Sciatic conditioned medium increases survival, proliferation and differentiation of retinal cells in...
10
Int. J. Devl Neuroscience 20 (2002) 11–20 Sciatic conditioned medium increases survival, proliferation and differentiation of retinal cells in culture Patr´ ıcia Maria Mendonça Torres a,b , Carla Valéria Vieira Guilarducci a , Alfred Sholl Franco a,b , Elizabeth Giestal de Araujo a,∗ a Departamento de Neurobiologia, Instituto de Biologia, Universidade Federal Fluminense, CP 100180, 24001-970 Niterói, RJ, Brazil b Centro de Ciˆ encias da Saúde, Instituto de Biof´ ısica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, 21949-900 Rio de Janeiro, RJ, Brazil Received 5 November 2001; received in revised form 21 December 2001; accepted 9 January 2002 Abstract Many evidences clearly demonstrate that Schwann cells provide trophic support for neurons. Different cytokines, including neurotrophins (NTs), are produced and released by Schwann cells. These trophic molecules play an important role on neuronal survival either during the development or during adult life. Cytokines have also a pivotal role on neuronal regeneration after lesions occurring during pathological conditions. The aim of this work was to study the effect of sciatic conditioned medium (SCM) on rat retinal cells maintained in culture. Our results show that treatment with SCM obtained after 14 days in vitro (SCM 14 day) induced a three-fold increase in protein content of the culture after 48 h in vitro and this value remained equally high up to 72 h. This effect was totally blocked either by addition of 30 M BAPTA-AM, an intracellular calcium chelator, 15 M fluorodeoxyuridine, an inhibitor of cell division, or 10 M genistein (geni) plus 1.25 M chelerythrine chloride (CC), the two last ones inhibitors of tyrosine kinases and protein kinase C, respectively. SCM induced an increase in [ 3 H]-choline uptake and [ 3 H]-thymidine incorporation of retinal cells. SCM also stimulated an increase in cytoplasmic processes outgrowth of retinal cells and survival of retinal ganglion cells. Our results clearly suggest that soluble molecules released by sciatic nerve fragments are able to increase the proliferation and survival of retinal cells in culture. © 2002 ISDN. Published by Elsevier Science Ltd. All rights reserved. Keywords: Cytokines; Retina; Schwann cells; Cellular survival; Cell adhesion 1. Introduction Neuronal differentiation and survival in the nervous sys- tem are supported by several microenvironmental signals throughout its development as well as in the adult phase (Svendsen and Sofroniew, 1996). Some of these signaling molecules are generically named cytokines, encompass- ing interleukins, growth factors, differentiation factors and neurotrophic factors (Patterson, 1993). Cytokines increase neuronal survival, promote neurite extension and induce neuronal differentiation (Weisenhorn et al., 1999). Neu- rons can receive trophic support from target cells, afferent cells, glial cells and also from themselves, in an autocrine mechanism (Korsching, 1993). After lesions to peripheral nerves, a series of cellular reac- tions occur in a process termed Wallerian degeneration (Re- ichert et al., 1994). This process is accompanied by changes in the expression of various molecules that have been shown ∗ Corresponding author. Tel.: +21-2717-1161; fax: +21-2719-5934. E-mail address: [email protected] (E.G. de Araujo). to support neuronal survival and neurite outgrowth, includ- ing cell adhesion molecules, extracellular matrix compo- nents and diffusible growth factors (Be’eri et al., 1998). Schwann cells, the glial elements of the peripheral nervous system (PNS), have several intrinsic features that con- tribute to successful axonal regeneration in the injured PNS. Schwann cells produce and release molecules of the extra- cellular matrix, like laminin (Cornbrooks et al., 1983). They also produce and release several cytokines including nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), ciliary neurotrophic factor (CNTF), glial cell line-derived neurotrophic factor (GDNF), interleukin-6 (IL-6), tumor necrosis factor alpha (TNF) and leukemia inhibitor factor (LIF) (Bunge, 1994; Thoenen, 1991; Bolin et al., 1995; Sendtner et al., 1992; Wagner and Myers, 1996; Kurek et al., 1996). In this context, these cytokines are usually named lesion factors (Thoenen, 1991). The ability of Schwann cells to experimentally promote remyelination and regeneration in central axons is of partic- ular interest. Some models using sciatic nerve grafts to optic nerves or to the spinal cord revealed axon regeneration of 0736-5748/02/$22 © 2002 ISDN. Published by Elsevier Science Ltd. All rights reserved. PII:S0736-5748(02)00005-9
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Sciatic conditioned medium increases survival, proliferation and differentiation of retinal cells in...

Int. J. Devl Neuroscience 20 (2002) 11–20
Sciatic conditioned medium increases survival, proliferationand differentiation of retinal cells in culture
Patrıcia Maria Mendonça Torresa,b, Carla Valéria Vieira Guilarduccia,Alfred Sholl Francoa,b, Elizabeth Giestal de Araujoa,∗
a Departamento de Neurobiologia, Instituto de Biologia, Universidade Federal Fluminense, CP 100180, 24001-970 Niterói, RJ, Brazilb Centro de Ciencias da Saúde, Instituto de Biofısica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, 21949-900 Rio de Janeiro, RJ, Brazil
Received 5 November 2001; received in revised form 21 December 2001; accepted 9 January 2002
Abstract
Many evidences clearly demonstrate that Schwann cells provide trophic support for neurons. Different cytokines, including neurotrophins(NTs), are produced and released by Schwann cells. These trophic molecules play an important role on neuronal survival either during thedevelopment or during adult life. Cytokines have also a pivotal role on neuronal regeneration after lesions occurring during pathologicalconditions. The aim of this work was to study the effect of sciatic conditioned medium (SCM) on rat retinal cells maintained in culture.Our results show that treatment with SCM obtained after 14 days in vitro (SCM 14 day) induced a three-fold increase in protein content ofthe culture after 48 h in vitro and this value remained equally high up to 72 h. This effect was totally blocked either by addition of 30�MBAPTA-AM, an intracellular calcium chelator, 15�M fluorodeoxyuridine, an inhibitor of cell division, or 10�M genistein (geni) plus1.25�M chelerythrine chloride (CC), the two last ones inhibitors of tyrosine kinases and protein kinase C, respectively. SCM inducedan increase in [3H]-choline uptake and [3H]-thymidine incorporation of retinal cells. SCM also stimulated an increase in cytoplasmicprocesses outgrowth of retinal cells and survival of retinal ganglion cells. Our results clearly suggest that soluble molecules released bysciatic nerve fragments are able to increase the proliferation and survival of retinal cells in culture. © 2002 ISDN. Published by ElsevierScience Ltd. All rights reserved.
Keywords:Cytokines; Retina; Schwann cells; Cellular survival; Cell adhesion
1. Introduction
Neuronal differentiation and survival in the nervous sys-tem are supported by several microenvironmental signalsthroughout its development as well as in the adult phase(Svendsen and Sofroniew, 1996). Some of these signalingmolecules are generically named cytokines, encompass-ing interleukins, growth factors, differentiation factors andneurotrophic factors (Patterson, 1993). Cytokines increaseneuronal survival, promote neurite extension and induceneuronal differentiation (Weisenhorn et al., 1999). Neu-rons can receive trophic support from target cells, afferentcells, glial cells and also from themselves, in an autocrinemechanism (Korsching, 1993).
After lesions to peripheral nerves, a series of cellular reac-tions occur in a process termed Wallerian degeneration (Re-ichert et al., 1994). This process is accompanied by changesin the expression of various molecules that have been shown
∗ Corresponding author. Tel.:+21-2717-1161; fax:+21-2719-5934.E-mail address:[email protected] (E.G. de Araujo).
to support neuronal survival and neurite outgrowth, includ-ing cell adhesion molecules, extracellular matrix compo-nents and diffusible growth factors (Be’eri et al., 1998).Schwann cells, the glial elements of the peripheral nervoussystem (PNS), have several intrinsic features that con-tribute to successful axonal regeneration in the injured PNS.Schwann cells produce and release molecules of the extra-cellular matrix, like laminin (Cornbrooks et al., 1983). Theyalso produce and release several cytokines including nervegrowth factor (NGF), brain-derived neurotrophic factor(BDNF), neurotrophin-3 (NT-3), ciliary neurotrophic factor(CNTF), glial cell line-derived neurotrophic factor (GDNF),interleukin-6 (IL-6), tumor necrosis factor alpha (TNF�)and leukemia inhibitor factor (LIF) (Bunge, 1994; Thoenen,1991; Bolin et al., 1995; Sendtner et al., 1992; Wagnerand Myers, 1996; Kurek et al., 1996). In this context, thesecytokines are usually named lesion factors (Thoenen, 1991).
The ability of Schwann cells to experimentally promoteremyelination and regeneration in central axons is of partic-ular interest. Some models using sciatic nerve grafts to opticnerves or to the spinal cord revealed axon regeneration of
0736-5748/02/$22 © 2002 ISDN. Published by Elsevier Science Ltd. All rights reserved.PII: S0736-5748(02)00005-9

12 P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20
central neurons (Dezawa and Nagano, 1993; Richardson etal., 1980; Tuszynski et al., 1998). Other experiments haveshown that sciatic conditioned media increases process out-growth and survival of dissociated neurons (Watabe et al.,1995; Dobresov et al., 1994; Kuffler and Megwinoff, 1994).
The neural retina is an excellent model system forstudying survival and differentiation of the CNS. It has awell-defined laminar structure with five major types of neu-rons including photoreceptors, horizontal, bipolar, amacrineand ganglion cells (Dowling, 1991). The retina has oneprincipal type of macroglial cell, the Müller cells. Oligoden-drocytes are completely absent in the retinae of most speciesand astrocytes are present only in the nerve fiber layer ofmammalian species (Newman and Reichenbach, 1996).
To study the action of factors released from peripheralnerve on developing central neurons, we analyzed the effectof sciatic conditioned medium (SCM) on neonatal rat retinalcells in vitro. Our data suggest that lesion factors releasedby fragments of sciatic nerve have a remarkable effect onneonatal rat retinal cells in culture. SCM increased cell ad-hesion, cytoplasmic processes outgrowth, neuronal survival,cholinergic differentiation and cell proliferation of retinalcell cultures.
2. Materials and methods
2.1. Materials
Medium 199 was purchased from Gibco, (Gaithers-burg, USA). Brefeldin A (BFA) came from BIOMOL(Plymouth Meeting, USA). The [methyl-3H]-thymidineand [methyl-3H]-choline were purchased from Amersham(Buckinghamshire, UK). Poly-l-ornithine, genistein (geni),chelerythrine chloride (CC), 1,2, bis (2-aminophenoxy,ethane-N,N,N′,N-tetracetic acid (BAPTA-AM), 5-fluoro-2′-deoxyuridine, glutamine, penicillin and streptomycin wereobtained from Sigma (St. Louis, USA). Trypsin was pur-chased from Worthington (Freehold, USA). All othersreagents were of analytical grade.
2.2. Cell culture
Primary cultures were prepared using procedures previ-ously described (Torres et al., 1998). Briefly, Lister Hoodedneonatal rats at postnatal day 1 (P1) were killed by decap-itation and their eyes removed. The pigmented epitheliumand attached tissues were separated from the neural retinawith fine forceps in a calcium- and magnesium-free balancedsalt solution (CMF) containing 100�g/ml streptomycin+100 U/ml penicillin. The retinae were incubated in CMFcontaining 0.1% trypsin for approximately 16 min at 37◦C.Cells were mechanically dissociated using a polished Pasteurpipette and added to plastic Petri dishes (35 mm) at a platingdensity of 106 cells per dish in 199 culture medium sup-plemented with 2 mM glutamine, 100�g/ml streptomycin+100 U/ml penicillin and 5% fetal calf serum (FCS). SCM and
the drugs to be tested were added immediately after platingof the cells. In some experiments, culture dishes were pre-viously treated with poly-l-ornithine (50�g/ml). Culturesused in experiments addressing ganglion cells survival wereincubated for 4 h to allow the cells to attach to the coverslips.Then, the culture medium and SCM were added. Cultureswere kept for different periods of incubation at 37◦C, in anatmosphere of 5% CO2/95% air.
All procedures using animals were performed accordingto the guidelines of the Society of Neuroscience.
2.3. SCM
SCM was obtained from adult Lister Hooded rats us-ing a method developed in our laboratory (Torres et al.,1998). Rats were killed by ether asphyxia and piecesof the sciatic nerves (approximately 2 cm) were dis-sected and placed in CMF solution containing 200�g/mlstreptomycin+ 200 U/ml penicillin. Nerves were minced in2 mm fragments and incubated in a 35 mm culture dish with2 ml of complete 199 culture medium at 37◦C, in an atmo-sphere of 5% CO2/95% air. The medium was completelychanged twice a week. Supernatants thus obtained after 1, 4,7 and 14 days in vitro were sterilized by filtration through amembrane of 0.2�m and stored at 4◦C. Cells were seededin 1 ml of medium 199 and immediately thereafter 1 ml ofSCM was added to the Petri dish (SCM 1:1).
2.4. SCM adsorption
For SCM adsorption experiments, the dishes were pre-treated as follows: (1) some dishes did not receive any treat-ment before culturing. (2) In another group, the dishes werepretreated with 1 ml of complete 199 culture medium for2 h. (3) A third group was pretreated with 1 ml of SCM for2 h. After this period, 199 medium or SCM was completelywithdrawn and the procedures thereafter were the same asfor the other experiments.
2.5. Protein assay
After maintenance of retinal cells in culture for selectedtime intervals, the protein content of the cultures was de-termined according to the method of Lowry (Lowry et al.,1951), using bovine serum albumin as standard.
2.6. Morphological analysis and quantificationof processes extension
The morphological changes induced by SCM on reti-nal cells were visualized and photographed in a Zeiss mi-croscope under phase contrast illumination. The numberof cells with processes smaller or longer than the diame-ter of the soma was evaluated in 10 fields of each dish.In these experiments, the plating density was 250.000 cellsper dish.

P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20 13
2.7. Proliferation assay
Incorporation of [3H]-thymidine into cell DNA was mea-sured in mixed cultures maintained in a density of 106 cellsper dish. Briefly, 0.5�Ci [3H]-thymidine (specific activity:47.0 Ci mmol−1) was incubated for 1 h at 37◦C in an atmo-sphere of 5% CO2/95% air. The DNA of monolayer cul-tures was precipitated with 0.4 M NaOH, dissolved in 5%trichloroacetic acid and radioactivity incorporated measuredby standard scintillation counting.
2.8. [3H ]-choline uptake
Choline uptake was evaluated by incubating thecultures with 0.2�Ci [3H]-choline (specific activity:50.0 Ci mmol−1) dissolved in 1 ml of Hank’s balanced saltsolution supplemented with 4 mM glucose (pH 7.4) for45 min at 37◦C. After incubation, the cultures were rapidlywashed with fresh incubation medium and the cells lysedwith 1 ml of H2O. The radioactivity was measured by liquidscintillation counting.
2.9. Quantification of retinal ganglion cells survival
Neonatal Lister Hooded rats were anaesthetized by hy-pothermia within the first 24 h after birth and 1.5�l of a30% horseradish peroxidase (HRP) solution in 2% dimethyl-sulphoxide (DMSO) was injected into each superior collicu-lus. After surgery, the animals survived for∼ 16 h beforethe procedures used for cell culture. This time period wasnecessary to allow for the uptake and retrograde transportof HRP to the soma of retinal ganglion cells. The pres-ence of HRP in the cytoplasm of retinal ganglion cells wasrevealed according to the protocol of Mesulam (Mesulan,1982). Briefly, the monolayers were fixed after 48 h in vitrowith a mixture of 1% paraformaldehyde and 2% glutaralde-hyde in 0.1 M sodium phosphate buffer for 5 min, washedin phosphate buffer and then reacted with tetramethylben-zidine. The number of retinal ganglion cells on each cover-slip was evaluated by counting 1/20 of the total area using aZeiss microscope at a magnification of 1000×, under brightfield. As an internal control for the variable percentage ofganglion cells labeled in distinct experiments, the numberof labeled cells at 4 h in culture was taken as 100% and theresults were reported as percentage of control.
2.10. Presentation of data and statistical analysis
All data are expressed as mean± S.E. of the mean fromexperiments performed in duplicate or triplicate that wererepeated at least twice. Statistical analysis of the resultswere performed using the one-way analysis of variance(ANOVA), and then all pairs of multiple groups of datawere assessed by Newman–Keuls comparison test.
Fig. 1. Effect of SCM obtained after different time of incubation (1, 4,7 and 14 days) on retinal cells kept in culture for 24, 48 and 72 h. CT:cultures incubated with control medium. Cells were plated at density of106 cells per dish previously treated with poly-l-ornithine (50�g/ml).Values of protein amount are expressed in�g of protein per dish andrepresent the mean± SEM (n = 3).
3. Results
In a first series of experiments, we analyzed the effect ofSCM on the protein content of the cultures. Fig. 1 showsthat protein level of control cultures increased by two-foldin the first 48 h of incubation and remained constant until72 h after the start of the cultures. Treatment of the cellswith SCM 1d did not change the protein content of thecultures. However, addition of SCM 4d, SCM 7d and SCM14d) greatly stimulated the cells and increased the proteinamount of the cultures. The greatest effect was observed withSCM 14d. After 48 and 72 h of stimulation, this treatmentelicited a three-fold increase in protein amount if comparedto control cultures. SCM 4d and SCM 7d also induced asimilar effect and a two-fold increase on protein amount wasobserved after 48–72 h.
The morphology of the cells treated with SCM 14d for72 h is shown in Fig. 2. We can notice the presence of largeclusters formed by glial cells firmly attached to the Petri dishwith neuronal cells lying above the glial cells (Fig. 2B). Onthe other hand, in control cultures retinal cells formed verysmall clusters and only a few cells with neuronal morphol-ogy can be identified (Fig. 2A). These observations stronglysuggest that SCM 14d induced an increase in neurite out-growth, neuronal survival and glial cell proliferation. Basedon these results, we decided to use only SCM 14d in theensuing experiments.
The morphology of cultures treated with SCM showedan increase in the number of cells with long processes.Fig. 3 illustrates the quantification of these results. In theselow-density cultures, the total number of cells in control cul-tures and in cultures treated with SCM did not change after

14 P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20
Fig. 2. Phase contrast photomicrographs showing the morphology of cultures after 72 h in vitro. (A) Control cultures (CT); (B): cultures treated withSCM obtained after incubation for 14 days. Cells were plated at density of 106 cells per dish previously treated with poly-l-ornithine (50�g/ml). Notethe presence of large clusters (asterisk) of neuronal cells (arrowhead) over glial cells (arrow) in cultures stimulated by SCM. Magnification bar: 50�m.
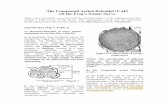
48 and 72 h (Fig. 3A). However, after treatment with SCMfor 48 h, there was an increase of∼ 50% in the number ofprocesses longer than soma after treatment with SCM for48 h (Fig. 3C). After 72 h of treatment with SCM, we ob-served a significant increase (∼160%) in the number of pro-cesses smaller than soma (Fig. 3B) and an increase of 190%in the number of cells presenting processes longer than soma,always in comparison with control cultures (Fig. 3C).
Some morphological changes induced by SCM, like cy-toplasmic processes outgrowth and formation of large clus-ters of cells (Fig. 2), suggest a possible effect of SCM oncell adhesion. In order to investigate that supposition, reti-nal cells were kept on untreated dishes in the presence ofSCM for several days. Fig. 4 illustrates the result of theseexperiments as evaluated by protein assay. We can observethat cultures treated with SCM 14d showed an increase inprotein amount over all the time studied. It is important tonotice that even in cultures with only 4 h in vitro, the effectof SCM was already observed. Based on these results, we
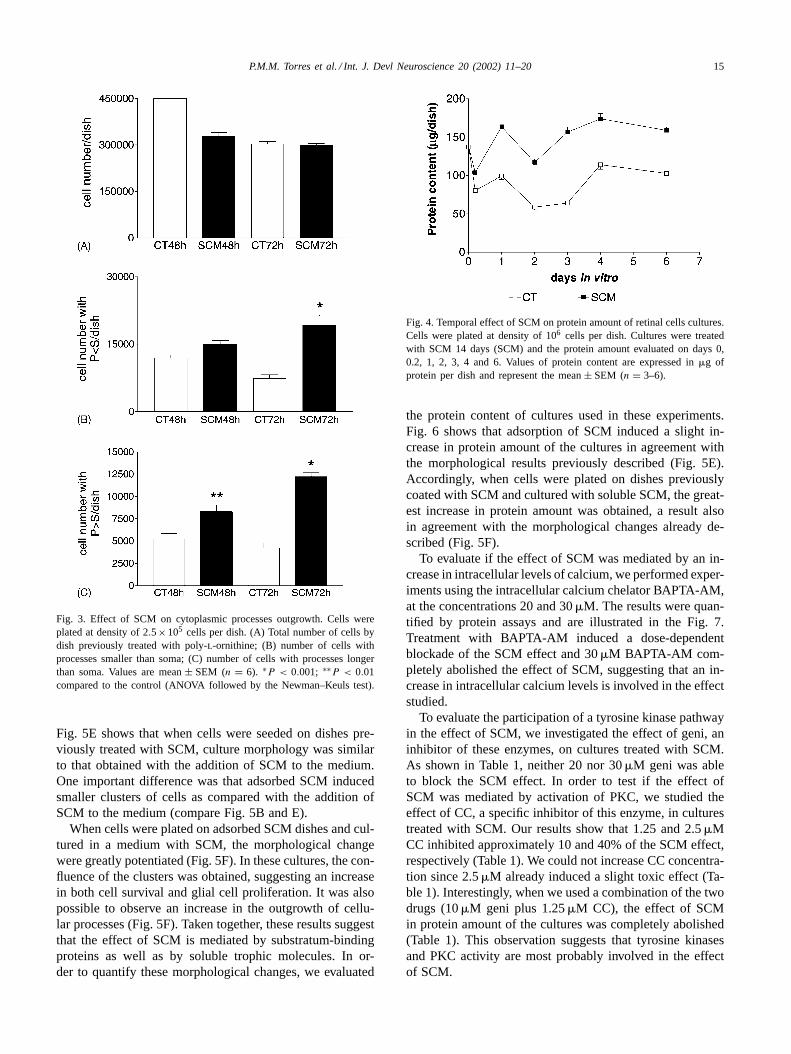
decided to evaluate whether only adhesion molecules medi-ated the SCM effect. Hence, we investigated the effect on themorphology of retinal cells in culture of SCM pre-adsorbedon culture dishes. The result is shown in Fig. 5. In controlcultures cells were seeded on untreated dishes. In anothergroup, cells were seeded on dishes previously treated for 2 h,either with complete medium or with SCM (1:1 dilution).Fig. 5A shows the morphology of control cultures whereonly small clusters of cells could be observed. The additionof SCM to the culture medium induced the formation oflarge and scattered clusters of cells (Fig. 5B). These clusterswere formed by glial cells firmly attached to the Petri dishas well as neuronal cells (dark cells lying on top of them).It is easy to pinpoint neurons in these clusters (Fig. 5B).When cells were seeded on dishes previously treated withcomplete medium, the cultures showed the same morpho-logical features observed in control cultures (Fig. 5C). Ascan be observed on Fig. 5D, the pre-adsorption of disheswith complete medium did not change the effect of SCM.
P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20 15
Fig. 3. Effect of SCM on cytoplasmic processes outgrowth. Cells wereplated at density of 2.5×105 cells per dish. (A) Total number of cells bydish previously treated with poly-l-ornithine; (B) number of cells withprocesses smaller than soma; (C) number of cells with processes longerthan soma. Values are mean± SEM (n = 6). ∗P < 0.001; ∗∗P < 0.01compared to the control (ANOVA followed by the Newman–Keuls test).
Fig. 5E shows that when cells were seeded on dishes pre-viously treated with SCM, culture morphology was similarto that obtained with the addition of SCM to the medium.One important difference was that adsorbed SCM inducedsmaller clusters of cells as compared with the addition ofSCM to the medium (compare Fig. 5B and E).
When cells were plated on adsorbed SCM dishes and cul-tured in a medium with SCM, the morphological changewere greatly potentiated (Fig. 5F). In these cultures, the con-fluence of the clusters was obtained, suggesting an increasein both cell survival and glial cell proliferation. It was alsopossible to observe an increase in the outgrowth of cellu-lar processes (Fig. 5F). Taken together, these results suggestthat the effect of SCM is mediated by substratum-bindingproteins as well as by soluble trophic molecules. In or-der to quantify these morphological changes, we evaluated
Fig. 4. Temporal effect of SCM on protein amount of retinal cells cultures.Cells were plated at density of 106 cells per dish. Cultures were treatedwith SCM 14 days (SCM) and the protein amount evaluated on days 0,0.2, 1, 2, 3, 4 and 6. Values of protein content are expressed in�g ofprotein per dish and represent the mean± SEM (n = 3–6).
the protein content of cultures used in these experiments.Fig. 6 shows that adsorption of SCM induced a slight in-crease in protein amount of the cultures in agreement withthe morphological results previously described (Fig. 5E).Accordingly, when cells were plated on dishes previouslycoated with SCM and cultured with soluble SCM, the great-est increase in protein amount was obtained, a result alsoin agreement with the morphological changes already de-scribed (Fig. 5F).
To evaluate if the effect of SCM was mediated by an in-crease in intracellular levels of calcium, we performed exper-iments using the intracellular calcium chelator BAPTA-AM,at the concentrations 20 and 30�M. The results were quan-tified by protein assays and are illustrated in the Fig. 7.Treatment with BAPTA-AM induced a dose-dependentblockade of the SCM effect and 30�M BAPTA-AM com-pletely abolished the effect of SCM, suggesting that an in-crease in intracellular calcium levels is involved in the effectstudied.
To evaluate the participation of a tyrosine kinase pathwayin the effect of SCM, we investigated the effect of geni, aninhibitor of these enzymes, on cultures treated with SCM.As shown in Table 1, neither 20 nor 30�M geni was ableto block the SCM effect. In order to test if the effect ofSCM was mediated by activation of PKC, we studied theeffect of CC, a specific inhibitor of this enzyme, in culturestreated with SCM. Our results show that 1.25 and 2.5�MCC inhibited approximately 10 and 40% of the SCM effect,respectively (Table 1). We could not increase CC concentra-tion since 2.5�M already induced a slight toxic effect (Ta-ble 1). Interestingly, when we used a combination of the twodrugs (10�M geni plus 1.25�M CC), the effect of SCMin protein amount of the cultures was completely abolished(Table 1). This observation suggests that tyrosine kinasesand PKC activity are most probably involved in the effectof SCM.
16 P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20
Fig. 5. Phase contrast photomicrographs showing the effect of pre-adsorption of SCM in cultures maintained for 48 h in vitro. (A) Control; (B) SCM;(C) pre-adsorbed 199 plus soluble 199; (D) pre-adsorbed 199 plus soluble SCM; (E) pre-adsorbed SCM plus soluble 199; (F) pre-adsorbed SCM plussoluble SCM. Magnification bar: 50�m.
To evaluate if the release of polypeptides in the culturemedium was involved in the effect of SCM, we used BFA,an inhibitor of polypeptides release. Our experiment showedthat 30 nM BFA was unable to block the effect of SCM(data not shown). This result indicates that the moleculespresent in SCM are directly inducing the effects observedin retinal cells.
In our cultures, an effect of SCM on the proliferation ofglial cells was evident (see Figs. 2 and 5). We then inves-tigated the effect of an inhibitor of cellular division, fluo-rodeoxyuridine, on the cultures treated with SCM. As shownin Fig. 8, 15�M fluorodeoxyuridine was able to completely
block the effect of SCM, suggesting that SCM stimulatesglial cells increasing their proliferation.
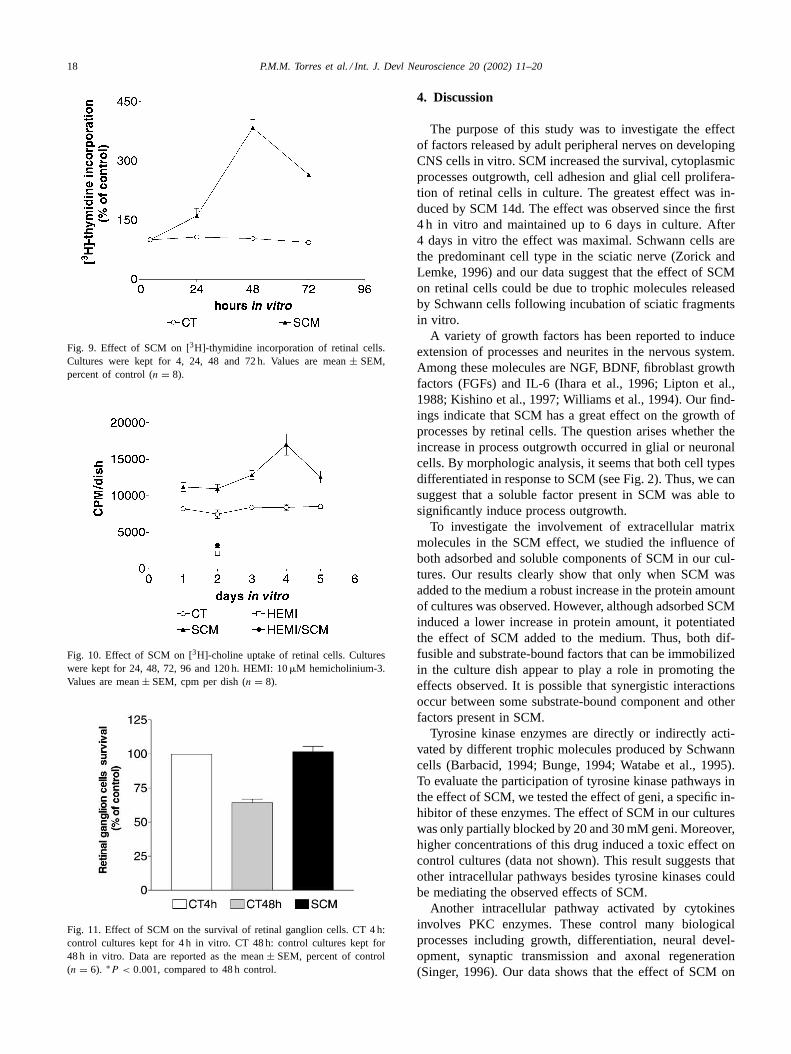
To quantify the proliferation of cells in the presenceof SCM, we measured the [3H]-thymidine incorporationto DNA in cultures after 4, 24, 48 and 72 h in vitro. Theresults of these experiments are shown in Fig. 9. Therewas an increase of 60.62% ± 4.42 in the incorporationof [3H]-thymidine in cultures treated with SCM for 24 hcompared with control cultures. Cultures treated with SCMafter 48 and 72 h revealed an increase of 284.12%± 7.45and 164.87% ± 5.88, respectively. These results indicatethat SCM increases the proliferation of retinal cells an

P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20 17
Fig. 6. The effect of pre-adsorption of SCM on protein amount. Retinalcells were cultured for 48 h in vitro. Data are reported as the mean± SEM,as a percent of control (n = 7–8). ∗P < 0.001 (ANOVA followed by theNewman–Keuls test). The protein content of control cultures was∼= 50�gper dish.
increase that is most evident after 48 h of incubation withSCM.
To investigate if SCM was able to influence the cholin-ergic system of retina, we measured the [3H]-cholineuptake in retinal cells kept in culture. The pattern ofcholinergic differentiation in our cultures was elicited by ahemicholinium-3-sensitive high-affinity choline transport.Fig. 10 shows that treatment with SCM induced a great in-crease on [3H]-choline uptake by retinal cells. We can noticethat the effect took place in the first 24 h. The maximal up-take was obtained after 96 h. The specificity of [3H]-cholinetransport was evaluated by measuring the uptake in culturesincubated with 10�M hemicholinium-3. This compoundwas able to completely block the [3H]-choline uptake incontrol cultures as well as in cultures treated with SCM.
Fig. 7. Influence of intracellular calcium level in the effect of SCM. Cellswere treated with 20 or 30�M BAPTA-AM and cultured for 48 h invitro. Values of protein assay are reported as the mean± SEM, percentof control (n = 5–7). ∗P < 0.001 compared to the control (ANOVAfollowed by the Newman–Keuls test). The protein content of controlcultures was∼= 50�g per dish.
Table 1Effect of SCM in the presence of inhibitors of tyrosine kinase and PKC
Treatment Protein assay (proteincontent (�g/dish), (%)of control ± SEM)
CT 98.7± 6.3 (n = 13)SCM 184.5± 6.3 (n = 13)∗20�M geni 86.5± 3.4 (n = 8)30�M geni 77.6± 7.4 (n = 8)SCM + 20�M geni 155.1± 15.1 (n = 8)∗SCM + 30�M geni 133.9± 9.9 (n = 8)∗∗1.25�M CC 96.5± 4.8 (n =13)2.5�M CC 87.2± 12.5 (n = 9)SCM + 1.25�M CC 165.0± 6.0 (n =13)∗SCM + 2.5�M CC 122.6± 5.3 (n = 9)∗∗10�M geni + 1.25�M CC 92.1± 5.9 (n = 9)SCM + 10�M geni + 1.25�M CC 115.1± 10.0 (n = 9)
The cultures were kept for 48 h in vitro. Values of protein assay areindicated by the mean± SEM, percent of control. The protein content ofcontrol cultures was∼= 50�g per Petri dish.
∗ P < 0.001 (compared to the control (ANOVA followed by theNewman–Keuls test)).
∗∗ P < 0.01.
This result indicates that SCM can influence the cholinergicdifferentiation of retinal cultures.
In our cultures, an effect of SCM on the survival of retinalcells was evident. The cultures treated with SCM showed ahigher number of glial and neuronal cells (see Fig. 2). Then,we decided to evaluate the effect of SCM on the survivalof ganglion cells. Fig. 11 shows that SCM increased thesurvival of these neurons. After 48 h, the number of ganglioncells in control cultures was reduced to∼50% of the initialnumber. However, in cultures treated with SCM, virtuallyall ganglion cells initially plated were kept alive. This resultindicates that SCM was able to support the survival of retinalganglion cells.
Fig. 8. Blockade of the increase in protein content induced by SCMby fluorodeoxyuridine. The cultures were kept for 48 h in vitro. Valuesof protein assay are reported as the mean± SEM, percent of control(n = 7). ∗P < 0.001 compared to the control (ANOVA followed by theNewman–Keuls test). The protein content of control cultures was∼= 50�gper dish.
18 P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20
Fig. 9. Effect of SCM on [3H]-thymidine incorporation of retinal cells.Cultures were kept for 4, 24, 48 and 72 h. Values are mean± SEM,percent of control (n = 8).
Fig. 10. Effect of SCM on [3H]-choline uptake of retinal cells. Cultureswere kept for 24, 48, 72, 96 and 120 h. HEMI: 10�M hemicholinium-3.Values are mean± SEM, cpm per dish (n = 8).
Fig. 11. Effect of SCM on the survival of retinal ganglion cells. CT 4 h:control cultures kept for 4 h in vitro. CT 48 h: control cultures kept for48 h in vitro. Data are reported as the mean± SEM, percent of control(n = 6). ∗P < 0.001, compared to 48 h control.
4. Discussion
The purpose of this study was to investigate the effectof factors released by adult peripheral nerves on developingCNS cells in vitro. SCM increased the survival, cytoplasmicprocesses outgrowth, cell adhesion and glial cell prolifera-tion of retinal cells in culture. The greatest effect was in-duced by SCM 14d. The effect was observed since the first4 h in vitro and maintained up to 6 days in culture. After4 days in vitro the effect was maximal. Schwann cells arethe predominant cell type in the sciatic nerve (Zorick andLemke, 1996) and our data suggest that the effect of SCMon retinal cells could be due to trophic molecules releasedby Schwann cells following incubation of sciatic fragmentsin vitro.
A variety of growth factors has been reported to induceextension of processes and neurites in the nervous system.Among these molecules are NGF, BDNF, fibroblast growthfactors (FGFs) and IL-6 (Ihara et al., 1996; Lipton et al.,1988; Kishino et al., 1997; Williams et al., 1994). Our find-ings indicate that SCM has a great effect on the growth ofprocesses by retinal cells. The question arises whether theincrease in process outgrowth occurred in glial or neuronalcells. By morphologic analysis, it seems that both cell typesdifferentiated in response to SCM (see Fig. 2). Thus, we cansuggest that a soluble factor present in SCM was able tosignificantly induce process outgrowth.
To investigate the involvement of extracellular matrixmolecules in the SCM effect, we studied the influence ofboth adsorbed and soluble components of SCM in our cul-tures. Our results clearly show that only when SCM wasadded to the medium a robust increase in the protein amountof cultures was observed. However, although adsorbed SCMinduced a lower increase in protein amount, it potentiatedthe effect of SCM added to the medium. Thus, both dif-fusible and substrate-bound factors that can be immobilizedin the culture dish appear to play a role in promoting theeffects observed. It is possible that synergistic interactionsoccur between some substrate-bound component and otherfactors present in SCM.
Tyrosine kinase enzymes are directly or indirectly acti-vated by different trophic molecules produced by Schwanncells (Barbacid, 1994; Bunge, 1994; Watabe et al., 1995).To evaluate the participation of tyrosine kinase pathways inthe effect of SCM, we tested the effect of geni, a specific in-hibitor of these enzymes. The effect of SCM in our cultureswas only partially blocked by 20 and 30 mM geni. Moreover,higher concentrations of this drug induced a toxic effect oncontrol cultures (data not shown). This result suggests thatother intracellular pathways besides tyrosine kinases couldbe mediating the observed effects of SCM.
Another intracellular pathway activated by cytokinesinvolves PKC enzymes. These control many biologicalprocesses including growth, differentiation, neural devel-opment, synaptic transmission and axonal regeneration(Singer, 1996). Our data shows that the effect of SCM on

P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20 19
the retinal cells is inhibited by 2.5 mM CC, suggestingthat a PKC transduction pathway is involved. Interestingly,when we used a combination of the two last drugs (10�Mgeni plus 1.25�M CC), the effect of SCM was completelyabolished. This observation suggests that tyrosine kinasesand PKC activity are most probably involved in the effectof SCM. However, a complete understanding of the mech-anisms by which SCM activates these pathways in retinalcells is not yet available.
Calcium is an important intracellular messenger in awide variety of cellular responses, including secretionof molecules and cell proliferation (Berninger and Poo,1996). The effect of SCM was totally blocked by 30 mMBAPTA-AM, an intracellular calcium chelator, suggestingthat SCM induces and increase in intracellular calciumthat in some way is involved in the culture modificationsobserved with SCM. Since activation of several PKC iso-forms is dependent on intracellular calcium mobilization,our results reinforce the possible role of PKC on retinalcell responses to SCM. However, we cannot exclude theinvolvement of other pathways that are also dependent oncalcium mobilization.
One of the most important mediators in the nervous sys-tem organization and plasticity are glial cells. Müller cellsare the principal glial cells in the retina, playing many ofthe roles carried out by astrocytes, oligodendrocytes andependymal cells in other CNS regions. Müller cells recog-nize a variety of neuronal signals, actively control levelsof K+, H+ and neurotransmitters in extracellular space andproduce and release several trophic molecules, thus regulat-ing the retinal microenviroment (Newman and Reichenbach,1996). In our cultures, most proliferative cells are repre-sented by glial cells. In our cultures, we observed an effectof SCM on glial cell proliferation, as analyzed by morpho-logical criteria (Fig. 2) and by [3H]-thymidine incorpora-tion (Fig. 9). Our present data show that addition of 15 mMfluorodeoxiuridine, an inhibitor of cellular division, com-pletely blocked the effect of SCM on retinal cells. This re-sult suggests that proliferation of glial cells is involved inthe SCM effects. The [3H]-thymidine incorporation assayrevealed that SCM induced a great effect on the prolifera-tion of retinal cells. Glial cells in our cultures, possibly areproviding the adequate substratum to neuronal adhesion aswell as the trophic support for neuronal survival and neuriteoutgrowth. Thus, the induction of glial cells proliferation bySCM could be the first step in the retinal cells response toSCM.
Besides their function in intercellular communication,trophic effects mediated by neurotransmitters have beenpostulated in the mammalian central nervous system (Drey-fus, 1998). Then, we decided to investigate possible ef-fects of SCM on the retinal cholinergic cell population.Acetylcholine is well established as the neurotransmitter ofan individual class of neurons in the retina, the starburstamacrine cells (Kolb, 1997). Our results indicate that SCMhas a great effect on the increase in [3H]-choline uptake by
retinal cells. As for other cellular metabolites with insuffi-cient synthesis, choline is taken up from the extracellularmedium by specific transport mechanisms. An ubiquitouslow affinity mechanism commonly thought to promote dif-fusion of choline across the cell membrane provides enoughcholine for phospholipid synthesis in most tissues, but ahigh-affinity choline transport mechanism is found at cholin-ergic nerve terminals that specifically use choline to synthe-size the neurotransmitter, acetylcholine (Kuhar and Murrin,1978). Like the neurotransmitter transporters (Shafqat et al.,1993), energy from the inwardly directed sodium gradientis used by the transporter to concentrate choline inside thenerve terminal. This form of choline transport is particu-larly sensitive to the drug hemicholinium-3 (Guyenet et al.,1973). The effect of SCM on [3H]-choline uptake by retinalcells was totally blocked by hemicholinium-3. This resultindicates that SCM induces cholinergic differentiation ofretinal cells, increasing the synthesis of acetylcholine.
SCM increased the survival of retinal ganglion cells inculture. A question on the nature of molecules that areinducing this effect naturally arises. The differentiation, sur-vival and death of neurons can be modulated by a series ofgrowth factors as well as by neurotransmitters. Moreover,a variety of studies suggest that neurotransmitters and NTsmay interact to regulate the development of neuronal pop-ulations (Dreyfus, 1998). Further experiments will be nec-essary to compare the effect of purified trophic moleculessuch as NGF, BDNF, interleukins, transforming growthfactor � (TGF�), among others, on the effect elicited bySCM in retinal cells. However, the robust effect induced bytreatment with SCM suggests that synergistic interactionsbetween cytokines may be involved in retinal cells responseto SCM. Schwann cells produce several neurotrophic fac-tors that support neuronal survival and differentiation.These include NGF, BDNF, NT-3, CNTF, glial-derivedneurotrophic factor (GDNF), IL-6, tumor necrosis factor�(TNF�), and LIF (Bolin et al., 1995; Be’eri et al., 1998;Bunge, 1994; Kurek et al., 1996; Sendtner et al., 1992;Wagner and Myers, 1996). Thus, a simple approach to iden-tify the trophic molecules responsible for the SCM effectswould be the use of cytokines neutralizing antibodies toblock the response of retinal cells. However, the effect ofother unknown factors produced by Schwann cells can notbe discarded. Combined effects of multiple cytokines fromSCM should also be considered (Torres and Araujo, 2001).
Taken together, these results enables us to suggestthat fragments of sciatic nerves in culture secrete trophicmolecules that have a pleiotropic effect on retinal cells inculture.
Acknowledgements
Technical assistance by Bernardino Matheus dos Santosand Alexandre José Fernandes is gratefully acknowledged.We also wish to thank Ana Lúcia Marques Ventura and Ar-naldo Paes de Andrade for critical reading of the manuscript.

20 P.M.M. Torres et al. / Int. J. Devl Neuroscience 20 (2002) 11–20
This work was supported by CAPES, CEG-UFF, FAPERJand FINEP.
References
Barbacid, M., 1994. The Trk family of neurotrophin receptors. J.Neurobiol. 25, 1386–1403.
Be’eri, H., Reichert, F., Saada, A., Rotshenker, S., 1998. The cytokinenetwork of Wallerian degeneration: IL-10 and GM-CSF. Eur. J.Neurosci. 10, 2707–2713.
Berninger, B., Poo, M., 1996. Fast actions of neurotrophic factors. Cur.Opin. Neurobiol. 6, 324–330.
Bolin, L.M., Verity, A.N., Silver, J.E., Shooter, E.M., Abrams, J.S., 1995.Interleukin-6 production by Schwann cells and induction in sciaticnerve injury. J. Neurochem. 64, 850–858.
Bunge, R.P., 1994. The role of the Schwann cell in trophic support andregeneration. J. Neurol. 241, S19–S21.
Cornbrooks, C.J., Carey, D.J., McDonald, J.A., Timple, R., Bunge, R.P.,1983. In vivo and in vitro observation on laminin production bySchwann cells. PNAS U.S.A. 80, 3850–3854.
Dezawa, M., Nagano, T., 1993. Contacts between regenerating axons andthe Schwann cells of sciatic nerve segments grafted to the optic nerveof adults rats. J. Neurol. 22, 1103–1112.
Dobresov, M., Dobresov, A., Kuffler, D.P., 1994. Influence of factorsreleased from sciatic nerve on adult dorsal root ganglion neurons. J.Neurobiol. 25, 1249–1266.
Dowling, J.E., 1991. Retina. Encyclop. Human Biol. 6, 615–631.Dreyfus, C.F., 1998. Neurotransmitters and neurotrophins collaborate to
influence brain development. Persp. Develop. Neurobiol. 5, 389–399.Guyenet, P., Lefresne, P., Rossier, J., Beaujouan, J.C., Glowinski, J.,
1973. Inhibition by hemicholinium-3 of{14C}acetylcholine synthesisand [3H]choline high-affinity uptake in rat striatal synaptosomes. Mol.Pharmacol. 9, 630–639.
Ihara, S., Iwamatsu, A., Fujiyoshi, T., Komi, A., Yamari, T., Fukui, Y.,1996. Identification of interleukin-6 as a factor that induces neuriteoutgrowth by PC12 cells primed with NGF. J. Biochem. 120, 865–868.
Kishino, A., Ishige, Y., Tatsuno, T., Nakayama, C., Noguchi, H., 1997.BDNF prevents and reverses adult rat motor neuron degeneration andinduces axonal outgrowth. Exp. Neurol. 144, 273–286.
Kolb, H., 1997. Amacrine cells of the mammalian retina: neurocircuitryand functional roles. Eye 11, 904–923.
Korsching, S., 1993. The neurotrophic factor concept: a reexamination.J. Neurosci. 13, 2739–2748.
Kuffler, D.P., Megwinoff, O., 1994. Neurotrophic influence of denervatedsciatic nerve on adult dorsal root ganglion neurons. J. Neurobiol. 25,1267–1282.
Kuhar, M.J., Murrin, L.C., 1978. Sodium-dependent, high-affinity cholineuptake. J. Neurochem. 30, 15–21.
Kurek, J.B., Austin, L., Cheema, S.S., Bartlett, P.F., Murphy, M.,1996. Up-regulation of leukaemia inhibitory factor and interleurkin-6in transected sciatic nerve and muscle following denervation.Neuromuscu. Disord. 6, 105–114.
Lipton, S.A., Wagner, T.A., Madison, R.D., D’Amore, P.A., 1988. Acidicfibroblast growth factor enhances regeneration of processes by postnatal
mammalian retinal ganglion cells in culture. PNAS U.S.A. 85, 2388–2392.
Lowry, O.H., Rosenbrough, N.J., Farr, A.L., Randall, R.J., 1951. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Mesulan, M.M., 1982. Tracing Neural Connections with HorseradishPeroxidase. Wiley, New York, p. 251.
Newman, E., Reichenbach, A., 1996. The Müller cell: a functional elementof the retina. TINS 19, 307–311.
Patterson, P.H., 1993. Cytokines and the function of the mature nervoussystem. Curr. Rev. Acad. Sci. Paris 316, 1150–1157.
Reichert, F., Saada, A., Rotshenker, S., 1994. Peripheral nerve injuryinduces Schwann cells to express two macrophage phenotypes:phagocytosis and the galactose-specific lectin MAC-2. J. Neurosci. 14,3231–3245.
Richardson, P.M., McGuinnes, U.M., Aguayo, A.J., 1980. Axons fromCNS neurones regenerate into PNS grafts. Nature 284, 264–265.
Sendtner, M., Sotöckli, K.A., Thoenen, H., 1992. Synthesis andlocalization of ciliary neurotrophic factor in the sciatic nerve of theadult rat after lesion and during regeneration. J. Cell Biol. 118, 139–148.
Shafqat, S., Velaz-Faircloth, M., Guadano-Ferraz, A., Fremeau, J.,1993. Molecular characterization of neurotransmitter transporters. Mol.Endocrinol. 7, 1517–1529.
Singer, H.A., 1996. Protein kinase C. Biochem. Smooth Muscle Contract.12, 155–165.
Svendsen, C.N., Sofroniew, M.V., 1996. Do central nervous systemneurons require target-derived neurotrophic support for survivalthroughout adult life and aging? Perspect. Dev. Neurobiol. 3, 133–142.
Thoenen, H., 1991. The changing scene of neurotrophic factors. TINS14, 165–170.
Torres, P.M.M., Araujo, E.G., 2001. Interleukin-6 increases the survivalof retinal ganglion cells in vitro. J. Neuroimmun. 117, 43–50.
Torres, P.M.M., Guilarducci, C.V.V., Araujo, E.G., 1998. Effects ofsciatic-conditioned medium on neonatal rat retinal cells in vitro. Braz.J. Med. Biol. Res. 31, 1439–1442.
Tuszynski, M.H., Weidner, N., McCormack, M., Miller, I., Powell, H.,Conner, J., 1998. Grafts of genetically modified Schwann cell to thespinal cord: survival, axon growth, and myelination. Cell Transpl. 7,187–196.
Wagner, R., Myers, R.R., 1996. Schwann cells produce tumornecrosis factor alpha: expression in injured and non-injured nerves.Neuroscience 73, 625–629.
Watabe, K., Fukuda, T., Tanaka, J., Honda, H., Toyohara, K., Sakai, O.,1995. Spontaneously immortalized adult mouse Schwann cell secreteautocrine and paracrine growth-promoting activities. J. Neurosci. Res.41, 279–290.
Weisenhorn, D.M.V., Roback, J., Yong, A.N., Wainer, B.H., 1999. Cellularaspects of trophic actions in the nervous system. Int. Res. Cytol. 189,177–265.
Williams, E.J., Furness, J., Walsh, F.S., Doherty, P., 1994. Characterizationof the second messenger pathway underlying neurite outgrowthstimulated by FGF. Development 120, 1685–1693.
Zorick, T.S., Lemke, G., 1996. Schwann cell differentiation. Curr. Opin.Cell Biol. 8, 870–876.