
A new specimen of Neuquensaurus australis , a Late Cretaceous saltasaurine titanosaur from north...
12
A NEW SPECIMEN OF NEUQUENSAURUS AUSTRALIS, A LATE CRETACEOUS SALTASAURINE TITANOSAUR FROM NORTH PATAGONIA LEONARDO SALGADO 1 , SEBASTIÁN APESTEGUÍA 2 , and SUSANA E. HEREDIA 3 1 CONICET-Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400, 8300 Neuquén, Argentina, [email protected]; 2 Museo Argentino de Ciencias Naturales “Bernadino Rivadavia”, Av. Ángel Gallardo 470, 1405 Capital Federal, Argentina, [email protected]; 3 CONICET-Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400, 8300 Neuquén, Argentina, [email protected] ABSTRACT—A new specimen of the sauropod titanosaur Neuquensaurus australis, collected in the locality of Cinco Saltos (Patagonia, Argentina), provides an opportunity to improve our knowledge of the anatomy of this dinosaur. The elements represented in this specimen include a complete cervical vertebra, most of the dorsal vertebrae, fifteen caudal vertebrae, the complete sacrum articulated to both ilia, one ischium, two femora, one tibia articulated to the fibula and astragalus, and two osteoderms. Surprisingly, the sacrum is composed of seven vertebrae, the last of which, unfused to the other six, is apparently biconvex. A third femur and one tibia were found associated with this specimen; these are morphologically similar but stouter than those belonging to the former specimen. INTRODUCTION Lydekker (1893:4) erected ‘Titanosaurus’ australis, one of the first known Patagonian dinosaurs, on the basis of an associated series of vertebrae, and a number of limb and girdle bones from Neuquén (North Patagonia, Argentina). Our current knowledge of the anatomy of this titanosaur sau- ropod is based mainly on Huene’s (1929) monograph on Creta- ceous dinosaurs of Argentina, wherein several of the hundreds of bones, brought from several Late Cretaceous localities of North Patagonia to the Museo de La Plata by Roth and Schiller by the beginning of the twentieth century, are described. The type specimen of ‘Titanosaurus’ australis was thus artificially con- structed by Huene on the base of mostly isolated materials. In his unpublished revision of the South American titanosaurs, Powell (1986) did not include new elements to those described by Huene (1929) and Bonaparte and Gasparini (1980). Thus, our understanding of the anatomy of this species has remained un- altered for decades. Powell considered it to be part of the Saltasaurinae, different enough from the Indian titanosaurs to support erection of the genus Neuquensaurus Powell, 1992, to contain the species, as Neuquensaurus australis (Lydekker, 1893). A 1998 field trip carried out by the museum of the Universidad del Comahue (Neuquén Province) and the Museo de Cinco Sal- tos (Río Negro Province), yielded a partially articulated skeleton of Neuquensaurus australis. In this paper, we provide a detailed description of this species, taking into account the new specimen as well as the many individuals (both published and unpublished) housed in the museums of La Plata (Buenos Aires Province) and Cipolletti (Río Negro Province). The new material improves substantially our understanding of the anatomy of Neuquensaurus australis in particular, and of titanosaurs in general. The new skeleton is very well preserved and includes parts until now poorly known, including the pelvis and the dorsal vertebrae. In addition, the present find allows a detailed analysis of the pattern of variation along the vertebral column of titanosaurs, the vertebrae of which are frequently found as isolated pieces. We adopted the nomenclature for the osseous laminae pro- posed by Wilson (1999). Institutional Abbreviations—FMNH, Field Museum of Natu- ral History, Chicago; MAL, Collection in the Shuler Museum of Paleontology (Dallas, Texas), but belonging to the Malawi De- partment of Antiquities; MCS, Museo de Cinco Saltos, Río Ne- gro Province; MCT, Collection of the Earth Science Museum of the National Department of Mineral Production, Río de Janeiro; MLP-Cs, Museo de La Plata, Cinco Saltos collection; MLP-Ly, Museo de La Plata, Lydekker collection; MPCA-Pv, Collection of vertebrate paleontology of the Museum of Cipolletti “Carlos Ameghino”, Cipolletti, Río Negro; PVL, Collection of verte- brate paleontology of the Instituto Miguel Lillo, Tucumán. GEOLOGICAL FRAMEWORK The Neuquén Basin, developed in Western Argentina (Fig. 1) has deposits ranging in age from Upper Triassic to Tertiary. The sediments filling this basin have been subdivided into three su- percycles (Groeber, 1946): “Jurassic” (Late Triassic to Lower Jurassic), “Ándico” (Lower Jurassic to Early Cretaceous) and “Riográndico” (Late Cretaceous to Paleocene). The latest su- percycle is also divided into the “Neuqueniano” Cycle or Neu- quén Group (Late Albian?/Lower Cenomanian to Lower Cam- panian) and the “Malalhueyano” Cycle or Malargüe Group (Up- per Campanian to Paleocene). The first one is composed of sediments deposited in a continental fluvial environment (Uliana and Dellapé, 1981). The second constitutes the first incursion of the Atlantic Ocean into the basin (Uliana and Dellapé, 1981). As stated by Andreis et al. (1974) and by Digregorio and Uliana (1979), a stratigraphic discordance occurs between the Malargüe and Neuquén Groups. During the Late Cretaceous, the eastern section of the basin became a foreland province that was dominated by molasse sedi- mentation related to the uplifting cordillera to the West (Ramos, 1981, 1985). Cinco Saltos Area The study area belongs to the eastern border of Neuquén Basin and conforms with Upper Cretaceous sedimentary out- crops exposed in the northwestern scarp near Cinco Saltos (Río Negro Province). Upper Cretaceous to Tertiary continental de- Journal of Vertebrate Paleontology 25(3):623–634, September 2005 © 2005 by the Society of Vertebrate Paleontology 623
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A new specimen of Neuquensaurus australis , a Late Cretaceous saltasaurine titanosaur from north...

A NEW SPECIMEN OF NEUQUENSAURUS AUSTRALIS, A LATE CRETACEOUSSALTASAURINE TITANOSAUR FROM NORTH PATAGONIA
LEONARDO SALGADO1, SEBASTIÁN APESTEGUÍA2, and SUSANA E. HEREDIA3
1CONICET-Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400,8300 Neuquén, Argentina, [email protected];
2Museo Argentino de Ciencias Naturales “Bernadino Rivadavia”, Av. Ángel Gallardo 470, 1405 Capital Federal, Argentina,[email protected];
3CONICET-Museo de Geología y Paleontología, Universidad Nacional del Comahue, Buenos Aires 1400,8300 Neuquén, Argentina, [email protected]
ABSTRACT—A new specimen of the sauropod titanosaur Neuquensaurus australis, collected in the locality of CincoSaltos (Patagonia, Argentina), provides an opportunity to improve our knowledge of the anatomy of this dinosaur. Theelements represented in this specimen include a complete cervical vertebra, most of the dorsal vertebrae, fifteen caudalvertebrae, the complete sacrum articulated to both ilia, one ischium, two femora, one tibia articulated to the fibula andastragalus, and two osteoderms. Surprisingly, the sacrum is composed of seven vertebrae, the last of which, unfused to theother six, is apparently biconvex. A third femur and one tibia were found associated with this specimen; these aremorphologically similar but stouter than those belonging to the former specimen.
INTRODUCTION
Lydekker (1893:4) erected ‘Titanosaurus’ australis, one of thefirst known Patagonian dinosaurs, on the basis of an associatedseries of vertebrae, and a number of limb and girdle bones fromNeuquén (North Patagonia, Argentina).
Our current knowledge of the anatomy of this titanosaur sau-ropod is based mainly on Huene’s (1929) monograph on Creta-ceous dinosaurs of Argentina, wherein several of the hundreds ofbones, brought from several Late Cretaceous localities of NorthPatagonia to the Museo de La Plata by Roth and Schiller by thebeginning of the twentieth century, are described. The typespecimen of ‘Titanosaurus’ australis was thus artificially con-structed by Huene on the base of mostly isolated materials.
In his unpublished revision of the South American titanosaurs,Powell (1986) did not include new elements to those describedby Huene (1929) and Bonaparte and Gasparini (1980). Thus, ourunderstanding of the anatomy of this species has remained un-altered for decades. Powell considered it to be part of theSaltasaurinae, different enough from the Indian titanosaurs tosupport erection of the genus Neuquensaurus Powell, 1992, tocontain the species, as Neuquensaurus australis (Lydekker,1893).
A 1998 field trip carried out by the museum of the Universidaddel Comahue (Neuquén Province) and the Museo de Cinco Sal-tos (Río Negro Province), yielded a partially articulated skeletonof Neuquensaurus australis. In this paper, we provide a detaileddescription of this species, taking into account the new specimenas well as the many individuals (both published and unpublished)housed in the museums of La Plata (Buenos Aires Province) andCipolletti (Río Negro Province).
The new material improves substantially our understanding ofthe anatomy of Neuquensaurus australis in particular, and oftitanosaurs in general. The new skeleton is very well preservedand includes parts until now poorly known, including the pelvisand the dorsal vertebrae. In addition, the present find allows adetailed analysis of the pattern of variation along the vertebralcolumn of titanosaurs, the vertebrae of which are frequentlyfound as isolated pieces.
We adopted the nomenclature for the osseous laminae pro-posed by Wilson (1999).
Institutional Abbreviations—FMNH, Field Museum of Natu-ral History, Chicago; MAL, Collection in the Shuler Museum ofPaleontology (Dallas, Texas), but belonging to the Malawi De-partment of Antiquities; MCS, Museo de Cinco Saltos, Río Ne-gro Province; MCT, Collection of the Earth Science Museum ofthe National Department of Mineral Production, Río de Janeiro;MLP-Cs, Museo de La Plata, Cinco Saltos collection; MLP-Ly,Museo de La Plata, Lydekker collection; MPCA-Pv, Collectionof vertebrate paleontology of the Museum of Cipolletti “CarlosAmeghino”, Cipolletti, Río Negro; PVL, Collection of verte-brate paleontology of the Instituto Miguel Lillo, Tucumán.
GEOLOGICAL FRAMEWORK
The Neuquén Basin, developed in Western Argentina (Fig. 1)has deposits ranging in age from Upper Triassic to Tertiary. Thesediments filling this basin have been subdivided into three su-percycles (Groeber, 1946): “Jurassic” (Late Triassic to LowerJurassic), “Ándico” (Lower Jurassic to Early Cretaceous) and“Riográndico” (Late Cretaceous to Paleocene). The latest su-percycle is also divided into the “Neuqueniano” Cycle or Neu-quén Group (Late Albian?/Lower Cenomanian to Lower Cam-panian) and the “Malalhueyano” Cycle or Malargüe Group (Up-per Campanian to Paleocene). The first one is composed ofsediments deposited in a continental fluvial environment (Ulianaand Dellapé, 1981). The second constitutes the first incursion ofthe Atlantic Ocean into the basin (Uliana and Dellapé, 1981). Asstated by Andreis et al. (1974) and by Digregorio and Uliana(1979), a stratigraphic discordance occurs between the Malargüeand Neuquén Groups.
During the Late Cretaceous, the eastern section of the basinbecame a foreland province that was dominated by molasse sedi-mentation related to the uplifting cordillera to the West (Ramos,1981, 1985).
Cinco Saltos Area
The study area belongs to the eastern border of NeuquénBasin and conforms with Upper Cretaceous sedimentary out-crops exposed in the northwestern scarp near Cinco Saltos (RíoNegro Province). Upper Cretaceous to Tertiary continental de-
Journal of Vertebrate Paleontology 25(3):623–634, September 2005© 2005 by the Society of Vertebrate Paleontology
623

posits constitute a continuous belt with horizontal disposition. Arevised formal stratigraphic terminology is used for this UpperCretaceous sedimentary succession in the Neuquén Basin(Leanza and Hugo, 2001) comprised, in ascending order, of the
Río Limay Subgroup, the Río Neuquén Subgroup and the RíoColorado Subgroup.
The Río Colorado Subgroup includes the Bajo de la Carpa andAnacleto Formations, both exposed on the central and eastern
FIGURE 1. Geological sketch of Pellegrini Lake–Cinco Saltos area (modified from Andreis et al, 1974) and stratigraphical section with Neuquen-saurus occurrence.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005624

regions of the basin. They show rather horizontal deposits ex-posed by erosion and known as ‘bardas.’ The age of the AnacletoFormation was recently confirmed as early Campanian by mag-netostratigraphy (Dingus et al., 2000).
In this area, the Río Colorado Subgroup is composed ofcoarse-grained, well-sorted, white to gray conglomerate sand-stones with intercalations of fine-grained sandstones and redlaminated siltstones. The light gray sandstone typically displaysmedium-scale tabular and trough cross stratification. Remains ofdinosaurs (including birds), crocodiles, snakes, lizards, turtles,and fishes appear in these rocks. The fine deposits are composedmainly of thinly bedded, red and green siltstones and mudstones,with abundant carbonate nodules. Well-preserved dinosaur re-mains come from these strata (Coria and Salgado, 1996; Salgadoet al., 1997b). The Río Colorado Subgroup outcrops have beeninterpreted as braidplain and floodplain deposits with develop-ment of soils (Salgado et al., 1997b; Leanza, 1999; Hugo andLeanza, 2001; Leanza and Hugo, 2001).
Neuquensaurus Section
This section is a heterolithic succession composed of mud-stones and siltstones alternating with sandstone beds (Fig. 1). Itis a 12 m-thick channel–floodplain deposit of red siltstones andred and green mudstones alternating with light yellow sand-stones displayed in thin beds. At the top, a thick conglomerate isdeveloped, and clay intraclasts and trough cross bedding areobserved. The thickening-upward and coarsening-upward bedsvary from 0.25 to 3 m in thickness. They appear massive, withcalcrete concretions, fine sediments, and trough cross-beddedsandstones. This evidence provides additional support for thepreviously proposed paleoenvironment in which Neuquensaurusremains were found, associated with floodplain deposits.
SYSTEMATIC PALEONTOLOGY
SAURISCHIA Seeley, 1883SAUROPODOMORPHA Huene, 1932
TITANOSAURIA Bonaparte and Coria, 1993SALTASAURINAE Powell, 1992
Genus NEUQUENSAURUS Powell, 1992
Type Species—Titanosaurus australis Lydekker, 1893
NEUQUENSAURUS AUSTRALIS (Lydekker, 1893)
Remarks—Powell (1986:161–162) diagnosed Neuquensaurusaustralis as follows: (1) small to medium-sized titanosaur, similarto Saltasaurus loricatus Bonaparte and Powell, 1980; (2) cervicalvertebrae relatively short and wide; (3) dorsal vertebrae withshort centra and reduced pleurocoels; (4) sacrum with six well-co-ossified vertebrae, sacral centra 2 to 5 narrowed; (5) biconvexfirst caudal; (6) caudal vertebrae relatively low, with wide ventralface of centrum deeply excavated, and limited by lateral roundedridges culminating in articular facets of hemapophyses; (7) lat-eral walls of vertebral centra minimally exposed in ventral view;(8) scapula with prominent area for muscular attachment oninner face, near antero-superior border, as in Saltasaurus; (9)sternal plates with very prominent ventral crests; (10) limb bonesrelatively short, more slender than in Saltasaurus; (11) radii sig-moid in outline; (12) short and robust metacarpals; (13) ilia withanterior projections laterally expanded, but somewhat shorterthan in Saltasaurus; (14) fibulae with simple but well-developedlateral tuberosities; (15) transversely short astragali.
Powell’s diagnosis of Neuquensaurus australis is a mixture of
characters present in other saltasaurines (characters 1, 2), or inother non-saltasaurine titanosaurs (characters 3, 4, 5, 8, 9, 10, 12,13, 15). We propose instead a list of six potential autapomorphiesof Neuquensaurus australis. These are included in the followingdiagnosis:
Revised Diagnosis—(1) Fibula with strong lateral tuberosityand bent shaft (Powell, 1986); (2) anteriormost dorsal vertebraelacking centroprezygapophyseal (cprl) and centropostzyg-apophyseal (cpol) laminae (Wilson, 2002:275); (3) seven sacralvertebrae; (4) sacral centra 3 to 5 narrowed (modified from Pow-ell, 1986); (5) mid and posterior caudal vertebrae bearing trans-versely wide, non-keeled ventral depression, limited by lateralrounded ridges culminating in articular facets of hemapophyses(Powell, 1986); and (6) lateral walls of caudal vertebral centralittle exposed in ventral view (Powell, 1986).
Characters 5 and 6 are observed in the holotype of Neuquen-saurus australis (MLP-Ly 1/2/3/4/5/6).
Material—MCS-5: One posterior cervical vertebra (MCS-5/17); six mid and posterior dorsal vertebrae (MCS-5/18-23); onesacrum articulated to both ilia (MCS-5/16); 15 caudal vertebrae(MCS-5/1-15), 14 of which (MCS-5/2-MCS-5/15) probably rep-resent a continuous series; one left ischium (MCS-5/24); bothfemora (the left one, MCS-5/27 and the right one, MCS-5/28),right tibia (MCS-5/25), right fibula (MCS-5/26), right astragalus(MCS-5/29), 3 haemal arches (MCS-5/30-32), 2 osteoderms(MCS-5/33 and 34) and many fragments of ribs.
Additionally, other elements were found associated withMCS-5: a right femur (MCS-9), a right tibia (MCS-6), a lefthumerus (MCS-8) and a metatarsal (MCS-10), all of which areprovisionally interpreted as belonging to the same genus. MCS-9and MCS-6 are thought to be part of a single specimen. A leftscapula (MCS-7) was associated with specimen MCS-5, but itdiffers from the scapulae assigned to Neuquensaurus at the Mu-seo de La Plata. However, because the bone was associated withthis skeleton, it will be described briefly here.
Occurrence—Cinco Saltos, Río Negro Province, Argentina.Río Colorado Subgroup (Cazau and Uliana, 1973), top of theAnacleto Formation (Heredia and Salgado, 1999; Leanza, 1999;Leanza and Hugo, 2001), early Campanian (Leanza, 1999;Leanza and Hugo, 2001).
DESCRIPTION
Cervical Vertebrae—A nearly complete posterior cervicalvertebra (possibly the 12th or 13th) is well preserved (MCS-5/17)(Fig. 2, Table 1). The centrum is elongate, dorsoventrally com-pressed, and strongly opisthocoelous. The cervical pleurocoel isoval and it is located behind the parapophysis. The neural spineculminates in a hemispherical protuberance, as in the ‘Peirópolistitanosaur’ (Powell, 1987a:“Serie A”).
There is a delicate intraprezygapophyseal lamina (tprl) (alsopresent in MLP-Cs-1374 and MLP-Cs-1375, which are referredto Neuquensaurus australis), and two spinoprezygapophyseallaminae (sprl), that delimit a shallow triangular concavity, whichis somewhat deeper in MLP-Cs-1374 and MLP-Cs-1375. Be-tween both sprl, there is a rugosity that may indicate a trace of aprespinal lamina. In the cervical vertebra MLP-Cs-1378, assignedquestionably to Neuquensaurus robustus by Huene (1929), thereare four diverging prespinal laminae.
The prezygapophyses are widely separated from the midline ofthe vertebra (Fig. 2B). These are robust and short, with subtri-angular articular surfaces that are slightly convex anteroposteri-orly. The postzygapophyses project outwards and backwards,and extend beyond the posterior margin of the vertebral cen-trum. This last character is thought to be a synapomorphy of theSaltasaurinae (Salgado et al., 1997a).
The stout transverse processes are deep, project outwards, andbend downward slightly. Conversely, in Isisaurus colberti, the
SALGADO ET AL.—NEW SPECIMEN OF NEUQUENSAURUS 625

processes seem to be shorter and thinner (Jain and Bandyo-padhyay, 1997:fig. 5). The transverse process is strengthened bythree laminae that converge towards its distal end: the first runsfrom the prezygapophysis (the prezygodiapophyseal lamina,prdl), the second one, from the postzygapophysis (the postzygo-diapophyseal lamina, podl), and the last one from the vertebralcentrum (the posterior centrodiaophyseal lamina, pcdl). The an-terior centrodiapophyseal lamina (acdl) is virtually nonexistent.
Unlike other titanosaur specimens, the parapophyses are lat-erally (rather than lateroventrally) projected, which is thought tobe due to post-mortem dorsoventral compression.
Dorsal Vertebrae—Two well-preserved mid-dorsal vertebrae(MCS-5/18-19) (Fig. 3) and four posterior dorsal vertebrae(MCS-5/20-23) (Figs. 4, 5; Table 1) were recovered. The wholesequence is possibly incomplete, but both series seem to be in-ternally continuous. Assuming a total of 10 dorsal vertebrae,which is the minimum number of dorsals reported in titanosaurs(Powell, 1987a), and taking into account the general morphologyof the centra, the relative pleurocoel size, the inclination of theneural spine and the relative placement of the parapophyses anddiapophyses, we interpret that the first two are the 4th and 5thdorsal vertebrae, and the others are the 7th, 8th, 9th, and 10thdorsal vertebrae.
All of the dorsal centra are strongly opisthocoelous. The pleu-rocoels are elliptical, slightly eye-shaped, with a well-developedsystem of internal divisions, as is usual in titanosaurs (Salgado etal., 1997a). The pleurocoels are proportionally approximately 15percent taller than in the ‘Peirópolis titanosaur’ ‘Serie B’ (Pow-ell, 1987a:lam. 2) and in Lirainosaurus astibiae (Sanz et al.,1999:pl. 1a).
The ventral face of the centrum has a subtle longitudinal keeldividing two shallow concavities. This smooth ridge can be seenin the 4th, 5th, and 7th dorsal vertebrae. It is still present in the8th, but only on the anterior half of the centrum. In the 9th and10th dorsal vertebrae, the keel is not well developed.
The neural arch is somewhat inclined posteriorly in the firstvertebrae of the series (Fig. 3), becoming progressively uprighttowards the 10th (Fig. 4). The transverse processes are invariablydirected up and backwards. All dorsal vertebrae lack the hypo-sphene-hypantrum accessory articulations, as in other titano-saurs (Wilson, 2002).
A system of osseous laminae unites different points of theneural arch and the centrum. In anterior view, the complex sprl+ prsl (see Wilson, 1999) is well developed throughout the series,and invariably reaches the base of the neural arch, between the
prezygapophyses (Figs. 3, 5). This condition is also observed inall the examined dorsal vertebrae of Neuquensaurus australishoused in the Museum of La Plata (MLP-Cs-1381, MLP-Cs-1373). In vertebra MLP-Cs-1385 the complex forks ventrally, asis thought to occur in the cervico-dorsal transition (Powell,1987a).
The neural spine is supported laterally by the spinodiapophy-seal laminae (spdl). These laminae reach the tip of the spine,below the lateral expansions that, in anterior view, give the spinea rhomboid configuration (Figs. 3, 5). An additional lamina thatruns from the diapophyses to the base of the sprl + prsl complexcan be seen in anterior view in the seventh (where it is poorlydeveloped), ninth, and tenth dorsal vertebrae (Fig. 5). This un-named diapophyseal lamina (which is provisionally designated asudl) has not been reported previously for sauropods. It is presentneither in the related species Rocasaurus muniozi (Salgado andAzpilicueta, 2000:fig. 3a, 4a) nor in Saltasaurus loricatus (Powell,1986).
The posterior centrodiapophyseal lamina (pcdl) is well devel-oped in all the dorsals. What was interpreted by Salgado et al.(1997a) as the anterior branch of the ventral bifurcation of the‘infradiapophyseal’ lamina is reinterpreted here as a new, acces-sory lamina (identified as the accessory posterior centrodi-apophyseal lamina, apcdl) that rises from the pcdl approximatelyat its junction with an unnamed parapophyseal lamina (upl, seebelow). The apcdl can be seen clearly in all the dorsals, exceptfor the fourth and tenth (Figs. 3, 4). This accessory lamina is notpresent in the posteriormost dorsals of Saltasaurus (PVL-4017-134) or in a posterior dorsal of the holotype of Rocasaurus mun-iozi (MPCA-Pv 46) (Salgado and Azpilicueta, 2000).
The fourth dorsal vertebrae of Saltasaurus loricatus (PVL4017-14) and Rapetosaurus krausei (FMNH-PR 2209) have ven-trally expanded pcdl. This is due to the fact that the apcdl,though present, is not clearly defined as an individual structure.In some vertebrae of Neuquensaurus australis (i.e., the fourth,MCS-5/18, and the seventh, MCS-5/20) the apcdl is more visibleon the left than on the right side. The apcdl, not just the ventrallyforked infradiapophyseal lamina, as claimed by Salgado et al(1997a), is considered here as a possible synapomorphy of Tita-nosauria.
The pcpl, well developed in all dorsals, runs obliquely from theparapophysis towards the centrum (Figs. 3, 4). It was considereda synapomorphy of Titanosauria by Salgado et al. (1997a) and asa synapomorphy of Titanosauriformes by Wilson and Sereno(1998). It was independently developed in diplodocids (Wilson,
FIGURE 2. Neuquensaurus australis, MCS-5/17, twelfth? cervical vertebra in lateral (A) and anterior (B) views. Abbreviations: pcdl, posteriorcentrodiapophyseal lamina; podl, postzygodiapophyseal lamina; prdl, prezygodiapophyseal lamina; spol, spinopostzygapophyseal lamina; sprl, spi-noprezygapophyseal lamina; tprl, intraprezygapophyseal lamina. Scale bar equals 10 cm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005626
1999). Another lamina runs parallel to the pcpl and dorsal to it(Figs. 3, 4). It is well developed also in Rocasaurus muniozi(Salgado and Azpilicueta, 2000:fig. 5), Saltasaurus loricatus(Powell, 1986:lam. 27c), Rapetosaurus krausei (Curry Rogers andForster, 2001:fig. 3b) and in the sixth dorsal of Opisthocoelicau-dia skarzynskii (Borsuk-Bialynicka, 1977:fig. 3A6). This un-named parapophyseal lamina (informally designated as upl) isseemingly present on the right side of the vertebra MAL-182 ofMalawisaurus dixeyi Haughton, 1928 (Gomani et al., 1999; pers.obs.). It runs from the parapophyses reaching up to the pcdl atthe origin point of the apcdl. In the ninth dorsal, the upl and thelower third of the pcdl are arranged in one line, forming a stronglamina in apparent detriment to the apcdl, which is weaker thanin the precedent vertebra (Fig. 4C). In this vertebra, the pcpl isvirtually absent. The upl constitutes the lower limit of the ‘post-parapophyseal fossa,’ seen in other titanosaurs such as Argyro-saurus Lydekker, 1893 (Bonaparte, 1999:fig. 35C).
In some titanosaurs (i.e., Saltasaurus, Rapetosaurus) the areaenclosed by the upl, the pcpl and the apcdl is shallow, and thusthe limiting laminae cannot be distinguished clearly.
A third lamina, the ppdl, arises from the parapophyses, estab-lishing an oblique connection with the diapophysis (Figs. 3, 4).This lamina cannot be seen clearly in dorsals 7 and 8, due to poorpreservation (Figs. 4A, B).
In most sauropods, the prezygapophyses are ventrally sup-ported by two stout, column-like pillars, the centroprezyg-apophyseal laminae (cprl) that bound the neural canal laterally,but in Neuquensaurus australis they are not as robust as in othertaxa (Fig. 5). Although the lamina linking the prezygapophysisand the diapophysis (the prdl) is not as well developed as inRocasaurus (Salgado and Azpilicueta, 2000:figs. 3a, 4a), it isclearly present in the fourth and the fifth vertebrae (Fig. 3).
In posterior view, the neural spine is supported by severallaminae: the middle one, the postspinal lamina (posl), forms the
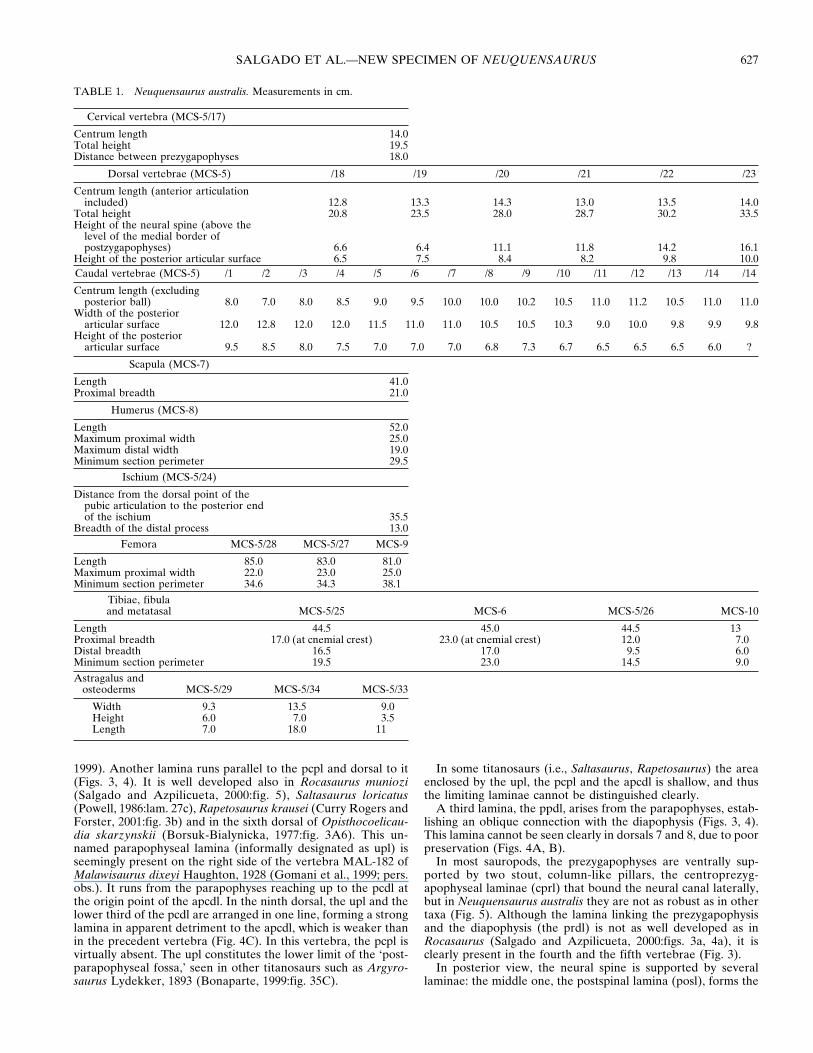
TABLE 1. Neuquensaurus australis. Measurements in cm.
Cervical vertebra (MCS-5/17)
Centrum length 14.0Total height 19.5Distance between prezygapophyses 18.0
Dorsal vertebrae (MCS-5) /18 /19 /20 /21 /22 /23
Centrum length (anterior articulationincluded) 12.8 13.3 14.3 13.0 13.5 14.0
Total height 20.8 23.5 28.0 28.7 30.2 33.5Height of the neural spine (above the
level of the medial border ofpostzygapophyses) 6.6 6.4 11.1 11.8 14.2 16.1
Height of the posterior articular surface 6.5 7.5 8.4 8.2 9.8 10.0Caudal vertebrae (MCS-5) /1 /2 /3 /4 /5 /6 /7 /8 /9 /10 /11 /12 /13 /14 /14
Centrum length (excludingposterior ball) 8.0 7.0 8.0 8.5 9.0 9.5 10.0 10.0 10.2 10.5 11.0 11.2 10.5 11.0 11.0
Width of the posteriorarticular surface 12.0 12.8 12.0 12.0 11.5 11.0 11.0 10.5 10.5 10.3 9.0 10.0 9.8 9.9 9.8
Height of the posteriorarticular surface 9.5 8.5 8.0 7.5 7.0 7.0 7.0 6.8 7.3 6.7 6.5 6.5 6.5 6.0 ?
Scapula (MCS-7)
Length 41.0Proximal breadth 21.0
Humerus (MCS-8)
Length 52.0Maximum proximal width 25.0Maximum distal width 19.0Minimum section perimeter 29.5
Ischium (MCS-5/24)
Distance from the dorsal point of thepubic articulation to the posterior endof the ischium 35.5
Breadth of the distal process 13.0
Femora MCS-5/28 MCS-5/27 MCS-9
Length 85.0 83.0 81.0Maximum proximal width 22.0 23.0 25.0Minimum section perimeter 34.6 34.3 38.1
Tibiae, fibulaand metatasal MCS-5/25 MCS-6 MCS-5/26 MCS-10
Length 44.5 45.0 44.5 13Proximal breadth 17.0 (at cnemial crest) 23.0 (at cnemial crest) 12.0 7.0Distal breadth 16.5 17.0 9.5 6.0Minimum section perimeter 19.5 23.0 14.5 9.0
Astragalus andosteoderms MCS-5/29 MCS-5/34 MCS-5/33
Width 9.3 13.5 9.0Height 6.0 7.0 3.5Length 7.0 18.0 11
SALGADO ET AL.—NEW SPECIMEN OF NEUQUENSAURUS 627

central axis in the eighth to tenth vertebrae. The spinopostzyg-apophyseal lamina (spol) encloses the posl distally. The postzyg-apophyses are short and the spinopostzygapophyseal laminae(spol) are moderately developed, in contrast to the condition inSaltasaurus. The articular facets of the postzygapophyses areoval and inclined mesio-ventrally, with their greater axes ori-ented transversely. Straight, tall, and parallel columns surroundthe neural canal, whereas the centropostzygapophyseal laminae(cpol) reach the postzygapophyses ventrally. Between the cpoland the pcdl, two deep posterior fossae are present in all theseries, the “latero-neural” fossae.
Sacrum—A nearly complete pelvis (sacrum articulated toboth ilia) was preserved (MCS-5/16) (Fig. 7A). The material has
been prepared only on its ventral side. Unlike other titanosaurs,wherein the number of sacral vertebrae is invariably six, thesacrum of MCS-5 is composed of seven vertebrae. The centra ofthe first six vertebrae are solidly co-ossified, but the seventhcentrum remains unfused. The last sacral is apparently biconvex.
Unlike other titanosaurs (MCT 1536-R) (Campos and Kellner,1999), the sacrum of Neuquensaurus australis includes stronglynarrowed third to fifth centra. Although this part of the titano-saur anatomy is not well known, we interpret this character as anautapomorphy of the species.
All of the sacral ribs except the first are preserved in MCS-5/16. Only the base of the narrow left first sacral rib is preserved.The second sacral rib, preserved only on the left side, is antero-
FIGURE 3. Neuquensaurus australis, MCS-5/18–19, fourth and fifth dorsal vertebrae in lateral (A, C) and anterior (B, D) views. Abbreviations:apcdl, accessory posterior centrodiapophyseal lamina; cpol, centropostzygapophyseal lamina; pcpl, posterior centroparapophyseal lamina; ppdl,paradiapophyseal lamina; prpl; prezygoparapophyseal lamina; upl, unnamed parapophyseal lamina; prsl, prespinal lamina; spdl, spinodiapophyseallamina. Other abbreviations as in Figure 2. Scale bar equals 10 cm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005628

laterally directed and distally expanded, as seen in the secondsacral of other titanosaurs (Kellner and Azevedo, 1999; Camposand Kellner, 1999). The rib of the third sacral is directed latero-posteriorly both in Neuquensaurus australis (MCS-5/16) and inPeirópolis sp. A (MCT 1536-R), almost reaching the base of thepubic peduncle. The fourth, fifth, and sixth ribs articulatedtightly to the acetabulum, distally fused in a sacro-costal yolk andthus participating in the articular surface of the acetabulum. This
has been proposed as a synapomorphy of the clade Barapasaurus+ Omeisaurus + Neosauropoda (Wilson and Sereno, 1998; Wil-son, 2002). The rib of the fourth sacral is still lateroposteriorlydirected. Those of the fifth and sixth are laterally directed, andthe sixth sacral rib is connected to the yolk at the level of theischiadic peduncle. The ribs of the last Neuquensaurus australissacral (seventh) are longer than those of the sixth and are later-ally directed. Distally, they are greatly expanded, articulatingwith the postacetabular region of the ilia. This indubitable con-nection with the ilia indicates that it is part of the sacrum.
Caudal Vertebrae—The vertebra considered here as the firstcaudal is anteroposteriorly short and strongly procoelous (Fig.6A; Table 1). The ventral and lateral faces of the centrum areslightly concave. The neural arch is high and the neural spineanteroposteriorly short and straight. The transverse processesare dorsoventrally expanded as in other sauropods.
The prezygapophyses, which are incompletely preserved inthis vertebra, were apparently short. Two centroprezygapophy-seal laminae (cprl) bound the neural canal, as occurs in the dorsalvertebrae. The postzygapophyses face lateroventrally. The spi-nopostzygapophyseal laminae (spol) are well developed, as is ashort lamina that connects the anterior postzygapophyseal mar-gin to the posterolateral part of the base of the neural spine, andis apparently homologous with the podl. Likewise, there are tworobust spinoprezygapophyseal laminae (sprl). In anterior view,the neural spine is pierced by many small foramina, the biggestof which is close to the prespinal lamina (prsl).
If the evidence provided by the caudal vertebra (MCS-5/1-15)is taken into account, it is obvious that one vertebra was locatedbetween Huene’s (1929) biconvex “first” caudal (actually, theseventh sacral, in lamina III, 3) and his “second” caudal (Huene,op. cit.: lamina III, 4, actually the third caudal). In turn, thevertebra MLP-Cs 1392, assigned by Huene (1929) to Neuquen-saurus robustus, would be the first caudal, not the second.
The centra of the subsequent vertebrae are all strongly pro-coelous and dorsoventrally compressed, with a wide and concaveventral surface (Fig. 6B, C, D). Conversely, although a slight
FIGURE 5. Neuquensaurus australis, MCS-5/22-23, ninth and tenthdorsal vertebrae in anterior view. Abbreviations: cprl, centroprezyg-apophyseal lamina; nc, neural canal; p, parapophysis; prz, prezygapophy-sis; tp, transverse process; udl, unnamed diapophyseal lamina. Otherabbreviations as in Figures 2–3. Scale bar equals 10 cm.
FIGURE 4. Neuquensaurus australis, MCS-5/20-23, A–D, seventh to tenth dorsal vertebrae in lateral view. Abbreviations as in Figures 2–3. Scalebar equals 10 cm.
SALGADO ET AL.—NEW SPECIMEN OF NEUQUENSAURUS 629

ridge is developed in the bottom of the ventral depression, nodefinite longitudinal ridge is present, unlike in other saltasau-rines (i.e., Saltasaurus and Rocasaurus).
The prezygapophyses are relatively short and the base of theneural arch remains implanted on the anterior half of the cen-trum, as in other Titanosauriformes (Salgado et al., 1997a). Thecaudal vertebrae are camerate, with inner open cavities distrib-uted along the laterodorsal portions of the centra. The neuralarch and transverse processes are composed mainly of spongytissue, made up of tiny cells uniformly distributed, a feature thatconstitutes a saltasaurine synapomorphy (Wilson, 2002).
The caudal vertebrae, beginning with the second, are typicallysaltasaurine in having a strongly caudally directed neural spine(Huene, 1929:lam. III), differing from the state in other titano-saurs where the spine is vertical, or even cranially inclined(Fig. 6B, C, D).
In the second and third centra, posterior to the transverseprocess, a short horizontal ridge separates two slightly concavesurfaces. The ridge lowers towards the fifth caudal, where it isbest developed. The last traces of the ridge are in the sixth andseventh vertebrae. Salgado and García (2002) observed identicalmorphological variation in other titanosaurs. They related theobserved variation to the differential origin of the caudofemora-lis longus muscle.
Posterior to the fourth caudal, the posterior inclination of theneural spines increases, and by the fifth vertebra, the spino-prezygapophyseal lamina becomes nearly horizontal. In the pos-teriormost caudals, the spinoprezygapophyseal laminae arefused distally. As in other titanosaurs, a well-developed prespinallamina is present. This lamina is also present in other specimensof Neuquensaurus australis (CS 1409, CS 1402), and it is a prob-able synapomorphy of titanosaurs, as stated by Wilson andSereno (1998:22).
Ilium—Both ilia are preserved (MCS-5/16) (Fig. 7A). Both thepreacetabular and postacetabular processes are outwardly ex-panded, as in other titanosaurs. As can be seen in other species(Campos and Kellner, 1999), the outward expansion of thepreacetabular process is accompanied by a lightening of the re-spective sacral ribs, although in the case of MCS-5/16, only thebase of first sacral rib is preserved.
If we consider a ratio between the spacing of the outermostpoints of the pubic peduncles of the ilia and the maximum iliacexpansion in the Brazilian titanosaur pelves, we obtain the fol-lowing results: in MCT 1488-R (sp. B), the ratio is 0.59; in MCT1536-R (sp. A) it is 0.63. In Neuquensaurus australis (MCS-5/16)it is 0.54 and in MLP 46-11-242, assigned to the same species, theratio is 0.57. These numbers indicate that the degree of expan-sion of the ilia is slightly greater in Neuquensaurus australis thanin the Brazilian titanosaurs. We don’t know if that difference isexplicable in terms of individual variation.
In the Brazilian pelvis MCT 1536-R (sp. A), both ilia are wing-like and laterally curved in ventral view, whereas in Neuquen-saurus (MCS-5/16) they are anterolaterally developed as straightoblique expansions. As a result, the outermost regions of theBrazilian pelvis are laterally directed, and located at the level ofthe S1-S2 boundary, whereas in Neuquensaurus they are antero-laterally directed and located right at the level of the S1.
In pelvis MCS-5/16, the preacetabular and postacetabularlobes are twisted differently, in such a way that the outer face ofthe preacetabular lobe faces upwards, whereas the outer face ofthe postacetabular lobe faces downwards. This condition occursalso in specimen MLP 46-11-242, assigned to Neuquensaurusaustralis.
The pubic peduncle is long and posteroventrally directed if thesacral axis is oriented horizontally. In ventral view, its distalsurface is subtriangular, with one of its vertices pointing inwards.The ischiadic peduncle is better developed than in Saltasaurusand Rocasaurus.
Ischium—The ischium is a flat and short bone (Fig. 7E, TableI). The iliac peduncle is well developed, and the ischiadic laminais relatively wide, but not to the extent in Rocasaurus. Likewise,the posterior process is somewhat shorter than in Rocasaurus,where its width is twice that of the iliac peduncle. The ischium ofSaltasaurus is similar to that of Neuquensaurus australis in itsrelatively narrow lamina, although it differs in that the iliac pe-duncle is relatively large relative to the whole bone (Powell,1986:lam. 43).
Scapula—A left scapula (MCS-7) associated with the Neu-quensaurus skeleton is nearly complete, although not well pre-served (Fig. 7F, Table I). However, its slender blade and smallsize do not agree with the stout limb bones of MCS-5 or withthose adult specimens housed in the Museo de La Plata Collec-tion. Taking into account these aspects, the bone will only bedescribed briefly here. Unlike the known scapulae of Neuquen-saurus, where the scapular blade has a strongly sigmoid dorsalridge and a distal expansion that resembles the condition in basaltitanosauriforms (e.g., Brachiosaurus), MCS-7 shows a straightdorsal margin, as in Lirainosaurus and Laplatasaurus araukani-cus (MLP CS 1031, MLP AV 1040). As in Rapetosaurus andmost derived titanosaurs, but differing from the condition inLirainosaurus, the scapular blade forms a 45° angle with thecoracoid boundary (Curry Rogers and Forster, 2001). However,
FIGURE 6. Neuquensaurus australis. A, MCS-5/1, first caudal verte-brae in left lateral and anterior views. B, MCS-5/2, second caudal verte-bra in left lateral, anterior, and posterior views. C, MCS-5/3-8, third toeighth caudal vertebrae. D, MCS-5/9-14, ninth to 14th caudal vertebrae.Scale bar equals 10 cm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005630

the acromion is not developed parallel to such a line, nor does itform a straight angle with the caudally displaced blade. In thisspecimen, the acromion merges with the blade to form a slightand perpendicular expansion, as is seen in Lirainosaurus. In
Laplatasaurus, on the other hand, the acromion developsabruptly from the blade. As in Isisaurus colberti and Saltasaurus,conspicuous dorsal or ventral ridges are absent on the inner side.The narrow blade resembles that of Lirainosaurus and
FIGURE 7. Neuquensaurus australis. A, MCS-5/16, pelvis in ventral view. B, MCS-5/33 and 34, osteoderms in dorsal view. C, MCS-5/28, rightfemur, MCS-5/25, tibia, MCS-5/26, fibula, and MCS-5/29, astragalus, in posterior view. D, MCS-9, right femur, and MCS-6, tibia, in posterior view.E, MCS-5/24, left ischium, in internal view. F, MCS- 7, left scapula, in lateral view. G, MCS-8, left humerus, in posterior view. H, MCS-5/25, tibia,MCS-5/26, fibula, and MCS-5/29, astragalus, in posterior and distal views. I, same, in anterior and proximal views. J, MCS-10, metatarsal, in proximaland anterior views. Abbreviations: S1 and S7, sacral vertebrae 1 and 7. Scale bars equal 10 cm except for A, C and D, where scale bars equal 50 cm.
SALGADO ET AL.—NEW SPECIMEN OF NEUQUENSAURUS 631

Laplatasaurus, and differs from the broad scapular blades ofIsisaurus colberti and Rapetosaurus, where the maximum acro-mial breadth is around one half of the total length (Jain andBandyopadhyay, 1997). Provisionally, this scapula is consideredto be Titanosauria indet.
Humerus—The left humerus (MCS-8; Fig. 7G; Table 1), un-doubtedly assigned to Neuquensaurus australis, was found asso-ciated with the bones of MCS-5, but its association with thatspecimen cannot be ensured. The bone is short and its proximalend is widely expanded. The humerus is similar to one assignedby Lydekker (1893) to Microcoelus patagonicus, which was laterreferred to Neuquensaurus australis by Huene (1929). It stronglyresembles Saltasaurus in most features. However, this humerusdoes not have the typical supracondylar ridges on the posteriorsurface, nor is the supraectepicondyilar ridge as well developedas in the species from Salta. These last features, however, couldbe due to individual variation (Powell, 1986:147).
The deltopectoral crest of the humerus is developed up to themid-length of the bone, as in Saltasaurus and Opisthocoelicaudia,and unlike those of most other titanosaurs in which the crest isdeveloped only on the proximal third of the humerus. The threegenera share a remarkably stout humerus with a wide proximalexpansion. This feature, combined with a distal expansion, wasused by Wilson and Sereno (1998) to diagnose Titanosauria.
Femur—The femora of Neuquensaurus australis are similar tothese of Saltasaurus, although they are less robust, especially theMCS-5/27 left femur and the MCS-5/28 right femur (Fig. 7C;Table 1), both found articulated to pelvis MCS-5/16. The leftfemur is slightly smaller than the right one. They are anteropos-teriorly compressed, as in most titanosaurs. As in the other tita-nosauriforms, there is a marked lateral bulge developed belowthe lesser trochanter (Salgado et al., 1997a). It is somewhat lessdeveloped in MCS-5/27 and MCS-5/28 than in MCS-9 (Fig. 7D).
As in Opisthocoelicaudia and Saltasaurus, the medial condyleis higher than the lateral one. This particular configuration of thedistal part of the bone led Wilson and Carrano (1999: fig. 5) topropose that the femora would have been slightly outwardlydirected relative to the axial plane. The distal end of the femurof Saltasaurus seems to be somewhat more expanded than inNeuquensaurus australis. This slight difference may be attributedto individual variation.
The femur MCS-9 (Fig. 7D; Table 1) is more robust thanMCS-5/27 and MCS-5/28. Owing to the fact that right tibiaMCS-6 articulates perfectly to this femur, we conclude that thesebelong to the same specimen. If so, the tibia/femur ratio wouldbe 0.55, almost identical to the value obtained with femur MCS-5/28 and tibia MCS-5/25 (0.54).
Tibia—Tibia (MCS-5/25) is short and relatively gracile [ro-bustness index (perimeter of minimum section/length) 0.44] ascompared to MCS-6, and the proximal end is massive and an-teroposteriorly expanded (Fig. 7C, H, I; Table 1). The cnemialcrest is very pronounced in anterior view. As in Saltasaurus andother titanosaurs (i.e., Gondwanatitan faustoi, Kellner and Aze-vedo, 1999:fig. 21), the lateral surface of the tibia behind thecnemial crest is slightly concave. The tibia-femur articular sur-face is rather flat. Distally the tibia expands transversely, a fea-ture seen in other titanosaurs (Wilson, 2002).
The tibia MCS-6 is stouter than MCS-5/25 (robustness index0.51), but morphologically identical to it in other respects. Asstated above, tibia MCS-6 seems to correspond to femur MCS-9(Fig. 7D; Table 1). Powell (1986) identified two different morphsof tibiae in Saltasaurus, one robust (PVL 4017-84), with arobustness index of 0.51, and the other gracile (PVL 4017-87and PVL 4017-88) with a robustness index of 0.47 (Powell, 1986:table 18).
Fibula—A fibula (MCS-5/26) was found articulated to tibiaMCS-5/25. This element is somewhat shorter than the tibia (Fig.7C, H, I; Table 1). It is relatively gracile and expanded at both
ends. As in other titanosaurs, the outer surface is convex,whereas the inner one is flat and slightly concave. The lateralprotuberance, in turn, is well demarcated.
Astragalus—The astragalus is high and transversely reduced,as in other titanosaurs (Borsuk-Bialynicka, 1977; Powell, 1986)(Fig. 7C, H, I; Table 1). In Opisthocoelicaudia, the breadth of theastragalus is 54% of the distal breadth of the tibia (Borsuk-Bialynicka [1977:8] listed “astragalus reduced in dimensions” asa probable diagnostic character of this species). In this manner,the transversely short astragalus is not a diagnostic character ofNeuquensaurus australis but possibly of a group of titanosaurs.The fact that the titanosaur astragalus is transversely reducedmeans that the transverse expansion of the distal end of thetitanosaur tibia was not accompanied by a widening of the as-tragalus. There is no evidence of an ossified calcaneum. The facefor the fibula is concave, whereas the broader tibial face is in-clined.
Metatarsals—Only one metatarsal (MCS-10) was found asso-ciated with this specimen. The bone, here interpreted as a thirdmetatarsal (Fig. 7J; Table 1), is stout with well-developed ends.In proximal view, the bone is rather triangular. The antero-proximal face bears a middle prominence bordered by two lat-eral grooves. In posterior view, the proximal half of the bone hasa deep central depression separated from two lateral and medialexternal depressions by two robust keels.
Osteoderms—two osteoderms have been recovered associ-ated with this skeleton (MCS-5) (Fig. 7B; Table 1). These piecesroughly coincide with the ones described collectively by Huene(1929) as “Type I”. The biggest piece has the typical shallowdepression on the dorsal face and a longitudinal ridge on theventral face. The smallest one lacks these features. The record ofthese bones in Neuquensaurus australis is important becausesimilar plates, found in the same locality, were previously as-signed to Titanosaurus araukanicus (Powell, 1980).
DISCUSSION
As stated by Powell (1986, 1987b, 1992), Neuquensaurus aus-tralis is most similar to Saltasaurus loricatus. The most obviousdifferences between the two species are in the caudal and sacralvertebrae and in the appendicular skeleton, mostly in the iliumand ischium. The most significant difference in the morphologyof the dorsal vertebrae is in the height of the neural spines.
A striking feature of Neuquensaurus australis, at least of thespecimen described herein (MCS-5), is the presence of sevensacral vertebrae. Taking this into account, we could state that thebiconvex centrum MLP-Cs 1390 (Huene, 1929:lam. III, 3) wouldnot be the first caudal at all but the last sacral centrum, which isusually not fused to the previous sacral, as is seen in other tita-nosaurs (Kellner and Azevedo, 1999).
Another interesting aspect of Neuquensaurus taxonomy is theapparent existence of two different morphs, one slender (repre-sented by skeleton MCS-5; Fig. 7C) and the other robust (rep-resented by MCS-6 and MCS-9; Fig. 7D). A similar situationoccurs in El Brete (Salta Province), where two different classesof bones are found in the quarry (Powell, 1986). A possibilitywould be to consider the coexistence of two different species.This was the interpretation of Huene, who erected a new species(Neuquensaurus robustus Huene, 1929) for the robust morph.However, Powell (1986) rejected this specific separation. Forhim, most of the differences noted between the morphs were notof taxonomic value. He left open the possibility that at least someof the dissimilarity could be due to individual or sexual variation.The evidence presented in this paper does not support eitherhypothesis.
ACKNOWLEDGMENTS
We would like to thank J. Calvo, W. Lagos, M. Poblete, I.Cerda, M. Gutierrez, N. Granchelli, and the people of Cinco
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005632

Saltos for their assistance during the fieldwork and for allowingus to study the material. Federico Poblete and William Barraprepared part of the material. Rodolfo Coria read the manu-script and provided technical assistance. Jorge A. González hasskillfully made the drawings. Kristi Curry Rogers, Jeff Wilson,Paul Upchurch, Matt Carrano, Michael Parrish, and Jenni Scottmade useful comments for improving the manuscript.
LITERATURE CITED
Andreis, R. R., A. M. Iñiguez Rodríguez, J. J. Lluch, and D. A. Sabio.1974. Estudio sedimentológico de las Formaciones del CretácicoSuperior del área del Lago Pellegrini (provincia de Río Negro,República Argentina). Revista de la Asociación Paleontológica Ar-gentina 29:85–104.
Bonaparte, J. F. 1999. Evolución de las vértebras presacras en Sau-ropodomorpha. Ameghiniana 36:115–187.
Bonaparte, J. F., and R. A. Coria. 1993. Un nuevo y gigantesco sau-rópodo titanosaurio de la Formación Río Limay (Albiense-Cenomaniense) de la Provincia del Neuquén, Argentina. Ameghini-ana 30:271–282.
Bonaparte, J. F., and Z. B. Gasparini. 1980. Los saurópodos de los gruposNeuquén y Chubut, y sus relaciones cronológicas. Actas V CongresoGeológico Argentino, Neuquén 2:393–406.
Bonaparte, J. F., and J. E. Powell. 1980. A continental assemblage oftetrapods from the Upper Cretaceous beds of El Brete, northwest-ern Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Eco-systèmes Continentaux du Mésozoïque. Mémoires de la SociétéGéologique de France (n.s.) 139:19–28.
Borsuk-Bialynicka, M. 1977. A new camarasaurid sauropod Opisthocoe-licaudia gen. n. sp. n. from the Upper Cretaceous of Mongolia.Paleontologia Polonica 37:5–64.
Campos, D. A., and A. W. A. Kellner. 1999. On some sauropod (Tita-nosauridae) pelves from the continental Cretaceous of Brazil; pp.143–166 in Y. Tomida, T. H. Rich, and P. Vickers-Rich (eds.), Pro-ceedings of the Second Gondwanan Dinosaur Symposium. NationalScience Museum Monograph No. 15, Tokyo.
Cazau, L. B., and M. A. Uliana. 1973. El Cretácico Superior continentalde la Cuenca Neuquina. Actas V Congreso Geológico Argentino3:131–163.
Coria, R. A. and L. Salgado. 1996. A basal iguanodontian (Ornithischia:Ornithopoda) from the Late Cretaceous of South America. Journalof Vertebrate Paleontology 16:445–457.
Curry Rogers, K., and C. Forster, 2001. The last of the dinosaur titans: anew sauropod from Madagascar. Nature 412: 530–534.
Digregorio, J. H., and M. A. Uliana. 1979. Cuenca Neuquina, pp.985–1032 in J. Turner (ed.), Segundo Simposio de Geología Re-gional Argentina. Academia Nacional de Ciencias de Córdoba,Tomo II.
Dingus, L., J. Clarke, G. R. Scott, C. Swisher, L. M. Chiappe, and R. A.Coria. 2000. Stratigraphy and magnetostratigraphic/faunal con-straints for the age of Sauropod embryo-bearing rocks in the Neu-quén Group (Late Cretaceous, Neuquén province, Argentina).American Museum Novitates 3290:1–11.
Gomani, E. M., L. L. Jacobs, and D. A. Winkler. 1999. Comparison of theAfrican titanosaurian, Malawisaurus, with a North American EarlyCretaceous sauropod; pp. 223–233 in Y. Tomida, T. H. Rich, and P.Vickers-Rich (eds.), Proceedings of the Second Gondwanan Dino-saur Symposium. National Science Museum Monographs, No. 15,Tokyo.
Groeber, P. 1946. Observaciones geológicas a lo largo del meridiano 70°1. Hoja Chos Malal. Revista de la Sociedad Geológica Argentina1:177–208.
Haughton, S. H. 1928. On some reptilian remains from the dinosaur bedsof Nyasaland. Transactions of the Royal Society of South Africa16:67–75.
Heredia, S. E., and L. Salgado. 1999. Posición estratigráfica de los estra-tos supracretácicos portadores de dinosaurios en Lago Pellegrini,Patagonia Septentrional, Argentina. Ameghiniana 36:229–234.
Huene, F. von. 1929. Los saurisquios y ornitisquios del Cretáceo Argen-tino. Anales del Museo de La Plata, Serie 2 3:1–196.
Huene, F. von. 1932. Die fossile Reptile-Ordnung Saurischia, ihre En-twicklung und Geschichte. Monographie Geologie und Palaeontolo-gie, Leipzig 1(4):1–361.
Hugo, C. A., and H. A. Leanza. 2001. Hoja Geológica 3969-IV, GeneralRoca, provincias del Neuquén y Río Negro. Instituto de Geologíay Recursos Naturales, SEGEMAR, Buenos Aires, Boletín 308:1–71.
Jain, S. L., and S. Bandyopadhyay. 1997. New titanosaurid (Dinosauria:Sauropoda) from the Late Cretaceous of Central India. Journal ofVertebrate Paleontology 17:114–136.
Kellner, A. W. A., and S. A. K. De Azevedo. 1999. A new sauropoddinosaur (Titanosauria) from the Late Cretaceous of Brazil; pp.111–142 in Y. Tomida, T. H. Rich, and P. Vickers-Rich (eds.), Pro-ceedings of the Second Gondwanan Dinosaur Symposium. NationalScience Museum Monographs, No. 15, Tokyo.
Leanza, H. A. 1999. The Jurassic and Cretaceous Terrestrial Beds fromSouthern Neuquén Basin, Argentina. Instituto Superior de Correl-ación geológica (Insugeo), Miscelánea 4, 30 pp.
Leanza, H. A., and C. A. Hugo. 2001. Cretaceous red beds from southernNeuquén Basin (Argentina): age, distribution and stratigraphic dis-continuities; pp. 117–122 in VII International Symposium on Meso-zoic Terrestrial Ecosystems, Asociación Paleontológica Argentina,Special Paper 7.
Lydekker, R. 1893. Contributions to the study of the fossil vertebrates ofArgentina. I, the dinosaurs of Patagonia. Anales del Museo de LaPlata, Paleontología 2:1–14.
Powell, J. E. 1980. Sobre la presencia de una armadura dérmica en al-gunos dinosaurios titanosáuridos. Acta Geológica Lilloana 15:41–47.
Powell, J. E. 1986. Revisión de los titanosáuridos de América del Sur.Unpublished Ph.D. dissertation, National University of Tucumán,Tucumán, 340 pp.
Powell, J. E. 1987a. Morfología del esqueleto axial de los dinosauriostitanosáuridos (Saurischia, Sauropoda) del Estado de Minas Gerais,Brazil; pp. 155–171 in Anais 10° Congresso Brasileiro de Paleonto-logia, Río de Janeiro, Brazil.
Powell, J. E. 1987b. Part VI. The titanosaurids; pp. 155–171 in Bonaparte,J. F. (ed.), The Late Cretaceous fauna of Los Alamitos, Patagonia,Argentina. Revista del Museo Argentino de Ciencias Naturales“Bernardino Rivadavia.” Paleontología III(3).
Powell, J. E. 1992. Osteología de Saltasaurus loricatus (Sauropoda-Titanosauridae) del Cretácico Superior del Noroeste Argentino;pp. 165–230 in J. L. Sanz and A. D. Buscalioni (eds.), Los Dinosau-rios y su Entorno biótico, Instituto “Juan de Valdéz,” Cuenca,Spain.
Ramos, V. 1981. Descripción Geológica de la Hoja 33 C. Los ChihuidosNorte, Provincia de Neuquén. Servicio Geológico Nacional, Boletín18:1–103.
Ramos, V. 1985. Mesozoico de la Alta Cordillera: facies y desarrolloestratigráfico, Argentina. 5to. Congreso Geológico Chileno (San-tiago). Actas 1:492–513.
Salgado, L., and C. Azpilicueta. 2000. Un nuevo saltasaurino (Sauropoda,Titanosauridae) de la provincia de Río Negro (Formación Allen,Cretácico Superior), Patagonia, Argentina. Ameghiniana 37:259–264.
Salgado, L., and R. García. 2002. Variación morfológica en la secuenciade vértebras caudales de algunos saurópodos titanosaurios. RevistaEspañola de Paleontología 17(2): 211–216.
Salgado, L., R. A. Coria, and J. O. Calvo. 1997a. Evolution of titanosauridsauropods. I: phylogenetic analysis based on the postcranial evi-dence. Ameghiniana 34:3–32.
Salgado, L., R. A. Coria, and S. E. Heredia. 1997b. New materials ofGasparinisaura cincosaltensis (Ornithischia, Ornithopoda) from theUpper Cretaceous of Argentina. Journal of Paleontology 71:933–940.
Sanz, J. L., J. E. Powell, J. Le Loeuff, R. Martínez, and X. Pereda-Suberbiola. 1999. Sauropod remains from the Upper Cretaceous ofLaño (northcentral Spain). Titanosaur phylogenetic relationships;pp. 235–255 in Estudios del Museo de Ciencias Naturales de Álava14, Special Paper 1.
SALGADO ET AL.—NEW SPECIMEN OF NEUQUENSAURUS 633

Seeley, H. G. 1883. On the classification of the fossil animals commonlycalled Dinosauria. Proceedings of the Royal Society, London 43:165–171.
Uliana, M. A., and D. A. Dellapé. 1981. Estratigrafía y evolución paleo-ambiental de la sucesión Maastrichtiano-Eoterciaria del Engolfa-miento Neuquino (Patagonia Septentrional). 8° CongresoGeológico Argentino (San Luis), Actas 3:673–711.
Wilson, J. A. 1999. A nomenclature for vertebral laminae in sauropodsand other saurischian dinosaurs. Journal of Vertebrate Paleontology19:639–653.
Wilson, J. A. 2002. Sauropod dinosaur phylogeny: critique and cladisticanalysis. Zoological Journal of the Linnean Society 136:217–276.
Wilson, J. A., and M. T. Carrano. 1999. Titanosaurs and the origin of“wide-gauge” trackways: a biomechanical and systematic perspec-tive on sauropod locomotion. Paleobiology 25:252–267.
Wilson, J. A., and P. C. Sereno. 1998. Early evolution and higher-levelphylogeny of sauropod dinosaurs. Society of Vertebrate Paleontol-ogy, Memoir 5:1–68.
Submitted 8 February 2005; accepted 19 February 2005.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 25, NO. 3, 2005634




















