
Seasonal distribution and duration of the planktonic stage of Dover sole, Solea solea, larvae in the...
14
Journal of Fish Biohgy (1993) 43 (Supplément A), 17-30 Seasonal distribution and duration of the planktonic stage of Dover sole, Solea solea, larvae in the Bay of Bîscay: an hypothesîs R. AMARA*, F. LAGARDERE! AND Y. DESAUNAY* *IFREMER Centre de Nantes. B.P. 1049. 44037 Nantes Cedex and ^Centre de Recherche en Ecologie Marine et Aquaculture (CNRS-ïFREMER), BP 5, F. 17135 L'Houmeau, France The possibility of dçriving a prediclion about the cflleci of seasonal variations în the duration of ihe planktonic larval phase on the dispersât of Jarval Dover sole was investigated. During six cruises, from February to May 1992, the distributioa of sole larvae was studied along a lOO-km transect, from the offshore spawning grounds to the coastal nurseries of the Bay of Biscay (France). Samples {n= 189) were collected with a suprabenthic samplcr, and vertical profiles of water tempcrature and salinily wcrt recorded simultaoeously. Counts of otolith incréments of [arval stage 4b (onset of metamorphosis) were used to esttmate the duration of the planktonic lifc. Duration of the larval phase decreases by about 15 days (57%) with water température increasc (between 8' in February aad 11-2' C in May). Sole larvae occur from the coastal area to 100 km offshore. Within the same cruise, no différence in the duration of ihe planktonic life was observed between the larvae caught in the onshore and the offshore area. In spile of seasonal différences in abundance, the extent and the shapc of îhc larval distributions during the period of study suggcst that the seasonal variations in the duration of the planktonic life did not affect the larval distribution. Key words: Soha solea; planktonic larval duration; distribution. I. INTRODUCTION Since Hjort (1914), many studies have focused on factors acting during the early life of fishes to explain recruitment variability. Survival and recruitment of young fishes dépend on the combined action of biologîcal and physical factors such as starvation, prédation, larval transport and température (Yoklavich & Bailey, 1990). To make significant advances in the study of fish population dynamics, it is necessary to define the interrelationships of biological and physical variables (Rothschild, 1986). For example, Boehiert & Mundy (1988) suggest that environmental variables such as light, température, and prey distribution, because they affect the vertical distribution of fish larvae, may control the passive drift. In fish species where spawning and nursery areas are spatially separated, the transport of eggs and larvae from areas of spawning/ hatching can be one of the most important factors affecting recruitment (Nelson et al, \9V\, 1981; Norcross & Shaw, 1984; Bergman et a/., 1989; K i m et al, 1989). A knowledge of factors affecting transport of larvae should help in the understanding of recruitment variability. Since 1984. the northenipartof the Bay of Biscay hasbeen the site ofextensive studies on spawning and early pelagic larval stages of Dover sole, Solea solea (L.) (for a review. see Koutsikopoulos & Lacroix, 1992). In this repon, where the spawning activity occurs at 40 to 80 km offshore, Koutsikopoulos et al (1991) 17 lOQ-; The Fishcries Society of the British Isies 0022-1112^93/43^017+14 SÛ8.00/0 © 1993 "me tisûenes y
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Seasonal distribution and duration of the planktonic stage of Dover sole, Solea solea, larvae in the...

Journal of Fish Biohgy (1993) 43 (Supplément A), 17-30
Seasonal distribution and duration of the planktonic stage of Dover sole, Solea solea, larvae in the Bay of Bîscay: an
hypothesîs
R . AMARA*, F . LAGARDERE! AND Y. DESAUNAY* *IFREMER Centre de Nantes. B.P. 1049. 44037 Nantes Cedex and ̂ Centre de
Recherche en Ecologie Marine et Aquaculture (CNRS-ïFREMER), BP 5, F. 17135 L'Houmeau, France
The possibility of dçriving a prediclion about the cflleci of seasonal variations în the duration of ihe planktonic larval phase on the dispersât of Jarval Dover sole was investigated. During six cruises, from February to May 1992, the distributioa of sole larvae was studied along a lOO-km transect, from the offshore spawning grounds to the coastal nurseries of the Bay of Biscay (France). Samples {n= 189) were collected with a suprabenthic samplcr, and vertical profiles of water tempcrature and salinily wcrt recorded simultaoeously. Counts of otolith incréments of [arval stage 4b (onset of metamorphosis) were used to esttmate the duration of the planktonic lifc. Duration of the larval phase decreases by about 15 days (57%) with water température increasc (between 8' in February aad 11-2' C in May). Sole larvae occur from the coastal area to 100 km offshore. Within the same cruise, no différence in the duration of ihe planktonic life was observed between the larvae caught in the onshore and the offshore area. In spile of seasonal différences in abundance, the extent and the shapc of îhc larval distributions during the period of study suggcst that the seasonal variations in the duration of the planktonic life did not affect the larval distribution.
Key words: Soha solea; planktonic larval duration; distribution.
I. INTRODUCTION
Since Hjort (1914), many studies have focused on factors acting during the early life of fishes to explain recruitment variability. Survival and recruitment of young fishes dépend on the combined action of biologîcal and physical factors such as starvation, prédation, larval transport and température (Yoklavich & Bailey, 1990). To make significant advances in the study of fish population dynamics, it is necessary to define the interrelationships of biological and physical variables (Rothschild, 1986). For example, Boehiert & Mundy (1988) suggest that environmental variables such as light, température, and prey distribution, because they affect the vertical distribution of fish larvae, may control the passive drift. In fish species where spawning and nursery areas are spatially separated, the transport of eggs and larvae from areas of spawning/ hatching can be one of the most important factors affecting recruitment (Nelson et al, \9V\, 1981; Norcross & Shaw, 1984; Bergman et a/., 1989; K i m et al, 1989). A knowledge of factors affecting transport of larvae should help in the understanding of recruitment variability.
Since 1984. the northenipartof the Bay of Biscay hasbeen the site ofextensive studies on spawning and early pelagic larval stages of Dover sole, Solea solea (L.) (for a review. see Koutsikopoulos & Lacroix, 1992). In this repon, where the spawning activity occurs at 40 to 80 km offshore, Koutsikopoulos et al (1991)
17 lOQ-; The Fishcries Society of the British Isies 0022-1112^93/43^017+14 SÛ8.00/0 © 1993 "me tisûenes y

18 R . AMARA ET AL.
noted the stability of eggs and pelagic larval distributions. They suggested that a small fraction of the pelagic larvae reached the coastal nursery grounds by diffusion, providing th'e continuity between spawning sites and juvénile nursery areas. In this area, the dispersai of sole larvae can be considered as an important déterminant in the distributional pattems and recruitment to the nursery grounds. The scale of dispersai would certainly dépend on movement of the water masses, larval behaviour, and duration of planktonic life.
Laboratory experiments (Laurence, 1975; Fonds, 1979; Policansky, 1982; Seikai et al., 1986), have shown that the duration of the planktonic larval stages in fishes varies inversely with température. I f larval duration is related to the utilization of transport or rétention processes, then the différence in the duration of larval stages due to seasonal or interannual variation in water température could affect the success of the transport and in this way the recruitment to the nurseries.
In this paper we use the results of six surveys to examine the seasonal distribution of sole larvae along an inshore-offshore transect, and the variation of the duration of the planktonic larval stage. Thèse results were analysed and discussed in relation to the question of dispersai during the season. The question is whether the observed variation in the duration of the planktonic stages affects the horizontal distribution and drift,
I L MA TERU LS AND METHODS
SAMPLE COLLECTIONS Sole larvae were collected during six cruises over a fixed transect as pari of the
National Research Programme on the Determinism of Recruitment. Collections were conducted from February to May 1992. Samples were collected continuously along the transects at irregular interval (mean space between stations 3-4 km) from the offshore spawning grounds (100 km) to the coastal nurseries of Bay of Vilaine, Bay of Biscay (Fig. I , Table I). Since the majority of larvae were distributed near the bottom in day time (Koutsikopoulos et al., 1991), collections were made during daytime with a Zebulon suprabenthic sledge. This siedge is particularly designed to coUect metamorphosing sole close to the bottom (0-1 m). The apparatus consists of a steel sledge loaded with a rectangular aluminium frame (2 m wide x I m high) to which two plankton nets (1-3-mm mesh size, 1 opening) were fastened. Water volume filtered was estimated with two General Oceanic flowmeters mounted inside each net mouth. The Zebulon was hauled on average for 15 min at 2-5 knots (4-6 km h~ *) and the total water volume filtered was about 3282 m"* ( ± 599). Vertical profiles of water température and salinity were recorded during ail cruises by conductivity température, depth (CTD) (Valeport model 600 MK2). Larvae were removed immediately follotting the haul and preserved in 95% ethanol for further otolith examination. Sole larvae were classified into five morphological stages as described by Al-Maghazachi Se Gibson (1984) for the turbot, Scophlhalmus maximus (L.), and reviewed by Lagardere for Dover sole (unpublished data). For each developmental stage the catches were expressed in numbers per 1000 m'. Stages 4 and 5 were subdivided into two substages; stage 4a (beginning of the urostylar torsion), stage 4b (onset of the metamorphosis when the left eye be^ns to migrate), stage 5a and 5b (respectively when the left eye is just past the centre of dorsum and eye migration complète). Variations in the horizontal distributions for the différent stages were evaluated by calculating the médian and the range of their distribution, considered respectively as the mean location and as an index of dispersion. We have preferred the médian to the centre of gravity, because the médian is a more représentative measure of the location, when the distributions are asymmelric (Sokal & Rohlf, 1981) as in our study.
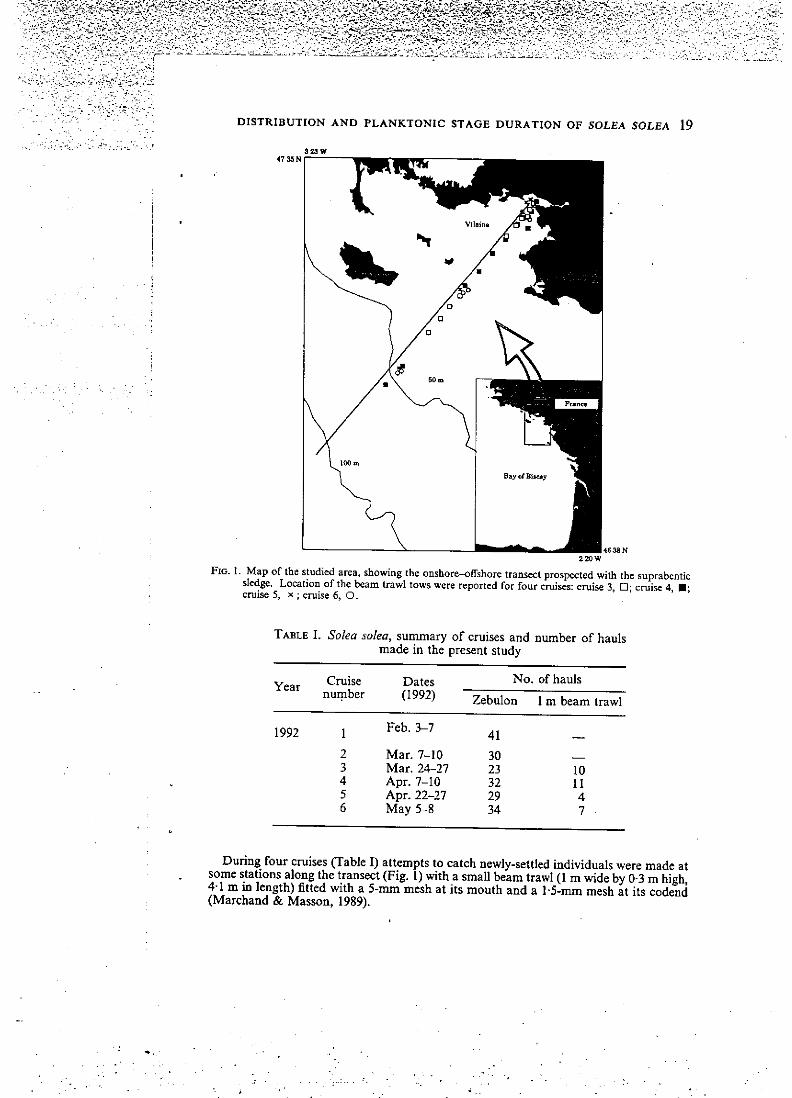
D I S T R I B U T I O N A N D P L A N K T O N I C S T A G E D U R A T I O N O F S O L E A S O L E A 19
32aW
! 2 0 W
F I G . L Map of the studied area, showîng the onshore-offshorc transect prospccted with the suprabcntic sledge. Location of the beam trawl tows were reported for four cniises: cruise 3, Q; cruisc 4, • ; cruise 5, x ; cruise 6, O.
T A B L E I . Solea solea, summary of cruises and number of hauls made in the présent study
Year Cruise number
Dates (1992)
No. of hauls
Zebulon 1 m beam trawl
1992 1 Feb. 3-7 41 — 2 Mar. 7-10 30 — 3 Mar. 24-27 23 10 4 Apr. 7-10 32 11 5 Apr. 22-27 29 4 6 May 5-8 34 7
During four cruises (Table I) attempts to catch newly-settled individuals were made at some stations along the transect (Fig. î) with a small beam trawl (1 m wide by 0-3 m high, 4-1 m in length) fitted with a 5-mm mesh at its mouth and a 1-5-mm mesh at ils codend (Marchand & Masson, 1989).

20 R . AMARA E T A t ,
TABLE I L Soïea solea, summary statistics for âge of larvae stage 3 and 4b (onset of metamorphosis)
Cruise no. Age at stage 3* n Age at stage 4b* n
1 — 40-7 (2-28) 15 2 — — 37-7 (2-03) 20 3 27-4 (1-96) 15 36-7 (2-98) 41 4 — — 35-4 (5-47) 8 5 18-8 (4-49) 15 28-7 (2-34) 31 6 15-4 (4-06) 15 25-5 (1-76) 19
•Age since mouth-opening (days). Average values (s.D.) are presented by cruises. n, number of agcd larvae.
OTOLITH PREPARATION
Sagittal otoliths were used for ageing after daily incréments had been validated (Lagardere, 1989; Lagardere & Troadec, 1992). Larvae were placed in a drop of water, the sagittae were located with the use of polarized light, removed from their otic capsule and mounted whole in Permount on a cover-slip. The sagittae were viewed by transmitted light with a magnification range of 600 to 1250. Otoliths with occluded incréments were mounted in cyanoacrylate glue and poHshed manually in the sagittal plane with a graded séries of aluminiimi papers (3 to OT jim) with periodic viewing to monitor progress until the primordia were exposed. OtoUths were photographed using a Leitz compound microscope on high contrast black and white film and incréments were counted on microphotographs. Because of faint incréments between the hatching and mouth-opening check (Lagardere & Chaumillon, 1988), âge was estimated by counting the numbers of incréments from the mouth-opening check to the edge of the otolith.
ESTIMATION OF THE DURATION OF THE PLANKTONIC LARVAL STAGE
In this study the duration of the planktonic larval stage was defined as days from the mouth-opening to onset of metamorphosis (stage 4b). One hundred and thirty-four larvae of stage 4b from différent cruises were aged (Table II). To assess the influence of environmental factors such as water température and food availability on the duration of the planktonic phase, the mean températures recorded between 10 and 80 km from the coast (areas where the majority of the larvae were caught) were reported for each cruise. For information about the seasonal availability of food, we have analysed the gut contents for 141 stage 4b larvae during the study period.
in. R E S U L T S
HYDROLOGICAL CONDITIONS Hydrological conditions observed along the transect indicated a pronounced
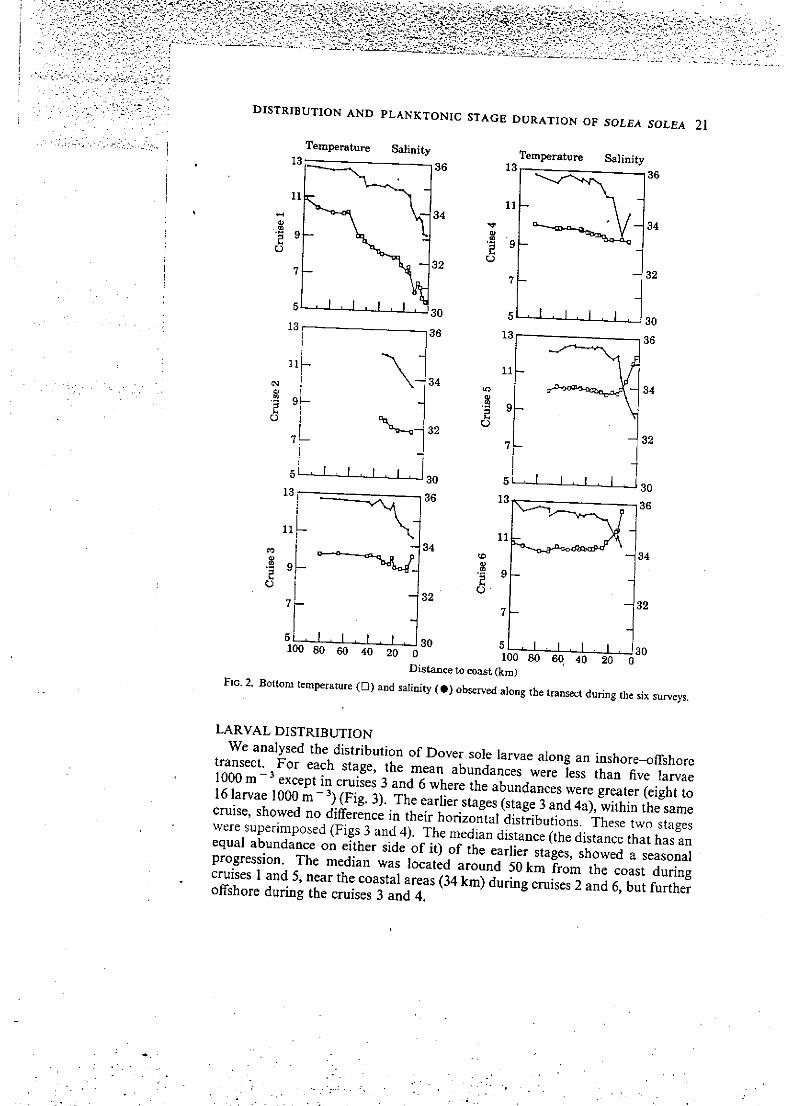
horizontal gradient (Fig. 2). The offshore waters were relatively stable, bottom température ranging from 10 to 13* C and bottom salinity staying higher than 35%o during the study period. In the coastal areas, bottom températures were low in February (<7° C) and early March (cruises 1 and 2). During the cruises 3 and 4 the horizontal thermal gradient disappeared (bottom températures were stable, from 9 to 10° C ail along the transect). From late April (cruise 5) bottom températures got warmer in the coastal area. Water salinity in the coastal area varied from 33 to 34%o during the study period.
DISTRIBUTION AND P L A N K T O N I C STAGE DURATION OF SOLEA SOLEA 21
Température Salinity 13— • 136
Température Salinity 13, 3̂6
100 80 60 40 20 0 100 80 60 40 20 0 Distance to coast (km)
F I G . 2. Bottom température (D) and salinity ( • ) observed along the transect during the six surveys.
LARVAL DISTRIBUTION We analysed the distribution of Dover sole larvae along an inshore-offshore
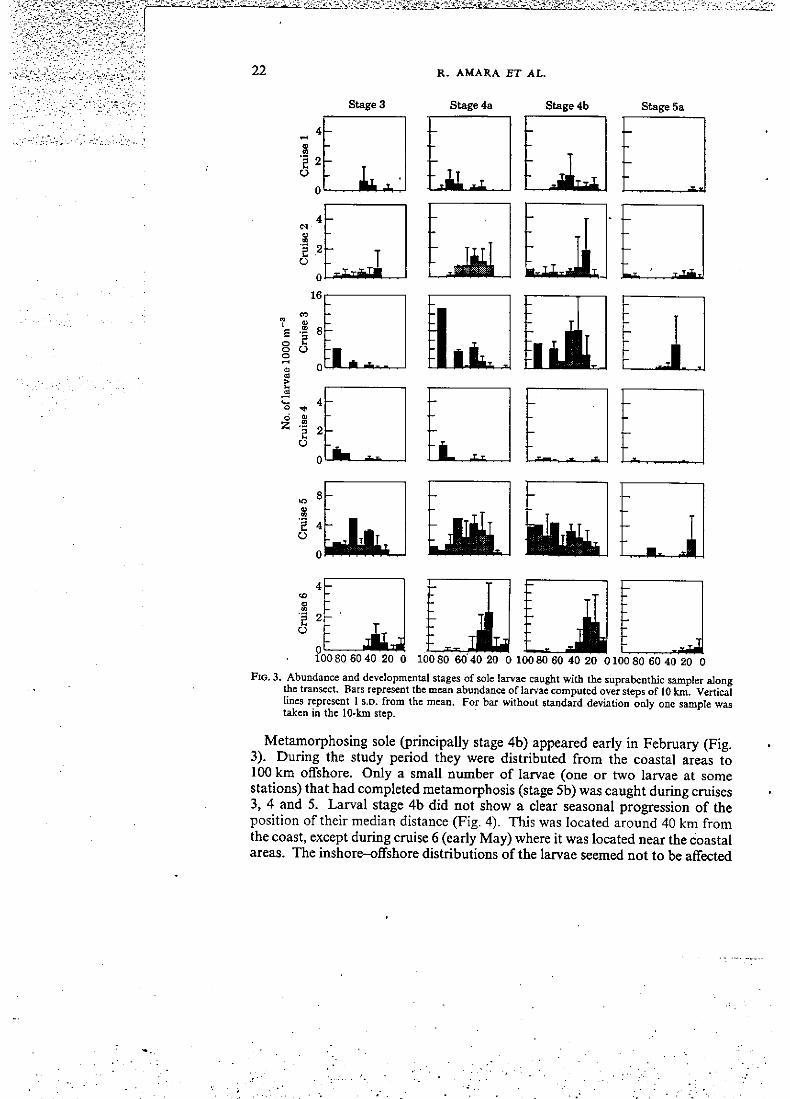
transect. For each stage, the mean abundances were less than five larvae 10(X) m " ' except in cruises 3 and 6 where the abundances were greater (eight to 16 larvae 1000 m " (Fig. 3). The earlier stages (stage 3 and 4a), within the same cruise, showed no diiference in their horizontal distributions. Thèse two stages were superimposed (Figs 3 and 4). The médian distance (the distance that has an equai abundance on either side of it) of the earlier stages, showed a seasonal progression. The médian was located around 50 km from the coast during cruises 1 and 5, near the coastal areas (34 km) during cruises 2 and 6, but further offshore during the cruises 3 and 4,
22 R . AMARA ET AL.
Stages Stage 4a Stage 4b Stage 5a
4 -
5 2 -
4 -iN • a) n e . 2 -
o r , T i T r * T i
16
7 a) -
g o P
-
TA,
-
^ J 1 ,
*S 4 -o
e 4 o
4 -
o 1 0 100 80 60 40 20 0 100 80 60 40 20 0 100 80 60 40 20 0100 80 60 40 20 0
Fio, 3. Abundance and developmental stages of sole larvae caught with the suprabenthic samplcr along the transect. Bars represent the mean abundance of larvae compulcd over steps of 10 km. Vertical lincs represent 1 S.D. from the mean. For bar without standard déviation only one sample was taken in the 10-kra step.
Metamorphosing sole (principally stage 4b) appeared early in February (Fig. 3). During the study period they were distributed from the coastal areas to 100 km offshore. Only a small nimiber of larvae (one or two larvae at some stations) that had completed metamorphosis (stage 5b) was caught during cruises 3, 4 and 5. Larval stage 4b did not show a clear seasonal progression of the position of their médian distance (Fig. 4). This was located around 40 km from the coast, except during cruise 6 (early May) where it was located near the coastal areas. The inshore-offshore distributions of the larvae seemed not to be aifected
D I S T R I B U T I O N A N D P L A N K T O N I C S T A G E D U R A T I O N O F SOLEA SOLEA 23
100
80 B
ë 60 a ta
S - g
40 -S
• • o 20 - o
o
0 1 2 3 4 5 6 Omise No,
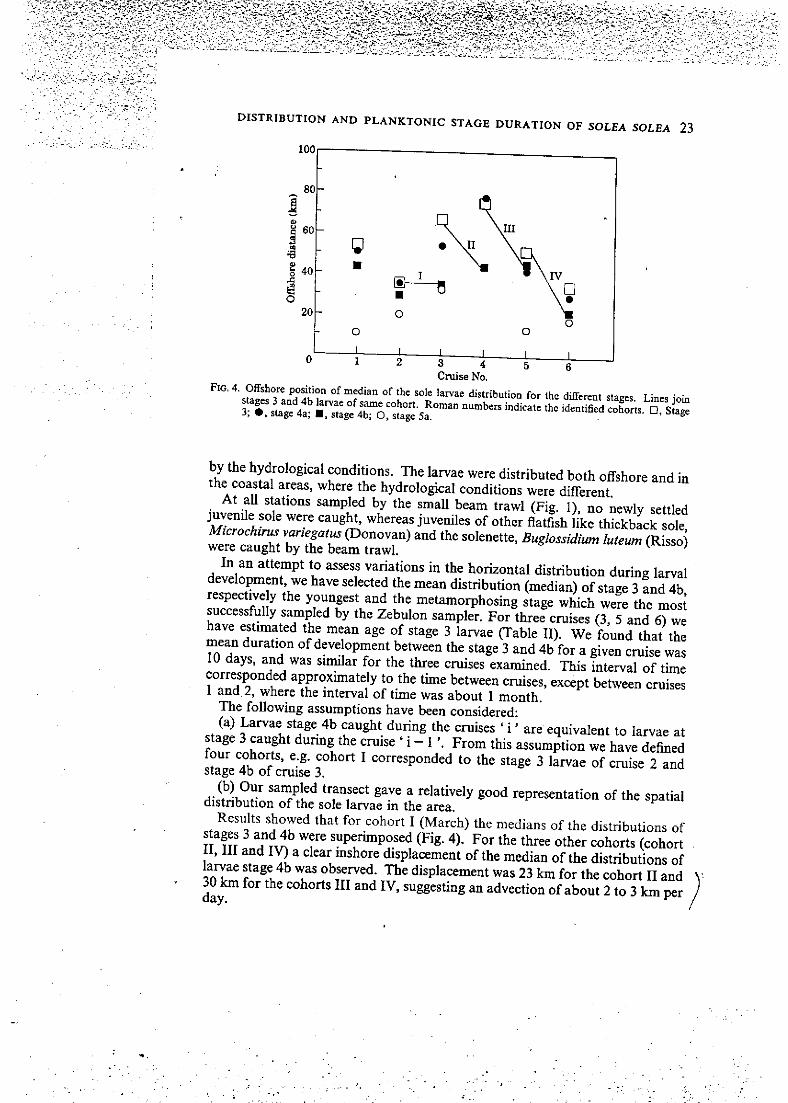
FIO. 4. Offshore position of médian of the sole larvae distribution for Ihc différent stages. Lines join stages 3 and 4b larvae of same cohort. Roman numbers indicatc the idcntified cohorts. • , Stage 3; stage 4a; • , stage 4b; O, stage 5a.
by the hydrological conditions. The larvae were distributed both offshore and in the coastal areas, where the hydrological conditions were différent,
At ail stations sampled by the small beam trawl (Fig. 1), no newly settled juvénile sole were caught, whereas juvéniles of other flatfîsh like thickback sole, Microchirus variegaius (Donovan) and the solenette, Bugîossidium îuteum (Risso) were caught by the beam trawl.
In an attempt to assess variations in the horizontal distribution during larval development, we have selected the mean distribution (médian) of stage 3 and 4b, respectively the youngest and the metamorphosing stage which were the most successfuUy sampled by the Zebulon sampler. For three cruises (3, 5 and 6) we have estimated the mean âge of stage 3 larvae (Table II) . We found that the mean duration of development between the stage 3 and 4b for a given cruise was 10 days, and was similar for the three cruises examined. This interval of time corresponded approximately to the time between cruises, except between cruises l and 2, where the interval of time was about l month.
The foUowing assumptions have been considered: (a) Larvae stage 4b caught during the cniises * i ' are équivalent to larvae at
stage 3 caught during the cruise * i — l '. From this assuraption we have defined four cohorts, e.g. cohort I corresponded to the stage 3 larvae of cruise 2 and stage 4b of cruise 3,
(b) Our sampled transect gave a relatively good représentation of the spatial distribution of the sole larvae in the area.
Results showed that for cohort I (March) the médians of the distributions of stages 3 and 4b were superimposed (Fig. 4). For the three other cohorts (cohort I I , I I I and IV) a clear inshore displacement of the médian of the distributions of larvae stage 4b was observed. The displacement was 23 km for the cohort I I and 30 km for the cohorts I I I and IV, suggesting an advection of about 2 to 3 km per day.
60 80 Days
100 120 140
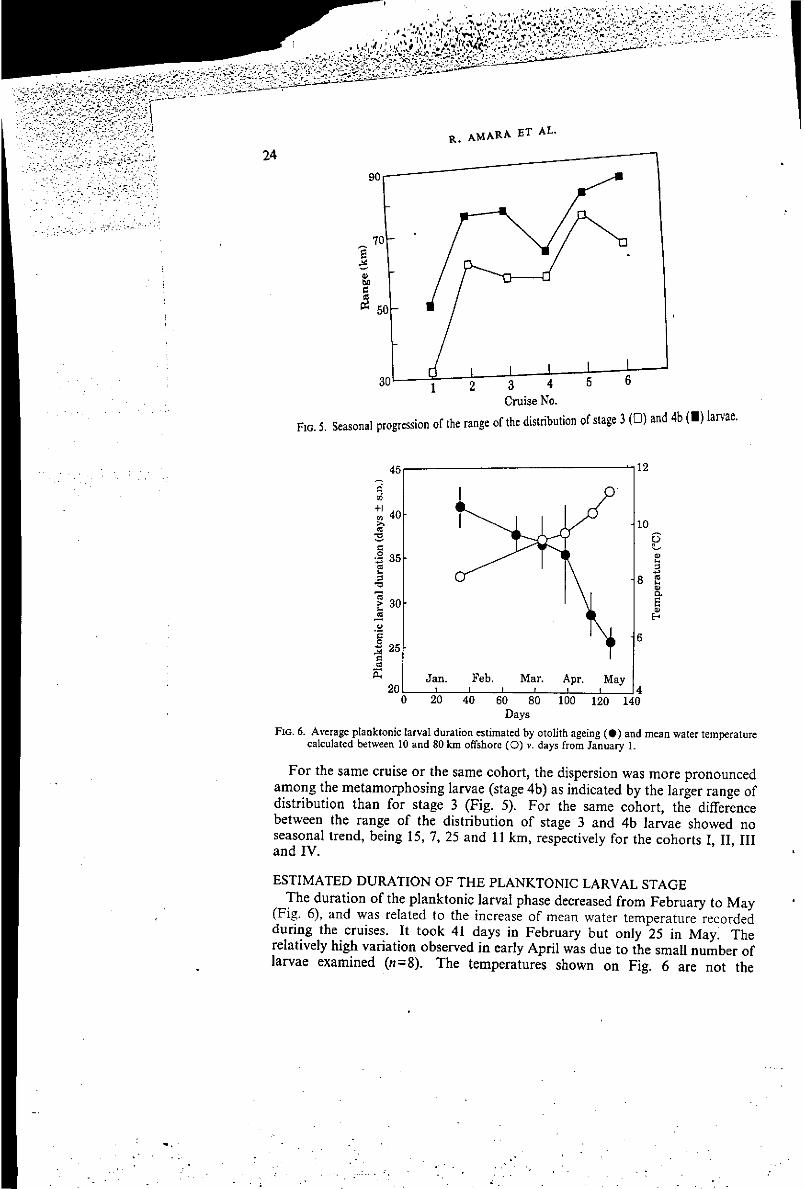
F I G , 6. Average plaaktorïc larval duration eslîmatcd by otolith ageing ( • ) and mean water température calculaied between 10 and 80 km offshore (O) v. days from January I .
For the same cruise or the same cohort, the dispersion was more pronounced among the metamorphosing larvae (stage 4b) as indicated by the larger range of distribution than for stage 3 (Fig. 5). For the same cohort, the différence between the range of the distribution of stage 3 and 4b larvae showed no seasonal trend, being 15, 7, 25 and 11 km, respectively for the cohorts I , I I , I I I and IV.
ESTIMATED DURATION OF THE PLANKTONIC LARVAL STAGE The duration of the planktonic larval phase decreased from February to May
(Fig. 6), and was related to the increase of mean water température recorded during the cruises. It took 41 days in February but only 25 in May! The relatively high variation observed in early April was due to the small number of larvae examined (rt=8). The températures shown on Fig. 6 are not the

D I S T R I B U T I O N A N D P L A N K T O N I C S T A G E D U R A T I O N O F SOLEA SOLEA 25
44
£ 36 3
24
• - a a
0 — n o
D 0 a n n a
r 0
— • • • a • • • •
- • • a c •
• - • • • • • • •
• • 001 • •
— • • • • • • • • • •
- •
1 1 1 1 1 1 100 80 60 40 20 0
Distance to coast (km) F I G . 8. Régression of larval planktonic duration on larval distribution from the coast for the cruises 3 ( • )
and S ( • ) . Sample size: cruise 3,41; cruise S, 31. Corrélations were not significant for either cruise.
températures that larvae had encountered during their development but those recorded during the cruises. Analysis of gut contents for 141 stage 4b larvae showed high différences between larvae that initiated metamorphosis early and late in the season (Fig. 7): 85% had an empty gut in February against 16 and 25% respectively in late April and early May. Copepods were the main component of their diet. During two cruises (3 and 5), there was no significant différence in the duration of the planktonic larval stage along the inshore-offshore transect, ANOVA test (cruise 3, r=0-003, n = 39, /'<0-001; cruise 5, r= -0-007, « = 30, /*<0-001) (Fig. 8). Thèse data suggest that ail stage 4b larvae caught along the transect had experienced the same environmental conditions during their development.

DISTRIBUTION AND P L A N K T O N I C STAGE DURATION OF 5 0 L E A SOLEA 25
44 -
a 3 40
e 36
1 32
28
C 24
20
•
- • • a
— 0 a c •
• n n
• n 0
— • Q C • • •
- C • • C •
— • _ • • • • a • 001
• • a a — • •
• • • • • a • •
-< 1 1
•
1 1 1
100 80 60 40 20 C Distance to coast (km)
F I G . 8. Régression of larvai planktonic duration on larval distribution from the coast for the cruises 3 ( • ) and 5 ( • ) . Sample size: cruise 3 , 4 1 ; cruise 5 , 3 1 . Corrélations were not significant for either cruise.
températures that larvae had encoumered during their development but those recorded during the cruises. Analysis of gut contents for 141 stage 4b larvae showed high différences between larvae that initiated metamorphosis early and late in the season (Fig. 7): 85% had an empty gut in February against 16 and 25% respectively in late April and early May. Copepods were the main component of their diet. During two cruises (3 and 5), there was no significant diJïerence in the duration of the planktonic larval stage along the inshore-offshore transect, ANOVA test (cruise 3, r=0-003, n=39, P<0-001; cruise 5, r = -0007 , « = 30, P<0-001) (Fig. 8). Thèse data suggest that ail stage 4b larvae caught along the transect had experienced the same environmental conditions during their development.

DISTRIBUTION AND P L A N K T O N I C STAGE DURATION OF SOLEA SOLEA 27
• The observed seasonal change in the médian of the distribution of stage 3 larvae might describe a seasonal variation in the spawning location due to the seasonal changes in the environmental conditions. However no clear trend in this pattem was found between the stage 3 larval distribution and either the température or salinity observed in the area. Koutsikopoulos & Lacroix (1992) suggested that the distribution of the spawning products of sole is not variable because ît is related to the topography of the area and especially to the nature of the sédiment.
The larval dispersai (the change in the extent and location of the médian of distribution) observed for each cohort, will resuit at first both in a net advection ' and diffusion. The différence in the extent of distribution between stage 3 and 4b (from 7 to 25 km) suggests that diffusion processes act on the larval distribution. The différence in the observed duration of planktonic life appeared to have no effect on the extent of the distribution of sole larvae. This was about 15 km for \ cohort I (March) and 11 km for cohort IV (late April-May). However, the ' validity of the onshore displacement of the médian of the distribution for cohorts I I and I I I is questionable. The relatively small number of larvae caught during cruise 4 might not represent the real distribution of the larvae, and in this way might affect the estimated displacement of the mean location of thèse two cohorts. The only clear inshore displacement of both the gênerai shape and the médian location of the distributions, was observed for cohort IV (late A p r i l -early May). However, during cruise 6 (May) ail the larvae staged were in the coastal area, suggesting a whole inshore transport of the water mass, rather than an ontogenic drift.
The assumptions made in the attempt to assess variations in horizontal distribution during larval development must be examined carefully. The sam-pling procédure is not well-adapted to study the transport process. The primary objective of the study was to investigate the fine inshore-offshore distribution of the metamorphosing larvae. A regular grid that covered ail the area, sampled during each survey, seemed appropriate for this purpose. The assumption that the sampled transect gave a real représentation of the spatial distribution of the larvae in the area, may not be valid. It is possible that alongshore movement of the spatial distribution of larvae (due to the alongshore residual circulation) occurs between cruises, and shifts the centre of mass of the distribution.
Our results show clearly a decrease of the duration of the planktonic stage as water températures increase (Fig. 6). However, high différences in stage 4b larvae with empty guts were observed between those caught early and late in the season (Fig. 7), which suggest that the availability of food had also contributed to the observed différences in the planktonic duration. Koutsikopoulos & Lacroix (1992) had observed a seasonal increase of the zooplankton biomass in the Bay of Biscay.
The observed mean location (médian) and exlent of the distributions during development suggest that both advective and diffusive processes act on the larval distribution. However, in spite of thèse observations no relation was found between the planktonic stage duration and the larval distribution. From stage 3, when sole larvae begïn vertical migration, they were distributed near the bottom in daytime and near the surface at night (Koutsikopoulos et al, 1991), Because the current speed decreased wîth depth (Koutsikopoulos et al, 1991) the degree

28 R . A M A R A E T A L .
of horizontal dispersai within the interval of time examined (10 days between stages 3 and 4b) was minhnized. In addition, because the prolonged planktonic duration increased the time of vulnerability to prédation, the constraints and/or advantages of an extended planktonic phase remained open to question. It is possible that eariy in the season, larvae suffered high mortality, due both to prédation and starvation, particularly in the inshore areas, and this masked the extent of dispersai.
In gênerai, species that recruit to estuaries from offshore areas do so at advanced developmental stages (Boehiert & Mundy, 1988). I f no transport occurs during or after the metamorphosis, as in English sole, Pawphys vetulus (Girard) (Boehiert & Mundy, 1987), and stone flounder, Kareius bicoîoraîus L . (Tsuruta, 1978), coastal recruits would be composed only of the small fraction of the pelagic population that have diffused there. However, Marchand & Masson (1989) speculated that the metamorphosing sole larvae located in the inshore areas, carry out sélective horizontal movements using bottom currents to reach the mouth of the estuary. Since 0-group juvéniles are concentrated in the estuaries and are not found offshore (Desaunay et ai, 1981; Koutsikopoulos et al, 1989, this study), according to the member/vagrant hypothesis (Sinclair, 1988), the majority of the larvae that settled offshore, would be lost to the population
Our results suggest that the planktonic duration did not affect larval dispersai.
We are grateful to the captain and the crew of the R. V. Gwen-Drez. P. Grellier, P. Bourriau, P. Beillois, D. Guérault, J. Huet, D. Dorel and S. I . Rogers helped at sea and in the laboratory. Thanks to C. Koutsikopoulos and D. Woehriing for discussion and/or critical reviews of the manuscript.
Références
Al-Maghazachi, S. J. & Gibson, R. (1984). The developmental stages of larval turbot, Scophlhalmus maximus (L . ) . Journal of Expérimental Marine Biology and Ecology 82, 35-51.
Bailey, K. M . (1981). Larval transport and recruitment of Pacific hake, Merluccius productus Marine Ecology Progress Séries 6, 1-9.
Bannister, R. C. A. (1978). Changes in plaice stock and plaice fisheries in the North Sea. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 172, 86-101.
Bergman, M. J. N., Van der Veer, H. W., Stam, A. & Zuidema, D. (1989). Transport mechanisms of larval plaice (Pîeuronectes platessa L.) from the coastal zone into the Wadden Sea nursery area. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 43-49.
Beverton, R. J. H. & Lee, A. J. (1965). Hydrographie fluctuations in the North Atlantic Océan and some biological conséquences. In The Biological Significance of Climatic Changes in Britain (Johnson, C. G. & Smith, L. P., eds). London; Académie Press.
Boehiert, G. W. & Mundy, B. C. (1987). Recruitment dynamics of metamorphosing English sole, Parophrys vetulus, to Yaquina Bay, Oregon. Estuarine Coastal and Shelf Science 25, 261-281.
Boehiert, G. W. & Mundy, B. C. (1988). Rôles of behavioral and physical factors in larval and juvénile fish recruitment to estuarine nursery areas. American Fisheries Society Symposium 3, 51-67.

DISTRIBUTION AND P L A N K T O N I C STAGE DURATION OF SOLEA SOLEA 29
Desaunay, Y., Perodou, J. B. & Beillois, P. (1981). Etude des nurseries de poissons du littoral de la Loire-Atlantique. Bulletin Institut des Pêches Maritimes 319, 1-23.
Dickson, R. R., Pope, J. G. & Holden. M. J. (1974). Environmental influence on the survival of the North Sea cod. In The Early Life History of Fish (Blaxter, J. H. S., éd.), pp. 69-80. Berlin: Springer-Verlag.
Fonds, M. (1979). Laboratory observation on the influence of température and salinity on the development of the eggs and growth of the larvae of Solea solea (Pisces). Marine Ecology Progress Séries 1, 91-99.
Haldorson, L. P. A. J., Sterritt, D. & Watts, J. (1989). Annual and seasonal variation in growth of larval walleye pollock and flathead sole in a southeastern Alaskan bay. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 220-225.
Hill, H. W. & Dickson. R. R. (1978). The largest year classes being associated with the exceptionally cold years of 1963 and 1966. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 172, 310-334.
Hjort, J. (1914). Fluctuations in the great fisheries of northem Europe viewed in the light of biological research. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 20, 1-228.
Houde, E. D. (1977). Food concentration and stocking deosity effects on survival and growth of laboratory-reared larvae of bay anchovy Anchoa mitcliilli and lined sole Achirus lineatus. Marine Biology 43, 333-341.
Houde, E. D. (1989). Comparative growth, mortality, and energetics of marine fish larvae: Température and implied latitudinal effects. Fishery Bulletin 87, 471-495.
Hovenkamp, F. (1989). Within-season variation in growth of larval plaice {Pîeuronectes platessa L.). Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 248-257.
Karakiri, M., Berghahn, R. & Westernhagen, H. W. (1989). Growth différences in 0-group plaice Pîeuronectes platessa as revealed by otolith microstructure analysis. Marine Ecology Progress Séries 49, 195-198.
Ketchen, K. S. (1956). Factors influencing the survival of lemon sole (Parophrys vetulus) in Hécate Strait, British Columbia. Journal of the Fisheries Research Board of Canada 13, 647-694.
Kim, S-, Kendall, M. & Arthur, W. Jr. (1989). Distribution and transport of larval walleye pollock (Tlieragra chakogramma) in Shelikof Strait, Gulf of Alaska, in relation to water movement. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 127-136.
Koutsikopoulos, C, Desaunay, Y., Dorel, D. & Marchand, J. (1989). The rôle of the coastal areas in the life history of sole {Solea solea L.) in the bay of Biscay. Topics in Marine Biology (Ros, J., éd.). Sciencia marina 53, 567-575.
Koutsikopoulos, C, Fortier, L. & Gagné, J. A. (1991). Cross-shelf dispersion of Dover sole eggs and larvae (Solea solea L.) in the Biscay Bay and recruitment to inshore nurseries. Journal of Plankton Research 13, 923-945.
Koutsikopoulos, C. & Lacroix, N. (1992). Distribution and abundance of sole Solea solea (L.) eggs and larvae in the Bay of Biscay between 1986 and 1989. Netherlands Journal of Sea Research 29, 81-91.
Lagardere, F. (1989). Influence of feeding conditions and température on Ihe growth rate and otohth-increment déposition of larval sole [Solea solea (L.)]. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 390-399.
Lagardere, F. & Chaumillon, G. (1988). Influence d'une première alimentation diflférce sur la croissance des larves de sole, Solea vulgaris, et l'accroissement de leur otolithes. Comptes Rendus de l'Académie des Sciences. Paris. Série 3, 306, 601-607.
Lagardere, F. & Troadec, H. (1992). L'estimation de l'âge des larves de poissons: vers une méthode objective par Tutilisation d'une technique de reconnaissance de

30 R. AMARA ET AL.
forme. In Tissus Durs et Âge Individuel des Vertébrés (Bagliniére, J. L., Castanet, J., Conand, F. & Meunier. F. J., eds). Paris: Colloques et séminaires ORSTOM-INRA: 79-91.
Laurence, C. G. (1975). Laboratory growth and metabolism of the winter flounder Pseudopleuronectes 'americanus from hatching through metamorphosis at three températures. Marine Biology 32, 223-229.
Marchand, J. & Masson, G. (1989). Process of estuarine colonisation by 0-group sole {Solea solea): hydrological conditions behaviour and feeding activity in the Vilaine estuary. Rapports et Procès- Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 191, 287-295.
Miller, J. M., Reed, J. P. & Pietrafesa, L. J. (1984). Pattems, mechanisms and approaches to the study of migration of estuarine dépendent 'fish larvae and juvéniles. In Mechanisms of Migration in Fishes (McCleave, J. D., Arnold, G. P., Dodson, J. J. & Neill, W. H., eds), pp. 209-225. New York: Plénum Press.
Minami, T. & Tanaka, M. (1992). Life history cycles in flatfish from the Northwestern Pacific, with particular référence to their early life historiés. Netherlands Journal of Sea Research 29, 35^8.
Mosegaard, H., Svedang, H. & Taberman, K. (1988). Uncoupling of somatic and otolith growth rates in Arctic charr (Salvelinus alpinus) as an effect of différences in température response. Canadian Journal of Fisheries and Aquatic Sciences 45, 1514-1524.
Nelson, W. R., Ingham, M. C. & Schaaf, W. E. (1977). Larval transport and year-class strength of Atlantic menhaden, Brevoortia tyrannus. Fishery Bulletin 75, 23-41.
Norcross, B. L. & Shaw, R. F. (1984). Oceanic and estuarine transport of fish eggs and larvae: a review. Transactions of the American Fisheries Society 113, 153-165.
Policansky, D. (1982), Influence of âge, size and température on metamorphosis in the starry flounder, Platichthys stellaîus. Canadian Journal of Fisheries and Aquatic Sciences 39, 517-517.
Postuma, K. H. (1971). The effect of température in the spawning and nursery areas on recruitment of autumn-spawning herring in the North Sea. Rapports et Proces-Verbaux des Réunions Conseil Permanent International pour l'Exploration de la Mer 160, 173-183.
Rijnsdorp, A. D., Van Beefc, F. A., Flatman, S., Millner, R. M., Riley, J. D., Giret, M. & De Clerck, R. (1992). Recruitment of sole stocks, Solea solea (L.), in Ihe northeasl Atlantic. Netherlands Journal of Sea Research 29, 173-192.
Rothschild, B. J. (1986). Dynamics of Marine Fish Populations. Cambridge, MA: Harvard University Press.
Seikai, T., Tanangonan, J. B. & Tanaka, K. (1986). Température influence on larval growth and metamorphosis of the japanese flounder, Paralichthys olivaceus in the laboratory. Bulletin of the Japanese Society of Scientific Fisheries 52, 977-982.
Sinclair, M. (1988). Marine Populations: An Essay on Population Régulation and Speciation. Seattle, WA: University of Washington Press.
Sokal, R. R. & Rohlf, J. F. (1981). Biometry. San Francisco: W. H. Freeman. Tsuruta, Y. (1978). Field observations on the immigration of larval stone flounder into
the nursery ground. Tohoku Journal of Agricultural Research 29, 10-13. Victor, B, C. (1986). Delayed metamorphosis with reduced larval growth in a coral reef
fish, Thalassoma bifasciatum. Canadian Journal of Fisheries and Aquatic Sciences 43, 1208-1213.
WeUington, G. M. & Victor, B. C. (1992). Régional différences in duration of the planktonic larval stage of reef fishes in the Eastern Pacific océan. Marine Biology 113, 491-498.
Yoklavich, M. M. & Bailey, K. M. (1990). Hatching period, growth and survival of young walleye pollock Theragra chalcogramma as determined from otolith analysis. Marine Ecology Progress Séries 64, 13-23.
ZijIstra, J. J. &. Witte, J. J. J. (1985). On the recruitment of 0-group plaice in the North Sea, Netherlands Journal of Zooîogy 35, 360-376.




















