
Rostral prefrontal cortex: What neuroimaging can learn from human neuropsychology
49
Jn: B. Levrne and F. I. M. Crafk (Eds.) Mfnd and the Frontaf Lobes- Cognition, Behavior, and Brain Imaging (pp. 47-92). New York- Oxford University Press. 2012. Rostral Prefrontal Cortex What Neuroimaging Can Learn from Human Neuropsychology PAUL W . B U R G E S S , G I L G 0 N E N - Y A A C 0 V I, A N D EMMANUELLE VOLLE Rostral prefrontal cortex (PFC) approximates area 10 of the human brain and is variously referred to as "anterior PFC" or "frontopolar cortex." It is a very large brain region in humans, covering approximately 25 to 30 cubic centimeters, and it is alarge area in humans relative to other animals (Holloway, 2002; Semendeferi et al., 2001). In volumetric terms it is probably the largest single architectonic region of the frontal lobes (Christoff et al., 2001), which themselves represent approximately one tilird of the total cortical mass. Rostral PFC has many other features that suggest that it might play a highly important role in human cogni- tion, espedally higher-order capacities. For instance, the supragranular layers of area 10 in humans have more space available for connections with other higher- order association areas than in other animals (Semendeferi et al., 2001). It also follows a complex and protracted course of development, with suggestions that full maturity might not be achieved even as late as early adulthood (see Dumontheil et al., 2008, for review). This process of change may continue through- out life: specific volume reductions (relative to total brain volume) have been found in frontopolar cortex between early adulthood and middle age (John et al., 2009). Until approximately 15 years ago, virtually nothing was known about the func- tions of this region. There were two principal reasons for this situation. First, the large difference in rostral PFC structure and relative volumes between animals and humans cast doubt on the transferability of findings derived from the former to the latter. Thus, the study of this region was less attractive to scientists con- ducting animal lesion studies than other prefrontal regions. Second, lesions to this region in humans typically cause impairments that are not as easily demon- strable in the clinic as those that cause basic impairments in language, motor, 47
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Rostral prefrontal cortex: What neuroimaging can learn from human neuropsychology

Jn: B. Levrne and F. I. M. Crafk (Eds.) Mfnd and the Frontaf Lobes-Cognition, Behavior, and Brain Imaging (pp. 47-92). New York-Oxford University Press. 2012.
Rostral Prefrontal Cortex
What Neuroimaging Can Learn from
Human Neuropsychology
P A U L W . B U R G E S S , G I L G 0 N E N - Y A A C 0 V I, A N D E M M A N U E L L E V O L L E
Rostral prefrontal cortex (PFC) approximates area 10 of the human brain and is variously referred to as "anterior PFC" or "frontopolar cortex." I t is a very large brain region i n humans, covering approximately 25 to 30 cubic centimeters, and i t is alarge area i n humans relative to other animals (Holloway, 2002; Semendeferi et al., 2001). I n volumetric terms i t is probably the largest single architectonic region of the f ronta l lobes (Christoff et al. , 2001), which themselves represent approximately one t i l i r d of the t o ta l cortical mass. Rostral PFC has many other features that suggest that i t m ight play a highly impor tant role i n human cognit i o n , espedally higher-order capacities. For instance, the supragranular layers of area 10 i n humans have more space available for connections w i t h other higher-order association areas t h a n i n other animals (Semendeferi et al., 2001). I t also follows a complex and protracted course of development, w i t h suggestions that f u l l m a t u r i t y might n o t be achieved even as late as early adulthood (see Dumonthe i l et al., 2008, for review). This process of change may continue throughout l i fe: specific volume reductions (relative to t o t a l bra in volume) have been found i n frontopolar cortex between early adulthood and middle age (John et al., 2009).
U n t i l approximately 15 years ago, v i r tua l ly no th ing was k n o w n about the functions of this region. There were two principal reasons for this s ituation. First, the large difference i n rostral PFC structure and relative volumes between animals and humans cast doubt on the transferability of findings derived f r o m the former to the latter. Thus, the study of this region was less attractive to scientists conducting animal lesion studies t h a n other prefrontal regions. Second, lesions to this region i n humans typically cause impairments that are no t as easily demonstrable i n the clinic as those that cause basic impairments i n language, motor,
47

48 M I N D A N D T H E F R O N T A L L O B E S
or perceptual systems (Burgess et al., 2009). This meant that researchers have tended to focus on the functional characteristics of other f ronta l lobe regions (e.g., dorsolateral PFC), especially those where comparison w i t h animal studies might be more straightforward. Consequently, the relatively small, b u t nevertheless s tr ik ing , body of evidence f r o m human neuropsychology thatcouldhave been used as the basis for theorizing about rostral PFC lay largely unconsidered.
The Advent of Neuroimaging: From Famine to Feast? I n the 1990s, the relatively new technique of funct ional neuroimaging began to exert real influence w i t h i n the field. W i t h this method, the question of what rost r a l PFC "is for," i n terms of cognition, was t h r u s t to the forefront of cognitive neuroscience. This was because the evidence f r o m neuroimaging seemed to i n d i cate that structures w i t h i n rostral PFC support processing that may be used i n a very wide variety of situations (e.g., Grady, 1999). Indeed, incidental (i.e., unplanned) findings of activations i n rostral PFC were found dur ing the performance of just about any k i n d of cognitive task, ranging f r o m the simplest (e.g., conditioning paradigms; Blaxton et al. , 1996) to highly complex tests involving memory and judgment (e.g., Burgess et al., 2001 , 2003; F r i t h & F r i t h , 2003; Koechlin et al. , 1999) or problem-solving (e.g., Christoff et al. , 2001 ; for review, see Ramnani & Owen, 2004).
Perhaps unsurprisingly, given the plethora of data, a range of theories emerged (see Burgess et al, 2006, for review). For instance, rostral PFC was associated w i t h episodic memory functions (e.g., Rugg et al. , 1996; Tulving et al. , 1996), w i t h Buckner (1996, p. 156) suggesting that "the common activation dur ing episodic retrieval is highly localised, fal l ing at or near Brodmann area 10." I t was also associated w i t h what might be termed "metacognition," such as reflecting on one's own thoughts, or tiiinking i n a very controlled, conscious, or goal-directed mode (e.g., Johnson et al. , 2002). For instance, Christoff and Gabrieli (2000, p. 183) described the role of this region as of "evaluation, moni tor ing , or manipulat ion of internal ly generated information," and "theory of m i n d " researchers suggested a role for medial rostral PFC "when we attend to our own mental states as wel l as the mental states of others" (Fr i th & F r i t h , 2003, p. 467). Further, there was a set of theories that Burgess and colleagues (2006) t e r m the "Sum Process" accounts. These, broadly, mainta in that rostral PFC supports processing involved i n the coordination of otherwise independent processing resources (e.g., Dreher, Koechlin, AH, & Grafman, 2002; Koechlin et al., 1999; Koechlin, Ody, & Kouneiher, 2003; Ramnani & Owen, 2004). Lending broad support to this type of account was the identif ication of a consistent association between BOLD signal changes i n rostral PFC and the performance of prospective memory tasks (e.g., Burgess eta l . , 2001, 2003), where performance of an ongoing task has to be dovetailed w i t h the detection of, and response to , a prospective memory cue.

Rostral Prefrontal Cortex 49
The prohferation of these theories was perhaps an improvement upon the complete dearth beforehand. But because of the inherent nature of neuroimaging data, these accounts have/ i n the main , been allowed to al l stand side by side. Indeed, they are continuing to proliferate, w i t h l i t t l e or no revision or dismissal of any existing accounts. I t is possible that neuroimaging data ( from one study alone, at least) cannot provide sufficient evidence to effect the dismissal of a cognitive model. There are a range of views on this matter, and a f u l l debate is beyond the scope of this chapter (for review see, e.g., Coltheart, 2006; Henson et al. , 2006). However, whether or no t i t is possible i n theory, i t is no t currently happening i n practice, except perhaps by the process of "natural selection" to which Henson (2006) refers (where, presumably, the randomness of science politics and chance forces sit alongside logical argument and data as influences). A potential serious contributor to this s i tuation is that , as Chambers and Matt ing ly (2005) have noted, neuroimaging analyses may detect activation i n regions of the bra in that are no t actually involved i n performance of the experimental paradigm under investigation. This is especially significant for the study of rostral PFC: there is good evidence for hemodynamic changes i n this region dur ing "mind-wandering" and other task-irrelevant processing (see Gilbert et al., 2007, for review).
On these grounds, theoriz ing about the cognitive functions supported by rost r a l PFC based on neuroimaging data w i l l be particularly convincing where i t is constrained by findings f r o m lesion studies. I n t ime, transcranial magnetic s t imulat ion (and related methods) may also provide useful data i n this respect (although there are technical issues to be overcome, especially w i t h respect to rostral PFC structures deeper w i t i i i n the brain [e.g., caudal medial rostral PFC]). However, data f r o m human lesion studies are b o t h directly relevant and already available. This chapter aims to outline the key aspects of these data and to show the ways i n which they can be potential ly of great use to the neuroimaging community.
Rostral PFC and "Intelligence" A key example of the way i n which data f r o m human lesion studies might constrain theoriz ing that has arisen out of neuroimaging studies is the issue of the role of rostral PFC i n "intelligence."
I t could be argued that over the past 50 years the major i ty of theorists w i t h i n the field of the psychology of individual differences have moved towards the view that there maybe no single construct that best represents "intelligence." Instead, they have concentrated more on what Spearman would have considered second-order or specific factors—those that Thomson (1939) pointed out may (on m a t h ematical grounds) be sufficient to explain "intelligence." O n this view, seeking stable patterns of activation i n the bra in that relate to a single "intelligence" construct might be fruitless. Congruent w i t h this view, and predating most, is the application of the most commonly used intelligence test i n neuropsychology,

50 MIND AND THE FRONTAL LOBES
the Wechsler intelligence scales (and variants; Wechsler, 1939). On this concept i o n of intelligence, the construct can encompass a wide range of abilities and aptitudes, and an I Q score is a simple average of a sample of them, most commonly used as a f o r m of baseline against which other test scores might be compared. The scores on any two subtests themselves, or between the larger groupings (e.g., Verbal I Q vs. "Performance," mainly non-verbal IQ) , can show considerable variat ion i n any one individual , even w i t h i n the healthy population (see, e.g., Spreen & Strauss, 1988, p. 121). The alternative common conception of I Q i n cognit ive neuroscience is i n the sense of "g," as typif ied, for example, i n the works of John Duncan (e.g., Duncan et al. , 2000).
Using the construct "intelligence" perhaps more i n the former way t h a n the latter, Jung, Haier, and colleagues claim that "central psychological constructs [such as general intelligence (g) and work ing memory capacity] . . . share common neural systems . . . a common anatomic framework for these constructs implicates Brodmann area 10" (Colom, Jung, & Haier, 2007; see also Jung & Haier, 2007). More specifically, the hypothesis is that the contr ibut ion to cognition made by processes supported by rostral PFC is "to hypothesis test various solutions to a given problem" (Jung & Haier, 2007, p. 138). A l though the specifics of this claim are no t outl ined i n great depth i n information-processing terms, i t seems l ikely that a decrement i n such an abil i ty might affect performance on a very wide range of tasks. Thus, lesions to this region should cause marked decrements on a wide range of tasks, including I Q tests.
This prediction unites this latter conception of intelligence w i t h the "g" view. The no t i on of "g" originally came f r o m Charles Spearman's examination of the performance of many individuals across lots of cognitive tests, using correlation matrices. He noticed an effect he called "positive manifo ld" (Spearman, 1904) i n these matrices. This is the tendency for any correlation m a t r i x of this k i n d to have more positive correlations t h a n negative ones. I n other words, there is a tendency, across lots of people, and across lots of tests, for those who are better at one test to also be better at another. Spearman's interpretat ion was t h a t this positive manifo ld was evidence of a hypothetical abi l i ty t h a t is required to per form many types of test, which he called "g" (short for "general intelligence"). Since that t ime , various psychometric tests have been designed to measure this construct, often now referred to as " f lu id intelligence" (following Cattell, 1963).
The key idea here is that there is a particular cognitive resource, or process, t h a t underpins performance on a wide range of tasks. This distinguishes the no t i on of fluid intelligence f r o m the one outl ined above, where I Q i s just an average of a range of different abilities. As Roca and colleagues (2010) note, funct ional imaging studies of tasks that supposedly measure fluid intelligence t end to find extensive activity on the lateral surface of the f ronta l cortex, especially around the inferior f ronta l sulcus and anterior insula / frontal operculum. Additionally, fluid intelligence tasks also regularly activate the dorsal anterior cingulate/pre-supplementary motor area and the intraparietal sulcus. Accordingly, Duncan and

Rostral Prefrontal Cortex 51
colleagues have proposed t h a t these activations reflect the neural operation of a cognitive system that supports fluid intelligence (e.g., Duncan, 2001 , 2005; Duncan et al., 2000; Duncan & Owen, 2000).
One way i n which this latter conception of intelligence is similar to the previous one considered here is i n the prediction i t makes for human lesion studies. I f rostral PFC supports some processing that corresponds i n some way to fluid intelligence (or Spearman's "g"), then this hypothesis also predicts that lesions to this region should cause decrements across a wide range of tasks. The extent of the decrement on any one task might reflect the degree to which that taskloads upon "g." However, unless the hypothesis applied to rostral PFC makes the prediction that lesions to this region should cause a wide range of decrement across many tasks (and more so than at least some other brain regions), then i t is essentially unfalsifiable.
Rostral PFC Lesions Need Not Cause Substantial Impairment on Traditional Tests of Intelligence The problem for many of the functional neuroimaging-based accounts of rostral PFC funct ion is that lesions to rostral PFC do no t typically cause substantial decrements on IQ. tests. Nor do they cause impairment on a wide range of other tasks, while simultaneously causing impairment on a quite restricted range of others. Moreover, this has been k n o w n for many years.
Probably the first group human lesion study of rostral PFC functions was carr ied out by Petrie (1952), although this was no t its principal a im. Petrie compared the effects of two different leucotomy procedures on cognition i n participants w i t h "neurotic" symptoms. The standard procedure at that t ime made an incision at a rostral-caudal plane that , i f extended to the outer surface of the brain, wou ld have bisected Brodmann areas (BAs) 45 ,47 , 46, 9, and 8. This was compared w i t h the effects of a rostral incision, where the cut was made more anteriorly, fal l ing just behind area 10.
The rostral operation led to improvements i n a range of tasks, notably manual dexterity and concentration, fewer perseverations, and better performance on WAIS (Wechsler Adu l t Intelligence Scale) D ig i t Symbol and Similarities subtests. There was no change, relative to preoperative levels, on many further tests (Porteus Mazes, WAIS Verbal I Q , Performance I Q , Ful l Scale I Q (FSIQ), Cattell Fluency, Proverb Interpretation) . However, there were decrements on just two tests: there was a varied response to distraction, and the sense that t ime passes more quickly (unfilled estimation of the passage of 60 seconds). I n this study, Petrie (1952) provided compelling evidence that lesions affecting rostral PFC lesions do not cause significant I Q decrement, although they may cause impairments i n some mental capacities that , to this day, are no t particularly wel l understood.
Since Petrie's study, a substantial body of fur ther evidence f r o m human lesion studies has accumulated, also demonstrating that patients w i t h rostral PFC

52 M I N D A N D T H E F R O N T A L L O B E S
damage are not typically impaired on tradi t ional tests of intelligence. Many single cases reported have scored i n the superior range on such tests, w i t h l i t t l e or no discernable change f r o m estimates of premorbid funct ioning, despite extensive rostral PFC involvement (e.g., Goel & Grafman, 2000; Goldstein et al. , 1993; j Shallice & Burgess, 1991; see Burgess, 2000, for review). Indeed, Uretzky and i Gilboa's (2010) case, Z.P. (WAIS-III Verbal I Q 121 , Performance I Q 104, FSIQ j 113), who has circumscribed r i ght front-polar atrophy fol lowing a traffic accident, ' gained degrees i n civil engineering and business administrat ion after his injury, which hardly suggests a marked I Q deficit.
Of course one might object that there may be something unusual about these single cases, or t h a t the estimates of premorbid funct ioning might be inaccurate. | However, group studies have also shown no difference i n IQs between patients w i t h rostral PFC lesions and those elsewhere. For instance, i n Dreher and coworkers' (2008) study, patients w i t h rostral PFC lesions (n = 7; called frontopolar lesions by Dreher and coworkers) had a mean number of years of education of 14.71 (SD 3.4), and the patients whose lesions affected the f ronta l lobes b u t no t the rostral aspects of i t (n = 5) had a very similar number (mean 14.2, SD 2.5). Likewise, there was no significant difference i n the performance of the two groups i n terms of W A I S - I I I Full-Scale IQscores (rostrals' mean = 110.43, SD = 20.4; n o n -rostrals = 103.20, SD = 8.35).
Another way to look at this issue would be to start w i t h the intelligence test rather t h a n the lesion: i n other words, to gather data on many patients w i t h v a r i ous lesions and to ask which lesion lo cation shows the strongest relationship w i t h I Q decrement. Warr ington , James, and Maciejewski (1986) provided by far the largest study of this k i n d to our knowledge, examining 656 patients w i t h un i la t eral cerebral lesions on a shortened version of the WAIS. They found that Verbal IQwas impaired fol lowing le f t hemisphere lesions, regardless of where the lesion was located w i t h i n the hemisphere. Performance IQwas impaired only fol lowing lesions affecting the r i ght parietal lobe. I n an analysis by indiv idual subtest, the only finding was a relationship between Block Design and Picture Arrangement decrement and r ight parietal involvement. More t h a n 20 years later, Glascher and associates (2009) carried out a similar k i n d of study, using the more sophisticated lesion imaging methods now available, and exarnining 2 4 1 patients w i t h focal bra in damage on the W A I S - I I I . They found a statistically significant lesion-deficit relationship i n le f t inferior f r onta l cortex for verbal comprehension scores, w i t h an association between lesions i n le f t lateral f r onta l and parietal cortex for the work ing memory index, and i n r i ght parietal cortex for perceptual organization. They found no single localization for processing speed.
From these studies we can conclude t h a t there is l i t t l e evidence for a preferent i a l l i n k between rostral PFC lesions and WAIS I Q decrements. But is this some artifact of the particular def init ion of intelligence encapsulated by the Wechsler Intelligence scales, or the way they measure them? Apparently not . Roca and colleagues (2010) studied the relationship between measures of f lu id intelligence

Rostral Prefrontal Cortex 53
(e.g., Cartel's Culture Fair Test) and performance on a range of measures of executive functions. I n the i r f irst experiment, the sample comprised 44 patients w i t h chronic lesions affecting a variety of subregions w i t h i n the f ronta l lobes (note: n o t just affecting rostral PFC). For two of the most commonly used executive funct ion tasks (Wisconsin Card Sorting Test [Nelson version] and Verbal Fluency), they found that although the patients were significantly poorer t h a n a group of matched healthy controls i n terms of raw performance on all three tests, once the variance attributable to f lu id inteUigence was accounted for by covariance, this difference disappeared. I n other words, for some executive tasks there seems to be an overlap i n the processing demands w i t h f lu id intelligence measures. But there was no apparent relation between where within the f ronta l lobes the lesion was located and deficits on the fluid intelligence measure (the Culture Fair Test). I n other words, Roca and associates d id no t find a stronger relationship between fluid intelligence deficits and lesions that affected rostral PFC t h a n for other parts of the f ronta l lobes.
However, i n a second experiment, they did make a discovery that informs us of the nature of the mental abilities that rostral PFC might support. Here, they administered a different, and wider, range of executive and social funct ion tasks to 2 1 patients. Most of these were performed more poorly by the patient group t h a n controls. This t ime, however, the investigators found that for some of the executive tasks, adjusting for fluid intelligence did not remove the group difference. These tests were go/no go (Dubois et al., 2000; Luria, 1966); proverb in ter pretat ion (Hodges, 1994); Hayling Sentence Completion Task (Burgess & Shallice, 1996, 1997); Hote l Task (Manly et al., 2002; a version of Shallice & Burgess's [1991] Six Element Task); and the Faux Pas test (Stone et al. , 1998). Roca and coworkers (2010) then devised a score that represented the mean residual f r o m these tests (i.e., a score that represented the degree of impairment on the tests that could no t be explained by fluid intelligence). When they examined the lesion overlaps for the six patients who showed the greatest negative value, they found that the location of the overlap was i n rostral PFC, especially i n the r i ght hemi sphere. I n other words, the rostral PFC lesions caused an impairment that was weE measured by a number of executive tasks, b u t that could no t be explained by changes i n f lu id intelligence. This study, together w i t h the evidence above, therefore provides good evidence t h a t rostral PFC lesions do no t typically affect performance substantially on tests of intelligence any more t h a n lesions elsewhere w i t h i n the brain . This finding holds whether I Q i s conceived of as an average across different domains, or as some homogeneous process, or set of them, operating across all cognitive domains.
A recent study, however, seems prima facie to r u n against the findings by Roca and coworkers. Glascher and associates (2010) reported a Schmid-Leiman factor transformation analysis of WAIS subtests i n their group of 241 patients w i t h brain damage of various etiologies (principally stroke). This analysis specified a higher-order factor common to three lower-order constructs (Verbal, Spatial,

54 MIND AND THE FRONTAL LOBES
Working Memory) . The investigators found that the factor scores for this higher-order construct related to the presence of lesions i n the left f ronta l pole, and concluded that this region plays some par t i n supporting "g." This is an interesting finding. However, there maybe many otherpossible interpretations of these data (as indeed, Thomson [1939] po inted out that there were for the positive mani fo ld i n correlation matrices that Spearman took as pr imary evidence of "g"). Glascher and associates did not use control or comparison tasks, so this l imits an interpretat ion i n information-processing terms. Linked to this, there may be influences that do not act equally as regards the possibility of general decrement i n rostral PFC patients: damage to this region tends to be neurologically "silent," so i t Is often no t detected u n t i l the lesions are large or have affected particular pathways or other PFC or subcortical regions, causing mult iple deficits. Nevertheless, although these l imitat ions make interpretat ion cUfficult, they do raise the mtr igu ing possib i l i t y of some f o r m of higher-order construct supported by a rostral PFC region.
The Challenge that Rostral PFC Presents to Neuroimaging I n l i ght of all the data above, especially findings f r o m single cases, the unequivocal assertion that there is a special role for rostral PFC i n "intelligence" (as t r a d i t i o n ally defined) seems plausible only i f the theorizing t h a t led to that conclusion had considered data f r o m neuroimaging alone. I f this is what has occurred, i t may t u r n out to be a useful example to historians of science. I t is already widely accepted i n the neuroimaging community that , as Poldrack and Wagner (2004) p u t i t : "Neuroimaging can demonstrate that bra in activity is correlated w i t h a cognitive task or process, b u t cannot demonstrate that the region is necessary for that task or process: This requires showing that disruption of the region results i n impairment of the funct ion" (p. 180). I t would seem that the issue of intelligence and rostral PFC is l ikely to be a test of the degree to which "involved i n , but n o t necessary for" w i l l prove useful i n characterizing brain-behavior relat ions—or at least, of the difficulty i n devising contrast conditions or other experimental manipulations t h a t control for al l possible non-relevant cognitive processing (see, e.g., Friston et al. , 1996)
Another problem for neuroimaging inferences i n reference to theoriz ing about rostral PFC funct ion concerns the specificity of findings; i n other words, the degree to which neural activation i n a particular region is specific to a particular task. For some more " informational ly encapsulated," routine-type processing, such as basic visual processing, there maybe such a dose correspondence between a particular local pat tern of activation and a particular stimulus (or thought) t h a t one is able w i t h some accuracy to predict the latter f r o m the former, at least w i t h i n a highly constrained experimental s i tuation. However, this seems less l ikely for

Rostral Prefrontal Cortex 55
more "central processes" and perhaps most problematic of all for systems supported by f ronta l lobe structures, which deal w i t h novelty (Burgess, 1997), and may have "adaptive coding"-type characteristics (Duncan, 2001). Where a consistent relationship is found between performance of task type A and activation i n region X, i t may be relatively easy to base one's theorizing about the functions of that region around information-processing models relating to task type A . However, i f region X is also active i n a wide range of tasks other t h a n A, this may prove unproductive.
I t may be that this k i n d of s i tuation has already occurred i n relation to rostral PFC theorizing. One of the earliest observations i n relation to rostral PFC activations f r o m neuroimaging was the large number of studies of episodic memory i n which activations i n B A 1 0 were found (e.g., Buckner et al., 1996). Grady (1999) provided an excellent review of the relevant findings available at the end of the 1990s. She reviewed 90 PET studies demonstrating regional cerebral blood flow (rCBF) changes w i t h i n the f ronta l lobes, and concluded that the most heavily represented funct ion of BA 10 is episodic memory. The basis for this conclusion was that most of the experiments report ing BA 10 activation were using episodic memory paradigms. This was an understandable conclusion, b u t may have been misleading since i t d id no t take into account the predominance of episodic memory investigations i n the Hterature at that time. I f one adjusts for the different pro portions of studies across different cognitive domains, a different picture emerges. Thus, 37/90 (41%) of the studies that Grady considered i n her review investigated episodic memory, and 47/90 (52%) of the studies she considered reported rCBF changes i n BA 10. However, only 68% of the episodic memory studies reported BA 10 activations, and only 25 (53%) of the paradigms that caused BA 10 activations were episodic memory ones. Furthermore, 7/90 of the studies that Grady considered were investigations of "working memory," and 6 of these (86%) showed BA 10 activation. Finally, 6/90 studies investigated conditioning or motor learning , and al l 6 (100%) reported BA 10 activation. Thus, on the basis of these data i t is doubtful that , whatever role BA 10 functions play i n cognition, they are any more active when people are involved i n episodic memory tasks t h a n when they are engaged i n other sorts of tasks. As MacLeod, Buckner, Miez in , Petersen, and Raichle (1998) p u t i t , "although . . . BA 10 is routinely activated by episodic memory tasks, i t is no t uniquely activated by episodic memory tasks" (p. 41).
As w i t h the "intelligence hypothesis" of rostral PFC function, caution might have been indicated by the human lesion evidence. The data f rom both single-case studies and group studies (see, e.g., Burgess etal . , 2000; Petrie, 1952) show that uncomplicated lesions to rostral PFC do not cause pervasive problems w i t h episodic recall and recognition. Indeed, many patients w i t h even extensive rostral PFC lesions may be completely normal on a wide range of clinical neuropsychological tests of episodic memory (e.g., Case A.P. f r om Shallice & Burgess, 1991). This is not , of course, t o say that rostral PFC is not involved i n some aspects of memory functioning—• i t probably is, as we shall see below—-the po int instead is that constraining the

56 MIND AND THE FRONTAL LOBES
theorizing based on neuroimaging data by requiring t i e accounts to be i n agreement simultaneously w i t h the human lesion evidence might have greatly accelerated the advance of our understanding. The question then might have been something like "what k i n d of processing might regularly be prompted by episodic memory tests (and other tests) b u t which, when damaged, would no t cause a defic i t on, e.g., a forced-choice recognition memory test?" Notably, those few authors who d id pursue a line of reasoning of this type alighted upon accounts that now seem remarkably prescient (e.g., Wheeler, Stuss, & Tulving, 1997).
Other Ways in which Lesion Evidence Is Critical for Theorizing I n this way (and others), one of the great uti l i t ies of lesion evidence is t h a t a negative finding can be as significant for theorizing as a positive one. This is, of course, demonstrated by perhaps the most significant pat tern of behavior as regards the orizing i n cognitive neuropsychology: the double dissociation. This is where, i n its classic example, one patient shows a marked decrement i n one domain of cognit i o n but unimpaired performance i n another. Then another patient shows the opposite pattern . Here, the decrements i n each case are noteworthy only because they are accompanied by a normal-level contrast.
I n this respect, one might ask for the broadest evidence we have of the patterns bo th of impairment and preservation fol lowing rostral PFC lesions. The wonderf u l set of group human lesion studies carried out over many years by Donald T. Stuss and Michael Alexander and their colleagues i n Toronto provide probably the best such database for the purpose, since similar methods were applied to the analysis of the data over a long period of t ime. Moreover, some of the cases were shared across studies, which for this purpose is useful.
W h a t we are t r y i n g to address i n this analysis is whether lesions affecting rost r a l PFC cause a breadth of deficits comparable w i t h lesions elsewhere w i t h i n the f ronta l cortex. I n this way, the a im is to answer the question of whether (a) theor iz ing should start by considering that rostral PFC supports processing that operates across a very wide range of tasks, or (b) whether one might instead consider performance on a more specific subset of t h e m as particularly good indicators of a "rostral PFC-supported" cognitive substrate. One might imagine that this is perhaps no t much more t h a n a formalized version of what occurred i n the early days of research into the functions of the pr imary visual cortex, for example, where a particular relation between deficit on a subset of tasks (i.e., those involving visual processing) and lesions to this region was noted. Fortunately, however, the procedures of Stuss and Alexander take into account what is already k n o w n ab out f r o n t a l lobe funct ion, i n that they do no t consider as experimental tasks sets of t h e m that have been shown no t to be relevant, and indeed, they go to some lengths to exclude patients w i t h compHcating presentations.

Rostral Prefrontal Cortex 57
Details about the precise populat ion contr ibut ing to these analyses are of course available i n the original research papers. Broadly, however, Stuss and Alexander followed the fol lowing procedures.
They tested patients whose lesions involved the f ronta l lobes (and sometimes outside the f ronta l lobes) and collected sets of non-patient control subjects matched as closely as possible to the patients for sex, age, and education. The lesions included infarction, hemorrhage (including ruptured aneurysms), trauma, and tumors. Most patients w i t h tumors had resection of meningiomas or l ow-grade gliomas and had no t been treated w i t h radiation. Stuss and Alexander avoided patients i n the very acute phase of their illness (e.g., less t h a n 2 months after onset), and also excluded people w i t h significant aphasia, visual neglect, motor control, or any other significant neurological or psychiatric disorders. Further, the patients tended to have IQ. scores w i t h i n the normal range. Incidentally, i t should be mentioned t h a t i n these studies, and comparable ones where rigorous neuropsychological screening is used to remove potential confounds (e.g., Burgess et al., 2000), no simple l i n k between etiology and cognitive results is found: i t is the location of the lesion that is the principal determining factor.
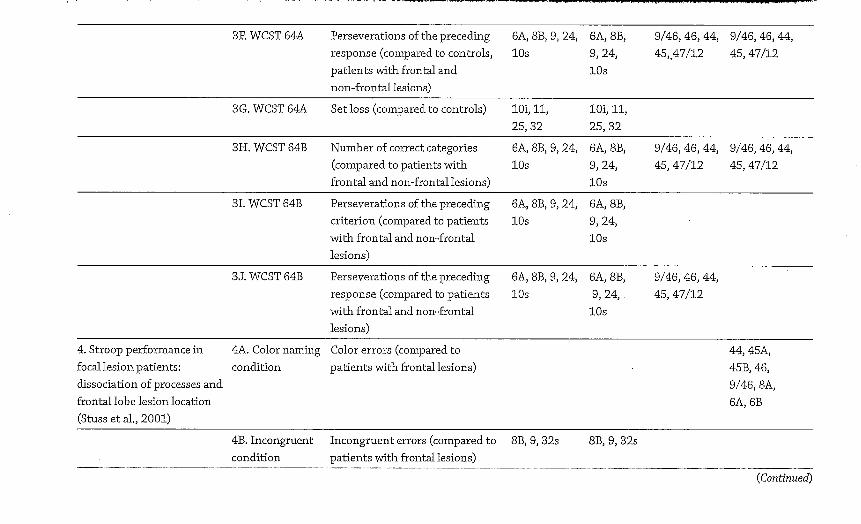
The results shown i n Tables 4 .1 and 4.2 and summarized i n Figure 4.1 demonstrate the number of variables, out of the t o t a l of 44 under consideration (see Table 4.1 for a l i s t of them), where impairments were documented by the Stuss/ Alexander group for each broad architectonic subdivision of the f ronta l lobes. N o t al l the group lesion studies conducted by Stuss and Alexander over this period are included because of technical differences i n their analysis method; the figure shows only those where comparability is sufficient for this purpose.
This simple meta-analysis of the Stuss/Alexander series of studies shows two results quite clearly. The first of these is that rostral PFC lesions do not cause deficits on a more widespread set of tasks t h a n lesions elsewhere w i t h i n the f ronta l lobes. For b o t h le f t and r i g h t hemispheres, lateral and medial surfaces, deficits were noted on the range of variables examined by Stuss and Alexander (see Table 4.2 for a list) less frequently t h a n for some other f ronta l lobe regions. A l though r ight medial aspects of superior area 10 were implicated (alongside others) i n groups of patients who performedpoorly on some variables, for the other regions, w i t h the possible exception of some aspects of ventral PFC, cognitive deficits associated w i t h area 10 lesions, regardless of hemisphere or surface, were consistent ly amongst the Zeast frequent findings i n this series of studies stretching over 10 years. Indeed, even considering the most frequently implicated subregion of area 10 (r ight medial superior), the equivalent region w i t i i n area 9 (i.e., r i g h t superior medial) was implicated more t h a n twice as frequently. This evidence would seem to suggest that i f "intelligence" is a construct that determines performance across a range of tasks, i t wou ld be most appropriate on this evidence to start first by considering area 9, n o t area 10 as the most l ikely supporting brain region.

58 M I N D A N D T H E F R O N T A L L O B E S
Figure 4.1. A visual i l l u s t r a t i o n o f the f indings f r o m a selection o f Stuss and colleagues' studies i n v o l v i n g pat ients w i t h f r o n t a l lobe lesions, between 1998 and 2008. Test results are compared to pat ients w i t h n o n - f r o n t a l lesions and /o r controls . The figure shows the frequency of a reported i m p a i r m e n t for each B r o d m a n n area based on the results f r o m 44 behavioral variables described i n Table 4 .1 . These frequencies are d iv ided i n t o six ranges, represented by di f ferent colors. Dark blue = the region was f o u n d to be impaired , to a statist ical ly s ignif icant degree, on 1 o f the 44 behavioral variables (relative to either controls or at least one other pat i ent group) . L ight blue = 2 /44 variables impaired . Green = 3 /44 variables impaired . Yellow = 4 to 1 1 variables impaired . Orange = 12 to 19 variables impaired . Red = 20 or more variables impaired . The results show t h a t rostra l PFC regions (area 10) are not impa i red on a larger n u m b e r of these behavioral indicators t h a n other PFC regions. These results provide a constra int for theor i z ing about the funct ions o f rostra l PFC. (See Color Plate Section for a color vers ion of this figure.)
O f course , t h e se l e c t i on o f tasks t h a t Stuss, A l e x a n d e r , a n d col leagues used i n t h i s series o f s tud ies was n o t r a n d o m . They w e r e chosen large ly because o f t h e i r p u t a t i v e s e n s i t i v i t y t o f r o n t a l lobe l es ions , o r w e r e d i s covered t o be so d u r i n g t h e course o f t h e e x p e r i m e n t . H o w e v e r , t h i s adds i n e x t r e m e l y u s e f u l ways t o t h e resu l t s a l ready o u t l i n e d above. We can i g n o r e a n y s u g g e s t i o n t h a t r o s t r a l PFC les ions u s u a l l y cause ser ious de f i c i t s i n p r i m a r y s ensory f u n c t i o n s , or i n t h e " r o u -t i n i z e d " l e a r n e d sk i l l s such as basic aspects o f language process ing , m o t o r or v i s u o -p e r c e p t u a l a b i l i t i e s , r e a d i n g , w r i t i n g , or s i m p l e aspects o f m e m o r y (e.g., k n o w l e d g e ;

Table 4.1. Summary of the Findings from Stuss and Colleagues Testing Patients with Lesions i n Frontal Lobes, 1998-2008
Study Conditio Variables Right Medial Left Medial Right Lateral Left Lateral
1. The effect of focal anterior 1A. Letter-based and posterior brain lesions fluency task on verbal fluency (Stuss et al., 1998)
Making errors (compared to controls)
9/46, 46, 44, 45, 47/12
I B . Letter-based fluency task
Produced fewer (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24
9/46, 46, 44, 45, 47/12
1C. Semantic fluency task
Produced fewer words (compared to controls)
9/46, 46, 44, 45, 47/12, 5, 7, 40
2. Humor appreciation: a role 2A. Verbal Humor Impairment in distinguishing of the right frontal lobe Statements test between humorous and neutral (Shammi & Stuss, 1999) verbal stimuli (compared to
controls)
8B, 9,10s 8B, 9,10s
2B. Verbal Humor Impairment in choosing correct, Statements test funny punchlines to incomplete
verbal joke items (compared to controls).
8B, 9,10s 8, 9,10s
(Continued)
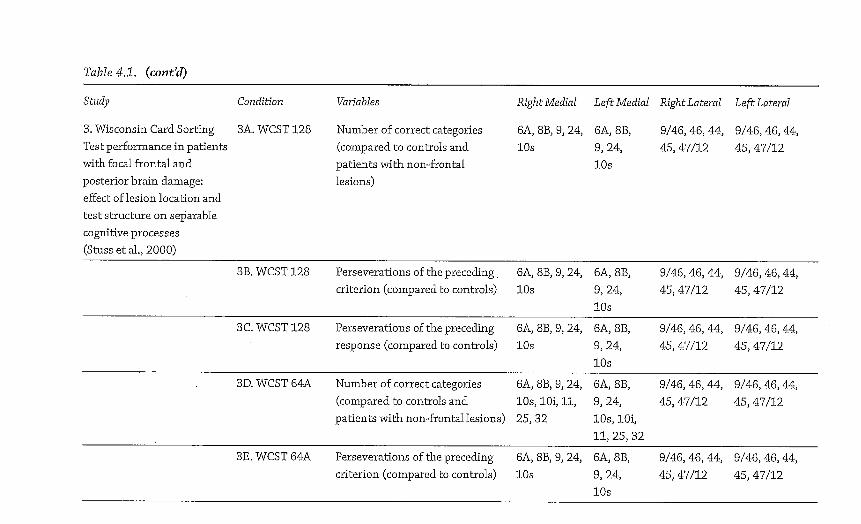
Table 4.1. (cont'd)
Study
3. Wisconsin Card Sorting Test performance i n patients with focal frontal and posterior brain damage: effect of lesion location and test structure on separable cognitive processes (Stuss et al., 2000)
Condition
3A. WCST128
Variables
Number of correct categories (compared to controls and patients w i t h non-frontal lesions)
Right Medial Left Medial Right Lateral Left Lateral
6A, 8B, 9, 24, 10s
6A, 8B, 9, 24, 10s
9/46, 46, 44, 9/46, 46, 44, 45, 47/12 45, 47/12
3B. WCST 128 Perseverations of the preceding 6A, 8B, 9, 24, 6A, 8B, 9/46, 46, 44, 9/46, 46, 44, criterion (compared to controls) 10s 9, 24, 45, 47/12 45, 47/12
10s
3C. WCST 128 Perseverations of the preceding 6A, 8B, 9, 24, 6A, 8B, 9/46, 46, 44, 9/46, 46, 44, response (compared to controls) 10s 9, 24, 45, 47/12 45, 47/12
10s
3D. WCST 64A Number of correct categories 6A, 8B, 9, 24, 6A, 8B, 9/46, 46, 44, 9/46, 46, 44, (compared to controls and 10s, lOi, 11, 9, 24, 45, 47/12 45, 47/12 patients w i t h non-frontal lesions) 25, 32 10s, lOi ,
11 , 25, 32
3E. WCST 64A Perseverations of the preceding 6A, 8B, 9, 24, 6A, 8B, 9/46,46, 44, 9/46, 46, 44, criterion (compared to controls) 10s 9, 24, 45, 47/12 45, 47/12
10s

3R WCST 64A Perseverations of the preceding response (compared to controls, patients wi th frontal and non-frontal lesions)
6A, 81 10s
5, 9, 24, 6A, 8B, 9, 24, 10s
9/46, 46, 44, 45,47/12
9/46, 46, 44, 45, 47/12
3G. WCST 64A Set loss (compared to controls) lOi , 11, 25, 32
lOi , 11, 25, 32
3H. WCST 64B Number of correct categories (compared to patients w i t h frontal and non-frontal lesions)
6A, 81 10s
5, 9, 24, 6A, 8B, 9, 24, 10s
9/46, 46, 44, 45, 47/12
9/46, 46, 44, 45, 47/12
31. WCST 64B Perseverations of the preceding criterion (compared to patients wi th frontal and non-frontal lesions)
6A, 81 10s
5, 9, 24, 6A, 8B, 9, 24, 10s
3 J. WCST 64B Perseverations of the preceding response (compared to patients w i t h frontal and non-frontal lesions)
6A, 81 10s
5, 9, 24, 6A, 8B, 9, 24, .
10s
9/46, 46, 44, 45, 47/12
4. Stroop performance in focal lesion patients: dissociation of processes and frontal lobe lesion location (Stuss et al., 2001)
4A. Color naming condition
Color errors (compared to patients wi th frontal lesions)
44, 45A, 45B, 46, 9/46, 8A, 6A, 6B
4B. Incongruent Incongruent errors (compared to 8B, 9, 32s 8B, 9, 32s condition patients wi th frontal lesions)
(Continued)

Table 4.1. (cont'd)
Study Condition Variables Right Medial Left Medial Right Lateral Left Lateral
5. Dissociation with in the 5A. Simple task Slowing down in reaction time 6A, 8B, 9, 24 6A, 8B, anterior attentional system: (compared to controls) 9, 24 effects of task complexity and irrelevant information on reaction time and accuracy (Stuss et al., 2002)
5B. Easy task Slowing down from simple to easy task (compared to controls)
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5C. Easy task Number of omissions (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24
44, 45A, 45B, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5D. Complex task Less proportional increase (compared to controls)
6A, 8B, 9, 24, 10,11, 25, 32
6A, 8B, 9, 24 ,10 ,11 , 25, 32
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5E. Complex task Number of omissions (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24
3F. WCST 64A Perseverations of the preceding response (compared to controls, patients wi th frontal and non-frontal lesions)
6A, 8B, 9, 24, 10s
6A, 8B, 9, 24, 10s
9/46, 46, 44, 45,47/12
9/46, 46, 44, 45, 47/12
3G. WCST 64A Set loss (compared to controls) lOi, 11, 25, 32
lOi , 11, 25, 32
3H. WCST 64B Number of correct categories (compared to patients wi th frontal and non-frontal lesions)
6A, 8B, 9, 24, 10s
6A, 8B, 9, 24, 10s
9/46, 46, 44, 45, 47/12
9/46, 46, 44, 45, 47/12
31. WCST 64B Perseverations of the preceding criterion (compared to patients wi th frontal and non-frontal lesions)
6A, 8B, 9, 24, 10s
6A, 8B, 9, 24, 10s
3 J. WCST 64B Perseverations of the preceding response (compared to patients wi th frontal and non-frontal lesions)
6A, 8B, 9, 24, 10s
6A, 8B, 9, 24, .
10s
9/46, 46, 44, 45, 47/12
4. Stroop performance in focal lesion patients: dissociation of processes and frontal lobe lesion location (Stuss et al., 2001)
4A. Color naming condition
Color errors (compared to patients wi th frontal lesions)
44, 45A, 45B, 46, 9/46, 8A, 6A, 6B
4B. Incongruent Incongruent errors (compared to 8B, 9, 32s 8B, 9, 32s condition patients w i t h frontal lesions)
(Continued)

Table 4.1. (cont'd)
Study Condition Variables Right Medial Left Medial Right Lateral Left Lateral
5. Dissociation wi th in the 5A. Simple task Slowing down in reaction time 6A, 8B, 9, 24 6A, 8B, anterior attentional system: (compared to controls) 9, 24 effects of task complexity and irrelevant information on reaction time and accuracy (Stuss et al., 2002)
5B. Easy task Slowing down from simple to easy task (compared to controls)
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5C. Easy task Number of omissions (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24
44, 45A, 45B, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5D. Complex task Less proportional increase (compared to controls)
6A, 8B, 9, 24, 10 ,11 , 25, 32
6A, 8B, 9, 24 ,10 ,11 , 25, 32
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5E. Complex task Number of omissions (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24

5R Complex task Number of false-positive responses (compared to controls)
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5G. Redundant Less proportional increase task (compared to controls)
6A, 8B, 9, 24 6A, 8B, 9, 24
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
5H. Redundant Number of omissions (compared 6A, 8B, 9, 24 6A, 8B, task to controls) 9,24
44, 45A, 45, 47/12,11, 10, 46, 9/46, 9, 8B, 8A
51. Second Number of omissions (compared 6A, 8B, 9, 24 6A, 8B, simple task to controls) 9,24
6. Impaired concentration 6A. Slowing in due to frontal lobe damage reaction time from two distinct lesion sites (compared to (Alexander et al., 2005) controls and
patients wi th frontal lesions)
9, 24, 32s 9/46
(Continued)

Table 4,1. (cont'd)
Study Condition Variables Right Medial Left Medial Right Lateral Left Lateral
6B. Number of errors (compared to controls [ in the first 100 trials] and patients wi th frontal lesions)
45A, 45B, 44, 47/12
7. Multiple frontal systems controlling response speed (Stuss et al., 2005)
7A. Choice Slowing i n reaction time reaction time test (compared to controls)
9, 24, 32s 9/46v, 46
7B. Prepare First and forth replications of reaction time prepare reaction time with 1
second of warning (compared to controls)
9, 24, 32s
7C. Prepare Second and th i rd replications of 9, 24, 32s reaction time prepare reaction time with 3
seconds of warning (compared to controls)
6A, 9/46d, 9/46v, 46, 45A
8. Effects of focal frontal lesions on response inhibition (Picton et al., 2006)
8A. Go/No Go Slowing i n reaction time (compared to controls)
9, 32s, 24s, 25

8B. Go/No Go Variability in reaction time (compared to controls)
9, 32s, 24s, 24i
44, 45A, 47/12
8C. Go/No Go False alarms in no-go blocks (compared to controls)
6A, 8B 6A, 8B
9. Inhibitory control is slowed i n patients with right superior frontal damage (Ploden & Stuss, 2006)
9A. Stop Signal Task
Slowing i n reaction time (compared to controls)
6A, 8B, 9
10. Keeping time: effects of focal frontal lesions (Picton et al., 2006)
10A. Tone paced condition
Variability to inter-response interval (compared to controls and patients wi th frontal lesions)
44, 45B
10B. Self-paced condition
Variability to inter-response interval (compared to controls and patients wi th frontal lesions)
45A, 8Ad
10C. Accuracy and Deterioration in performance variability of (compared to controls) t iming
9,10s, 32s 6A, 8B, 24s, 32s
9/46d, 10s 9, 8B, 8Ad, 9/46d, 45B
11. Regional frontal injuries cause distinct impairments in cognitive control
11A. Modified Stroop test
Slowing i n reaction time (compared to controls and patients wi th frontal lesions)
6A, 9, 32s, 24s, 25
9, 25 8Av, 9/46d, 9
(Alexander et al., 2007)
(Continued)

Table 4.1. (cont'd)
Study Condition
11B. Modified Stroop test
Variables
False-positive errors (compared to controls and patients w i t h frontal lesions)
Right Medial Left Medial Right Lateral Left Lateral
9/46v, 47/12, 45B
12. Multiple effects of performance lesions on task-switching (Shallice et al., 2008)
12A. Single task condition
Number of errors (compared to controls)
6A 9/46v
12B. Long Cue and Short Cue conditions combined
Number of errors (compared to controls)
9,14
13. The multiple dimensions of sustained attention (Shallice etal. , 2008)
13A. Slow condition
Deterioration of performance (compared to patients with frontal lesions)
10s, 9, 32s, 24s
9, 24s 10s
13B. Fast condition
Deterioration from low number of targets to high number of targets in the fast condition (compared to patients wi th frontal lesions)
9, 32s, 24s 6A, 8Av, 6B, 44,45A, 46

Notes: The first column specifies the paper names. The second column specifies 44 conditions in which patients w i t h frontal lobe lesions were found to be impaired. The third column specifies the variables measured for each of the conditions.
The remaining four columns specify Brodmann areas that represent the localization of the lesions in the impaired conditions in respect to two brain surfaces (i.e., lateral and medial) and the two hemispheres (i.e., left and right). Typically, lesions encompassed more than one region in any one patient. In all conditions the significance level wasp < 0.05.
Abbreviations: v = ventral, d = dorsal, I = inferior, s = superior. Technical note: I n most of the papers there was no specific classification of the lesion localization in terms of Brodmann areas. To include as many conditions as possible
but at the same time include conditions where the localization is accurate and specific, we used different criteria for the lesion localization across the papers as follows: 1. In papers 1 and 5 lesion localization were not specified i n terms of Brodmann areas. As suggested to us by Prof. Stuss and Dr. Alexander, the classification of the
lesion localization was as follows: Lesions in "inferior medial PFC" were referred to in this analysis as BA 10,11, 25, and 32 on the medial surface, while lesions in the "superior medial" region were classified as BA 6A, 8B, 9, and 24 on the medial surface. This procedure was adopted from Levine et al. (1998). Lesions on the left or right lateral surface were classified as BA 44,45A, 45,47/12,11,10,46, 9/46, 9, 8B and 8A, lesions involving dorsolateral PFC were classified as BA 9/46,46,44, 45, 47/12 on the lateral surface, and lesions i n the parietal lobe were classified as BA 5, 7, 40 on the lateral surface.
2. I n paper 2 for the two conditions reported, a specific classification of the patients' lesions in terms of Brodmann areas was given (BA 8B, 9,10s in the right hemisphere).
4. For paper 3 a classification in terms of Brodmann areas was given to two groups of patients: "Superior Medial" (BA 6, 8, 9,10s, and 24) and "Inferior Medial" (BA lOi, 11,12, 25, and 32). For the other groups of patients (i.e., right and left dorsolateral PFC) the classification was similar to those i n papers 1 and 5.
5. I n paper 4 a specific classification i n terms of Brodmann areas was reported, in accordance with the cytoarchitectonic areas adopted by Petrides and Pandya (1994), to the following groups: the left or right lateral surfaces (BA 9/46, 44, 45, 46, 6A, 6B, 8A), "Superior Medial" (BA 8B, 9, 32s), and "Inferior Medial" (BA 10,11,14).
6. In papers 6-13 there was no classification in terms of Brodmann areas for the groups of patients. Thus, for these papers only conditions in which lesion localizations were specified in terms of Brodmann areas (e.g., usually i n a figure) could be included i n the analysis.
7. I n all papers brain areas representing localization of lesions were illustrated in the figure in the following way: BA 24,10, and 32 were divided into the inferior and superior subregions, and BA 9/46, and 8A were divided into the ventral and dorsal subregions.

68 MIND AND THE FRONTAL LOBES
episodic recognition memory) , and so f o r t h : al l the studies so far reviewed here detail patients w i t h rostral PFC damage but whose abilities i n these kinds of domains is unaffected. Nor does i t seem likely t h a t the screening procedure used by Stuss, Alexander, and their colleagues led to an abnormal degree of rejection of rostral PFC patients: the clinical wisdom of many years is that rostral PFC is relat ively silent i n terms of neurological symptoms (indeed, this was one of the reasons for the relative lack of progress made i n this area u n t i l recent times).
We can also dismiss the suggestion that rostral PFC supports some k i n d of processing related to "intelligence" as defined as a "g"-type processing resource. Such a construct must surely have a strong general influence upon behaviors such as choice reaction t ime and other "hard" tasks. Neither would rostral PFC be wel l characterized as relating to some processing reflected i n some "average" of i n t e l ligence tasks. Further, what the Stuss/Alexander findings show is t h a t there appears to be no special relation between rostral PFC lesions and deficits on a range of t radi t ional "executive" tasks either.
From this , we can learn that explanations i n terms of the macro-level constructs tradit ional ly used i n the field (but rarely w i t h detailed formulat ion i n information-processing theory) , such as "planning" or "problem-solving" or "cognit ive control," may be inadequate s tart ing points for theoriz ing about rostral PFC funct ion (see also Burgess, 2000; Burgess et al. , 2000). While i t maybe that rostral PFC supports processing that plays a role i n the performance of tasks that have been labeled as, for example, "analogical reasoning," i t is doubtful that this construct adequately defines the characteristics of the task that particularly require that processing (Voile et al., 2010). Indeed, i n many cases i t may be a matter of ident i fy ing the common processing across tasks that prima facie look quite different (i.e., use different s t imul i , largely different instructions, different responses, etc.) rather t h a n the defining features of a particular task (for neuroimaging evidence see the studies of Burgess and colleagues, who have made extensive use of "cognitive conjunction"-type designs; e.g., Burgess et al. , 2001 , 2003; Gilbert et al. , 2005; Simons et al. , 2005).
The second finding f r o m the Stuss/Alexander meta-analysis is that the deficits that can most precisely be at tr ibuted to rostral PFC lesions seem to relate to quite specific situations. None of the Stuss/Alexander studies implicated only area 10 as critical to performance of a particular variable: instead, they identi f ied bra in regions where area 10 was one of the regions that , when impaired, caused impair ment . However, this makes the results even more remarkable. I f one considers just those variables where lesions t h a t included rostral PFC (area 10) caused an impairment that was significantly worse t h a n i n patients w i t h lesions elsewhere, the result is a very restricted subset of the 44 variables indeed. Patients w i t h lesions t h a t included rostral PFC showed impairments i n humor judgment, i n 2 (only) of the 10 Wisconsin Card Sorting Test (WCST) variables considered by Stuss and Alexander, and i n mainta in ing performance i n time-keeping tasks, and were worst dur ing slow conditions of sustained at tent ion tasks. W i t h this latter

Rostral Prefrontal Cortex 69
finding, there is an interesting concordance w i t h the findings of Petrie (1952), more t h a n 50 years earlier, which suggested that rostral PFC lesions can lead to a variable response to distraction, and the sense that t ime passes more quickly, b u t leaving other cognitive abilities intact .
Towards a Theory of Rostral PFC Function, Integrating Human Lesion Data with Functional Neuroimaging So, i f the wide array of cognitive constructs relating to I Q and many tradi t ional executive abilities are to be dismissed as a start ing po int for theoriz ing about rost r a l PFC funct ion, where instead might one start? Burgess and colleagues (2009) argue that the most te l l ing finding is that many people w i t h rostral PFC pass t r a dit ional tests b u t show handicaps i n everyday behavior. We argue that the reason for this is that there are many skills required by everyday life activities that are no t captured by tradi t ional neuropsychological (and IQ) tests, and i t may be these skills that are principally affected by rostral PFC lesions.
So what might these abilities be? I n fact, when one starts to analyze the demands of what makes a human effective i n real-life situations, i t quickly b ecomes apparent t h a t there are very many mental abilities that might no t be well estimated by tradi t ional neuropsychological tests. The most obvious are those relating to social and emotional behavior. But other impor tant abilities relate to insight and introspective ability, and mentalizing more generally (mduding "theory of mind" ) . To this we might add source and context memory, and a whole raf t of capadties relating to one's abil ity to organize behavior over long periods of t ime, and i n the absence of external cues (e.g., mmti task ing , prospective memory) . No doubt there are also many additional mental capadties that exist that we have no t even yet identif ied.
Impairments in Everyday Life in the Context of High IQ: A Common Occurrence Following Rostral PFC Lesions Perhaps unsurprisingly, therefore, impairments i n the abi l i ty to cope w i t h the demands of everyday life fo l lowing f ronta l lobe involvement are n o t uncommon. They also can occur i n the context of preserved I Q (for review, see Burgess et al. , 2009).
Probably the first case of this k i n d for which good neurological documentation exists was reported by Penfield and Evans (1935). They described the symptoms

Table 4.2. Comparisons of Number of Variables Involved Across Brain Regions
Table 1. Frontal
Condition 10s lO i 11 14 47/12 25 46 32i 32s 45A 45B
1A. Letter-based fluency task
LI LI LI LI
I B . Letter -based fluency task
LI LI LI LI
Lm
1C. Semantic fluency task
LI LI LI LI
2A. Verbal Humour Statement
Rm
test* Rl
2B. Verbal Humour Statements
Rm
test* Rl
3A. WCST variants: 3A. WCST 128
Rm
Lm
Rl
LI
Rl
LI
Rl
LI
Rl
LI
3B.WCST 128 Rm Rl Rl Rl Rl
Lm LI LI LI LI
3C. WCST 128 Rm Rl Rl Rl Rl
Lm LI LI LI LI
3D. WCST 64A Rm Rm Rm Rl Rm Rl Rm Rm Rl Rl
Lm Lm Lm LI Lm LI Lm Lm LI LI
3E.WCST 64A Rm Rl Rl Rl Rl
Lm LI LI LI LI
3R WCST 64A Rm Rl Rl Rl Rl
Lm LI LI LI LI
3G.WCST64A* Rm Rm Rm Rm Rm
Lm Lm Lm Lm Lm
70

Parietal
:>9 9/46v 9/46d 24i 24s 44 8B 8Av 8Ab 6A 6B 5 7 40
i n LI LI
LI Rm Rm LI Rm Rm
Lm Lm Lm Lm
I : L1 LI LI LI LI LI
liRm Rm
|m Rl
f . Rm
p i RL
iem Rl . Rl Rm Rm Rl Rm Rm
| l m LI LI Lm Lm LI Lm Lm
P i n Rl Rl Rm Rm Rl Rm Rm
fern LI LI Lm Lm LI Lm Lm
fern Rl Rl Rm Rm Rl Rm Rm
ifcm LI LI Lm Lm LI Lm Lm
pRm Rl Rl Rm Rm Rl Rm Rm
p,ra LI LI Lm Lm LI Lm Lm
&m Rl Rl Rm Rm Rl Rm Rm
fLm LI LI Lm Lm LI Lm Lm
fem Rl Rl Rm Rm Rl Rm Rm
| t m LI LI Lm Lm LI Lm Lm
*— (Continued)
71

Table 4.2. (cont'd)
Table 1. Frontal
Condition 10s lOi 11 14 47/12 25 46 32i 32s 45A 45B
3H. WCST 64B Rm Rl Rl Rl Rl
Lm LI LI LI LI
31. WCST 64B* Rm
Lm
3 J. WCST 64B Rm
Lm
Rl Rl Rl Rl
4A. Color naming condition
LI LI LI
4B. Incongruent condition
Rm
Lm
5A. Simple task
5B. Easy task*
LI LI LI LI LI LI LI
SC. Easy task
Rl Rl Rl Rl Rl Rl Rl
5D. Complex task
Rm Rm
Lm Lm
Rm
Lm
Rm
Lm
Rm Rm
LI LI LI LI LI LI LI
5E. Complex task
5P. Complex task * Rl Rl Rl Rl Rl Rl Rl
5G. Redundant task
LI LI LI LI LI LI LI
72

Parietal
9 9/46v 9/46d 24i 24s 44 8B 8Av 8Ab 6A 6B 5 7 40
Rm
Lm
Rl
LI
Rl
LI
Rm
Lm
Rm
Lm
Rl
LI
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rl Rl Rm
Lm
Rm
Lm
Rl Rm
Lm
Rm
Lm
LI LI LI LI LI LI LI
Rm Rm
Lm Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
LI LI LI LI LI LI LI
Rm
Lm
Rl Rl Rl
Rm
Lm
Rm
Lm
Rl
Rm
Lm
Rl Rl Rl
Rm
Lm
Rm Rm Rm Rm Rm
Lm
LI LI LI
Lm Lm
LI
Lm
LI LI LI
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rm
Lm
Rl Rl Rl Rl Rl Rl Rl
Rm Rm Rm Rm Rm
Lm
LI LI LI
Lm Lm
LI
Lm
LI LI LI
Lm
(Continued)
73

Table 4.2. (cont'd)
Table 1. Frontal
Condition 10s lOi 11 14 47/12 25 46 32i 32s 45A 45B
5H. Redundant task
51. Second simple task
6A. Slowing inRT
Rm
6B. Number of Errors
LI LI LI
7A. Choice RT test
Rl Rm
7B. Prepare RT Rm
7C. Prepare RT Rl Rm Rl '
8A. Go-no Go Rm Rm
8B. Go-no Go Rl Rm Rl
8C. Go-no Go
9A. Stop Signal Task
10A. Tone paced Rl
10B. Self-paced condition
Rl
IOC. Accuracy and variability of
Rm Rm
tinting Rl Lm
LI
74

Parietal
9 9/46v 9/46d 24i 24s 44 8B 8Av 8Ab 6A 6B 5 7 40
Rm Rm Rm Rm Rm
Lm Lm Lm Lm Lm
Rm Rm Rm Rm Rm
Lm Lm Lm Lm Lm
Rm Rl Rl Rm Rm
Lm
LI
Rm Rl Rm Rm
Rm Rm Rm
Rm Rl Rl Rm Rm Rl
Rm Rm
Rm Rm Rm Rl
Lm
LI
Lm
LI
Rm Rm Rm
Rl
Rl
Rm
Rl Lm Lm Lm
LI LI LI LI
(Continued)
75

Table 4.2. (cont'd)
Table 1. Frontal
Condition 10s lOi 11 14 47/12 25 46 32i 32s 45A 45B
11A. Modified Stroop
Rm Rm
Lm
11B. Modified Stroop test
LI LI
12A. Single task
12B. Long Cue + Lm Short Cue
13A. Slow condition *
Rm Rm
Rl
13B. Fast condition
Rl Rm Rl
Notes: Data are from the Stuss/Alexander series (see Table 4.1 for details of studies and variables). Each patient may have involvement of multiple brain regions. Each suhregion is split into hemisphere (R or L), and lateral or medial surface (1 or m). These studies identified, i n group comparisons, an aspect of area 10 as being part of some larger region specifically involved i n 7/44 variables (marked i n italics, and with an asterisk). For variables 2A, 2B, 3G, 5B, and 5F, the comparison was made between patients whose damage may well have (according to the Stuss/Alexander method of lesion localization; see footnote to Table 4.1 for details) included area 10 and controls (with no other patient group being impaired). For variable 31 the comparison was made between patients whose damage may have included area lOandofher patient groups (the control group was not tested on this condition). Finally, for variable 13A the comparison was made between patients whose damage included (but was not restricted to) area 10 and other patient groups (with no other patient group being impaired relative to controls).
76

Parietal
9/46v 9/46d 24i 24s 44 8B 8Av 8Ab 6A 6B 5 7 40
Rm Rm
Rl Rl
LI
LI Rl
Rm
Lm
Rm Rl Rl 'Rl Rl
77

78 MIND AND THE FRONTAL LOBES
that Penfield's sister was experiencing after the removal of a r ight f ronta l oligodendroglioma: "She had planned to get a simple supper for one guest (Wilder Penfield) and four members of her own family. She looked forward to i t w i t h pleasure and had the whole day for preparation. This was a t h i n g she could have done w i t h ease . . . before. W h e n the appointed hour arrived she was i n the kitchen, the food was all there, one or two things were on the stove, b u t the salad was no t ready, the meat had not been started and she was distressed and confused by her long continued effort alone. I t seemed evident that she would never be able to get everything ready at once" (p. 131).
Penfield's sister was no t suffering f r o m a marked and generalized cognitive dedine, and d id n o t have serious disabilities i n basic cognitive systems (e.g., das-sic dense amnesia, visuo-spatial/perceptual or agnosic problems, disorders of motor control and so f or th ) . Similar cases were soon reported (e.g., Ackerly & Benton, 1947; Brickner, 1936). These established, at least on the grounds of c l in i cal observation alone, that behavioral disorganization can be seen i n the absence of gross cognitive dedine, harking back to the most famous case of f ronta l lobe damage: Harlow's Pbineas Gage.
However, i t was no t u n t i l the mid-1980s that an attempt was made to characterize the nature of the critical cognitive deficit underpinning this type of disorder. Eslinger and Damasio (1985) described the case of E.V.R., who had undergone surgical removal of a large bilateral f ronta l meningioma. A t the time of his operation E.V.R. was a finandal officer w i t h a small company and a respected member of his community. He was marr ied and the father of two children; his brothers and sisters considered h i m a role model and a natural leader. After the operation, however, E.V.R. lost his job, went bankrupt , was divorced by his wife, and moved i n w i t h his parents. He subsequently marr ied a prost itute and was divorced again w i t h i n 2 years. Extensive psychological evaluations demonstrated that he was superior or above average on most intelligence tests (e.g., Verbal I Q of 125; Performance I Q of 124) and, indeed, on most other kinds of cognitive test as well . Despite these normal findings, however, E.V.R. was often unable to make simple everyday decisions, such as which toothpaste to buy, what restaurant to go to , or what to wear. He would instead make endless comparisons and contrasts, often being completely unable to come to a dedsion at all . Further, Eslinger and Damasio report prospective memory problems: " i t was as i f he forgot to remember short -and intermediate- t e r m goals" (1985, p. 1737).
Eslinger and Damasio's paper was impor tant because i t was the first to present a formal neuropsychological examination of this k i n d of presentation, thus adding i n an impor tant way to Penfield and Evans, and the others who had presented only formal neurological data. However, Eslinger and Damasio's study s t i l l d id n o t quanti fy the most critical aspect of the presentation: the nature of the cognitive impairment .
A study by Shallice and Burgess (1991) addressed this lacuna. They presented three cases who had all suffered f ronta l lobe damage fol lowing traumatic

Rostral Prefrontal Cortex 79
brain in jury . None had any significant impairment on formal tests of perception, language, and intelligence, and two performed wel l on a variety of t radi t ional tests of executive funct ion. Indeed, one of these cases (A.P.) was probably the best example of the syndrome so far reported. A.P. sustained an open head i n j u r y i n a road traffic accident when he was i n his early 20s. The i n j u r y caused a v i r tua l ly complete removal of the rostral prefrontal cortex bilaterally plus damage to surrounding regions. On standard neuropsychological measures of intellectual funct i on ing , memory, and perception and even tradi t ional tests of executive funct ion, A.P. performed w i t h i n the superior range.
But A.P. d id show impairment i n everyday life. The most obvious was a marked mult i tasking and prospective memory problem. This manifested itself as t a r d i ness and disorganization, the severity of which ensured that despite his excellent intellect and social skills, he never managed to r e t u r n to work at the level he had enjoyed premorbidly. Shallice and Burgess (1991) invented two new tests of m u l t i tasking to assess these problems. The first, the "Mult ip le Errands Test," is a real-life mult i tasking test carried out i n a shopping m a l l or similar area. Participants have to complete a number of tasks, principally involving shopping, while fol lowi n g a set of rules (e.g., no shop should be entered other t h a n to buy something). The tasks vary i n terms of complexity (e.g., buy a small brown loaf vs. discover the exchange rate of the Euro yesterday), and there are a number of "hidden" problems i n the tasks t h a t have to be appreciated and the possible course of action evaluated. For instance, one i t e m asks that participants wr i te and send a postcard, yet they are given no pen, and although they cannot use anything no t bought on the street to help them, they are also t o l d that they need to spend as l i t t l e money as possible. I n this way, the task is quite "open-ended" or "ffl-structured" (i.e., there are many possible courses of action, and i t is up to the individual to decide which one to choose). (For fur ther studies using versions of this task see, e.g., Alderman et al., 2003; Dawson et al., 2009 Knight et al., 2002.)
The second task that Shallice and Burgess invented was a more controlled experimental task, the "Six Element Test." This requires participants to swap efficiently between three simple subtasks, each divided into two sections w i t h i n 15 minutes, while fo l lowing some arbitrary rules (e.g., "you cannot do par t A of a subtask followed immediately by part B of the same subtask"). There are no cues signaling when to switch tasks, and although a clock is present, i t is covered, so that checking i t has to be a deliberate action. Thus, this paradigm has a strong component of vo luntary time-based task switching (i.e., one f o r m of prospective memory) . (A version of this task is now par t of the Behavioral Assessment of the Dysexecutive Syndrome [BADS] assessment battery [Wilson et al. , 1996].)
Despite their excellent general cognitive skills, A.P. and the other patients reported by Shallice and Burgess all performed these tasks below the 5 t h percentile of age- and IQ-matched controls. O n the Mul t ip le Errands Task the subjects made a range of types of error. Many of these could be interpreted as problems w i t h prospective memory. For instance, they would find themselves having to go

80 MIND AND THE FRONTAL LOBES
in to the same shop more t h a n once to buy items that could all have been bought at one vis i t ; they forgot to carry out tasks that they had previously learned that they needed to do, or to fol low task rules. They also made a range of social behavior errors (e.g., leaving a shop w i t h o u t paying; offering sexual favors i n l ieu of payment) . Shallice and Burgess (1991) termed this k i n d of behavioral disorganization i n the context of preserved intellect and other cognitive functions the "Strategy Appl icat ion Disorder."
ShalHce and Burgess's patients gave l i t t l e due as to the anatomical localization of the lesion critical for this pat tern of def idt , since the patients had suffered large traumatic lesions that invaded many subregions. But 2 years later, Goldstein and colleagues (1993) described an instructive case. G.N., a 51-year-old right-handed man, had undergone a le f t f ronta l lobectomy 2.5 years earlier fo l lowing the discovery of a f ronta l lobe t u m o r (mixed astrocytoma-oligodendroglioma). A 5-cm resection of le f t f ronta l lobe f r o m the f ronta l pole was undertaken. This surgery made l i t t l e difference to his general cognitive abilities (e.g., WAIS-R Verbal IQ. 129, Performance I Q 111 ; story recall, immediate 7 5 - 9 0 t h percentile, delayed 5 0 - 7 0 t h ; Rey Osterreith delayed figure recall 8 0 - 9 0 f h percentile; Tran-making 7 0 - 7 5 t h percentile). However, i t was neverthdess dear f r o m his everyday behavior that something was seriously wrong. He had held a senior management posit i o n w i t h i n an international company, but 2 years after surgery he had to take medical retirement because of increasing lethargy. He worked f r o m home as a freelance management consultant b u t had difficulty making dedsions. For instance, he took 2 weeks to dedde which slides to use for a work presentation, b u t never actually came to a condusion. He also experienced anger-control difficulties.
Goldstein and colleagues (1993) administered the Mul t ip le Errands Test. G.N. made significantly more errors t h a n controls, being less efficient (e.g., having to r e t u r n to a shop), breaking rules (e.g., using a stamp that another customer gave h im) , mis interpret ing tasks (e.g., sticking the stamp on the wrong card), as wel l as fa i l ing to complete some tasks altogether, report ing that he had k n o w n he had to do t h e m b u t somehow "forgot" them. He also showed some "sodal ru le" breaks. For instance, he had omit ted to f ind out the price of tomatoes while earlier i n the greengrocers, and realizing t h a t he should no t go back into the shop unless he was to buy something, he very conspicuously climbed onto the f r u i t display outside the shop and peered i n the shop window.
This case and others reported i n the l iterature show a remarkably similar patt e r n of neuropsychological test performance. Burgess (2000) summarized the performance of eight wel l -known patients. None of t h e m had any language or visuo-perceptual impairment , and all scored w i t h i n the superior range on tests of current intellectual functions. Four of the seven showed no impairment on any memory test. But, most remarkably, two showed no impairment on a range of c l inical executive funct ion tests k n o w n to be sensitive to frontallobe lesions. Moreover, no executive test has been failed by more than 2/8 cases. This contrasts w i t h the observation that all of the reported cases of "strategy application disorder"

Rostral Prefrontal Cortex 81
who have been given either the Mult ip le Errands or Six Element Test have failed at least one of them.
Burgess and colleagues (2009) po inted out that the mult i tasking situations presented to a partic ipant by the Mult ip le Errands and Six Element Tests share a number of similarities. These are:
1 . A number of discrete and different tasks have to be completed. 2. Performance on these tasks needs to be dovetailed i n order to be t i m e -
effective. 3. Due to either cognitive or physical consfraints, only one task can be performed
at any one t ime. 4. The times for r e t u r n to task are no t signaled directly by the situation. 5. There is no moment-by-moment performance feedback of the sort that par
ticipants i n many laboratory experiments w i l l receive. Typically, failures are no t signaled at the t ime they occur.
6. Unforeseen interrupt ions , sometimes of h igh pr ior i ty , w i l l occasionally occur, and things w i l l no t always go as planned.
7. Tasks usually differ i n terms of pr ior i ty , difficulty, and the length of t ime they w i l l occupy.
8. People decide for themselves what constitutes adequate performance.
None of these features are characteristic of t radi t ional neuropsychological tests, nor the types of procedures that are typically used i n a neurological examinat ion . They are, however, very common aspects of everyday situations.
These patients show a single dissociation between performance on tradi t ional neuropsychological tests and abil i ty to cope w i t h situations requir ing complex behavioral organization. However, some might argue that these everyday situations are somehow "harder" or "more complex" or simply sample behavior over a longer t ime frame (and thus might more easily detect problems t h a t occur only occasionally). On these grounds one might argue t h a t the dissociation is artifac-tual . To refute this possibility, Burgess and colleagues (2009) reported two patients who performed well on the Mult ip le Errands Test despite showing marked impair ment on tests of memory and IQ. The to ta l i ty of the evidence f r o m single cases therefore suggests a double dissociation between memory, I Q , and complex behavioral organization, at least under certain conditions.
Localization of Deficits in Behavioral Organization that Cannot Be Attributed to IQ Impairment Stuss and Alexander's group are not the only ones to have reported group studies indicating the nature of the impairment caused by rostral PFC lesions. Burgess and

82 MIND AND THE FRONTAL LOBES
coworkers (2000) examined 60 acute neurological patients f r o m an unselected series of referrals (approximately three quarters of w h o m were suffering f r o m brain tumors) . Their performance was compared w i t h a group of 60 age- and IQ-matched healthy controls. A l l participants were given a mult i tasking test called the Greenwich Test. This is a mult i tasking test t h a t follows the principles of the Six Element Test, but i n contrast the major i ty of the variance i n performance of the test comes f r o m rule infractions rather t h a n task-switching problems. Participants were presented w i t h three different simple tasks and t o l d that they had to attempt at least some of each of the tasks i n 10 minutes, while fo l lowing a set of rules. One of these rules relates to all subtests ("in al l three tasks, completi n g a red i t e m w i l l gain you more points t h a n completing an i t e m of any other color") and there are four task-specific rules (e.g., " i n the tangled lines test you must no t mark the paper other t h a n to wr i te your answers down"). The task was administered i n a f o r m that allowed consideration of the relative contributions of task rule learning and remembering, planning, plan-fol lowing, and remembering one's actions to overall mult i tasking performance. Specifically, before participants began the test, the ir abi l i ty to learn the task rules (by b o t h spontaneous and cued recall) was measured; this measure was called "Learn." They were then asked how they intended to do the test, and a measure of the complexity and appropriateness of the ir plans was gained (a variable called "Plan"). This enabled us to look at whether their failures could be due to poor p lanning (see, e.g., Kliegel et al., 2005). The participants then performed the task itself, and by comparing what they d id w i t h what they had planned to do, a measure of "Plan Eollowfng" was made. Mul t i task ing performance (the number of task switches minus the number of rule breaks) was referred to as the test "Score." After these stages were finished, subjects were asked to recollect their own actions by describing i n detail what they had done ("Recount"). Finally, delayed memory for the task rules was examined ("Remember").
Burgess and colleagues (2000) f ound that lesions i n different bra in regions were associated w i t h impairment at different stages i n the mult i tasking procedure. For instance, lesions to a large region of superior posterior medial cortex including the left posterior cingulate and forceps major gave deficits on all measures except planning. Remembering task contingencies after a delay was also affected by lesions i n the region of the anterior cingulate. Critically, however, Burgess and colleagues found that patients w i t h le f t hemisphere rostral PFC lesions, compared w i t h patients w i t h lesions elsewhere, showed a significant m u l titasking impairment ("Score" variable) despite no significant impairment on remembering task rules ("Remember" variable). Indeed, the le f t rostral prefrontal cases showed no significant impairment on any variable except the one reflecting mult i tasking performance. I n other words, despite being able to learn the task rules, f o r m a plan, remember their actions, and say what they should have done, they nevertheless d id no t do what they said they intended to do. These impair ments appeared independently of any I Q decrement.

Rostral Prefrontal Cortex 83
But perhaps these results were specific to this particular mult i tasking test rather t h a n being true of a class of s i tuation w i t h the previously described characteristics? This appears n o t to be the case. Burgess, Veitch, and Costello (submitted; reported i n Burgess et al. , 2006) administered a new version of Burgess and colleagues' (1996) Six Element Test to 69 acute neurological patients w i t h circumscribed focal lesions and 60 healthy individuals using the administrat ion framework of Burgess and colleagues (2000). The Six Element Test differs f r o m the Greenwich Test i n that the mmti task ing score reflects mainly vo luntary t i m e -based switching rather t h a n rule-fol lowing. Compared w i t h other patients, those whose lesions involved the rostral prefrontal regions of the r i ght hemisphere made significantly fewer vo luntary task switches, attempted fewer subtasks, and spent far longer on mdividual subtasks. As w i t h the study by Burgess and colleagues (2000), these mult i tasking deficits could no t be at tr ibuted to deficits i n general intellectual funct ioning, rule knowledge, p lanning, or retrospective memory. Furthermore, there was no obvious relation between Six Element Test performance and concomitant failure on several tradi t ional tests of executive funct ion.
W h e n one considers the earlier case studies report ing relatively isolated deficits of behavioral organization i n the l i ght of the results f r o m these group studies, i t seems that that there is a remarkably consistent fmcling of involvement of rost r a l PFC. For instance, i n the six cases reviewed by Burgess (2000) for w h o m good brain scan data were available, al l of t h e m had rostral PFC involvement of either the le f t or r i ght hemispheres (or both) . I n addit ion to these cases, we might also add the more recent patient reported by Bird and associates (2004) who had suffered a rare f o r m of stroke affecting the medial aspects of area 10 bilaterally, and who failed the Six Element Test despite passing some other executive tests (e.g., the WCST).
From Human Neuropsychology to Neuroimaging: Constraints for Theorizing I f one uses these results f r o m human neuropsychology to pr ior i t ize the potent ia l functions that one might seek to explore using functional neuroimaging, which functions, more precisely, might one target?
Burgess and colleagues (2001, 2003) hypothesized that a key cognitive deficit behind t h a t behavioral organization impairment shown by patients w i t h rostral PFC damage might concern prospective memory. Prospective memory is the abi l i t y to remember to carry out an intended action (or thought) after a delay period where one has been occupied w i t h another task. A f u l l review of the conclusions f r o m these studies is given i n Burgess and colleagues (2008) and is beyond the scope of this chapter. However, i n brief, we (and others; see, e.g., Reynolds, West, & Braver, 2009) have found a consistent relation between hemodynamic changes

84 MIND AND THE FRONTAL LOBES
i n b o t h medial and lateral rostral PFC and performance of prospective memory-tasks. Moreover, recent data f r o m our lab f r o m human group lesion studies of prospective memory tasks that are very similar to the ones we have used i n our neuroimaging studies are also showing a relation between rostral PFC lesions and failures on these tasks (Voile, Costello, Gonen-Yaacovi, Gilbert, & Burgess, 2011). Broadly, we f ind that lateral rostral PFC activation increases dur ing prospective memory conditions, w i t h higher medial rostral PFC activation during the ongoing task only. These activation patterns can occur regardless of the precise f o r m (e.g., words, pictures, numbers, etc.) of the s t imul i ( w i t h the potential ly highly impor t a n t caveat that we, and others, have so far used only visual presentation of our st imul i ) . Some regions of rostral PFC also appear hemodynamically insensitive to the demands of the intended action or how difficult i t is to spot the prospective memory target. However, rostral PFC activation patterns do seem highly sensitive to even very small changes i n task instruct ion (Gilbert et al. , 2009).
Here, then, is one example of broad congruence between findings f r o m neuroimaging and neuropsychology, and where each method seems to be providing hypotheses and tests relevant to the other method. Another potential funct ion is mentaiizing, and related functions. "Mental iz ing" is a broad t e r m referring to a range of cognitive processing relating to t h i n k i n g about one's own thoughts or characteristics and those of others (e.g., Theory of M i n d and introspection; for review see F r i t h & F r i t h , 2003). There is a consistent relationship i n the funct ional neuroimaging literature between tasks that stress these functions, and related ones such as deception and moral behavior, and activation of medial rost r a l PFC (e.g., Eslinger et al., 2009; Gilbert et al. , 2007). There is also supportive evidence f r o m human lesion studies (e.g., Stuss et al. , 2001) and transcranial direct current s t imulat ion (Kar im et al. , 2010). O n these grounds, i t seems plausible that the root of at least some of the deficits i n social behavior f ound i n the cases of "strategy application disorder" described previously might also be traced to impairments i n these abilities, although to our knowledge this has no t yet been formally exarnined.
Another understudied area of human cognition where there may be potential for f r u i t f u l cross-method interaction concerns higher-level memory control, by which we mean the abil i ty to manipulate and introspect upon episodic memory traces. Examples include memory for Context (remembering details of an event that were no t central to i t at the t ime of encoding), Source (remembering aspects of events relating to its provenance, such as when something occurred, where i t occurred, etc.), and Reality Monitoring (distinguishing between imagined and perceived events). There is a very consistent relation i n the l iterature between performance of these tasks and activation i n rostral PFC (see Burgess et al., 2007, for review) and elsewhere w i t h i n the prefrontal cortex (e.g., dorsolateral and ventrolateral PFC; see Turner et al. , 2008). A n d i t has been k n o w n for some t ime that patients w i t h f ronta l lobe lesions have difficulties on a range of source m o n i tor ing tasks (e.g., Duarte et al. , 2010; Janowsky, Shimamura, & Squire, 1989;

Rostral Prefrontal Cortex 85
Kopelman, Stanhope, & Kingsley, 1997; Milner, Petrides, & Smith, 1985; Shimamura, Janowsky, &c Squire, 1990). However, i t seems on present evidence that these kinds of higher-level memory control tasks require the operation of a network of PFC and extra-PFC regions, and the precise role that rostral PFC may play w i t h i n this network is unclear.
The Gateway Hypothesis of Rostral PFC Function One account that attempts to help, and that seeks to explain simultaneously the involvement of rostral PFC i n a wide range of tasks, is the "gateway hypothesis" of Burgess and colleagues (Burgess et al. , 2005, 2006, 2007). Indeed, the hypothesis tries to provide a uni fy ing framework for understanding common processing requirements across the many different types of tasks where rostral PFC involvement has been implicated. I t suggests t h a t one impor tant funct ion of rostral PFC is i n enabling a person to effect control over the degree to which he or she attends to externally available s t imul i , or alternatively to stimulus-independent thought (i.e., self-generated thoughts i n our heads).
That we can effect such control can hardly be i n doubt: there are many situations where one needs to exert such control, ranging f r o m preventing mind -wandering (or deliberately engaging i n i t ) , to having to attend dur ing bor ing tasks, and also including many i f no t most situations, even i f only momentarily, where one is faced w i t h a novel s i tuation and does no t know how to proceed. This type of attentional control is l ikely to also be involved where one is dovetailing two or more activities (so has to "bear sometMng i n m i n d " while per forming another task) or where one has to reflect deeply upon one's experience i n order to choose a way to respond i n an ambiguous or open-ended situation (as may occur i n memory control tasks, or complex social situations), or to view that memory trace i n a new l ight . Burgess and his colleagues have invented tasks that measure this "attentional gateway" (i.e., switcl i ing between stimulus-oriented and st imulus-independent attending, or source switching) and have shown a consistent relation between performance of these tasks and rostral PFC activation, and that that these patterns of activation seem to overlap and temporally coincide w i t h those provoked by tasks such as prospective memory tasks (e.g., Beno i te ta l , submitted; Dumonthei l , Gilbert, Burgess, & Otten, 2010; Dumonthei l , Gilbert, F r i t h , & Burgess, 2010; Gilbert et al., 2005, 2006).
I f rostral PFC supports such a mental ability, then one might expect the fol lowing: (1) t h a t funct ional activation i n rostral PFC wou ld be found i n a wide variety of tasks, and no t restricted to one class; (2) these activations might be quite insensitive to the nature of the s t imul i presented (because i t is no t what the person is attending to, i t is the direction of attending that is crucial), b u t may be quite sensitive instead to, for example, changes i n task instruct ion (ditto) , or to aspects no t often considered by experimenters (e.g., h o w b o r i n g t h e task is; degree of novelty

86 MIND AND THE FRONTAL LOBES
w i t h i n partic ipant) ; and (3) that I Q would be le f t relatively unchanged by many intelligence tests (since they do not typically heavily tap aspects such as prospective memory, mmti tasking , high-level memory control, mental izing and i n t r o spection, and so f o r t h . Neither are they typically open-ended, ambiguous, monotonous, or have any of the other task characteristics mentioned here i n relat i o n to good tests of rostral PFC funct ion. Indeed, when one makes a l ist , l ike th is , of the characteristics that I Q tasks do not have, one can begin to see how part i cular, perhaps even peculiar, are typical intelligence tests i n relation to the demands made by everyday li fe.
Conclusion Cognitive neuroscience must be an inherently m u l t i - or inter-ctisdplinary enterprise because there is no single level at which the instant iat ion of mental phenomena can be understood. To understand ful ly how the bra in facilitates thought , we must understand simultaneously how chemical, electrical, hemodynamic, and other properties and changes occurring w i t h i n the bra in can be related to in for mation-processing and phenomenological levels of explanation. I t seems likely therefore that where one particular method does no t constrain the inferences i t makes f r o m one level to another by reference to converging evidence f r o m other methods, the possibility of inferential error increases. This chapter has h i g h l ighted one such way i n which this may be currently occurring. The evidence f r o m human lesion studies of rostral PFC is hard to reconcile w i t h a sizeable proport ion of the theorizing that is emerging f r o m neuroimaging. Here we have t r i ed to pro vide some insights and references that may provide a start towards such reconcil iation by constraining neuroimaging evidence w i t h human lesion evidence.
There is l i t t l e doubt about the enormous increase i n our knowledge of the functions and organization of rostral PFC that functional neuroimaging has afforded. Indeed, as mentioned at the start of this chapter, determined theoriz ing on this topic was kick-started by numerous findings of rostral PFC activations i n neuroimaging experiments, and most of what we currently believe about rostral PFC comes f r o m this method. However, there is a danger, when a new sdenidfic method becomes overwhelmingly predominant, that highly relevant material f r o m more established methods might no t mainta in the influence that i t deserves.
As an investigative method i n cognitive neuroscience, funct ional neuroimaging has many advantages over lesion studies. However, human lesion evidence has two principal advantages over neuroimaging data. The first is the causal advantage for theorizing that lesion evidence provides: inference is much more straightforward. Second, human lesion data often also have an advantage i n terms of understanding the to ta l i ty of influence that perturbat ion of a particular cognitive system evinces. A neurological pat ient can often be observed i n a wide variety of situations, and performing a wide variety of tasks, so i f a particular construct is

Rostral Prefrontal Cortex 87
impaired, one can see the extent of its influence across them. This is no t typically possible w i t h neuroimaging designs, but may be particularly critical for theoriz ing about "central" systems that may operate across many situations and w i t h many types of s t imul i . A n d indeed, i t would appear that some systems supported by prefrontal cortex have just these characteristics. I n this way, the data i n particular f r o m Donald T. Stuss and his colleagues are of critical importance, and they have provided a database that w i l l serve as a guide to theorists for very many years to come. For that accomplishment, those of us who study rostral PFC are extremely grateful.
Acknowledgments We would l ike to thank Donald T. Stuss and Michael Alexander for their detailed and extremely helpful comments on technical matters relating to the metaanalyses of the ir results presented i n this chapter. We are also very grateful to Brian Levine and Fergus Craik for their comments on an earlier draft.
References Ackerly, S. S., & Benton, A. L. (1947). Report of a case of bilateral frontal lobe defect. Research
Publications: Association for Research in Nervous and Mental Disease, 27, 479-504. Alderman, N., Burgess, P. W., Knight, C, & Henman, C. (2003). Ecological validity of a simplified
version of the Multiple Errands Test. Journal of the International Neuropsychological Society, 9, 31-44.
Alexander, M . P., Stuss, D. T., Picton, T., Shallice, T., & Gillingham, S. (2007). Regional frontal in ju ries cause distinct impairments i n cognitive control. Neurology, 68,1515-1523.
Alexander, M . P., Stuss, D. T., Shallice, T., Picton, T. W., & Gillingham, S. (2005). Impaired concentration due to frontal lobe damage from two distinct lesion sites. Neurology, 23, 572-579.
Bird, C. M . , Castelli, E, Malik, O., Fr i th , U., & Husain, M . (2004). The impact of extensive medial frontal lobe damage on "Theory of Mind" and cognition. Brain, 127, 914-928.
Blaxton, T. A., Zeffiro, T. A., Gabrieli, J. D. E., Bookheimer, S. Y., Carrillo, M . C, Theodore, W. H., & Disterhoft, J. F. (1996). Functional mapping of human learning: a positron emission tomography activation study of eyeblink conditioning. Journal of Neuroscience, 16, 4032-4040.
Brickner, R. M . (1936). Tfte intellectual functions of the frontal lobes: a study based upon observation of a man after partial bilateral frontal lobectomy. New York: Macmillan.
Buckner, R. L. (1996). Beyond HERA: contributions of specific prefrontal brain areas to long-term memory retrieval. Psychonomic Bulletin and Review, 3,149-158.
Burgess, P. W. (1997). Theory and methodology i n executive function research. I n : P. Rabbitt (Ed.), Theory and methodology of frontal and executive function (pp. 81-116). Hove, U.K.: Psychology Press.
Burgess, P. W. (2000). Strategy application disorder: the role of the frontal lobes i n human m u l t i -tRsking. Psychological Research, 63, 279-288.
Burgess, P. W., Alderman, N., Voile, E., Benoit, R. G., 8c Gilbert, S. J. (2009). Mesulam's frontal lobe mystery re-examined. Restorative Neurology and Neuroscience, 27(5), 493-506.
Burgess, P. W., Dumontheil, I . , 8c Gilbert, S. J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Sciences, 11, 290-298.

88 MIND AND THE FRONTAL LOBES
Burgess, P. W., Dumontheil, I . , Gilbert, S. J., Okuda, J., Scholvinck, M . L., & Simons, J. S. (2008). On the role of rostral prefrontal cortex (area 10) in prospective memory. I n : M . Kliegel, M . A. McDaniel, 8c G. O. Einstein (Eds.), Prospective memory: cognitive, neuroscience, developmental, and applied perspectives (pp. 235-260). Mahwah, NJ : Erlbaum.
Burgess, P. W„ Gilbert, S. J., Okuda, J., & Simons, J. S. (2006). Rostral prefrontal brain regions (area 10): a gateway between inner thought and the external world? I n : W. Prinz & N. Sebanz (Eds.), Disorders of volition (pp. 373-396). Cambridge, MA: M I T Press.
Burgess, P. W., Quayle, A., & Fri th , C. D. (2001). Brain regions involved i n prospective memory as determined by positron emission tomography. Neuropsychologia, 39, 545-555.
Burgess, P. W., Scott, S. K., 8c Frith, C. D. (2003). The role of the rostral frontal cortex (area 10) in prospective memory: a lateral versus medial dissociation. Neuropsychologia, 41, 906-918.
Burgess, P. W., & Shallice, T. (1996). Response suppression, init iation and strategy use following frontal lobe lesions. Neuropsychologia, 34, 263-273.
Burgess, P. W., & Shallice, T. (1997). The Hayling and Brixton Tests. Thurston, Suffolk: Thames Valley Test Company.
Burgess, P. W., Simons, J. S., Dumontheil, I . , & Gilbert, S. J. (2005). The gateway hypothesis of rostral PFC function. I n : J. Duncan, L. Phillips, & P. McLeod (Eds.), Measuring the mind: speed, control and age (pp. 215-246). Oxford University Press.
Burgess, P. W., Veitch, E., Costello, A., & Shallice, T. (2000). The cognitive and neuroanatomical correlates of multitasking. Neuropsychologia, 38, 848-863.
Cattell, R. B. (1963). Theory of fluid and crystallized intelligence: a critical experiment. Journal of Educational Psychology, 54(1), 1-22.
Chambers, C. D., 8c Mattingley, J. B. (2005). Neurodisruption of selective attention: insights and implications. Trends in Cognitive Sciences, 9, 542-550.
Christoff, K., 8c Gabrieli, J. D. E. (2000).The frontopolar cortex and human cognition: evidence for a rostrocaudal hierarchical organization wi th in the human prefrontal cortex. Psychohiology, 28,168-186.
Christoff, K., Prabhakaran, V , Dorfman, J., Zhao, Z., Kroger, J. K., Holyoak, K. J., 8c Gabrieli J. D. E. (2001). Rostrolateral prefrontal cortex involvement i n relational integration during reasoning. Neuroimage, 14,113 6-1149.
Colom, R., Jung, R. E., & Haier, R. J. (2007). General intelligence and memory span: evidence for a common neuroanatomy: framework. Cognitive Neuropsychology, 24(8), 867-878.
Coltheart, M . (2006) What has functional neuroimaging told us about the mind (so far)? Cortex 42(3), 323-331.
Dawson, D. R., Anderson, N . D., Burgess, P. W., Cooper, E., Krpan, K. M . , & Stuss, D. T. (2009). Further development of the Multiple Errands Test: standardized scoring, reliabiUty, and ecological validity for the Baycrest version. Archives of Physical Medicine and Rehabilitation, 90, S I , 41 -51 .
Dreher, J. C, Koechlin, E., AH, S. O., & Grafman, J. (2002). The roles of t iming and task order during task switching. Neuroimage, 17, 95-109.
Dreher, J. C, Koechlin, E., Tiemey, M . , 8c Grafman, J. (2008). Damage to the fronto-polar cortex is associated w i t h impaired multitasking. PLoS ONE, 3(9), e3227.
Duarte, A., Henson, R. N., Knight, R.T., Emery, T., & Graham, K. S. (2010). Orbito-frontal cortex is necessary for temporal context memory. Journal of Cognitive Neurosdence, 22(8), 1819-1831.
Dubois, B., Slachevsky, A., Litvan, I , , 8c Pillon B. (2000). The Fab: a frontal assessment battery at bedside. Neurology, 55,1621-1626.
Dumontheil, I . , Burgess, P. W., 8c Blakemore S-J. (2008). Development of rostral prefrontal cortex and cognitive and behavioural disorders. Developmental Medicine and Child Neurology, 50, 1-14.
Dumontheil, I . , Gilbert, S. J., Burgess, P. W., & Otten, L. J. (2010). Neural correlates of task and source switching: similar or different? Biological Psychology, 83(3), 239-249.
Dumontheil, I . , Gilbert, S. J., Frith, C. D., 8c Burgess, P. W. (2010). Recruitment of lateral rostral prefrontal cortex i n spontaneous and task-related thoughts. Quarterly Journal of Experimental Psychology, 63(9), 1740-1756.

Rostral Prefrontal Cortex 89
Duncan, J. (2001). A n adaptive coding model of neural function i n prefrontal cortex. Nature Reviews Neuroscience, 2, 820-829.
Duncan, J. (2005). Frontal lobe function and general intelligence: why i t matters. Cortex, 41(2), 215-217.
Duncan, J., 8c Owen, A. (2000). Consistent response of the human frontal lobe to diverse cognitive demands. Trends in Neurosciences, 23, 475-483.
Duncan, J., Riidiger, J. S., Kolodny, J., Bor, D., Herzog, H. , Ahmed, A., Newell, F. N. , & Emslie, H . (2000). A neural basis for general intelligence. Science, 289(5478), 457-460.
Eslinger, P. J., 8c Damasio, A. R. (1985). Severe disturbance of higher cognition after bilateral frontal lobe ablation: patient E.V.R. Neurology, 35,1731-17'41.
Eslinger, P. J., Robinson-Long, M . , Realmuto, J., Moll , J., deOliveira-Souza, R., Tovar-Moll, F., Wang, J., 8c Yang, Q. X. (2009). Developmental frontal lobe imaging in moral judgment: Arthur Benton's enduring influence 60 years later. Journal of Clinical and Experimental Neuropsychology, 31(2), 158-169.
Floden, D., 8c Stuss, D. T. (2006). Inhibitory control is slowed i n patients w i t h right superior medial frontal damage. Journal of Cognitive Neuroscience, 18,1843-1849.
Friston, K. J., Price, C. J., Fletcher, P., Moore, C, Frackowiak, R. S. J., 8c Dolan, R. J. (1996). The trouble w i t h cognitive subtraction. Neuroimage, 4, 97-104.
Frith, TJ., 8c Fr i th , C. D. (2003). Development and neurophysiology of mentalizing. Philosophical Transactions of the Royal Society of London B, 358(1431), 459-473.
Gilbert, S. J., Dumontheil, I . , Simons, J. S., Frith, C. D., 8c Burgess, P. W. (2007). Comment on "Wandering minds: the default network and stimulus-mdependent thought." Science, 317(5834), 43.
Gilbert, S. J., Frith, C. D., & Burgess, P. W. (2005). Involvement of rostral prefrontal cortex in select ion between stimulus-oriented and stimulus-mdependent thought. European Journal of Neuroscience, 21,1423-1431.
Gilbert, S. J., Gollwitzer, P. M . , Cohen, A. L., Oettingen, G., 8c Burgess, P. W. (2009). Separable brain systems supporting cued versus self-initiated realization of delayed intentions. Journal of Experimental Psychology: Learning, Memory, and Cognition, 35(4), 905-915.
Gilbert, S. J., Simons, J. S., Frith, C. D., 8c Burgess, P. W. (2006). Performance-related activity i n medial rostral prefrontal cortex (area 10) during low demand tasks. Journal of Experimental Psychology: Human Perception and Performance, 32, 45-58.
Gilbert, S. J., Williamson, I . D. M . , Dumontheil, I . , Simons, J.S., Frith, C. D., 8c Burgess, P. W. (2007). Distinct regions of medial rostral prefrontal cortex supporting social and nonsocial functions. Social Cognitive and Affective Neuroscience 2(3), 206-216.
Glascher, J., Rudrauf, D., Colom, R., Paul, L. K., Tranel, D., Damasio, H. , 8c Adolphs, R. (2010). Distributed neural system for general intelligence revealed by lesion mapping. Proceedings of the National Academy of Sciences of the United States of America, 107(10), 4705-4709.
Glascher, J., Tranel, D., Paul, L. K. Rudrauf, D., Rorden, C, Hornaday, A., Grabowski, T., Damasio, H., 8c Adolphs, R. (2009). Lesion mapping of cognitive abilities linked to intelligence. Neuron, 61(5), 681-691.
Goel, V., 8c Grafman, J. (2000). The role of the right prefrontal cortex i n ill-structured problem solving. Cognitive Neuropsychology, 17(5), 415-436.
Goldstein, L. H. , Bernard, S., Fenwick, P. B. C, Burgess, P. W., 8c McNeil, J. (1993). Unilateral frontal lobectomy can produce strategy application disorder. Journal of Neurology, Neurosurgery & Psychiatry, 56, 274-276.
Grady, C, L. (1999). Neuroimaging and activation of the frontal lobes. I n B. L. Miller 8c J. L. Cummings (Eds.), The human frontal lobes: function and disorders (pp. 196-230). New York: Guilford Press.
Henson, R. N . (2006). What has (neuro)psychology told us about the mind (so far)? A reply to Coltheart. Cortex, 42, 387-392.
Hodges, J. R. (1994). Cognitive assessment for clinicians. Oxford University Press. Holloway, R. L. (2002). Brief communication: how much larger is the relative volume of area 10 of
the prefrontal cortex i n humans? American Journal of Physical Anthropology, 118, 399-401.

90 MIND AND THE FRONTAL LOBES
Janowsky, J, S., Shimamura, A. P., & Squire, L. R. (1989). Source memory impairments i n patients w i t h frontal lobe lesions. Neuropsychologia, 27,1043-1056.
John, J. P., Burgess, P. W., Yashavantha, B. S., Shakeel, M . K., Halahaffi, H. N., & Jain, S. (2009). Differential relationship of frontal pole and whole brain volumetric measures wi th age in neu-roleptic-naive schizophrenia and healthy subjects. Schizophrenia Research, 109(1-3), 148-158.
Johnson, S. C, Baxter, L. C, Wilder, L. S., Pipe, J. G.,Heiserman, J. E., 8c Prigatano, G. P. (2002). Neural correlates of self-reflection. Brain, 125,1808-1814.
Jung, R. E., 8c Haier, R. J. (2007). The Parieto-Frontal Integration Theory (P-FIT) of intelligence: converging neuroimaging evidence. Behavioral & Brain Sciences, 30(2), 35-54.
Karim, A. A., Schneider, M . , Lotze, M . , Veit, R., Sauseng, P., Braun, C, 8c Birbaumer, N . (2010). The t r u t h about lying: inhibit ion of the anterior prefrontal cortex improves deceptive behavior. Cerebral Cortex, 20(1), 205-213.
RTiegel, M . , Phillips, L. H. , Lemke, U., 8c Kopp, U. A. (2005). Planning and reaHsation of complex intentions i n patients w i t h Parkinson's disease. Journal of Neurology, Neurosurgery & Psychiatry, 76(11), 1501-1505.
Knight, C, Alderman, N., 8c Burgess, P. W. (2002). Development of a simplified version of the m u l tiple errands test for use i n hospital settings. Neuropsychological Rehabilitation, 12, 231-255.
Koechlin, E., Basso, G., Pietrmi, P., Panzer, S., 8c Grafman, J. (1999). The role of the anterior prefrontal cortex i n human cognition. Nature, 399,148-151.
Koechlin, E., Ody, C, 8c Kouneiher, F. (2003). The architecture of cognitive control i n the human prefrontal cortex. Science, 302(5648), 1181-1185.
Kopelman, M . D., Stanhope, N., 8c Kingsley, D. (1997). Memory for temporal and spatial context i n patients w i t h focal diencephalic, temporal lobe and frontal lobe lesions. Neuropsychologia, 35,1533-1545.
Levine, B., Stuss, D. T., Milberg, W. P., Alexander, M . P., Schwartz, M . , 8c Macdonald, R. (1998). The effects of focal and diffuse brain damage on strategy application: evidence from focal lesions, traumatic brain injury, and normal aging. Journal of the International Neuropsychological Society, 4, 247-264.
Luria, A . R. (1966). Higher cortical function in man. London: Tavistock. MacLeod, A. K., Buckner, R. L., Miezin, F. M . , Petersen, S. E„ & Raichle, M . E. (1998). Right anterior
prefrontal cortex activation during semantic monitoring and working memory. Neuroimage, 7,41-48.
Manly, T., Hawkins, K., Evans, J., Woldt, K., 8c Robertson, I . H , (2002). Rehabilitation of executive function: a facilitation of effective goal management on complex tasks using periodic auditory alerts. Neuropsychologia, 40, 2671-2681.
Milner, B., Petrides, M . , 8c Smith, M . L. (1985). Frontal lobes and the temporal organization of memory. Human Neurobiology, 4,137-142.
Penfield, W., 8c Evans, J. (1935). The frontal lobe i n man: a clinical study of maximum removals. Brain, 58,115-133.
Petrides, M . , 8c Pandya, D. M . (1994). Comparative architectonic analysis of the human and macaque frontal cortex. I n : F. Boiler 8c J. Grafman (Eds.), Handbook of neuropsychology (vol. 9, pp. 17-57). Amsterdam: Elsevier.
Petrie, A . (1952). Personality and the frontal lobes. London, UK: Routledge 8c Kegan Paul. Picton, T. W , Stuss, D. T., Alexander, M . P., Shallice, T., Binns, M . A. 8c Gillingham, S. (2006). Effects
of focal frontal lesions on response inhibit ion. Cerebral Cortex, 17, 826-838. Picton, T. W., Stuss, D. T., Shallice, T., Alexander, M . P., 8c Gillingham, S. (2006). Keeping time:
effects of focal frontal lesions. Neuropsychologia, 44,1195-1209. Poldrack, R. A., 8c Wagner, A. D. (2004). What can neuroimaging tell us about the mind? Insights
from prefrontal cortex. Current Directions in Psychological Science 13(5), 177-181. Ramnani, N. , 8c Owen, A. M . (2004). Anterior prefrontal cortex: Insights into function from anat
omy and neuroimaging. Nature Reviews Neuroscience, 5,184-194. Reynolds, J. R., West, R., 8c Braver, T. (2009). Distinct neural circuits support transient and
sustained processes i n prospective memory and working memory. Cerebral Cortex, 19(5), 1208-1221.

Rostral Prefrontal Cortex 91
Roca, M . , Parr, A., Thompson, R., Woolgar, A., Torralva, T., Antoun, N., Manes, E, & Duncan J. (2010). Executive function and fluid intelligence after frontal lobe lesions. Brain, 133(1), 234-247.
Rugg, M . D., Fletcher, P. C, Frith, C. D., Frackowiak, R. S. J., & Dolan, R. J.(1996). Differential activation of the prefrontal cortex i n successful and unsuccessful memory retrieval. Brain, 119,2073-2084.
Semendeferi, K., Armstrong, E., Schleicher, A., Zffles, K., 8c Van Hoesen, G. W. (2001). Prefrontal cortex i n humans and apes: a comparative study of area 10. American Journal of Physical Anthropology, 114, 224-241.
Shallice, T., 8c Burgess, P. W. (1991). Deficits i n strategy application following frontal lobe damage in man. Brain, 114, 727-741.
Shallice, T., Stuss, D. T., Alexander, M . P., Picton, T. W., 8c Derkzen, D. (2008). The multiple dimensions of sustained attention. Cortex, 44, 794-805.
Shallice, T., Stuss, D. T., Picton, T. W., Alexander, M . P., 8c Gillingham, S. (2008). Multiple effects of prefrontal lesions on task-switching. Frontier in Human Neuroscience, 1,1-11.
Shammi, P., 8c Stuss, D. T. (1999). Humour appreciation: a role of the right frontal lobe. Brain, 122, 657-666.
Shimamura, A. P., Janowsky, J. S., 8c Squire, L. R. (1990). Memory for the temporal order of events i n patients wi th frontal lobe lesions and amnesic patients. Neuropsychology, 28, 803-813.
Simons, J. S., Gilbert, S. J., Owen, A. M . , Fletcher, P. C, 8c Burgess, P. W. (2005). Distinct roles for lateral and medial anterior prefrontal cortex in contextual recollection. Journal of Neurophysiology, 94, 813-820.
Spearman, C. (1904). "General intelligence" objectively determined and measured. American Journal of Psychology, 15,201-293.
Spreen, G., 8c Strauss, E. (1988). A compendium of neuropsychological tests: administration, norms and commentary. New York: Oxford University Press.
Stone, V. E., Baron-Cohen, S., 8c Knight, R. T. (1998). Frontal lobe contributions to theory of mind. Journal of Cognitive Neuroscience, 10, 640-656.
Stuss, D. T., Alexander, M . P., Hamer, L., Palumbo, C, Dempster, R., Binns, M . , Levine, B., & Izukawa, D. (1998). The effects of focal anterior and posterior brain lesions on verbal fluency. Journal of International Neuropsychological Society, 4, 265-278.
Stuss, D. T., Alexander, M . P., Shallice, T., Picton, T. W., Binns, M . A., Macdonald, R., Borowiec, A., 8c Katz, D. I . (2005). Multiple frontal systems controlling response speed. Neuropsychologia, 43, 396-417.
Stuss, D. T., Binns, M . A., Murphy, K. J., & Alexander, M . P. (2002). Dissociations wi th in the anterior attentional system: effects of task complexity and irrelevant information on reaction time speed and accuracy. Neuropsychology, 16, 500-513.
Stuss, D. T., Floden, D., Alexander, M . P., Levine, B., 8c Katz, D. (2001). Stroop performance i n focal lesion patients: dissociation of processes and frontal lobe lesion location. Neuropsychologia, 39, 771-786.
Stuss, D. T., Gallup, G. G. Jr., 8c Alexander, M . P. (2001) The frontal lobes are necessary for "theory of mind." Brain, 124(Pt 2), 279-286.
Stuss, D. T., Levine, B., Alexander, M . P., Hong, J., Palumbo, C, Hamer, L., Murphy, K. J., 8c Izukawa, D. (2000). Wisconsin Card Sorting Test performance i n patients w i t h focal frontal and posterior brain damage: effects of lesion location and test structure on separable cognitive processes. Neuropsychologia, 38, 388-402.
Thomson, G. H . (1939). The factorial analysis of human ability. London, UK: University of London Press.
Tulving, E., Markowitsch, H . J., Criak, F. I . M . , Habib, R., 8c Houle, S. (1996). Novelty and familiarity activations i n PET studies of memory encoding and retrieval. Cerebral Cortex, 6, 71-79.
Turner, M . S., Simons, J. S., Gilbert, S. J., Frith, C. D„ 8c Burgess, P. W. (2008). Distinct roles for lateral and medial rostral prefrontal cortex i n source monitoring of perceived and imagined events. Neuropsychologia, 46,1442-1453.

92 MIND AND THE FRONTAL LOBES
Uretzky, S., & Gilboa, A. (2010). Knowing your lines but missing your cue: rostral prefrontal lesions impair prospective memory cue detection, but not action-intention superiority. Journal of Cognitive Neuroscience, 22(12), 2745-2757.
VoEe, E., Gilbert, S. I , Benoit, R. G., & Burgess, P. W. (2010). Specialization of the rostral prefrontal cortex for distinct analogy processes. Cerebral Cortex, 20(11), 2647-2659.
Voile E, Gonen-Yaacovi G, de Lacy Costello A, Gilbert SJ, Burgess PW. (2011). The role of rostral prefrontal cortex i n prospective memory: A voxel-based lesion study. Neuropsychologia. Mar 1. [Epub ahead of print]
Warrington, E. K., James, M . , ScMaciejewski, C. (1986). The WAIS as alateralizingandlocalizing diagnostic instrument: a study of 656 patients wi th unilateral cerebral lesions. Neuropsychologia, 24(2), 223-239.
Wechsler, D. (1939). The measurement of adult intelligence. Baltimore: Williams & Wilkins. Wheeler, M . A., Stuss, D. T., & Tulving, E. (1997). Towards a theory of episodic memory: the frontal
lobes and autonoetic consciousness. Psychological Bulletin, 121(3), 331-354. Wilson, B. A., Alderman, N., Burgess, P. W , Emslie, H. , 8: Evans, J. (1996). Behavioural assessment of
the dysexecutive syndrome. Bury St. Edmunds, U.K.: Thames Valley Test Company.

• 1 condition • 2 conditions • 3 conditions 11 conditions
• 12-19 conditions 20 conditions or above
Figure 4.1. A v isual i l l u s t r a t i o n of the findings f r o m a selection of Stuss and colleagues' studies invo lv ing pat ients w i t h f r o n t a l lobe lesions, between 1998 and 2008. Test results are compared to pat ients w i t h n o n - f r o n t a l lesions and /o r controls . The figure shows the frequency of a reported i m p a i r m e n t for each B r o d m a n n area based on the results f r o m 44 behavioral variables described i n Table 4 .1 . These frequencies are d iv ided i n t o six ranges, represented by di f ferent colors. Dark blue = the region was f ound to be impaired , to a statist ical ly s ignif icant degree, on 1 o f the 44 behavioral variables (relative to either controls or at least one other pat ient group) . L ight blue = 2 /44 variables impaired . Green = 3/44 variables impaired . Yellow = 4 to 1 1 variables impaired . Orange = 12 to 19 variables impaired . Red = 20 or more variables impaired. The results show t h a t rostral PFC regions (area 10) are not impaired on a larger number of these behavioral indicators than other PFC regions. These results provide a constra int for theor i z ing about the funct ions o f rostral PFC.




















