
Seasonal and cell type specific expression of sulfate transporters in the phloem of Populus reveals...
19
Seasonal and cell type specific expression of sulfate transporters in the phloem of Populus reveals tree specific characteristics for SO 4 22 storage and mobilization Jasmin Du ¨rr • Heike Bu ¨ cking • Susanne Mult • Henning Wildhagen • Klaus Palme • Heinz Rennenberg • Franck Ditengou • Cornelia Herschbach Received: 5 August 2009 / Accepted: 2 December 2009 Ó Springer Science+Business Media B.V. 2010 Abstract The storage and mobilization of nutrients in wood and bark tissues is a typical feature of trees. Sulfur can be stored as sulfate, which is transported from source to sink tissues through the phloem. In the present study two tran- scripts encoding sulfate transporters (SULTR) were identi- fied in the phloem of grey poplar (Populus tremula 9 P. alba). Their cell-specific expression was analyzed throughout poplar in source tissues, such as mature leaves, and in sink tissues, such as the wood and bark of the stem, roots and the shoot apex. PtaSULTR1;1 mRNA was detected in companion cells of the transport phloem, in the phloem of high-order leaf veins and in fine roots. PtaSULTR3;3a mRNA was found exclusively in the transport phloem and here in both, companion cells and sieve elements. Both sulfate transporter transcripts were located in xylem parenchyma cells indicating a role for PtaSULTR1;1 and PtaSULTR3;3a in xylem unloading. Changes in mRNA abundance of these and of the sulfate transporters PtaSULTR4;1 and PtaSULTR4;2 were analyzed over an entire growing season. The expression of PtaSULTR3;3a and of the putative vacuolar efflux transporter PtaSULTR4;2 correlated negatively with the sulfate content in the bark. Furthermore, the expression pattern of both PtaSULTR3;3a and PtaSULTR4;2 correlated significantly with temperature and day length. Thus both SULTRs seem to be involved in mobilization of sulfate during spring: PtaSULTR4;2 medi- ating efflux from the vacuole and PtaSULTR3;3a mediating loading into the transport phloem. In contrast, the abundance of PtaSULTR1;1 and PtaSULTR4;1 transcripts was not affected by environmental changes throughout the whole season. The transcript abundance of all tested sulfate transporters in leaves was independent of weather condi- tions. However, PtaSULTR1;1 abundance correlated nega- tively with sulfate content in leaves, supporting its function in phloem loading. Taken together, these findings indicate a transcriptional regulation of sulfate distribution in poplar trees. Keywords Sulfate transporter Phloem loading Poplar In situ gene expression Microautoradiographics Seasonal changes Real-time RT-PCR Introduction Sulfur is one of the six macronutrients that plants require for growth and development. It is available to plants mostly in Electronic supplementary material The online version of this article (doi:10.1007/s11103-009-9587-6) contains supplementary material, which is available to authorized users. J. Du ¨rr S. Mult H. Wildhagen H. Rennenberg C. Herschbach (&) Chair of Tree Physiology, Institute of Forest Botany and Tree Physiology, Albert-Ludwigs-University Freiburg, Georges- Ko ¨hler-Allee 053/054, 79110 Freiburg, Germany e-mail: [email protected] H. Bu ¨cking Biology and Microbiology Department, South Dakota State University, Northern Plains Biostress (SNP, 251B), Brookings, SD 57007, USA K. Palme F. Ditengou Institute of Biology II, Faculty of Biology, Albert-Ludwigs- Universiy Freiburg, Scha ¨nzlestrasse 1, 79104 Freiburg, Germany K. Palme F. Ditengou Centre for Biological Signaling Studies, Albert-Ludwigs- University Freiburg, Albertstrasse 19, 79104 Freiburg, Germany K. Palme Freiburg Institute of Advanced Sciences, Albert-Ludwigs- University Freiburg, Albertstrasse 19, 79104 Freiburg, Germany 123 Plant Mol Biol DOI 10.1007/s11103-009-9587-6
-
Upload
global-studies -
Category
Documents
-
view
1 -
download
0
Transcript of Seasonal and cell type specific expression of sulfate transporters in the phloem of Populus reveals...

Seasonal and cell type specific expression of sulfate transportersin the phloem of Populus reveals tree specific characteristicsfor SO4
22 storage and mobilization
Jasmin Durr • Heike Bucking • Susanne Mult •
Henning Wildhagen • Klaus Palme • Heinz Rennenberg •
Franck Ditengou • Cornelia Herschbach
Received: 5 August 2009 / Accepted: 2 December 2009
� Springer Science+Business Media B.V. 2010
Abstract The storage and mobilization of nutrients in
wood and bark tissues is a typical feature of trees. Sulfur can
be stored as sulfate, which is transported from source to sink
tissues through the phloem. In the present study two tran-
scripts encoding sulfate transporters (SULTR) were identi-
fied in the phloem of grey poplar (Populus tremula 9
P. alba). Their cell-specific expression was analyzed
throughout poplar in source tissues, such as mature leaves,
and in sink tissues, such as the wood and bark of the stem,
roots and the shoot apex. PtaSULTR1;1 mRNA was detected
in companion cells of the transport phloem, in the phloem of
high-order leaf veins and in fine roots. PtaSULTR3;3a
mRNA was found exclusively in the transport phloem and
here in both, companion cells and sieve elements. Both
sulfate transporter transcripts were located in xylem
parenchyma cells indicating a role for PtaSULTR1;1 and
PtaSULTR3;3a in xylem unloading. Changes in mRNA
abundance of these and of the sulfate transporters
PtaSULTR4;1 and PtaSULTR4;2 were analyzed over an
entire growing season. The expression of PtaSULTR3;3a
and of the putative vacuolar efflux transporter PtaSULTR4;2
correlated negatively with the sulfate content in the bark.
Furthermore, the expression pattern of both PtaSULTR3;3a
and PtaSULTR4;2 correlated significantly with temperature
and day length. Thus both SULTRs seem to be involved in
mobilization of sulfate during spring: PtaSULTR4;2 medi-
ating efflux from the vacuole and PtaSULTR3;3a mediating
loading into the transport phloem. In contrast, the abundance
of PtaSULTR1;1 and PtaSULTR4;1 transcripts was not
affected by environmental changes throughout the whole
season. The transcript abundance of all tested sulfate
transporters in leaves was independent of weather condi-
tions. However, PtaSULTR1;1 abundance correlated nega-
tively with sulfate content in leaves, supporting its function
in phloem loading. Taken together, these findings indicate a
transcriptional regulation of sulfate distribution in poplar
trees.
Keywords Sulfate transporter � Phloem loading �Poplar � In situ gene expression �Microautoradiographics �Seasonal changes � Real-time RT-PCR
Introduction
Sulfur is one of the six macronutrients that plants require for
growth and development. It is available to plants mostly in
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11103-009-9587-6) contains supplementarymaterial, which is available to authorized users.
J. Durr � S. Mult � H. Wildhagen � H. Rennenberg �C. Herschbach (&)
Chair of Tree Physiology, Institute of Forest Botany and Tree
Physiology, Albert-Ludwigs-University Freiburg, Georges-
Kohler-Allee 053/054, 79110 Freiburg, Germany
e-mail: [email protected]
H. Bucking
Biology and Microbiology Department, South Dakota State
University, Northern Plains Biostress (SNP, 251B), Brookings,
SD 57007, USA
K. Palme � F. Ditengou
Institute of Biology II, Faculty of Biology, Albert-Ludwigs-
Universiy Freiburg, Schanzlestrasse 1, 79104 Freiburg, Germany
K. Palme � F. Ditengou
Centre for Biological Signaling Studies, Albert-Ludwigs-
University Freiburg, Albertstrasse 19, 79104 Freiburg, Germany
K. Palme
Freiburg Institute of Advanced Sciences, Albert-Ludwigs-
University Freiburg, Albertstrasse 19, 79104 Freiburg, Germany
123
Plant Mol Biol
DOI 10.1007/s11103-009-9587-6

the form of sulfate in the soil. Sulfate is taken up by the
roots and distributed within the plant by short- and long-
distance transport (Herschbach and Rennenberg 2001a, b).
Short-distance sulfate transport over membranes is medi-
ated by sulfate transporters (SULTR) (Buchner et al. 2004a).
The Arabidopsis thaliana and Oryza sativa genomes both
encode 14 putative transporters (Buchner et al. 2004a)
which cluster into five sub-groups (Hawkesford 2003).
Sulfate uptake into root cells is mediated by high affinity
transporters of group 1 (Smith et al. 1997; Vidmar et al.
1999, 2000; Yoshimoto et al. 2002; Shibagaki et al. 2002)
and has been described to operate through a sulfate/proton
co-transport mechanism (Lass and Ullrich-Eberius 1984;
Hawkesford et al. 1993). In roots, sulfate is transported out
of xylem parenchyma cells into the xylem, where it is
transported to the shoot. Some sulfate transporter sequences
of group 2 are transcribed in vascular tissues: for example
AtSULTR2;1 in the xylem parenchyma, in root pericycle
cells and in the phloem of leaves (Takahashi et al. 2000).
Recent studies have showed co-expression of AtSULTR3;5
and AtSULTR2;1 in xylem parenchyma and root pericycle
cells. It has been demonstrated that AtSULTR3;5 is
important for sulfate loading into the xylem during sulfur
starvation (Kataoka et al. 2004a), but its involvement in
sulfate efflux has not been shown.
GFP fusion proteins have demonstrated the localization
of AtSULTR4;1 and AtSULTR4;2 sulfate transporters in
the tonoplast (Kataoka et al. 2004b). Furthermore, double
knockout mutants showed that both transporters contribute
to xylem loading by releasing sulfate from the vacuole.
These transporters are, thus, involved in controlling the
storage of sulfate in the vacuole (Kataoka et al. 2004b).
Sulfate reaching the shoot via the transpiration stream can
be transported into mesophyll cells and further into plastids
for sulfate assimilation (Saito 2004), or into the vacuoles
for storage (Bell et al. 1994). Alternatively, sulfate can be
loaded into the phloem and transported back to the roots
(Smith and Lang 1988; Adiputra and Anderson 1995).
In phloem exudates of deciduous trees, sulfate is the
predominant sulfur compound (Herschbach and Rennen-
berg 2001b). The loading of sulfate into the phloem of leaf
veins and sulfate transport to trunk and root tissues has
been demonstrated in beech (Herschbach and Rennenberg
1996), oak (Schulte et al. 1998) and poplar (Hartmann et al.
2000). For example, 35S-sulfate fed to young mature and
mature poplar leaves has been detected in positions basal to
the fed leaf in wood and bark tissues and in phloem exu-
dates (Hartmann et al. 2000). Sulfate has to be taken up
into the sieve element/companion cell complex for trans-
port via the phloem by mass flow. In Arabidopsis, one
sulfate transporter AtSULTR1;3, was detected in the sieve
element/companion cell complex of cotyledons and roots
(Yoshimoto et al. 2003), but leaves from vegetative plants
were not investigated. During seedling development
AtSULTR1;3 is important for the loading of remobilized
sulfate into the phloem of the cotyledons. Although
AtSULTR2;2 is expressed in the root phloem and in the
vascular bundle sheath of leaves (Takahashi et al. 2000) its
contribution to phloem loading has not been demonstrated.
One important feature of trees is their seasonal growth
pattern. This requires storage and mobilization of reserve
metabolites including sulfate (Herschbach and Rennenberg
1996; Hartmann et al. 2000; Rennenberg et al. 2007).
During spring, sulfate is mobilized and transported into the
developing buds and leaves (Herschbach and Rennenberg
1996). Hence, sulfate has to be transported from paren-
chyma cells into the xylem and/or phloem. This can be
achieved by symplastic translocation or by sulfate release
into the apoplast followed by subsequent uptake into the
sieve element/companion cell complex.
Many studies have revealed that the abundance of sev-
eral sulfate transporter transcripts change as a response to
the sulfur demand. For example, increasing expression
levels were found under sulfur limitation when the sulfate
uptake capacity increased (Smith et al. 1997; Vidmar et al.
2000; Buchner et al. 2004b; Hopkins et al. 2004; Kataoka
et al. 2004b; Rouached et al. 2008; Koralewska et al.
2009). Moreover, reduced or increased expression was also
observed during heavy metal application (Heiss et al. 1999;
Nocito et al. 2006). Transcriptome profiling of dormant and
active cambial meristem revealed a significant reduction in
complexity of the cambial transcriptome during the dor-
mant state (Schrader et al. 2004). All these findings support
the idea that sulfate distribution within plants, particularly
seasonal changes in phloem loading and unloading, is
regulated at the transcriptional level. Therefore, the present
study aimed (1) to identify sequences encoding phloem-
specific sulfate transporters from the genome of Populus
tremula 9 P. alba; (2) to characterize the cell type-specific
expression pattern of transcripts by in situ hybridization in
source and sink tissues; and (3) to profile seasonal changes
in transcript abundance of phloem-specific, and of putative
vacuolar, sulfate transporters.
Materials and methods
Plant material
Poplars of the hybrid Populus tremula 9 Populus alba,
clone 717 1B4 (Institute National de la Recherche
Agronomique, INRA, France) were micropropagated as
described by Strohm et al. (1995) and Noctor et al. (1996).
Four-week old cuttings were transferred into a soil mixture
containing commercial soil, silica sand, and perlite (1:1:1)
and further grown in a greenhouse (26 ± 5�C) under long
Plant Mol Biol
123

day conditions (Hartmann et al. 2000). After 10 weeks, the
poplar seedlings were harvested and separated into (1) apex
with two visible leaves, (2) the 10th and 11th leaf counted
from the apex (separated into the main leaf vein including
the petiole and the leaf lamina), (3) bark and wood from the
corresponding stem section, main roots and, (4) fine roots
with a diameter up to 1 mm.
For seasonal analyses, poplar cuttings were grown from
February 2005 in the greenhouse, trimmed in autumn 2005
and transferred into the field for acclimation during winter.
The seedlings were planted in a field containing humus in
April 2006 near the institute. They were fertilized at the
beginning once with 120 g of a commercial long-time
fertilizer (Basa cote Plus 12 M, COMPO, Austria). During
drought periods in summer, trees were watered with tap
water. From the end of August 2006 until mid September
2007 leaf and bark samples were harvested between 10 am
and 11 am (sampling dates see Supplemental Table S1).
Two leaves between the seventh and ninth leaf counted
from the twig apex as well as the bark between the fifth and
15th leaf from one twig were rotationally harvested from
three of 16 trees at each sampling date. Dormant buds were
always removed from the twigs. Plant material was frozen
in liquid N2 and stored at -80�C for further analyses.
Preparation of RNA and cDNA
Tissue samples were first homogenized in liquid nitrogen.
Total RNA was then either extracted with an extraction kit
(RNeasy Plant Mini Kit; QIAGEN) according to the
instructions of the manufacturer or, alternatively from
1.2 g homogenized tissue as described by Kolosova et al.
(2004) for northern-blot analyses, or from 120 mg
homogenized tissue for the real-time PCR analyses.
For real-time PCR analyses of bark samples, genomic
DNA was digested with DNase I (Fermentas) prior RNA
transcription. In leaf samples genomic DNA was digested
using the On-Column RNAase-Free DNase Set (Qiagen)
during RNA extraction. The amount and purity of RNA were
determined with a nano-spectrophotometer (NanoDrop�
ND-1000, Peqlab). First strand cDNA was synthesized from
1 or 2 lg RNA using a final concentration of 500 nM random
primer R12 [50-NNN NNN NNN NNN-30] and Superscript II
Reverse Transcriptase (Invitrogen) according to the manu-
facturer’s instructions. No RT was used as a control reaction
and means the omission of the transcriptase enzyme.
Cloning of putative sulfate transporters
Based on in silico research of the Populus trichocarpa
genome database (http://genome.jgi-psf.org/Poptr1/Poptr1.
home.html), 18 putative sulfate transporter (SULTR)
sequences were selected and parts of their open reading
frames were cloned from the poplar hybrid Populus tremula 9
P. alba. The amplification reaction was performed using Taq-
DNA Polymerase (Promega) following the standard protocol
of the manufacturer with cDNA of different tissues. With the
exception of three sequences, all SULTR sequences were
cloned by nested PCR. Primers for PCR were designed based
on the sequence information from Populus trichocarpa
(Supplemental Table S2). The first PCR included the specific
forward 1 primer and PolyT-mix as reverse primer. The sec-
ond PCR contained 1 ll of the first PCR solution as template
and the sequence specific forward 2 and reverse 2 primers.
Amplified DNA segments were separated on a 0.8% agarose
gel, extracted with the QIAquick gel extraction kit (Qiagen),
incorporated into the pCR2.1 vector (Invitrogen) and trans-
formed into competent E. coli cells (INVF0a, Invitrogen).
Segments of the putative SULTR genes were verified by
sequencing (MWG Biotech AG, Martinsried, Germany) and
subsequent multiple sequence alignment using ClustalW
(SDSC Biology WorkBench, http://workbench.sdsc.edu/,
Thompson et al. 1994).
Characterization of putative sulfate transporters
The homology of the selected SULTR amino acid sequences
to the SULTR sequences from Arabidopsis thaliana and
Oryza sativa was determined by neighbor-joining tree
analyses (MEGA 4; Tamura et al. 2007) with gap open
penalty of 50 and gap extension penalty of 0.2 with the score
matrix BLOSUM62. Poisson correction was used as substi-
tution model. Identity and similarity between Arabidopsis or
Oryza and Populus tremula 9 P. alba genes were deter-
mined by local alignment (Tatusova and Madden 1999; http://
www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi). Full length
amino acid sequences of PtaSULTR1;1, PtaSULTR3;3a and of
the group 4 proteins PtaSULTR4;1, PtaSULTR4;2 were
aligned with the Arabidopsis AtSULTR1;1 sequence using
ClustalW2 (http://www.ebi.ac.uk/Tools/clustalw2/index.html).
A combined analysis with MEMSAT (http://saier-144-37.
ucsd.edu/memsat.html) (Jones et al. 1994) and TMAP (http://
bioinfo4.limbo.ifm.liu.se/tmap/index.html) (Persson and Argos
1994) was used to determine the locations of probable membrane
spanning domains (MSDs). Predicted transit peptide regions
were calculated using the ChloroP 1.1 (Emanuelsson et al. 1999)
program (http://www.cbs.dtu.dk/services/ChloroP/).
Northern-blot analyses
For each tissue, 15 lg of total RNA was loaded per gel slot
and separated on 1% formaldehyde-agarose gels at 100 V
for 80 min by electrophoresis. RNA was transferred onto
Hybond-XL nylon membranes (Amersham) with 109 SSC
Plant Mol Biol
123

and fixed at 80�C for 2 h. Prehybridization with
40 lg ml-1 herring sperm DNA was performed at 65�C in
a 7% SDS-FSB-buffer (FSB: 0.05 M NaP2O7, 0.115 M
NaH2PO4, 0.5 M EDTA). Specific cDNA probes between
150 and 360 bp long including 30UTR parts were prepared
for each isoform. Primers and the lengths of the fragments
are given in Supplemental Table S3. Hybridization with32P-labelled cDNA probes containing 32P-dATP insertion
was performed in 7% SDS-FSB-buffer over night using the
Strip-EZTM DNA kit (Ambion). Northern blots treated with
5.8S rRNA (Acc. No. AY781281) probe were used as
loading controls. After hybridization the membranes were
washed three times under high stringency conditions with
1% SDS-FSB-buffer and examined by a Phosphor Imager
(Molecular Imager FX, Bio-Rad Laboratories).
In situ hybridization
Approximately 3 mm sections of different tissues were
harvested from 10-week old Populus tremula 9 P. alba
trees. These were: (1) the apex, (2) the 11th leaf counted
from the apex, (3) young stem parts with young developing
leaves (fourth and fifth leaf counted from the apex), stems
with young mature leaves (tenth and 11th leaf) and older
stem parts with mature leaves (25th and 27th leaf), (4) the
main root and, (5) fine roots. Tissues were fixed in 4%
paraformaldehyde phosphate buffer (PBS: 140 mM NaCl,
10 mM KCl, 6.4 mM Na2HPO4, 2 mM NaH2PO4, pH 7)
containing 0.01% Triton x-100 overnight at 4�C after
vacuum infiltration according to Cnops et al. (2006).
Samples were dehydrated in a graded ethanol series and
subsequently infiltrated in a Roticlear (Carl Roth) series
described by Deeken et al. (2008). The samples were
embedded in paraplast (Sigma–Aldrich) at 60�C by
changing the solution four times and subsequent vacuum
infiltration with a self-made infiltration device according to
Deeken et al. (2008). Eight to 10 lm sections were cut with
a RM2245 microtome (Leica) and were transferred onto
charged glass slides (SuperFrost� Plus, Menzel, Germany).
Paraplast was removed with a series of HistoClear and
ethanol (Deeken et al. 2008). The proteins were digested by
treating the sections with 125 lg ml-1 Pronase (Sigma–
Aldrich) in PBS-buffer for 10 min at 37�C. Subsequently,
the samples were post-fixed with 4% paraformaldehyde
in PBS.
Gene specific probes were generated by PCR for both
PtaSULTR1;1 and PtaSULTR3;3a (primers and probe length
see Supplemental Table S3). The forward primer carried
the T3 [50-AATTAACCCTCACTAAAGGGAGA-30] and
the reverse primer the T7 RNA polymerase binding site
[50-GCTTCTAATACGACTCACTATAGGGAGA-30] at
the 50 end. Labeling with digoxigenin (DIG) was performed
by in vitro transcription using DIG RNA labeling mix
(Roche Diagnostics) with either T3 or T7 RNA polymerase
(Fermentas) according to the instruction of the manufac-
turer. Labeled probes were dissolved in 100 ll water and the
quality was tested by electrophoresis on a 1%-agarose-TBE-
buffer gel. The RNA concentration was determined spec-
trophotometrically at 260 nm.
Each slide was incubated for 1 h at 50�C in a humid
chamber with 100 ll prehybridization buffer [50% form-
amide and 29 SSC buffer (209 SSC was 3 M NaCl, 0.3 M
sodium citrate) and 40 U ml-1 RNase inhibitor (Fermen-
tas)]. Hybridization with equal concentrations (20–30
lg ml-1) of antisense or sense probe was performed on
slides with 100 ll hybridization buffer [50% formamide,
49 SSC buffer, 109 Denhardt’s solution, 0.5 mg/ml yeast
t-RNA, 10% dextran sulfate and 40 U ml-1 RNase inhib-
itor (Fermentas) in water] overnight at 50�C in a humid
chamber. The following wash steps were performed in
glass boxes at 50�C each for 30 min: 39 SSC, 39 SSC,
1.59 SSC, 0.759 SSC and two further wash steps with
0.39 SSC and 0.19 SSC for 20 min each. After the second
wash step, the sections were treated with RNase (RNase A
20 lg ml-1, Carl Roth) for 20 min at 37�C in TE-buffer
(pH 8.0). Non-specific binding sites were blocked with
BSA (bovine serum albumin) in blocking buffer (100 mM
Tris–HCl, pH 7.5, 150 mM NaCl, 0.5% BSA, 0.02%
Tween 20) for 1 h at room temperature. Incubation with
anti-DIG-UTP-alkaline phosphatase FAB fragments
(1:2,000, Roche Diagnostics) and staining with nitroblue
tetrazolium chloride and 5-bromo-4-chloro-3-indolyl
phosphate (Roche Diagnostics) was carried out according
to Cnops et al. (2006). The slides were mounted in 50%
glycerol and inspected with an inverted microscope
(Axiovert 200 M and Stemi SV 11 Apo, Zeiss). Images
were taken with a digital camera (Zeiss) and processed
with AxioVision 4.6 software (Zeiss).
Microautoradiographic studies
Mature leaves from 8 to 10-week old poplar plants were
fed with 150 lCi carrier free [35S]-SO42- via flap-feeding
(Biddulph 1956) as previously described (Hartmann et al.
2000) to allow a direct penetration of the feeding solution
into the phloem. The stem sections below the fed leaf were
harvested after 24 h. The stem sections were cryofixed by
plunging into melting nitrogen and freeze-dried under low
temperature and high vacuum conditions (CFD, Leica,
Bensheim, Germany). To prevent a re-crystallization of
water in the plant tissues (*-80�C), the samples were first
freeze-dried for 7 days at -100�C, and then for 2 days
each at -90, -80, and -60�C and then slowly warmed up
to room temperature. The samples were then pressure infil-
trated according to the method described by Fritz (1980) in
an epoxy resin (Spurr 1969) using dried diethyl ether as an
Plant Mol Biol
123

inter-medium. After polymerization, dry sections of 1 lm
were cut with Teflon-coated glass knifes and mounted on
filmed microscope slides. The sections were overlaid in the
dark room with a thin layer of the nuclear research emulsion
L4 (Ilford, Dreieich, Germany) with a crystal size of
0.11 lm. The thickness of the film layer was approximately
5 lm. After exposure in the darkness at 4�C for several
weeks (dependent on the radioactivity of the samples), the
film was developed with the fine grain developer D19 A/S
(Sanderson 1981), rinsed in water and than fixed with a
commercial b/w fixer (Tetanol, Norderstedt, Germany). As
controls, slides without sections were overlaid with film and
processed in the same way as described above. The micro-
graphs were analyzed using Nomarski DIC microscopy
(Olympus, Hamburg, Germany).
Quantitative real-time RT PCR
Quantitative PCR measurements of PtaSULTR1;1,
PtaSULTR3;3a, of the putative vacuole localized sulfate
transporters PtaSULTR4;1, PtaSULTR4;2 and of the refer-
ence gene, the elongation factor 1-beta (EF1beta; Acc.
No.: FJ372570), were performed using the following gene-
specific forward and reversed primers: (50-TTTATAACCC
GTGCAGATAAGGAC-3) and (50-CCTTTTAGCAAATG
GTCACCAC-30) for PtaSULTR1;1, (50-GCCCCTCTTGT
GTCTGTGATC-30) and (50-TCCACGAGGGAGGATT-
TAGC-30) for PtaSULTR3;3a, (50-GGCACTGCGTATA-
TATGATATCTGTC-30) and (50-AAACCTTACGACAAG
TATTGCATTG-30) for PtaSULTR4;1, (50-GAGGCAGGG
CGTAGATTG-30) and (50-GGAAGCAAGCCTTACAAT
GC-30) for PtaSULTR4;2 and, (50-TGAGGATCTCTGGT
GTCGAAG-30) and (50-GTCTCAGCAGATGGAGGAGT
G-30) for EF1beta. The resulting PCR fragments were
113 bp (PtaSULTR1;1), 115 bp (PtaSULTR3;3a), 114 bp
(PtaSULTR4;1), 132 bp (PtaSULTR4;2) and 100 bp
(EF1beta). The specificity of the primer pairs was tested by
RT-PCR and amplified segments were controlled by
sequencing after purification from agarose gels. mRNA
abundance in leaf and bark tissues was determined by real
time RT PCR using the LightCycler 480 SYBR Green I
Master mix (Roche, Germany) and the Multiwell Plate 384
(Roche, Germany). The 10 ll reaction volume contained
5 ll 29 SYBR Green I Master mix, 2.5 ll (25 ng) cDNA
and a final primer concentration of 0.8 or 1.2 lM. After a
hot start for 2 min at 95�C, 45 PCR cycles were performed
with a 15 s melting step at 95�C, 15 s annealing time at
55�C, and 15 s extension time at 72�C on a LightCycler�
480 system (Roche, Germany). Transcripts per lg RNA
were calculated based on standard curves generated with
serial dilutions of linearized plasmids containing an insert
of the gene of interest.
Determination of sulfate contents
Sulfate concentration was determined by anion exchange
chromatography from 50 mg powdered bark and leaf tissue
as described by Herschbach et al. (2000).
Statistical analyses
Bivariate data correlations of the seasonal sulfate trans-
porter expression were performed with transformed (ln)
values of the transcript means and the sulfate means from
particular sampling dates using the Pearson linear corre-
lation function. The data transformation resulted in nor-
mality and homogeneity of variances of data. The weather
data (maximum, minimum and mean temperature, relative
humidity and sun shine duration) were available from a
climate station in close vicinity (Station 10803 of the
Deutscher Wetterdienst, Freiburg Airport, Germany).
Meteorological data were mean values from 24 h prior to
the sampling time. Statistical analyses were performed
using the statistical software package (SPSS� GmbH
software, version 16, Munich, Germany). Significant dif-
ferences in transcript abundances and sulfate contents
among sampling dates were tested by analysis of variance
(ANOVA) using a significance level of P \ 0.05. Post-hoc
tests (Turkey test and Games–Howell-test) were used to
determine significant differences between adjacent sam-
pling dates.
Accession numbers
Sequences can be found in the NCBI database (http://
www.ncbi.nlm.nih.gov) under the following accession
numbers for Populus tremula 9 P. alba: DQ906929
(PtaSULTR1;1), DQ174472 (PtaSULTR1;2), DQ906931
(PtaSULTR2;1a), DQ906933 (PtaSULTR2;1b), DQ174473
(PtaSULTR2;2), DQ174470 (PtaSULTR3;1a), DQ906928
(PtaSULTR3;1b), DQ174469 (PtaSULTR3;2a), DQ906934
(PtaSULTR3;2b), DQ906924 (PtaSULTR3;3a), DQ906926
(PtaSULTR3;3b), DQ174467 (PtaSULTR3;4a), DQ174466
(PtaSULTR3;4b), DQ906927 (PtaSULTR3;5), DQ906930
(PtaSULTR4;1), DQ906935 (PtaSULTR4;2), DQ174477
(PtaSULTR5;2), DQ174475 (PtaSULTR5;1), AY781281
(5.8 S rRNA) and for Arabidopsis: AB018695 (AtSULTR1;1),
AB042322 (AtSULTR1;2), AB049624 (AtSULTR1;3),
AB003591 (AtSULTR2;1), D85416 (AtSULTR2;2),
D89631 (AtSULTR3;1), AB004060 (AtSULTR3;2),
AB023423 (AtSULTR3;3), AB054645 (AtSULTR3;4),
AB061739 (AtSULTR3;5), AB008782 (AtSULTR4;1),
AB052775 (AtSULTR4;2), NP_178147 (AtSULTR5;1),
NP_180139 (AtSULTR5;2) and for Oryza: AF493790
(OsSULTR1;1), XP_470587 (OsSULTR1;2), AF493790
(OsSULTR1;3), AAN59769 (OsSULTR2;1), AAN59770
Plant Mol Biol
123

(OsSULTR2;2), NP_921514 (OsSULTR3;1), AAN06871
(OsSULTR3;2), AK104831 (OsSULTR3;3), BAD68396
(OsSULTR3;4), NM_192602 (OsSULTR3;5), NM_191791
(OsSULTR3;6), AF493793 (OsSULTR4;1), BAC05530
(OsSULTR5;1), BAD03554 (OsSULTR5;2).
Results
Identification and expression analyses of SULTR genes
Phylogenetic analysis of the 18 putative SULTR (sulfate
transporter) gene sequences from Populus tremula 9
P. alba revealed that they can be classified into the five sub-
groups proposed by Hawkesford (2003) (Fig. 1a). Sequence
similarity to the homologous Arabidopsis sequences was
between 80 and 90% (Supplemental Table S4). All putative
SULTR sequences showed a tissue-specific expression pat-
tern (Supplemental Fig. S1). Two sulfate transporter tran-
scripts, PtaSULTR1;1 of group 1 and PtaSULTR3;3a of
group 3, were abundant in the bark of the stem, in the main
root and in leaf veins. Small amounts of PtaSULTR1;1
mRNA were also found in the stem wood (Fig. 1c). As the
bark includes the phloem we further focused on these two
transporters. We included the two sequences of group 4,
PtaSULTR4;1 and PtaSULTR4;2 in our analyses as members
of group 4 are putative vacuolar sulfate transporters. Both;
PtaSULTR4;1 and PtaSULTR4;2, were expressed ubiqui-
tously throughout all tissues (Fig. 1c). A combined analysis
with MEMSAT and TMAP showed that the open reading
frames of PtaSULTR1;1, PtaSULTR3;3a, PtaSULTR4;1,
and PtaSULTR4;2 each encode 12 predicted membrane
spanning domains (MSDs) (Fig. 1b). At the carboxyl-ter-
minus, a conserved STAS (sulfate transporter anti sigma
factor antagonist) domain was identified in the PtaSULTR
sequences from groups 1, 2, 3, and 4 by comparing these
sequences with the Arabidopsis SULTR1;1 sequence pre-
viously examined by Shibagaki and Grossman (2004)
(Fig. 1b).
Cell type-specific expression of PtaSULTR1;1
and PtaSULTR3;3a analyzed by in situ hybridization
As the phloem connects mature leaves with different sink
tissues such as young developing leaves, roots and storage
tissues of the trunk, we used in situ hybridization to detect
the expression of both transporters (1) in transverse sec-
tions of leaves and leaf veins, (2) in longitudinal sections of
the apex, (3) in transverse sections of stems at different
developmental stages, of roots with secondary growth, and
(4) in fine roots. The sections were analyzed with antisense
probes. Sense probes used as controls showed no blue
staining (Figs. 2a, g, 3a, g, 4a, 5a, 6a, c). Brown spots
visible in the apex (developing leaves) and in roots repre-
sent the staining of phenolic compounds (green arrows in
Figs. 3e, 6c, d). Thus the staining observed with the anti-
sense probes clearly shows the mRNA abundance of
PtaSULTR1;1 or PtaSULTR3;3a.
Transverse sections of leaves and leaf veins
PtaSULTR1;1 expression was found in major leaf veins,
especially in companion cells of the phloem and in xylem
parenchyma cells (Fig. 2c, d). In minor leaf veins, the entire
phloem was labeled (Fig. 2e, f). Palisade parenchyma,
spongy mesophyll cells, and upper epidermis cells
were weakly stained (Fig. 2e, f). The expression of
PtaSULTR3;3a differed from that of PtaSULTR1;1 (Fig. 2h–m).
The phloem and xylem parenchyma cells of major leaf veins
showed PtaSULTR3;3a expression. Both, sieve elements and
companion cells were labeled with comparable intensity
(Fig. 2i). PtaSULTR3;3a transcripts were not detectable in
minor veins and also not in other leaf cells (Fig. 2l, m).
Longitudinal sections of the apex
Both PtaSULTR1;1 and PtaSULTR3;3a transcripts were
detected in the shoot apex but with clear differences in their
expression patterns (Fig. 3). Whereas a high expression
of PtaSULTR1;1 was found in leaf primordia (Fig. 3b, c) and
in meristem cells of the leaf margin (LMM; Fig. 3d, e),
PtaSULTR3;3a was only weakly detected in the shoot apical
meristem (SAM, Fig. 3h). Both genes were expressed in the
provascular strands (Fig. 3b, h).
Transverse sections of stems
Stem sections with mature leaves (tenth to 11th) (Fig. 4)
showed PtaSULTR1;1 expression in several cell types. Most
significant is the expression in companion cells of the
phloem (Fig. 4e, f) beside a strong mRNA abundance in ray
initials of the vascular cambium and, to a lower extent, in
fusiform cells (Fig. 4b, c, e). PtaSULTR3;3a was expressed
in companion cells but, staining with a comparable intensity
was also found in sieve elements (Fig. 5e, f). Irrespective of
the stem age PtaSULTR3;3a transcripts were not detected in
the vascular cambium (Fig. 5e, Supplemental Fig. S3).
Both, PtaSULTR1;1 and PtaSULTR3;3a were expressed
in developing ray pith cells (Figs. 4c, e, 5c, e), in ray cells
connecting xylem vessels in the mature wood (Figs. 4e, g,
5e, g) and in parenchyma cells of the primary xylem
(Figs. 4h, 5h).
Furthermore, PtaSULTR1;1 transcripts were visible in
cells just below the epidermis (Fig. 4d) while transverse
sections of young trunks (fourth and fifth leaf counted from
the apex) revealed strong PtaSULTR1;1 expression in the
Plant Mol Biol
123

(C)
(A) (B)
PtaSULTR1;1
PtaSULTR3;3a
5.8S rRNA
5.8S rRNAPtaSULTR4;2
PtaSULTR4;1
5.8S rRNA
5.8S rRNA
PtaSULTR1.1
PtaSULTR1.2
AtSULTR1.2
AtSULTR1.3
AtSULTR1.1
OsSULTR1.3
OsSULTR1.1
OsSULTR1.2
PtaSULTR2.1a
PtaSULTR2.1b
AtSULTR2.1
PtaSULTR2.2
AtSULTR2.2
OsSULTR2.1
OsSULTR2.2
PtaSULTR3.1a
PtaSULTR3.1b
PtaSULTR3.2a
PtaSULTR3.2b
AtSULTR3.1
AtSULTR3.2
OsSULTR3.1
OsSULTR3.2
PtaSULTR3.5
AtSULTR3.5
OsSULTR3.5
OsSULTR3.6
PtaSULTR3.3a
PtaSULTR3.3b
AtSULTR3.3
PtaSULTR3.4a
PtaSULTR3.4b
AtSULTR3.4
OsSULTR3.4
PtaSULTR4.1
PtaSULTR4.2
AtSULTR4.1
AtSULTR4.2
OsSULTR4.1
PtaSULTR5.1
PtaSULTR5.2
AtSULTR5.2
OsSULTR5.2
AtSULTR5.1
OsSULTR5.1
94
69
98
95
100
100
99
97
100
100
100
100
100
100
100
100
100
100
100
98
88
96
100
100
98
64
42
100
98
100
97
93
97
98
92
82
100
89
100
98
53
98
0.2 substitutions per site
Fig. 1 Characteristics of the sulfate transporter gene family from
Populus tremula 9 P. alba. a Phylogenetic analysis of predicted
sulfate transporter amino acid sequences from Populus tremula 9
P. alba. The bootstrap values, expressed as percentage, were obtained
from 1,000 replicate trees. b Full length amino acid sequences of
PtaSULTR1;1, PtaSULTR3;3a, PtaSULTR4;1, and PtaSULTR4;2
were aligned with the Arabidopsis AtSULTR1;1 sequence. Predicted
12 membrane-spanning domains (MSDs) were marked with blackbars. Consensus amino acids are highlighted in yellow while those
with similar properties are highlighted in grey. A conserved basic
amino acid residue (Arg) between MSD 9 and 10 is indicated by a
solid circle. The conserved STAS (sulfate transporter anti sigma
factor antagonist) domain is indicated by a red box. Predicted transit
peptide regions are marked with blue bold letters. c Northern blots
from PtaSULTR sequences. A 15 lg total RNA was extracted from
different tissues from 10-week old poplars as indicated: the apex, the
10th and 11th leaf counted from the apex separated into major leaf
vein including the petiole and the remaining leaf lamina, the
corresponding stem section separated into bark and wood, the main
root separated into bark, wood, and fine roots. Northern blots
were hybridized with isoform-specific, [32P] labeled probes for
PtaSULTR1;1, PtaSULTR3;3a, PtaSULTR4;1, and PtaSULTR4;2sequences. 5.8 S rRNA probe was used as a loading control. Data
presented are from pooled samples from three independent plants
from one of two experiments with comparable results
Plant Mol Biol
123

epidermis and the cells around lenticels (Supplemental
Figs. S2, S4). The cortex cells that are located between
sclerenchymatous fibers and the epidermis showed weak
PtaSULTR1;1 expression (Fig. 4b) while only periderm
cells revealed a slight PtaSULTR3;3a staining (Fig. 5c, d).
The expression was also analyzed along the entire stem
to identify age dependent differences. But, the expression
patterns observed in stem sections bearing young mature
leaves (tenth to 11th counted from the apex) was repre-
sentative for the entire stem (Supplemental Figs. S2, S3).
However, whereas PtaSULTR1;1 mRNA was expressed
in roots with secondary growth (Supplemental Fig. S2),
PtaSULTR3;3a was not (data not shown).
Fine roots
PtaSULTR3;3a transcripts were not visible in fine roots
(Supplemental Fig. S5) but PtaSULTR1;1 mRNA was here
expressed in the phloem (Fig. 6d, f). This was also found
when lateral root appear (Fig. 6e). The phloem was stained
in diarch, i.e., near the root tip, as well as in tetrarch
vascular root bundles (Fig. 6f). In longitudinal sections,
PtaSULTR1;1 transcripts were detected in the stele and in
the root meristem of the emerging lateral root (Fig. 6g, h).
Microautoradiographic 35S localization
To test whether transcript abundance of SULTR genes
correlates with sulfate transport activity, a mature leaf was
fed with 35S-sulfate. The radioactivity was examined at the
cellular level by microautoradiographic analysis in stem
sections basal to the fed leaf. 35S distribution is visualized
as silver grains in the photographs. Accumulated 35S was
found in ray initials of the vascular cambium (Fig. 7c),
companion cells of the phloem (Fig. 7b) and pith ray cells
of wood (Fig. 7d). These results clearly demonstrate the
transport of 35S from source leaves to more basal regions of
the stem, and thus the unloading of sulfate from the
phloem, and further transport into ray pith cells. The latter
is indicated by an enrichment of silver grains in pith ray
cells (Fig. 7d).
Seasonal changes in sulfate content and of SULTR
expression in bark
The seasonal expression of PtaSULTR1;1, PtaSULTR3;3a,
and the putative vacuolar sulfate transporters PtaSULTR4;1
and PtaSULTR4;2 was analyzed in leaves and bark
throughout an annual growth cycle (Fig. 8). An elongation
Fig. 2 In situ localization of PtaSULTR1;1 and PtaSULTR3;3a in
leaves. Transverse sections of major leaf veins (a–d, g–k) and of leaf
lamina with minor leaf veins (e, f and l, m) were hybridized with the
sense (a) or antisense (b–f) probe for PtaSULTR1;1 as well as with the
sense (g) or antisense probe (h–m) for PtaSULTR3;3a. Enlarged parts
are indicated by squares. Bars are 200 lm for a, b, g and h and
20 lm for all other. Abbreviations: cc companion cell, le lower
epidermis, p phloem, pp palisade parenchyma, px protoxylem,
se sieve element, sm spongy mesophyll, ue upper epidermis, x xylem,
xp xylem parenchyma, xv xylem vessel
Plant Mol Biol
123

factor gene family sequence (elongation factor 1-beta;
EF1beta) was used as a reference gene (Brunner et al.
2004; Nicot et al. 2005). In bark, the expression of EF1beta
in autumn and spring was twice as abundant as during
dormancy (Fig. 8f). The expression of PtaSULTR1;1 in the
bark decreased in autumn when tree growth switched to
dormancy (mean temperatures below 5�C) (Fig. 8b). Dur-
ing bud break the expression of PtaSULTR1;1 increased
significantly: twofold week-on-week between 3rd April
and 18th April 2007 (P \ 0.05; 3rd April \ 11th
April \ 18th April). At the same time, the sulfate that had
accumulated in the bark during autumn 2006 disappeared
(Fig. 8a). The expression level of PtaSULTR3;3a was
comparable to that of PtaSULTR1;1 during autumn 2006
and spring 2007 and increased significantly during bud
swelling (P \ 0.05; 12th March \ 3rd April, P \ 0.068;
3rd April \ 18th April) (Fig. 8c). During dormancy,
PtaSULTR3;3a mRNA was not detected but the transcript
level increased again at the end of March and remained
high throughout leaf expansion and early leaf development.
The expression of PtaSULTR4;2 decreased during autumn
but, in contrast to PtaSULTR3;3a, low expression levels
were still detected during dormancy (Fig. 8c, e). In spring,
mRNA abundance of PtaSULTR4;2 increased, starting
shortly before bud break (P \ 0.05; 12th March \ 27th
March). PtaSULTR4;1 expression increased significantly
during winter with a maximum on 21st December
(P \ 0.05; 25th October \ 22nd November \ 21st
December [ 22nd January). Subsequently, it remained
constant until bud break, which occurred between 3rd April
and 11th April (Fig. 8d). The expression of all genes tested
was high in the bark of young twigs developed in 2007 and
declined thereafter.
Bivariate Pearson-correlation analyses were used to
calculate the correlation between the expression of differ-
ent sulfate transporters, the sulfate content and meteoro-
logical parameters (maximum, minimum and mean
temperature, day length, relative humidity, wind, rain, and
hours of sunshine). PtaSULTR3;3a and PtaSULTR4;2
expression was positively correlated in bark tissue. Both
genes were negatively correlated to the sulfate content
(Table 1). PtaSULTR3;3a and PtaSULTR4;2 were posi-
tively and the sulfate content negatively correlated to mean
temperature, maximum temperature, minimum temperature
and day lengths (Table 1, see also Supplemental Fig. S6).
The expression of PtaSULTR1;1 and PtaSULTR4;1 corre-
lated positively, but neither showed any correlation to
meteorological parameters.
Fig. 3 In situ localization of PtaSULTR1;1 and PtaSULTR3;3a in the
shoot apex. Longitudinal sections from the shoot apex were hybrid-
ized with the sense (a) or antisense (b–e) probe for PtaSULTR1;1 and,
with the sense (g) or antisense (h) probe for PtaSULTR3;3a. a, b, g,
and h longitudinal sections from the apex; c shoot apical meristem
(SAM); d a lateral bud with leaf primordium; e leaf margin meristem
(LMM). A green arrow in e indicates phenolic compounds. Bars are
200 lm for a, b, g, and h and 50 lm for c, d, and e. Abbreviations:
lp leaf primordium, ps provascular strand
Plant Mol Biol
123

Seasonal changes in sulfate content and of SULTR
expression in leaves
In leaves, the average expression of EF1beta was tenfold
higher than in bark. One peak of EF1beta transcript abun-
dance was detected just after bud break when leaves started
to expand (Fig. 8f). In comparison, the mean expression
level of PtaSULTR1;1 was 100-fold lower, but was compa-
rable in bark and leaves (Fig. 8b). A peak value of
PtaSULTR1;1 and PtaSULTR4;1 mRNA was observed in
autumn 2006 before leaf fall. During leaf expansion, i.e.,
after bud break in spring 2007, expression of PtaSULTR1;1
and PtaSULTR4;2 increased until the leaves were fully
expanded (in May) and decreased thereafter (P \ 0.05; 18th
April \ 18th May [ 14th August, 28th August, 13th Sep-
tember). In contrast, mRNA abundance of PtaSULTR3;3a,
was low during early leaf expansion, increased in June 2007
when the leaves were fully developed and decreased later in
the season (P \ 0.05; 18th May \ 29th June [ 13th Sep-
tember). The expression of PtaSULTR4;1 was constant in
spring and early summer but decreased in late summer. The
sulfate content increased in the developing leaves until late
summer 2007 to a comparable amount to that measured in the
previous year (Aug 2006) (Fig. 8a). Sulfate decreased during
late summer in the leaves developed in 2006 and remained
low during autumn.
The bivariate analyses showed a linear negative corre-
lation between the transcript levels of PtaSULTR1;1 and
the sulfate content in the leaf tissue (Table 2). The
expression levels of PtaSULTR1;1, PtaSULTR4;1, and
PtaSULTR4;2 were positively correlated (Table 2) but only
mRNA values of PtaSULTR4;2 correlated negatively to
maximum temperature.
Discussion
Characteristics of SULTR sequences
In the present study, the sulfate transporter-encoding gene
family of Populus tremula 9 P. alba was investigated. The
overall topology of the SULTR sequences PtaSULTR1;1,
PtaSULTR3;3a, PtaSULTR4;1, and PtaSULTR4;2 fits with
the proposed 12 membrane spanning domain (MSD) model
(Smith et al. 1995; Hawkesford 2003). One Arg (R-391/390/
363/361) residue located between MSD 9 and 10, is con-
served among all sulfate transporter sequences from poplar
(this study), Arabidopsis thaliana, Stylosanthes hamata, and
Hordeum vulgare (Takahashi et al. 1997). As previously
proposed after work in other species this residue may be
involved in the binding of sulfate (Smith et al. 1995). In
Arabidopsis, the STAS domain, which extends into the
Fig. 4 In situ localization of
PtaSULTR1;1 in stem sections
where the 10th to 11th leaves
(counted from the apex) are
attached. Transverse stem
sections were hybridized with
the sense (a) or antisense (b–h)
probe for PtaSULTR1;1. a, bOverview of the transverse
section; c bark and cambial
zone; d epidermis and periderm
of the bark; e cambium and
phloem; f phloem; g primary
wood with pith rays; h primary
xylem at the pith. Enlarged parts
are indicated by squares. Barsare 200 lm for a–c and e and
20 lm for d, f–h.
Abbreviations: br phloem ray
cells, c cambium, cc companion
cell, co cortex, cu cuticle,
ep epidermis, f fusiform cell,
p phloem, ph periderm, pi pith,
pr pith ray cells, px primary
xylem, ri ray initials, se sieve
element, sf sclerenchymatous
fibres, sp sieve plate, x xylem,
xp xylem parenchyma, xv xylem
vessel
Plant Mol Biol
123
cytoplasm (Aravind and Koonin 2000), is necessary for the
correct localization of SULTR proteins within the mem-
brane; it also determines the kinetic characteristics of sulfate
transport (Rouached et al. 2005; Shibagaki and Grossman
2006). All putative poplar sulfate transporters of group 1, 2,
3, and 4 contain this STAS domain and are illustrated
for PtaSULTR1;1, PtaSULTR3;3a, PtaSULTR4;1, and
PtaSULTR4;2 (Fig. 1b) which exhibited significant identi-
ties (E \ 0.01 Pearson 2000) with genes encoding sulfate
transporters in Arabidopsis thaliana (http://blast.ncbi.nlm.
nih.gov/Blast.cgi). The poplar sequence PtaSULTR1;1
showed highest homology to the Arabidopsis sequence
AtSULTR1;3 of group 1, but a lower identity to the
AtSULTR1;1 and AtSULTR1;2 sequences. AtSULTR1;3 is
specifically expressed in the phloem of cotyledons and has
been functionally characterized (Yoshimoto et al. 2003).
PtaSULTR4;1 and PtaSULTR4;2; amino acid sequences
revealed highest identity with, and shared many character-
istics with, AtSULTR4;1 including a transit peptide
sequence (Fig. 1b). As AtSULTR4;1 is located in the tono-
plast and generates sulfate efflux (Kataoka et al. 2004b) it is
assumed that both SULTRs of group 4 from poplar also
functions as a sulfate efflux transporter in the tonoplast. The
two poplar sequences with highest similarity to Arabidopsis
AtSULTR5;2 lack characteristics of sulfate transporters such
as the STAS domain (Aravind and Koonin 2000). This is not
surprising since recently AtSULTR5;2 was identified as a
molybdate transporter (Tomatsu et al. 2007).
In terms of genome evolution, analyses of Arabidopsis
thaliana and poplar ESTs have revealed a whole genome
duplication event in poplar (salicoid duplication) (Tuskan
et al. 2006). Thus, a single-copy gene in Arabidopsis tha-
liana is typically represented by two copies in Populus
species (Jansson and Douglas 2007). This could be the
reason for the observation that two highly similar
PtaSULTR sequences (a and b; Fig. 1) showed significant
identity with only one sulfate transporter in Arabidopsis
thaliana. The possibility that closely related genes are
derived from the two combined genomes of the used hybrid
Populus tremula 9 P. alba can be exclude because corre-
sponding sequences are also represented in the Populus
trichocarpa genome.
Expression of sulfate transporters relevant for sulfate
loading into the phloem of leaves
Sulfate transporters are dependent on the proton motive
force and function as proton-symport carrier (Lass and
Ullrich-Eberius 1984; Hawkesford et al. 1993; Kataoka
et al. 2004b). During active growth, sulfate reaches the
leaves via xylem transport. Therefore, the expression of
PtaSULTR1;1 in leaf mesophyll cells may indicate that
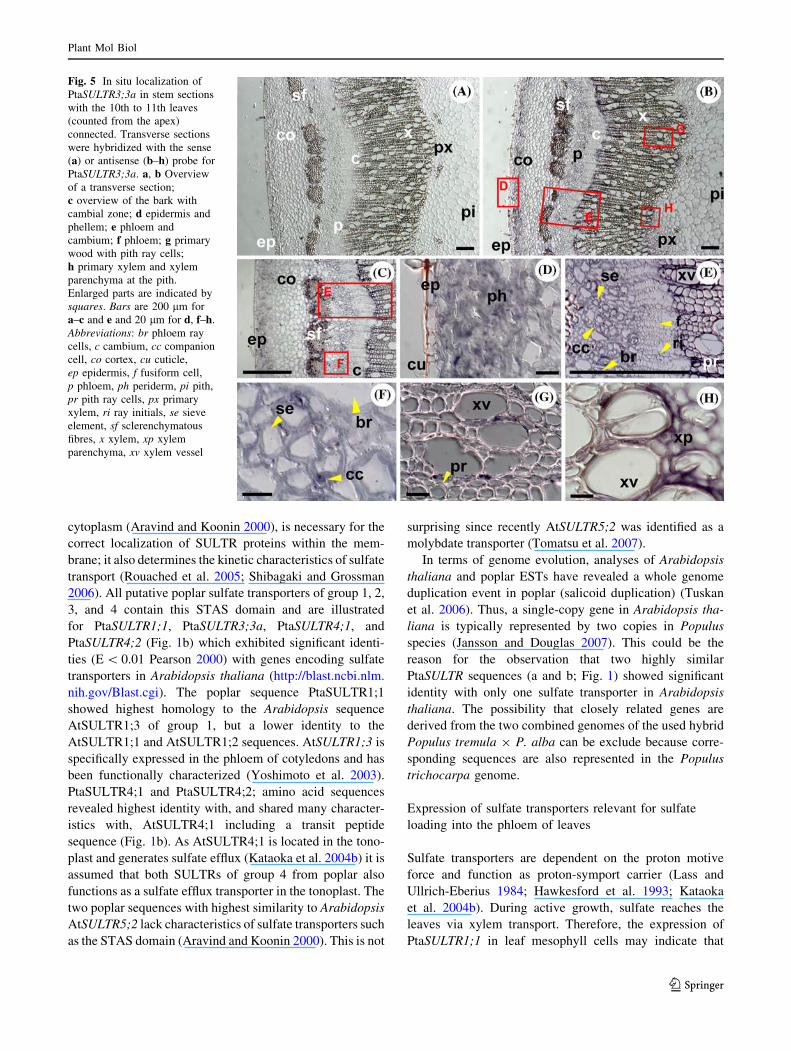
Fig. 5 In situ localization of
PtaSULTR3;3a in stem sections
with the 10th to 11th leaves
(counted from the apex)
connected. Transverse sections
were hybridized with the sense
(a) or antisense (b–h) probe for
PtaSULTR3;3a. a, b Overview
of a transverse section;
c overview of the bark with
cambial zone; d epidermis and
phellem; e phloem and
cambium; f phloem; g primary
wood with pith ray cells;
h primary xylem and xylem
parenchyma at the pith.
Enlarged parts are indicated by
squares. Bars are 200 lm for
a–c and e and 20 lm for d, f–h.
Abbreviations: br phloem ray
cells, c cambium, cc companion
cell, co cortex, cu cuticle,
ep epidermis, f fusiform cell,
p phloem, ph periderm, pi pith,
pr pith ray cells, px primary
xylem, ri ray initials, se sieve
element, sf sclerenchymatous
fibres, x xylem, xp xylem
parenchyma, xv xylem vessel
Plant Mol Biol
123

PtaSULTR1;1 transports sulfate from the apoplast to the
symplast. Mature poplar leaves are sources of carbohy-
drates (Dickson 1991) and sulfate (Hartmann et al. 2000).
Minor leaf veins are associated with the collection phloem
and major leaf veins contribute to the transport phloem
(van Bel 2003). Thus the abundance of PtaSULTR1,1
mRNA in the phloem of minor leaf veins indicates the
importance of PtaSULTR1,1 in loading sulfate into the
phloem. A comparable function can be assumed for
PtaSULTR1;1 and PtaSULTR3;3a in major leaf veins, since
both transcripts were detected in the phloem. Accumulation
of sulfate in the bark starts in late autumn when the
expression of PtaSULTR1;1 in senescent leaves is high.
This indicates an increase in phloem loading of sulfate in
the leaves for sulfate storage in the stem (Fig. 9a). This was
supported by the observation, that the sulfate content in
leaves negatively correlated with the PtaSULTR1;1 mRNA
level. Moreover, the sulfate content in senescent leaves was
significantly lower compared to late summer. However,
whether other sulfur sources, for example thiols or proteins,
are mobilized and converted to sulfate still needs to be
investigated.
An interesting point is that, in leaves, the expression of
sulfate transporters is independent of weather parameters.
Only PtaSULTR4;2 showed a negative correlation with
maximum temperature (Table 2). Hence SULTR gene
expression seems mainly determined by a developmental
program in leaves. Environmental factors may become
more important 1 month after bud break and later in
summer (Wissel et al. 2003). Accumulation of sulfate, and
of PtaSULTR1;1 and PtaSULTR3;3a mRNAs in the leaves
was not constant throughout leaf development. After bud
break, sulfate decreased in expanding leaves, probably due
to a dilution effect caused by leaf expansion or by high
rates of sulfate assimilation. Simultaneously, PtaSULTR1;1
expression increased and reached a maximum when leaves
were fully expanded (Fig. 9b, c). Sulfate uptake into leaf
mesophyll cells, probably mediated by PtaSULTR1;1, may
remain high during leaf development until leaves are fully
expanded. The sulfate content in mature leaves increased
during early summer when the expression of PtaSULTR4;2
and PtaSULTR1;1 decreased. It can be assumed that the
decline in PtaSULTR4;2 and PtaSULTR1;1 mRNA restricts
both sulfate efflux from the vacuole and sulfate loading
into the phloem that enables sulfate accumulation in mature
leaves. Expression of the second phloem-specific sulfate
transporter, PtaSULTR3;3a, which is only expressed in
major leaf veins, increased in early summer when the
expression of PtaSULTR1;1 was low. Hence, phloem
re-loading gets relevant during the summer.
Fig. 6 In situ localization of
PtaSULTR1;1 in fine roots.
Transverse sections of fine roots
were hybridized with the sense
(a, c) or antisense (b, d–h)
probe for PtaSULTR1;1. a, bOverview of a transverse
section of fine roots; c, d cross
section of the stele with a
tetrarch xylem; e transverse
section of a tetrarch stele with
side root; f fine root whose stele
contains a diarch xylem. g, hlongitudinal sections of fine root
with developing side root.
Enlarged parts are indicated by
squares. A green arrow in c, dindicates phenolic compounds.
Bars are 200 lm for a and b and
50 lm for c–h. Abbreviations:
co cortex, ed endodermis,
ep epidermis, mx metaxylem,
p phloem, px protoxylem,
sr side root, st stele
Plant Mol Biol
123

Expression of sulfate transporters in stem tissues
relevant for sulfate storage and mobilization
PtaSULTR3;3a mRNA was detected in sieve elements and
companion cells along the entire stem. Since mature sieve
elements do not possess translation machinery (van Bel et al.
2002), the presence of PtaSULTR3;3a mRNA in sieve ele-
ments seems surprising. However, transcripts of several
cellular proteins including a sucrose transporter have been
found in sieve elements by in situ hybridization (Kuhn et al.
1997) or in phloem exudates (Doering-Saad et al. 2006;
Lough and Lucas 2006; Omid et al. 2007; Le Hir et al. 2008;
Kehr and Buhtz 2008). Leakage of PtaSULTR3;3a mRNA
from the companion cells into the sieve element for move-
ment by phloem mass flow may therefore possible. Such a
movement has been demonstrated by grafting experiments
for several sequences (referred in Kehr and Buhtz 2008).
PtaSULTR1;1 transcripts were strongly detected in
companion cells and ray initials throughout the stem. Fur-
thermore, 35S accumulation was observed in these cells after35S-sulfate was fed to mature leaves (Fig. 7). The accumu-
lation of 35S-sulfate in companion cells below the fed leaf
indicates sieve element unloading. van Bel and Kempers
(1990) found that the sieve element/companion cell com-
plex is symplastically isolated from the surrounding paren-
chyma in Salix alba and Ricinus. If this is also true for
poplar, sulfate must be transported out of the complex into
the apoplast. This unloading could be mediated by voltage
dependent anion channels (Frachisse et al. 1999; Roberts
2006; Fig. 9a). Phloem unloading via PtaSULTR1;1 seems
unlikely because sulfate transport into cells via the plasma
membrane depends on a proton motive force in plants (Lass
and Ullrich-Eberius 1984; Hawkesford et al. 1993) and, a
high Em of the sieve element/companion cell complex (van
Bel and Kempers 1990) probably facilitates sulfate uptake
from the apoplast. Correspondingly, in Populus tricho-
carpa, companion cells but not sieve elements possess
plasma membrane ATPases (Arend et al. 2002). Thus
PtaSULTR1;1 and PtaSULTR3;3a transporters might be
involved in the retrieval of sulfate leaking out of the phloem.
A comparable function has been postulated for sucrose
transporters similarly expressed along the transport phloem
(Williams et al. 2000; van Bel 2003; Carpaneto et al. 2005).
However, an accumulation of sulfate, which comes from
sieve elements, in companion cells cannot be achieved by
diffusion process through plasmodesmata between the two
cell types. This discrepancy can only be investigated by
further studies. For instance, by creating poplar plants
deficient in PtaSULTR1;1 and/or PtaSULTR3;3a expression.
However, a possible complementation of sulfate transport
into the phloem by other sulfate transporters of the gene
family has to be taken into account. This effect is evident
from double and triple sultr knockout mutants of Arabid-
opsis (Lydiate and Higgins, personal communication).
Fig. 7 Microautoradiographic
detection of 35S in stem sections
basal from a mature leaf to
which 35SO42- was fed into the
phloem via flap feeding.
a Overview of a transverse
section of the stem basal to the
fed leaf (209). b Phloem with
strong staining in companion
cells; c cambium with strong
staining in ray initials; d young
wood with pith rays. c to d were
taken at 50-fold extension. The
black grains indicate 35S sulfur.
Bars are 50 lm for a and 20 lm
for b–d. Abbreviations: cccompanion cell, f fusiform cell,
pr pith ray, ri ray initials, sesieve element, x xylem, xvxylem vessel
Plant Mol Biol
123

The phloem-specific sulfate transporter PtaSULTR3;3a
and the putative vacuolar efflux transporter PtaSULTR4;2
are positively correlated and both are negatively correlated
with the sulfate content in bark. The sulfate level in the bark
reached maximum values at the end of December, when
both PtaSULTR3;3a and PtaSULTR4;2 showed nearly no
expression. Since PtaSULTR4;2 is involved in sulfate
mobilization from the vacuole (Kataoka et al. 2004b), and
PtaSULTR3;3a is relevant for phloem loading (see above)
these observations are consistent (Fig. 9a). During bud
break, sulfur was mobilized from soluble and insoluble
storage pools in bark and wood tissues of beech (Herschbach
and Rennenberg 1996). Phloem transport enables swelling
buds to be supplied with metabolites during insufficient
xylem sap flow. The increased level of PtaSULTR4;2
mRNA, encoding a putative vacuolar sulfate transporter,
during bud swelling and early leaf development indicates
sulfate mobilization from the vacuole of bark cells (Fig. 9b).
Table 1 Correlation matrix of sulfate transporter transcript levels, sulfate content of bark and weather parameters
PtaSULTR1;1 PtaSULTR3;3a PtaSULTR4;1 PtaSULTR4;2 Sulfate
PtaSULTR3;3a 0.301
PtaSULTR4;1 0.546** 0.013
PtaSULTR4;2 0.363 0.881** 0.238
Sulfate -0.010 -0.621** 0.375 -0.521**
T mean -0.068 0.749** -0.454 0.596** -0.682**
T max -0.100 0.703** -0.421 0.583** -0.529**
T min 0.008 0.691** -0.399 0.491** -0.750**
Day length -0.067 0.814** -0.138 0.866** -0.639**
Values presented are Pearson correlation coefficients. Two asterisks represent significant positive or negative correlation at P \ 0.01
Fig. 8 Quantitative analyses of SULTR expression and sulfate
contents in bark (left column) and leaf (right column) tissue during
seasonal growth. a Sulfate content of leaves and bark from twigs
which developed in 2006 (light gray squares) and from twigs
which developed in 2007 (dark gray circles). PtaSULTR1;1
(b), PtaSULTR3;3a (c), PtaSULTR4;1 (d), PtaSULTR4;2 (e) and
expression of the elongation factor 1-beta (f) was analyzed in leaves
and bark from twigs developed in 2006 (light gray bars) and in 2007
(dark gray bars). Expression of mRNA copies was related to total
RNA
Plant Mol Biol
123
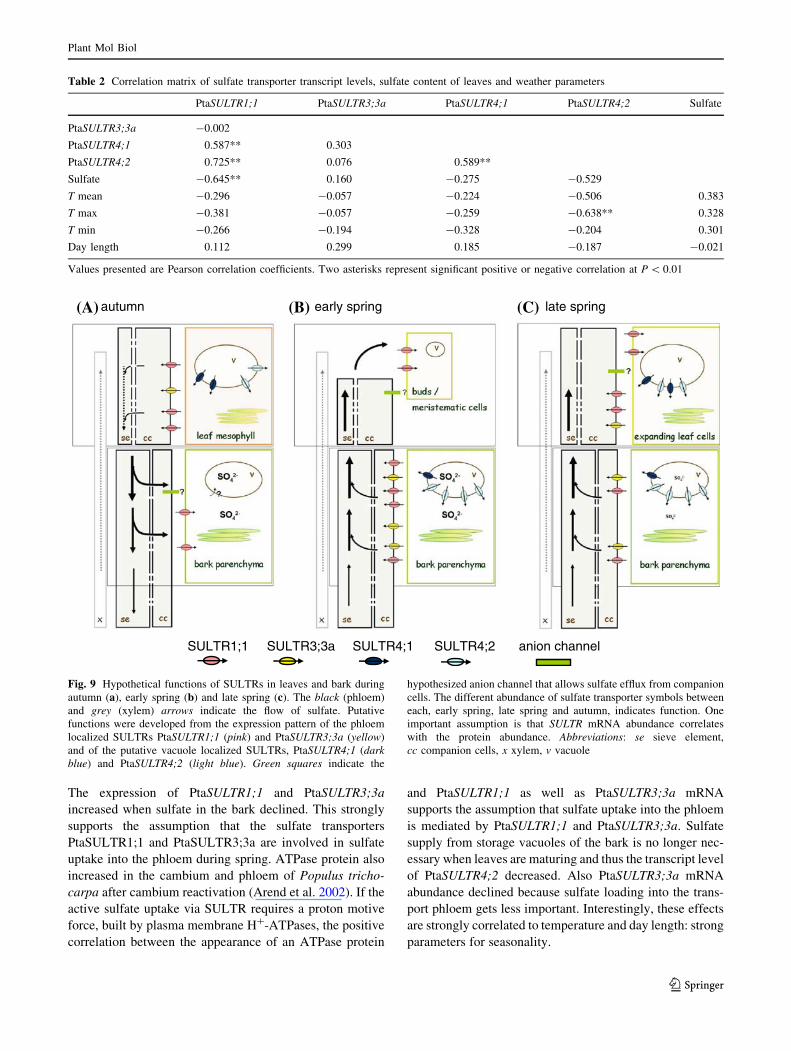
The expression of PtaSULTR1;1 and PtaSULTR3;3a
increased when sulfate in the bark declined. This strongly
supports the assumption that the sulfate transporters
PtaSULTR1;1 and PtaSULTR3;3a are involved in sulfate
uptake into the phloem during spring. ATPase protein also
increased in the cambium and phloem of Populus tricho-
carpa after cambium reactivation (Arend et al. 2002). If the
active sulfate uptake via SULTR requires a proton motive
force, built by plasma membrane H?-ATPases, the positive
correlation between the appearance of an ATPase protein
and PtaSULTR1;1 as well as PtaSULTR3;3a mRNA
supports the assumption that sulfate uptake into the phloem
is mediated by PtaSULTR1;1 and PtaSULTR3;3a. Sulfate
supply from storage vacuoles of the bark is no longer nec-
essary when leaves are maturing and thus the transcript level
of PtaSULTR4;2 decreased. Also PtaSULTR3;3a mRNA
abundance declined because sulfate loading into the trans-
port phloem gets less important. Interestingly, these effects
are strongly correlated to temperature and day length: strong
parameters for seasonality.
Table 2 Correlation matrix of sulfate transporter transcript levels, sulfate content of leaves and weather parameters
PtaSULTR1;1 PtaSULTR3;3a PtaSULTR4;1 PtaSULTR4;2 Sulfate
PtaSULTR3;3a -0.002
PtaSULTR4;1 0.587** 0.303
PtaSULTR4;2 0.725** 0.076 0.589**
Sulfate -0.645** 0.160 -0.275 -0.529
T mean -0.296 -0.057 -0.224 -0.506 0.383
T max -0.381 -0.057 -0.259 -0.638** 0.328
T min -0.266 -0.194 -0.328 -0.204 0.301
Day length 0.112 0.299 0.185 -0.187 -0.021
Values presented are Pearson correlation coefficients. Two asterisks represent significant positive or negative correlation at P \ 0.01
SULTR4;1SULTR1;1 SULTR3;3a SULTR4;2 anion channel
autumn early spring late spring(A) (B) (C)
Fig. 9 Hypothetical functions of SULTRs in leaves and bark during
autumn (a), early spring (b) and late spring (c). The black (phloem)
and grey (xylem) arrows indicate the flow of sulfate. Putative
functions were developed from the expression pattern of the phloem
localized SULTRs PtaSULTR1;1 (pink) and PtaSULTR3;3a (yellow)
and of the putative vacuole localized SULTRs, PtaSULTR4;1 (darkblue) and PtaSULTR4;2 (light blue). Green squares indicate the
hypothesized anion channel that allows sulfate efflux from companion
cells. The different abundance of sulfate transporter symbols between
each, early spring, late spring and autumn, indicates function. One
important assumption is that SULTR mRNA abundance correlates
with the protein abundance. Abbreviations: se sieve element,
cc companion cells, x xylem, v vacuole
Plant Mol Biol
123

After 35S-sulfate feeding to a mature leaf, 35S became
enriched in ray initials basal to the fed leaf. This indicates the
origin of stored sulfate by sulfate uptake into ray cells.
Hence, the high expression of PtaSULTR1;1 supports sulfate
uptake from the apoplast into ray initials. Further transport
within the ray pith could occur by cell to cell transport via
plasmodesmata. Sulfate storage along the stem in pith ray
cells is also possible after its removal from the xylem. In a
similar manner as the putative sucrose transporter JrSUT1
(Decourteix et al. 2006) PtaSULTR1;1 and PtaSULTR3;3a
encoding mRNAs are detected in pith cells that are con-
nected to xylem vessels. A prerequisite for sucrose and sul-
fate uptake into these cells again requires the presence of a
proton motive force. ATPase was detected in ray cells con-
nected with xylem vessels of Populus trichocarpa twigs after
cambial reactivation during spring (Arend et al. 2002). Both,
the expression of PtaSULTR1;1 and PtaSULTR3;3a as well
as the presence of a plasma membrane H?-ATPase support
an unloading of sulfate from the xylem into vessel-associated
ray cells. The same process could also be relevant for both
transporters in major leaf veins. However, several further
sulfate transporters were found in wood of poplar (see Sup-
plemental Fig. S1) that may contribute to xylem unloading of
sulfate during vegetative growth.
Expression of sulfate transporters in developing tissues
(apex and lateral roots)
Previous investigations showed that sulfate is transported
from leaves to the shoot apex and fine roots (Hartmann
et al. 2000). Whereas PtaSULTR3;3a transcripts were only
visible in the apical meristem of the shoot (SAM),
PtaSULTR1,1 transcripts were highly abundant in leaf
primordia, leaf margin meristems (LMM) and provascular
strands. Both sulfate transporters could be responsible for
sulfate uptake into dividing cells providing sulfate for
assimilation. The relevance of sulfate assimilation in het-
erotrophic and growing tissues is still unknown. Hartmann
et al. (2000) concluded that sulfate assimilation in the apex
is possible and has been demonstrated by 35S-sulfate
feeding to a separated apex (Herschbach 2003).
In fine roots from maize, highest APS reductase activity,
the regulatory step of sulfate reduction (Kopriva and
Koprivova 2004; Martin et al. 2005), has been detected in
the root tips (Kopriva et al. 2001). High PtaSULTR1;1
transcript levels in the root meristem of developing lateral
roots are consistent with these findings. Sulfate uptake via
PtaSULTR1;1 into dividing and growing cells supplies
sulfate for the sulfate reduction and assimilation pathway
relevant in poplar roots (Scheerer et al. 2009). The abun-
dance of PtaSULTR1;1 mRNA in the protophloem of fine
roots supports the idea of loading sulfate into the phloem
for further allocation to the root tip.
Conclusion
Our results provide novel evidence for the seasonal regu-
lation of sulfate storage, mobilization and distribution via
the phloem at the transcription level of sulfate transporters.
In the perennial model species Populus two sulfate trans-
porters, PtaSULTR3;3a and PtaSULTR1;1, are expressed in
the phloem of leaf veins and along the stem. Important
processes of deciduous trees such as poplar are sulfate
storage during winter and sulfate mobilization during
spring (Herschbach and Rennenberg 1996; Rennenberg
et al. 2007). These processes cannot be investigated in
Arabidopsis. The mRNA of PtaSULTR3;3a and of the
putative vacuolar located sulfate efflux transporter
PtaSULTR4;2 in the bark increased in spring during bud
break (summarized in Fig. 9) and correlated with the day
length, temperature and, most importantly, inversely with
sulfate content. In Arabidopsis, one function of the sulfate
transporters of group 4 seems to be the delivery of sulfate
from the vacuole for further loading into the xylem (Kat-
aoka et al. 2004b). Here, we present evidence for the
seasonal function of PtaSULTR4;2 and for its role in sul-
fate mobilization from the vacuole, and of PtaSULTR3;3a
and PtaSULTR1;1 for sulfate loading into the phloem.
Moreover, the clear correlation with temperature and day
length which are characteristic for seasonal changes
revealed environmental control for the expression of
PtaSULTR4;2 and PtaSULTR3;3a in the bark. In contrast,
the expression of sulfate transporters seems mainly under
developmental control in leaves. Therefore, the regulatory
signals, inducing changes in gene expression, may vary
among tissues and different sulfate transporters.
Acknowledgments This work was financial supported by the Deut-
sche Forschungsgemeinschaft (DFG) under the contract numbers HE
3003/2 & 3. The Excellence Initiative of the German Federal and State
Governments (EXC 294), SFB 592 and the Landesstiftung are grate-
fully acknowledged. The authors thank Simone Sikora and Katja Rapp
for technical assistance and Dr. William Teale to improve language.
References
Adiputra IGK, Anderson JW (1995) Effect of sulfur nutrition on
redistribution of sulfur in vegetative barely. Physiol Plant
95:643–650
Aravind L, Koonin EV (2000) The STAS domain a link between
anion transporters and antisigma-factor antagonists. Curr Biol
10:53–55
Arend M, Weisenseel MH, Brummer M, Osswald W, Fromm J (2002)
Seasonal changes of plasmamembrane H?-ATPase and endog-
enous ion current during cambial growth in poplar plants. Plant
Physiol 129:1651–1663
Bell CI, Cram WJ, Clarkson DT (1994) Compartmental analysis of35SO4
2- exchange kinetics in roots and leaves of a tropical
legume Macroptilium atropurpureum cv. Siratro. J Exp Bot
45:879–886
Plant Mol Biol
123

Biddulph SF (1956) Visual indications of 35S and 32P translocation in
the phloem. Am J Bot 43:143–148
Brunner AM, Yakovlev IA, Strauss SH (2004) Validating internal
controls for quantitative plant gene expression studies. BMC
Plant Biol 4:14
Buchner P, Takahashi H, Hawkesford M (2004a) Plant sulfate
transporters: co-ordination of uptake, intracellular and long
distance transport. J Exp Bot 55:1765–1773
Buchner P, Stuiver EE, Westerman S, Wirtz M, Hell R, Hawkesford
MJ, De Kok LJ (2004b) Regulation of sulfate uptake and
expression of sulphate transporter genes in Brassica oleracea as
affected by atmospheric H2S and pedospheric sulphate nutrition.
Plant Physiol 136:3396–3408
Carpaneto A, Geiger D, Bamberg E, Sauer NM, Fromm J, Hedrich R
(2005) Phloem-localized, proton-coupled sucrose carrier
ZmSUT1 mediates sucrose efflux under the control of the sucrose
gradient and the proton motive force. J Biol Chem 280:21437–
21443
Cnops G, Neyt P, Raes J, Petrarulo M, Nelissen H, Malenica N,
Luschnig C, Tietz O, Ditengou F, Palme K, Azmi A, Prinsen E,
van Lijsebettens M (2006) The TORNADO1 and TORNADO2
genes function in several patterning processes during early leaf
development in Arabidopsis thaliana. Plant Cell 18:852–866
Decourteix M, Alves G, Brunel N, Ameglio T, Guilliot A, Lemoine
R, Peter G, Sakr S (2006) JrSUT1, a putative xylem sucrose
transporter, could mediate sucrose influx into xylem parenchyma
cells and be up-regulated by freeze-thaw cycles over the autumn-
winter period in walnut tree (Juglans regia L.). Plant Cell
Environ 29:36–47
Deeken R, Ache P, Kajahn I, Klinkenberg J, Bringmann G, Hedrich R
(2008) Identification of Arabidopsis thaliana RNAs provides a
search criterion for phloem-based transcripts hidden in complex
datasets of microarray experiments. Plant J 55:746–759
Dickson RE (1991) Assimilate distribution and storage. In: Ragha-
vendra AS (ed) Physiology of trees. Wiley, New York, pp 51–85
Doering-Saad C, Newbury HJ, Couldridge CE, Bale JS, Pritchard J
(2006) A phloem-enriched cDNA library from Ricinus: insights
into phloem function. J Exp Bot 57:3183–3193
Emanuelsson O, Nielsen H, von Heijne G (1999) ChloroP, a neural
network-based method for predicting chloroplast transit peptides
and their cleavage sites. Prot Sci 8:978–984
Frachisse J-M, Thomine S, Colcombet J, Guern J, Barbier-Brygoo H
(1999) Sulfate is both a substrate and an activator of the voltage-
dependent anion channel of Arabidopsis hypocotyl cells. Plant
Physiol 121:253–261
Fritz E (1980) Microautoradiographic localization of assimilates in
phloem: problems and new method. Ber Dtsch Bot Ges 93:109–121
Hartmann T, Mult S, Suter M, Rennenberg H, Herschbach C (2000)
Leaf age-dependent differences in sulfur assimilation and
allocation in poplar (Populus tremula 9 P. alba) leaves. J Exp
Bot 51:1077–1088
Hawkesford MJ (2003) Transporter gene families in plants: the sulfate
transporter gene family—redundancy or specialization? Physiol
Plant 117:155–163
Hawkesford MJ, Davidian J-C, Grignon C (1993) Sulfate/proton
cotransport in plasma-membrane vesicles isolated from roots of
Brassica napus L.: increased transport in membranes isolated
from sulfur starves plants. Planta 190:297–304
Heiss S, Schafer HJ, Haag-Kerwer A, Rausch T (1999) Cloning sulfur
assimilation genes of Brassica juncea L.: cadmium differentially
affects the expression of putative low-affinity sulphate trans-
porter and isoforms of ATP sulphurylase and APS reductase.
Plant Mol Biol 39:847–857
Herschbach C (2003) Whole plant regulation of sulfur nutrition of
deciduous trees—influences of the environment. Plant Biol
5:233–244
Herschbach C, Rennenberg H (1996) Storage and remobilization of
sulfur in beech trees (Fagus sylvatica). Physiol Plant 98:
125–132
Herschbach C, Rennenberg H (2001a) Significance of phloem-
translocated organic sulfur compounds for the regulation of
sulfur nutrition. Prog Bot 62:177–193
Herschbach C, Rennenberg H (2001b) Sulfur nutrition of deciduous
trees. Naturwissenschaften 88:25–36
Herschbach C, van der Zalm E, Schneider A, Jouanin L, De Kok LJ,
Rennenberg H (2000) Regulation of sulfur nutrition in wild-type
and transgenic poplar over-expressing c-glutamylcysteine syn-
thetase in the cytosol as affected by atmospheric H2S. Plant
Physiol 124:461–473
Hopkins L, Parmar S, Bouranis DL, Howarth JR, Hawkesford MJ
(2004) Coordinated expression of sulphate uptake and compo-
nents of the sulphate assimilatory pathway in maize. Plant Biol
6:408–414
Jansson S, Douglas CJ (2007) Populus: a model system for plant
biology. Annu Rev Plant Biol 58:435–458
Jones DT, Taylor WR, Thorton JM (1994) A model recognition
approach to the prediction of all-helical membrane-protein
structure and topology. Biochemistry 33:3038–3049
Kataoka T, Hayashi N, Yamaya T, Takahashi H (2004a) Root-to-
shoot transport of sulfate in Arabidopsis. Evidence for the role of
SULTR3;5 as a component of low-affinity sulfate transport
system in the root vasculature. Plant Physiol 136:4198–4204
Kataoka T, Watanabe-Takahashi A, Hayashi N, Ohnishi M, Mimura
T, Buchner P, Hawkesford MJ, Yamaya T, Takahashi H (2004b)
Vacuolar sulfate transporters are essential determinants control-
ling internal distribution of sulfate in Arabidopsis. Plant Cell
16:2693–2704
Kehr J, Buhtz A (2008) Long distance transport and movement of
RNA through the phloem. J Exp Bot 59:85–92
Kolosova N, Miller B, Ralph S, Ellis BE, Douglas C, Ritland K,
Bohlmann J (2004) Isolation of high-quality RNA from gym-
nosperm and angiosperm trees. Biotechniques 36:821–824
Kopriva S, Koprivova A (2004) Plant adenine 50-phosphosulfate
reductase: the past, the present, and the future. J Exp Bot
55:1775–1783
Kopriva S, Jones S, Koprivova A, Suter M, von Ballmoos P, Brander
K, Fluckiger J, Brunold C (2001) Influence of chilling stress on
the intercellular distribution of assimilatory sulfate reduction and
thiols in Zea mays. Plant Biol 3:24–31
Koralewska A, Buchner P, Stuiver CEE, Posthumus FS, Kopriva S,
Hawkesford MJ, De Kok LJ (2009) Expression and activity of
sulphate transporters and APS reductase in crly kale in response
to sulphate deprivation and re-supply. J Plant Physiol 166:
168–179
Kuhn C, Franceschi VR, Schulz A, Lemoine R, Frommer WB (1997)
Macromolecular trafficking indicated by locatization and turn-
over of sucrose transporters in enucleated sieve elements.
Science 275:1298–1300
Lass B, Ullrich-Eberius CI (1984) Evidence for proton/sulfate
cotransport and its kinetics in Lemna gibba G1. Planta 161: 53–60
Le Hir R, Beneteau J, Bellini C, Vilaine F, Dinant S (2008) Gene
expression profiling: keys for investigating phloem functions.
Trends Plant Sci 13:273–280
Lough TJ, Lucas WJ (2006) Integrative plant biology: role of phloem
long-distance macromolecular trafficking. Annu Rev Plant Biol
57:203–232
Martin MN, Tarczynski MC, Shen B, Leustek T (2005) The role of 50-adenylylsulfate reductase in controlling sulfate reduction in
plants. Photosynth Res 86:309–329
Nicot N, Hausman JF, Hoffmann L, Evers D (2005) Housekeeping
gene selection for real-time RT-PCR normalization in potato
during biotic and abiotic stress. J Exp Bot 56:2907–2914
Plant Mol Biol
123

Nocito FF, Lancilli C, Crema B, Fourcroy P, Davidian J-C, Sacchi
GA (2006) Heavy metal stress and sulfate uptake in maize roots.
Plant Physiol 141:1138–1148
Noctor G, Strohm M, Jouanin L, Kunert K-J, Foyer CH, Rennenberg
H (1996) Synthesis of glutathione in leaves of transgenic poplar
overexpression c-glutamylcysteine synthetase. Plant Physiol
112:1071–1078
Omid A, Keilin T, Glass A, Leshkowitz D, Wolf S (2007)
Characterisation of phloem-sap transcription profile in melon
plants. J Exp Bot 58:3645–3656
Pearson WR (2000) Flexible sequence similarity searching with the
FASTA3 program package. Methods Mol Biol 132:185–219
Persson B, Argos P (1994) Prediction of transmembrane segments in
proteins utilizing multiple sequence alignments. J Mol Biol
237:182–192
Rennenberg H, Herschbach C, Harberer K, Kopriva S (2007) Sulfur
metabolism in plants: are trees different? Plant Biol 9:620–637
Roberts SK (2006) Plasma membrane anion channels in higher plants
and their putative functions in roots. New Phytol 169:647–666
Rouached H, Berthomieu P, El Kassis E, Cathala N, Catherinot V,
Labesse G, Davidian JC, Fourcroy P (2005) Structural and
functional analysis of the C-terminal STAS (sulfate transporter
and anti-sigma antagonist) domain of the Arabidopsis thalianasulfate transporter SULTR1.2. J Biol Chem 280:15976–15983
Rouached H, Wirtz M, Alary R, Hell R, Arpat AB, Davidian J-C,
Fourcroy P, Berthomieu P (2008) Differentialregulation of the
expression of two high-affinity sulphate transporters, SULTR1.1
and SULTR1.2, in Arabidopsis. Plant Physiol 147:897–911
Saito K (2004) Sulfur assimilatory metabolism. The long and
smelling road. Plant Physiol 136:2443–2450
Sanderson J (1981) Modified development to improve the perfor-
mance of AR-10 stripping emulsions for use the more energetic
isotopes. J Microsc 124:177–182
Scheerer U, Haensch R, Mendel R, Kopriva S, Rennenberg H,
Herschbach C (2009) Sulphur flux through the sulphate assim-
ilation pathway is differently controlled by adenosine 50-phosphosulphate reductase under stress and in transgenic poplar
plants overexpressing c-ECS, SO or APR. J Exp Bot. doi:
10.1093/jxb/erp327
Schrader J, Moyle R, Bhalerao R, Hertzberg M, Lundeberg J, Nilsson P,
Bhalerao RP (2004) Cambial meristem dormancy in trees involves
extensive remodelling of the transcriptome. Plant J 40:173–187
Schulte M, Herschbach C, Rennenberg H (1998) Interactive effects of
elevated atmospheric CO2, mycorrhization and drought on long-
distance transport of reduced sulfur in young pedunculate oak
trees (Quercus robur L.). Plant Cell Environ 21:917–926
Shibagaki N, Grossman AR (2004) Probing the function of STAS
domains of the Arabidopsis sulfate transporters. J Biol Chem
279:30791–30799
Shibagaki N, Grossman AR (2006) The role of the STAS domain in
the function and biogenesis of a sulfate transporter as probed by
random mutagenesis. J Biol Chem 281:22964–22973
Shibagaki N, Rose A, McDermott JP, Fujiwara T, Hayashi H, Yoneyama
T, Davies JP (2002) Selenate-resistant mutants of Arabidopsisthaliana identify Sultr1;2 a sulfate transporter required for efficient
transport of sulfate into roots. Plant J 29:475–489
Smith IK, Lang AL (1988) Translocation of sulfate in soybean
(Glycine max L. Merr). Plant Physiol 86:798–802
Smith FW, Ealing PM, Hawkesford MJ, Clarkson DT (1995) Plant
members of a family of sulfate transporters reveal functional
subtypes. Proc Natl Acad Sci USA 92:9373–9377
Smith FW, Hawkesford MJ, Ealing PM, Clarkson DT, Vanden Berg
PJ, Belcher AR, Warrilow GS (1997) Regulation of a cDNA
from barley roots encoding a high affinity sulfate transporter.
Plant J 12:875–884
Spurr AR (1969) A low-viscosity epoxy resin embedding medium for
electron microscopy. J Ultrastruct Res 26:31–43
Strohm M, Jouanin L, Kunert KJ, Pruvost C, Polle A, Foyer C,
Rennenberg H (1995) Regulation of GSH synthesis in leaves of
transgenic poplar (Populus tremula 9 P. alba) overexpressing
GSH synthetase. Plant J 7:141–145
Takahashi H, Yamazaki M, Sasakura N, Watanabe A, Leustek T,
Engler JA, Engler G, van Montagu M, Saito K (1997)
Regulation of sulfur assimilation in higher plants: a sulfate
transporter induced in sulfate-starved roots plays a central
role in Arabidopsis thaliana. Proc Natl Acad Sci USA
94:11102–11107
Takahashi H, Watanabe-Takahashi A, Smith FW, Blake-Kalff M,
Hawkesford MJ, Saito K (2000) The roles of three functional
sulfate transporters involved in uptake and translocation of
sulfate in Arabidopsis thaliana. Plant J 23:171–182
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol 24:1596–1599
Tatusova TA, Madden TL (1999) BLAST 2 Sequences, a new tool
for comparing protein and nucleotide sequences. FEMS 174:
247–250
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:
4673–4680
Tomatsu H, Takano J, Takahashi H, Watanabe-Takahashi A,
Shibagaki N, Fujiwara T (2007) An Arabidopsis thalianahigh-affinity molybdate transporter required for efficient
uptake of molybdate from soil. Proc Natl Acad Sci USA
104:18807–18812
Tuskan GA, DiFazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten
U, Putnam N, Ralph S, Rombauts S, Salamov A, Schein J, Sterck
L, Aerts A, Bhalerao RR, Bhalerao RP, Blaudez D, Boerjan W,
Brun A, Brunner A, Busov V, Campbell M, Carlson J, Chalot M,
Chapman J, Chen GL, Cooper D, Coutinho PM, Couturier J,
Covert S, Cronk Q, Cunningham R, Davis J, Degroeve S,
Dejardin A, Depamphilis C, Detter J, Dirks B, Dubchak I,
Duplessis S, Ehlting J, Ellis B, Gendler K, Goodstein D,
Gribskov M, Grimwood J, Groover A, Gunter L, Hamberger B,
Heinze B, Helariutta Y, Henrissat B, Holligan D, Holt R, Huang
W, Islam-Faridi N, Jones S, Jones-Rhoades M, Jorgensen R,
Joshi C, Kangasjarvi J, Karlsson J, Kelleher C, Kirkpatrick R,
Kirst M, Kohler A, Kalluri U, Larimer F, Leebens-Mack J, Leple
JC, Locascio P, Lou Y, Lucas S, Martin F, Montanini B, Napoli
C, Nelson DR, Nelson C, Nieminen K, Nilsson O, Pereda V,
Peter G, Philippe R, Pilate G, Poliakov A, Razumovskaya J,
Richardson P, Rinaldi C, Ritland K, Rouze P, Ryaboy D,
Schmutz J, Schrader J, Segerman B, Shin H, Siddiqui A, Sterky
F, Terry A, Tsai CJ, Uberbacher E, Unneberg P, Vahala J, Wall
K, Wessler S, Yang G, Yin T, Douglas C, Marra M, Sandberg G,
Van de Peer Y, Rokhsar D (2006) The genome of black
cottonwood, Populus trichocarpa (Torr & Gray). Science
131:1596–1604
Van Bel AJE (2003) The phloem, a miracle of ingenuity. Plant Cell
Environ 26:125–149
Van Bel AJE, Kempers R (1990) Symplastically isolation of the sieve
element-companion cell complex in the phloem of Ricinuscommunis and Salix alba stems. Planta 183:69–76
Van Bel AJE, Ehlers K, Knoblauch M (2002) Sieve elements caught
in the act. Trends Plant Sci 7:126–132
Vidmar JJ, Schjoerring JK, Touraine B, Glass ADM (1999) Regu-
lation of the hvst1 gene encoding a high-affinity sulfate
transporter from Hordeum vulgare. Plant Mol Biol 40:883–892
Plant Mol Biol
123

Vidmar JJ, Tagmount A, Cathala N, Tourain B, Davidian J-V (2000)
Cloning and characterization of a root specific high-affinity
sulfate transporter from Arabidopsis thaliana. FEBS Lett
475:65–69
Williams LE, Lemoine R, Sauer N (2000) Sugar transporters in higher
plants—a diversity of roles and complex regulation. Trends Plant
Sci 5:283–290
Wissel K, Pettersson F, Berglund A, Jansson S (2003) What affects
mRNA levels in leaves of field-grown aspen? A study of
developmental and environmental influences. Plant Physiol
133:1190–1197
Yoshimoto N, Takahashi H, Smith FW, Yamaya T, Saito K (2002)
Two distinct high-affinity sulfate transporters with different
inducibilities mediate uptake of sulfate in Arabidopsis roots.
Plant J 29:465–473
Yoshimoto N, Inoue E, Saito K, Yamaya T, Takahashi H (2003)
Phloem-localizing sulfate transporter, Sultr1;3, mediates re-
distribution of sulfur from source to sink organs in Arabidopsis.
Plant Physiol 131:1511–1517
Plant Mol Biol
123




















