (1999). Ebf1 con-trols early cell differentiation in the embryonic striatum. Development126, 5285–5294.
34. Lin, H., andGrosschedl, R. (1995). Failure of B-cell differentiation inmicelacking the transcription factor EBF. Nature 376, 263–267.
35. Garel, S., and Rubenstein, J.L. (2004). Intermediate targets in formationof topographic projections: inputs from the thalamocortical system.Trends Neurosci. 27, 533–539.
36. Ghanem, N., Yu, M., Poitras, L., Rubenstein, J.L., and Ekker, M. (2008).Characterization of a distinct subpopulation of striatal projection neu-rons expressing the Dlx genes in the basal ganglia through the activityof the I56ii enhancer. Dev. Biol. 322, 415–424.
37. Zerucha, T., Stuhmer, T., Hatch, G., Park, B.K., Long, Q., Yu, G.,Gambarotta, A., Schultz, J.R., Rubenstein, J.L., and Ekker, M. (2000).A highly conserved enhancer in the Dlx5/Dlx6 intergenic region is thesite of cross-regulatory interactions between Dlx genes in the embry-onic forebrain. J. Neurosci. 20, 709–721.
38. Gyory, I., Boller, S., Nechanitzky, R., Mandel, E., Pott, S., Liu, E., andGrosschedl, R. (2012). Transcription factor Ebf1 regulates differentiationstage-specific signaling, proliferation, and survival of B cells. GenesDev. 26, 668–682.
39. Madisen, L., Zwingman, T.A., Sunkin, S.M., Oh, S.W., Zariwala, H.A., Gu,H., Ng, L.L., Palmiter, R.D., Hawrylycz, M.J., Jones, A.R., et al. (2010). Arobust and high-throughput Cre reporting and characterization systemfor the whole mouse brain. Nat. Neurosci. 13, 133–140.
40. Little, G.E., Lopez-Bendito, G., Runker, A.E., Garcıa, N., Pinon, M.C.,Chedotal, A., Molnar, Z., and Mitchell, K.J. (2009). Specificity andplasticity of thalamocortical connections in Sema6A mutant mice.PLoS Biol. 7, e98.
41. Nahmani, M., and Erisir, A. (2005). VGluT2 immunochemistry identifiesthalamocortical terminals in layer 4 of adult and developing visualcortex. J. Comp. Neurol. 484, 458–473.
42. Rebsam, A., Seif, I., and Gaspar, P. (2002). Refinement of thalamocort-ical arbors and emergence of barrel domains in the primary somatosen-sory cortex: a study of normal and monoamine oxidase a knock-outmice. J. Neurosci. 22, 8541–8552.
43. Lebrand, C., Cases, O., Wehrle, R., Blakely, R.D., Edwards, R.H., andGaspar, P. (1998). Transient developmental expression of monoaminetransporters in the rodent forebrain. J. Comp. Neurol. 401, 506–524.
44. Rebsam, A., Seif, I., and Gaspar, P. (2005). Dissociating barrel develop-ment and lesion-induced plasticity in the mouse somatosensory cortex.J. Neurosci. 25, 706–710.
45. Ince-Dunn, G., Hall, B.J., Hu, S.C., Ripley, B., Huganir, R.L., Olson, J.M.,Tapscott, S.J., and Ghosh, A. (2006). Regulation of thalamocorticalpatterning and synaptic maturation by NeuroD2. Neuron 49, 683–695.
46. Welker, E., Armstrong-James, M., Bronchti, G., Ourednik, W.,Gheorghita-Baechler, F., Dubois, R., Guernsey, D.L., Van der Loos, H.,and Neumann, P.E. (1996). Altered sensory processing in the somato-sensory cortex of the mouse mutant barrelless. Science 271, 1864–1867.
47. Abdel-Majid, R.M., Leong, W.L., Schalkwyk, L.C., Smallman, D.S.,Wong, S.T., Storm, D.R., Fine, A., Dobson, M.J., Guernsey, D.L., and
Neumann, P.E. (1998). Loss of adenylyl cyclase I activity disruptspatterning of mouse somatosensory cortex. Nat. Genet. 19, 289–291.
48. Shibuki, K., Hishida, R., Murakami, H., Kudoh, M., Kawaguchi, T.,Watanabe, M., Watanabe, S., Kouuchi, T., and Tanaka, R. (2003).Dynamic imaging of somatosensory cortical activity in the rat visualizedby flavoprotein autofluorescence. J. Physiol. 549, 919–927.
49. Murakami, H., Kamatani, D., Hishida, R., Takao, T., Kudoh, M.,Kawaguchi, T., Tanaka, R., and Shibuki, K. (2004). Short-term plasticityvisualized with flavoprotein autofluorescence in the somatosensorycortex of anaesthetized rats. Eur. J. Neurosci. 19, 1352–1360.
50. Ferezou, I., Bolea, S., and Petersen, C.C. (2006). Visualizing the corticalrepresentation of whisker touch: voltage-sensitive dye imaging in freelymoving mice. Neuron 50, 617–629.
51. Iwasato, T., Datwani, A., Wolf, A.M., Nishiyama, H., Taguchi, Y.,Tonegawa, S., Knopfel, T., Erzurumlu, R.S., and Itohara, S. (2000).Cortex-restricted disruption of NMDAR1 impairs neuronal patterns inthe barrel cortex. Nature 406, 726–731.
52. Cases, O., Vitalis, T., Seif, I., De Maeyer, E., Sotelo, C., and Gaspar, P.(1996). Lack of barrels in the somatosensory cortex of monoamineoxidase A-deficient mice: role of a serotonin excess during the criticalperiod. Neuron 16, 297–307.
53. Li, H., and Crair, M.C. (2011). How do barrels form in somatosensorycortex? Ann. N Y Acad. Sci. 1225, 119–129.
54. Narboux-Neme, N., Evrard, A., Ferezou, I., Erzurumlu, R.S., Kaeser, P.S.,Laine, J., Rossier, J., Ropert, N., Sudhof, T.C., and Gaspar, P. (2012).Neurotransmitter release at the thalamocortical synapse instructs barrelformation but not axon patterning in the somatosensory cortex.J. Neurosci. 32, 6183–6196.
55. Staiger, J.F., Bisler, S., Schleicher, A., Gass, P., Stehle, J.H., and Zilles,K. (2000). Exploration of a novel environment leads to the expression ofinducible transcription factors in barrel-related columns. Neuroscience99, 7–16.
56. Gosse, N.J., Nevin, L.M., and Baier, H. (2008). Retinotopic order in theabsence of axon competition. Nature 452, 892–895.
57. Imai, T., Yamazaki, T., Kobayakawa, R., Kobayakawa, K., Abe, T.,Suzuki, M., and Sakano, H. (2009). Pre-target axon sorting establishesthe neural map topography. Science 325, 585–590.
58. Imai, T., and Sakano, H. (2011). Axon-axon interactions in neuronalcircuit assembly: lessons from olfactory map formation. Eur. J.Neurosci. 34, 1647–1654.
59. Triplett, J.W., Pfeiffenberger, C., Yamada, J., Stafford, B.K., Sweeney,N.T., Litke, A.M., Sher, A., Koulakov, A.A., and Feldheim, D.A. (2011).Competition is a driving force in topographic mapping. Proc. Natl.Acad. Sci. USA 108, 19060–19065.
Current Biology Vol 23 No 9816




















