
Rule-dependent Activity for Prosaccades and Antisaccades in the Primate Prefrontal Cortex
14
Rule-dependent Activity for Prosaccades and Antisaccades in the Primate Prefrontal Cortex Stefan Everling and Joseph F. X. DeSouza Abstract & Everyday life typically requires behavior that involves far more than simple stimulus–response associations. Environ- mental cues are often ambiguous and require different actions depending on the situation. The prefrontal cortex (PFC) is thought to be crucial for this flexible control of behavior. An important task that probes this ability is the antisaccade task in which subjects have to suppress a glance towards a suddenly presented peripheral stimulus and instead look away from the stimulus to its mirror location. Here we recorded the activity of PFC neurons in monkeys trained to alternate between blocks of prosaccade and antisaccade trials with no external instruc- tion cues. We found that the activity of many neurons was different between the two tasks during the fixation period before the peripheral stimulus was presented. These differ- ences were already present on the first correct trials after a task switch. The activity of these neurons also discriminated between correct responses and errors. We hypothesize that the PFC provides bias signals to saccade-related areas that are necessary to preset the oculomotor system for different tasks. & INTRODUCTION The ability to alter behavioral responses to identical stimuli in the face of changing contingencies is especially well developed in primates. A classic neuropsychological task that probes this ability is the Wisconsin Card Sorting Task in which subjects are asked to sort cards according to the color, shape, or number of symbols appearing on them (Milner, 1963). Subjects are provided with feed- back after each match which enables them to acquire the correct classification scheme. After a fixed number of matches, the rule changes without notice and the subject must shift to a new rule of classification. Human patients with lesions of the prefrontal cortex (PFC) can acquire the initial rule but are unable to shift to a new classification rule. Functional neuroimaging studies have confirmed a role of the PFC in set-shifting in humans (Monchi, Petrides, Petre, Worsley, & Dagher, 2001) and nonhuman primates (Nakahara, Hayashi, Konishi, & Miyashita, 2002). Further, nonhuman primates with PFC lesions are impaired in analogous tasks (Dias, Robbins, & Roberts, 1996). Recent single-neuron recording studies in monkeys have begun to reveal neural correlates for rules in the PFC (Wallis, Anderson, & Miller, 2001; Asaad, Rainer, & Miller, 2000; White & Wise, 1999) and in the posterior parietal cortex (Stoet & Snyder, 2004). Neural responses to pictures and during delay periods were found to be different in the PFC between a delayed match-to-sample and a delayed nonmatch-to-sample task (Wallis et al., 2001). The stimuli and the delay periods were identical between the two tasks with the only difference being the behavioral rules that required the monkeys in the first task to decide whether two successively presented pictures were the same and in the second task whether they were different. Indeed, many PFC neurons show differences already in their baseline activity between different tasks. Asaad et al. (2000) recorded single- neuron activity in the PFC while monkeys performed a spatial, an object, and an association task. Neural activity was different for many cells between the three tasks while monkeys were looking at a central fixation point (FP) before a stimulus was presented. The authors suggest that these differences may reflect task-specific activation that might allow conflicting sensory informa- tion to be mapped to the appropriate motor output. All studies so far have investigated the role of the PFC in maintaining or alternating between two or more arbitrary stimulus–response (SR) associations. In every- day life, however, some SR associations are strong and might be executed in a more ‘‘automatic’’ manner, whereas others are weak and require a more ‘‘con- trolled’’ execution (Baddeley, 1986). Strong SR associa- tions are usually those in which the stimulus and the response are compatible, whereas incompatible SR asso- ciations are usually weak. It is, for example, well known from manual SR compatibility tasks that involve spatial stimuli and spatial responses that reaction times are faster and responses are more accurate when the stim- ulus and the response are compatible rather than in- University of Western Ontario, London, Canada D 2005 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 17:9, pp. 1483–1496
Transcript of Rule-dependent Activity for Prosaccades and Antisaccades in the Primate Prefrontal Cortex

Rule-dependent Activity for Prosaccades andAntisaccades in the Primate Prefrontal Cortex
Stefan Everling and Joseph F X DeSouza
Abstract
amp Everyday life typically requires behavior that involves farmore than simple stimulusndashresponse associations Environ-mental cues are often ambiguous and require different actionsdepending on the situation The prefrontal cortex (PFC) isthought to be crucial for this flexible control of behavior Animportant task that probes this ability is the antisaccade task inwhich subjects have to suppress a glance towards a suddenlypresented peripheral stimulus and instead look away from thestimulus to its mirror location Here we recorded the activity ofPFC neurons in monkeys trained to alternate between blocks
of prosaccade and antisaccade trials with no external instruc-tion cues We found that the activity of many neurons wasdifferent between the two tasks during the fixation periodbefore the peripheral stimulus was presented These differ-ences were already present on the first correct trials after a taskswitch The activity of these neurons also discriminatedbetween correct responses and errors We hypothesize thatthe PFC provides bias signals to saccade-related areas that arenecessary to preset the oculomotor system for differenttasks amp
INTRODUCTION
The ability to alter behavioral responses to identicalstimuli in the face of changing contingencies is especiallywell developed in primates A classic neuropsychologicaltask that probes this ability is the Wisconsin Card SortingTask in which subjects are asked to sort cards accordingto the color shape or number of symbols appearing onthem (Milner 1963) Subjects are provided with feed-back after each match which enables them to acquirethe correct classification scheme After a fixed number ofmatches the rule changes without notice and thesubject must shift to a new rule of classification Humanpatients with lesions of the prefrontal cortex (PFC) canacquire the initial rule but are unable to shift to a newclassification rule Functional neuroimaging studies haveconfirmed a role of the PFC in set-shifting in humans(Monchi Petrides Petre Worsley amp Dagher 2001) andnonhuman primates (Nakahara Hayashi Konishi ampMiyashita 2002) Further nonhuman primates withPFC lesions are impaired in analogous tasks (DiasRobbins amp Roberts 1996)
Recent single-neuron recording studies in monkeyshave begun to reveal neural correlates for rules in thePFC (Wallis Anderson amp Miller 2001 Asaad Rainer ampMiller 2000 White amp Wise 1999) and in the posteriorparietal cortex (Stoet amp Snyder 2004) Neural responsesto pictures and during delay periods were found to bedifferent in the PFC between a delayed match-to-sample
and a delayed nonmatch-to-sample task (Wallis et al2001) The stimuli and the delay periods were identicalbetween the two tasks with the only difference being thebehavioral rules that required the monkeys in the firsttask to decide whether two successively presentedpictures were the same and in the second task whetherthey were different Indeed many PFC neurons showdifferences already in their baseline activity betweendifferent tasks Asaad et al (2000) recorded single-neuron activity in the PFC while monkeys performed aspatial an object and an association task Neural activitywas different for many cells between the three taskswhile monkeys were looking at a central fixation point(FP) before a stimulus was presented The authorssuggest that these differences may reflect task-specificactivation that might allow conflicting sensory informa-tion to be mapped to the appropriate motor output
All studies so far have investigated the role of the PFCin maintaining or alternating between two or morearbitrary stimulusndashresponse (SR) associations In every-day life however some SR associations are strong andmight be executed in a more lsquolsquoautomaticrsquorsquo mannerwhereas others are weak and require a more lsquolsquocon-trolledrsquorsquo execution (Baddeley 1986) Strong SR associa-tions are usually those in which the stimulus and theresponse are compatible whereas incompatible SR asso-ciations are usually weak It is for example well knownfrom manual SR compatibility tasks that involve spatialstimuli and spatial responses that reaction times arefaster and responses are more accurate when the stim-ulus and the response are compatible rather than in-University of Western Ontario London Canada
D 2005 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 179 pp 1483ndash1496
compatible (Munoz amp Everling 2004) A popular oculo-motor task that requires an incompatible SR associationis the antisaccade task (Munoz amp Everling 2004 Hallettamp Adams 1980 Hallett 1978) In contrast to a morelsquolsquoautomaticrsquorsquo prosaccade task that requires subjects tolook towards a suddenly presented stimulus in theperipheral visual field the antisaccade task is morelsquolsquocontrolledrsquorsquo and requires subjects to suppress thisprepotent response and instead to look away from thestimulus to its mirror location Therefore alternatingbetween prosaccade trials and antisaccade trials requiressubjects to switch between an lsquolsquoautomaticrsquorsquo task and alsquolsquocontrolledrsquorsquo task
To explore the role of the PFC in maintaining andswitching between prosaccades and antisaccades we
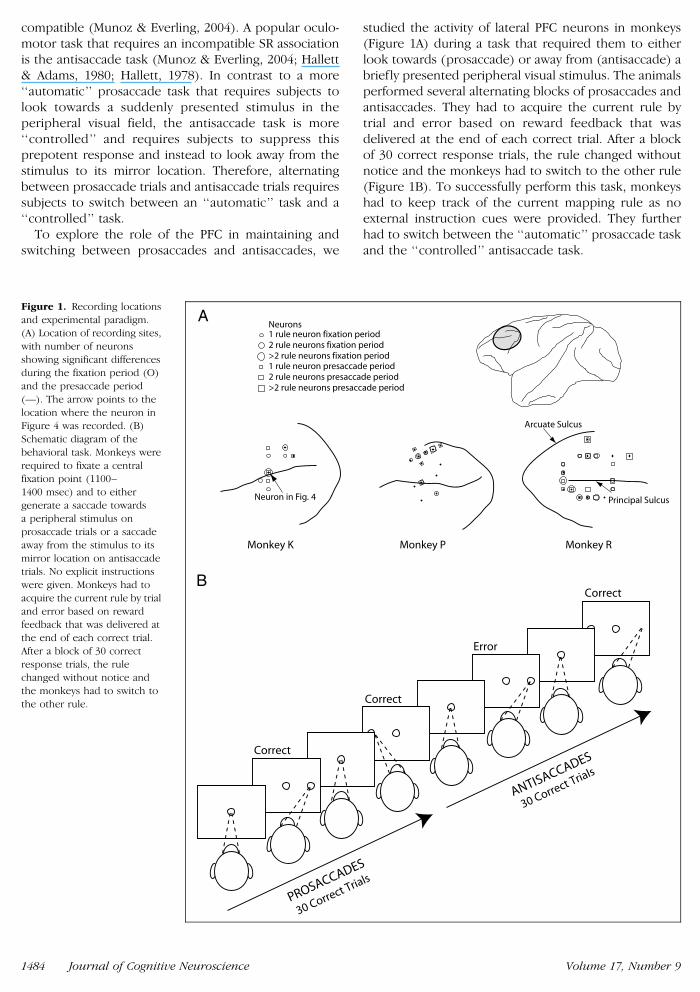
studied the activity of lateral PFC neurons in monkeys(Figure 1A) during a task that required them to eitherlook towards (prosaccade) or away from (antisaccade) abriefly presented peripheral visual stimulus The animalsperformed several alternating blocks of prosaccades andantisaccades They had to acquire the current rule bytrial and error based on reward feedback that wasdelivered at the end of each correct trial After a blockof 30 correct response trials the rule changed withoutnotice and the monkeys had to switch to the other rule(Figure 1B) To successfully perform this task monkeyshad to keep track of the current mapping rule as noexternal instruction cues were provided They furtherhad to switch between the lsquolsquoautomaticrsquorsquo prosaccade taskand the lsquolsquocontrolledrsquorsquo antisaccade task
Figure 1 Recording locations
and experimental paradigm(A) Location of recording sites
with number of neurons
showing significant differencesduring the fixation period (O)
and the presaccade period
(mdash) The arrow points to the
location where the neuron inFigure 4 was recorded (B)
Schematic diagram of the
behavioral task Monkeys were
required to fixate a centralfixation point (1100ndash
1400 msec) and to either
generate a saccade towardsa peripheral stimulus on
prosaccade trials or a saccade
away from the stimulus to its
mirror location on antisaccadetrials No explicit instructions
were given Monkeys had to
acquire the current rule by trial
and error based on rewardfeedback that was delivered at
the end of each correct trial
After a block of 30 correct
response trials the rulechanged without notice and
the monkeys had to switch to
the other rule
1484 Journal of Cognitive Neuroscience Volume 17 Number 9
RESULTS
Neural and behavioral data were recorded from threemonkeys over a total of 46 experimental sessions (12 formonkey K 24 for monkey P 10 for monkey R)
Behavior
All three monkeys were able to successfully alternatebetween pro- and antisaccade trials Figure 2A showstheir average performance for successive blocks of trialsMonkeys required approximately 10 trials to reach fourout of four correct prosaccade responses (the arbitrary
criterion that we employed) and about 15 trials to reachthe same behavioral criterion for antisaccades Figure 2Billustrates the monkeysrsquo performance before and after atask switch The animals performed prosaccades andantisaccades at ~85 before a task switch The 100performance at the trial immediately preceding the taskswitch is an effect of the task design that switchedbetween tasks after 30 correct trials Thus every blockended with a correct trial Performance dropped dras-tically at the first trial following a task switch (~15)This is not surprising because the monkeys were notexplicitly informed when the task would switch andtherefore almost always continued to execute thepreviously rewarded behavior Indeed the perform-ance after the switch would have been identical tothe last trials in the previous block had the task notswitched The performance then gradually recoveredover the next 5 to 10 trials Interestingly monkeysperformed slightly better for a few trials after thebehavioral rule switched to antisaccades compared towhen it switched to prosaccades (red line above blueline after the task switch) This indicates that it waseasier for the animals to switch from the lsquolsquoautomaticrsquorsquoprosaccade task to the lsquolsquocontrolledrsquorsquo antisaccade taskthan vice versa
Monkey R had faster saccadic reaction times (SRTs)for prosaccades than antisaccades (median pro-SRT181 msec median anti-SRT 255 msec) whereas mon-key K (median pro-SRT 223 msec median anti-SRT221) and monkey P (median pro-SRT 203 msecmedian anti-SRT 188 msec) showed no differencesin SRTs between correct pro- and antisaccades Thisresult is somewhat surprising giving the well-knownfinding of longer SRTs for antisaccades than prosac-cades (Munoz amp Everling 2004) A possible explana-tion might be that monkey R was experimentally naıveprior to this study whereas monkeys K and P weresubjects in a number of other paradigms (attentionparadigm Desouza amp Everling 2004 randomly inter-leaved prosaccade and antisaccade paradigm Desouzaamp Everling unpublished data delayed match-to-sampleand conditional pro- and antisaccade paradigm Johnstonamp Everling unpublished data) before participating inthis study
Monkeys often generated prosaccades on antisaccadetrials (monkey K 19 monkey P 11 monkey R 39)and many of these prosaccades had short reaction timesin the range of express saccades In line with previousreports (Bell Everling amp Munoz 2000 Everling Dorrisamp Munoz 1998) monkeys did not generate antisaccadeswith express saccade latencies In contrast with previousstudies all monkeys also made a large number ofantisaccades on prosaccade trials (monkey K 14monkey P 26 monkey R 21) This is not surprisingas the animals in this study were not given an explicitinstruction cue of whether to generate a prosaccade oran antisaccade on a given trial
Figure 2 Performance over blocks and at task reversal (A) Number oftrials (mean plusmn SEM ) required by the three monkeys to reach four out
of four correct responses after a task reversal on prosaccade blocks
(blue) and antisaccade blocks (red) Data are averaged across all
recording sessions (B) Performance at the point of task reversal Theproportion of correct responses (mean plusmn SEM ) is plotted for the 10
trials prior to a task reversal to the 20 trials after a task reversal The
first trial following task reversal is at 0 Data are averaged across all
recording sessions from all three monkeys
Everling and DeSouza 1485
Neural Activity
We recorded the activity of a total of 195 neurons fromthe lateral PFC of three monkeys (36 from monkey K 82from monkey P and 77 from monkey R) while theyperformed between 8 and 27 (median 14) task switchesof alternating blocks of pro- and antisaccade trials andgenerated 388 to 912 correct responses (median 666)
Our initial analysis of the peristimulus histogramsshowed that many neurons had different dischargepatterns between pro- and antisaccade trials Figure 3shows the activity of a PFC neuron that was more activeon prosaccade trials than on antisaccade trials Thisneuron exhibited a higher activity after acquiring centralfixation on prosaccade trials than on antisaccade trialsand it maintained a higher activity level throughout thetrial The neuronrsquos response to contralateral stimulus
presentations was also stronger on prosaccade trialsthan on antisaccade trials
To examine the time-course of the mapping ruleeffect we performed a receiver operating characteristic(ROC) analysis using a sliding 500-msec time windowthat was incremented by 100 msec steps for each of the195 PFC neurons (Figure 4) These ROC values are basedon the absolute difference in discharge rate betweencorrect prosaccade trials and correct antisaccade trialspooled across directions Therefore the values in thisanalysis could range from 05 (no rule information) to10 (perfect discrimination between pro- and antisac-cade trials) To test whether these ROC values weresignificant from chance we also conducted a bootstrapanalysis (see Methods for details) The population ofPFC neurons exhibited significant rule selectivity (solidline above 95th percentile dashed line of shuffled data)
Figure 3 Single PFC neuronwith rule selectivity The
neuron was more active on
prosaccade trials than on
antisaccade trials Each dotindicates the time of an action
potential relative to peripheral
stimulus presentation andeach row represents one trial
Trials are sorted according to
saccadic reaction times (open
circles) Open trianglesindicate the onset of fixation
Superimposed on the rasters
are the average spike density
waveforms (30 msec Gaussian)(see Figure 1 for the recording
location of this neuron)
1486 Journal of Cognitive Neuroscience Volume 17 Number 9
throughout the entire trial period even prior to fixationonset This finding indicates that the activity of PFCneurons is significantly different between prosaccadeand antisaccade trials throughout the trial
Fixation Activity
Next we identified those PFC neurons that exhibitedsignificant differences in their activity between prosac-cade and antisaccade trials during the fixation periodbefore the peripheral stimulus was presented We usedthe last 500 msec immediately prior to peripheral stim-ulus presentation as a measurement of neural fixationactivity During this period the monkeys were fixating acentrally presented white dot on both prosaccade andantisaccade trials Therefore the visual stimulation andthe eye positions were identical between prosaccade andantisaccade trials during this period For each neuronwe computed an ROC value on the activity in this epochand compared it with the 95th percentile ROC value for
randomly shuff led data in a bootstrap analysis (seeMethods for details) In this analysis 26 of the neurons(51195) showed a significant difference between fixa-tion activity on prosaccade trials compared with antisac-cade trials ( p lt 05) These PFC neurons had relativelylow discharge rates during the fixation period (026ndash139 spikessec mean = 414 SD = 355 spikessec) andthe differences in neural activity between prosaccadeand antisaccade trials were therefore usually only in theorder of a few spikes per second These changes never-theless represented a considerable modulation in theneuronsrsquo discharge rate between prosaccade and anti-saccade trials The percent change in fixation activity forthese neurons ranged between 70 and 882 with amean difference of 288 (SD = 184)
We found slightly more neurons that preferred pro-saccades than antisaccades (32 of 51 or 627 weremore active for prosaccades 19 of 51 or 373 weremore active for antisaccades) This difference ap-proached significance (x2 test p = 069) We repeated
Figure 4 Time-course of rule
selectivity (A) The time-course
was determined by the sliding
ROC analysis (window500 msec centered at time
point) across all 195 neurons
from which we recorded ROCvalues for the comparison of
correct prosaccades and
correct antisaccade trials
Neurons were sortedaccording to their mean ROC
value in the period from
1000 msec before fixation
onset to 1500 msec afterfixation onset The first and
second vertical lines indicate
the randomly chosen intervalwhen the monkeys started
central fixation and the third
vertical line indicates the time
point when the peripheralstimulus was presented (B)
Average time-course of ROC
values for all 195 neurons
(solid line) and for the medianand 95th percentile of the
shuff led data (dashed lines)
The average ROC value for thepopulation was significantly
different from chance ( p lt
05) throughout the entire
analysis period
Everling and DeSouza 1487
this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9
compatible (Munoz amp Everling 2004) A popular oculo-motor task that requires an incompatible SR associationis the antisaccade task (Munoz amp Everling 2004 Hallettamp Adams 1980 Hallett 1978) In contrast to a morelsquolsquoautomaticrsquorsquo prosaccade task that requires subjects tolook towards a suddenly presented stimulus in theperipheral visual field the antisaccade task is morelsquolsquocontrolledrsquorsquo and requires subjects to suppress thisprepotent response and instead to look away from thestimulus to its mirror location Therefore alternatingbetween prosaccade trials and antisaccade trials requiressubjects to switch between an lsquolsquoautomaticrsquorsquo task and alsquolsquocontrolledrsquorsquo task
To explore the role of the PFC in maintaining andswitching between prosaccades and antisaccades we
studied the activity of lateral PFC neurons in monkeys(Figure 1A) during a task that required them to eitherlook towards (prosaccade) or away from (antisaccade) abriefly presented peripheral visual stimulus The animalsperformed several alternating blocks of prosaccades andantisaccades They had to acquire the current rule bytrial and error based on reward feedback that wasdelivered at the end of each correct trial After a blockof 30 correct response trials the rule changed withoutnotice and the monkeys had to switch to the other rule(Figure 1B) To successfully perform this task monkeyshad to keep track of the current mapping rule as noexternal instruction cues were provided They furtherhad to switch between the lsquolsquoautomaticrsquorsquo prosaccade taskand the lsquolsquocontrolledrsquorsquo antisaccade task
Figure 1 Recording locations
and experimental paradigm(A) Location of recording sites
with number of neurons
showing significant differencesduring the fixation period (O)
and the presaccade period
(mdash) The arrow points to the
location where the neuron inFigure 4 was recorded (B)
Schematic diagram of the
behavioral task Monkeys were
required to fixate a centralfixation point (1100ndash
1400 msec) and to either
generate a saccade towardsa peripheral stimulus on
prosaccade trials or a saccade
away from the stimulus to its
mirror location on antisaccadetrials No explicit instructions
were given Monkeys had to
acquire the current rule by trial
and error based on rewardfeedback that was delivered at
the end of each correct trial
After a block of 30 correct
response trials the rulechanged without notice and
the monkeys had to switch to
the other rule
1484 Journal of Cognitive Neuroscience Volume 17 Number 9
RESULTS
Neural and behavioral data were recorded from threemonkeys over a total of 46 experimental sessions (12 formonkey K 24 for monkey P 10 for monkey R)
Behavior
All three monkeys were able to successfully alternatebetween pro- and antisaccade trials Figure 2A showstheir average performance for successive blocks of trialsMonkeys required approximately 10 trials to reach fourout of four correct prosaccade responses (the arbitrary
criterion that we employed) and about 15 trials to reachthe same behavioral criterion for antisaccades Figure 2Billustrates the monkeysrsquo performance before and after atask switch The animals performed prosaccades andantisaccades at ~85 before a task switch The 100performance at the trial immediately preceding the taskswitch is an effect of the task design that switchedbetween tasks after 30 correct trials Thus every blockended with a correct trial Performance dropped dras-tically at the first trial following a task switch (~15)This is not surprising because the monkeys were notexplicitly informed when the task would switch andtherefore almost always continued to execute thepreviously rewarded behavior Indeed the perform-ance after the switch would have been identical tothe last trials in the previous block had the task notswitched The performance then gradually recoveredover the next 5 to 10 trials Interestingly monkeysperformed slightly better for a few trials after thebehavioral rule switched to antisaccades compared towhen it switched to prosaccades (red line above blueline after the task switch) This indicates that it waseasier for the animals to switch from the lsquolsquoautomaticrsquorsquoprosaccade task to the lsquolsquocontrolledrsquorsquo antisaccade taskthan vice versa
Monkey R had faster saccadic reaction times (SRTs)for prosaccades than antisaccades (median pro-SRT181 msec median anti-SRT 255 msec) whereas mon-key K (median pro-SRT 223 msec median anti-SRT221) and monkey P (median pro-SRT 203 msecmedian anti-SRT 188 msec) showed no differencesin SRTs between correct pro- and antisaccades Thisresult is somewhat surprising giving the well-knownfinding of longer SRTs for antisaccades than prosac-cades (Munoz amp Everling 2004) A possible explana-tion might be that monkey R was experimentally naıveprior to this study whereas monkeys K and P weresubjects in a number of other paradigms (attentionparadigm Desouza amp Everling 2004 randomly inter-leaved prosaccade and antisaccade paradigm Desouzaamp Everling unpublished data delayed match-to-sampleand conditional pro- and antisaccade paradigm Johnstonamp Everling unpublished data) before participating inthis study
Monkeys often generated prosaccades on antisaccadetrials (monkey K 19 monkey P 11 monkey R 39)and many of these prosaccades had short reaction timesin the range of express saccades In line with previousreports (Bell Everling amp Munoz 2000 Everling Dorrisamp Munoz 1998) monkeys did not generate antisaccadeswith express saccade latencies In contrast with previousstudies all monkeys also made a large number ofantisaccades on prosaccade trials (monkey K 14monkey P 26 monkey R 21) This is not surprisingas the animals in this study were not given an explicitinstruction cue of whether to generate a prosaccade oran antisaccade on a given trial
Figure 2 Performance over blocks and at task reversal (A) Number oftrials (mean plusmn SEM ) required by the three monkeys to reach four out
of four correct responses after a task reversal on prosaccade blocks
(blue) and antisaccade blocks (red) Data are averaged across all
recording sessions (B) Performance at the point of task reversal Theproportion of correct responses (mean plusmn SEM ) is plotted for the 10
trials prior to a task reversal to the 20 trials after a task reversal The
first trial following task reversal is at 0 Data are averaged across all
recording sessions from all three monkeys
Everling and DeSouza 1485
Neural Activity
We recorded the activity of a total of 195 neurons fromthe lateral PFC of three monkeys (36 from monkey K 82from monkey P and 77 from monkey R) while theyperformed between 8 and 27 (median 14) task switchesof alternating blocks of pro- and antisaccade trials andgenerated 388 to 912 correct responses (median 666)
Our initial analysis of the peristimulus histogramsshowed that many neurons had different dischargepatterns between pro- and antisaccade trials Figure 3shows the activity of a PFC neuron that was more activeon prosaccade trials than on antisaccade trials Thisneuron exhibited a higher activity after acquiring centralfixation on prosaccade trials than on antisaccade trialsand it maintained a higher activity level throughout thetrial The neuronrsquos response to contralateral stimulus
presentations was also stronger on prosaccade trialsthan on antisaccade trials
To examine the time-course of the mapping ruleeffect we performed a receiver operating characteristic(ROC) analysis using a sliding 500-msec time windowthat was incremented by 100 msec steps for each of the195 PFC neurons (Figure 4) These ROC values are basedon the absolute difference in discharge rate betweencorrect prosaccade trials and correct antisaccade trialspooled across directions Therefore the values in thisanalysis could range from 05 (no rule information) to10 (perfect discrimination between pro- and antisac-cade trials) To test whether these ROC values weresignificant from chance we also conducted a bootstrapanalysis (see Methods for details) The population ofPFC neurons exhibited significant rule selectivity (solidline above 95th percentile dashed line of shuffled data)
Figure 3 Single PFC neuronwith rule selectivity The
neuron was more active on
prosaccade trials than on
antisaccade trials Each dotindicates the time of an action
potential relative to peripheral
stimulus presentation andeach row represents one trial
Trials are sorted according to
saccadic reaction times (open
circles) Open trianglesindicate the onset of fixation
Superimposed on the rasters
are the average spike density
waveforms (30 msec Gaussian)(see Figure 1 for the recording
location of this neuron)
1486 Journal of Cognitive Neuroscience Volume 17 Number 9
throughout the entire trial period even prior to fixationonset This finding indicates that the activity of PFCneurons is significantly different between prosaccadeand antisaccade trials throughout the trial
Fixation Activity
Next we identified those PFC neurons that exhibitedsignificant differences in their activity between prosac-cade and antisaccade trials during the fixation periodbefore the peripheral stimulus was presented We usedthe last 500 msec immediately prior to peripheral stim-ulus presentation as a measurement of neural fixationactivity During this period the monkeys were fixating acentrally presented white dot on both prosaccade andantisaccade trials Therefore the visual stimulation andthe eye positions were identical between prosaccade andantisaccade trials during this period For each neuronwe computed an ROC value on the activity in this epochand compared it with the 95th percentile ROC value for
randomly shuff led data in a bootstrap analysis (seeMethods for details) In this analysis 26 of the neurons(51195) showed a significant difference between fixa-tion activity on prosaccade trials compared with antisac-cade trials ( p lt 05) These PFC neurons had relativelylow discharge rates during the fixation period (026ndash139 spikessec mean = 414 SD = 355 spikessec) andthe differences in neural activity between prosaccadeand antisaccade trials were therefore usually only in theorder of a few spikes per second These changes never-theless represented a considerable modulation in theneuronsrsquo discharge rate between prosaccade and anti-saccade trials The percent change in fixation activity forthese neurons ranged between 70 and 882 with amean difference of 288 (SD = 184)
We found slightly more neurons that preferred pro-saccades than antisaccades (32 of 51 or 627 weremore active for prosaccades 19 of 51 or 373 weremore active for antisaccades) This difference ap-proached significance (x2 test p = 069) We repeated
Figure 4 Time-course of rule
selectivity (A) The time-course
was determined by the sliding
ROC analysis (window500 msec centered at time
point) across all 195 neurons
from which we recorded ROCvalues for the comparison of
correct prosaccades and
correct antisaccade trials
Neurons were sortedaccording to their mean ROC
value in the period from
1000 msec before fixation
onset to 1500 msec afterfixation onset The first and
second vertical lines indicate
the randomly chosen intervalwhen the monkeys started
central fixation and the third
vertical line indicates the time
point when the peripheralstimulus was presented (B)
Average time-course of ROC
values for all 195 neurons
(solid line) and for the medianand 95th percentile of the
shuff led data (dashed lines)
The average ROC value for thepopulation was significantly
different from chance ( p lt
05) throughout the entire
analysis period
Everling and DeSouza 1487
this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9
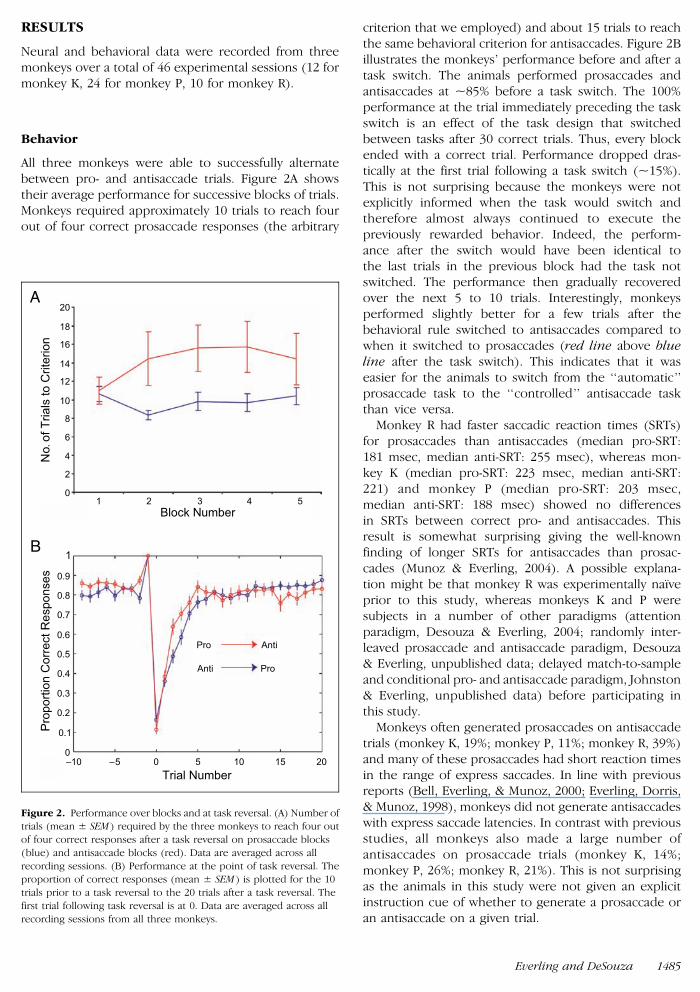
RESULTS
Neural and behavioral data were recorded from threemonkeys over a total of 46 experimental sessions (12 formonkey K 24 for monkey P 10 for monkey R)
Behavior
All three monkeys were able to successfully alternatebetween pro- and antisaccade trials Figure 2A showstheir average performance for successive blocks of trialsMonkeys required approximately 10 trials to reach fourout of four correct prosaccade responses (the arbitrary
criterion that we employed) and about 15 trials to reachthe same behavioral criterion for antisaccades Figure 2Billustrates the monkeysrsquo performance before and after atask switch The animals performed prosaccades andantisaccades at ~85 before a task switch The 100performance at the trial immediately preceding the taskswitch is an effect of the task design that switchedbetween tasks after 30 correct trials Thus every blockended with a correct trial Performance dropped dras-tically at the first trial following a task switch (~15)This is not surprising because the monkeys were notexplicitly informed when the task would switch andtherefore almost always continued to execute thepreviously rewarded behavior Indeed the perform-ance after the switch would have been identical tothe last trials in the previous block had the task notswitched The performance then gradually recoveredover the next 5 to 10 trials Interestingly monkeysperformed slightly better for a few trials after thebehavioral rule switched to antisaccades compared towhen it switched to prosaccades (red line above blueline after the task switch) This indicates that it waseasier for the animals to switch from the lsquolsquoautomaticrsquorsquoprosaccade task to the lsquolsquocontrolledrsquorsquo antisaccade taskthan vice versa
Monkey R had faster saccadic reaction times (SRTs)for prosaccades than antisaccades (median pro-SRT181 msec median anti-SRT 255 msec) whereas mon-key K (median pro-SRT 223 msec median anti-SRT221) and monkey P (median pro-SRT 203 msecmedian anti-SRT 188 msec) showed no differencesin SRTs between correct pro- and antisaccades Thisresult is somewhat surprising giving the well-knownfinding of longer SRTs for antisaccades than prosac-cades (Munoz amp Everling 2004) A possible explana-tion might be that monkey R was experimentally naıveprior to this study whereas monkeys K and P weresubjects in a number of other paradigms (attentionparadigm Desouza amp Everling 2004 randomly inter-leaved prosaccade and antisaccade paradigm Desouzaamp Everling unpublished data delayed match-to-sampleand conditional pro- and antisaccade paradigm Johnstonamp Everling unpublished data) before participating inthis study
Monkeys often generated prosaccades on antisaccadetrials (monkey K 19 monkey P 11 monkey R 39)and many of these prosaccades had short reaction timesin the range of express saccades In line with previousreports (Bell Everling amp Munoz 2000 Everling Dorrisamp Munoz 1998) monkeys did not generate antisaccadeswith express saccade latencies In contrast with previousstudies all monkeys also made a large number ofantisaccades on prosaccade trials (monkey K 14monkey P 26 monkey R 21) This is not surprisingas the animals in this study were not given an explicitinstruction cue of whether to generate a prosaccade oran antisaccade on a given trial
Figure 2 Performance over blocks and at task reversal (A) Number oftrials (mean plusmn SEM ) required by the three monkeys to reach four out
of four correct responses after a task reversal on prosaccade blocks
(blue) and antisaccade blocks (red) Data are averaged across all
recording sessions (B) Performance at the point of task reversal Theproportion of correct responses (mean plusmn SEM ) is plotted for the 10
trials prior to a task reversal to the 20 trials after a task reversal The
first trial following task reversal is at 0 Data are averaged across all
recording sessions from all three monkeys
Everling and DeSouza 1485
Neural Activity
We recorded the activity of a total of 195 neurons fromthe lateral PFC of three monkeys (36 from monkey K 82from monkey P and 77 from monkey R) while theyperformed between 8 and 27 (median 14) task switchesof alternating blocks of pro- and antisaccade trials andgenerated 388 to 912 correct responses (median 666)
Our initial analysis of the peristimulus histogramsshowed that many neurons had different dischargepatterns between pro- and antisaccade trials Figure 3shows the activity of a PFC neuron that was more activeon prosaccade trials than on antisaccade trials Thisneuron exhibited a higher activity after acquiring centralfixation on prosaccade trials than on antisaccade trialsand it maintained a higher activity level throughout thetrial The neuronrsquos response to contralateral stimulus
presentations was also stronger on prosaccade trialsthan on antisaccade trials
To examine the time-course of the mapping ruleeffect we performed a receiver operating characteristic(ROC) analysis using a sliding 500-msec time windowthat was incremented by 100 msec steps for each of the195 PFC neurons (Figure 4) These ROC values are basedon the absolute difference in discharge rate betweencorrect prosaccade trials and correct antisaccade trialspooled across directions Therefore the values in thisanalysis could range from 05 (no rule information) to10 (perfect discrimination between pro- and antisac-cade trials) To test whether these ROC values weresignificant from chance we also conducted a bootstrapanalysis (see Methods for details) The population ofPFC neurons exhibited significant rule selectivity (solidline above 95th percentile dashed line of shuffled data)
Figure 3 Single PFC neuronwith rule selectivity The
neuron was more active on
prosaccade trials than on
antisaccade trials Each dotindicates the time of an action
potential relative to peripheral
stimulus presentation andeach row represents one trial
Trials are sorted according to
saccadic reaction times (open
circles) Open trianglesindicate the onset of fixation
Superimposed on the rasters
are the average spike density
waveforms (30 msec Gaussian)(see Figure 1 for the recording
location of this neuron)
1486 Journal of Cognitive Neuroscience Volume 17 Number 9
throughout the entire trial period even prior to fixationonset This finding indicates that the activity of PFCneurons is significantly different between prosaccadeand antisaccade trials throughout the trial
Fixation Activity
Next we identified those PFC neurons that exhibitedsignificant differences in their activity between prosac-cade and antisaccade trials during the fixation periodbefore the peripheral stimulus was presented We usedthe last 500 msec immediately prior to peripheral stim-ulus presentation as a measurement of neural fixationactivity During this period the monkeys were fixating acentrally presented white dot on both prosaccade andantisaccade trials Therefore the visual stimulation andthe eye positions were identical between prosaccade andantisaccade trials during this period For each neuronwe computed an ROC value on the activity in this epochand compared it with the 95th percentile ROC value for
randomly shuff led data in a bootstrap analysis (seeMethods for details) In this analysis 26 of the neurons(51195) showed a significant difference between fixa-tion activity on prosaccade trials compared with antisac-cade trials ( p lt 05) These PFC neurons had relativelylow discharge rates during the fixation period (026ndash139 spikessec mean = 414 SD = 355 spikessec) andthe differences in neural activity between prosaccadeand antisaccade trials were therefore usually only in theorder of a few spikes per second These changes never-theless represented a considerable modulation in theneuronsrsquo discharge rate between prosaccade and anti-saccade trials The percent change in fixation activity forthese neurons ranged between 70 and 882 with amean difference of 288 (SD = 184)
We found slightly more neurons that preferred pro-saccades than antisaccades (32 of 51 or 627 weremore active for prosaccades 19 of 51 or 373 weremore active for antisaccades) This difference ap-proached significance (x2 test p = 069) We repeated
Figure 4 Time-course of rule
selectivity (A) The time-course
was determined by the sliding
ROC analysis (window500 msec centered at time
point) across all 195 neurons
from which we recorded ROCvalues for the comparison of
correct prosaccades and
correct antisaccade trials
Neurons were sortedaccording to their mean ROC
value in the period from
1000 msec before fixation
onset to 1500 msec afterfixation onset The first and
second vertical lines indicate
the randomly chosen intervalwhen the monkeys started
central fixation and the third
vertical line indicates the time
point when the peripheralstimulus was presented (B)
Average time-course of ROC
values for all 195 neurons
(solid line) and for the medianand 95th percentile of the
shuff led data (dashed lines)
The average ROC value for thepopulation was significantly
different from chance ( p lt
05) throughout the entire
analysis period
Everling and DeSouza 1487
this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

Neural Activity
We recorded the activity of a total of 195 neurons fromthe lateral PFC of three monkeys (36 from monkey K 82from monkey P and 77 from monkey R) while theyperformed between 8 and 27 (median 14) task switchesof alternating blocks of pro- and antisaccade trials andgenerated 388 to 912 correct responses (median 666)
Our initial analysis of the peristimulus histogramsshowed that many neurons had different dischargepatterns between pro- and antisaccade trials Figure 3shows the activity of a PFC neuron that was more activeon prosaccade trials than on antisaccade trials Thisneuron exhibited a higher activity after acquiring centralfixation on prosaccade trials than on antisaccade trialsand it maintained a higher activity level throughout thetrial The neuronrsquos response to contralateral stimulus
presentations was also stronger on prosaccade trialsthan on antisaccade trials
To examine the time-course of the mapping ruleeffect we performed a receiver operating characteristic(ROC) analysis using a sliding 500-msec time windowthat was incremented by 100 msec steps for each of the195 PFC neurons (Figure 4) These ROC values are basedon the absolute difference in discharge rate betweencorrect prosaccade trials and correct antisaccade trialspooled across directions Therefore the values in thisanalysis could range from 05 (no rule information) to10 (perfect discrimination between pro- and antisac-cade trials) To test whether these ROC values weresignificant from chance we also conducted a bootstrapanalysis (see Methods for details) The population ofPFC neurons exhibited significant rule selectivity (solidline above 95th percentile dashed line of shuffled data)
Figure 3 Single PFC neuronwith rule selectivity The
neuron was more active on
prosaccade trials than on
antisaccade trials Each dotindicates the time of an action
potential relative to peripheral
stimulus presentation andeach row represents one trial
Trials are sorted according to
saccadic reaction times (open
circles) Open trianglesindicate the onset of fixation
Superimposed on the rasters
are the average spike density
waveforms (30 msec Gaussian)(see Figure 1 for the recording
location of this neuron)
1486 Journal of Cognitive Neuroscience Volume 17 Number 9
throughout the entire trial period even prior to fixationonset This finding indicates that the activity of PFCneurons is significantly different between prosaccadeand antisaccade trials throughout the trial
Fixation Activity
Next we identified those PFC neurons that exhibitedsignificant differences in their activity between prosac-cade and antisaccade trials during the fixation periodbefore the peripheral stimulus was presented We usedthe last 500 msec immediately prior to peripheral stim-ulus presentation as a measurement of neural fixationactivity During this period the monkeys were fixating acentrally presented white dot on both prosaccade andantisaccade trials Therefore the visual stimulation andthe eye positions were identical between prosaccade andantisaccade trials during this period For each neuronwe computed an ROC value on the activity in this epochand compared it with the 95th percentile ROC value for
randomly shuff led data in a bootstrap analysis (seeMethods for details) In this analysis 26 of the neurons(51195) showed a significant difference between fixa-tion activity on prosaccade trials compared with antisac-cade trials ( p lt 05) These PFC neurons had relativelylow discharge rates during the fixation period (026ndash139 spikessec mean = 414 SD = 355 spikessec) andthe differences in neural activity between prosaccadeand antisaccade trials were therefore usually only in theorder of a few spikes per second These changes never-theless represented a considerable modulation in theneuronsrsquo discharge rate between prosaccade and anti-saccade trials The percent change in fixation activity forthese neurons ranged between 70 and 882 with amean difference of 288 (SD = 184)
We found slightly more neurons that preferred pro-saccades than antisaccades (32 of 51 or 627 weremore active for prosaccades 19 of 51 or 373 weremore active for antisaccades) This difference ap-proached significance (x2 test p = 069) We repeated
Figure 4 Time-course of rule
selectivity (A) The time-course
was determined by the sliding
ROC analysis (window500 msec centered at time
point) across all 195 neurons
from which we recorded ROCvalues for the comparison of
correct prosaccades and
correct antisaccade trials
Neurons were sortedaccording to their mean ROC
value in the period from
1000 msec before fixation
onset to 1500 msec afterfixation onset The first and
second vertical lines indicate
the randomly chosen intervalwhen the monkeys started
central fixation and the third
vertical line indicates the time
point when the peripheralstimulus was presented (B)
Average time-course of ROC
values for all 195 neurons
(solid line) and for the medianand 95th percentile of the
shuff led data (dashed lines)
The average ROC value for thepopulation was significantly
different from chance ( p lt
05) throughout the entire
analysis period
Everling and DeSouza 1487
this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

throughout the entire trial period even prior to fixationonset This finding indicates that the activity of PFCneurons is significantly different between prosaccadeand antisaccade trials throughout the trial
Fixation Activity
Next we identified those PFC neurons that exhibitedsignificant differences in their activity between prosac-cade and antisaccade trials during the fixation periodbefore the peripheral stimulus was presented We usedthe last 500 msec immediately prior to peripheral stim-ulus presentation as a measurement of neural fixationactivity During this period the monkeys were fixating acentrally presented white dot on both prosaccade andantisaccade trials Therefore the visual stimulation andthe eye positions were identical between prosaccade andantisaccade trials during this period For each neuronwe computed an ROC value on the activity in this epochand compared it with the 95th percentile ROC value for
randomly shuff led data in a bootstrap analysis (seeMethods for details) In this analysis 26 of the neurons(51195) showed a significant difference between fixa-tion activity on prosaccade trials compared with antisac-cade trials ( p lt 05) These PFC neurons had relativelylow discharge rates during the fixation period (026ndash139 spikessec mean = 414 SD = 355 spikessec) andthe differences in neural activity between prosaccadeand antisaccade trials were therefore usually only in theorder of a few spikes per second These changes never-theless represented a considerable modulation in theneuronsrsquo discharge rate between prosaccade and anti-saccade trials The percent change in fixation activity forthese neurons ranged between 70 and 882 with amean difference of 288 (SD = 184)
We found slightly more neurons that preferred pro-saccades than antisaccades (32 of 51 or 627 weremore active for prosaccades 19 of 51 or 373 weremore active for antisaccades) This difference ap-proached significance (x2 test p = 069) We repeated
Figure 4 Time-course of rule
selectivity (A) The time-course
was determined by the sliding
ROC analysis (window500 msec centered at time
point) across all 195 neurons
from which we recorded ROCvalues for the comparison of
correct prosaccades and
correct antisaccade trials
Neurons were sortedaccording to their mean ROC
value in the period from
1000 msec before fixation
onset to 1500 msec afterfixation onset The first and
second vertical lines indicate
the randomly chosen intervalwhen the monkeys started
central fixation and the third
vertical line indicates the time
point when the peripheralstimulus was presented (B)
Average time-course of ROC
values for all 195 neurons
(solid line) and for the medianand 95th percentile of the
shuff led data (dashed lines)
The average ROC value for thepopulation was significantly
different from chance ( p lt
05) throughout the entire
analysis period
Everling and DeSouza 1487
this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

this analysis with different periods (eg 200 and1000 msec prior to peripheral stimulus onset) and ob-tained very similar results
Neurons with differences in their activity during thefixation period were found both dorsal and ventral ofthe principal sulcus with no clear topography (Figure 1A)
To investigate how the rule selectivity evolved overthe course of a block we computed the activity in10 bins of three successive correct trials for the 19 neu-rons that preferred antisaccades (Figure 5A) and for the32 neurons that preferred prosaccades (Figure 5B) Thefigure shows that these neurons had a different activitybetween pro- and antisaccade blocks already on thefirst three trials after a task switch One-way repeated-measures of analysis of variance (ANOVA) showed nodifferences between the activity in the 10 bins for thepopulation of 19 neurons that preferred antisaccadeson prosaccade blocks [F(19) = 079 p = 62] or antisac-cade blocks [F(19) = 055 p = 84] or the populationof 32 neurons that preferred prosaccades on prosac-cade blocks [F(19) = 099 p = 45] or antisaccadeblocks [F(19) = 059 p = 81] This indicates that rule-selectivity did not change over the course of task block
We next tested whether PFC neurons with differencesin their fixation period activity between blocks of correctpro- and correct antisaccades also showed differences inactivity between correct trials and error trials withinthese blocks Figure 6A and B shows the results of thisanalysis for the 19 neurons that had higher fixationperiod activity levels for antisaccades than prosaccadesThese neurons had the same activity on error antisac-cade trials (423 plusmn 094 spikessec) compared with cor-rect antisaccade trials (443 plusmn 085 spikessec) (pairedt test p = 65) Their activity levels were significantlyhigher on error antisaccade trials than on correct pro-saccade trials (345 plusmn 073 spikessec) (paired t testp lt 005) Error prosaccade trials were associated withmore fixation activity (427 plusmn 086 spikessec) thancorrect prosaccade trials (paired t test p lt 05) Theactivity did not differ between error prosaccades andcorrect antisaccades (paired t test p = 85)
The analysis of the fixation activity levels of the32 neurons with higher activity levels for prosaccadesthan antisaccades yielded similar results (Figure 6C andD) The neurons were significantly more active (pairedt test p lt 05) on error antisaccade trials (401 plusmn059 spikessec) than on correct antisaccade trials(341 plusmn 050 spikessec) The activity was not differentbetween error antisaccade trials and correct prosaccadetrials (441plusmn 062 spikessec) ( p = 18) On errorprosaccade trials neurons were significantly less active(375 plusmn 055 spikessec) than on correct prosaccadetrials (paired t test p lt 05) but the activity did notdiffer between error prosaccade trials and correct anti-saccade trials (paired t test p = 47)
It should be noted that in all these comparisonsneural activity was highly correlated between the con-ditions (see correlation coefficients r in Figure 6) Thisindicates that neural activity was modulated fairly con-sistently within the population of rule-selective PFCneurons
To test how well the fixation activity of these neuronson a single trial could predict the performance of themonkey within a block of trials we computed ROCvalues for the 51 neurons with rule-selective fixationactivity In this analysis we tested whether a neuronthat for example preferred prosaccade trials would alsobe less active on error prosaccade trials on whichmonkeys generated antisaccades than on correct pro-saccade trials The ROC values could therefore rangefrom 0 indicating that the neuron is more active forerror prosaccades than correct prosaccades on all thetrials to 1 indicating that the neuron is less active forerror prosaccades than correct prosaccades on all thetrials An ROC value of 05 indicates that the neuron hadthe same fixation activity for correct prosaccade trialsand error prosaccade trials Figure 7A and B shows thedistribution of ROC values for all 51 neurons forprosaccade blocks (Figure 7A) and antisaccade blocks(Figure 7B) Both distributions were significantly differ-
Figure 5 Activity of rule-selective neurons on correct trials over thecourse of blocks (A) Mean activity for the 19 neurons that were more
active for antisaccades than prosaccades in bins of three successive
correct trials for antisaccade blocks (red line circles) and prosaccade
blocks (blue line squares) Error bars are standard errors of thedifferences (B) Same as (A) but for the 32 neurons which were more
active for prosaccades
1488 Journal of Cognitive Neuroscience Volume 17 Number 9
ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

ent from 5 (Wilcoxon signed-rank tests p lt 0001 forprosaccades p lt 0005 for antisaccades) indicating thatthe population of PFC neurons carries informationabout the behavior of monkeys on a block of trials
In the error trial analysis presented above we com-pared all correct trials with all error trials within a blockThe first 5ndash10 trials in each block however may becontaminated by a trial-and-error strategy To testwhether PFC neurons display differences between thisearly period and the late period we compared correcttrials and error trials separately in the period from thetask switch to the 10th correct trial (early period) and inthe remainder of the block (late period) The distribu-tions of the ROC values for the early period and lateperiod are presented in Figure 7CndashF In the early periodthe distribution of ROC values was different from 5for prosaccades (Wilcoxon signed-rank test p lt 05Figure 7C) but not for antisaccades (Wilcoxon signed-rank test p = 14 Figure 7D) In the late period both dis-tributions were significantly different from 5 (Wilcoxonsigned rank tests p lt 0001 for prosaccades Figure 7Ep lt 0005 for antisaccades Figure 7F) These findings
indicate that the activity of PFC neurons is predictive oferrors on prosaccade trials (ie antisaccades) through-out the whole block whereas PFC neurons do notpredict errors on antisaccade trials (ie prosaccades)in the early period following the task switch
To test whether differences in the SRTs of antisaccadeerrors could account for these findings we comparedSRTs of errors on antisaccade trials between the earlyperiod and the late period Errors were significantlyfaster in the early period (mean 232 plusmn 3 msec median212 msec) than in the late period (mean 244 plusmn 3 msecmedian 229 msec) (t test p lt 005)
Presaccade Activity
The prosaccade task and the antisaccade task requirediametrically opposite mapping rules In the prosaccadetask the stimulus location is also the saccade targetlocation whereas it acts as both a cue for the antisac-cade to its mirror location and as a distractor for anincorrect prosaccade To discern whether the presac-cade activity of PFC neurons ref lected the stimulus
Figure 6 Comparison of the
fixation activity on correct trials
and error trials (A) Mean
activity (plusmnSEM ) in the last500 msec prior to stimulus
presentation of 19 individual
neurons that had significantlyhigher activity level on correct
antisaccade than on correct
prosaccade trials (B) The
activity is plotted beforeerrors (abscissa) against the
activity levels before correct
responses (ordinate) The
dashed lines indicate the unityline (slope = 1) and the solid
lines indicate the least square
fit (CD) Same as (AB) butfor 32 neurons that had
significantly higher activity
level on correct prosaccade
than on correct antisaccadetrials
Everling and DeSouza 1489
locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

locations saccade directions and or the rules wecomputed two-way ANOVAs on the activity in the250 msec immediately prior to saccade onset for eachneuron We chose this period to exclude any activityafter the saccade (eg visual feedback) The factors inthis analysis were Stimulus location (ipsilateral or con-tralateral) and Saccade direction (ipsilateral or contra-lateral) A potential problem with this analysis is thatdifferent SRTs between the task conditions result indifferent amounts of visual activation to be included inthe analysis epoch To reduce this possibility we choosethe rather long analysis epoch of 250 msec whichcaptured the visual activation for the majority of trials
A few neurons exhibited activity selective for the stim-ulus location but not for saccade direction Figure 8Ashows the activity of such a neuron The neuron re-sponded to the presentation of the peripheral stimulusin the contralateral visual field but not the presentation inthe ipsilateral field The activity of this neuron was notmodulated by the direction of the saccade A small num-ber of neurons showed a significant effect of stimuluslocation but not of saccade direction or the interaction(11195 or 56 evaluated at p lt 05) This number wasnot significantly different from chance (x2 test p gt 05)
Some other neurons showed activity selective for thesaccade direction independent of the location of the
Figure 7 Discrimination
between correct trials and
error trials (A B) Distribution
of ROC values for 51 rule-selective neurons in the period
500 msec prior to stimulus
onset for the comparison of allcorrect trials with all error trials
on prosaccade blocks (A) and
antisaccade blocks (B) An ROC
value of 05 indicates thatneurons had the same activity
on correct trials and error
trials (CD) Same as (AB) but
for the early period after a taskswitch (beginning of block to
10th correct trial) (EF) Same
as (AB) but for the late periodafter a task switch (after the
10th correct trial to the 30th
correct trial)
1490 Journal of Cognitive Neuroscience Volume 17 Number 9
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9
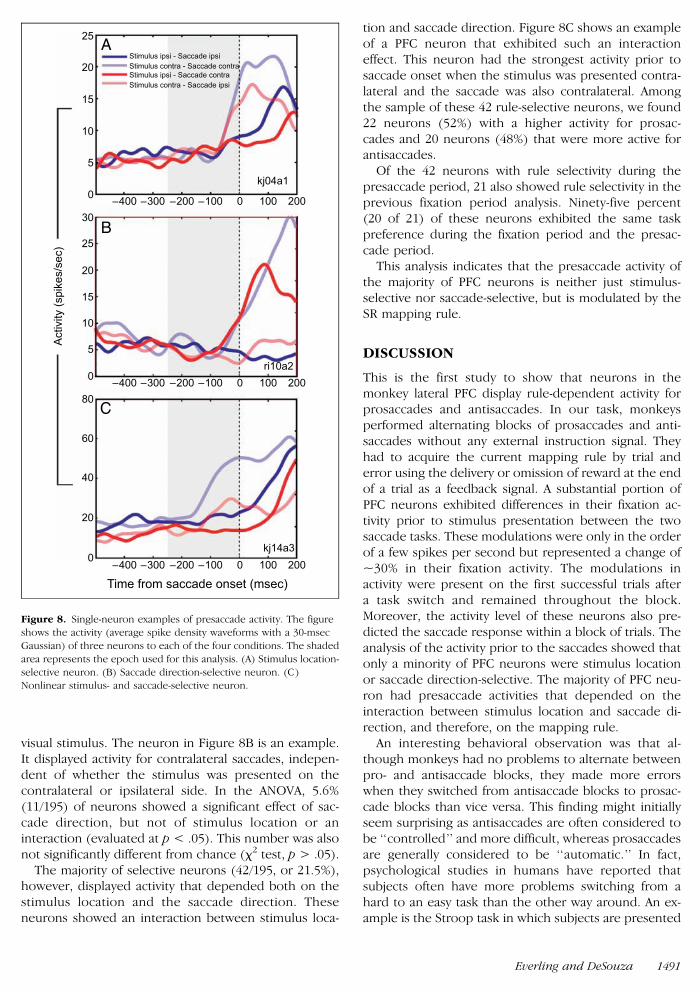
visual stimulus The neuron in Figure 8B is an exampleIt displayed activity for contralateral saccades indepen-dent of whether the stimulus was presented on thecontralateral or ipsilateral side In the ANOVA 56(11195) of neurons showed a significant effect of sac-cade direction but not of stimulus location or aninteraction (evaluated at p lt 05) This number was alsonot significantly different from chance (x2 test p gt 05)
The majority of selective neurons (42195 or 215)however displayed activity that depended both on thestimulus location and the saccade direction Theseneurons showed an interaction between stimulus loca-
tion and saccade direction Figure 8C shows an exampleof a PFC neuron that exhibited such an interactioneffect This neuron had the strongest activity prior tosaccade onset when the stimulus was presented contra-lateral and the saccade was also contralateral Amongthe sample of these 42 rule-selective neurons we found22 neurons (52) with a higher activity for prosac-cades and 20 neurons (48) that were more active forantisaccades
Of the 42 neurons with rule selectivity during thepresaccade period 21 also showed rule selectivity in theprevious fixation period analysis Ninety-five percent(20 of 21) of these neurons exhibited the same taskpreference during the fixation period and the presac-cade period
This analysis indicates that the presaccade activity ofthe majority of PFC neurons is neither just stimulus-selective nor saccade-selective but is modulated by theSR mapping rule
DISCUSSION
This is the first study to show that neurons in themonkey lateral PFC display rule-dependent activity forprosaccades and antisaccades In our task monkeysperformed alternating blocks of prosaccades and anti-saccades without any external instruction signal Theyhad to acquire the current mapping rule by trial anderror using the delivery or omission of reward at the endof a trial as a feedback signal A substantial portion ofPFC neurons exhibited differences in their fixation ac-tivity prior to stimulus presentation between the twosaccade tasks These modulations were only in the orderof a few spikes per second but represented a change of~30 in their fixation activity The modulations inactivity were present on the first successful trials aftera task switch and remained throughout the blockMoreover the activity level of these neurons also pre-dicted the saccade response within a block of trials Theanalysis of the activity prior to the saccades showed thatonly a minority of PFC neurons were stimulus locationor saccade direction-selective The majority of PFC neu-ron had presaccade activities that depended on theinteraction between stimulus location and saccade di-rection and therefore on the mapping rule
An interesting behavioral observation was that al-though monkeys had no problems to alternate betweenpro- and antisaccade blocks they made more errorswhen they switched from antisaccade blocks to prosac-cade blocks than vice versa This finding might initiallyseem surprising as antisaccades are often considered tobe lsquolsquocontrolledrsquorsquo and more difficult whereas prosaccadesare generally considered to be lsquolsquoautomaticrsquorsquo In factpsychological studies in humans have reported thatsubjects often have more problems switching from ahard to an easy task than the other way around An ex-ample is the Stroop task in which subjects are presented
Figure 8 Single-neuron examples of presaccade activity The figureshows the activity (average spike density waveforms with a 30-msec
Gaussian) of three neurons to each of the four conditions The shaded
area represents the epoch used for this analysis (A) Stimulus location-
selective neuron (B) Saccade direction-selective neuron (C)Nonlinear stimulus- and saccade-selective neuron
Everling and DeSouza 1491
with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

with color words printed in colors (Stroop 1935) Sub-jects generally have more problems to ignore the wordsand name the color (color-naming task) than to ignorethe color and name the words (word-naming task)Allport Styles and Hsieh (1994) however showed thatsubjects show larger switch costs when they have toswitch from color-naming to word-naming than viceversa The authors therefore proposed the task-setinertia (TSI) hypothesis According to this hypothesisswitch costs result from the persistent activation ofprevious task processes In the case of our taskmonkeys had to strongly suppress prosaccades towardsthe peripheral stimulus on antisaccade blocks and thisinhibition then persisted for a few trials The TSI hy-pothesis does not predict similar switch costs for pro-saccade to antisaccade blocks because monkeys did notnormally have to inhibit antisaccades during the prosac-cade blocks Therefore there should be no persistinginhibition of antisaccades from the prosaccade blocks
The PFC has been implicated in the performance ofantisaccades by many studies Patients with discretelesions of the dorsolateral PFC have difficulties suppress-ing the prepotent prosaccade in the antisaccade task(Pierrot-Deseilligny Muri et al 2003 Walker HusainHodgson Harrison amp Kennard 1998 Pierrot-DeseillignyRivaud Gaymard amp Agid 1991) Several functional imag-ing studies have also reported an increased activation ofthe dorsolateral PFC for antisaccades compared with pro-saccades (Desouza Menon amp Everling 2003 Doricchiet al 1997 Sweeney et al 1996) A recent functional mag-netic resonance imaging study that compared BOLDsignals associated with antisaccade task performancein schizophrenic patients and healthy controls foundan increased BOLD signal in the dorsolateral PFC forcontrol subjects but not for schizophrenic patients(McDowell et al 2002) suggesting that abnormalitiesin dorsolateral PFC may play an important role in im-paired antisaccade task performance
Single-neuron recording studies in monkeys per-forming an antisaccade task indicate that correct taskperformance requires the activation of fixation neuronsand the suppression of saccade neurons in the frontaleye fields (FEFs) (Everling amp Munoz 2000) and the su-perior colliculus (SC) (Everling Dorris Klein amp Munoz1999 Everling Dorris amp Munoz 1998) before stimuluspresentation It has been proposed that without suffi-cient inhibition the incoming visual signal could sumwith an elevated prestimulus activity and directly passthe neuronrsquos threshold for saccade initiation (Munoz ampEverling 2004) The result would be a reflexive saccadethat moves the eyes toward the stimulus instead ofaway from it Several brain areas in the frontal lobe(Pierrot-Deseilligny Muri et al 2003 Gaymard PlonerRivaud Vermersch amp Pierrot-Deseilligny 1998 GuittonBuchtel amp Douglas 1985) and basal ganglia (BriandStrallow Hening Poizner amp Sereno 1999 Lasker amp Zee1997) have been proposed as putative sources of this
top-down suppression signal A possible role of thesupplementary eye fields in the suppression of reflexivesaccades has been demonstrated by single-neuron re-cordings in monkeys (Amador Schlag-Rey amp Schlag2003 Schlag-Rey Amador Sanchez amp Schlag 1997)
All previous antisaccade studies in monkeys have usedexternal cues (either the color or the shape of thefixation stimulus) to instruct the animals either togenerate a prosaccade or an antisaccade on a given trial(Amador et al 2003 Zhang amp Barash 2000 2003 Bellet al 2000 Everling amp Munoz 2000 Everling DorrisKlein et al 1999 Gottlieb amp Goldberg 1999 AmadorSchlag-Rey amp Schlag 1998 Schlag-Rey et al 1997Funahashi Chafee amp Goldman-Rakic 1993) In our taskdesign no external cues were given and monkeys wererequired to keep track of which response behavior wascurrently rewarded This requirement is somewhat sim-ilar to the pro- and antisaccades tasks employed in mosthuman studies that are run in separate blocks with onlyan explicit instruction (usually verbal) before partici-pants begin The modulations of neural active prior tostimulus presentation during the fixation period andoften even before the monkeys started fixation mayreflect a neural correlate for the maintenance of thecurrent rule in place by PFC neurons Our analysisfurther showed that PFC neurons modulated their ac-tivity already on the first correct trials following a taskswitch and they maintained this modulation throughoutthe entire block of trials
Further the activity level of PFC neurons did not onlydiffer between prosaccade trials and antisaccade trialsbut also between correct trials and error trials Thisfinding supports the idea that the activity levels of theseneurons influenced the monkeysrsquo task performanceInterestingly the activity of PFC neurons did not distin-guish between correct antisaccades and errors duringthe early trial period This suggests that processes inother areas than the PFC were involved in the genera-tion of these incorrect saccades towards the stimulus onantisaccade trials The findings of faster SRTs of errors inthe early period suggest that they might have been theresult of an increased motor preparation in the earlyperiod after the switch from prosaccades to antisac-cades There is some evidence that the preparatoryprestimulus activity in the SC changes continuously ina manner consistent with the task demands (DorrisPare amp Munoz 2000) In accordance with these findingswe hypothesize that SC saccade neurons have highlevels of preparatory activity at the end of a prosaccadeblock and it is likely that it requires several trials beforethis activity is reduced An increased level of preparatorysaccade activity might be responsible for some of theerrors in the early period and not a wrong representa-tion of the current rule of PFC neurons
Our finding of differences in neural activity during thefixation period is similar to findings reported by Asaadet al (2000) who showed differences in the fixation
1492 Journal of Cognitive Neuroscience Volume 17 Number 9
activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

activity of PFC neurons between an object task anassociative task and a spatial task Indeed the averagemodulations of fixation activity in task-selective neu-rons were comparable to those we found betweenprosaccades and antisaccades (320 by Asaad andcolleagues and 288 in this study) The authorshypothesized that these differences could providetop-down signals that bias the activity of sensorysystems toward the representation of relevant informa-tion (Desimone amp Duncan 1995) or it could beinvolved in the retrieval of information from long-termmemory (Tomita Ohbayashi Nakahara Hasegawa ampMiyashita 1999)
The PFC however has also direct projections to theSC (Leichnetz Spencer Hardy amp Astruc 1981 Goldmanamp Nauta 1976) and FEFs (Selemon amp Goldman-Rakic1988) Single-neuron recordings have demonstratedelevated fixation neuron activity and reduced saccadeneuron activity during the fixation period on antisaccadetrials compared with prosaccade trials in both areas Itseems conceivable that different fixation period activitylevels in PFC neurons could directly modulate theactivity of neurons in the SC and FEFs via cortico-tectaland cortico-cortical projections This hypothesis is inline with a recent model of the PFC in cognitive control(Miller amp Cohen 2001) According to this model thePFC provides bias signals to other brain areas which helpto guide the flow of neural activity along the pathwaysthat are needed for correct task performance In thecase of antisaccade this would require the increase offixation neuron activity in the SC and FEFs and thereduction of preparatory saccade neuron activity beforethe stimulus appears (Everling amp Munoz 2000 EverlingDorris Klein et al 1999 Everling Dorris amp Munoz1998) In the case of a prosaccade a reduced fixationactivity and an increased preparatory saccade activitywould be beneficial as it would decrease SRTs (MunozDorris Pare amp Everling 2000) We hypothesize thatsome of those PFC neurons with higher activity forantisaccades project predominately to fixation neuronswhereas those PFC neurons with higher activity forprosaccades send projections to saccade neurons Anti-dromic identification studies are necessary to directlytest this hypothesis
The correct performance of the antisaccade task doesnot just require the suppression of the prosaccade butalso the inversion of the saccade target vector Thisprocess is not needed for prosaccades as the visualstimulus activates in this case the correct population ofsaccade neurons in the SC and FEFs In the antisaccadetask the initial visual activity must be suppressed andsaccade neurons must be activated in the oppositehemisphere Our findings demonstrate that the presac-cadic activity of the majority of PFC neurons was neitherjust visual-location nor saccade-direction selective butrule-dependent This activity could provide the signal tothe SC and FEFs that allows a rule-dependent mapping
of sensory information to the appropriate motor output(Miller amp Cohen 2001 Fuster 1991)
It is also possible that our findings reflect a moregeneral role of the PFC in encoding the task that is notspecific for the saccadic motor system This interpreta-tion is supported by several studies that found correlatesfor abstract rules and categories in the PFC (FreedmanRiesenhuber Poggio amp Miller 2001 Wallis et al 2001)Wallis et al (2001) tested monkeys with two successivelypresented pictures and the animals either had to use amatch rule or a nonmatch rule depending on a priorinstruction They found that the majority of neuronscoded the abstract rule and not the objects Freedmanet al (2001) trained monkeys to categorize computer-generated stimuli into the categories lsquolsquocatsrsquorsquo and lsquolsquodogsrsquorsquoPFC neurons showed a sharp categorization of thestimuli irrespective of their exact physical attributes Inthese two studies monkeys had to indicate their re-sponse by either holding or releasing a lever The activityof PFC neurons however was not related to the actualmotor response but to the representation of the abstractrule or category
Taken together our results show rule-dependentactivity for prosaccades and antisaccades in the primatePFC throughout the trial The data clearly demonstratethat the activity of PFC neurons is not just fixation-stimulus- or saccade-related but that it depends on themapping rule necessary for a correct task performanceWe believe that the poor antisaccade task performanceof patients with lesions or dysfunctions of the PFC(Pierrot-Deseilligny Muri et al 2003 Everling amp Fischer1998 Walker et al 1998 Pierrot-Deseilligny Rivaudet al 1991) is the result of a disturbance of these rule-dependent bias signals to saccade-related areas
METHODS
Subjects
Three male rhesus monkeys (Macaca mulatta) weigh-ing between 4 and 9 kg were the subjects in this studyUsing previously described methods the three monkeyswere implanted with head restraints and with recordingchambers (Desouza amp Everling 2004) One monkey wasimplanted with a scleral search coil to monitor eyemovements (Fuchs amp Robinson 1966) whereas aninfrared video eye tracker (ISCAN Boston MA) wasused for the other two monkeys
A recording chamber was placed over the principalsulcus (left hemisphere in two animals right hemi-sphere in one animal AndashP 31 mm MndashL 18 mm) suchthat the principal sulcus and the surrounding cortex wasreadily accessible (Figure 1A) Animals received antibi-otics and analgesics postoperatively and were handled inaccordance with the guidelines of the Canadian Councilon Animal Care policy on the use of laboratory animalsand a protocol approved by the Animal Use Subcom-
Everling and DeSouza 1493
mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

mittee of the University of Western Ontario Council onAnimal Care Monkeys were under the close supervisionof the university veterinarians
Behavioral Task
Monkeys were trained to perform alternating blocks ofprosaccades and antisaccades (Figure 1B) The experi-mental paradigm was presented by running the CORTEXprogram on two Pentium PCs The program also mon-itored the animalsrsquo behavior and controlled the rewarddelivery Each trial started with the presentation of an FP(white dot 173 cdm2 0158) at the center of the blackscreen (001 cdm2) Monkeys were given 2 sec toacquire it and maintain steady fixation within a 058 058 window for a random period of 1100 to 1400 msecA peripheral visual stimulus (0158) was then presentedpseudorandomly with equal probability 88 to the left or88 to the right of the FP The peripheral stimulus was red(159 cdm2 CIEx = 0614 CIEy = 0345) for two of theanimals and green (174 cdm2 CIEx = 0303 CIEy =0585) for the third animal To obtain a reward monkeyshad to generate a saccade within 500 msec after stimuluspresentation to the stimulus location on prosaccadetrials or to its mirror location on antisaccade trials(58 58) They were required to maintain fixation at thetarget location for 200 msec On successful trials a juicereward was given 200 msec after the end of the saccade
Error trials were repeated at the end of each blockAfter 30 correct responses the task switched (eg fromprosaccades to antisaccades and vice versa) without anyexplicit signal to the animals Monkeys received juiceuntil satiation after which they were returned to theirhome cage Records were kept of the weight and healthstatus of the monkeys and additional water and fruitwere provided
Recording Technique
Monkeys were seated comfortably in a primate chairwithin a sound-attenuating chamber with their headsrestrained and a juice-spout placed at their mouth forcomputer-controlled reward delivery (Crist InstrumentsHagerstown MD) The stimuli were presented on a21-in colored computer screen 42 cm in front of theanimals For monkey R horizontal and vertical eyepositions were sampled at 1000 Hz using a magneticsearch coil system (David Northmore Inst Newark DE)whereas a video eye tracker was used to monitor theeye positions at 120 Hz in monkeys P and K
Extracellular recordings were made from the lateralPFC with commercially available dura-puncturing tung-sten microelectrodes (FHC Inst Bowdoinham ME)Arrays of two to eight electrodes were driven withinthe recording chamber by custom-designed screw mini-microdrives that were mounted on a Delrin grid (CristInstruments) The grid array had a 1-mm spacing be-
tween adjacent locations and was visualized in situby MRI and registered with the anatomy (Desouza ampEverling 2004) To ensure a relatively unbiased samplingof PFC neural activity we did not prescreen neurons fortask-related responses Instead we advanced the elec-trodes until the activity of one or more neurons was wellisolated and then data collection started Neural activitywas amplified filtered and stored for off-line clusterseparation applying principal component analysis in 2-Dand 3-D with the Plexon MAP system (Plexon DallasTX) Horizontal and vertical eye positions and theoccurrence of behavioral events (eg start of trial onsetof fixation stimulus presentation) were also stored inthe Plexon MAP system
Data Analysis
Data analysis was performed using custom-designedsoftware programmed in Matlab (Mathworks NatickMA) Saccade onset was defined as the time at whichthe horizontal velocity exceeded 308sec and the end ofthe saccade was defined as the time when the velocityfell below 308sec Incomplete trials (no fixation brokenfixation no response) were excluded automatically fromanalyses of neural activity Each trial was visually in-spected and trials with saccades during the requiredfixation interval were excluded from further analysisTrials with SRTs below 80 msec were excluded as antici-pations Trials with SRTs above 500 msec were excludedas no-response trials
To evaluate the time-course of rule selectivity in thepopulation of PFC neurons we performed a sliding ROCanalysis An ROC analysis measures the degree of over-lap between two distributions For each neuron theactivity on correct prosaccade trials was compared withthe activity on correct antisaccade trials yielding twodistributions of neuronal activity Each point on the ROCcurve was created by plotting the proportion of thedistribution with the higher mean activity that exceededa criterion against the proportion of the distributionwith the lower mean activity To generate the entireROC curve the criterion level was incremented fromzero to the maximal discharge rate in 100 steps The areaunder the ROC curve is a quantitative measure of theseparation of the two distributions A value of 05indicates that the two distributions completely overlapA value of 10 indicates that the two distributions arecompletely separate Starting from 2000 msec prior toperipheral stimulus presentation the ROC value wascalculated for a 500-msec epoch (centered around thetime point) This analysis was repeated in 100-msecincrements until 500 msec after stimulus presentationAn ROC time-course was calculated for each neuron andthen averaged across all 195 PFC to obtain the averageROC time-course To test whether the ROC values weresignificant at any time points we conduced a bootstrapanalysis The ROC analysis described above was repeated
1494 Journal of Cognitive Neuroscience Volume 17 Number 9
1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

1000 times for each time point each time assigning acondition (prosaccade or antisaccade) at random toeach trial Each of the 1000 repetitions of the analysisperformed on all 195 neurons yielded a single averagetime-course The distribution of the 1000 average ROCvalues at each point in time along the time-course wasthen used to determine the 50th and 95th percentilevalues Both were plotted in a figure together with theaverage ROC time-course for the unshuffled data The50th percentile showed the expected result in the caseof the null hypothesis (no encoding of the behavioralrule by the population of PFC neurons) and the 95thpercentile indicated the 5 significance criterion
To test whether individual PFC neurons showed rule-specific activity for prosaccades and antisaccades wefirst calculated an ROC value for the 500-msec epochprior to stimulus presentation To test whether this ROCvalue was significantly different from chance at p lt 05we performed a bootstrap analysis for each neuron TheROC analysis was repeated 10000 times for each neu-ron each time the trials were shuffled and a condition(prosaccade or antisaccade) was assigned randomly Aneuron was classified as being rule-selective if the un-shuffled ROC value exceeded the 95th percentile ROCvalue from the shuffled data
Acknowledgments
We thank Melanie Brimson for technical and surgical supportand Tyrrel DeLangley for excellent veterinary support KevinJohnston and Brian Corneil provided very helpful commentsand suggestions We gratefully acknowledge Joe Gatirsquos and RaviMenonrsquos assistance during MRI scans We also thank theanonymous reviewer for helpful advice on the ROC bootstrapanalysis This research was supported by grants from theCanadian Institutes of Health Research the National Alliancefor Research on Schizophrenia and Depression and thePremierrsquos Research Excellence Program S Everling is aCanadian Institutes of Health Research New Investigator andan EJLB Foundation research scholar
Reprint requests should be sent to S Everling Centre for Brainand Mind Robarts Research Institute 100 Perth Drive LondonOntario N6A 5K8 Canada or via e-mail severlinuwoca
REFERENCES
Allport A Styles E A amp Hsieh S (1994) Shifting intentionalset Exploring the dynamic control of tasks In C Umilta ampM Moscovitch (Eds) Attention and performance XV(pp 421ndash452) Cambridge MIT Press
Amador N Schlag-Rey M amp Schlag J (1998) Primateantisaccades I Behavioral characteristics Journal ofNeurophysiology 80 1775ndash1786
Amador N Schlag-Rey M amp Schlag J (2003) Primateantisaccade II Supplementary eye field neuronal activitypredicts correct performance Journal of Neurophysiology91 1672ndash1689
Asaad W F Rainer G amp Miller E K (2000) Task-specificneural activity in the primate prefrontal cortex Journal ofNeurophysiology 84 451ndash459
Baddeley A (1986) Working memory London OxfordUniversity Press
Bell A H Everling S amp Munoz D P (2000) Influence ofstimulus eccentricity and direction on characteristics ofpro- and antisaccades in non-human primates Journal ofNeurophysiology 84 2595ndash2604
Briand K A Strallow D Hening W Poizner H amp SerenoA B (1999) Control of voluntary and ref lexive saccades inParkinsonrsquos disease Experimental Brain Research 12938ndash48
Desimone R amp Duncan J (1995) Neural mechanisms ofselective visual attention Annual Review of Neuroscience18 193ndash222
Desouza J F amp Everling S (2004) Focused attentionmodulates visual responses in the primate prefrontal cortexJournal of Neurophysiology 91 855ndash862
Desouza J F Menon R S amp Everling S (2003) Preparatoryset associated with pro-saccades and anti-saccades inhumans investigated with event-related FMRI Journal ofNeurophysiology 89 1016ndash1023
Dias R Robbins T W amp Roberts A C (1996) Primateanalogue of the Wisconsin Card Sorting Test Effects ofexcitotoxic lesions of the prefrontal cortex in the marmosetBehavioral Neuroscience 110 872ndash886
Doricchi F Perani D Incoccia C Grassi F Cappa S FBettinardi V Galati G Pizzamiglio L amp Fazio F (1997)Neural control of fast-regular saccades and antisaccades Aninvestigation using positron emission tomographyExperimental Brain Research 116 50ndash62
Dorris M C Pare M amp Munoz D P (2000) Immediateneural plasticity shapes motor performance Journal ofNeuroscience 20 RC52
Everling S Dorris M C Klein R M amp Munoz D P (1999)Role of primate superior colliculus in preparation andexecution of antisaccades and prosaccades Journal ofNeuroscience 19 2740ndash2754
Everling S Dorris M C amp Munoz D P (1998) Reflexsuppression in the antisaccade task is dependent onprestimulus neural processes Journal of Neurophysiology80 1584ndash1589
Everling S amp Fischer B (1998) The antisaccade A review ofbasic research and clinical studies Neuropsychologia 36885ndash899
Everling S amp Munoz D P (2000) Neuronal correlates forpreparatory set associated with pro-saccades andanti-saccades in the primate frontal eye field Journal ofNeuroscience 20 387ndash400
Freedman D J Riesenhuber M Poggio T amp Miller E K(2001) Categorical representation of visual stimuli in theprimate prefrontal cortex Science 291 312ndash316
Fuchs A F amp Robinson D A (1966) A method formeasuring horizontal and vertical eye movementchronically in the monkey Journal of Applied Physiology21 1068ndash1070
Funahashi S Chafee M V amp Goldman-Rakic P S (1993)Prefrontal neuronal activity in rhesus monkeys performing adelayed anti-saccade task Nature 365 753ndash756
Fuster J M (1991) The prefrontal cortex Anatomyphysiology and neuropsychology of the frontal lobeNew York Raven
Gaymard B Ploner C J Rivaud S Vermersch A I ampPierrot-Deseilligny C (1998) Cortical control of saccadesExperimental Brain Research 123 159ndash163
Goldman P S amp Nauta W J (1976) Autoradiographicdemonstration of a projection from prefrontal associationcortex to the superior colliculus in the rhesus monkeyBrain Research 116 145ndash149
Gottlieb J amp Goldberg M E (1999) Activity of neurons in thelateral intraparietal area of the monkey during anantisaccade task Nature Neuroscience 2 906ndash912
Everling and DeSouza 1495
Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9

Guitton D Buchtel H A amp Douglas R M (1985) Frontallobe lesions in man cause difficulties in suppressing ref lexiveglances and in generating goal-directed saccadesExperimental Brain Research 58 455ndash472
Hallett P E (1978) Primary and secondary saccades to goalsdefined by instructions Vision Research 18 1279ndash1296
Hallett P E amp Adams B D (1980) The predictability ofsaccadic latency in a novel voluntary oculomotor taskVision Research 20 329ndash339
Lasker A G amp Zee D S (1997) Ocular motor abnormalitiesin Huntingtonrsquos disease Vision Research 37 3639ndash3645
Leichnetz G R Spencer R F Hardy S G amp Astruc J(1981) The prefrontal corticotectal projection in themonkey an anterograde and retrograde horseradishperoxidase study Neuroscience 6 1023ndash1041
McDowell J E Brown G G Paulus M Martinez A StewartS E Dubowitz D J amp Braff D L (2002) Neural correlatesof refixation saccades and antisaccades in normal andschizophrenia subjects Biological Psychiatry 51 216ndash223
Miller E K amp Cohen J D (2001) An integrative theory ofprefrontal cortex function Annual Review of Neuroscience24 167ndash202
Milner B (1963) Effects of different brain lesions on cardsorting Archives of Neurology 9 90ndash100
Monchi O Petrides M Petre V Worsley K amp Dagher A(2001) Wisconsin Card Sorting revisited Distinct neuralcircuits participating in different stages of the task identifiedby event-related functional magnetic resonance imagingJournal of Neuroscience 21 7733ndash7741
Munoz D P Dorris M C Pare M amp Everling S (2000) Onyour mark get set Brainstem circuitry underlying saccadicinitiation Canadian Journal of Physiology andPharmacology 78 934ndash944
Munoz D P amp Everling S (2004) Look away The anti-saccade task and the voluntary control of eye movementNature Reviews Neuroscience 5 218ndash228
Nakahara K Hayashi T Konishi S amp Miyashita Y (2002)Functional MRI of macaque monkeys performing a cognitiveset-shifting task Science 295 1532ndash1536
Pierrot-Deseilligny C Muri R M Ploner C J Gaymard BDemeret S amp Rivaud-Pechoux S (2003) Decisional role ofthe dorsolateral prefrontal cortex in ocular motor behaviourBrain 126 1460ndash1473
Pierrot-Deseilligny C Rivaud S Gaymard B amp Agid Y(1991) Cortical control of reflexive visually-guided saccadesBrain 114 1473ndash1485
Schlag-Rey M Amador N Sanchez H amp Schlag J (1997)Antisaccade performance predicted by neuronal activity inthe supplementary eye field Nature 390 398ndash401
Selemon L D amp Goldman-Rakic P S (1988) Commoncortical and subcortical targets of the dorsolateral prefrontaland posterior parietal cortices in the rhesus monkeyEvidence for a distributed neural network subservingspatially guided behavior Journal of Neuroscience 84049ndash4068
Stoet G amp Snyder L H (2004) Single neurons in posteriorparietal cortex of monkeys encode cognitive set Neuron 421003ndash1012
Stroop J R (1935) Studies of interference in serial verbalreactions Journal of Experimental Psychology 18 643ndash662
Sweeney J A Mintun M A Kwee S Wiseman M BBrown D L Rosenberg D R amp Carl J R (1996) Positronemission tomography study of voluntary saccadic eyemovements and spatial working memory Journal ofNeurophysiology 75 454ndash468
Tomita H Ohbayashi M Nakahara K Hasegawa I ampMiyashita Y (1999) Top-down signal from prefrontal cortexin executive control of memory retrieval Nature 401699ndash703
Walker R Husain M Hodgson T L Harrison J ampKennard C (1998) Saccadic eye movement and workingmemory deficits following damage to human prefrontalcortex Neuropsychologia 36 1141ndash1159
Wallis J D Anderson K C amp Miller E K (2001) Singleneurons in prefrontal cortex encode abstract rules Nature411 953ndash956
White I M amp Wise S P (1999) Rule-dependent neuronalactivity in the prefrontal cortex Experimental BrainResearch 126 315ndash335
Zhang M amp Barash S (2000) Neuronal switching ofsensorimotor transformations for antisaccades Nature 408971ndash975
Zhang M amp Barash S (2003) Persistent LIP activity inmemory-antisaccades Working memory for a sensorimotortransformation Journal of Neurophysiology 911424ndash1441
1496 Journal of Cognitive Neuroscience Volume 17 Number 9




















