
Microbial Colonization and Competition on the Marine Alga Ulva australis
10
10.1128/AEM.00449-06. 2006, 72(8):5547. DOI: Appl. Environ. Microbiol. Dhana Rao, Jeremy S. Webb and Staffan Kjelleberg Ulva australis the Marine Alga Microbial Colonization and Competition on http://aem.asm.org/content/72/8/5547 Updated information and services can be found at: These include: REFERENCES http://aem.asm.org/content/72/8/5547#ref-list-1 at: This article cites 58 articles, 26 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on November 12, 2013 by guest http://aem.asm.org/ Downloaded from on November 12, 2013 by guest http://aem.asm.org/ Downloaded from
Transcript of Microbial Colonization and Competition on the Marine Alga Ulva australis

10.1128/AEM.00449-06.
2006, 72(8):5547. DOI:Appl. Environ. Microbiol. Dhana Rao, Jeremy S. Webb and Staffan Kjelleberg
Ulva australisthe Marine Alga Microbial Colonization and Competition on
http://aem.asm.org/content/72/8/5547Updated information and services can be found at:
These include:
REFERENCEShttp://aem.asm.org/content/72/8/5547#ref-list-1at:
This article cites 58 articles, 26 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 2006, p. 5547–5555 Vol. 72, No. 80099-2240/06/$08.00�0 doi:10.1128/AEM.00449-06Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Microbial Colonization and Competition on the Marine Alga Ulva australisDhana Rao, Jeremy S. Webb,† and Staffan Kjelleberg*
School of Biotechnology and Biomolecular Sciences and Centre for Marine Biofouling and Bio-innovation,University of New South Wales, Sydney, New South Wales 2052, Australia
Received 23 February 2006/Accepted 10 May 2006
Pseudalteromonas tunicata and Roseobacter gallaeciensis are biofilm-forming marine bacteria that are oftenfound in association with the surface of the green alga Ulva australis. They are thought to benefit the plant hostby producing inhibitory compounds that are active against common fouling organisms. We investigated factorsthat influence the ability of P. tunicata and R. gallaeciensis to attach to and colonize the plant surface and alsothe competitive interactions that occur between these organisms and other isolates from U. australis duringbiofilm formation on the plant surface. A surprisingly high number of P. tunicata cells, at least 108 cells ml�1,were required for colonization and establishment of a population of cells that persists on axenic surfaces of U.australis. Factors that enhanced colonization of P. tunicata included inoculation in the dark and pregrowth ofinocula in medium containing cellobiose as the sole carbon source (cellulose is a major surface polymer of U.australis). It was also found that P. tunicata requires the presence of a mixed microbial community to colonizeeffectively. In contrast, R. gallaeciensis effectively colonized the plant surface under all conditions tested. Studiesof competitive interactions on the plant surface revealed that P. tunicata was numerically dominant comparedwith all other bacterial isolates tested (except R. gallaeciensis), and this dominance was linked to production ofthe antibacterial protein AlpP. Generally, P. tunicata was able to coexist with competing strains, and eachstrain existed as microcolonies in spatially segregated regions of the plant. R. gallaeciensis was numericallydominant compared with all strains tested and was able to invade and disperse preestablished biofilms. Thisstudy highlighted the fact that microbial colonization of U. australis surfaces is a dynamic process anddemonstrated the differences in colonization strategies exhibited by the epiphytic bacteria P. tunicata and R.gallaeciensis.
Biofouling is caused initially by bacterial growth and biofilmformation on natural and artificial surfaces. Bacterial biofilmsfacilitate the attachment and growth of a range of other foulingorganisms, such as diatoms, invertebrate larvae, and algalspores (18). Strategies to prevent bacterial biofilms, therefore,represent a powerful approach to the prevention of biofouling.
Biofouling communities on surfaces of marine plants andanimals can have detrimental effects on the host organism.Physical damage to the host results from the production oftoxins, digestive enzymes, and waste products by the microbialcommunity. However, the extent of biofouling on marine or-ganisms is markedly less than the extent of biofouling on in-animate structures. Often the later stages of biofouling, such asthe attachment of algae and barnacles, do not occur (2). Incontrast to inanimate surfaces, which are colonized in a rapidand predictable manner by a diverse assemblage of marinemicrobes (62), biotic surfaces frequently harbor species-spe-cific microbial communities (22, 61, 62). These communitiescan be variable and distinct from those found in the surround-ing environment. Thus, many algal and invertebrate species areable to regulate the bacterial colonization of their surfaces (29,32, 62).
Microalgal surfaces are typically covered by bacteria at den-
sities of approximately 107 bacteria cm�2 (3). Studies of theepiphytic microbial communities present on macroalgae haveemphasized the spatial distribution of bacteria, with specificparts of the thallus hosting specific bacterial populations (9). Insome cases the bacterial populations change with the season orthe age of the host (55, 56). Associations between algae andbacteria are common, and studies have generally focused onthe benefits provided to the bacteria, such as support of bac-terial growth by dissolved organic carbon released by algal cells(30, 34, 49).
Although some of the bacterium-alga interactions have beencharacterized, the ecological significance of most naturally oc-curring epiphytic bacterial communities is unclear, and inmany cases the bacterial species involved have not been iden-tified (17). It has been demonstrated that bacterial biofilms arepresent on the surface of Ulva australis (formerly known asUlva lactuca) (54), and it has been speculated that the seaweeduses microbial defense to protect against fouling (24, 27). U.australis has no known physical or chemical defense systemsagainst fouling organisms, and it has been suggested that thehost may manipulate the bacterial community on its surface,which in turn protects the host by interfering with the devel-opment of a mature biofouling community. Such interactionsare not uncommon in the marine environment (for reviews seereferences 4, 7, 15, 19, 26, and 44).
One important species of marine bacteria that is found inassociation with U. australis is Pseudoalteromonas tunicata.This bacterium produces a diverse range of biologically activecompounds that specifically target marine fouling organisms(25). One of these compounds is a 190-kDa multisubunit an-
* Corresponding author. Mailing address: School of Biotechnologyand Biomolecular Sciences and Centre for Marine Biofouling andBio-innovation, Biological Sciences Building, University of New SouthWales, Sydney, NSW 2052, Australia. Phone: 61 (2) 9385 2102. Fax: 61(2) 9385 1779. E-mail: [email protected].
† Present address: School of Biological Sciences, University ofSouthampton, Southampton SO16 7PX, United Kingdom.
5547
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from

tibacterial protein designated AlpP which is effective againstboth gram-negative and gram-positive bacteria from a range ofenvironments (28, 37). Recent work has demonstrated thatAlpP can provide a competitive advantage to P. tunicata duringbiofilm growth in laboratory biofilm experiments (48).
Roseobacter gallaeciensis is also frequently isolated from thesurface of U. australis. Roseobacter spp. are cosmopolitan andhave been isolated, for example, from green seaweeds (53),marine snow particles (22), and dinoflagellates (1, 33, 39).Roseobacter spp. are able to metabolize dimethylsulfoniopro-pionate (DMSP), and their presence and activity on algal sur-faces are significantly correlated with DMSP-producing algae,including dinoflagellates and prymnesiophytes (20).
The extensive, highly diverse microbial community associ-ated with U. australis makes it an interesting study organism foraddressing questions of surface colonization and host associa-tion. The dynamics of surface colonization in natural systems,particularly during the early stages of biofilm establishment,are poorly understood for marine bacteria. For example, anassessment of the ability of distinct bacteria to colonize algalsurfaces would be useful for identifying bacterial traits thatcontribute to epiphytic fitness. We recently described compet-itive biofilm interactions of P. tunicata and other marine strainsin a laboratory glass flow cell system (48). However, little isknown about the ecology of colonization and competition on aliving surface, including whether P. tunicata is a dominantcompetitor in ecologically relevant settings.
In this study we aimed to investigate, for the first time, thecolonization biology of marine bacteria on the surface of amarine plant, in this case U. australis. We investigated thehypothesis that P. tunicata and R. gallaeciensis are effectivecolonizers of U. australis and are able to compete with anddominate other marine bacterial isolates during biofilm forma-tion on the plant surface. We found that P. tunicata requiresthe presence of a natural seawater community to colonizeeffectively, whereas R. gallaeciensis is an aggressive colonizerunder all conditions tested. Below we highlight the differencesin colonization strategies exhibited by the epiphytic strains ofP. tunicata and R. gallaeciensis and demonstrate that antibac-terial compounds have a role in the colonization of U. australis.This study demonstrated that microbial colonization of theplant surfaces is a dynamic process in which differences inattachment, colonization, and competitive biofilm formationcan markedly affect the establishment and organization ofepiphytic microbial communities.
MATERIALS AND METHODS
Collection of plants and generation of axenic U. australis. The common marinealga U. australis was collected from the rocky intertidal zone (latitude, 151.2572;longitude, �33.9121) near Sydney, Australia. The method used to make U.australis axenic was modified from the method described by Scheffel (51). Briefly,plants were rinsed in 50 ml autoclaved seawater and cut into 5-cm-long frag-ments. These fragments were swabbed with cotton buds, and 0.6-cm disks werepunched out of the thallus using a cork borer. The disks were rinsed in auto-claved seawater and incubated in 0.012% NaOCl for 5 min. The disks wereallowed to recover in sterile seawater for 1 h before they were incubated in anantibiotic cocktail (300 mg liter�1 ampicillin, 30 mg liter�1 polymyxin B, and 60mg literl�1 gentamicin dissolved in sterile seawater) for 18 h in 24-well tissueculture plates (Sarstedt). Disks were then incubated in 0.008% NaOCl for 5 min.These disks were allowed to recover in sterile seawater for at least 3 h to removetraces of oxidants, suspended in 2 ml of sterile seawater, and agitated at roomtemperature at 60 rpm.
Plant tissue viability was examined by staining treated U. australis disks with0.25% (wt/vol) Evans blue (60). This blue dye penetrates into dead and damagedplant cells, while intact cells are able to exclude the dye. All colonization andcompetition experiments were done in triplicate, and three to five individualdisks were randomly sampled at each sampling time.
Colonization experiments. Bacterial strains were isolated from the surface ofU. lactuca as described previously (48). Cultures were stored at �80°C in 50%(vol/vol) glycerol in VNSS medium (38) and maintained on VNSS mediumplates. P. tunicata and R. gallaeciensis were labeled with a green fluorescentprotein (GFP) color tag as described previously (48). Bacteria were cultured for24 h at 25°C in VNSS broth for preparation of inocula. Cells were harvested byspinning down the culture and resuspending the pellet in seawater. The cellconcentration was estimated by counting cells by epifluorescence microscopyusing a hemocytometer and was adjusted to the desired level by dilution withseawater. Experiments were conducted in Sarstedt 24-well plates with axenicdisks, and the bacteria were applied by immersing the disks in a suspension of P.tunicata and R. gallaeciensis for 3 h. The disks were then rinsed twice in filteredseawater and transferred to fresh Sarstedt 24-well plates containing 2 ml offiltered seawater. The plates were incubated on a shaker at 60 rpm and 25°C for16 h in the presence of light.
Factors affecting colonization. In order to investigate some of the factors thatwere essential for colonization of P. tunicata, we tested a range of conditions.
(i) Density of P. tunicata required for attachment. P. tunicata was inoculatedonto axenic U. australis disks in 24-well tissue culture plates at a range ofconcentrations (104 to 108 cells ml�1). The disks were incubated for 3 h, rinsedtwice in sterile seawater, and then incubated at 25°C and 60 rpm for the durationof the experiment.
(ii) Axenic versus nonaxenic plant surfaces. The attachment of P. tunicatacells which had been inoculated at a concentration of 107 cells ml�1 was com-pared on the both axenic and nonaxenic surfaces of U. australis disks.
(iii) U. australis disks versus whole plants. Attachment was tested on axenicdisks in 24-well plates, as well as on axenic whole plants (which had been treatedin the same way as the disks). The plants were oriented upright in beakers andimmersed in filtered seawater.
(iv) Dark versus light. Preparations were inoculated in the dark at a concen-tration of 107 cells ml�1 and then incubated in the dark for 3 h. This procedurewas compared to inoculation at the same density and incubation for 3 h indaylight.
(v) Carbon source. The standard culture conditions for marine strains includedVNSS medium containing glucose. However, we also grew cells for 48 h inminimal medium (45) with cellobiose as the sole carbon source in a still culture.The cells were inoculated onto U. australis disks, and attachment was comparedto the attachment of cells grown in minimal medium with trehalose and glucoseas carbon sources.
(vi) Incubation time. Disks were inoculated with 107 cells ml�1 and incubatedfor 1, 3, 12, and 24 h.
(vii) Effect of multispecies consortia on colonization. Axenic algal disks wereinoculated with overnight cultures of GFP-labeled P. tunicata and 17 unlabeledstrains isolated from the surface of U. australis (48) in filtered seawater. P.tunicata was also inoculated into a natural seawater community. The resultingmixed-species biofilm was visualized by staining with acridine orange. Attach-ment and colonization of GFP-labeled P. tunicata within the mixed biofilm werevisualized by observing the unstained biofilm with a confocal microscope.
To test all factors, a density of 107 cells ml�1 was used unless otherwise stated,and P. tunicata cells were suspended in filtered seawater. Attachment, biofilmformation, and microcolony development were monitored by epifluorescencemicroscopy. The times when preparations were examined were immediately afterinoculation and 1, 3, and 7 days following inoculation. At each sampling time,three disks were randomly selected, and a total of 12 random fields of view werecounted for each disk. Samples were viewed by epifluorescence microscopy withan Axiophot microscope equipped with a 10� 1.30-numerical-aperture objective(Leica).
Although preliminary experiments indicated that R. gallaeciensis was an ag-gressive colonizer of U. australis, we also tested attachment and colonization ofthis organism under all the experimental conditions used for P. tunicata.
Competition in dual-species biofilms on U. australis. Other marine organismsused in dual-species competition experiments were Pseudalteromonas gracilis,Alteromonas sp., and Cellulophaga fucicola, all of which were isolated from U.australis as described previously (48). The strains were labeled with red fluores-cent protein (DsRed) and GFP as described previously (48). The growth rates ofwild-type marine isolates and their labeled derivatives on the surface of axeni-cally treated U. australis did not differ significantly (data not shown).
Overnight cultures of bacteria were inoculated onto U. australis at a density of
5548 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from
108 cells ml�1 in filtered and natural seawater and incubated for 3 h withoutshaking at room temperature. Disks were rinsed three times with filtered sea-water, transferred to sterile seawater in Sarstedt 24-well plates, and incubated at25°C at 60 rpm for the duration of the experiment. Unlabeled biofilms werevisualized on the surface of the plant by staining with acridine orange. Biofilmswere grown in filtered seawater and visualized with a confocal laser scanningmicroscope (Olympus) using fluorescein isothiocyanate and tetramethyl rhoda-mine isocyanate optical filters for green and red fluorescent proteins, respec-tively. Labeled P. tunicata was inoculated onto the surface with competing ma-rine strains, including R. gallaeciensis. To investigate whether the antibacterialprotein produced by P. tunicata improved the competitiveness of this organismon the surface of U. australis, competition studies were also performed with theAlpP mutant of P. tunicata (37).
To cultivate mixed biofilms, disks were inoculated with a 500-�l suspension ofstationary-phase cultures of P. tunicata and the competitive marine strain infiltered seawater. In order to ensure that the initial ratio of attached cells of thetwo competing strains was 1:1, we first monitored the initial attachment of themixed culture after a 3-h adhesion period. In most cases, the initial levels ofattachment to the plant surface were deemed to be approximately equal, asdetermined by counting the numbers of red- and green-labeled cells by epiflu-orescence microscopy.
To investigate whether P. tunicata and R. gallaeciensis were able to invade andcolonize an established biofilm, marine strains (P. tunicata, C. fucicola, andAlteromonas sp.) were allowed to preestablish a biofilm for 48 h on U. australis.The preformed biofilm was inoculated with �108 cells of wild-type P. tunicata orR. gallaeciensis, incubated for 3 h without shaking, and then incubated at 60 rpmfor the duration of the experiment. After the resumption of shaking, the biofilmwas examined for red and green fluorescence. Experiments were repeated inthree separate rounds with three independent 24-well plates run in parallel. Theviability of bacterial cells was determined using a Live-Dead staining kit (Mo-lecular Probes Inc., Eugene, Oreg.).
RESULTS
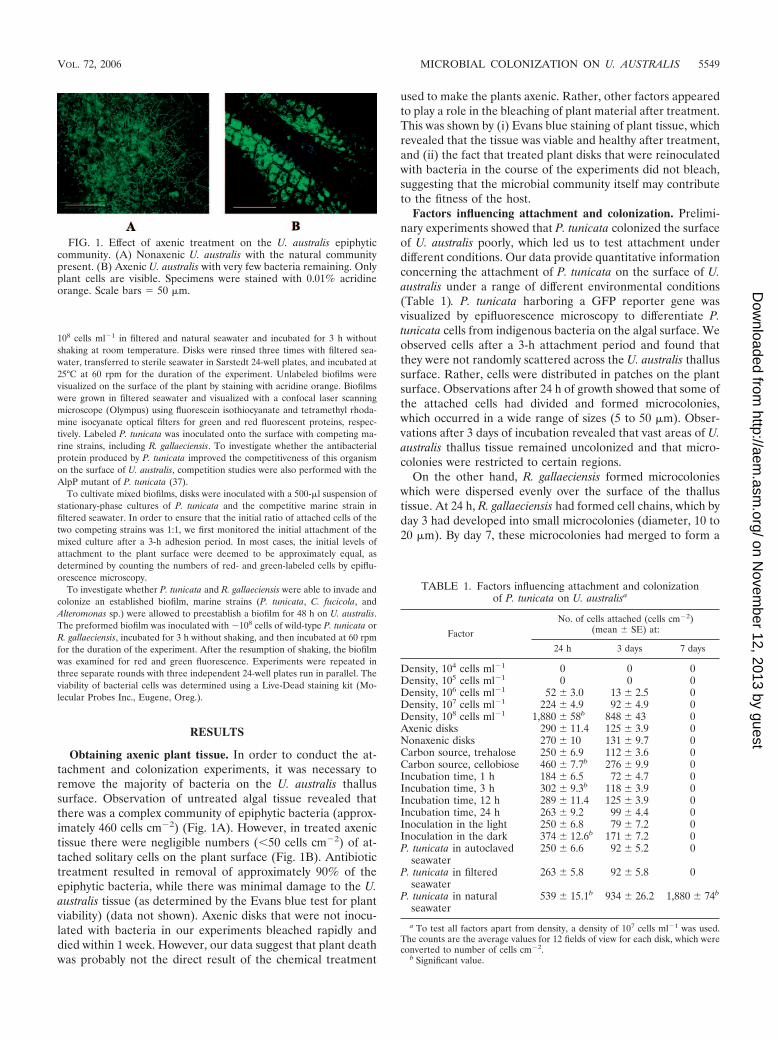
Obtaining axenic plant tissue. In order to conduct the at-tachment and colonization experiments, it was necessary toremove the majority of bacteria on the U. australis thallussurface. Observation of untreated algal tissue revealed thatthere was a complex community of epiphytic bacteria (approx-imately 460 cells cm�2) (Fig. 1A). However, in treated axenictissue there were negligible numbers (�50 cells cm�2) of at-tached solitary cells on the plant surface (Fig. 1B). Antibiotictreatment resulted in removal of approximately 90% of theepiphytic bacteria, while there was minimal damage to the U.australis tissue (as determined by the Evans blue test for plantviability) (data not shown). Axenic disks that were not inocu-lated with bacteria in our experiments bleached rapidly anddied within 1 week. However, our data suggest that plant deathwas probably not the direct result of the chemical treatment
used to make the plants axenic. Rather, other factors appearedto play a role in the bleaching of plant material after treatment.This was shown by (i) Evans blue staining of plant tissue, whichrevealed that the tissue was viable and healthy after treatment,and (ii) the fact that treated plant disks that were reinoculatedwith bacteria in the course of the experiments did not bleach,suggesting that the microbial community itself may contributeto the fitness of the host.
Factors influencing attachment and colonization. Prelimi-nary experiments showed that P. tunicata colonized the surfaceof U. australis poorly, which led us to test attachment underdifferent conditions. Our data provide quantitative informationconcerning the attachment of P. tunicata on the surface of U.australis under a range of different environmental conditions(Table 1). P. tunicata harboring a GFP reporter gene wasvisualized by epifluorescence microscopy to differentiate P.tunicata cells from indigenous bacteria on the algal surface. Weobserved cells after a 3-h attachment period and found thatthey were not randomly scattered across the U. australis thallussurface. Rather, cells were distributed in patches on the plantsurface. Observations after 24 h of growth showed that some ofthe attached cells had divided and formed microcolonies,which occurred in a wide range of sizes (5 to 50 �m). Obser-vations after 3 days of incubation revealed that vast areas of U.australis thallus tissue remained uncolonized and that micro-colonies were restricted to certain regions.
On the other hand, R. gallaeciensis formed microcolonieswhich were dispersed evenly over the surface of the thallustissue. At 24 h, R. gallaeciensis had formed cell chains, which byday 3 had developed into small microcolonies (diameter, 10 to20 �m). By day 7, these microcolonies had merged to form a
TABLE 1. Factors influencing attachment and colonizationof P. tunicata on U. australisa
Factor
No. of cells attached (cells cm�2)(mean � SE) at:
24 h 3 days 7 days
Density, 104 cells ml�1 0 0 0Density, 105 cells ml�1 0 0 0Density, 106 cells ml�1 52 � 3.0 13 � 2.5 0Density, 107 cells ml�1 224 � 4.9 92 � 4.9 0Density, 108 cells ml�1 1,880 � 58b 848 � 43 0Axenic disks 290 � 11.4 125 � 3.9 0Nonaxenic disks 270 � 10 131 � 9.7 0Carbon source, trehalose 250 � 6.9 112 � 3.6 0Carbon source, cellobiose 460 � 7.7b 276 � 9.9 0Incubation time, 1 h 184 � 6.5 72 � 4.7 0Incubation time, 3 h 302 � 9.3b 118 � 3.9 0Incubation time, 12 h 289 � 11.4 125 � 3.9 0Incubation time, 24 h 263 � 9.2 99 � 4.4 0Inoculation in the light 250 � 6.8 79 � 7.2 0Inoculation in the dark 374 � 12.6b 171 � 7.2 0P. tunicata in autoclaved
seawater250 � 6.6 92 � 5.2 0
P. tunicata in filteredseawater
263 � 5.8 92 � 5.8 0
P. tunicata in naturalseawater
539 � 15.1b 934 � 26.2 1,880 � 74b
a To test all factors apart from density, a density of 107 cells ml�1 was used.The counts are the average values for 12 fields of view for each disk, which wereconverted to number of cells cm�2.
b Significant value.
FIG. 1. Effect of axenic treatment on the U. australis epiphyticcommunity. (A) Nonaxenic U. australis with the natural communitypresent. (B) Axenic U. australis with very few bacteria remaining. Onlyplant cells are visible. Specimens were stained with 0.01% acridineorange. Scale bars � 50 �m.
VOL. 72, 2006 MICROBIAL COLONIZATION ON U. AUSTRALIS 5549
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from
relatively flat, mat-like biofilm, which covered a large propor-tion of the algal surface. Also in contrast to P. tunicata, R.gallaeciensis cells attached and colonized effectively under allconditions tested. At cell densities greater than 106 cells ml�1,colonization of U. australis by R. gallaeciensis appeared to besimilar in all situations (data not shown). Furthermore, at-tached cells persisted indefinitely on algal disks. Compared tothe analysis of P. tunicata, the smaller cell size and the highrates of cell attachment of R. gallaeciensis made counting dif-ficult; hence, cell attachment was not quantified.
(i) Effects of inoculum cell density on colonization of U.australis. Direct in situ observation of epiphytic bacteria re-vealed that the density of cells in the inoculum affected micro-colony formation. After an initial attachment period (3 h), weincubated disks for 24 h and then examined the plant surfacefor bacterial cells and microcolony formation. Surprisingly, noattachment took place when P. tunicata was inoculated at lowdensities (104 or 105 cells ml�1) (Table 1). At a density of 106
cells ml�1 we observed that few cells attached (52 cells cm�2).These cells were mostly solitary; no microcolonies formed, andcells usually did not persist for more than 3 days. At a densityof 107 cells ml�1 some of the attached cells were clusteredtogether and formed small microcolonies consisting of about15 to 20 cells after 24 h. At a density of 108 cells ml�1, micro-colonies were considerably larger (up to 100 �m across) afterthe same growth period (24 h). In spite of the large sizes ofmicrocolonies obtained with high cell densities, P. tunicata didnot persist for more than 5 days. Although R. gallaeciensis wasable to colonize at lower densities (104 and 105 cells ml�1), thepersistence was much greater at higher densities (106 to108cells ml�1). At higher densities R. gallaeciensis rapidlyformed microcolonies that survived and persisted indefinitelyon the plant, compared to the colonies that formed at a lowdensity, which did not remain for more than 5 or 6 days. Thus,
both organisms exhibited density-dependent colonization andenhanced microcolony formation at high cell densities.
(ii) Colonization in the dark. On the basis of our observationof surprisingly low levels of attachment of P. tunicata on theplant surface, we investigated the possibility that P. tunicataattachment may be affected by the presence of DMSP on theplant surface. It is known that DMSP is released in senescentalgae or when algae undergo oxidative stress, particularly un-der high light intensities (31). We therefore examined coloni-zation of U. australis by P. tunicata in the dark. We observedthat attachment of P. tunicata improved when cells were inoc-ulated in the dark, and the number of cells that attachedincreased from 250 cells cm�2 to 374 cells cm�2 (Table 1).However, the cells did not persist and form biofilms. Most ofthe microcolonies formed were small, although the numberwas greater (about 16 to 20 microcolonies per cm�2 with 12 to25 cells in each microcolony, compared to about 3 to 6 micro-colonies per cm�2 with 50 to 90 cells in each microcolony forplant tissue incubated in the presence of light). We repeatedthese sets of experiments using the Live-Dead stain to establishwhether cells in smaller microcolonies died or whether theysimply failed to attach. However, the results were inconclusiveas it was difficult to visualize the dead cells against the redautofluorescence of the algae. In contrast, R. gallaeciensis wasable to attach and colonize at a density of 106 cells ml�1
regardless of whether the preparations were inoculated in thelight or in the dark.
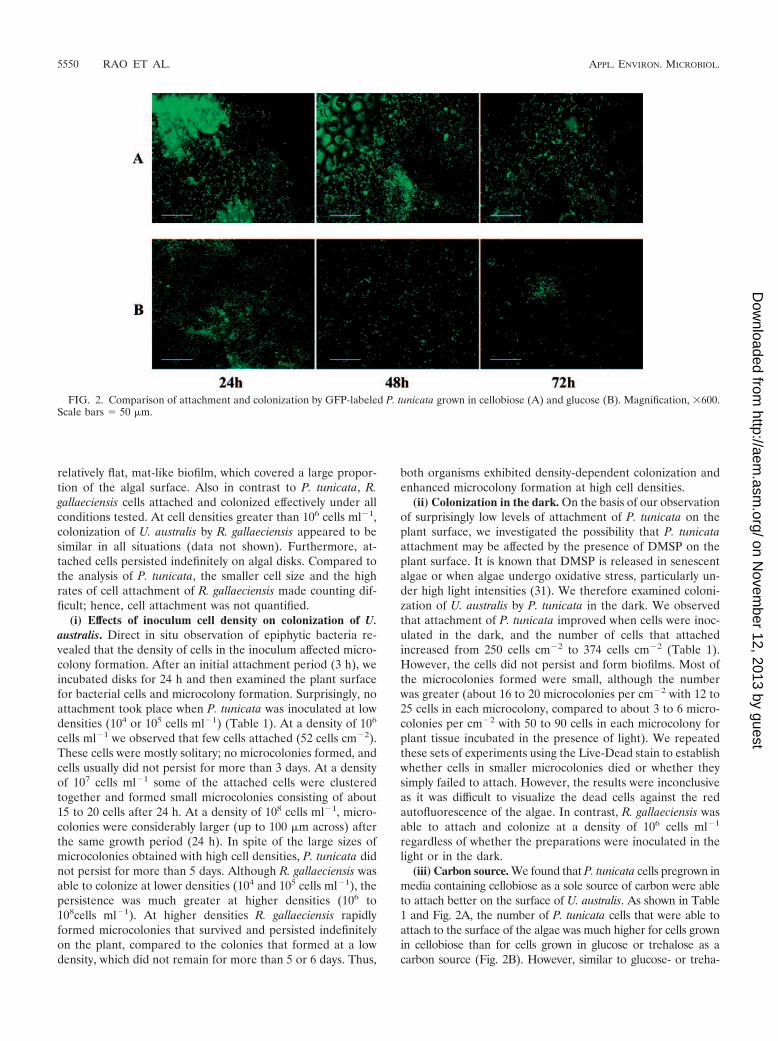
(iii) Carbon source. We found that P. tunicata cells pregrown inmedia containing cellobiose as a sole source of carbon were ableto attach better on the surface of U. australis. As shown in Table1 and Fig. 2A, the number of P. tunicata cells that were able toattach to the surface of the algae was much higher for cells grownin cellobiose than for cells grown in glucose or trehalose as acarbon source (Fig. 2B). However, similar to glucose- or treha-
FIG. 2. Comparison of attachment and colonization by GFP-labeled P. tunicata grown in cellobiose (A) and glucose (B). Magnification, �600.Scale bars � 50 �m.
5550 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from
lose-grown cells, the cellobiose-grown cells did not persist formore than 5 days on the plant surface.
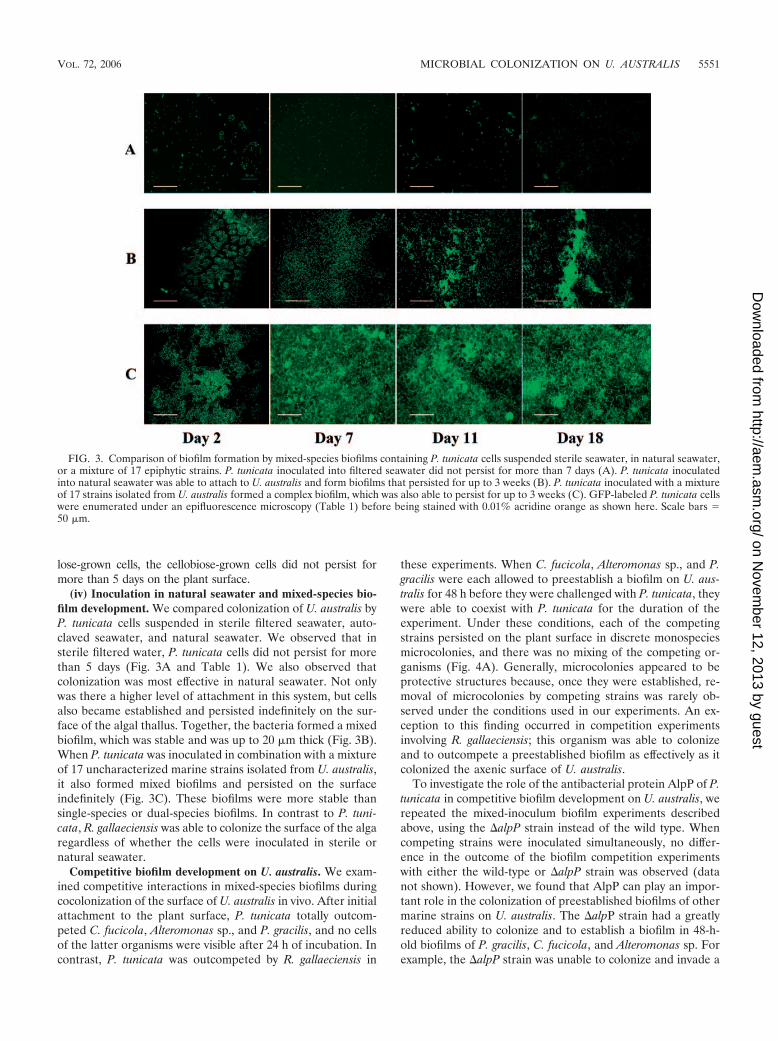
(iv) Inoculation in natural seawater and mixed-species bio-film development. We compared colonization of U. australis byP. tunicata cells suspended in sterile filtered seawater, auto-claved seawater, and natural seawater. We observed that insterile filtered water, P. tunicata cells did not persist for morethan 5 days (Fig. 3A and Table 1). We also observed thatcolonization was most effective in natural seawater. Not onlywas there a higher level of attachment in this system, but cellsalso became established and persisted indefinitely on the sur-face of the algal thallus. Together, the bacteria formed a mixedbiofilm, which was stable and was up to 20 �m thick (Fig. 3B).When P. tunicata was inoculated in combination with a mixtureof 17 uncharacterized marine strains isolated from U. australis,it also formed mixed biofilms and persisted on the surfaceindefinitely (Fig. 3C). These biofilms were more stable thansingle-species or dual-species biofilms. In contrast to P. tuni-cata, R. gallaeciensis was able to colonize the surface of the algaregardless of whether the cells were inoculated in sterile ornatural seawater.
Competitive biofilm development on U. australis. We exam-ined competitive interactions in mixed-species biofilms duringcocolonization of the surface of U. australis in vivo. After initialattachment to the plant surface, P. tunicata totally outcom-peted C. fucicola, Alteromonas sp., and P. gracilis, and no cellsof the latter organisms were visible after 24 h of incubation. Incontrast, P. tunicata was outcompeted by R. gallaeciensis in
these experiments. When C. fucicola, Alteromonas sp., and P.gracilis were each allowed to preestablish a biofilm on U. aus-tralis for 48 h before they were challenged with P. tunicata, theywere able to coexist with P. tunicata for the duration of theexperiment. Under these conditions, each of the competingstrains persisted on the plant surface in discrete monospeciesmicrocolonies, and there was no mixing of the competing or-ganisms (Fig. 4A). Generally, microcolonies appeared to beprotective structures because, once they were established, re-moval of microcolonies by competing strains was rarely ob-served under the conditions used in our experiments. An ex-ception to this finding occurred in competition experimentsinvolving R. gallaeciensis; this organism was able to colonizeand to outcompete a preestablished biofilm as effectively as itcolonized the axenic surface of U. australis.
To investigate the role of the antibacterial protein AlpP of P.tunicata in competitive biofilm development on U. australis, werepeated the mixed-inoculum biofilm experiments describedabove, using the alpP strain instead of the wild type. Whencompeting strains were inoculated simultaneously, no differ-ence in the outcome of the biofilm competition experimentswith either the wild-type or alpP strain was observed (datanot shown). However, we found that AlpP can play an impor-tant role in the colonization of preestablished biofilms of othermarine strains on U. australis. The alpP strain had a greatlyreduced ability to colonize and to establish a biofilm in 48-h-old biofilms of P. gracilis, C. fucicola, and Alteromonas sp. Forexample, the alpP strain was unable to colonize and invade a
FIG. 3. Comparison of biofilm formation by mixed-species biofilms containing P. tunicata cells suspended sterile seawater, in natural seawater,or a mixture of 17 epiphytic strains. P. tunicata inoculated into filtered seawater did not persist for more than 7 days (A). P. tunicata inoculatedinto natural seawater was able to attach to U. australis and form biofilms that persisted for up to 3 weeks (B). P. tunicata inoculated with a mixtureof 17 strains isolated from U. australis formed a complex biofilm, which was also able to persist for up to 3 weeks (C). GFP-labeled P. tunicata cellswere enumerated under an epifluorescence microscopy (Table 1) before being stained with 0.01% acridine orange as shown here. Scale bars �50 �m.
VOL. 72, 2006 MICROBIAL COLONIZATION ON U. AUSTRALIS 5551
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from
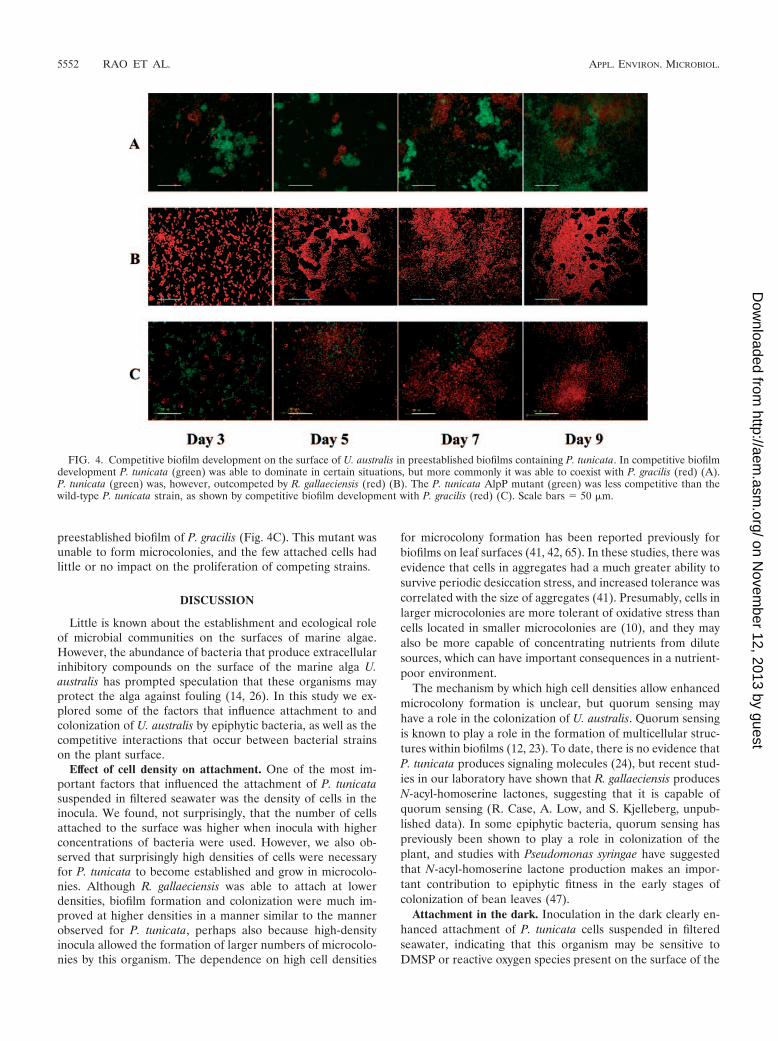
preestablished biofilm of P. gracilis (Fig. 4C). This mutant wasunable to form microcolonies, and the few attached cells hadlittle or no impact on the proliferation of competing strains.
DISCUSSION
Little is known about the establishment and ecological roleof microbial communities on the surfaces of marine algae.However, the abundance of bacteria that produce extracellularinhibitory compounds on the surface of the marine alga U.australis has prompted speculation that these organisms mayprotect the alga against fouling (14, 26). In this study we ex-plored some of the factors that influence attachment to andcolonization of U. australis by epiphytic bacteria, as well as thecompetitive interactions that occur between bacterial strainson the plant surface.
Effect of cell density on attachment. One of the most im-portant factors that influenced the attachment of P. tunicatasuspended in filtered seawater was the density of cells in theinocula. We found, not surprisingly, that the number of cellsattached to the surface was higher when inocula with higherconcentrations of bacteria were used. However, we also ob-served that surprisingly high densities of cells were necessaryfor P. tunicata to become established and grow in microcolo-nies. Although R. gallaeciensis was able to attach at lowerdensities, biofilm formation and colonization were much im-proved at higher densities in a manner similar to the mannerobserved for P. tunicata, perhaps also because high-densityinocula allowed the formation of larger numbers of microcolo-nies by this organism. The dependence on high cell densities
for microcolony formation has been reported previously forbiofilms on leaf surfaces (41, 42, 65). In these studies, there wasevidence that cells in aggregates had a much greater ability tosurvive periodic desiccation stress, and increased tolerance wascorrelated with the size of aggregates (41). Presumably, cells inlarger microcolonies are more tolerant of oxidative stress thancells located in smaller microcolonies are (10), and they mayalso be more capable of concentrating nutrients from dilutesources, which can have important consequences in a nutrient-poor environment.
The mechanism by which high cell densities allow enhancedmicrocolony formation is unclear, but quorum sensing mayhave a role in the colonization of U. australis. Quorum sensingis known to play a role in the formation of multicellular struc-tures within biofilms (12, 23). To date, there is no evidence thatP. tunicata produces signaling molecules (24), but recent stud-ies in our laboratory have shown that R. gallaeciensis producesN-acyl-homoserine lactones, suggesting that it is capable ofquorum sensing (R. Case, A. Low, and S. Kjelleberg, unpub-lished data). In some epiphytic bacteria, quorum sensing haspreviously been shown to play a role in colonization of theplant, and studies with Pseudomonas syringae have suggestedthat N-acyl-homoserine lactone production makes an impor-tant contribution to epiphytic fitness in the early stages ofcolonization of bean leaves (47).
Attachment in the dark. Inoculation in the dark clearly en-hanced attachment of P. tunicata cells suspended in filteredseawater, indicating that this organism may be sensitive toDMSP or reactive oxygen species present on the surface of the
FIG. 4. Competitive biofilm development on the surface of U. australis in preestablished biofilms containing P. tunicata. In competitive biofilmdevelopment P. tunicata (green) was able to dominate in certain situations, but more commonly it was able to coexist with P. gracilis (red) (A).P. tunicata (green) was, however, outcompeted by R. gallaeciensis (red) (B). The P. tunicata AlpP mutant (green) was less competitive than thewild-type P. tunicata strain, as shown by competitive biofilm development with P. gracilis (red) (C). Scale bars � 50 �m.
5552 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from

algae. DMSP occurs in many diverse species of algae, includingU. australis (14), and is released in senescent algae or underoxidative stress conditions, particularly under high light inten-sities (31, 59). There is evidence that DMSP provides protec-tion against photooxidation for algae (59), but its effects onepiphytic bacterial communities on Ulva spp. are not fullyunderstood (46). We observed that attachment of P. tunicata inthe light resulted in fewer but larger microcolonies, whereasmicrocolonies established from cells inoculated in the darkwere smaller and more numerous. One possibility is that cellsin larger more resistant microcolonies survived exposure tooxidative stress, which increased with higher light intensities.R. gallaeciensis was able to attach and colonize effectively re-gardless of the light conditions, suggesting that it may be ableto tolerate and/or metabolize DMSP (59). In studies of thediversity of dimethyl sulfide-producing bacteria from oceanicand estuarine waters it was found that all the isolates belongingto the Roseobacter group tested were dimethyl sulfide produc-ers and therefore could utilize DMSP (21). Another proposedecological role for DMSP is that it acts as a precursor of cuesfor chemosensory attraction between algae and certain specificbacteria (57). Thus, DMSP may be one of the chemical signalsreleased by U. australis which is recognized by R. gallaeciensis.Furthermore, it has been proposed that the substrate for thesulfur-based inhibitory compounds produced by R. gallaeciensis(thiotropocin and tropodithietic acid) is DMSP (8).
Effect of cellobiose on attachment. It is evident that P. tuni-cata attachment improved considerably when cells were grownin cellobiose and then suspended in filtered seawater, but thecells did not persist on the surface of U. australis. Both U.australis and Ciona intestinalis, from which P. tunicata is mostcommonly isolated, contain accessible cellulose polymers intheir cell walls (5, 13). A cellulose binding protein with a highbinding affinity for microcrystalline cellulose was discovered inP. tunicata (11). Clearly, cellobiose plays a role in attachment,but its effects may be restricted to the early stages of coloni-zation, in which it has been suggested to function as an an-chorage or cue for P. tunicata attachment (58). Recent studieshave found that cellulose stimulated the production of pigmentin this organism, which is known to be coregulated with ex-pression of AlpP (58), and also induced the expression ofmannose-sensitive hemagglutinin (type IV-like) pili in P. tuni-cata (11). Thus, cellulose appears to play an important role inthe colonization and responses of P. tunicata on the surface ofU. australis.
Synergistic biofilm formation. P. tunicata persisted on thesurface of the algae only if it was inoculated in seawater thatcontained a natural seawater community or if it was inoculatedin filtered seawater that contained a mixture of epiphyticstrains isolated from U. australis. This suggests that P. tunicatarequires the presence of diverse bacteria in the inoculum foreffective colonization, which may allow succession and/or co-operative colonization of the surface. Cocolonizing bacteriacould modify the habitat and create a “microenvironment” thatencourages the attachment and growth of other colonizingmicroorganisms. We tested whether a preexisting biofilm en-hances the attachment and persistence of P. tunicata by allow-ing it to colonize nonaxenic U. australis disks with the intactnatural community on the surface. However, P. tunicata wasunable to colonize under these conditions, indicating that an
established biofilm of other bacteria does not facilitate thecolonization. Another possibility is that cooperative interac-tions during the process of colonization (for example, come-tabolism or coaggregation) can occur between P. tunicata andother cocolonizing marine strains. The phenomenon of coop-erative biofilm formation in mixed consortia has recently beendescribed for other bacteria (16, 52). Of the 17 strains, it is notclear which strains or mixtures of strains are responsible forallowing P. tunicata to colonize. The underlying mechanisms ofcooperative biofilm formation are the basis of ongoing exper-iments in our laboratory.
Attachment and colonization of U. australis by P. tunicataappears to be influenced by many factors. Attachment andcolonization are particularly poor at low cell densities. The com-bination of environmental factors that appear to enhance col-onization at a concentration of 107 cells ml�1 does not seem tobe effective at lower cell densities (104 to 105 cells ml�1). Itappears that colonization is limited by nutrient availability, butit is also possible that unknown defensive chemicals producedby the plant play a role.
Competition in biofilms on U. australis. In coinoculationcompetition studies, we observed that P. tunicata suspended infiltered seawater outcompeted all of the other marine isolatestested except R. gallaeciensis. The results are similar to ourprevious findings of competition on glass surfaces in a labora-tory flow cell model. In that study, we demonstrated that com-petition was largely controlled by the production of inhibitorycompounds and the relative sensitivity of competing strains tothe inhibitors (48).
However, we did observe some important differences be-tween bacterial competition on the surface of the marine plantand the competition in laboratory systems in the preestab-lished biofilm studies. Unlike laboratory flow cells, which allowa continuous flow of nutrients, the surface of algae likely pre-sents nutrient-limited conditions. Our studies demonstratedthat exogenous application of nutrients resulted in a rapidincrease in the microcolony size of P. tunicata (data notshown), suggesting that microcolonies on the plant surface areindeed nutrient limited. Furthermore, our observation of thepatchy distribution of microcolonies might reflect the spatialheterogeneity of nutrients available on algal surfaces. Severalstudies conducted with higher terrestrial plants suggested thatmost areas of a leaf harbor only small amounts of nutrients (35,63), and on bean plants inoculated with P. syringae strainB728A aggregates of bacteria were distributed nonrandomly ina wide range of cluster sizes, which roughly corresponded tonutrient availability (36). Our observation that P. tunicata col-onized a preestablished biofilm poorly suggest that the initialcolonizers could deplete a large percentage of carbon sourcesand make it more difficult for the invading bacteria to seques-ter themselves in the existing biofilm community. This has beendescribed for epiphytic bacteria on bean leaves, on which apreestablished biofilm can deplete resources, leading to pre-emptive exclusion (64).
In competition studies in which P. tunicata was allowed toinvade a preestablished biofilm, it did not outcompete theother strains and coexisted with the competing strain for theduration of the experiment. When invading a preestablishedbiofilm, P. tunicata tends to colonize and establish microcolo-nies in areas which remain free from colonization by a prees-
VOL. 72, 2006 MICROBIAL COLONIZATION ON U. AUSTRALIS 5553
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from

tablished biofilm. This results in spatially segregated micro-colonies of competing strains with limited interactions andleads to coexistence of strains. Our observations of competitiveinteractions on the surface of U. australis are supported byrecent reports on spatial segregation of epiphytic bacteria onleaf surfaces (40, 43). The observations differ from the resultsof flow cell experiments in which P. tunicata was able to dom-inate and eventually remove certain competing strains (48).The evidence of a reduced ability to colonize a preestablishedbiofilm was even more pronounced for the AlpP mutant. Theability of the mutant to form microcolonies in a preestablishedbiofilm was impaired, and the mutant had minimal impact onthe competing strain, which remained dominant. Thus, ourresults indicate that AlpP enhances colonization of a preestab-lished biofilm by P. tunicata.
In contrast, R. gallaeciensis did not seem to be constrained bynutrient limitations and formed a biofilm which eventuallycovered most of the surface of the alga. An ability to utilize awide range of carbon sources (50) and an ability to metabolizeDMSP may enable this organism to colonize regions of theplant that are inhospitable to other bacteria. Even when colo-nizing a preestablished biofilm, R. gallaeciensis might be able toaccess nutrients that are inaccessible to other epiphytic bacte-ria, and this ability might contribute to its success and epiphyticfitness (6).
Conclusions. P. tunicata and R. gallaeciensis were found tohave different colonization and competition behaviors duringcolonization of U. australis. While R. gallaeciensis is capable ofcolonizing U. australis under a range of conditions, coloniza-tion by P. tunicata is enhanced by high cell densities, the pres-ence of cellobiose in the preculture, inoculation in the dark,and interactions with a natural seawater community to attachand persist on the surface of the algae. The epiphytic fitness ofR. gallaeciensis may be attributed to several factors, includingits versatility in utilizing a number of carbon sources (particu-larly those not available to competing strains) and the produc-tion of antibacterial compounds and signaling molecules.
Competition in which a preestablished biofilm is challengedwith P. tunicata results in the coexistence of competitors. Thismay be due in part to the protective nature of microcolonies,which may resist invasion. There is a diffusion gradient inmicrocolonies, so that metabolically active cells at the outeredge of the microcolony may perish while cells in the deeperregions are likely to be protected from the antibacterial pro-tein. Limited nutrients on the surface of U. australis may leadstrains to inhabit distinct niches on the plant, and this may alsoresult in the coexistence of competing strains. Microcolonies ofcompeting strains are spatially separated, which may limit mi-crobial interactions and provide refuges for competitors. R.gallaeciensis does not seem to be constrained by low-nutrientconditions and is able to invade and disperse competingstrains, suggesting that its antibacterial protein is able to dif-fuse through microcolonies.
The environmental factors that influence nutrient accumu-lation on the surface of the seaweed and in turn microbialcolonization and competitive interactions are complex. Wepropose that these factors may have a profound impact on thecomposition and activity of the epiphytic community.
ACKNOWLEDGMENTS
This work was supported by a USP-AusAID scholarship, by theAustralian Research Council, by the Leverhulme Trust, United King-dom, and by the Centre for Marine Biofouling and Bio-innovation,University of New South Wales, Sydney, Australia.
REFERENCES
1. Alavi, M., T. Miller, K. Erlandson, R. Schneider, and R. Belas. 2001. Bac-terial community associated with Pfiesteria-like dinoflagellate cultures. En-viron. Microbiol. 3:380–396.
2. Armstrong, E., K. G. Boyd, and J. G. Burgess. 2000. Prevention of marinebiofouling using natural compounds from marine organisms. Biotechnol.Annu. Rev. 6:221–241.
3. Armstrong, E., K. G. Boyd, A. Pisacane, C. J. Peppiatt, and J. G. Burgess.2000. Marine microbial natural products in antifouling coatings. Biofouling16:215–224.
4. Armstrong, E., L. Yan, K. G. Boyd, P. C. Wright, and J. G. Burgess. 2001.The symbiotic role of marine microbes on living surfaces. Hydrobiologia461:37–40.
5. Baldan, B., P. Andolfo, L. Navazio, C. Tolomio, and P. Mariani. 2001.Cellulose in algal cell wall: an “in situ” localisation. Eur. J. Histochem.45:51–56.
6. Beattie, G. A., and S. E. Lindow. 1999. Bacterial colonization of leaves: aspectrum of strategies. Phytopathology 89:353–359.
7. Boyd, K. G., D. R. Adams, and J. G. Burgess. 1999. Antibacterial andrepellent activities of marine bacteria associated with algal surfaces. Biofoul-ing 14:227–236.
8. Bruhn, J. B. B., K. F. Nielsen, M. Hjelm, M. Hansen, J. Bresciani, S. Schulz,and L. Gram. 2005. Ecology, inhibitory activity, and morphogenesis of amarine antagonistic bacterium belonging to the Roseobacter clade. Appl.Environ. Microbiol. 71:7263–7270.
9. Corre, S., and D. Prieur. 1990. Density and morphology of epiphytic bacteriaon the kelp Laminaria digitata. Bot. Mar. 33:515–523.
10. Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms:a common cause of persistent infections. Science 284:1318–1322.
11. Dalisay-Saludes, D. 2004. Antibacterial activity and mechanisms of coloni-sation of Pseudoalteromonas tunicata. Ph.D. thesis. University of New SouthWales, Sydney, Australia.
12. Davies, D. G., M. R. Parsek, J. P. Pearson, B. H. Iglewski, J. W. Costerton,and E. P. Greenberg. 1998. The involvement of cell-to-cell signals in thedevelopment of a bacterial biofilm. Science 200:295–298.
13. de Leo, G., E. Patricolo, and G. D’Ancona Lunetta. 1977. Studies on thefibrous component of the test of Ciona intestinalis Linnaeus. I. Cellulose-likepolysaccharide. Acta Zool. 58:135–141.
14. Dickson, D. M. J., and G. O. Kirst. 1986. The role of -dimethylsulfonio-propionate, glycine betaine and homarine in the osmoacclimation of Platy-monas subcordiformes. Planta 167:536–543.
15. Dobretsov, S. V., and P. Y. Qian. 2002. Effect of bacteria associated with thegreen alga Ulva reticulata on marine micro- and macrofouling. Biofouling18:217–228.
16. Filoche, S. K., M. Zhu, and C. D. Wu. 2004. In situ Biofilm formation bymulti-species oral bacteria under flowing and anaerobic conditions. J. Dent.Res. 83:802–806.
17. Fisher, M. M., L. W. Wilcox, and L. E. Graham. 1998. Molecular character-ization of epiphytic bacterial communities on charophycean green algae.Appl. Environ. Microbiol. 64:4384–4389.
18. Fuhrman, J. A., K. Mccallum, and A. A. Davis. 1993. Phylogenetic diversityof subsurface marine microbial communities from the Atlantic and Pacificoceans. Appl. Environ. Microbiol. 59:1294–1302.
19. Gil-Turness, M. S., and W. Fenical. 1992. Embryos of Homarus americanusare protected by epibiotic bacteria. Bio. Bull. 182:105–108.
20. Gonzalez, J., and M. Moran. 1997. Numerical dominance of a group ofmarine bacteria in the alpha-subclass of the class Proteobacteria in coastalseawater. Appl. Environ. Microbiol. 63:4237–4242.
21. Gonzalez, J. M., R. P. Kiene, and M. A. Moran. 1999. Transformation ofsulfur compounds by an abundant lineage of marine bacteria in the alpha-subclass of the class Proteobacteria. Appl. Environ. Microbiol. 65:3810–3819.
22. Gram, L., H. P. Grossart, A. Schlingloff, and T. Kiorboe. 2002. Possiblequorum sensing in marine snow bacteria: production of acylated homoserinelactones by Roseobacter strains isolated from marine snow. Appl. Environ.Microbiol. 68:4111–4116.
23. Heydorn, A., B. Ersboll, J. Kato, M. Hentzer, M. R. Parsek, T. Tolker-Nielsen, M. Givskov, and S. Molin. 2002. Statistical analysis of Pseudomonasaeruginosa biofilm development: impact of mutations in genes involved intwitching motility, cell-to-cell signaling, and stationary-phase sigma factorexpression. Appl. Environ. Microbiol. 68:2008–2017.
24. Holmstrom, C., S. Egan, A. Franks, S. McCloy, and S. Kjelleberg. 2002.Antifouling activities expressed by marine surface associated Pseudoaltero-monas species. FEMS Microbiol. Ecol. 41:47–58.
25. Holmstrom, C., S. James, B. A. Neilan, D. C. White, and S. Kjelleberg. 1998.
5554 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from

Pseudoalteromonas tunicata sp. nov., a bacterium that produces antifoulingagents. Int. J. Syst. Bacteriol. 48:1205–1212.
26. Holmstrom, C., and S. Kjelleberg. 1994. The effect of external biologicalfactors on settlement of marine invertebrate and new antifouling technology.Biofouling 8:147–160.
27. Holmstrom, C., D. Rittschof, and S. Kjelleberg. 1992. Inhibition of settle-ment by larvae of Balanus amphitrite and Ciona intestinalis by a surface-colonizing marine bacterium. Appl. Environ. Microbiol. 58:2111–2115.
28. James, S., C. Holmstrom, and S. Kjelleberg. 1996. Purification and charac-terisation of a novel antibacterial protein from the marine bacterium D2.Appl. Environ. Microbiol. 62:2783–2788.
29. Jensen, P. R., C. A. Kauffman, and W. Fenical. 1996. High recovery ofculturable bacteria from the surfaces of marine algae. Mar. Biol. 126:1–7.
30. Jones, A. K., and R. C. Cannon. 1986. The release of micro-algal photosyn-thate and associated bacterial uptake and heterotrophic growth. Br. Phycol.J. 21:341–358.
31. Karsten, U., C. Wiencke, and G. O. Kirst. 1990. The effect of light intensityand daylength on the -dimethylsulfoniopropionate (DMSP) content of ma-rine green macroalgae from Antarctica. Plant Cell Environ. 13:989–993.
32. Kjelleberg, S., P. Steinberg, M. Givskov, L. Gram, M. Manefield, and R.deNys. 1997. Do marine natural products interfere with prokaryotic AHLregulatory systems? Aquat. Microb. Ecol. 13:85–93.
33. Lafay, B., R. Ruimy, C. de Traubenberg, V. Breittmayer, M. Gauthier, andR. Christen. 1995. Roseobacter algicola sp. nov., a new marine bacteriumisolated from the phycosphere of the toxin-producing dinoflagellate Proro-centrum lima. Int. J. Syst. Bacteriol. 45:290–296.
34. Lange, W. 1967. Effect of carbohydrates on the symbiotic growth of plank-tonic blue-green algae with bacteria. Nature 215:1277–1278.
35. Leben, C. 1988. Relative humidity and the survival of epiphytic bacteria withbuds and leaves of cucumber plants. Phytopathology 78:179–185.
36. Leveau, J. H. J., and S. E. Lindow. 2001. Inaugural article. Appetite of anepiphyte: quantitative monitoring of bacterial sugar consumption in thephyllosphere. Proc. Natl. Acad. Sci. USA 98:3446–3453.
37. Mai-Prochnow, A., F. Evans, D. Dalisay-Saludes, S. Stelzer, S. Egan, S.James, J. S. Webb, and S. Kjelleberg. 2004. Biofilm development and celldeath in the marine bacterium Pseudoalteromonas tunicata. Appl. Environ.Microbiol. 70:3232–3238.
38. Marden, P., A. Tunlid, K. Malmcrona-Friberg, G. Odham, and S. Kjelle-berg. 1985. Physiological and morphological changes during short term star-vation of marine bacteria isolates. Arch. Microbiol. 142:326–332.
39. Miller, T. R., and R. Belas. 2004. Dimethylsulfoniopropionate metabolism byPfiesteria-associated Roesobacter spp. Appl. Environ. Microbiol. 70:3383–3391.
40. Monier, J.-M., and S. E. Lindow. 2005. Aggregates of resident bacteriafacilitate survival of immigrant bacteria on leaf surfaces. Microb. Ecol. 49:343–352.
41. Monier, J.-M., and S. E. Lindow. 2003. Differential survival of solitary andaggregated bacterial cells promotes aggregate formation on leaf surfaces.Proc. Natl. Acad. Sci. USA 100:15977–15982.
42. Monier, J.-M., and S. E. Lindow. 2004. Frequency, size, and localization ofbacterial aggregates on bean leaf surfaces. Appl. Environ. Microbiol. 70:346–355.
43. Monier, J.-M., and S. E. Lindow. 2005. Spatial organization of dual-speciesbacterial aggregates on leaf surfaces. Appl. Environ. Microbiol. 71:5484–5493.
44. Nakasono, S., J. G. Burgess, K. Takashi, M. Koike, C. Murayama, S.Nakamura, and T. Matsunaga. 1993. Electrochemical prevention of marinebiofouling with a carbon-chloroprene sheet. Appl. Environ. Microbiol. 59:3757–3762.
45. Neidhart, F. C., P. L. Bloch, and D. F. Smith. 1974. Culture medium forenterobacteria. J. Bacteriol. 119:736–747.
46. Nishiguchi, M. K. 1994. The role of dimethylsulfoniopropionate (DMSP) inmarine macroalgae of the Monterey Bay. Ph.D. thesis. University of Cali-fornia at Santa Cruz, Santa Cruz.
47. Quinones, B., C. J. Pujol, and S. E. Lindow. 2004. Regulation of AHLproduction and its contribution to epiphytic fitness in Pseudomonas syringae.Mol. Plant-Microbe Interact. 17:521–531.
48. Rao, D., J. S. Webb, and S. Kjelleberg. 2005. Competitive interactions inmixed-species biofilms containing the marine bacterium Pseudoalteromonastunicata. Appl. Environ. Microbiol. 71:1729–1736.
49. Rier, S. T., and R. J. Stevenson. 2002. Effects of light, dissolved organiccarbon, and inorganic nutrients on the relationship between algae and het-erotrophic bacteria in stream periphyton. Hydrobiologia 489:179–184.
50. Ruiz-Ponte, C., V. Cilia, C. Lambert, and J. L. Nicolas. 1998. Roseobactergallaeciensis sp. nov., a new marine bacterium isolated from rearings andcollectors of the scallop Pecten maximus. Int. J. Syst. Bacteriol. 48:537–542.
51. Scheffel, A. 2003. Protocol for generating axenic Ulva lactuca and studies oftype IV pili mediated attachment of Pseudoalteromonas tunicata to the algalsurface. Diploma thesis. University of New South Wales, Sydney, Australia.
52. Sharma, A., S. Inagaki, W. Sigurdson, and H. K. Kuramitsu. 2005. Synergybetween Tannerella forsythia and Fusobacterium nucleatum in biofilm forma-tion. Oral Microbiol. Immunol. 20:39–42.
53. Shiba, T. 1992. The genus Roseobacter, 2nd ed. Springer-Verlag, Berlin,Germany.
54. Sieburth, J. M. 1975. Microbial seascapes: a pictoral essay on marine mi-croorganisms and their environments. University Park Press, Baltimore, Md.
55. Sieburth, J. M., and J. T. Conover. 1965. Sargassum tannin, an antibioticwhich retards fouling. Nature 208:52–53.
56. Sieburth, J. M., and J. L. Tootle. 1981. Seasonality of microbial fouling onAscophyllum nodosum (L.) Lejol, Fucus vesiculosus L., Polysiphonia lanosa(L.) Tandy and Chondrus crispus Stackh. J. Phycol. 17:57–64.
57. Simo, R. 2001. Production of atmospheric sulfur by oceanic plankton: bio-geochemical, ecological and evolutionary links. Trends Ecol. Evol. 16:287–294.
58. Skovhus, T. L. 2005. Occurrence and function of marine antifoulingPseudoalteromonas species. Ph.D. thesis. University of Aarhus, Aarhus, Den-mark.
59. Sunda, W., D. J. Kieber, R. P. Kiene, and S. Huntsman. 2002. An antioxidantfunction for DMSP and DMS in marine algae. Nature 418:317–320.
60. Swain, D., and D. N. De. 1994. Vital staining, a technique for rapid screeningof plant protoplast viability. Ind. J. Exp. Biol. 32:501–506.
61. Taylor, M. W., P. J. Schupp, I. Dahllof, S. Kjelleberg, and P. Steinberg. 2004.Host specificity in marine sponge-associated bacteria, and potential implica-tions for marine microbial diversity. Environ. Microbiol. 6:121–130.
62. Wahl, M. 1989. Marine epibosis. 1 Fouling and antifouling: some basicaspects. Mar. Ecol. Progr. Ser. 58:175–189.
63. Weller, D. M., and A. W. Saettler. 1980. Colonisation and distribution ofXanthomonas phaseoli and Xanthomonas phaseoli var. fuscans in field-grownnavy beans. Phytopathology 70:500–506.
64. Wilson, M., and S. Lindow. 1994. Ecological similarity and coexistence ofepiphytic ice-nucleating (ice�) Pseudomonas syringae strains and a non-ice-nucleating (ice�) biological control agent. Appl. Environ. Microbiol. 60:3128–3137.
65. Wilson, M., and S. E. Lindow. 1994. Inoculum density-dependent mortalityand colonization of the phyllosphere by Pseudomonas syringae. Appl. En-vrion. Microbiol. 60:2232–2237.
VOL. 72, 2006 MICROBIAL COLONIZATION ON U. AUSTRALIS 5555
on Novem
ber 12, 2013 by guesthttp://aem
.asm.org/
Dow
nloaded from




















