
SEASONAL VARIABILITY OF PHYSICOCHEMICAL AND RHEOLOGICAL PROPERTIES OF ULVAN IN TWO ULVA SPECIES...
12
SEASONAL VARIABILITY OF PHYSICOCHEMICAL AND RHEOLOGICAL PROPERTIES OF ULVAN IN TWO ULVA SPECIES (CHLOROPHYTA) FROM THE BRITTANY COAST 1 Audrey Robic 2 Centre d’Etudes et de Valorisation des Algues, Presqu’ı ˆle de Pen Lan, BP3, 22610 Pleubian, France Institut National de la Recherche Agronomique, Unite ´ Biopolyme `res, Interactions, Assemblages, UR1268, Rue de la Ge ´raudie `re, BP 71627, 44316 Nantes, France Jean-Franc ¸ois Sassi 3 , Patrick Dion, Yannick Lerat Centre d’Etudes et de Valorisation des Algues, Presqu’ı ˆle de Pen Lan, BP3, 22610 Pleubian, France and Marc Lahaye Institut National de la Recherche Agronomique, Unite ´ Biopolyme `res, Interactions, Assemblages, UR1268, Rue de la Ge ´raudie `re, BP 71627, 44316 Nantes, France The seasonal variability in the extraction yield, physicochemical characteristics, and rheological properties of ulvan from two Ulva species contribut- ing to Brittany ‘‘green tides’’ has been studied. These seaweeds were collected in the water column for Ulva armoricana Dion, de Reviers et Coat and on hard substrata for Ulva rotundata Bliding. The maximum ulvan extraction efficiency was not related to the maximum ulvan content in the seaweeds, but with the active growth period of the seaweeds. Ulvan chemical structure, macromolecular character- istics, and rheological properties were affected by both species and seasons. The proportion of high- molecular-weight ulvan was the major factor positively correlated with the gelling properties. Characteristics of ulvan from U. rotundata subjected to tides were more affected by seasons than ulvan from U. armoricana living in a more constant environment. These results point to several useful recommendations concerning Ulva sp. biomass collected with regard to ulvan characteristics and uses. Key index words: extraction yield; gelling proper- ties; green tides; physicochemical characteristics; seasonal variability; Ulva armoricana; Ulva rotun- data; ulvan Abbreviations: dwt, dry weight; Gal, galactose; Glc, glucose; HPAEC, high-performance anion exch- ange chromatography; HPSEC, high-performance size exclusion chromatography; Rha, rhamnose; UA, uronic acids; w ⁄ w, weight ⁄ weight; Xyl, xylose Species of Ulva are abundant seaweeds occurring in the intertidal zone. They colonize hard substrata, anchored by an attachment disk, but some species may give rise to a free-living form drifting on soft substrata. Ulva are fast-growing algae and are oppor- tunistic for space and nutrient uptake. Growth in the water column is particularly prevalent in eutrophic coastal and lagoon waters where Ulva sp. proliferate in the form of ‘‘green tides’’ (Fletcher 1996). This phenomenon often results in mass pro- duction and harmful accumulations (Morand and Briand 1996). To date, this biomass has very low added value and ways to use it include compost (Maze ´ et al. 1993, Cuomo et al. 1995), methane pro- duction (Briand and Morand 1997), human food consumption (Pe ´rez 1997), and potentially paper making (Nicolucci and Monegato 1993). Ulva sp. represent a source of a complex sulfated polysaccha- ride, ulvan, which is drawing increasing interest as a potential source of new functional biopolymer (Lahaye and Robic 2007). Ulvans represent 8%–29% of the algal dry weight. They are mainly made of rhamnose, glucuronic acid, iduronic acid, xylose, and sulfate. Minor amounts of glucose, galactose, and mannose can also be found (Percival and McDowell 1967, Quem- ener et al. 1997). Most of the constitutive sugars are distributed as repeating sequences. The two major ones are aldobiuronic acid disaccharides designated as type A (ulvanobiuronic acid 3-sulfate, A 3s ) and type B (ulvanobiuronic acid 3-sulfate, B 3s ) (Fig. 1). Partially sulfated xylose residues at O-2 can also occur in place of uronic acids. In addition, glucu- ronic acid can branch at O-2 of rhamnose 3-sulfate (Ray and Lahaye 1995a,b). Ulvan is characterized by physicochemical and biological properties linked to its peculiar chemical structures, charge density, and molecular weight dis- tribution. Among them, ulvan yields low viscous 1 Received 5 September 2008. Accepted 25 February 2009. 2 Author for correspondence: e-mail [email protected]. 3 Author for correspondence: e-mail [email protected]. J. Phycol. 45, 962–973 (2009) ȑ 2009 Phycological Society of America DOI: 10.1111/j.1529-8817.2009.00699.x 962
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of SEASONAL VARIABILITY OF PHYSICOCHEMICAL AND RHEOLOGICAL PROPERTIES OF ULVAN IN TWO ULVA SPECIES...

SEASONAL VARIABILITY OF PHYSICOCHEMICAL AND RHEOLOGICAL PROPERTIESOF ULVAN IN TWO ULVA SPECIES (CHLOROPHYTA) FROM THE BRITTANY COAST1
Audrey Robic2
Centre d’Etudes et de Valorisation des Algues, Presqu’ıle de Pen Lan, BP3, 22610 Pleubian, France
Institut National de la Recherche Agronomique, Unite Biopolymeres, Interactions, Assemblages, UR1268, Rue de la Geraudiere,
BP 71627, 44316 Nantes, France
Jean-Francois Sassi 3, Patrick Dion, Yannick Lerat
Centre d’Etudes et de Valorisation des Algues, Presqu’ıle de Pen Lan, BP3, 22610 Pleubian, France
and Marc Lahaye
Institut National de la Recherche Agronomique, Unite Biopolymeres, Interactions, Assemblages, UR1268, Rue de la Geraudiere,
BP 71627, 44316 Nantes, France
The seasonal variability in the extraction yield,physicochemical characteristics, and rheologicalproperties of ulvan from two Ulva species contribut-ing to Brittany ‘‘green tides’’ has been studied.These seaweeds were collected in the water columnfor Ulva armoricana Dion, de Reviers et Coat andon hard substrata for Ulva rotundata Bliding. Themaximum ulvan extraction efficiency was not relatedto the maximum ulvan content in the seaweeds,but with the active growth period of the seaweeds.Ulvan chemical structure, macromolecular character-istics, and rheological properties were affected byboth species and seasons. The proportion of high-molecular-weight ulvan was the major factorpositively correlated with the gelling properties.Characteristics of ulvan from U. rotundata subjectedto tides were more affected by seasons than ulvanfrom U. armoricana living in a more constantenvironment. These results point to several usefulrecommendations concerning Ulva sp. biomasscollected with regard to ulvan characteristics anduses.
Key index words: extraction yield; gelling proper-ties; green tides; physicochemical characteristics;seasonal variability; Ulva armoricana; Ulva rotun-data; ulvan
Abbreviations: dwt, dry weight; Gal, galactose; Glc,glucose; HPAEC, high-performance anion exch-ange chromatography; HPSEC, high-performancesize exclusion chromatography; Rha, rhamnose;UA, uronic acids; w ⁄ w, weight ⁄ weight; Xyl, xylose
Species of Ulva are abundant seaweeds occurringin the intertidal zone. They colonize hard substrata,anchored by an attachment disk, but some speciesmay give rise to a free-living form drifting on softsubstrata. Ulva are fast-growing algae and are oppor-tunistic for space and nutrient uptake. Growth inthe water column is particularly prevalent ineutrophic coastal and lagoon waters where Ulva sp.proliferate in the form of ‘‘green tides’’ (Fletcher1996). This phenomenon often results in mass pro-duction and harmful accumulations (Morand andBriand 1996). To date, this biomass has very lowadded value and ways to use it include compost(Maze et al. 1993, Cuomo et al. 1995), methane pro-duction (Briand and Morand 1997), human foodconsumption (Perez 1997), and potentially papermaking (Nicolucci and Monegato 1993). Ulva sp.represent a source of a complex sulfated polysaccha-ride, ulvan, which is drawing increasing interest as apotential source of new functional biopolymer(Lahaye and Robic 2007).
Ulvans represent 8%–29% of the algal dry weight.They are mainly made of rhamnose, glucuronicacid, iduronic acid, xylose, and sulfate. Minoramounts of glucose, galactose, and mannose canalso be found (Percival and McDowell 1967, Quem-ener et al. 1997). Most of the constitutive sugars aredistributed as repeating sequences. The two majorones are aldobiuronic acid disaccharides designatedas type A (ulvanobiuronic acid 3-sulfate, A3s) andtype B (ulvanobiuronic acid 3-sulfate, B3s) (Fig. 1).Partially sulfated xylose residues at O-2 can alsooccur in place of uronic acids. In addition, glucu-ronic acid can branch at O-2 of rhamnose 3-sulfate(Ray and Lahaye 1995a,b).
Ulvan is characterized by physicochemical andbiological properties linked to its peculiar chemicalstructures, charge density, and molecular weight dis-tribution. Among them, ulvan yields low viscous
1Received 5 September 2008. Accepted 25 February 2009.2Author for correspondence: e-mail [email protected] for correspondence: e-mail [email protected].
J. Phycol. 45, 962–973 (2009)� 2009 Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2009.00699.x
962

aqueous solutions able to form thermoreversiblegels in the presence of divalent cations and borate(Haug 1976, Lahaye and Robic 2007). Its peculiarion-exchange and gelling characteristics can providenew functionalities for applications in which metalchelation is required and ⁄ or in the conception ofmaterials such as nanostructured composites (Lazaet al. 2007). Ulvan also presents several potentiallyvaluable biological properties for agricultural, food,and pharmaceutical applications (Ivanova et al.1994, Bobin-Dubigeon et al. 1997, Lahaye andKaeffer 1997, Kaeffer et al. 1999, Barcelo et al.2000, Cluzet et al. 2004, Mao et al. 2006).
To establish the industrial use of Ulva for ulvanproduction, several requirements regarding biomassavailability and quality have to be met. The proce-dure used to stabilize collected Ulva had a markedimpact on ulvan extraction yield and physicochemi-cal and rheological properties (Robic et al. 2008).The best gelling properties were obtained afterrapid stabilization treatment of seaweeds right aftercollection, but an inverse relationship was deter-mined between gelling properties and ulvan extrac-tion yield. Other sources of ulvan variability canarise from confusion in identifying the species(Lahaye and Ray 1996). Ulva comprises a group ofuniform algal morphologies colonizing diversehabitats due to their tolerance of a wide range ofenvironmental conditions (Smith 1947, Littler andLittler 1980, Beach et al. 1995). They show littlefunctional thallus differentiation, and precise identi-fication in the field without genetic tools is difficult(Blomster et al. 1998, Coat et al. 1998). Ecophysio-logical factors and environmental conditions areother major potential causes of ulvan quantitativeand qualitative variations. Environmental parametershave been reported to affect the development ofred and brown seaweeds (growth, reproduction)with consequences on agar, carrageenans, and algi-nate biosynthesis and their functional properties(Kloareg and Quatrano 1988, Craigie 1990, Lahaye
2001). To date, studies on species and seasonalimpacts on green algal polysaccharide compositionand properties are limited (Abdel-Fattah and Edrees1973, Medcalf et al. 1975, Lahaye et al. 1999, Siddh-anta et al. 2001).
In this work, we explored parameters generatingthe variability of ulvan extraction yield and physico-chemical and rheological properties of two abundantUlva species on the Brittany coast. The algae werecollected at different seasons from the water columnfor the drifting life-form of U. armoricana, presentonly in summer blooms on the soft bottom ofeutrophic bays, and over several months for fixedpopulations of U. rotundata, present along the coaston hard substrata. The two samplings differ both atthe species level and in environmental conditions toenhance potential variability in chemical content.
MATERIALS AND METHODS
Material. Algae were collected monthly when available.U. armoricana was collected from July 2006 to October 2006 andin July 2007 at Saint Michel en Greve (3�34¢28.23¢¢ W,48�40¢34.2¢¢ N) at low tide. U. rotundata was collected from July2006 to October 2006 and from February 2007 to July 2007 inPleubian (3�4¢12.61¢¢ W, 48�51¢35.39¢¢ N) at low tide. Algaewere immediately frozen at )30�C.
Extraction of ulvan. After thawing, seaweeds were ground topieces of �0.5 cm using a grinder (Minipimer, MR 6500 BC;Braun, Kronberg, Deutschland) and refluxed (2 h, 85�C) insodium oxalate (0.05 M in deionized water, dry solid content:6.8% w ⁄ w) under constant stirring. The suspension was filteredon a strainer with a pore size of �1 mm, and the residues wereextracted by deionized water (dry initial solid content: 6.8%w ⁄ w) for 1 h at room temperature. The last slurry was filtered,and the combined extracts were concentrated (5·) by ultrafil-tration (Mw 10 kDa, Amicon, Beverly, MA, USA). The retentatesolution was diafiltered with five volumes of deionized waterand then freeze-dried. Each extraction was performed induplicate. A sample from each seaweed was dried at 50�C inventilated ovens (Memmert D06061 Modell 500; MemmertGmbH + Co. KG, Schwabach, Germany) and ground to�100 lm particle size with a Retsch mill (model 5657 type 2;Retsch, Dusseldorf, Germany) prior to analysis.
Fig. 1. Structure of the mainrepeating disaccharides in Ulvaulvan: ulvanobiuronic acids A3S andB3S, and ulvanobioses U3S andU2’S,3S.
ULVAN VARIABILITY 963

Chemical analysis. Dry solid content corresponded to theweight of samples remaining after 24 h at 103�C. Organicmatter content was quantified gravimetrically after 12 h at550�C. Sulfate content was measured according to Tabatabai(1974) for the seaweed samples and according to Quemeneret al. (1997) for the ulvan samples. Protein content wasestimated as N Kjelhdahl · 6.25. Total uronic acid contentwas measured by the m-phenyl phenol colorimetric method(Thibault 1979) using glucuronic acid as standard. Neutralsugars and uronic acid composition in extracts were deter-mined after methanolysis in MeOH-HCl and HPLC analysis(Absorbosphere RP18, 5 lm, 4 · 250 mm, Grace, Deerfield,IL, USA; Quemener et al. 2000). Chromatographic peaks wereidentified by comparison with reference sugars. Quantificationof neutral sugars in seaweeds was performed by gas–liquidchromatography (GC) after sulfuric acid degradation (Englystand Cummings 1988, Hoebler et al. 1989). Insoluble sampleswere dispersed in 13 M sulfuric acid for 30 min at 25�C, priorto dilution to 1 M and hydrolysis (2 h, 100�C). For calibration,standard monosaccharide solutions and inositol (internalstandard) were used. Starch content was determined afteramylolysis of the ulvan extracts following an adaptation of themethod of McCleary et al. 1994. The extracts were firstsubmitted to an a-amylase and then to an amyloglucosidase.The resulting free glucose was determined after high-perfor-mance anion exchange chromatography (HPAEC) analysis(Carbopac PA1, 4 · 250 mm; Dionex Corporation, Sunnyvale,CA, USA). The starch glucose content was subtracted from theglucose measured by methanolysis to yield cell-wall glucosecontent. All measurements were done in triplicate.
Molecular weight determinations. Ulvan solution (4 mg ÆmL)1) in 50 mM NaNO3 containing 0.02% NaN3 passedthrough 0.22 lm membrane filter was injected at 25�C on ahigh-performance size exclusion chromatography system(HPSEC) constituted of two Shodex OH-pack SB HQ 804and 805 columns (8 · 300 mm, Guard columns: OHpak SB-800P, 6 · 50 mm, Shodex; Showa Denko KK, Miniato, Japan)eluted at 0.7 mL Æ min)1 with 50 mM NaNO3 containing 0.02%NaN3. Elution was monitored by online intrinsic viscosity ([g])determination using a differential viscometer (T-50A; ViscotekEurope, Irigny, France), a differential refractometer (ERC 7517A; Erma Inc., Tokyo, Japan), and a UV detector at 280 nm(LDC ⁄ Milton Roy SpectroMonitor 3000; LDC S.A., Paris,France). Weight average molecular weight was calculated fromintrinsic viscosity using the Trisec software (Viscotek). Thesedeterminations were made on material eluted from 16.4 to20.0 min and from 20.0 to 25.0 min. The proportion ofpopulations eluted from 16.4 to 20.0 min, from 20.0 to25.0 min, and from 25.0 to 30.0 min was determined usingthe refractometric data.
Rheological measurements. Dynamic viscosity analysis wasperformed using a Carri-med CS-100 stress-controlled rheom-eter (TA Instrument Division de Waters SAS, Guyancourt,France) with a temperature-regulated cone-plate device(radius: 2 cm; cone-plate angle: 4�1¢22¢¢). The polysaccharideconcentration in salt solutions (CuSO4 7 mM and H3BO3
30 mM) was 1.6% (w ⁄ v), and experiments were carried out at20�C. Copper ions and boric acid were added simultaneouslyjust before pouring the polysaccharide solution onto the Carri-med plate. Paraffin oil was layered onto the cone and sample toprevent evaporation. The kinetics of gel formation was mon-itored for 45 min by measuring the complex modulus at0.159 Hz (1 rad Æ s)1) with a low deformation of 3%.
Structural analysis. 13C NMR spectra were recorded on aBruker ARX 400 spectrometer (Bruker BioSpin S. A., Wissem-bourg, France). A sample of �70 mg was dissolved in 1 mL ofD2O (99.9%) and 2 mL of H2O with a trace of acetone asinternal reference (31.45 ppm). Analyses were realized at333 K.
Enzymatic degradation. Crude ulvan-lyase was prepared as a60%–80% ammonium sulfate precipitate of a liquid culturefrom a gram-negative ulvanolytic bacterium isolated frommarine sediments (Lahaye et al. 1997). Ulvan (0.6% w ⁄ v indeionized water, pH 7.5) was degraded at room temperature byaddition of crude ulvan-lyase (2.1 nkat). After 8 h, the samequantity of enzyme was added, and incubation was continuedfor an additional 15 h. Degradation was stopped by heating theulvan solution in a boiling water bath for 10 min. Nondegrad-ed ulvan was precipitated in four volumes of ethanol, andoligosaccharides were recovered from the ethanol supernatantafter evaporation, dissolved in water, and freeze-dried.
Gel permeation chromatography. Freeze-dried oligosaccharidesfrom ulvan enzymatic degradation were dissolved in a smallamount of 50 mM NaNO3 containing 0.02% NaN3 andchromatographed through Shodex OH Pak SB-802.5 HQcolumns (8 · 300 mm) by NaNO3 (50 mM, 0.7 mL Æ min)1).Elution was followed by a differential refractometer (ERC 7517A). The proportion of peak 1 to 6 was expressed as apercentage of the sum of the area of all peaks detected.
Data analyses. Statistical differences (expressed as P-values)were evaluated by analysis of variance using Statgraphics PlusVersion 5.1 (Statistical Graphics Corp., Warrenton, VA, USA).Principal component analysis (PCA) was performed andPearson correlation (r) was determined using MATLABsoftware, version 7 (Math Works, Natick, MA, USA). The 95%confidence limit corresponded to |r| values > 0.5 for seaweedsamples, 0.4 for U. rotundata ulvan, 0.6 for U. armoricana ulvan,and 0.4 for the combined collection of ulvan.
RESULTS
Chemical composition of U. rotundata and U. armor-icana. The total sugar, sulfate, and protein con-tents on the dry algae weight basis are given inTable 1. Using only the data of the commonmonths, the mean total sugar (26.0 ± 3.5% forU. rotundata and 22.5 ± 6.7% for U. armoricana)and protein (13.4 ± 4.3% for U. rotundata and15.1 ± 2.8% for U. armoricana) contents did not dif-fer significantly between the two species. U. rotunda-ta contained a significantly (P = 0.0002) highermean sulfate content (8.7 ± 1.7%) than U. armorica-na (4.2 ± 1.3%). On the whole, the total sugar con-tent was inversely correlated with the proteincontent (r = )0.6).
PCA of the chemical data differentiated speciesalong the first principal component (Fig. 2). Totalsugar, rhamnose, xylose, and sulfate contents char-acterized U. rotundata, whereas protein and glucosecontents distinguished U. armoricana.
Total sugar content was the lowest in U. rotundatacollected in March and April 2007, and the richestfrom July to September 2006 and from May to July2007. For U. armoricana, the October 2006 samplewas the poorest in total sugars. The highest proteincontent in U. rotundata was determined for theOctober 2006 and February 2007 samples. ForU. armoricana, the protein content in the July2007 sample was higher than that of July 2006.U. rotundata and U. armoricana collected in July 2007were the richest in sulfate (12.6% and 6.4%,respectively).
964 AUDREY ROBIC ET AL.
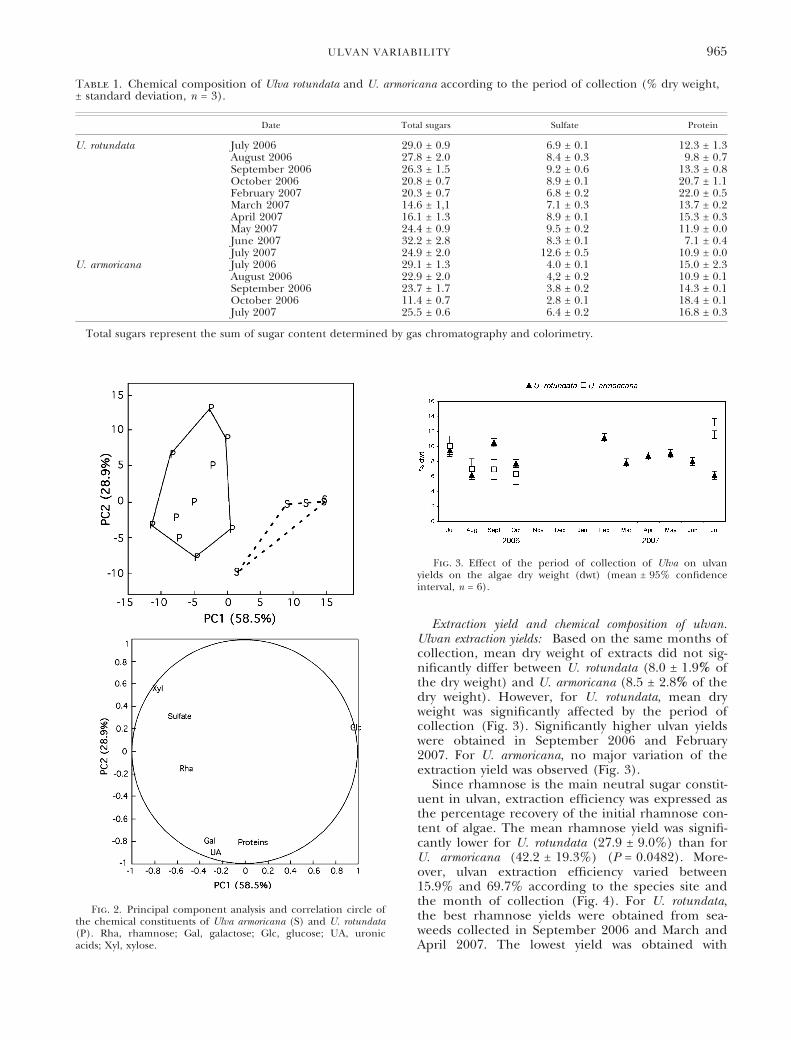
Extraction yield and chemical composition of ulvan.Ulvan extraction yields: Based on the same months ofcollection, mean dry weight of extracts did not sig-nificantly differ between U. rotundata (8.0 ± 1.9% ofthe dry weight) and U. armoricana (8.5 ± 2.8% of thedry weight). However, for U. rotundata, mean dryweight was significantly affected by the period ofcollection (Fig. 3). Significantly higher ulvan yieldswere obtained in September 2006 and February2007. For U. armoricana, no major variation of theextraction yield was observed (Fig. 3).
Since rhamnose is the main neutral sugar constit-uent in ulvan, extraction efficiency was expressed asthe percentage recovery of the initial rhamnose con-tent of algae. The mean rhamnose yield was signifi-cantly lower for U. rotundata (27.9 ± 9.0%) than forU. armoricana (42.2 ± 19.3%) (P = 0.0482). More-over, ulvan extraction efficiency varied between15.9% and 69.7% according to the species site andthe month of collection (Fig. 4). For U. rotundata,the best rhamnose yields were obtained from sea-weeds collected in September 2006 and March andApril 2007. The lowest yield was obtained with
Table 1. Chemical composition of Ulva rotundata and U. armoricana according to the period of collection (% dry weight,± standard deviation, n = 3).
Date Total sugars Sulfate Protein
U. rotundata July 2006 29.0 ± 0.9 6.9 ± 0.1 12.3 ± 1.3August 2006 27.8 ± 2.0 8.4 ± 0.3 9.8 ± 0.7September 2006 26.3 ± 1.5 9.2 ± 0.6 13.3 ± 0.8October 2006 20.8 ± 0.7 8.9 ± 0.1 20.7 ± 1.1February 2007 20.3 ± 0.7 6.8 ± 0.2 22.0 ± 0.5March 2007 14.6 ± 1,1 7.1 ± 0.3 13.7 ± 0.2April 2007 16.1 ± 1.3 8.9 ± 0.1 15.3 ± 0.3May 2007 24.4 ± 0.9 9.5 ± 0.2 11.9 ± 0.0June 2007 32.2 ± 2.8 8.3 ± 0.1 7.1 ± 0.4July 2007 24.9 ± 2.0 12.6 ± 0.5 10.9 ± 0.0
U. armoricana July 2006 29.1 ± 1.3 4.0 ± 0.1 15.0 ± 2.3August 2006 22.9 ± 2.0 4,2 ± 0.2 10.9 ± 0.1September 2006 23.7 ± 1.7 3.8 ± 0.2 14.3 ± 0.1October 2006 11.4 ± 0.7 2.8 ± 0.1 18.4 ± 0.1July 2007 25.5 ± 0.6 6.4 ± 0.2 16.8 ± 0.3
Total sugars represent the sum of sugar content determined by gas chromatography and colorimetry.
Fig. 2. Principal component analysis and correlation circle ofthe chemical constituents of Ulva armoricana (S) and U. rotundata(P). Rha, rhamnose; Gal, galactose; Glc, glucose; UA, uronicacids; Xyl, xylose.
Fig. 3. Effect of the period of collection of Ulva on ulvanyields on the algae dry weight (dwt) (mean ± 95% confidenceinterval, n = 6).
ULVAN VARIABILITY 965
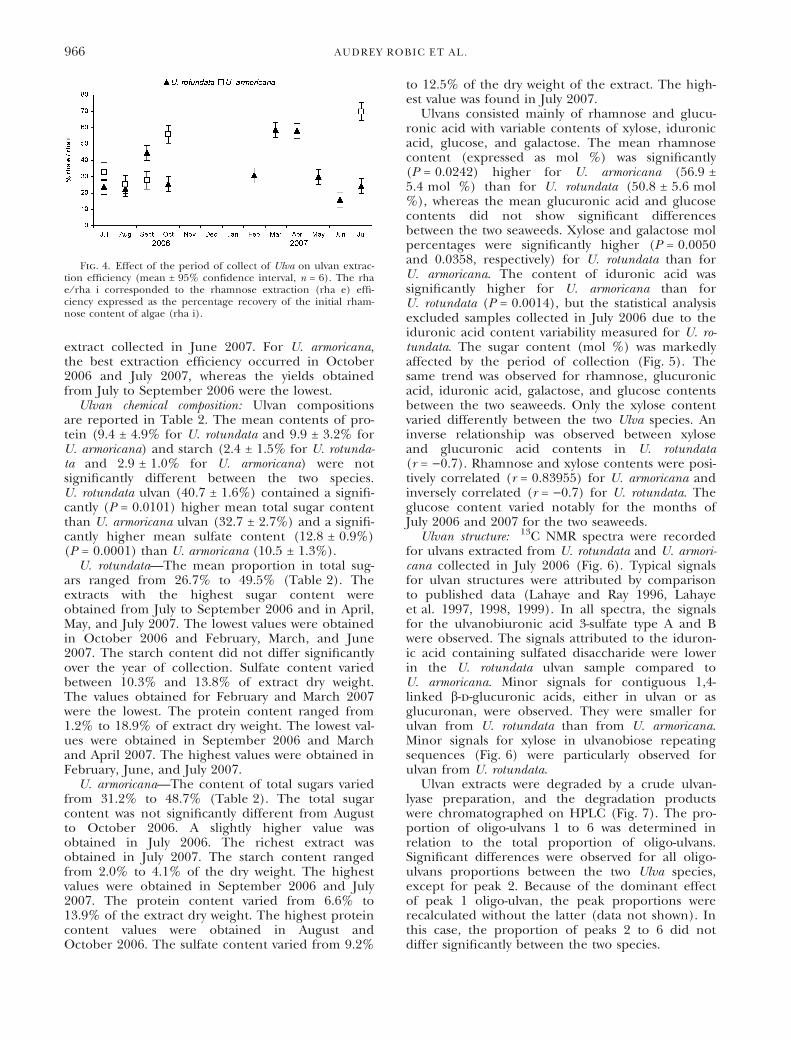
extract collected in June 2007. For U. armoricana,the best extraction efficiency occurred in October2006 and July 2007, whereas the yields obtainedfrom July to September 2006 were the lowest.
Ulvan chemical composition: Ulvan compositionsare reported in Table 2. The mean contents of pro-tein (9.4 ± 4.9% for U. rotundata and 9.9 ± 3.2% forU. armoricana) and starch (2.4 ± 1.5% for U. rotunda-ta and 2.9 ± 1.0% for U. armoricana) were notsignificantly different between the two species.U. rotundata ulvan (40.7 ± 1.6%) contained a signifi-cantly (P = 0.0101) higher mean total sugar contentthan U. armoricana ulvan (32.7 ± 2.7%) and a signifi-cantly higher mean sulfate content (12.8 ± 0.9%)(P = 0.0001) than U. armoricana (10.5 ± 1.3%).
U. rotundata—The mean proportion in total sug-ars ranged from 26.7% to 49.5% (Table 2). Theextracts with the highest sugar content wereobtained from July to September 2006 and in April,May, and July 2007. The lowest values were obtainedin October 2006 and February, March, and June2007. The starch content did not differ significantlyover the year of collection. Sulfate content variedbetween 10.3% and 13.8% of extract dry weight.The values obtained for February and March 2007were the lowest. The protein content ranged from1.2% to 18.9% of extract dry weight. The lowest val-ues were obtained in September 2006 and Marchand April 2007. The highest values were obtained inFebruary, June, and July 2007.
U. armoricana—The content of total sugars variedfrom 31.2% to 48.7% (Table 2). The total sugarcontent was not significantly different from Augustto October 2006. A slightly higher value wasobtained in July 2006. The richest extract wasobtained in July 2007. The starch content rangedfrom 2.0% to 4.1% of the dry weight. The highestvalues were obtained in September 2006 and July2007. The protein content varied from 6.6% to13.9% of the extract dry weight. The highest proteincontent values were obtained in August andOctober 2006. The sulfate content varied from 9.2%
to 12.5% of the dry weight of the extract. The high-est value was found in July 2007.
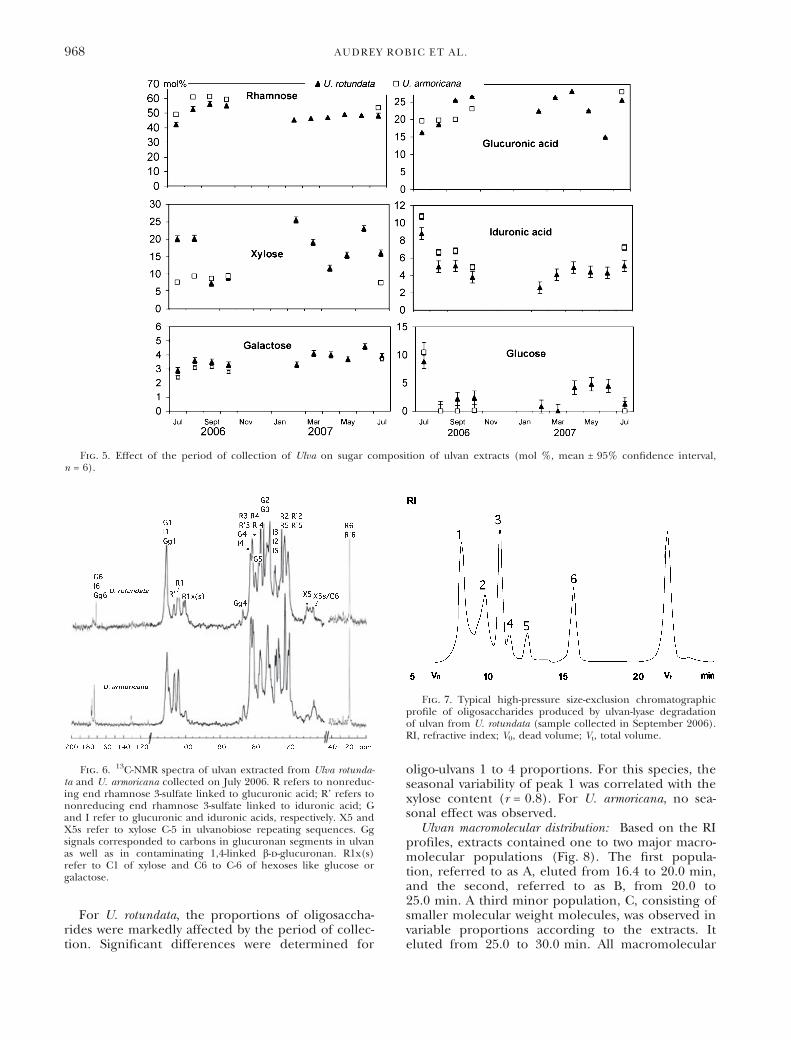
Ulvans consisted mainly of rhamnose and glucu-ronic acid with variable contents of xylose, iduronicacid, glucose, and galactose. The mean rhamnosecontent (expressed as mol %) was significantly(P = 0.0242) higher for U. armoricana (56.9 ±5.4 mol %) than for U. rotundata (50.8 ± 5.6 mol%), whereas the mean glucuronic acid and glucosecontents did not show significant differencesbetween the two seaweeds. Xylose and galactose molpercentages were significantly higher (P = 0.0050and 0.0358, respectively) for U. rotundata than forU. armoricana. The content of iduronic acid wassignificantly higher for U. armoricana than forU. rotundata (P = 0.0014), but the statistical analysisexcluded samples collected in July 2006 due to theiduronic acid content variability measured for U. ro-tundata. The sugar content (mol %) was markedlyaffected by the period of collection (Fig. 5). Thesame trend was observed for rhamnose, glucuronicacid, iduronic acid, galactose, and glucose contentsbetween the two seaweeds. Only the xylose contentvaried differently between the two Ulva species. Aninverse relationship was observed between xyloseand glucuronic acid contents in U. rotundata(r = )0.7). Rhamnose and xylose contents were posi-tively correlated (r = 0.83955) for U. armoricana andinversely correlated (r = )0.7) for U. rotundata. Theglucose content varied notably for the months ofJuly 2006 and 2007 for the two seaweeds.
Ulvan structure: 13C NMR spectra were recordedfor ulvans extracted from U. rotundata and U. armori-cana collected in July 2006 (Fig. 6). Typical signalsfor ulvan structures were attributed by comparisonto published data (Lahaye and Ray 1996, Lahayeet al. 1997, 1998, 1999). In all spectra, the signalsfor the ulvanobiuronic acid 3-sulfate type A and Bwere observed. The signals attributed to the iduron-ic acid containing sulfated disaccharide were lowerin the U. rotundata ulvan sample compared toU. armoricana. Minor signals for contiguous 1,4-linked b-d-glucuronic acids, either in ulvan or asglucuronan, were observed. They were smaller forulvan from U. rotundata than from U. armoricana.Minor signals for xylose in ulvanobiose repeatingsequences (Fig. 6) were particularly observed forulvan from U. rotundata.
Ulvan extracts were degraded by a crude ulvan-lyase preparation, and the degradation productswere chromatographed on HPLC (Fig. 7). The pro-portion of oligo-ulvans 1 to 6 was determined inrelation to the total proportion of oligo-ulvans.Significant differences were observed for all oligo-ulvans proportions between the two Ulva species,except for peak 2. Because of the dominant effectof peak 1 oligo-ulvan, the peak proportions wererecalculated without the latter (data not shown). Inthis case, the proportion of peaks 2 to 6 did notdiffer significantly between the two species.
Fig. 4. Effect of the period of collect of Ulva on ulvan extrac-tion efficiency (mean ± 95% confidence interval, n = 6). The rhae ⁄ rha i corresponded to the rhamnose extraction (rha e) effi-ciency expressed as the percentage recovery of the initial rham-nose content of algae (rha i).
966 AUDREY ROBIC ET AL.

Ta
ble
2.Se
aso
nal
vari
atio
no
fu
lvan
chem
ical
com
po
siti
on
,m
ole
cula
rw
eigh
to
fp
op
ula
tio
ns
Aan
dB
(Mw
A,
Mw
B),
mo
lecu
lar
wei
ght
dis
trib
uti
on
(%A
,%
B,
%C
),an
dco
nse
rvat
ion
mo
du
lus
of
gels
(G’)
fro
mU
lva
rotu
nda
taan
dU
.ar
mor
ican
a(%
dry
wei
ght,
±st
and
ard
dev
iati
on
,n
=6)
.
Per
iod
of
coll
ecti
on
2006
2007
P-v
alu
eJu
lyA
ugu
stSe
pte
mb
erO
cto
ber
Feb
ruar
yM
arch
Ap
ril
May
Jun
eJu
ly
U.
rotu
nda
taT
ota
lsu
gars
42.8
±1.
345
.4±
5.5
46.0
±2.
836
.2±
1.1
26.7
±1.
738
.0±
1.5
44.7
±3.
142
.8±
2.9
34.6
±1.
449
.5±
2.5
0.00
02Su
lfat
e12
.5±
0.7
12.8
±3.
313
.8±
0.6
13.2
±0.
310
.3±
0.0
11.3
±2.
012
.6±
1.1
13.6
±0.
411
.9±
0.4
11.5
±2.
10.
0049
Pro
tein
11.2
±1.
28.
1±
0.9
2.4
±0.
48.
9±
0.5
18.9
±0.
45.
7±
0.7
1.2
±0.
310
.4±
0.7
13.9
±4.
716
.4±
3.0
0.00
00St
arch
2.9
±0.
32.
9±
0.9
1.2
±0.
10.
9±
0.0
1.5
±0.
12.
1±
0.0
1.6
±0.
71.
5±
0.5
2.8
±0.
24.
1±
2.4
0.07
47M
wA
·103
(gÆm
ol)
1)
1,05
098
079
063
092
083
065
096
01,
200
800
0.00
02M
wB
·103
(gÆ
mo
l)1)
140
100
8587
120
120
110
110
115
110
0.10
67%
A50
.752
.756
.050
.937
.049
.958
.657
.441
.254
.00.
0040
%B
27.5
32.2
34.8
34.9
32.9
29.2
24.8
27.6
31.4
31.7
0.01
23%
C21
.815
.19.
214
.230
.020
.916
.615
.027
.414
.30.
0014
G’
(NÆ
m)
2)
145
220
200
955
260
165
245
2540
50.
0000
Per
iod
of
coll
ecti
on
2006
2007
P-v
alu
eJu
lyA
ugu
stSe
pte
mb
erO
cto
ber
July
U.
arm
oric
ana
To
tal
suga
rs36
.2±
1.0
32.2
±1.
931
.6±
0.7
31.2
±0.
548
.7±
2.6
0.00
04Su
lfat
e10
.6±
3.1
10.2
±0.
310
.1±
0.2
9.2
±0.
612
.5±
0.2
0.00
01P
rote
in9.
9±
1.1
12.3
±0.
16.
6±
1.5
13.9
±2.
56.
8±
1.2
0.01
47St
arch
2.5
±0.
62.
1±
0.3
4.1
±0.
32.
0±
0.0
4.0
±0.
00.
0036
Mw
A·1
03(g
Æmo
l)1)
980
100
800
730
660
0.00
94M
wB
·103
(gÆ
mo
l)1)
9090
9090
100
0.07
17%
A49
.344
.735
.923
.860
.70.
0000
%B
31.1
32.3
3945
.729
.80.
0008
%C
19.6
22.9
25.1
30.5
9.5
0.00
06G
’(N
Æm
)2)
275
175
120
2560
00.
0000
To
tal
suga
rsre
pre
sen
tth
esu
mo
fsu
gar
con
ten
td
eter
min
edb
ym
eth
ano
lysi
s.
ULVAN VARIABILITY 967
For U. rotundata, the proportions of oligosaccha-rides were markedly affected by the period of collec-tion. Significant differences were determined for
oligo-ulvans 1 to 4 proportions. For this species, theseasonal variability of peak 1 was correlated with thexylose content (r = 0.8). For U. armoricana, no sea-sonal effect was observed.
Ulvan macromolecular distribution: Based on the RIprofiles, extracts contained one to two major macro-molecular populations (Fig. 8). The first popula-tion, referred to as A, eluted from 16.4 to 20.0 min,and the second, referred to as B, from 20.0 to25.0 min. A third minor population, C, consisting ofsmaller molecular weight molecules, was observed invariable proportions according to the extracts. Iteluted from 25.0 to 30.0 min. All macromolecular
Fig. 5. Effect of the period of collection of Ulva on sugar composition of ulvan extracts (mol %, mean ± 95% confidence interval,n = 6).
Fig. 6. 13C-NMR spectra of ulvan extracted from Ulva rotunda-ta and U. armoricana collected on July 2006. R refers to nonreduc-ing end rhamnose 3-sulfate linked to glucuronic acid; R’ refers tononreducing end rhamnose 3-sulfate linked to iduronic acid; Gand I refer to glucuronic and iduronic acids, respectively. X5 andX5s refer to xylose C-5 in ulvanobiose repeating sequences. Ggsignals corresponded to carbons in glucuronan segments in ulvanas well as in contaminating 1,4-linked b-d-glucuronan. R1x(s)refer to C1 of xylose and C6 to C-6 of hexoses like glucose orgalactose.
Fig. 7. Typical high-pressure size-exclusion chromatographicprofile of oligosaccharides produced by ulvan-lyase degradationof ulvan from U. rotundata (sample collected in September 2006).RI, refractive index; V0, dead volume; Vt, total volume.
968 AUDREY ROBIC ET AL.

populations absorbed UV, which was indicative ofprotein coelution.
The mean proportion of the macromolecularpopulation A was significantly (P = 0.0327) higherfor U. rotundata (52.8 ± 3.5%) than for U. armoricana(42.9 ± 13.1%). The mean proportion of populationB did not differ significantly between the two sea-weeds, whereas the proportion of smaller molecularweight molecules was higher (P = 0.0292) forU. armoricana (21.5 ± 7.5%) than for U. rotundata(14.9 ± 4.6%). For both species, the proportion ofthe different ulvan populations was markedlyaffected by the period of collection (Fig. 9 andTable 2). For U. rotundata, February and June 2007samples were characterized by a low proportion of Aand a high proportion of C. The proportion of pop-ulation C was positively correlated with the xylosecontent (r = 0.8). For U. armoricana, the proportionof A decreased from July to October 2006, whilethat of B and C increased simultaneously. The
proportion of A was markedly higher in July 2007than in July 2006. There was a good correlationbetween the proportion of populations B and C andthe xylose content (r = 0.6 and 0.7, respectively).
Mean weight average molecular weights of ulvanfrom populations A and B did not differ signifi-cantly between U. rotundata and U. armoricana. Thelowest molecular weights of population A fromU. rotundata were found in October 2006 and April2007, whereas the highest value was observed inJune 2007 (Table 2). The molecular weight of popu-lation A from U. armoricana decreased significantlyfrom August to October 2006 (Table 2). The molec-ular weight of population A in July 2007 wasmarkedly higher than in July 2006. For both species,the molecular weight of population B was not signif-icantly affected by the period of collection.
Gelling characteristics of ulvan: Conservation mod-ulus (G’) of ulvan gels made in the presence of cop-per (7 mM) and borate (30 mM) are given inTable 2. Mean storage modulus for U. rotundata andU. armoricana (respectively, 176 and 239 N Æ m)2)did not differ significantly (P = 0.7344). U. rotundataulvan gels gave values ranging from 5 to405 N Æ m)2. The highest value was obtained withthe sample collected in July 2007, whereas the low-est values were obtained in February and June 2007.U. armoricana ulvan gels had a conservation modulusranging from 25 to 600 N Æ m)2. The highest valuewas for the sample collected in July 2007. Other val-ues ranged from 25 to 275 N Æ m)2 and decreasedslightly from July to October.
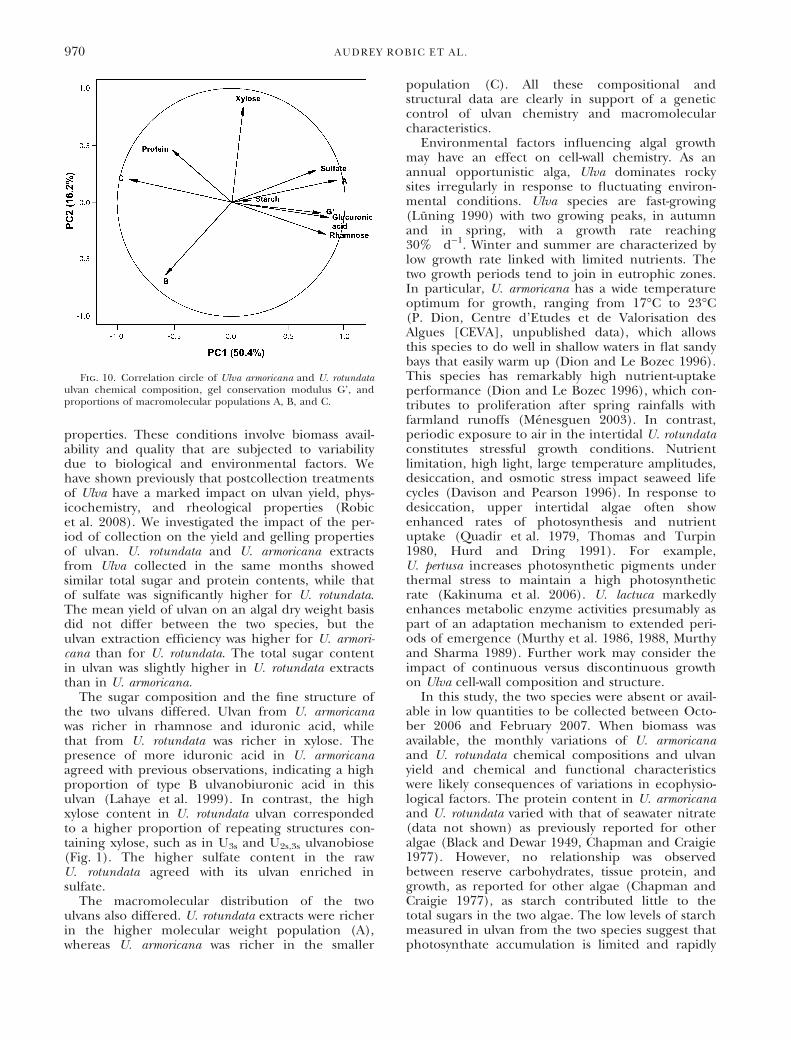
To identify which physicochemical and chemicalvariables were the most important contributors tothe rheological properties of ulvan, the storagemodulus; the proportion of populations A, B, andC; and the contents of rhamnose, glucuronic acid,xylose, sulfate, protein, and starch were submittedto PCA (Fig. 10). The first two principal compo-nents took 50.3% and 16.2% of the total inertia intoaccount and grouped G’ with population A, rham-nose, glucuronic acid, and sulfate contents. Theconservation modulus was positively correlated withthe proportion of high molecular weight and to thecontents of rhamnose, glucuronic acid, and sulfate.It was negatively correlated with the proportion ofC. It should be noted that, although no direct corre-lation was found, protein content seemed to havean impact on the rheological properties of ulvan.When the proportion in high molecular weight mol-ecules is nearly the same as for May and July 2007for U. rotundata, the sample with the highest storagemodulus is characterized by the highest proteincontent.
DISCUSSION
The development of industrial exploitation ofUlva biomass for ulvan uses requires constant orpredictive polysaccharide quantities and functional
Fig. 8. Typical high-pressure size-exclusion chromatographicprofiles of ulvan extracted from U. armoricana (sample collectedin August 2006). RI: refractive index detector; UV: UV detector;V0: dead volume; Vt: total volume.
Fig. 9. Effect of the period of collection of Ulva on the pro-portions of the macromolecular populations of ulvans deter-mined using refractometric data (mean ± 95% confidenceinterval, n = 6).
ULVAN VARIABILITY 969
properties. These conditions involve biomass avail-ability and quality that are subjected to variabilitydue to biological and environmental factors. Wehave shown previously that postcollection treatmentsof Ulva have a marked impact on ulvan yield, phys-icochemistry, and rheological properties (Robicet al. 2008). We investigated the impact of the per-iod of collection on the yield and gelling propertiesof ulvan. U. rotundata and U. armoricana extractsfrom Ulva collected in the same months showedsimilar total sugar and protein contents, while thatof sulfate was significantly higher for U. rotundata.The mean yield of ulvan on an algal dry weight basisdid not differ between the two species, but theulvan extraction efficiency was higher for U. armori-cana than for U. rotundata. The total sugar contentin ulvan was slightly higher in U. rotundata extractsthan in U. armoricana.
The sugar composition and the fine structure ofthe two ulvans differed. Ulvan from U. armoricanawas richer in rhamnose and iduronic acid, whilethat from U. rotundata was richer in xylose. Thepresence of more iduronic acid in U. armoricanaagreed with previous observations, indicating a highproportion of type B ulvanobiuronic acid in thisulvan (Lahaye et al. 1999). In contrast, the highxylose content in U. rotundata ulvan correspondedto a higher proportion of repeating structures con-taining xylose, such as in U3s and U2s,3s ulvanobiose(Fig. 1). The higher sulfate content in the rawU. rotundata agreed with its ulvan enriched insulfate.
The macromolecular distribution of the twoulvans also differed. U. rotundata extracts were richerin the higher molecular weight population (A),whereas U. armoricana was richer in the smaller
population (C). All these compositional andstructural data are clearly in support of a geneticcontrol of ulvan chemistry and macromolecularcharacteristics.
Environmental factors influencing algal growthmay have an effect on cell-wall chemistry. As anannual opportunistic alga, Ulva dominates rockysites irregularly in response to fluctuating environ-mental conditions. Ulva species are fast-growing(Luning 1990) with two growing peaks, in autumnand in spring, with a growth rate reaching30% Æ d)1. Winter and summer are characterized bylow growth rate linked with limited nutrients. Thetwo growth periods tend to join in eutrophic zones.In particular, U. armoricana has a wide temperatureoptimum for growth, ranging from 17�C to 23�C(P. Dion, Centre d’Etudes et de Valorisation desAlgues [CEVA], unpublished data), which allowsthis species to do well in shallow waters in flat sandybays that easily warm up (Dion and Le Bozec 1996).This species has remarkably high nutrient-uptakeperformance (Dion and Le Bozec 1996), which con-tributes to proliferation after spring rainfalls withfarmland runoffs (Menesguen 2003). In contrast,periodic exposure to air in the intertidal U. rotundataconstitutes stressful growth conditions. Nutrientlimitation, high light, large temperature amplitudes,desiccation, and osmotic stress impact seaweed lifecycles (Davison and Pearson 1996). In response todesiccation, upper intertidal algae often showenhanced rates of photosynthesis and nutrientuptake (Quadir et al. 1979, Thomas and Turpin1980, Hurd and Dring 1991). For example,U. pertusa increases photosynthetic pigments underthermal stress to maintain a high photosyntheticrate (Kakinuma et al. 2006). U. lactuca markedlyenhances metabolic enzyme activities presumably aspart of an adaptation mechanism to extended peri-ods of emergence (Murthy et al. 1986, 1988, Murthyand Sharma 1989). Further work may consider theimpact of continuous versus discontinuous growthon Ulva cell-wall composition and structure.
In this study, the two species were absent or avail-able in low quantities to be collected between Octo-ber 2006 and February 2007. When biomass wasavailable, the monthly variations of U. armoricanaand U. rotundata chemical compositions and ulvanyield and chemical and functional characteristicswere likely consequences of variations in ecophysio-logical factors. The protein content in U. armoricanaand U. rotundata varied with that of seawater nitrate(data not shown) as previously reported for otheralgae (Black and Dewar 1949, Chapman and Craigie1977). However, no relationship was observedbetween reserve carbohydrates, tissue protein, andgrowth, as reported for other algae (Chapman andCraigie 1977), as starch contributed little to thetotal sugars in the two algae. The low levels of starchmeasured in ulvan from the two species suggest thatphotosynthate accumulation is limited and rapidly
Fig. 10. Correlation circle of Ulva armoricana and U. rotundataulvan chemical composition, gel conservation modulus G’, andproportions of macromolecular populations A, B, and C.
970 AUDREY ROBIC ET AL.

metabolized and ⁄ or converted to cell-wall polysac-charides.
Ulvan extraction yield expressed on the basis ofalgal dry weight varied with month of collection butwas likely biased by coextracted proteins for winter-collected U. rotundata samples. Ulvan accessibility wasalso observed to vary. For U. rotundata, it increasedfrom February to April 2007 during the active growthperiod of the alga and decreased thereafter. This vari-ation is interpreted on the basis of cell-wall polysac-charide interactions. During active growth of Ulva,newly deposited ulvan may not be tightly linked toother cell-wall components, whereas as growth rateslows, linkages are formed, and ulvan become lessprone to extraction by hot aqueous solutions ofsodium oxalate or hot water. Ulvan accessibility fromU. armoricana was particularly high in autumn, at theend of the proliferation period. This period coin-cided with the beginning of algal disintegration asso-ciated with modifications of cell-wall polysaccharidesinteractions. For the July 2007 sample, the high ulvanextraction efficiency was attributed to an active algalgrowth period. Reports from the ‘‘green-tide’’ moni-toring network of CEVA showed that algal prolifera-tion began late this year due to particular weatherand oceanographic conditions. In 2007 there was avery high nitrogen flux in comparison with previousyears (P. Dion, S. Ballu, and T. Perrot [CEVA],personal communication).
Seasonal variations in ulvan sugar compositionwere small and similar for the two species, exceptfor xylose in U. rotundata. The lowest xylose content(September–October and April) corresponded toperiods of active growth of U. rotundata. This find-ing indicates that actively growing algae synthesizemore A3S units and that synthesis of U units is devel-opmentally regulated. Glucuronic and iduronic acidcontents varied distinctly in the two algae, and glu-curonic acid proportion tended to be higher duringactive growth periods. This tendency suggests thatthe putative epimerase converting glucuronic acidto iduronic acid as proposed for animal glycosami-noglycan biosynthesis (Lahaye and Robic 2007) hasslow kinetics or its expression is also regulated. Incontrast with U. rotundata ulvan, no seasonal impactwas observed for U. armoricana ulvan fine structureas evaluated by ulvan lyase degradation profiles. Thewater column may dampen environmental variationsfor drifting seaweeds whose growth may be moreregular. Eutrophic conditions induced the persis-tence of spring conditions during a long periodwith a low C ⁄ N ratio.
The physicochemical and rheological propertiesof ulvans were markedly affected by the period ofcollection. The proportion of the high-molecular-weight population was the highest during theperiod of active growth of algae and then decreasedto the benefit of the medium- and low-molecular-weight populations. Taking into account all thechemical and physicochemical data together, the
ulvan gel storage modulus depends on the propor-tion of rhamnose, glucuronic acid, and sulfate andon the proportion of the high-molecular-weightpopulation A. Xylose-containing structures had nomajor impact on gelling characteristics probablydue to the overwhelming effect of molecular weightdistribution on the storage modulus. However, fur-ther work is needed to study the impact of the fineulvan structure and of proteins to ulvan rheologicalproperties.
This study allows recommendations to be pro-posed for an industrial ulvan production objective.If high-molecular-weight ulvans are required forgelling ⁄ thickening applications, seaweeds shouldthen be collected during their active growth peri-ods. Inversely, if ulvan production is for applicationsconcerning more specifically rare sugars andoligosaccharides, proliferating seaweeds could becollected at the end of the biomass productionperiod.
CONCLUSION
Like other texturing algal polysaccharides, ulvanchemical and macromolecular structures are geneti-cally controlled and modulated by growth condi-tions. U. armoricana and U. rotundata ulvans mainlydiffer in their proportion of ulvanobiose and in typeB ulvanobiuronic acid and molecular weight distri-butions. These differences may be genetic, but theyneed to be further evaluated in relation to possibleadaptations of the algae to their specific habitatsand growth conditions. Furthermore, consideringthe seasonal effects on ulvan chemistry, molecularweight, and functional properties, and in the lightof variations in red algal galactan chemistry andfunctional properties with the algal reproductive sta-tus (Craigie 1990), further work should consider theimpact of Ulva gametophyte and sporophyte lifephases on ulvan quality. From the viewpoint ofindustrial application, collection of Ulva for the pro-duction of gelling ulvan would be best during theactive growth periods of the algae and particularlyin spring. The gelling characteristics of an ulvanextract from a mixed population of Ulva speciescontributing to Brittany ‘‘green tides’’ would be lessaffected by small chemical structure differencesthan by variations in molecular weight distribution.
The authors wish to thank Laurine Buon from CEVA for hercontribution to this study. We also thank the RNE Service inCEVA for their helpful comments in the environmental analy-ses. We are grateful to INRA-BIA personnel: Dr. J.-L. Doublierand S. Durand for discussion and technical assistance for rhe-ological measurements, B. Quemener and R. Looten for assis-tance in carrying out the HPLC analyses and the starchanalysis, J. Vigouroux and M.-J. Crepeau for the technicalassistance, and Dr. D. Legland and Dr. M.-F. Devaux for theirhelpful advice on statistical analyses. This work was supportedin part by the French National Association for Research andTechnology (ANRT) and the General Council of Cotesd’Armor.
ULVAN VARIABILITY 971

Abdel-Fattah, A. F. & Edrees, M. 1973. Seasonal changes in theconstituents of Ulva lactuca. Phytochemistry 12:481–5.
Barcelo, A., Claustre, J., Moro, F., Chayvialle, J. A., Cuber, J. C. &Plaisancie, P. 2000. Mucin secretion is modulated by luminalfactors in the isolated vascularly perfused rat colon. Gut46:218–24.
Beach, K. S., Smith, C. M., Michael, T. M. & Shin, H. W. 1995.Photosynthesis in reproductive cells of Ulva fasciata andEnteromorpha flexuosa: implications for ecological success. Mar.Ecol. Prog. Ser. 125:129–37.
Black, W. A. P. & Dewar, E. T. 1949. Correlation of some of thephysical and chemical properties of the sea with the chemicalconstitution of the algae. J. Mar. Biol. Assoc. U. K. 28:673–99.
Blomster, J., Maggs, C. A. & Stanhope, M. J. 1998. Molecular andmorphological analysis of Enteromorpha intestinalis and E.compressa (Chlorophyta) in the British Isles. J. Phycol. 34:319–40.
Bobin-Dubigeon, C., Lahaye, M. & Barry, J. L. 1997. Human colonicbacterial degradability of dietary fibres from sea-lettuce (Ulvasp). J. Sci. Food Agric. 73:149–59.
Briand, X. & Morand, P. 1997. Anaerobic digestion of Ulva sp. 1.Relationship between Ulva composition and methanisation.J. Appl. Phycol. 9:511–24.
Chapman, A. R. O. & Craigie, J. S. 1977. Seasonal growth in Lam-inaria longicruris: relations with dissolved inorganic nutrientsand internal reserves of nitrogen. Mar. Biol. 40:197–205.
Cluzet, S., Torregrosa, C., Jacquet, C., Lafitte, C., Fournier, J.,Mercier, L., Salamagne, S., Briand, X., Esquerre-Tugaye, M. T.& Dumas, B. 2004. Gene expression profiling and protectionof Medicago truncatula against a fungal infection in response toan elicitor from green algae Ulva spp. Plant Cell Environ.27:917–28.
Coat, G., Dion, P., Noailles, M.-C., De Reviers, B., Fontaine, J.-M.,Berger-Perrot, Y. & Loiseaux-De Goer, S. 1998. Ulva armoricana(Ulvales, Chlorophyta) from the coasts of Brittany (France).II. Nuclear rDNA ITS sequence analysis. Eur. J. Phycol. 33:81–6.
Craigie, J. S. 1990. Cell walls. In Cole, K. M. & Sheath, R. G. [Eds.]Biology of the Red Algae. Cambridge University Press, Cam-bridge, UK, pp. 221–57.
Cuomo, V., Perretti, A., Palomba, I., Verde, A. & Cuomo, A. 1995.Utilisation of Ulva rigida biomass in the Venice Lagoon (Italy):biotransformation in compost. J. Appl. Phycol. 7:479–85.
Davison, I. R. & Pearson, G. A. 1996. Stress tolerance in intertidalseaweeds. J. Phycol. 32:197–211.
Dion, P. & Le Bozec, S. 1996. The French Atlantic Coasts. In Sch-ramm, W. & Nienhuis, P. H. [Eds.] Marine Benthic Vegetation.Springer, Berlin, pp. 251–64.
Englyst, H. N. & Cummings, J. H. 1988. Improved method ofmeasurement of dietary fiber as non-starch polysaccharides inplant foods. J. AOAC 71:808–14.
Fletcher, R. L. 1996. Recent changes and the effects of eutrophi-cation. In Schramm, W. & Nienhuis, P. H. [Eds.] Marine Ben-thic Vegetation. Springer, Berlin, pp. 7–43.
Haug, A. 1976. The influence of borate and calcium on the gelformation of a sulfated polysaccharide from Ulva lactuca. ActaChem. Scand. 30:562–6.
Hoebler, C., Barry, J.-L., David, A. & Delort-Laval, J. 1989.Rapid acid hydrolysis of plant cell wall polysaccharides andsimplified quantitative determination of their neutral mono-saccharides by gas–liquid chromatography. J. Agric. Food. Chem.37:360–5.
Hurd, C. L. & Dring, M. J. 1991. Desiccation and phosphate uptakeby intertidal fucoid algae in relation to zonation. Br. Phycol. J.26:327–33.
Ivanova, V., Rouseva, R., Kolarova, M., Serkedjieva, R., Rachev, R. &Manolova, N. 1994. Isolation of a polysaccharide with antiviraleffect from Ulva lactuca. Prep. Biochem. 24:83–97.
Kaeffer, B., Benard, C., Lahaye, M., Blottiere, H. M. & Cherbut, C.1999. Biological properties of ulvan, a new source of greenseaweed sulfated polysaccharides, on cultured normal andcancerous colonic epithelial cells. Planta Med. 65:527–31.
Kakinuma, M., Coury, D. A., Kuno, Y., Itoh, S., Kozawa, Y., Inagaki,E., Yoshiura, Y. & Amano, H. 2006. Physiological and
biochemical responses to thermal and salinity stresses in asterile mutant of Ulva pertusa (Ulvales, Chlorophyta). Mar. Biol.149:97–106.
Kloareg, B. & Quatrano, R. S. 1988. Structure of the cell wallsof marine algae and ecophysiological functions of thematrix polysaccharides. Oceanogr. Mar. Biol. Annu. Rev. 2:259–315.
Lahaye, M. 2001. Chemistry and physico-chemistry of phycocol-loids. Cah. Biol. Mar. 42:137–57.
Lahaye, M., Alvarez-Cabal Cimadevilla, E., Kuhlenkamp, R.,Quemener, B., Lognone, V. & Dion, P. 1999. Chemicalcomposition and 13C NMR spectroscopic characterisation ofulvans from Ulva (Ulvales, Chlorophyta). J. Appl. Phycol. 11:1–7.
Lahaye, M., Brunel, M. & Bonnin, E. 1997. Fine chemical structureanalysis of oligosaccharides produced by an ulvan-lyase deg-radation of the water-soluble cell wall polysaccharides fromUlva sp. (Ulvales, Chlorophyta). Carbohydr. Res. 304:325–33.
Lahaye, M., Inizan, F. & Vigouroux, J. 1998. NMR analysis of thechemical structure of ulvan and of ulvan-boron complex for-mation. Carbohydr. Polym. 36:239–49.
Lahaye, M. & Kaeffer, B. 1997. Seaweed dietary fibres: structure,physico-chemical and biological properties relevant to intesti-nal physiology. Sci. Aliments 17:563–84.
Lahaye, M. & Ray, B. 1996. Cell-wall polysaccharides from themarine green alga Ulva ‘‘rigida’’ (Ulvales, Chlorophyta). NMRanalysis of ulvan oligosaccharides. Carbohydr. Res. 283:161–73.
Lahaye, M. & Robic, A. 2007. Structure and functional properties ofUlvan, a polysaccharide from green seaweeds. Biomacromole-cules 8:1765–74.
Laza, A. L., Jaber, M., Miehe-Brendle, J., Demais, H., Le Deit, H.,Delmotte, L. & Vidal, L. 2007. Green nanocomposites: syn-thesis and characterization. J. Nanosci. Nanotechnol. 7:3207–13.
Littler, M. M. & Littler, D. S. 1980. The evolution of thallus formand survival strategies in benthic marine macroalgae: fieldand laboratory tests of a functional form model. Am. Nat.116:25–44.
Luning, K. 1990. Seaweeds, Their Environment, Biogeography, andEcophysiology. John Wiley & Sons, New York, 527 pp.
Mao, W., Zang, X., Li, Y. & Zhang, H. 2006. Sulfated polysaccharidefrom marine green algae Ulva conglobata and their anticoagu-lant activity. J. Appl. Phycol. 18:9–14.
Maze, J., Morand, P. & Potoky, P. 1993. Stabilisation of ‘green tides’Ulva by a method of composting with a view to pollutionlimitation. J. Appl. Phycol. 5:183–90.
McCleary, B. V., Solah, V. & Gibson, T. S. 1994. Quantitative mea-surement of total starch in cereal flours and products. J. CerealSci. 20:51–8.
Medcalf, D. G., Lionel, T., Brannon, J. H. & Scott, J. R. 1975. Sea-sonal variation in the mucilaginous polysaccharides from Ulvalactuca. Bot. Mar. 18:67–70.
Menesguen, A. 2003. Les ‘‘marees vertes’’ en Bretagne, la responsabilitedu nitrate. IFREMER, Plouzane, France, 11 pp.
Morand, P. & Briand, X. 1996. Excessive growth of macroalgae:a symptom of environmental disturbance. Bot. Mar. 39:491–516.
Murthy, M. S., Rao, A. S. & Faldu, P. J. 1988. Invertase and totalamylase activities of Ulva lactuca from different tidal levels,under desiccation. Bot. Mar. 31:53–6.
Murthy, M. S., Rao, A. S. & Reddy, E. R. 1986. Dynamics of nitratereductase activity in two intertidal algae under desiccation. Bot.Mar. 24:471–4.
Murthy, M. S. & Sharma, C. L. N. S. 1989. Peroxidase activity in Ulvalactuca under desiccation. Bot. Mar. 32:511–3.
Nicolucci, C. & Monegato, A. 1993. Procedure for manufacturingpaper from seaweed and paper thus obtained. European Pat-ent, EP 0 565 920 A1.
Percival, E. & McDowell, R. H. 1967. Chemistry and Enzymology ofMarine Algal Polysaccharides. Academic Press, London, 219 pp.
Perez, R. 1997. Ces algues qui nous entourent. Conception actuelle, roledans la biosphere, utilisations, culture. Ed. IFREMER, Plouzane,France, 272 pp.
Quadir, A., Harrison, P. J. & DeWreede, R. E. 1979. The effects ofemergence and submergence on the photosynthesis and res-piration of marine macrophytes. Phycologia 18:83–8.
972 AUDREY ROBIC ET AL.

Quemener, B., Lahaye, M. & Bobin-Dubigeon, C. 1997. Sugardetermination in ulvans by a chemical-enzymatic methodcoupled to high performance anion exchange chromatogra-phy. J. Appl. Phycol. 9:179–88.
Quemener, B., Marot, C., Mouillet, L., Da Riz, V. & Diris, J. 2000.Quantitative analysis of hydrocolloids in food systems bymethanolysis coupled to reverse HPLC. Part 1. Gelling car-rageenans. Food Hydrocoll. 14:9–17.
Ray, B. & Lahaye, M. 1995a. Cell-wall polysaccharides from the mar-ine green alga Ulva rigida (Ulvales, Chlorophyta). Extractionand chemical composition. Carbohydr. Res. 274:251–61.
Ray, B. & Lahaye, M. 1995b. Cell-wall polysaccharides from themarine green alga Ulva rigida (Ulvales, Chlorophyta). Chemi-cal structure of ulvan. Carbohydr. Res. 274:313–8.
Robic, A., Sassi, J.-F. & Lahaye, M. 2008. Impact of stabilizationtreatments of the green seaweed Ulva rotundata (Chlorophyta)
on the extraction yield, the physico-chemical and rheologicalproperties of ulvan. Carbohydr. Polym. 74:344–52.
Siddhanta, A. K., Goswami, A. M., Ramavat, B. K., Mody, K. H. &Mairh, O. P. 2001. Water soluble polysaccharides of marinealgal species of Ulva (Ulvales, Chlorophyta) of Indian waters.Indian J. Mar. Sci. 30:166–72.
Smith, G. M. 1947. On the reproduction of some Pacific coastspecies of Ulva. Am. J. Bot. 34:80–7.
Tabatabai, M. A. 1974. Determination of sulfate in water samples.Sulphur Institute 10:11–3.
Thibault, J.-F. 1979. Automatisation du dosage des substancespectiques par la methode au meta-hydroxydiphenyl. Lebensm-Wiss. Technol. 12:247–51.
Thomas, T. E. & Turpin, D. H. 1980. Desiccation enhanced nutri-ent uptake rates in the intertidal alga Fucus distichus. Bot. Mar.23:479–81.
ULVAN VARIABILITY 973




















