
Immunohistochemical study of human retinal astroglia
12
Pergamon 004269I39(93)EOO49-D Yision Res. Vol. 34, No. 15, 19351946, pp. 1994 Copyright 0 1994 Elsevier Science Ltd Printed in Great Britain. All rights reserved 0042-6989/94 $7.00 + 0.00 Tmmunohistochemical Study of Human Retinal Astroglia JOSE M. RAMIREZ,* ALBERT0 TRIVINO,* ANA I. RAMIREZ,* JUAN J. SALAZAR,* JULIAN GARCIA-SANCHEZ* Received 22 JuIy 1993; in revised form 21 October 1993 Immunocytochemical localization of glial fibrillary acidic protein (GFAP) has been used to study astrocyte distribution and morphology in whole mouuted humau retinas and vertical sections. Two types of astrocytes can be distlnffuishedz elongated astrocytes are located in the nerve Abre layer (NFL); and star-shaped astroqtes are found in the gauglion cell layer (GCL). Astroglial processes join to form bundles. The bundles formed by the elongated askqtes lie along and separate the nerve fibre bundles. Processes from star-shaped astrocytes reach towards other star-shaped astrocytes and towards the vessels to form a morphologically honeycombed plexus. These astrocytes also send other processes towards the interual nuclear layer (INL), forming an irre@ar plexus which accompanies the GCL capillaries that extend into the INL. Often, the cell bodies of the star-shaped GCL astrocytes lie over vessels and form cell clusters. Finally, none of the retinas examined for this study evidenped the “perivascular astrocytes” described by Wolter in the human retina. Human Retina Glia fibrillary acidic protein Astrocytes Immunohistochemistry 1. INTRODUCTION There are two types of astroglial cells in mammal retinas: Miiller glia, which extend radially across the entire retina; and astrocytes, which are only found in the vascularized areas. Astroglial cells were originally studied using silver techniques (Ramon y Cajal, 1892, 1911; Marchesani, 1926; Polyak, 1941). More recently the use of GFAP has made it possible to label these cells selectively in different animal species (Dahl, Bjijrklund & Bignami, 1986). Anti-GFAP morphological studies of human retinal astrocytes are scarce and most of those that do exist are focused on the study of epiretinal membranes and retinoblastomas (Lane & Klintworth, 1983; Hiscott, Grierson, Trombetta, Rahl, Marshall & McLeod, 1984; Kivell, Tarkkanen & Virtanen, 1986; Hui, Goodnight, Zhang, Sorgente & Ryan, 1988; Sramek, Wallow, Stevens & Nork, 1989; Guerin, Wolfshagen, Eifrig & Anderson, 1990). The most thorough study of human retinal astrocytes was made by Wolter (1955a,b, 1957, 1959, 1961), who used the Scharenberg modification of Del Rio Hortega’s silver carbonate method on horizontal retina sections. He observed several types of astroglial cells that formed a very elaborate net. The cells extended through all of the vascularized areas of the retina and contacted both axons and blood vessels. Wolter also described a special type of astroglial cell, which he called “perivascular astrocyte”, that could only be found surrounding blood vessels. However, later electron microscope studies have not been able to demonstrate ultrastructural differences between the different astrocytes in the human retina and have cast doubt on the existence of perivascular astro- cytes (Ikui, Uga & Kohno, 1976). 1935 Given the controversy on perivascular astrocytes, and the absence of anti-GFAP morphological studies of whole-mounts, the present paper undertakes a morpho- immunohistochemical study of astroglial cells in the human retina that also determines their distribution in the different retinal regions. Thirty-four normal adult human eyes (age range 2@40 yr), enucleated about 2-4 hr post mortem for cornea1 transplantation were obtained from the Spanish Eye Bank for this study. One to two hours after enucle- ation and cornea1 processing, the eyes were fixed with 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS) at pH 7.4 for 4 hr at 4°C. Thirty eyes were processed as retinal whole-mounts and four eyes were sectioned vertically. *Institute de Investigaciones Oftalmol6gicas “Ram6n Castroviejo”, Facultad de Medicina, Pab. VI, 4” Planta, UniversidadCom- plutense,28040 Madrid, Spain. After 15 min fixation, the retina was removed from the eyecup for whole-mounts and then reintroduced into the fixing solution for 4 hr. Afterwards the retinas were washed in PBS for several hours at 4°C. 2. MATERIALSAND METHODS
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Immunohistochemical study of human retinal astroglia

Pergamon 004269I39(93)EOO49-D
Yision Res. Vol. 34, No. 15, 19351946, pp. 1994 Copyright 0 1994 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0042-6989/94 $7.00 + 0.00
Tmmunohistochemical Study of Human Retinal Astroglia JOSE M. RAMIREZ,* ALBERT0 TRIVINO,* ANA I. RAMIREZ,* JUAN J. SALAZAR,* JULIAN GARCIA-SANCHEZ*
Received 22 JuIy 1993; in revised form 21 October 1993
Immunocytochemical localization of glial fibrillary acidic protein (GFAP) has been used to study astrocyte distribution and morphology in whole mouuted humau retinas and vertical sections. Two types of astrocytes can be distlnffuishedz elongated astrocytes are located in the nerve Abre layer (NFL); and star-shaped astroqtes are found in the gauglion cell layer (GCL). Astroglial processes join to form bundles. The bundles formed by the elongated askqtes lie along and separate the nerve fibre bundles. Processes from star-shaped astrocytes reach towards other star-shaped astrocytes and towards the vessels to form a morphologically honeycombed plexus. These astrocytes also send other processes towards the interual nuclear layer (INL), forming an irre@ar plexus which accompanies the GCL capillaries that extend into the INL. Often, the cell bodies of the star-shaped GCL astrocytes lie over vessels and form cell clusters. Finally, none of the retinas examined for this study evidenped the “perivascular astrocytes” described by Wolter in the human retina.
Human Retina Glia fibrillary acidic protein Astrocytes Immunohistochemistry
1. INTRODUCTION
There are two types of astroglial cells in mammal retinas: Miiller glia, which extend radially across the entire retina; and astrocytes, which are only found in the vascularized areas.
Astroglial cells were originally studied using silver techniques (Ramon y Cajal, 1892, 1911; Marchesani, 1926; Polyak, 1941). More recently the use of GFAP has made it possible to label these cells selectively in different animal species (Dahl, Bjijrklund & Bignami, 1986).
Anti-GFAP morphological studies of human retinal astrocytes are scarce and most of those that do exist are focused on the study of epiretinal membranes and retinoblastomas (Lane & Klintworth, 1983; Hiscott, Grierson, Trombetta, Rahl, Marshall & McLeod, 1984; Kivell, Tarkkanen & Virtanen, 1986; Hui, Goodnight, Zhang, Sorgente & Ryan, 1988; Sramek, Wallow, Stevens & Nork, 1989; Guerin, Wolfshagen, Eifrig & Anderson, 1990).
The most thorough study of human retinal astrocytes was made by Wolter (1955a,b, 1957, 1959, 1961), who used the Scharenberg modification of Del Rio Hortega’s silver carbonate method on horizontal retina sections. He observed several types of astroglial cells that formed a very elaborate net. The cells extended through all of the vascularized areas of the retina and contacted both axons and blood vessels. Wolter also described a special type of astroglial cell, which he called “perivascular astrocyte”, that could only be found surrounding blood vessels. However, later electron microscope studies have not been able to demonstrate ultrastructural differences between the different astrocytes in the human retina and have cast doubt on the existence of perivascular astro- cytes (Ikui, Uga & Kohno, 1976).
1935
Given the controversy on perivascular astrocytes, and the absence of anti-GFAP morphological studies of whole-mounts, the present paper undertakes a morpho- immunohistochemical study of astroglial cells in the human retina that also determines their distribution in the different retinal regions.
Thirty-four normal adult human eyes (age range 2@40 yr), enucleated about 2-4 hr post mortem for cornea1 transplantation were obtained from the Spanish Eye Bank for this study. One to two hours after enucle- ation and cornea1 processing, the eyes were fixed with 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS) at pH 7.4 for 4 hr at 4°C. Thirty eyes were processed as retinal whole-mounts and four eyes were sectioned vertically.
*Institute de Investigaciones Oftalmol6gicas “Ram6n Castroviejo”, Facultad de Medicina, Pab. VI, 4” Planta, Universidad Com- plutense, 28040 Madrid, Spain.
After 15 min fixation, the retina was removed from the eyecup for whole-mounts and then reintroduced into the fixing solution for 4 hr. Afterwards the retinas were washed in PBS for several hours at 4°C.
2. MATERIALS AND METHODS

1936 JOSE M. RAMIREZ ct al.
FIGURE I. Vertical retina section showing GFAP( +) fundamentally within the NFL and the GCL; some capillaries reach the INL surrounded by astroglial processes (arrow) I*, vitreo; ONL, outer nuclear layer; NNL, internal nuclear layer; GCL,
ganglion cell layer; NFL, nerve fiber layer; 250x).
The posterior segment was separated from the eyes to prepare vertical sections. The tissue was dehydrated, embedded in paraffin and cut into 6 and 20 pm sections.
2.1. r~~unocytoche~~cal procedure
The peroxidase antiperoxidase method was used with the following procedure for the whole-mounts. The retinas were pretreated with 0.3% hydrogen peroxide in PBS for 30 min at room temperature before washing in PBS (3 x 10 min). They were then incubated for 24 hr at 4°C in 10% normal goat serum (NGS) (Sigma, U.S.A.) and 0.2% Triton X-100 (Merck, Germany) diluted in PBS. Whole retinas were incubated in monocional mouse antibody directed against giial fibrillary acidic protein (GFAP clone GA-5) (Biomakor, Israel) in a l/250 dilution at 4°C for 34 days and then washed in
PBS (3 x 3 hr). The retinas were incubated at 4°C for 2 days with the immunoglobulin fraction from goat anti- mouse serum (Sigma) diluted l/100, and then washed in PBS (3 x 3 hr), After this, they were incubated at 4°C for 24 hr with mouse peroxidase antiperoxidase complex (PAP) (Sigma) diluted l/700, and rinsed in PBS (3 x 3 hr). The whole retinas were treated with 0.03% DAB (diamino~~dine tetrahydrochl~de~ (Sigma) in PBS for 5 min, followed by 0.03% ~AB~O.~l% hydro- gen peroxide in PBS for an additional 7 min. After final rinsing in PBS (3 x lOmin), the retinas were flat- mounted and fixed onto gelatin chrome-alum subbed slides, dehydrated in ascending alcohols, cleared in xylene and mounted in Canada balsam (Merck).
In addition to PAP technique, indirect immuno- fluorescence was also used. In this technique the
FIGURE 2 (facing page). Morphology and distribution of the anti-GFAP labelled retinal astrocytes. (A, 8) Star-shaped astrocyte (arrow) located within a honeycomb in the GCL [(A) 250 x , *blood vessels; (B) 500 x]; (C, D) Astrocyte clusters (arrow) located on a vessel wall in the GCL [(C) 400 x ; (D) 200 x *blood vessels]; (E) star-shaped astrocyte (arrow) forming part of a GCL astroglial bundle (*b&d vessels, 400 x ); (F) star-shaped astrocytes of the horizontal raphe in the NFL, these c&Is form bundles that lie concentric&y around the fovea (the fovea is located OSmm from the lower right edge of the micrograph (125 x ); elongated astrocyte with some processes reaching into the GCL (arrowhead) and others running parallel to NFL axons (arrow) (400 x ); (H) elongated astrocyte sending a process towards a nearby NFL astroglial bundle (arrowhead) (400 x ). [Whole mount preparations: (B) immunofluorescence; (A.C. D, E, F, G, H) PAP; (C, E, F, G, H) Nomarski Optics.]
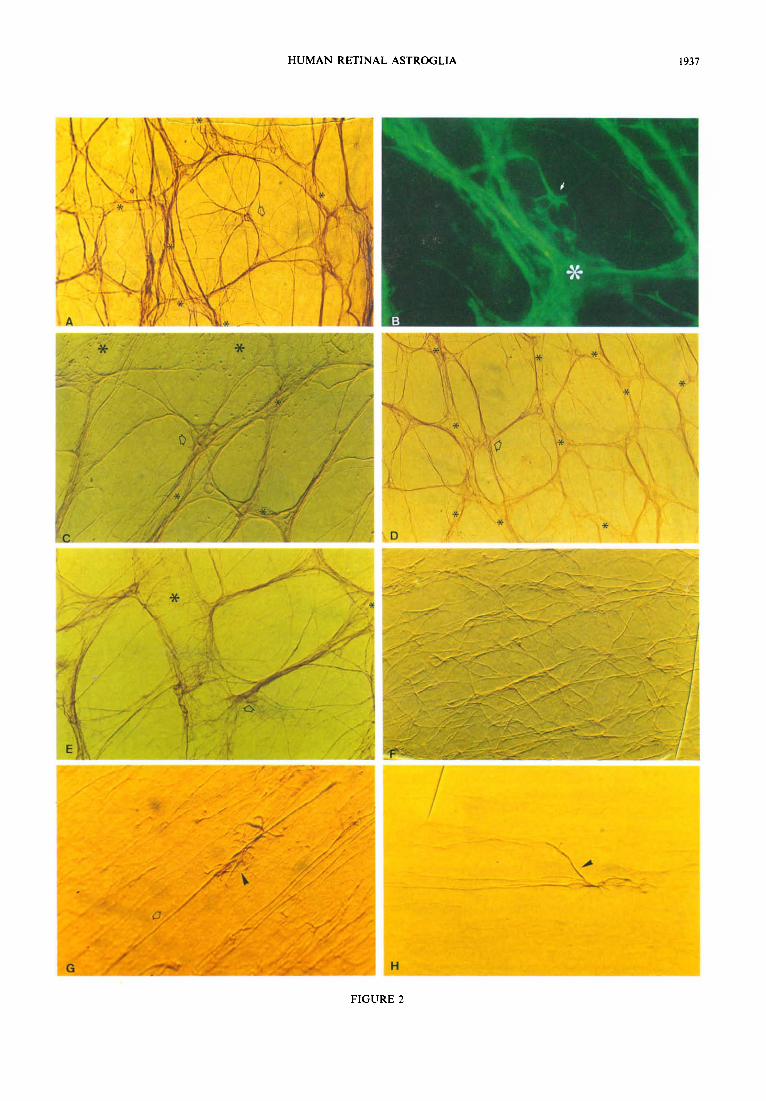
HUMAN RETINAL ASTROGLIA 1937
FIGURE 2

1938 JOSE M. RAMIREZ ef al.
GFAP antiserum (diluted l/l 50) was followed by goat antimouse antibodies conjugated to fluorescein- isothiocyanate (FITC) (Sigma) diluted 1 /SO for 24 hr. In this case, retinas were mounted without dehydration in Citifluor (Agar, U.K.).
The procedure for vertical sections was basically the same except for the incubation and washing times: 1 hr in normal goat serum, 4 hr in anti-GFAP, 2 hr in goat antimouse serum, 2 hr in PAP complex, 2 hr in FITC- conjugated antimouse goat serum, and three 15 min washes.
All antibody dilutions were carried out in 0.2% Triton X-100, 1% NGS in O.lM PBS at pH 7.4. Controls were treated the same as above except that the incubation in the primary antibody solution was eliminated to demon- strate that the secondary antibodies reacted only with their respective primary antibody.
3. RESULTS
3.1. Location of the retinal astrocytes
The distribution of the GFAP( +) astrocytes co- incided with that of retinal vessels in the whole-mount retinas. These cells coursed alongside the blood vessels from the optic disk to the capilla;es proximal to the ora serrata and were found throughout the retina with the exception of a zone about 0.4 mm in diameter in the centre of the fovea (the only avascular area).
On vertical retina sections the GFAP( +) astrocytes were mostly located in the ganglion cell layer (GCL) and nerve fibre layer (NFL); however, some processes and a few cell bodies accompanied the capillaries as far as the internal nuclear layer (INL) (Fig. 1).
Like the blood vessels, most of the astroglial cells were found in the areas close to the optic disk; the number of astrocytes decreased towards the periphery, while, in the avascular zone of the fovea, they abruptly disappeared.
3.2. Astrocyte morphology
Depending on the differing distribution of the cell processes, human retinal astrocytes can be classified as elongated or starshaped.
3.2.1. Star-shaped astrocytes. These cells show high GFAP( +) immunoreactivity. The oval cell body is small and has a prominent, ovoid nucleus. Normally, 4-8 primary processes extend out from the cell body and
divide into thinner, secondary processes [Fig. 2(A,B)]. On occasions the latter extend into the retinal par- enchyma alongside the blood vessels that reach the INL.
The astrocyte processes extending radially out from the cell body give it a stellate appearance. These pro- cesses, which can be very long, reach other astrocytes or blood vessels and form an astroghal plexus with a honeycomb morphology [Fig. 2(A, B)].
The cell body is generally located over or in the proximity of blood vessels [Fig. 2(A,B,C,D,E)]. This type of astrocyte can be observed in the GCL and is distributed in this layer throughout the retina. Some star-shaped astrocytes were observed in the NFL in the region of the horizontal raphe temporal to fovea along the horizontal meridian [Fig. 2(F)].
3.2.2. Elongated astrocytes. These cells also show high GFAP( + ) immunoreactivity . Their small, oval-shaped cell body has an ovoid nucleus. Two to four very long primary processes extend straight out from the two poles of the cell body parallel to the axons to join other elongated astrocytc procuses in the NFL {Fig. Z(G,H)].
These cells also have other probes that may reach through the NFL as far as the GCL [r;ig. 2(G)]. Yet other processes may extond perpendicubeity to the axons towards other nearby astrocyte processes [Fig. 2(H)].
The cell body is located in the NFL between the axonal bundles and the radial @pap&try capillaries [described by Michaelson and Campbell (1940) and Henkind (1967)]. The processes separate the axon bun- dles from each other and come close enough to touch the capillaries.
3.3. Retinal astrocyte distribution
The astrocyte processes tend to join together to form astroglial bundles. The distritmtion of these bundles varies depending on the retinal layer in which they are found.
3.3.1. Ganglion cell layer astrogfiaf bwwk. The pro- cesses from the star-shaped astrocytes in the GCL form bundles that are regularly distributed throughout the retina in a honeycombed pkxus that surrounds the blood vessels [Fig. 3(A)]. Usually, the bodies of the ganglion cells lie within the honeycomb chambers.
The processes that form the bundles accompany the different blood vessels and follow a trajectory that is
FIGURE 3 (/acing page). Morphology and distribution of the anti-GFAP labelkd retinal astrocytea. (A) GCL astroghal
bundles with a honeycomb morphology (*blood vessels, 125 x ); (B) formation of the hvmb stntdmon~avcwL the bundles fan out, some turning 90 deg to form part of the vascular astroghal net (urrowa), rrad thn rertpiaiss otbetproarse~
to form a new astroglial bundle (arrowhead) (*blood vessels, 300 x ). [(A, B) Whok-mormt pr@ma&~, PAP, ~Nomarski Optics.]
FIGURE 5 (facing page). Morphology and distribution of the anti-GFAP lab&d retinal aetrocytee. (A) Lo&udinal
astroghal bundles within the NFL (250 x ). (B) Vertical retina section which shows the pamILd asnay of the straight -tad astrocytes in the NFL (arrows) and the honeycomb lay-out of the GCL star-shaped a~mcy&~ (#rrow~) ti v&po; *Mood
vessel; 250 x ). (C) Retinal whole-mount showing the progressive disappearance of the NFL astroghal bundles (arrows); the
micrograph shows an area inferior to the optic disk (the optic disk is located 6 mm beyond the right of the arrows) (32 x ).
(D) Star-shaped astrocytc located on the boundary between the avascular and vascular zones of the fovea (the arrow marks
the astrocyte nucleus: upper part. vascular zone; lower part. avascular zone: 500 x ). [(A.C.D) Whole-mount preparations. PAP; (B) vertical sections. immunofluorescence: (A) Nomarski Optics.]

HUMAN RETINAL ASTROGLIA 1939
FIGURES 3 and 5

I940 JOSE M. RAMIREZ er cd
more-or-less parallel to the course of the axons of the NFL, independently of whether the vessel is an artery or vein. On reaching a vessel, the processes within the bundles fan out; some turn 90 deg. to form an arch-like structure that forms part of the vascular-astroglial net; the rest join other processes and form a new astroglial bundle that extends towards another vessel [Fig. 3(B)].
The cell bodies of the star-shaped astrocytes are usually located over the blood vessels; occasionally the cell bodies can be seen clustered in some areas on the vessels [Fig. 2(C,D)]. At other points a cell body can lie within the chambers of the combs [Fig. 2(A,B)] or else form part of an astroglial bundle, in which case the cell appears less stellate [Fig. 2(E)].
The astroglial plexus varies in the different zones of the retina. It is much denser in the region surrounding the fovea and the optic disk [Fig. 4(A)] than in the retinal periphery [Fig. 4(B)]. Thus, the honeycombs forming the plexus are quite small in the vicinity of the optic disk and progressively increase in size towards the ora serrata. This is due to both the progressive decrease in the number of astrocytes and blood vessels and the progress- ive increase in the distance between the latter. The honeycomb structure is difficult to make out in the proximity of the ora serrata, which is only reached by a few capillaries {Fig. 4(C)].
The processes of the GCL astrocytes can reach as far into the retinal parenchyma as the INL, where a very few astrocyte cell bodies have also been seen [Fig. 4(D)].
FIGURE 4. Astrocyte distribution in the human retina (anti-GFAP staining). (A, B) GCL astroghal retina: the plexus ia much denser in the central retina (A) than in the periphery (B) [(A, B) 50 x )I. (c) R;ieiaa
honeycomb structure is difficult to observe in the proximity of the ora serrata (125 x ). (D) Vertical retinaI-sect&n: the GCL
astrocytes surround a blood vessel (arrowhead); some astroglial processes accompany the external retinal capillaries into the INL (arrow) (* vitreo, 250 x ). (E, F) Irregularly-shaped plexus of the internal plexiform layer and INL. (E) Focus at the GCL,
and (F) focus at the INL, in this micrograph the irregularly shaped plexus is clearly visible (arrows) (200 x ). [(A, B, C) Whole
mount preparation. PAP; (D) vertical sections. immunofluorescence: (E. F) whole mount, immunofluorescence.]

HUMAN RETINAL ASTROGLIA 1941
These processes accompany the capillaries and constitute an irregularly-shaped plexus, which is difficult to observe in whole-mounts because of its location deep within the retinal tissue [Fig. 4(E, F)].
3.3.2. NerveJibre layer astroglial bundles. The longest processes of the elongated NFL astrocytes join to make up astroglial bundles that run parallel to the trajectory of the axons [Fig. 5(A,B)]. These bundles are only distributed in an area of 175 mm2 around the optic disk and the fovea. Since the thickness of the NFL decreases on the periphery of this area, the astrocyte processes from the NFL fade into the astroglial plexus of the GCL [Fig. 5(C)].
The astrocytes disappear in the avascular zone of the fovea; those on the boundary between the vascular and avascular zones have a star-shaped morphology [Fig. 5(D)].
From the exterior edge of the perifoveal region, the bundles lie in a band approx. 2 mm wide concentric to the centre of the fovea [Fig. 6(A)], except along the papillomacular bundle where the compact bundles fol- low to a straight trajectory between the fovea and the optic disk [Fig. 6(B)].
In the horizontal raphe, 2.5 mm temporally from centre of the fovea, the NFL astrocytes are more star-shaped, but still lie concentrically to the fovea [Fig. 2(F)].
The results are schematically presented in Figs 7, 8, 9, which give a three-dimensional reconstruction of astro- cyte distribution in three retinal regions: near the optic disk (Fig. 7); the retinal periphery (Fig. 8); and the perifoveal-fovea1 regions (Fig. 9).
4. DISCUSSION
Astrocyte morphology and distribution have been studied in anti-GFAP stained whole-mount and vertical section human retina preparations. The earliest morpho- logical studies of these cells were made with silver- stained horizontal sections. Compared with the less specific silver techniques, anti-GFAP marking allows astrocytes to be labelled more selectively (Dahl et al., 1986) and improves cell visualization. This is possible because GFAP is the major component of the filaments found in astrocytes (Lazarides, 1980; Dahl et al., 1986). So, since Miillerian glia in normal retinas do no.t react to GFAP (Newman, 1986; Schnitzer, 1988b), and there are no other known GFAP( +) structures in the retina, any GFAP immunoreactivity found there must correspond to astrocytes.
4.1. Retinal astrocyte distribution
Human GFAP(+) retinal astrocytes are mainly lo- cated in the NFL and GCL, although their processes can reach into the INL, where a very few astrocyte cell bodies can occasionally be found in association with capillaries. The ceils are found throughout the entire retina except for a rim adjacent to the ora serrata and a circular area centred on the fovea with a diameter of 0.4 mm.
FIGURE 6. Perifoveal astrocytes. (A) Astrocytes lying concentrically to the fovea, which is beyond the right edge of the micrograph (125 x ). (B) Region of the papillomacular bundle where the astrocytes form compact parallel bundles (the optic disk is beyond the left edge of the
micrograph) (250 x ). (Whole mount preparations, PAP.)
In the human retina astrocytes are found in the same regions as the blood vessels. These cells are situated in the GCL and form a plexus that surrounds the blood vessels. The astrocyte processes can also accompany the capillaries as far as the INL. In the NFL the astrocytes run parallel to the trajectory of the axons and can reach the radial peripapillary capillaries. This capillary plexus stops abruptly in the vicinity of the main arterial trunks at 4mm nasal and 7 mm temporal to the optic disk (Toussain, Kuwabara & Cogan, 1961; Henkind, 1967, 1969; Hogan, Alvarado & Weddell, 1971) and in this layer the number of astrocytes gradually decreases to the point of disappearing at approx. 6 mm nasal, 10 mm temporal, 5 mm superior and 6 mm inferior from the optic disk.
The astrocytes also disappear from a zone with a diameter of about 0.4 mm in the centre of the fovea, in a region where the blood vessels are also absent.
The association between astrocytes and blood vessels has been reported previously. Using electronmicroscopy, Ikui et al. (1976) observed the absence of astroglia in the avascular zones of the human retina (avascular zone of the fovea and proximity of the ora serrata) and suggested a relation between vascularization and astrocyte distri- bution. In addition, in animals with partially vascular- ized retinas [i.e. rabbits (Schnitzer, 1985, 1988a, b; Schnitzer & Karschin, 1986; Stone & Dreher, 1987; Triviiio, Ramirez, Ramirez, Salazar & Garcia, 1988, 1990, 1992; Robinson & Dreher, 1989)] or nearly

JOSE M. RAMIREZ et al.
FIGURE 7. ~r~~mensional scheme of retinal astrocyte distribution in the region near the optic disk. The astrocytes are shown in the levels of the NFL, GCL and INL. In the NFL the astrocytes are elongated and their processes join to form bundles that lie between the axonal bundles and radial peripapillary capillaries (I). In the GCL the astrocytes are star-shaped; their processes reach other astrocyte processes or blood vessels and form a honeycombed astroglial plexus (2). Accompanying capillary processes from astrocytes in the GCL can reach the INL, and these processes form an irregularly shaped plexus (3).
(OD, optic disk.)
FIGURE 8. Three-dimensional scheme of retinal astrocyte distribution in the peripheral zone. At this level the NFL is quite thin and the longitudinal astroglial bundles have disappeared (arrowhead); a few astrocytes lie between the ganglionary cell layer capillaries (arrow). The marked decrease in the number of astrocytes and blood vessels in this part of the retina make
it difficult to make out the honeycombed astroglial plexus.

HUMAN RETINAL ASTROGLIA 1943
FIGURE 9. Three-dimensional scheme of retinal astrocyte distribution in the perifoveal and fovea1 regions at the level of the papillomacular bundle. In the perifoveal region (1) astrocytes are found in all three retinal layers: NFL, GCL and INL. In the NFL, the astrocyte processes form straight compact bundles that lie between the axonal bundles of the papillomacular bundle. In the GCL, the star-shaped astrocytes and their processes form a honeycombed plexus around and between the blood vessels; the capillaries from the GCL that reach the INL are also accompanied by processes from the star-shaped astrocytes. In the fovea1 region (2), where the retinal layers become much thinner, the elongated NFL astrocytes disappear and only a few star-shaped GCL astrocytes, which accompany the capillaries, remain. At 0.4 mm from the centre of the fovea, the last
capillary and astrocyte, have disappeared. (OD, optic disk.)
avascular retinas [i.e. horses, hamsters and possums (Stone & Dreher, 1987; Schnitzer, 1987, 1988a, b)], astro- cytes are only found in zones with blood vessels; in animals with total retinal vascularization (cats, rats and primates) these cells are distributed throughout the retina except for the avascular zones (Ogden, 1978; Bussow, 1980; Shaw & Weber, 1983; Karschin, Wassle & Schnitzer, 1986a, b; Stone & Dreher, 1987; Schnitzer, 1987, 1988a,b; Distler, Weigel & Hoffmann, 1993).
The astrocyte-blood vessel relation begins during embryological development. Stone and Dreher (1987) proposed that cat retina astrocytes could penetrate from the optic nerve at the same time as the blood vessels; and later studies (Ling & Stone, 1988; Chang-Ling, Halasz & Stone, 1990; Chang-Ling & Stone, 1991a,b) suggested that molecules secreted by spindle cells (retinal vascular- ization precursors) during retinal development guided astrocyte migration from the optic disk in the retina. However, a recent study (Jiang, Liou & Caldwell, 1993) has suggested that retinal microvascular endothelial cell growth, structural phenotype and fibroenectin pro- duction are modulated by the astrocytes.
4.2. Retinal astrocyte morphology
Morphologically, two types of astrocytes can be ob- served in two different parts of the retina. Basically, star-shaped astrocytes are found in the GCL and elongated astrocytes are found in the NFL.
The morphology of these cells, is determined by the adaptation of their processes to the surrounding structures (Chang-Ling & Stone, 1991b; Holllnder, Makarov, Dreher, van Driel, Chang-Ling & Stone, 1991; Distler et al., 1993). Astrocytes in NFL, which are located between the axonal bundles and the radial peripapillary capillaries, have an elongated morphology; astrocytes located in the GCL, which interconnect the
blood vessels within this layer, have a more starry appearance.
Different authors have observed the elongated astro- cytes in the NFL (Wolter, 1955a,b, 1957, 1959, 1961; Ogden, 1978). Bussow (1980) suggested that the astroglia in the monkey and cat retina would be a cell type that associated specifically with the axons of the ganglion cells. Karschin et al. (1986a), supported this theory when they observed that, after induced axonal degeneration, the elongated astrocytes of the cat retina lost this form and sent processes in all directions. These observations suggested to Chang-Lin and Stone (1991b) that the typical star shape of the astrocytes was, in fact, a “lack of form” adopted by these cells in the absence of other structures.
According to HollHnder et al. (1991), the force that would determine the lengthened morphology of the NFL astrocytes would be the affinity between the astrocyte processes and the axonal node-like sites. This hypothesis would also explain findings in rat retina showing that astrocytes approaching axons were star-shaped (Stone & Dreher, 1987); these astrocytes were not apposed to the node-like sites on the axons, which were actually ap- proached by Miiller glia processes (HollHnder et al., 1991).
Like Chang-Ling and Stone (1991b) and Distler et al. (1993), we think that all- retinal astrocytes would be variations or subtypes of the same basic astroglial type that had different degrees of specialization. Thus, elongated astrocytes would be adapted to connections with the axonal nodes while the star-shaped astrocytes would be more involved in vascular connections.
4.3. Astroglial relations
Like Ikui et al. (1976), we have observed that processes from human retinal astrocytes tend to join

1944 JOSE M. RAMIREZ CI tri.
together and form astroglial bundles. At the level of the GCL, these bundles connect the different blood vessels forming the honeycomb plexus that was already de- scribed by Wolter 1955a, b, 1957, 1959, 1961) using silver techniques.
The cell bodies of the astrocytes that make up this plexus can be found within the chambers of the honey- combs, or as part of the astroglial bundles, but, more normally, they lie over blood vessels. On many occasions we have observed clustered astrocyte cell bodies, a formation that has not been described before in man or other mammalian species.
In the GCL the astroglial plexus is denser in the proximity of the optic disk than in the periphery, corresponding to the gradual decrease in blood vessels in the direction of the ora serrata.
The astrocytes in this layer send processes towards the INL forming an irregular plexus that accompanies the external retinal capillaries. This plexus coincides with the one Wolter (1955a, b, 1961) described in the IPL and INL of human retina; but it has not been found in the retina of the monkey (Distler et at., 1993).
Within the NFL the astroglial processes tend to group into bundles that are aligned along the axons and may contact adjacent radial peripapillary capillaries. These astrocytes can also send processes to the GCL astroglial plexus.
In the NFL, the parallel astroglial bundles radiate straight out from the optic disk. Temporal to the optic disk the bundles course concentrically around the centre of the fovea and an axis in the area of the papillomacular bundle between the optic disk and the fovea. The large number of axons in the papillomacular bundle (Pollock & Miller, 1986) results in densely packed astrocyte bundles. The NFL disappears in the vicinity of the fovea (Hogan et al., 1971), and, thus, the astrocytes located in the fovea are the star-shaped astrocytes of the GCL which accompany the scarce capillaries of the vascular- ized rim of the fovea. Star-shaped astrocytes have also been observed in monkey fovea (Distler er al., 1993). At the horizontal raphe, although located in the NFL, the astrocytes are star-shaped, possibly because of the dis- tinctive distribution of the axons in this area. Axons from ganglion cells located slightly below the horizontal raphe course superiorly, while axons from other ganglion cells located slightly above the horizontal raphe course inferiorly (Hogan et al., 1971; Sigelman & Ozanics, 1982; Pollock et al., 1986); these mingled axons could explain the starry shape of horizontal raphe astrocytes in the NFL.
The existence of an astroglial net in the vascularized zones of the retina has also been observed in other species (Karschin et at., 1986a, b; Stone & Dreher, 1987; Distler et al., 1993), although the structure of this net can be quite different from that in humans. The cat has a single continuous astroglial net in which the shape of the astrocytes changes from stellated to elongated as the position of the cell approaches the optic disk (Karschin er al., 1986a, b). There are two astroglial cell layers in monkeys: in the NFL the appearance of the astrocytes
changes from star shaped on the periphery to elongated in more central parts of the retina; the astrocytes of the GCL are all star shaped (Distler et al., 1993). In man, as in the monkey, there are two layers of astroghal cells. one in the GCL and the other in the NFL, but. differently from the descriptions of the monkey retina (Distler et al., 1993), in man the astrocytes in each layet have their own characteristic morphology.
Chang-Ling and Stone (199lb) hypothesized that astrocyte distribution in the cat retina was determined by a “contact-spacing” relation between adjacent astrocytes through which the processes could meet but a distance would be maintained between cell bodies. This inter- action would require repulsive forces to separate the astrocyte cell bodies and adhesive forces that would maintain process contact so that a net would be formed (Tout, Dreher, Chang-Ling & Stone, 1993). However. this theory cannot be applied in the human retina since, as has been described here, the astrocyte cell bodies often cluster in certain vascular zones.
Wolter (1955a,b, 1957) described a special type of astroglial cell in the human retina. It was only related with blood vessels and he called it “‘perivascular astro- cytes”. These cells were described as lying over the blood vessels, which they surrounded with their processes, although they never contacted other structures. Most later authors also accepted the existence of these cells in the human retina (Erskine, 1963; Wolff, 1968). However, when Ikui et al. (1976) could find no ultrastructural difference between the astrocytes that were located over the vessels and those that were not, they questioned the existence of perivascular astrocytes. Later studies using immunohist~hemical techniques on rabbit retinas have detected the presence of perivascular glia (Sehnitzer, 1985, 1988a,b; Schnitzer & Karschin, 1986; Stone & Dreher, 1987; Triviiio, Ramirez, Ramirez, Salazar & Garcia-Sanchez, 1988, 1992; Robinson & Dreher, 1989); these reports also implicity accepted the existence of these cells in the human retina.
Using anti-GFAP on human retinas WE have not been able to observe Wolter’s perivas~ular astrocytes over the blood vessels in any of the numerous whole-mounts or sections examined for this study. Perhaps Woker’s silver impregnations (Wolter, 1955a, b), which stained superfi- cially, only marked the network of fibres lying over the vessels and gave it a cellular appearance which would have lead to his interpretation of the images.
In addition, no author has reported observing Walter’s perivascular glia in any retina from cats, rats, or monkeys, where astroglial distribution is like that in humans.
The presence of these cells in rabbit retinas could be explained by the position of the blood vessels, which are epiretinal (Hyvlrinen, 1967); in men, monkeys, cats and rats, the vessels are intraretinally located (Michaelson, 1954). An epiretinal vascular position would favour the appearance of glial types that are specialized in

anchoring vessels to the retinal parenchyma. An earlier paper using rabbit retinas has reported three types of astroglial cells associated with the vascular system (Triviiio et al., 1992). Type I and Type II astrocytes would be responsible for anchoring the vessels to the retina, and would therefore be authentic perivascular astrocytes. Type III astrocytes, located in the retinal parenchyma, would contact the intraparenchymal reti- nal capillaries. This last cell type would be the equivalent to the star-shaped astrocytes found in the GCL of the human retina.
Sharing and division of labour between astrocytes and Miiller cells.
Journal of Comparative Neurology, 313, 581-603.
Hui, Y. N., Goodnight, R., Zhang, X. J., Sorgente, N. & Ryan, S. J.
(1988). Glial epiretinal membranes and contraction. Immunohisto-
chemical and morphological studies. Archives of Ophthalmology,
106, 128&1285.
Hyviirinen, L. (1967). Vascular structures of the rabbit retina. Acta
Ophthalmologica, 45, 852-861.
Ikui, H., Uga, S. & Kohno, T. (1976). Electron microscope study on
astrocytes in the human retina using ruthenium red. Ophthalmic
Research, 8, 10&110.
Thus, the existence of “perivascular glia” that func- tion as vascular anchors in intraretinally vascularised retinas would be unnecessary. These cells might exist in pathological conditions, like retinopathy of prematurity or in epiretinal membranes (Hiscott et al., 1984; Hui et al., 1988; Sramek et al., 1989), in which intravitreal vessels can appear. However, the question of whether the astroglial cells that appear in these pathological con- ditions are actually like the perivascular astrocytes of the rabbit retina or are a different glial type must still be determined.
Jiang, B., Liou, G. & Caldwell, R. B. (1993). Astrocytes modulate
endothelial cell growth differentiation, and fibronectin expression.
Investigative Ophthalmology and Visual Science (Suppi.), 34, P987.
Karschin, A., Wissle, H. & Schnitzer, J. (1986a). Shape and distri-
bution of astrocytes in the cat retina. Investigative Ophthalmology
and Visual Science, 27, 828-831.
Karschin, A., Wlssle, H. & Schnitzer, J. (1986b). Immunocyto-
chemical studies on astroglia of the cat retina under normal and
pathological conditions. Journal of Comparative Neurology, 249,
564-576.
REFERENCES
Kivelii, T., Tarkkanen, A. & Virtanen, I. (1986). Intermediate filaments
in the human retina and retinoblastoma. An immunohistochemical
study of vimentin, glial fibrillary acidic protein, and neurofilaments.
Investigative Ophthalmology and Visual Science, 27, 1075-1084.
Lane, J. C. & Klintworth, G. H. (1983). A study of astrocytes in
retinoblastomas using the immunoperoxidase technique and anti-
bodies to glial fibrillary acidic protein. American Journal of Ophthal-
mology, 95, 197-207.
Biissow, H. (1980). The astrocytes in the retina and optic nerve head
of mammals: A special glia for the ganglion cell axons. Cellular and
Tissue Research, 206, 367-378.
Chang-Ling, T. & Stone, J. (1991a). Factor determining the migration
of astrocytes into the developing retina: Migration does not depend
on intact axons or patent vessels. Journal of Comparative Neurology,
303, 375-386.
Lazarides, E. (1980). Intermediate filaments as mechanical integrator
of cellular space. Nature, 283, 249-256.
Ling, T. L. & Stone, J. (1988). The development of astrocytes in the
cat retina: Evidence of migration from the optic nerve. Deuelopmen-
tal Brain Research, 44, 73-85.
Chang-Ling, T. & Stone, J. (1991b). Factors determining the mor-
phology and distribution of astrocytes in the cat retina: A “contact-
spacing” model of astrocyte interaction. Journal of Comparative
Neurology, 303, 387-399.
Chang-Ling, T., Halasz, P. & Stone, J. (1990). Development of retinal
vasculature in the cat: Processes and mechanisms. Current Eye
Research, 9, 459478.
Marchesani, 0. (1926). Die morphologie der glia im nervus opti-
cus und in der retina, dargestellt nach den neusten unter-
suchungsmethoden und untersuchungsergebnissen. Archives ftir
Ophthalmologie (Graefe’s), 117, 575405.
Michaelson, I. C. (1954). Retinal circulation in man and animals
(pp. 29-38). Springfield, Ill.: Charles C. Thomas.
Michaelson, I. C. &Campbell, A. C. (1940). The anatomy of the retinal
vessels, and some observations on their significance in certain retinal
diseases. Transactions of the Ophthalmological Society of the United
Kingdom, 60, 71L112. Dahl, D., Bjiirklund, H. & Bignami, A. (1986). Immunological mark-
ers in astrocytes. In Federoff, S. & Vernadakis, A. (Eds), Astrocytes,
Vol. III: Cell biology and pathology of astrocytes (pp. I-19).
London: Academic Press.
Distler, C., Weigel, H. & Hoffman, K. P. (1993). Glia cells of the
monkey retina. I. Astrocytes. Journal of Comparative Neurology,
333, 134-147.
Newman, E. A. (1986). The Miiller cell. In Federoff, S. & Vernadakis,
A. (Eds), Astrocytes, Vol. I: Development, morphology, and regional
specialization of astrocytes (pp. 149-171). London: Academic Press.
Ogden, T. E. (1978). Nerve fiber layer astrocytes of the primate retina:
Morphology, distribution, and density. Investigative Ophthalmology
and Visual Science, 17, 499-5 10.
Erskine, C. A. (1963). Neuroglial relationships in the human retinal
capillary network. British Journal of Ophthalmology, 47, 601410.
Gutrin, C. J., Wolfshagen, R. W., Eifrig, D. E. & Anderson, D. H.
(1990). Immunocytochemical identification of Miiller’s glia as a
component of human epiretinal membranes. Investigative Ophthal-
mology and Visual Science, 31, 1483-1491.
Henkind, P. (1967). Radial peripapillary capillaries of the retina. I.
Anatomy: Human and comparative. British Journal of Ophthal-
mology, 51, 115-123.
Pollock, S. C. & Miller, N. R. (1986). The retinal nerve fiber layer.
International Ophthalmology Clinics, 26, 201-221.
Polyak, S. L. (1941). The retina. Chicago, Ill.: University of Chicago
Press.
Ram6n y Cajal, S. (1982). La rktine des verttbr&. La Cellule, 9,
121-255.
Henkind, P. (1969). Microcirculation of the peripapillary retina.
Transactions of the American Academy of Ophthalmology and
Otolaryngology, 73, 890-897.
Hiscott, P. S., Grierson, I., Trombetta, C. J., Rahl, H. S.,
Marshall, J. & McLeod, D. (1984). Retinal and epiretinal glia:
An immunohistochemical study. British Journal of Ophthalmology,
68, 698-707.
Ram6n y Cajal, S. (1911). Histologie du SystPme de I’homme et des
vertebr& (Vol. II, pp. 296367). Paris: A. Maloine fiditeur.
Robinson, S. R. & Dreher, Z. (1989). Evidence for three morphological
classes of astrocyte in the adult rabbit retina: Functional and
developmental implications. Neuroscience Letters, 106, 261-268.
Schnitzer, J. (1985). Distribution and immunoreactivity of glia in the
retina of the rabbit. Journal of Comparative Neurology, 240,
128-142.
Schnitzer, J. (1987). Retinal astrocytes: Their restriction to vascu-
larized parts of the mammalian retina. Neuroscience Letters, 78, 29-34.
Hogan, M. J., Alvarado, J. A. & Weddel, J. E. (1971). Histology of the
human eye: An atlas and textbook (pp. 393-522). Toronto: W. B. Saunders & Co.
Schnitzer, J. (1988a). The development of astrocytes and blood vessels
in the postnatal rabbit retina. Journal of Neurocytology, 17,
433449. Hollinder, H., Makarov, F., Dreher, Z., van Driel, D., Chang-Ling, Schnitzer, J. (1988b). Astrocytes in mammalian retina. Progress in
T. L. & Stone, J. (1991). Structure of the macroglia of the retina: Retinal Research, 7, 209-23 1.
HUMAN RETINAL ASTROGLIA 1945

1946 JOSE M. RAMIREZ er (11.
Schnitzer, J. SC Karschin, A. (1986). The shape and distribution of Trivirio, A., Ramirez, A. 1.. Salazar, J. J.. Ramirer, J. M. & astrocytes in the retina of the adult rabbit. Cellular and Tissue Garcia-Sanchez, J. (1990). Astrocitos retinianos relacionados Research, 246, 91-102. con 10s axones de las celulas ganglionares. Estudio inmuno-
Shaw, G. & Weber K. (1983). The structure and development of the histoquimico. Archives de la Sociedad Espariola de Ofiabnologio Invest, 3, 87-92. rat retina: An immunofluorescence microscopical study using anti-
bodies specific for intermediate filament proteins. European Journal of Cellular Biology, 30, 219-232.
Sigelman, J. & Ozanics, V. (1982). Retina. In Jakobiec, F. A. (Ed.), Ocular anatomy, embryology and teratology (pp. 44-506). Philadel- phia, Pa: Harper & Row.
Sramek, S. J., Wallow, I. H., Stevens, T. S. & Nork, T. M. (1989). Immunostaining of preretinal membranes for actin, fibronectin, and glial fibrillary acidic protein. Ophthalmology, 96, 835-841.
Stone, J. & Dreher, Z. (1987). Relationship between astrocytes, ganglion cells and vasculature of the retina. Journal of Comparative Neurology, 255, 3549.
Toussaint, D., Kuwabara, T. & Cogan, D. G. (1961): Retinal vascular patterns. Part II. Human retinal vessels studied in three dimensions. Archives of Ophthalmology, 65, 575-581.
Tout, S., Dreher, Z., Chang-Ling, T. L. & Stone J. (1993). Contact- spacing among astrocytes is independent of neighbouring struc- tures: In vivo an in vitro evidence. Journal of Comparative
Neurology, 332, 433443. Trivifio, A., Ramirez, A. I., Ramirez, J. M., Salazar, J. J. & Garcia-
Sanchez, J. (1988). Estudio de las relaciones vaso-gliales en la retina del conejo albino (Oryctolagus cunicuius). Archives de la Sociedad Espaiiola de Qftalmologia Invest, I, 103-I 10.
Wollf. E. (1968). Anatomy of the eye and orbit (in&ding the central connections, development, and comparative anatomy of the visual apparatus) (pp. 135.-138), 6th edn (revised by Last, R. J.). Philadelphia, Pa: W. B. Saunders & Co.
Walter, J. R. (1955a). The astroglia of the human retina: And other glial elements of the retina under normal and pathologic con- ditions. American Journal of Ophthalmology, 40, 888100.
Wolter, J. R. (1955b). The cells of Remark and the astroglia of the normal human retina. Archives of Ophthalmology. 53, 832-838.
Wolter, J. R. (1957). Perivascular glia of the blood vessels of the human retina. American Journal of Ophthalmology, 44, 766.-773.
Wolter, J. R. (1959). Glia of the human retina. American Journal of Ophthalmology, 48, 370-393.
Wolter, J. R. (1961). Silver carbonate techniques for the demonstration of ocular histology. In Smelser, G. K. (Eds) The structure of the eye (pp. 117-138). New York: Academic Press.
Triviio, A., Ramirez, J. M., Ramirez, A. I., Salazar, J. J. & Garcia- Acknowledgements-We thank C. F. Warren for her linguistic Sanchez, J. (1992). Retinal perivascular astroglia: An immunoper- assistance. This .work was supported by CAICYT grant PM89-0200 oxidase study. Vision Research, 32, 1601-1607. and PB92-0185.




















