
The Monkeying of the Americas: Primate Biogeography in the ...

We studied the relationship between vascularization and neuronalactivity in the visual cortex during postnatal development in theprimate. Analyses were focused on layer IVC that displays asequential pattern of maturation for the magno- and parvocellularsystems in separate sublayers, respectively IVCalpha and IVCbeta.Cytochrome oxidase and endogeneous alkaline phosphatase histo-chemistry was used to analyse, on the same sections, the laminarpatterns of cortical activity and vessel density in the primary visualcortex of the marmoset (Callithrix jacchus). Experiments werecarried out in five young and two adult animals. We showed that thetemporal pattern of angiogenesis differs in layer IVCalpha andIVCbeta. During the first postnatal month, vessel density is higher inIVCalpha than in IVCbeta and runs parallel to cytochrome oxidaseintensity. In 2-month-old animals, both vessel densities and cyto-chrome oxidase activity are similar in IVCalpha and IVCbeta. Inadults, the vessel densities in IVCalpha and IVCbeta are the reverseof those observed during the first postnatal month. Vessel diameterdoes not account for this evolution in vascular patterns. In thediscussion, we suggest that such a developmental time-course ofangiogenesis might be linked to the synaptogenesis requirementsthat proceed differently for the magno- and parvocellular systems inthe primate striate cortex.
IntroductionThe existence of cortical modules, both neuronal and vascular,has been demonstrated in the supragranular layers of theprimary visual cortex (blobs) (Zheng et al., 1991) and in thesomatosensory cortex (barrels) (Woosley et al., 1996). However,the mechanisms by which these modules develop have not beenaddressed in primates. If the anatomical and physiologicalproperties of the primary visual cortex have been largelyinvestigated in several primate species, only a few works studiedits vascular architecture (Bell and Ball, 1985, 1990; Zheng et al.,1991). Generally speaking, brain vascular architecture, thatdetermines cerebral blood f low and oxygen utilization by thecerebral tissue, is not extensively analysed. Relationshipsbetween blood oxygenation level and neuronal activity arethoroughly investigated (Vanzetta and Grinvald, 1999; Logothetiset al., 1999, 2001) since numerous investigators associateneuronal functional activation with tissue metabolic require-ments and temporary modifications in the regional blood f lowto study the neuronal basis of cognitive functions in adults.During development, hemodynamic changes have moreover tobe considered in a changing space, since brain development stilloccurs in childhood and adolescence.
To examine the development of cortical modules, we adoptthe hypothesis that neural activity drives the maturation ofmicrovascular circuitry in the developing brain. During thepostnatal period, brain metabolism is energy-demanding toinsure synaptogenesis and neural maturation. The formation of agiven vascular pattern is likely to be controlled by intrinsicmechanisms, neuronal activity and sensory experience. As a
direct consequence, alterations in neuronal inputs may preventthe ‘normal’ vascular pattern from establishing. Sensorydeprivation would thus disturb angiogenesis processes and, ifirreversible, retardation in angiogenesis might become severeenough to impair neural plasticity. These mechanisms inpathological brain development are rarely considered. In rats,complex environments produce an increase in capillary densityin the visual cortex (Sirevaag et al., 1988). Visual deprivationprovokes modification of the pial angioarchitecture of theparieto-occipital cortex (Wolff et al., 1992) and delaysmaturation of the vascular support (Argandoña and Lafuente,1996). Similar studies have not, to our knowledge, been carriedout in the primates and the first step to be considered is toestablish the correlation between neural maturation andangiogenesis in the developing brain in control monkeys.
In the present paper, we examine the correlation betweenvascular organization, analysed by alkaline phosphatase revela-tion, and cortical metabolism, revealed by cytochrome oxidaseactivity, in the developing primary visual cortex in the primateCallithrix jacchus. Mitochondrial cytochrome oxidase enzymeactivity reveals laminar and modular neuronal activation patternsin the cerebral cortex (Carroll and Wong Riley, 1984; WongRiley, 1989). Its levels adjust to changes in regional neuronalactivity (Wong Riley, 1979; Wong Riley and Welt, 1980). Linksbetween brain oxidative metabolism, assessed by cytochromeoxidase activity and vessel density, have been established indifferent grey matter structures in the adult rat brain. In thehippocampus and the olfactory cortex there is a clear corres-pondence between vessel density, glucose utilization and cyto-chrome oxidase activity (Borowsky and Collins, 1989). In thesomatic sensory cortex, differential distribution of microvesselsbetween barrels and the surrounding cortex is correlated withthe distribution of mitochondrial enzymes (such as cytochromeoxidase) and with electrical and metabolic neuronal activities(Riddle et al., 1993). In the rabbit parietal cortex, cytochromeoxidase activity and capillary density are not correlated in veryyoung animals, the cytochrome oxidase pattern being moremature compared with that of vessel density, but are closelylinked in older rabbits (Tuor et al., 1994).
A possible correlation between neural maturation and angio-genesis in the developing brain needs further investigation in theprimate that presents specific features of cortical development,compared to rodents (Kennedy and Dehay, 1993). Developmentof primate cortex is unique in its early cortical specification, inboth areal and laminar definition. Thalamic afferents exert anearly inf luence on the prenatal developing cortex in monkeys,especially on the development of connections. In the rodentcortex, modular features are formed postnatally, whereas thecortex of newborn primates already presents adult-like modules.We focus our study on layer IVC of the primary visual cortexbecause it represents an adequate model for analysing the
Vascularization in the Primate VisualCortex during Development
Caroline Fonta and Michel Imbert
Centre de Recherche Cerveau et Cognition, UMR 5549 CentreNational de la Recherche Scientifique/Université Paul Sabatier,Faculté de Médecine Rangueil, 133 route de Narbonne, 31062Toulouse cedex, France
Cerebral Cortex Feb 2002;12:199–211; 1047–3211/02/$4.00© Oxford University Press 2002. All rights reserved.
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

development of a vascular pattern in relation to neural matura-tion. Layer IVC represents the main target of the visual thalamo-cortical axons. It is generally described as composed of twodistinct layers, layer IVCalpha and layer IVCbeta, that are therespective targets of the magnocellular and the parvocellularsystems in the striate cortex (Hubel and Wiesel, 1972; Blasdeland Lund, 1983; Freund et al., 1989). Thalamic afferents aresegregated and organized into an adult-like pattern in layerIVCalpha and IVCbeta in newborn macaques (Rakic, 1977;Horton and Hocking, 1996) and very young marmosets(Chappert et al., 2001). Both layers display similarly highcytochrome oxidase staining in adults, but not in juveniles. Inyoung primates, layer IVCalpha shows a dense cytochromeoxidase staining, while layer IVCbeta is only a lightly stainedzone (Horton, 1984; Kennedy et al., 1985; Hendrickson et al.,1991; Spatz et al., 1993, 1994; Horton and Hocking, 1996; Fontaet al., 1997). It is proposed that this developmental pattern ofcytochrome oxidase in layer IVC corresponds to the differentialmaturation time-course between the magno- and parvocellularpathways. The hypothesis that vascularization of the primaryvisual cortex is correlated with the differential maturation of thetwo systems appears pertinent, but had not yet been tested.
The present work analyses laminar vessel density in theprimary visual cortex of the New World monkey Callithrix
jacchus, during early postnatal life and in the adult. Analysisfocuses on the two sublayers of layer IVC that display asequential maturation and a high sensitivity to visual deprivationin the first two postnatal months (Spatz et al., 1993; Fonta et al.,1997, 2000). The anatomical identification of a vascular territorycan be made by various techniques, some being direct, such astransillumination (Zheng et al., 1991), but not always adequateto detect in parallel other markers by immunohistochemistryor histochemistry (e.g. intravascular contrasting agents). Weobtained good quality resolution of vessel distribution byhistochemical detection of an endogenous endothelial enzyme,alkaline phosphatase. This method had been previously used tovisualize adult brain vascular architecture in humans andmacaques (Friede, 1966; Bell and Scarrow, 1984; Bell and Ball,1985, 1990). Cytochrome oxidase activity was used to visualize,on the same sections, the relative levels of laminar corticalactivity. The results show that angiogenesis occurs in parallel toneural maturation in layer IVC.
Material and MethodsThe animals were born in a marmoset rearing facility maintained at theCentre de Recherche Cerveau et Cognition, Toulouse, France. Sevenmarmosets were used: two adults, one 7 years old (98.4) and one 11 yearsold (97.7); two animals 60 postnatal days old (PND) (99.9 and 00.5); oneanimal 24 PND (97.12); one 17 PND animal (97.9); and one neonate, a fewhours old (00.3).
Animals were transcardially perfused under deep pentobarbitalanesthesia (50 mg/kg) with a rinse of NaCl 0.9% with heparin followed by4% paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4. The brainwas then postfixed in 4% paraformaldehyde for 2 h and cryoprotectedovernight in 30% sucrose in 0.1 M PB. One animal (97.9) was perfusedwith 10% formol.
Cortical blocks were sectioned at a thickness of 30 µm with a cryostat.Experiments were carried out on free-f loating frontal sections.
All animal experiments were carried out in accordance with TheNational Institute of Health guidelines.
Histochemistry
Double staining was carried out on sections devoted to quantitativeanalyses. The sections were stained first for cytochrome oxidase, then foralkaline phosphatase. In these double-stained sections, the cytochromeoxidase brown background was used to distinguish borders of layers.Vessels, revealed by endothelial alkaline phosphatase, were easily
identified as dark blue or black areas. Single staining (cytochrome oxidaseor alkaline phosphatase) was performed on the other sections.
For cytochrome oxidase histochemistry, sections were incubatedovernight in a solution containing 50 mg 3–3′-diaminobenzidine tetra-chloride (DAB), 35 mg cytochrome C, 20 mg catalase and 4 g D(+) sucroseper 100 ml PB (0.1 M, pH 7.4) at room temperature in the dark withagitation. Sections were then rinsed in several PB baths before staining ofendothelial cells.
In some cases (if sections were not treated additionally for alkalinephosphatase staining), cytochrome oxidase reaction was preceded by anincubation in a cobalt chloride Tris solution.
Endogenous alkaline phosphatase reactions were performed byincubating sections in a 100 mM Tris–HCl pH 9.5 solution containing100 mM NaCl, 50 mM MgCl2, 0.53 mM 4-nitro blue tetrazolium chloride(NBT) and 0.38 mM 5-bromo-4-chloro-3-indolyl-phosphate 4-toluidine salt(BCIP). The reaction was carried out for 10 min at room temperature. Thereaction was stopped by transferring sections to a 10 mM Tris pH 7.5solution containing 1 mM EDTA and 10 mM levamisole. Sections werethen rinsed in Tris and mounted.
Quantitative Analyses of Vessel Density
Mounted sections were examined and selected slides were digitized witha Leica microscope equipped with a 3 CCD camera (Sony DXC930P),interfaced with an IBM-compatible computer, Imascan Board (Imagraph)and Optimas software (Media Cybernetics). Morphometric analyses wereperformed with Optimas image analysis software. Images were firstconverted into grey values.
Measurements were made on the sampled images in 1 mm widestripes extending from layer I to white matter on coronal sections. Twosets of measurements were performed. One set was made in the upperand lower banks of the calcarine sulcus, representing the peripheral visualfield (Fritsches and Rosa, 1996). Ten or eleven stripes were analysed intwo or three sections from the same hemisphere for each animal. Thesecond set concerned the dorsal and dorso-lateral surfaces of the occipitallobe, corresponding to part of area 17 representing the central visual field(Fritsches and Rosa, 1996). Ten stripes were defined in two sections fromthe same hemisphere for each animal studied. Ventral and dorsal partswere equally represented in each set of measures.
The same side was investigated for age-matched animals to avoidpossible effects of left/right asymmetry, even if cerebral dominance isgenerally not significant in adult macaques as far as density of neurons andcortical morphometric features are concerned. In any case, there seems tobe no hemispheric specialization (O’Kusky and Colonnier, 1982b;Gottlieb et al., 1985; Suner and Rakic, 1996).
Inside each 1 mm wide stripe, quantification of vascular parameterswas performed within each layer or sublayer, defined by its cytochromeoxidase staining. The numbers assigned to the layers conform to thenomenclature used by Brodmann.
For all the animals, common areas of interest were as follows.
1. The supragranular layers corresponding to layers II and III (a depth of200–400 µm below layer I was investigated).
2. Sublayers IVCalpha and IVCbeta — in the youngest animals (firstpostnatal month, i.e. animals 00.3, 97.9 and 97.12), densecytochrome oxidase staining was restricted to layer IVCalpha. In2-month-old animals (99.9 and 00.5), layer IVC displayed anhomogeneous staining; however, a thin pale band separated theouter (IVCalpha) and inner (layer IVCbeta) parts of layer IVC. Thisband was more obvious with alkaline phosphatase staining.Therefore, the two sublayers of IVC could be identified, at highmagnification, in juveniles. In the adults (97.7 and 98.4), the innerpart (layer IVCbeta) showed a stronger cytochrome oxidase signalthan the outer part of layer IVC. However, the boundary betweenthe two sublayers was blurred. The innermost part of layer IVC(100–200 µm) was considered as layer IVCbeta. The 100–200 µmabove, presenting a weaker cytochrome oxidase staining, wereattributed to layer IVCalpha.
3. Layer V.4. Layer VI — in 2-month-old animals, cytochrome oxidase staining
enabled the two sublayers of layer VI to be clearly distinguished(with an outer sublayer darker than the inner sublayer).
200 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

When defining the individual layers, the extreme borders werediscarded from the selection, as was the intermediate zone betweenIVCalpha and IVCbeta, that appears lightly labelled in cytochromeoxidase and more obviously in alkaline phosphatase staining, in 2-month-
old animals. In fact, neurons located in this transition zone presentconnections different from those of neurons in layers IVCalpha andIVCbeta and they may, therefore, represent a separate entity (Boyd et al.,2000).
Figure 1. Double staining [cytochrome oxidase (CO) in brown; alkaline phosphatase (AP) vessels in blue] in the primary visual cortex of a newborn marmoset (coronal section). LayerIVCalpha and layer Va are more CO reactive than layer IVCbeta. AP strongly stains the white matter (wm). (B) Magnified part of the ventral calcarine cortex in (A); cs, calcarine sulcus.
Cerebral Cortex Feb 2002, V 12 N 2 201
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

Two parameters were quantified: (i) vascular density defined as thearea occupied by vessel profiles per unit area of cortex and (ii) vesseldiameter.
Vascular density was analysed at a final magnification of 180× by anautomatic sequence of commands created to identify and measure all field
areas with grey levels within a range of grey values indicative of vesselstaining. This range was manually adjusted for each analysis becausestaining intensities might provide different grey ranges from one sectionto another. Thus, a semi-automatic procedure separated stained areasfrom background. The selection was visually checked and corrections
Figure 2. Cytochrome oxidase (CO, in A) and alkaline phosphatase (AP,in B) staining in the 24 PND animal. CO activity is stronger in layer IVCalpha than in layer IVCbeta. Apart fromstaining endothelial cells, AP produces a blue background in layer IVC and in the white matter (wm). In both stainings, layer IVCalpha is more labeled than layer IVCbeta.
202 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

made to discard selection of artefacts or to achieve a better selection. Forthis purpose, a higher magnification could be obtained using a zoomfunction. The area of interest (layer or sublayer) was delimited. Finally,the individual values of the selected areas and the value of the area ofinterest were extracted and stored in a database (Excel). The total vesselarea was expressed as a percentage of the reference area (area of interest).
Data were analysed using non parametric tests (Wilcoxon test forpaired samples).
Vessel diameter was defined at a final magnification of 360×.Measurements were performed in the same area of interest used forvascular density analyses in layer IVC. The diameter was measuredperpendicular to the long axis of the vessel profile. The same vessel couldprovide several measures: (i) if it could be traced without branching andif its diameter varied along this distance, two measurements of diameterwere made, or (ii) if it branched, measurements were made between twosuccessive branching points. Diameter measures were sorted into classesof 1 µm increment. Diameter distributions were compared by means of anon-parametric test (Kolmogorov–Smirnov test).
Values in the tables are indicated as mean and standard error of themean (SEM) for each parameter. In statistical analyses, made with SPSS 9.0software, P < 0.05 was regarded as significant.
Results
Metabolic and Vascular Stainings
Cytochrome oxidase staining presents the characteristic laminarpatterns previously described in marmosets (Spatz et al., 1993,1994; Fonta et al., 1997)
In the very young animals (during the first postnatal weeks),dense cytochrome oxidase staining was restricted to the outerpart of layer IVC, layer IVCalpha. A narrow band of densestaining was observed at the inner part of layer IVC, named Va(Spatz et al., 1993) (Figs 1, 2A). In 2-month-old marmosets, layersIVCalpha and IVCbeta were heavily stained (Fig. 3). Layer VI wasalso labeled, more strongly in its upper part. Layer V was faintlystained. Layers II and III were moderately labeled. In adults thelower part of IVC appears darker (Fig. 4).
Endothelial alkaline phosphatase clearly outlines vessels in allthe animals studied. But in juveniles, a diffuse background couldalso be observed in layer IVC (Fig. 5). This staining provided astrong labeling of white matter (Fig. 1A) and layer IVCalpha inthe youngest animals. Layer IVCbeta presented a lighter back-ground staining (Fig. 2B). In the 2-month-old marmosets, astrong background was seen in layer IVC and, to a lesser extent,in layers IVA and VI (Fig. 3B). White matter was devoid of anybackground staining in 2-month-old animals and in adults (Figs3B, 4). The background staining was weaker over the innermostpart of layer IVCbeta in adult preparations (Fig. 6A,B).
Thus, in young animals, the strong cytochrome oxidasestaining and the high background level of alkaline phosphatasestaining in layer IVC matched (Fig. 3). However, a transitionband with pale staining in the middle part of layer IVC wasclearly apparent with alkaline phosphatase detection and notwith cytochrome oxidase staining in 2-month-old marmosets(Fig. 3).
Vessel Density Analyses
We first concentrated the analyses on the calcarine visual cortexthat had been investigated for quantitative analyses of the areal,cellular and synaptic components during postnatal developmentin marmosets (Missler et al., 1993a,b). We then analysed vasculardensity in the dorsolateral part to compare the development ofvascularization in the peripheral-field and central-field striatecortex.
The mean vascular density increases as a function of age(∼ 10% of the cortical surface in the neonate and ∼ 22.5% in the
adult). We observed differences between animals of a same age(Table 1). Individual specificities or experimental factors (suchas depth of anesthesia, level of anoxia, inadequate pressureduring perfusion resulting in collapsed blood vessels) mayexplain such differences. However, the relative vasculardensities in the different cortical layers are reproduciblebetween age-matched animals (e.g. 2-month-old marmosets 99.9and 00.5; Fig. 7).
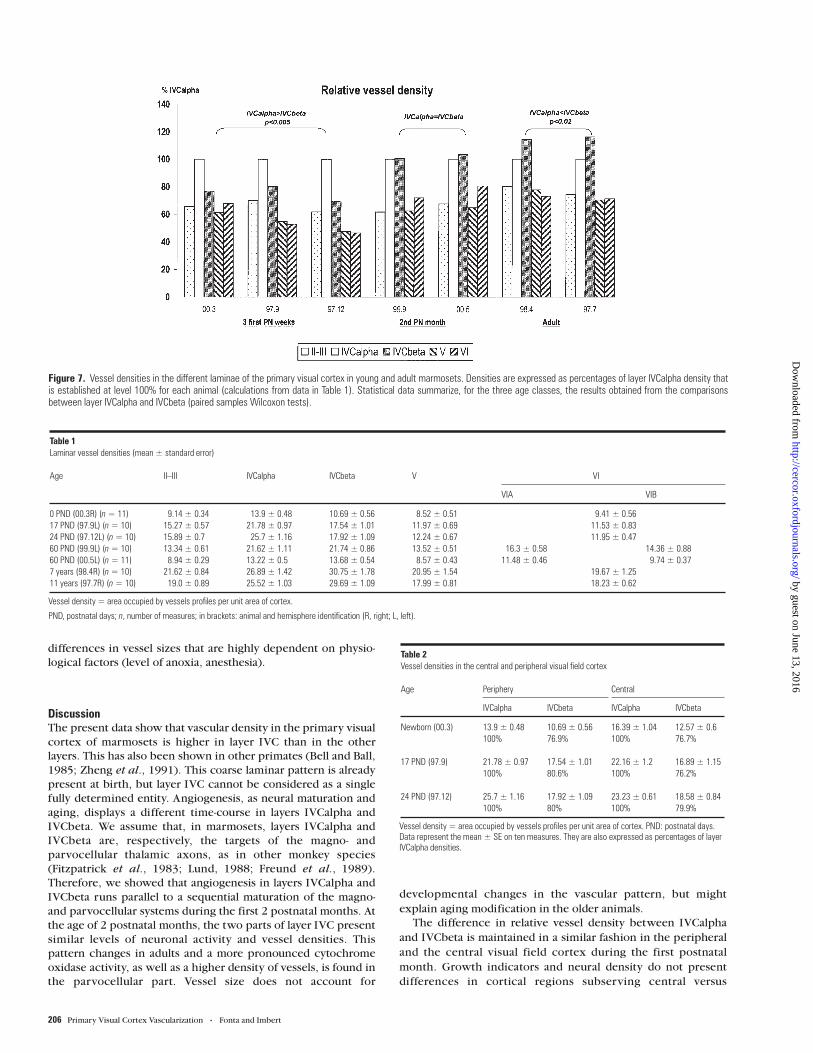
Figure 7 details the relative percentages of vessel densitycompared to layer IVCalpha, for each animal. Data werenormalized by expressing values as percentages of layerIVCalpha vessel density. For each animal, statistical analyseswere performed on raw data (i.e. vessel density) to comparevessel densities between two different laminae.
Whatever the age of the animal, vessel density is always higherin the two sublayers of layer IVC than in the infra- (P < 0.03) andsupragranular (P < 0.033) layers. Layer IVCbeta displays aspecific evolution relative to layer IVCalpha. In the newbornmarmoset (003) and in the 3-week-old marmosets (97.9 and97.12), vessel density is higher in layer IVCalpha than in layerIVCbeta (P ≤ 0.005). There is a 20–30% difference between
Figure 3. Cytochrome oxidase (CO, in A) and alkaline phosphatase (AP, in B) staining ofserial sections in marmoset 99.9 (60 PND). Arrowheads and asterisks indicate commonvessels. The high CO activity and the strong AP background staining clearly match inlayer IVC. At this age, AP does not stain the white matter (wm).
Cerebral Cortex Feb 2002, V 12 N 2 203
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

IVCalpha and IVCbeta vessel densities. In 2-month-old animals(99.9 and 00.5), vessel densities in IVCalpha and IVCbeta do notpresent any significant difference. In adults, vessel density issignificantly higher in layer IVCbeta than in IVCalpha [P = 0.02
(98.4) and P = 0.005 (97.7)]. Vessel density is 15% higher inIVCbeta. The densities of vessels in the two sublayers of layerIVC appear, then, to follow the intensity of cytochrome oxidasestaining (cf. Figs 1–4).
Figure 4. Double staining [cytochrome oxidase (CO) in brown; alkaline phosphatase (AP) vessels in blue] in the primary visual cortex of an adult marmoset. (B) Magnified part of thedorsal calcarine cortex in (A). CO activity is stronger in the lower tier of layer IVC; wm, white matter.
204 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

Vessel densities in layers VI and V are similar, except in the2-month-old animals (Table 1). In these animals, layer VI can bedivided into two parts that differ in their cytochrome oxidasestaining. The outer part, layer VIA, with a stronger staining,presents a significantly higher vascular density than the innerpart, layer VIB [P = 0.037 (9.99) and P = 0.026 (0.05)]. Vasculardensities in layers VIB and V are similar.
At birth (marmoset 003), the vascular density in layers II andIII (9.1%) is not significantly different from those in layers V andVI (8.5 and 9.4%, respectively; Table 1). In 3-week-old juveniles,vascular density is higher in the supragranular layers than in theinfragranular layers (P ≤ 0.007). In 2-month-old animals, layersII/III and V show similar vascular densities that are significantlylower than density in layer VI (0.006 ≤ P ≤ 0.037). In adults,vascular densities, although higher in supragranular layers, arenot significantly different from densities in infragranular layers.
The difference in vascular density between IVCalpha andIVCbeta is also found in the dorsal external part of the occipitalpole (Table 2). Vascular density is significantly higher in layerIVCalpha than in IVCbeta in the neonate (P = 0.007), as in the3-week-old animals (P = 0.005). The relative vessel density oflayer IVCbeta compared to IVCalpha is not different in thecalcarine cortex and in the dorsolateral cortex, with a 19–23%difference between the two layers.
Cytochrome oxidase activity also showed a partition of layerIVC in the dorsolateral visual cortex, with dark staining only inIVCalpha (not shown).
Vessel Diameter Analyses
Considering that the two sublayers of IVC present the samegrowth rate during postnatal development (Missler et al.,1993a), the difference in evolution of vessel densities inIVCalpha and IVCbeta could be due to a difference in thenumber and/or in the size of vessels. We measured the vesseldiameters in these two sublayers in the calcarine cortex.Measurements were made in all seven marmosets (Table 3).
The distributions of vessel diameters, sorted into classes of1 µm, appear similar in layers IVCalpha and IVCbeta (Fig. 8).They were not significantly different except in one adult — theoldest marmoset (97.7), where vessels in layer IVCbeta wereslightly larger than those in IVCalpha (P = 0.004).
Differences between age-matched animals (e.g. 00.5 and99.9) clearly appear. Vessel diameter and density were bothlower in marmoset 00.5 than in marmoset 99.9. It can besuggested that the differences in vessel density observedbetween the two animals (cf. Table 1) were supported by
Figure 5. Alkaline phosphatase staining in the primary visual cortex of a 60 PND-oldmarmoset. The enzyme is revealed in the vessel walls and in the surrounding corticaltissue in layer IVC. A narrow band is devoided of strong background staining betweenlayer IVCalpha and IVCbeta.
Figure 6. Alkaline phosphatase staining in the primary visual cortex of an adult animal.Background staining is absent in the white matter (wm) and appears lightly in IVCbeta.In (A), asterisks indicate the position of the area enlarged in (B), with a 90° clockwiserotation.
Cerebral Cortex Feb 2002, V 12 N 2 205
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from
differences in vessel sizes that are highly dependent on physio-
logical factors (level of anoxia, anesthesia).
DiscussionThe present data show that vascular density in the primary visual
cortex of marmosets is higher in layer IVC than in the other
layers. This has also been shown in other primates (Bell and Ball,
1985; Zheng et al., 1991). This coarse laminar pattern is already
present at birth, but layer IVC cannot be considered as a single
fully determined entity. Angiogenesis, as neural maturation and
aging, displays a different time-course in layers IVCalpha and
IVCbeta. We assume that, in marmosets, layers IVCalpha and
IVCbeta are, respectively, the targets of the magno- and
parvocellular thalamic axons, as in other monkey species
(Fitzpatrick et al., 1983; Lund, 1988; Freund et al., 1989).
Therefore, we showed that angiogenesis in layers IVCalpha and
IVCbeta runs parallel to a sequential maturation of the magno-
and parvocellular systems during the first 2 postnatal months. At
the age of 2 postnatal months, the two parts of layer IVC present
similar levels of neuronal activity and vessel densities. This
pattern changes in adults and a more pronounced cytochrome
oxidase activity, as well as a higher density of vessels, is found in
the parvocellular part. Vessel size does not account for
developmental changes in the vascular pattern, but might
explain aging modification in the older animals.
The difference in relative vessel density between IVCalpha
and IVCbeta is maintained in a similar fashion in the peripheral
and the central visual field cortex during the first postnatal
month. Growth indicators and neural density do not present
differences in cortical regions subserving central versus
Figure 7. Vessel densities in the different laminae of the primary visual cortex in young and adult marmosets. Densities are expressed as percentages of layer IVCalpha density thatis established at level 100% for each animal (calculations from data in Table 1). Statistical data summarize, for the three age classes, the results obtained from the comparisonsbetween layer IVCalpha and IVCbeta (paired samples Wilcoxon tests).
Table 1Laminar vessel densities (mean ± standard error)
Age II–III IVCalpha IVCbeta V VI
VIA VIB
0 PND (00.3R) (n = 11) 9.14 ± 0.34 13.9 ± 0.48 10.69 ± 0.56 8.52 ± 0.51 9.41 ± 0.5617 PND (97.9L) (n = 10) 15.27 ± 0.57 21.78 ± 0.97 17.54 ± 1.01 11.97 ± 0.69 11.53 ± 0.8324 PND (97.12L) (n = 10) 15.89 ± 0.7 25.7 ± 1.16 17.92 ± 1.09 12.24 ± 0.67 11.95 ± 0.4760 PND (99.9L) (n = 10) 13.34 ± 0.61 21.62 ± 1.11 21.74 ± 0.86 13.52 ± 0.51 16.3 ± 0.58 14.36 ± 0.8860 PND (00.5L) (n = 11) 8.94 ± 0.29 13.22 ± 0.5 13.68 ± 0.54 8.57 ± 0.43 11.48 ± 0.46 9.74 ± 0.377 years (98.4R) (n = 10) 21.62 ± 0.84 26.89 ± 1.42 30.75 ± 1.78 20.95 ± 1.54 19.67 ± 1.2511 years (97.7R) (n = 10) 19.0 ± 0.89 25.52 ± 1.03 29.69 ± 1.09 17.99 ± 0.81 18.23 ± 0.62
Vessel density = area occupied by vessels profiles per unit area of cortex.
PND, postnatal days; n, number of measures; in brackets: animal and hemisphere identification (R, right; L, left).
Table 2Vessel densities in the central and peripheral visual field cortex
Age Periphery Central
IVCalpha IVCbeta IVCalpha IVCbeta
Newborn (00.3) 13.9 ± 0.48 10.69 ± 0.56 16.39 ± 1.04 12.57 ± 0.6100% 76.9% 100% 76.7%
17 PND (97.9) 21.78 ± 0.97 17.54 ± 1.01 22.16 ± 1.2 16.89 ± 1.15100% 80.6% 100% 76.2%
24 PND (97.12) 25.7 ± 1.16 17.92 ± 1.09 23.23 ± 0.61 18.58 ± 0.84100% 80% 100% 79.9%
Vessel density = area occupied by vessels profiles per unit area of cortex. PND: postnatal days.Data represent the mean ± SE on ten measures. They are also expressed as percentages of layerIVCalpha densities.
206 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

peripheral visual fields in macaques (Powell and Hendrickson,1981; Gottlieb et al., 1985). Maturation of both magno- andparvocellular retinogeniculate axon arbors occurs 1 week aheadfor central vision compared to peripheral vision in the galago(Lachica and Casagrande, 1988). As we sampled animals at agesspaced 1 month apart, a timing difference of a week cannot bedetected.
Cytochrome Oxidase Staining
The laminar pattern of cytochrome oxidase activity in theprimary visual cortex of young and adult marmosets, alreadydescribed (Spatz et al., 1993, 1994; Fonta et al., 1997),resembles that found in the macaque (Horton, 1984; Kennedy et
al., 1985; Hendrickson et al., 1991) and human (Wong-Riley et
al., 1993) developmental patterns. Dense cytochrome oxidasestaining is restricted to layer IVCalpha and to a thin layer at theboundary between layers IVCbeta and V in neonates. In adultanimals, layers IVCalpha and IVCbeta are both highly cyto-chrome oxidase reactive. We presently show that the evolutionbetween the newborn and the adult-like patterns occurs duringthe second postnatal month in marmosets. The adult pattern isreached from the fourth postnatal month in humans (Wong-Rileyet al., 1993) and from the second postnatal month in macaques(Hendrickson et al., 1991; Endo et al., 2000). In the adult, thehigher cytochrome oxidase reactivity of layer IVCbeta,compared to IVCalpha, observed in marmosets in the presentpaper and elsewhere (Spatz et al., 1994) has also been described
in macaques (Wong-Riley and Carroll, 1984) and humans(Wong-Riley et al., 1993).
We noticed that layer VI was more cytochrome oxidasereactive in its outer part than in its inner part in 2-month-oldmarmosets. This could be linked to the concentration of thalamo-cortical terminals in the upper part of layer VI for New Worldprimates (Hendrickson et al., 1978; Spatz, 1979; Fitzpatrick et
al., 1983). The matching of intracortical and cortico-thalamicconnections for neurons in the upper and lower parts of layer VIwith the parvo- and magnocellular system, respectively, isdocumented in macaques (Fitzpatrick et al., 1985; Wiser andCallaway, 1996). To our knowledge, partition of layer VI intosublayers associated with the magno- or the parvocellular systemhas not been described in New World monkeys.
Alkaline Phosphatase Staining
With the alkaline phosphatase method, apart from stainingendothelial cells, we obtained a lamina-specific backgroundstaining that is found mainly in juvenile preparations. Inter-estingly, this background staining is found in the layers that arethe targets of thalamic afferents (layers IV and VI) and in thewhite matter where axons from the thalamus (LGN) accumulatebefore birth. Moreover, in layer IVC, alkaline phosphatasebackground intensity parallels the evolution of cytochromeoxidase activity, being high first in IVCalpha, then in IVCalphaand beta. Several explanations can be proposed, as backgroundstaining can be due either to endothelial cells themselves, or toother cortical elements. Artefactual presence of alkaline phos-phatase outside vessels due to physical or chemical alterations ofthe vessels can be discarded, since background staining is notgeneral. For the same reasons, enzyme leakage in the vesselvicinity by immaturity and absence of blood–brain barrierproperties seems unlikely. We propose, instead, that alkalinephosphatase reveals neuronal or glial properties involved inpostnatal development of thalamo-cortical afferents. Alkalinephosphatase activity in neocortical neuropile, especially insensory primary areas, has been found in macaques (Friede,1966). Tissue non-specific alkaline phosphatase has been foundin glia and neurons, apart from blood capillaries and pericytes(Ovtscharoff, 1973). Its activity increases during the fetal andearly postnatal periods and then decreases (Ng and Tam, 1986).This enzyme had been localized on the pre- and postsynapticmembranes in the rat central nervous system (Sugimura andMizutani, 1979) and its presence is related to the formation ofneuronal connections in the central nervous system (Marani et
al., 1995), cell differentiation and onset of synapse developmentin the retina (Araki and Saito, 1986). In vitro studies suggest thatthe putative role of alkaline phosphatase in tissue formation maybe through cell–cell interaction and proliferation (Hui et al.,1993). These different findings document the possible activity ofalkaline phosphatase in developmental neural circuitry; how-ever, its cellular substrates have not yet been identified.
Figure 8. Distributions of vessel sizes in layer IVCalpha and IVCbeta, in the differentmarmosets studied. Diameters have been sorted into classes of 1 µm increment.Numbers of measures realized in each case are presented in Table 3.
Table 3Vessel diameters in layer IVC (µm)
Age 0 PND (00.3) 17 PND (97.9) 24 PND (97.12) 60 PND (99.9) 60 PND (00.5) 7 years (98.4) 11 years (97.7)
Alpha Beta Alpha Beta Alpha Beta Alpha Beta Alpha Beta Alpha Beta Alpha Beta
Mean 7.44 7.12 8.69 8.56 10.51 10.19 9.93 9.68 7.36 7.32 10.51 10.40 11.03 11.37SEM 0.11 0.10 0.12 0.13 0.17 0.16 0.17 0.12 0.08 0.10 0.12 0.14 0.13 0.12n 736 554 901 701 813 658 774 859 964 1020 1044 1057 849 929
n: number of measures.
Cerebral Cortex Feb 2002, V 12 N 2 207
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

Angiogenesis and Neuronal Development
The present study shows that vascular density increases as afunction of age from birth to adulthood. However, this evolutionmay result from two different processes. First, during postnataldevelopment, angiogenesis occurs and supplies an increasingand maturing neuropile. In marmosets, the size of the primaryvisual cortex increases after birth and shows an overshoot at∼ 3 postnatal months (Missler et al., 1993a). Then, as the corticalneuropile is reduced in adults, blood vessels concentrate. Thesize of the visual cortex decreases from the third month until9–12 months after birth, to reach adult values that are not muchdifferent from those of newborns — 105% for the corticalthickness and 112% for the volume (Missler et al., 1993a).
The laminar distribution of cortical vessels, with a strikinglyhigh density in layer IVC, that abruptly ends at the boundarybetween the primary (V1)/secondary (V2) visual cortex as inadult human vascular architecture (Bell and Ball, 1985), mayref lect the cytoarchitectonic features of the cortex. Density ofneuron somata is higher in layer IVC, in the adult as in the youngmarmoset (Fritschy and Garey, 1986). However, the develop-mental variation in vessel density cannot ref lect modifications inthe numerical density of neurons in individual laminae: neuronnumber does not vary greatly during postnatal development, asthere is no evidence for important neuronal loss or mitosisduring postnatal development in marmoset visual cortex(Fritschy and Garey, 1986). Moreover, neuron density is higherin layer IVCbeta than in layer IVCalpha in both young and adults[in macaques (O’Kusky and Colonnier, 1982a,b)], while vesseldensity is first higher only in layer IVCalpha.
The differential increase in vascular supply in layers IVCalphaand IVCbeta during postnatal development is more likely to beassociated with corresponding different metabolic requirementsin neurites. As maturation or changes in cortical function aregenerally associated with modifications in synaptic circuits,angiogenesis and synaptogenesis might be closely linked. In rats,vascular proliferation takes place during synaptogenesis in thevisual cortex (Rowan and Maxwell, 1981; Bakkum et al., 1991).In the marmoset visual cortex, a high rate of synaptogenesisoccurs during the first three postnatal months (Missler et al.,1993b). Unfortunately, the authors of that study did not show thedata for individual layers. In macaques, an overproduction ofsynapses is reached at three postnatal months, followed by aplateau of high synaptic density (Bourgeois and Rakic, 1993).These last authors did not distinguish layers IVCalpha andIVCbeta, but others (O’Kusky and Colonnier, 1982a) have founda higher synaptic density in IVCbeta than in IVCalpha at sixpostnatal months. Data on the time-course of synaptogenesisduring the first three postnatal months are unfortunately un-available.
In the adult, the total density of synapses (O’Kusky andColonnier, 1982a,b), both asymmetric and symmetric (Latawiecet al., 2000), is maintained higher in IVCbeta than in IVCalpha.This result can parallel the higher density of capillaries presentlyestablished in adult marmoset layer IVCbeta.
The postnatal development of dendritic spines is betterdocumented than that of synapses. In macaques, dendritic spinedensity on layer IVC spiny stellate neurons, that represent 95% oflayer IVC neuropile (Mates and Lund, 1983), shows a markedincrease during the first two postnatal months, spine frequencyand spine maturation displaying different time-courses inIVCalpha and IVCbeta (Boothe et al., 1979; Lund and Holbach,1991). The majority of new synapses during postnatal develop-ment form on dendritic spines and represent 75% of thesynapses in the macaque (Bourgeois and Rakic, 1993). From this
pool of data we may then expect a parallel between develop-ment of dendritic spines and synapse densities during the firstpostnatal months. Therefore, if synaptogenesis is shown to occurdifferently in IVCalpha and IVCbeta, it can be hypothesized thatsynaptogenesis produces angiogenesis in layer IVC and that thisangiogenesis proceeds asynchronously in layers IVCalpha andIVCbeta, because of different evolution in the maturation inthese two layers.
From all the elements presented above, it can be proposedthat angiogenesis is due to an increased period of synaptogenesisin the cerebral cortex. If this is so, the initial formation ofthe vascular pattern, as with the initial formation of synapses(Bourgeois and Rakic, 1996), is dominated by intrinsic mechan-isms and stimulation from the periphery would have no effect onangiogenesis. However, an early deprivation of retinal inputincreases the proportion of asymmetrical synapses in layer IVC(Bourgeois and Rakic, 1996). As asymmetrical synapses areputatively excitatory and as the main excitatory (glutamatergic)neurons and asymmetric synapses have a higher oxidative metab-olism than inhibitory (GABAergic) neurons and symmetricsynapses (Carroll and Wong-Riley, 1984; Hertz et al., 1988), onewould expect higher vascularization in layer IVC in the visualcortex of deprived animals than in controls and, in consequence,a higher metabolic activity. This is consistent with positronemission tomography (PET) analyses of visual cortex metabolismin human subjects with early retinal loss, who present a higherglucose utilization than normal subjects (Veraart et al., 1990).
Magnocellular/Parvocellular Maturation
We show in the present paper that vascularization of the primaryvisual cortex is correlated with the differential maturation oflayers IVCalpha and IVCbeta. These two anatomical modulessustain different visual functions. Classically, the occipito-parietal visual stream, dominated by the magnocellular system,deals with spatial localization, motion analyses and luminancesensitivity, and the occipitotemporal stream with patterndiscrimination, object recognition and chromatic sensitivity.Data show that these functional pathways present a differenttime-course of development in both humans and monkeys. Thegeneral trend is that the functions involving the magnocellularpathway mature earlier (Mikami and Fujita, 1992; Kovacs et al.,1999; Dobkins et al., 1999). Physiological studies measuringenergy metabolism in monkeys by means of 2DG auto-radiography reveal that areas of motion analysis (occipitoparietalstream) mature earlier than those of the object recognitionpathway (Bachelier et al., 1991; Distler et al., 1996). If weconsider the different steps of the anatomical pathways from theretina to the primary visual cortex, it appears that, apart from theretinal ganglion cells step (Meissirel et al., 1997), the magno-cellular system matures earlier than the parvocellular system(Gottlieb et al., 1985; Lachica and Casagrande, 1988; Pospichalet al., 1994; Hawken et al., 1997; Kogan et al., 2000). Therefore,convergence of data, through different approaches (anatomical,functional and metabolic), suggests a more precocious magno-cellular system.
Glia Partner
Blobs in the primary visual cortex, revealed by a high level ofcytochrome oxidase activity, coincide with areas expressinghigh density of astroglia (Colombo et al., 1999). As blobs are alsovascular modules (Zheng et al., 1991), we may suggest that themodular organization of the cortex represents, in fact, the closemetabolic coupling between neurons, glial cells and endothelialcells. It is tempting to associate glia, that is a crucial element in
208 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

cortical tissue postnatal development, with the neuron–endothelial partnership.
Glia has a pivotal role in coupling neuronal activity to energymetabolism. A pool of recent data show that astrocytes display anactive part in processing and conveying neuronal informationand may then be involved in plasticity processes. Moreover,astroglia induces blood–brain barrier characteristics, of whichalkaline phosphatase is an indicator, in cerebral endothelial cells,through soluble factors or direct contact (Meyer et al., 1991;Takemoto et al., 1994). Reciprocally, glial differentiation intoastrocytes or oligodendrocytes may result from interactions withblood vessels in the neocortex during postnatal development(Zerlin and Goldmann, 1997). In rats, gliogenesis, angiogenesisand synaptogenesis occur over the same period — the first threepostnatal weeks (Rowan and Maxwell, 1981; Bakkum et al.,1991; Zerlin and Goldmann, 1997). In primates, too, gliogenesismay occur in parallel to synaptogenesis and angiogenesis. Glialcell density f luctuates and glial maturation occurs during earlypostnatal development in marmosets (Fritschy and Garey, 1986;Missler et al., 1994), differently in IVCalpha and IVCbeta inmacaques (O’Kusky and Colonnier, 1982b, Mates and Lund,1983).
If cytological relationships and metabolic coupling betweenglia and neurons, and glial and endothelial cells are documented,the mechanisms of angiogenesis in the cerebral cortex duringpostnatal development and the part of neuronal activity in thisprocess are still poorly understood. Reciprocal neuron–endothelial cell interactions during brain development areexpected, since one receptor to vascular endothelial growthfactor, that stimulates blood vessel growth, was found to be alsothe receptor of semaphorin III, an axonal guidance molecule(Soker et al., 1998). This could represent a coordination systembetween the developing nervous and circulatory systems.
NotesWe are grateful to Pierre Carrechio for his helpful advice in imageanalyses. We thank Pascal Barone for his comments on this manuscript,Catherine Chappert, Catherine Marlot and Frédéric Sarrato for theirtechnical help and Sebastien Aragones for his care in marmoset breeding.Supported by Centre National de la Recherche Scientifique.
Address correspondence to Caroline Fonta, Centre de RechercheCerveau et Cognition, UMR 5549 Centre National de la RechercheScientifique/Université Paul Sabatier, Faculté de Médecine Rangueil,133 route de Narbonne, 31062 Toulouse cedex, France. Email: [email protected].
ReferencesAraki M, Saito T (1986) Alterations in the differentiation of chick retina
caused by an intraocular injection of an alkaline phosphataseinhibitor. Exp Eye Res 43:713–728.
Argandoña EG, Lafuente JV (1996) Effects of dark rearing on thevascularization of the developmental rat visual cortex. Brain Res732:43–51.
Bachevalier J, Hagger C, Mishkin M (1991) Functional maturation of theoccipitotemporal pathway in infant rhesus monkeys. In: Brain workand mental activity (Lassen NA, Ingvar DH, Raichle MEF, Friberg L,eds), pp. 231–240. Copenhagen: Munskgaard.
Bakkum BW, Benevento LA, Cohen RS (1991) Effects of light/dark- anddark-rearing on synaptic morphology in the superior colliculus andvisual cortex of the postnatal and adult rat. J Neurosci Res 28:65–80.
Bell MA, Ball MJ (1985) Laminar variation in the microvascular architec-ture of normal human visual cortex (area 17). Brain Res 335:139–143.
Bell MA, Ball MJ (1990) Neuritic plaques and vessels of visual cortex inaging and Alzheimer’s dementia. Neurobiol Aging 11:359–370.
Bell MA, Scarrow WG (1984) Staining for microvascular alkalinephosphatase in thick celloidin sections of nervous tissue: morpho-metric, and pathological applications. Microvasc Res 27:189–203.
Blasdel GG, Lund JS (1983) Termination of afferent axons in macaquestriate cortex. J Neurosci 3:1389–1413.
Boothe RG, Greenough WT, Lund JS, Wrege K (1979) A quantitativeinvestigation of spine and dendrite development of neurons in visualcortex (area 17) of Macaca nemestria monkeys. J Comp Neurol186:473–490.
Borowsky IW, Collins RC (1989) Metabolic anatomy of brain: a com-parison of regional capillary density, glucose metabolism, and enzymeactivities. J Comp Neurol 288:401–413.
Bourgeois JP, Rakic P (1993) Changes of synaptic density in the primaryvisual cortex of the macaque monkey from fetal to adult stage. JNeurosci 13:2801–2820.
Bourgeois JP, Rakic P (1996) Synaptogenesis in the occipital cortex ofmacaque monkey devoid of retinal input from early embryonic stages.Eur J Neurosci 8:942–950.
Boyd JD, Mavity-Hudson JA, Casagrande VA (2000) The connections oflayer 4 subdivisions in the primary visual cortex (V1) of the owlmonkey. Cereb Cortex 10:644–662.
Carroll EW, Wong-Riley M (1984) Quantitative light and electronmicroscopic analysis of cytochrome oxidase-rich zones in the striatecortex of the squirrel monkey. J Comp Neurol 222:1–17.
Chappert C, Fonta C, Malecaze F, Imbert M (2001) Ocular dominancecolumns in the adult New World Monkey Callithrix jacchus. VisNeurosci (in press).
Colombo JA, Schleicher A, Zilles K (1999) Patterned distribution ofimmunoreactive astroglial processes in the striate (V1) cortex of NewWorld monkeys. Glia 25:85–92.
Distler C, Bachevalier J, Kennedy C, Mishkin M, Ungerleider LG (1996)Functional development of the corticocortical pathway for motionanalysis in the macaque monkey: a 14C-2-deoxyglucose study. CerebCortex 6:185–194.
Dobkins KR, Anderson CM, Lia B (1999) Infant temporal contrastsensitivity functions (tCSFs) mature earlier for luminance than forchromatic stimuli: evidence for precocious magnocellular develop-ment? Vision Res 39:3223–3239.
Endo M, Kaas JH, Jain N, Smith EL, Chino Y (2000) Binocular cross-orientation suppression in the primary visual cortex (V1) of infantrhesus monkeys. Invest Ophthalmol Vis Sci 1:4022–4031.
Fitzpatrick D, Itoh K, Diamond IT (1983) The laminar organization of thelateral geniculate body and the striate cortex in the squirrel monkey(Saimiri sciureus). J Neurosci 3:673–702.
Fitzpatrick D, Lund JS, Blasdel GG (1985) Intrinsic connections ofmacaque striate cortex: afferent and efferent connections oflamina4C. J Neurosci 5:3329–3349.
Fonta C, Chappert C, Imbert M (1997) N-Methyl-D-aspartate subunit R1involvement in the postnatal organization of the primary visual cortexof Callithrix jacchus. J Comp Neurol 386:260–276.
Fonta C, Chappert C, Imbert M (2000) Effect of monocular deprivation onNMDAR1 immunostaining in ocular dominance columns of themarmoset Callithrix jacchus. Vis Neurosci 17:345–352.
Freund TF, Martin KA, Soltesz I, Somogyi P, Whitteridge D (1989)Arborisation pattern and postsynaptic targets of physiologicallyidentified thalamocortical afferents in striate cortex of the macaquemonkey. J Comp Neurol 289:315–336.
Friede RL (1966) A quantitative mapping of alkaline phosphatase in thebrain of the rhesus monkey. J Neurochem 13:197–203.
Fritsches KA, Rosa MGP (1996) Visuotopic organisation of striate cortexin the marmoset monkey (Callithrix jacchus). J Comp Neurol372:264–282.
Fritschy JM, Garey LJ (1986) Quantitative changes in morphologicalparameters in the developing visual cortex of the marmoset monkey.Dev Brain Res 29:173–188.
Gottlieb MD, Pasik P, Pasik T (1985) Early postnatal development of themonkey visual system. I. Growth of the lateral geniculate nucleus andstriate cortex. Brain Res 349:53–62.
Hawken MJ, Blakemore C, Morley JW (1997) Development of contrastsensitivity and temporal-frequency selectivity in primate lateralgeniculate nucleus. Brain Res 14:86–98.
Hendrickson AE, Wilson JR, Ogren MP (1978) The neuroanatomicalorganization of pathways between the dorsal lateral geniculatenucleus and visual cortex in old world and new world primates. JComp Neurol 182:123–136.
Hendrickson AE, Van Brederode JMF, Mulligan KA, Celio MR (1991)Development of the calcium-binding proteins parvalbumin andcalbindin in monkey striate cortex. J Comp Neurol 307:626–646.
Cerebral Cortex Feb 2002, V 12 N 2 209
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

Hertz L, Drejer J, Schousboe A (1988) Energy metabolism in glutama-tergic neurons, GABAergic neurons and astrocytes in primarycultures. Neurochem Res 13:605–610.
Horton JC (1984) Cytochrome oxidase patches: a new cytoarchitectonicfeature of monkey visual cortex. Phil Trans R Soc Lond 304:199–253.
Horton JC, Hocking DR (1996) An adult-like pattern of ocular dominancecolumns in striate cortex of newborn monkeys prior to visualexperience. J Neurosci 16:1791–1807.
Hubel DH, Wiesel TN (1972) Laminar and columnar distribution ofgeniculo-cortical fibers in the macaque monkey. J Comp Neurol146:421–450.
Hui M, Hu M, Tenenbaum HC (1993) Changes in cell adhesion and cellproliferation are associated with expression of tissue non-specificalkaline phosphatase. Cell Tissue Res 274:429–437.
Kennedy H, Bullier J, Dehay C (1985) Cytochrome oxidase activity in thestriate cortex and lateral geniculate nucleus of the new born and adultmacaque monkey. Exp Brain Res 61:204–209.
Kennedy H, Dehay C (1993) Cortical specification of mice and men.Cereb Cortex 3:171–186.
Kogan CS, Zangenehpour S, Chaudhuri A (2000) Developmental profilesof SMI-32 immunoreactivity in monkey striate cortex. Brain Res DevBrain Res 119:85–95.
Kovacs I, Kozma P, Feher A, Benedek G (1999) Late maturation ofvisual spatial integration in humans. Proc Natl Acad Sci USA96:12204–12209.
Lachica EA, Casagrande VA (1988) Development of primate retino-geniculate axon arbors. Vis Neurosci 1:103–123.
Latawiec D, Martin KA, Meskenaite V (2000) Termination of thegeniculocortical projection in the striate cortex of macaque monkey:a quantitative immunoelectron microscopic study. J Comp Neurol419:306–319.
Logothetis NK, Guggenberger H, Peled S, Pauls J (1999) Functionalimaging of the monkey brain. Nat Neurosci 2:555–562.
Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A (2001)Neurophysiological investigation of the basis of the fMRI signal.Nature 412:150–157.
Lund JS (1988) Anatomical organization of macaque monkey striate visualcortex. Annu Rev Neurosci 11:253–288.
Lund JS, Holbach SM (1991) Postnatal development of thalamic recipientneurons in the monkey striate cortex: I. Comparison of spineacquisition and dendritic growth of layer 4C alpha and beta spinystellate neurons. J Comp Neurol 309:115–128.
Marani E, Van Der Veeken JGPM, Lakke EAJF (1995) Simultaneousdemonstration of CD15 and alkaline phosphatase activity in cryostatsections of rat fetuses: a detailed technical description for thedeveloping brain. Eur J Morphol 33:137–147.
Mates SL, Lund JS (1983) Neuronal composition and development inlamina 4C of monkey striate cortex. J Comp Neurol 221:60–90.
Meissirel C, Wikler KC, Chalupa LM, Rakic P (1997) Early evidence ofmagnocellular and parvocellular functional subsystems in the embry-onic primate visual system. Proc Natl Acad Sci USA 94:5900–5905.
Meyer J, Rauh J, Galla HJ (1991) The susceptibility of cerebral endothelialcells to astroglial induction of blood–brain barrier enzymes dependson their proliferative state. J Neurochem 57:1971–1977.
Mikami A, Fujita K (1992) Development of the ability to detect visualmotion in infant macaque monkeys. Dev Psychobiol 25:345–354.
Missler M, Eins S, Merker HJ, Rothe H, Wolff JR (1993a) Pre- and postnataldevelopment of the primary visual cortex of the common marmoset.I. A changing space for synaptogenesis. J Comp Neurol 333:41–52.
Missler M, Wolff A, Merker HJ, Wolff JR (1993b) Pre- and postnataldevelopment of the primary visual cortex of the common marmoset.II. Formation, remodelling, and elimination of synapses as over-lapping processes. J Comp Neurol 333:53–67.
Missler M, Eins S, Böttcher H, Wolff JR (1994) Postnatal development ofglial fibrillary acidic protein, vimentin and S100 protein in monkeyvisual cortex: evidence for a transient reduction of GFAP immuno-reactivity. Dev Brain Res 82:103–117.
Ng TB, Tam PP (1986) Changes of acid and alkaline phosphatase activitiesin the developing mouse brain. Biol Neonate 50:107–113.
O’Kusky J, Colonnier M (1982a) Postnatal changes in the number ofneurons and synapses in the visual cortex (area 17) of the macaquemonkey: a stereological analysis in normal and monocularly deprivedanimals. J Comp Neurol 210:291–306.
O’Kusky J, Colonnier M (1982b) A laminar analysis of the number of
neurons, glia, and synapses in the visual cortex (area 17) of adultmonkeys. J Comp Neurol 210:278–290.
Ovtscharoff W (1973) Ultracytochemische Lokalisierung der alkalischenPhosphatase im Cortex cerebri bei neugeborenen Ratten. Histo-chemie 37:93–95.
Pospichal MW, Florence SL, Kaas JH (1994) The postnatal development ofgeniculocortical axon arbors in owl monkeys. Vis Neurosci 11:71–90.
Powell TP, Hendrickson AE (1981) Similarity in number of neuronsthrough the depth of the cortex in the binocular and monocular partsof area 17 of the monkey. Brain Res 216:409–413.
Rakic P (1977) Prenatal development of the visual cortex of the rhesusmonkey. Philos Trans R Soc Lond B Biol Sci 278:254–260.
Riddle DR, Gutierrez G, Zheng D, White LE, Richards A, Purves D (1993)Differential metabolic and electrical activity in the somatic sensorycortex of juvenile and adult rats. J Neurosci 13:4193–4213.
Rowan R A, Maxwell DS (1981) Patterns of vascular sprouting in thepostnatal development of the cerebral cortex of the rat. Am J Anat160:247–255.
Sirevaag AM, Black JE, Shafron D, Greenough WT (1988) Direct evidencethat complex experience increases capillary branching and surfacearea in visual cortex of young rats. Dev Brain Res 43:299–304.
Soker S, Takashima S, Miao HQ, Neufeld G, Klagsbrun M (1998)Neuropilin-1 is expressed by endothelial and tumor cells as anisoform-specific receptor for vascular endothelial growth factor. Cell92:735–745.
Spatz WB (1979) The retino-geniculo-cortical pathway in Callithrix. II.The geniculo-cortical projection. Exp Brain Res 36:401–410.
Spatz WB, Vogt DM, Illing RB (1993) Distribution of cytochrome oxidasein layers IV and V of the striate cortex in neonate monkeys. Exp BrainRes 95:183–186.
Spatz WB, Illing RB, Vogt Weisenhorn DM (1994) Distribution ofcytochrome oxydase and parvalbumin in the primary visual cortex ofthe adult and neonate monkey, Callithrix jacchus. J Comp Neurol339:519–534.
Sugimura K, Mizutani A (1979) Histochemical and cytochemical studiesof alkaline phosphatase activity in the synapses of rat brain.Histochemistry 61:123–129.
Suner I, Rakic P (1996) Numerical relationship between neurons in thelateral geniculate nucleus and primary visual cortex in macaquemonkeys. Vis Neurosci 13:585–590.
Takemoto H, Kaneda K, Hosokawa M, Ide M, Fukushima H (1994)Conditioned media of glial cell lines induce alkaline phosphataseactivity in cultured artery endothelial cells. Identification ofinterleukin-6 as an induction factor. FEBS Lett 350: 99–103.
Tuor UI, Kurpita G, Simone C (1994) Correlation of local changes incerebral blood f low, capillary density, and cytochrome oxidase duringdevelopment. J Comp Neurol 342:439–448.
Vanzetta I, Grinvald A (1999) Increased cortical oxidative metabolism dueto sensory stimulation: implications for functional brain imaging.Science 286:1555–1558.
Veraart C, De Volder AG, Wanet-Defalque MC, Bol A, Michel C, GoffinetAM (1990) Glucose utilization in human visual cortex is abnormallyelevated in blindness of early onset but decreased in blindness of lateonset. Brain Res 510:115–121.
Wiser AK, Callaway EM (1996) Contributions of individual layer 6pyramidal neurons to local circuitry in macaque primary visualcortex. J Neurosci 16:2724–2739.
Wolff JR, Toldi J, Siklos L, Feher O, Joo F (1992) Neonatal enucleationinduces correlated modification in sensory responsive areas and pialangioarchitecture of the parietal and occipital cortex of albino rats. JComp Neurol 317:187–197.
Wong-Riley M (1979) Changes in the visual system of monocularlysutured or enucleated cats demonstrable with cytochrome oxidasehistochemistry. Brain Res 171:11–28.
Wong-Riley M (1989) Cytochrome oxidase: an endogenous metabolicmarker for neuronal activity. Trends Neurosci 12:94–101.
Wong-Riley M, Carroll EW (1984) Effect of impulse blockage oncytochrome oxidase activity in monkey visual system. Nature307:262–264.
Wong-Riley M, Welt C (1980) Histochemical changes in cytochromeoxidase of cortical barrels after vibrissal removal in neonatal and adultmice. Proc Natl Acad Sci USA 77: 2333–2337.
Wong-Riley MT, Hevner RF, Cutlan R, Earnest M, Egan R, Frost J, NguyenT (1993) Cytochrome oxidase in the human visual cortex: distributionin the developing and the adult brain. Vis Neurosci 10:41–58.
210 Primary Visual Cortex Vascularization • Fonta and Imbert
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from

Woosley TA, Rovainen CM, Cox SB, Henegar MH, Liang GE, Liu D,Moskalenko YE, Sui J, Wei L (1996) Neuronal units linked to micro-vascular modules in cerebral cortex: response elements for imagingthe brain. Cereb Cortex 6:647–660.
Zerlin M, Goldmann JE (1997) Interactions between glial progenitors and
blood vessels during early postnatal corticogenesis: blood vesselcontact represents an early stage of astrocyte differentiation. J CompNeurol 387:537–546.
Zheng D, LaMantia A-S, Purves D (1991) Specialized vascularization of theprimate visual cortex. J Neurosci 11:2622–2629.
Cerebral Cortex Feb 2002, V 12 N 2 211
by guest on June 13, 2016http://cercor.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















