
Role of TNF in the Altered Interaction of Dormant Mycobacterium tuberculosis with Host Macrophages
10
ORIGINAL ARTICLE Characterization of an rpoN mutant of Mesorhizobium ciceri U.S. Gautam 1 , A. Jajoo 2 , A. Singh 1 , P.K. Chakrabartty 3 and S.K. Das 4 1 National Research Center on Plant Biotechnology, Indian Agricultural Research Institute, Pusa, New Delhi, India 2 School of Life Sciences, Devi Ahilya Vishwavidyalaya, Indore, India 3 Madhyamgram Experimental Farm and Department of Microbiology, Bose Institute, Kolkata, India 4 Institute of Life Sciences, Department of Biotechnology, Nalco Square, Bhubaneswar, India Introduction The members of the soil bacteria Rhizobiaceae infect the legume roots and incite the formation of nodules. Meso- rhizobium ciceri induces the formation of nitrogen-fixing nodules on the roots of chickpea (Cicer arietinum). Within the infected cells of the nodules, the bacteria are enveloped in a membrane of plant origin, called the peribacteroid membrane (PBM), and divide and differ- entiate into nitrogen-fixing bacteroids. In the nitrogen- fixation process of rhizobium–legume symbiosis, a large amount of energy is required for the enzyme nitrogenase to reduce atmospheric nitrogen to ammonia. The energy becomes available from the oxidation of reduced carbon supplied by the host to the bacteroid. C 4 -dicarboxylates are the major carbon source taken up by bacteroids and dicarboxylate transport (Dct) system is essential for an effective symbiosis (Ronson et al. 1981). Strains of Rhizobium leguminosarum bv. viciae, Rhizobium legumi- nosarum bv. trifolii, Sinorhizobium meliloti with nonfunc- tional Dct system formed ineffective nodules, and their ability to fix nitrogen was dependent on and influenced by functional dct system (Ronson et al. 1981; Finan et al. 1983; Bolton et al. 1986). Genetic analyses of Dct ) mutants showed that the gene for rhizobial C 4 -dicarb- oxylate permease (dctA) is regulated by the alternative sigma factor, RpoN, and the two-component regulatory system DctB ⁄ D (Jiang et al. 1989). The alternative sigma factor, RpoN, also known as r 54 , is encoded by the gene rpoN (ntrA, glnF). RpoN is essen- tial for transcription initiation by RNA polymerase at promoters characterized by an invariant GG doublet at 24 and a GC doublet at 12 upstream of the transcriptional start site (Dixon 1987). The gene rpoN works in Keywords Cicer arietinum (L), dicarboxylate transport, Mesorhizobium ciceri, nitrogen fixation, rpoN gene, transposon mutagenesis. Correspondence Subrata K. Das, Institute of Life Sciences, Department of Biotechnology, Nalco Square, Bhubaneswar – 751 023, India. E-mail: [email protected] 2006 ⁄ 1736: received 11 December 2006, revised 13 March 2007 and accepted 21 March 2007 doi:10.1111/j.1365-2672.2007.03432.x Abstract Aims: To study the genetic basis of C 4 -dicarboxylate transport (Dct) in relation to symbiotic nitrogen fixation in Mesorhizobium ciceri. Methods and Results: A Tn5-induced mutant strain (TL16) of M. ciceri, unable to grow on C 4 -dicarboxylates, was isolated from the wild-type strain TAL 620. The mutant lacked activities of the enzymes, which use C 4 -dicarboxylates as substrate. The sequencing of the 3Æ2kb EcoRI fragment, which was the site of Tn5 insertion, revealed three complete and two partial open reading frames. In the mutant, Tn5 interrupted the rpoN gene, of which only one copy was there. Complementation and biochemical studies suggest that the M. ciceri rpoN activ- ity is required for C 4 -Dct, maturation of bacteroids and symbiotic nitrogen fix- ation. The fine structure of the ineffective nodules produced by TL16 on Cicer arietinum L changed in comparison with those produced by the wild type. Conclusions: The mutant strain TL16 suffered a disruption in the rpoN gene. Only one copy of rpoN gene is present in M. ciceri. The mutation abolishes Dct activity. It additionally abolishes the symbiotic nitrogen fixation activity of the bacteroids in the nodules. Significance and Impact of the Study: This first document in M. ciceri shows that a functional rpoN gene is essential for the transport of dicarboxylic acids and symbiotic nitrogen fixation. Journal of Applied Microbiology ISSN 1364-5072 1798 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 ª 2007 The Authors
Transcript of Role of TNF in the Altered Interaction of Dormant Mycobacterium tuberculosis with Host Macrophages

ORIGINAL ARTICLE
Characterization of an rpoN mutant of Mesorhizobium ciceriU.S. Gautam1, A. Jajoo2, A. Singh1, P.K. Chakrabartty3 and S.K. Das4
1 National Research Center on Plant Biotechnology, Indian Agricultural Research Institute, Pusa, New Delhi, India
2 School of Life Sciences, Devi Ahilya Vishwavidyalaya, Indore, India
3 Madhyamgram Experimental Farm and Department of Microbiology, Bose Institute, Kolkata, India
4 Institute of Life Sciences, Department of Biotechnology, Nalco Square, Bhubaneswar, India
Introduction
The members of the soil bacteria Rhizobiaceae infect the
legume roots and incite the formation of nodules. Meso-
rhizobium ciceri induces the formation of nitrogen-fixing
nodules on the roots of chickpea (Cicer arietinum).
Within the infected cells of the nodules, the bacteria are
enveloped in a membrane of plant origin, called the
peribacteroid membrane (PBM), and divide and differ-
entiate into nitrogen-fixing bacteroids. In the nitrogen-
fixation process of rhizobium–legume symbiosis, a large
amount of energy is required for the enzyme nitrogenase
to reduce atmospheric nitrogen to ammonia. The energy
becomes available from the oxidation of reduced carbon
supplied by the host to the bacteroid. C4-dicarboxylates
are the major carbon source taken up by bacteroids and
dicarboxylate transport (Dct) system is essential for an
effective symbiosis (Ronson et al. 1981). Strains of
Rhizobium leguminosarum bv. viciae, Rhizobium legumi-
nosarum bv. trifolii, Sinorhizobium meliloti with nonfunc-
tional Dct system formed ineffective nodules, and their
ability to fix nitrogen was dependent on and influenced
by functional dct system (Ronson et al. 1981; Finan et
al. 1983; Bolton et al. 1986). Genetic analyses of Dct)
mutants showed that the gene for rhizobial C4-dicarb-
oxylate permease (dctA) is regulated by the alternative
sigma factor, RpoN, and the two-component regulatory
system DctB ⁄ D (Jiang et al. 1989).
The alternative sigma factor, RpoN, also known as r54,
is encoded by the gene rpoN (ntrA, glnF). RpoN is essen-
tial for transcription initiation by RNA polymerase at
promoters characterized by an invariant GG doublet at 24
and a GC doublet at 12 upstream of the transcriptional
start site (Dixon 1987). The gene rpoN works in
Keywords
Cicer arietinum (L), dicarboxylate transport,
Mesorhizobium ciceri, nitrogen fixation, rpoN
gene, transposon mutagenesis.
Correspondence
Subrata K. Das, Institute of Life Sciences,
Department of Biotechnology, Nalco Square,
Bhubaneswar – 751 023, India.
E-mail: [email protected]
2006 ⁄ 1736: received 11 December 2006,
revised 13 March 2007 and accepted 21
March 2007
doi:10.1111/j.1365-2672.2007.03432.x
Abstract
Aims: To study the genetic basis of C4-dicarboxylate transport (Dct) in relation
to symbiotic nitrogen fixation in Mesorhizobium ciceri.
Methods and Results: A Tn5-induced mutant strain (TL16) of M. ciceri, unable
to grow on C4-dicarboxylates, was isolated from the wild-type strain TAL 620.
The mutant lacked activities of the enzymes, which use C4-dicarboxylates as
substrate. The sequencing of the 3Æ2kb EcoRI fragment, which was the site of
Tn5 insertion, revealed three complete and two partial open reading frames. In
the mutant, Tn5 interrupted the rpoN gene, of which only one copy was there.
Complementation and biochemical studies suggest that the M. ciceri rpoN activ-
ity is required for C4-Dct, maturation of bacteroids and symbiotic nitrogen fix-
ation. The fine structure of the ineffective nodules produced by TL16 on Cicer
arietinum L changed in comparison with those produced by the wild type.
Conclusions: The mutant strain TL16 suffered a disruption in the rpoN gene.
Only one copy of rpoN gene is present in M. ciceri. The mutation abolishes
Dct activity. It additionally abolishes the symbiotic nitrogen fixation activity of
the bacteroids in the nodules.
Significance and Impact of the Study: This first document in M. ciceri shows
that a functional rpoN gene is essential for the transport of dicarboxylic acids
and symbiotic nitrogen fixation.
Journal of Applied Microbiology ISSN 1364-5072
1798 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807
ª 2007 The Authors

conjunction with the NtrC class of transcriptional activa-
tors to control the expression of diverse genes, including
those responsible for nitrogen fixation, Dct, assimilation
and dissimilation of nitrate and catabolism of aromatic
compounds (Kustu et al. 1989; Merrick 1993). The rpoN
has been reported from the symbiotically nitrogen-fixing
bacteria Rhizobium etli (Michiels et al. 1998b), Sinorhizo-
bium meliloti (Ronson et al. 1987), Rhizobium sp. strain
NGR234 (Van Slooten et al. 1990) and Bradyrhizobium
japonicum (Kullik et al. 1991) for its role in nitrogen
metabolism. In S. meliloti, the rpoN locus has been
mutated and sequenced (Ronson et al. 1987). The mutant
formed Fix- nodules on alfalfa and failed to grow on
C4-dicarboxylates (Albright et al. 1989), while rpoN
mutant of Rhizobium sp. strain NGR234 showed pleio-
trophic phenotype, including an effect on nodulation
gene expression (Van Slooten et al. 1990). In R. etli, two
rpoN genes encoding the r54 have been characterized and
shown to be differentially regulated (Michiels et al.
1998a). During free living growth, rpoN1 is required for
growth on several nitrogen and carbon sources. There is a
severe decrease in nitrogen fixation after inactivation of
rpoN2 (Michiels et al. 1998a), indicating the essential role
of this gene in nitrogen fixation by bacteroids. To date,
no information, whatsoever, is available on the rpoN regu-
lon of M. ciceri. The genetic basis of C4-Dct, in relation
to formation of nodules and nitrogen fixation activity in
this species, has not been studied.
In the present study, we report the isolation of a Tn5-
induced mutant of M. ciceri, strain TL16. The mutant is
phenotypically nonfunctional in Dct and forms ineffective
nodules. Cloning of the mutant gene and analysis of the
nucleotide sequence of the gene provide evidence that, in
the mutant strain TL16, the rpoN gene is disrupted by
Tn5. We conclude that rpoN-encoded alternative sigma
factor is required for symbiotic functions: nodule organo-
genesis, and nodule functions like C4-Dct and nitrogen
fixation in M. ciceri. To the best of our knowledge, this is
the first report suggesting a role of rpoN in symbiotic
functions, including transport of dicarboxylate and
synthesis of the enzymes which use C4-dicarboxylate as
substrate in M. ciceri.
Materials and methods
Bacterial strains, plasmids and media
Bacterial strains and plasmids used in this study are listed
in Table 1. Mesorhizobium ciceri was grown in yeast
extract-mannitol (YM) medium (Vincent 1970) or in rhiz-
obium minimal medium containing: (g l)1) K2HPO4, 2Æ0;
KH2PO4, 1Æ5; NaCl, 0Æ15; NH4Cl, 0Æ5; MgSO4.7H2O, 0Æ5;
Table 1 Bacterial strains and plasmids used
Bacterial strains Relevant phenotype* Reference
Escherichia coli S17Æ1 Pro, hsdR, recA [RP4.2 (Tc::Mu) (Km::Tn7)] Simon et al. (1983)
DH5a E. coli host strain, endA1 hsdR17 supE44
thi-1 recA1 gyrA96 relA1 D (argF-lac ZYA)
Bethesda Research
Laboratories, Inc.
HB101 F-hsdS20(rB- mB
-) recA13 ara-14 proA2 LacY1
galK2 rpsL20 xyl-5 mtl-1 supE44k-
Promega
Mesorhizobium ciceri
TAL620 (wild type)
Nxr, Apr, cmr, Neos, Cms, Tcs, Rifs NifTAL*
TL16 (mutant) Nxr, dct, rpoN::Tn5, Neor This study
Plasmids
pSUP5011 Apr, Cmr, Neor, pBR322 derivative containing
Tn5-mob
Simon (1984)
pRK2013 Tra+, Kmr, ColE1 replicon Figurski and Helinski (1979)
pLAFR1 Cosmid cloning vector, Tcr Friedman et al. (1982)
pBlueScriptKS+ Apr, LacZ’, T7 Phil10 promoter, f1 ori Stratagene
pGEM-TEasy Apr, cloning vector Promega
pUS1 10Æ9-kb EcoRI fragment of rpoN::Tn5 insert
of strain TL16 in pBlueScriptKS+
This study
pUS2 1Æ527-kb PCR amplified fragment containing
rpoN gene of strain TAL620 in pGEM-TEasy vector
This study
pUS3 2Æ1-kb PCR amplified fragment containing rpoN
gene of strain TAL620 in a cosmid cloning vector pLAFR1
This study
*NifTAL, NifTAL project and MIRCEN, University of Hawaii, Paia, Hawaii, USA.
s, sensitive; r, resistant; Nx, nalidixic acid; Neo, neomycin; Km, kanamycin; Ap, ampicillin; Rif, rifampicin; Cm, chloramphenicol; Tp, trimethoprim;
Tc, tetracycline; Sm, streptomycine; dct, dicarboxylic acid transport defective.
U.S. Gautam et al. Mesorhizobium ciceri rpoN mutant
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 1799

CaCl2.2H2O, 0Æ01, and sodium succinate, 5Æ0 or mannitol,
10Æ0 as carbon source, pH 6Æ8. Escherichia coli was grown in
Luria broth (Miller 1972). Antibiotic concentrations for
E. coli strain S17Æ1 ⁄ pSUP5011::Tn5-mob were as follows:
streptomycin, 50 lg ml)1; neomycin, 50 lg ml)1; and either
chloramphenicol, 30 lg ml)1 or ampicillin, 50 lg ml)1. For
growing M. ciceri, nalidixic acid was added at 25 lg ml)1
to the medium. The media were solidified with agar-agar
powder (Difco) at 15Æ0 g l)1 when necessary.
For enzyme assays, M. ciceri strains were grown in YM
medium for 2 days, washed and resuspended in 200 ml of
rhizobium minimal medium containing 10-mmol l)1 suc-
cinate and incubated for 6 h at 28�C on a shaker (Adolf
Kuhner AG, ISF-1-V, Schweiz) at 200 rev min)1 for the
induction of Dct activity.
Tn5 mutagenesis
For mutagenesis, the mating of the recipient strain
M. ciceri TAL620, grown in YM medium for 2 days at
28�C and the donor E. coli strain S17Æ1, harbouring the
suicide plasmid pSUP5011:: Tn5-mob (Simon et al. 1983;
Simon 1984), grown in Luria broth for 12 h at 37�C, was
carried out (Das et al. 2006). Transconjugants were selec-
ted on rhizobium minimal medium containing glucose
and succinate to which 2,3,5-triphenyl tetrazolium
chloride (30 lg ml)1), nalidixic acid (25 lg ml)1) and
neomycin (100 lg ml)1) were added. After 6 days of
incubation at 28�C, mutants defective in the utilization of
dicarboxylic acids were isolated by screening the nalidixic
acid–neomycin-resistant transconjugants, which appeared
as white colonies on succinate–glucose–tetrazolium plates
(Ronson et al. 1981), but failed to grow on succinate.
Growth
Wild-type M. ciceri TAL620 and the rpoN mutant strain
TL16 were analysed for their growth in rhizobium min-
imal medium containing mannitol (10 g l)1) as carbon
source. The cells were initially grown in YM medium,
harvested, washed with minimal medium and were used
as inocula. The growth of cells was monitored by measur-
ing the optical density at 600 nm in a SPECORD210
spectrophotometer, Analytik Jena, Germany.
Dicarboxylate uptake assay
For the induction of Dct activity, the cells were incubated
for 4 h in rhizobium minimal medium containing
10 mmol l)1 of either succinate or malate as appropriate.
Dicarboxylate uptake assays with induced M. ciceri
cells (300 lg of cell protein) were carried out at 30�C in
2-ml minimal medium (Bolton et al. 1986) containing
40 lmol l)1 [1,4-14C] succinate (specific activity 130 mci
;mmol)1) or 60 l mol l)1 [U-14C] malate (specific activity
192 mci mmol)1) in the presence of 5-mmol l)1 unlabelled
succinate or malate as appropriate. During incubation, an
aliquot of the sample was withdrawn at regular time inter-
val, passed through a filter (pore size, 0Æ45 lm; Millipore
Corp., Billerica, MA, USA), and washed twice each time
with 10 ml of cold minimal medium. The filters containing
the cells were air-dried and placed into vials containing
10 ml of scintillation fluid. 14C incorporation was counted
in a Beckman LS 8000 scintillation counter.
Preparation of cell-free extract
Cells grown in minimal medium were washed and suspen-
ded in 50-mmol l)1 tris-HCl buffer containing 1 mmol l)1
MgCl2, pH 7Æ4, for extraction of malate dehydrogenase or
50mmol l)1 potassium phosphate buffer (pH 7Æ4) for the
extraction of fumarase or succinate dehydrogenase. The
cells were disrupted by sonication with a Labsonic M
sonicator (B. Braun Biotech International, Melsungen,
Germany). The extract was centrifuged at 10 000 g for
30 min at 4�C and the clear supernatant was used as the
source of enzymes.
Enzyme assays
Enzyme assays were done in a spectrophotometer at 25�C.
Malate dehydrogenase was assayed following the method
of Yoshida (1969). Fumarase was assayed according to
Hill and Bradshaw (1969). Succinate dehydrogenase was
assayed following the protocol of Veeger et al. (1969). Dur-
ing enzyme assays, appropriate corrections were made for
substrate and enzyme blanks. Specific activities were
expressed as nanomoles of product formed per minute per
milligram protein. Protein was estimated using bovine
serum albumin as the standard (Bradford 1976).
DNA manipulation, cloning and sequencing
DNA was isolated according to standard protocol (Meade
et al. 1982). Routine manipulation of DNA, plasmid
isolation, construction of recombinant plasmids, electro-
phoresis of DNA and transformation were carried out
according to standard procedures (Sambrook et al. 1989).
Digestions with restriction enzymes and DNA ligation
were performed according to the manufacturers’ instruc-
tions (Promega, Inc. and New England Biochemical,
Beverly, MA, USA). Southern blotting was performed by
using a standard Southern blot protocol (Southern 1975)
as described previously (Das and Mishra 1996).
DNA sequencing was performed as described (Das
et al. 2006). Open reading frames (ORF) were identified
Mesorhizobium ciceri rpoN mutant U.S. Gautam et al.
1800 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807
ª 2007 The Authors

using BLASTX (Altschul et al. 1990). Comparisons of
DNA sequences and their derivative amino acid sequences
with sequences in the European Molecular Biology
Laboratory (EMBL) ⁄ GenBank were performed using
BLAST (Altschul et al. 1997) and CLUSTALX (Thompson
et al. 1994).
PCR amplification and cloning of the rpoN gene of
Mesorhizobium ciceri for complementation
The complete coding region of the rpoN gene from
M. ciceri strain TAL620 was amplified using the primers F1
(5¢-GAA GTC CGG ATG CGT AGA CGA GGC 3¢) and
R1 (5¢ TGG CTT GTG GAA AAT CGA CCC GCC-3¢),
and a 1527-bp product was obtained. The primers were
designed from the sequences of the flanking regions of
the rpoN genes of other bacteria in the database. PCR was
conducted, and the resulting DNA fragment was purified
using a QIAquick PCR purification kit (Qiagen, Hilden,
Germany) and cloned into pGEM-TEasy plasmid vector
(Promega) for sequencing as described (Das et al. 2006).
The oligonucleotide primers rpoNF (5¢- GCG GAA
TTC TGA AAT TTG TTT CCT GAC -3¢) and rpoNR (5¢-CCG GAA TTC AGG CCA ACG TCG TGA CGG -3¢)were used for PCR amplification of about 2Æ1 kb of
genomic DNA containing the rpoN gene (ORF2), intro-
ducing EcoRI site (underlined) into the PCR product.
After digestion with EcoRI, the PCR product was cloned
into the cosmid cloning vector pLAFR1 to yield pUS3.
Cosmid pSU3 was used to complement the mutant TL16.
Triparental spot mating was performed using the recipi-
ent TL16, the donor pSU3 and the helper strain
HB101 ⁄ pRK2013. The exconjugants were identified by
streaking on rhizobium minimal medium supplemented
with succinate or malate.
Genomic context analysis
Genomic context of rpoN gene of M. ciceri strain
TAL620 in comparison with the genomes of related
nitrogen-fixing members of rhizobiales were analysed
using the GeConT programme. This programme is avail-
able at: http://www.ibt.unam.mx/biocomputo/gecont.html
(Ciria et al. 2004).
Plant tests
Plant tests were carried out using surface-sterilized seeds
of chickpea (C. arietinum L) cv. Pusa C-235, as described
previously (Raychaudhuri et al. 2005). Each test was con-
ducted with three plants, and four replicates were consid-
ered for each test. At 35 days of inoculation, the shoot
portions of the plants were cut at the stem–root junction
and placed in an oven at 70�C for 5 days for dry weight
determination. Acetylene reduction assays were performed
(Ronson and Primrose 1979) using the nodules as des-
cribed previously (Das et al. 2006).
Microscopy
Root nodules were excised from the plants at 35 days of
inoculation and washed with 0Æ05-mol l)1 potassium phos-
phate buffer, pH 6Æ8. Slices from the central area of root
nodules were fixed in 2Æ5% glutaraldehyde in 0Æ05 mol l)1
potassium phosphate buffer, pH 6Æ8. Subsequent steps of
tissue preparations were carried out according to standard
protocol (Karnovsky 1965; Finan et al. 1983). Sections for
light microscopy were stained with 0Æ01% toluidine blue.
Ultra-thin sections for electron microscopy were stained
with 1% uranyl acetate, pH 4Æ5, followed by lead citrate
(Reynolds 1963) and examined with a Philips CM10
electron microscope.
Nucleotide sequence accession number
The nucleotide sequence data of M. ciceri reported in this
communication have been submitted to the GenBank
nucleotide sequence database under accession nos: ORF1,
EF079828; ORF2, AY850034; ORF3, EF460345; ORF4,
EF460347; and ORF5, EF460346.
Results
Transposon mutagenesis and isolation of mutant
Tn5 was introduced into M. ciceri strain TAL620 using the
suicide plasmid pSUP5011 containing Tn5-Mob from
E. coli S17Æ1 developed by Simon et al. (1983). Transconju-
gants occurred at a frequency of 4Æ2 · 10)4 per donor.
Spontaneous neomycin (100 lg ml)1)-resistant colonies of
M. ciceri strain TAL620 occurred at a very low frequency
(<10)9). The selection for the M. ciceri transconjugants was
carried out on YM-agar plates containing neomycin and
nalidixic acid. Over 5000 nalidixic acid–neomycin-resistant
transconjugants were obtained. Screening on minimal
medium containing glucose and succinate as carbon
sources and 2,3,5-triphenyl tetrazolium chloride allowed
the isolation of several white colonies of mutants, of which,
TL16 (Table 1) was selected for further studies.
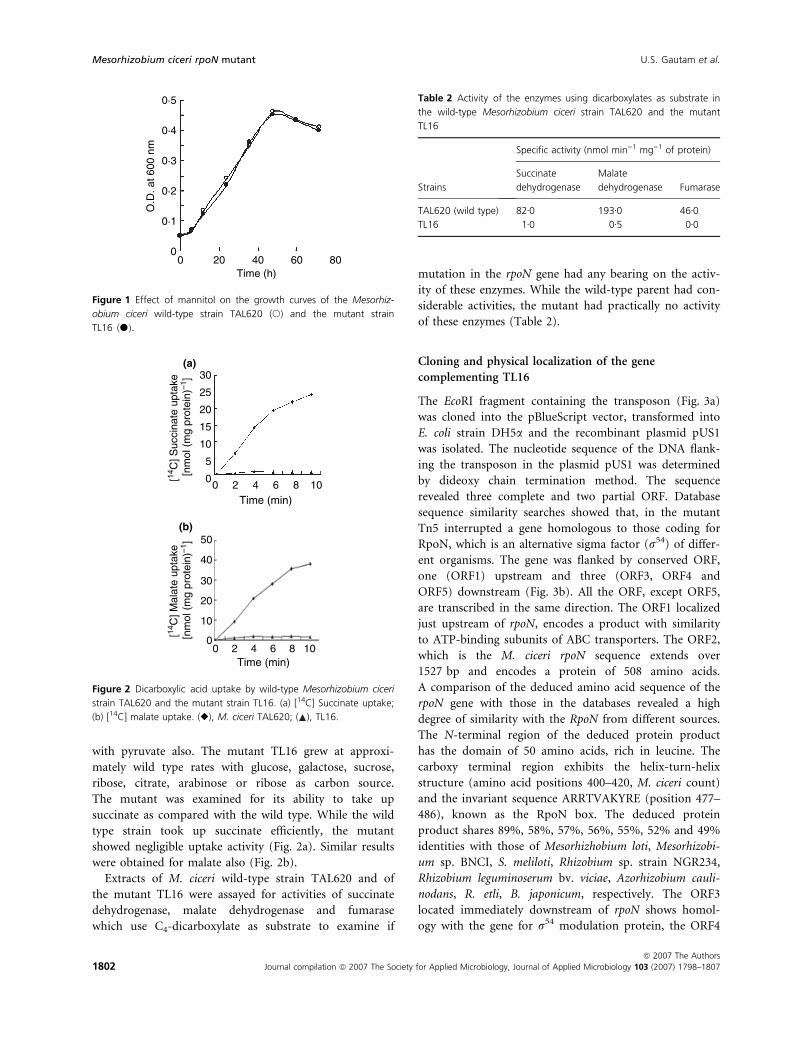
Phenotypes of Mesorhizobium ciceri mutant
A phenotype of the M. ciceri mutant TL16 was its failure
to grow with succinate, malate or fumarate as carbon
source; whereas growth with mannitol was similar to that
of the wild type (Fig. 1). Similar results were obtained
U.S. Gautam et al. Mesorhizobium ciceri rpoN mutant
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 1801
with pyruvate also. The mutant TL16 grew at approxi-
mately wild type rates with glucose, galactose, sucrose,
ribose, citrate, arabinose or ribose as carbon source.
The mutant was examined for its ability to take up
succinate as compared with the wild type. While the wild
type strain took up succinate efficiently, the mutant
showed negligible uptake activity (Fig. 2a). Similar results
were obtained for malate also (Fig. 2b).
Extracts of M. ciceri wild-type strain TAL620 and of
the mutant TL16 were assayed for activities of succinate
dehydrogenase, malate dehydrogenase and fumarase
which use C4-dicarboxylate as substrate to examine if
mutation in the rpoN gene had any bearing on the activ-
ity of these enzymes. While the wild-type parent had con-
siderable activities, the mutant had practically no activity
of these enzymes (Table 2).
Cloning and physical localization of the gene
complementing TL16
The EcoRI fragment containing the transposon (Fig. 3a)
was cloned into the pBlueScript vector, transformed into
E. coli strain DH5a and the recombinant plasmid pUS1
was isolated. The nucleotide sequence of the DNA flank-
ing the transposon in the plasmid pUS1 was determined
by dideoxy chain termination method. The sequence
revealed three complete and two partial ORF. Database
sequence similarity searches showed that, in the mutant
Tn5 interrupted a gene homologous to those coding for
RpoN, which is an alternative sigma factor (r54) of differ-
ent organisms. The gene was flanked by conserved ORF,
one (ORF1) upstream and three (ORF3, ORF4 and
ORF5) downstream (Fig. 3b). All the ORF, except ORF5,
are transcribed in the same direction. The ORF1 localized
just upstream of rpoN, encodes a product with similarity
to ATP-binding subunits of ABC transporters. The ORF2,
which is the M. ciceri rpoN sequence extends over
1527 bp and encodes a protein of 508 amino acids.
A comparison of the deduced amino acid sequence of the
rpoN gene with those in the databases revealed a high
degree of similarity with the RpoN from different sources.
The N-terminal region of the deduced protein product
has the domain of 50 amino acids, rich in leucine. The
carboxy terminal region exhibits the helix-turn-helix
structure (amino acid positions 400–420, M. ciceri count)
and the invariant sequence ARRTVAKYRE (position 477–
486), known as the RpoN box. The deduced protein
product shares 89%, 58%, 57%, 56%, 55%, 52% and 49%
identities with those of Mesorhizhobium loti, Mesorhizobi-
um sp. BNCI, S. meliloti, Rhizobium sp. strain NGR234,
Rhizobium leguminoserum bv. viciae, Azorhizobium cauli-
nodans, R. etli, B. japonicum, respectively. The ORF3
located immediately downstream of rpoN shows homol-
ogy with the gene for r54 modulation protein, the ORF4
10
Time (min)
30(a)
25
20
15
10
5
00 108642
[14C
] Suc
cina
te u
ptak
e[n
mol
(m
g pr
otei
n)–1
]
(b)50
40
30
20
10
00 8642
Time (min)
[14C
] Mal
ate
upta
ke[n
mol
(m
g pr
otei
n)–1
]
Figure 2 Dicarboxylic acid uptake by wild-type Mesorhizobium ciceri
strain TAL620 and the mutant strain TL16. (a) [14C] Succinate uptake;
(b) [14C] malate uptake. (r), M. ciceri TAL620; ( ), TL16.
Time (h)
O.D
. at 6
00 n
m
0·5
0·4
0·3
0·2
0·1
00 20 40 60 80
Figure 1 Effect of mannitol on the growth curves of the Mesorhiz-
obium ciceri wild-type strain TAL620 (s) and the mutant strain
TL16 (d).
Table 2 Activity of the enzymes using dicarboxylates as substrate in
the wild-type Mesorhizobium ciceri strain TAL620 and the mutant
TL16
Strains
Specific activity (nmol min)1 mg)1 of protein)
Succinate
dehydrogenase
Malate
dehydrogenase Fumarase
TAL620 (wild type) 82Æ0 193Æ0 46Æ0
TL16 1Æ0 0Æ5 0Æ0
Mesorhizobium ciceri rpoN mutant U.S. Gautam et al.
1802 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807
ª 2007 The Authors

with the gene of phosphotransferase system enzyme II
and the ORF5 with that of NADH oxidase.
A 1Æ527-kb stretch of chromosomal DNA containing the
ORF2 (rpoN gene) was amplified from the wild-type strain
TAL620 by PCR. It was cloned into pGEM-TEasy vector;
subsequently, the recombinant plasmid pUS2 was isolated
and the rpoN gene in the clone was sequenced. The location
of the Tn5 mutation was determined from the sequence of
the transposon insertion junction in the rpoN gene frag-
ment, which was earlier amplified by PCR from the DNA
of the wild-type strain, TAL620 and sequenced. It was
found that Tn5 insertion was within the codons 150–153.
The genomic context of rpoN gene in M. ciceri, as com-
pared with that in the related nitrogen-fixing members of
Rhizobiales shows that it is almost identical to that of
M. loti, although considerable homogeneity in composi-
tion was also observed with those of B. japonicum, M. loti
and R. etli. All the four genomes are observed to have
gene orthologs for ATP-binding protein, sigma 54 pro-
tein, sigma 54 modulation protein and phosphotransf-
erase system enzyme II in their genome (Fig. 3c).
For confirmation, complementation of mutant TL16
with the ORF coding for rpoN gene was carried out.
A 2Æ1 kb DNA fragment containing rpoN gene was ampli-
fied by PCR from the strain TAL620 and cloned into
pLAFR1 to yield pUS3. The cosmid pSU3 was used to
complement the mutant TL16 by triparental mating to
yield the exconjugant TL16 ⁄ US3 (Table 3). The cosmid
pUS3 was able to complement TL16, and exhibited pheno-
type similar to that of the wild type in respect to growth
B. japonicum
Tn5-mob (7·7 kb)
EcoRI
EcoRI
Allur1
Pst1
Sma1
Bgl1Sal1 Sac2
Pvu1
Sal1
Msc1
EcoRI
EcoRI
2·0 kb
(a)
(b)
(c)
10·9 kb
3·2 kb3210
ORF3ORF1
blr0722
RHE
ABC
rpoN2 bl
ml
RHE
smp
m
p
NADH
NADH
RH
p b
mll3196
rpoNch
rpoN ptsll
mll3
ORF2 (rpoN) ORF4 ORF5
M. loti
R. etli
M. ciceri
Figure 3 Genetic organization and genomic context of the Mesorhizobium ciceri mutant strain TL16 rpoN DNA region. Panel a, the EcoRI-diges-
ted DNA fragment containing the transposon Tn5; panel b, the restriction sites and transcriptional directions of ORF1, ORF2 (rpoN), ORF3, ORF4
and ORF5; panel c, genomic context and organization conservation of the rpoN region in the genome of M. ciceri in comparison with genomes
of related nitrogen-fixing members of Rhizobiales. Arrows represent genes with their relative orientation in the genomes. Genes are coloured
according to their functional categories and labelled according to their original gene annotation in the database. Yellow, ATP-binding protein;
orange, sigma 54 protein; red, sigma 54 modulation protein; green, phosphotransferase system enzyme II; grey, putative permease protein; light
brown, NADH oxidase; and pink, acetyl transferase.
Table 3 Symbiotic properties of the wild-type Mesorhizobium ciceri
TAL620 and the rpoN mutant TL16
Strains
Shoot dry
weight (g)*
Acetylene
reduction�
TAL620 1Æ402 ± 0Æ27 1959 ± 128Æ4
TL16 0Æ517 ± 0Æ11 –
TL16 ⁄ US3 (TL16 containing pUS3) 1Æ29 ± 0Æ18 1928 ± 119
Uninoculated control 0Æ496 ± 0Æ13 –
*Dry weight is expressed as the mean values in g ± the standard devi-
ation of four replicates, each with three plants.
�Acetylene reduction activities are expressed as the mean of nano-
moles of ethylene formed per hour ± the standard deviation of four
replicates, each with three nodulated plant roots.
–, not detectable.
U.S. Gautam et al. Mesorhizobium ciceri rpoN mutant
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 1803

on succinate or malate and the acetylene reduction
activity (Table 3).
When genomic DNA from the strain TL16 was digested
with EcoRI restriction enzyme and hybridized with 32P-
labelled internal HindIII fragment (�5Æ4 kb) of Tn5-mob
as probe, a positive signal was obtained on a 10Æ9 kb
DNA fragment of TL16 (Fig. 4a). To examine the copy
number of rpoN gene present in M. ciceri, genomic DNA
of the strain TAL620 or TL16 was digested with EcoRI
and hybridized with the 1Æ527 kb PCR amplified rpoN
gene sequence as probe. Only one hybridization band
signal was observed either on a 10Æ9 kb EcoRI DNA
fragment of TL16 or on a 3Æ2-kb EcoRI DNA fragment
of TAL620 (Fig. 4b).
Symbiotic properties of the rpoN mutant
The symbiotic properties of the mutant TL16 was investi-
gated after inoculation of chickpea seedlings growing
under nitrogen-deficient condition. Plants inoculated with
the strain TL16 were pale green. At 35 days, the mean dry
weight of the shoots of the plants (1Æ402 g per three
plants) inoculated with the wild-type strain TAL620 was
almost three times that of the plants (0Æ517 g per three
plants) inoculated with the mutant strain TL16. The
mean dry weight of the plants inoculated with the mutant
was, however, not appreciably different from that of the
uninoculated control (Table 3). The nodules formed by
the wild-type strain had a high acetylene reductase activ-
ity (Table 3). In contrast, the acetylene reduction activity
of the nodules formed by the rpoN mutant was absent
(Table 3).
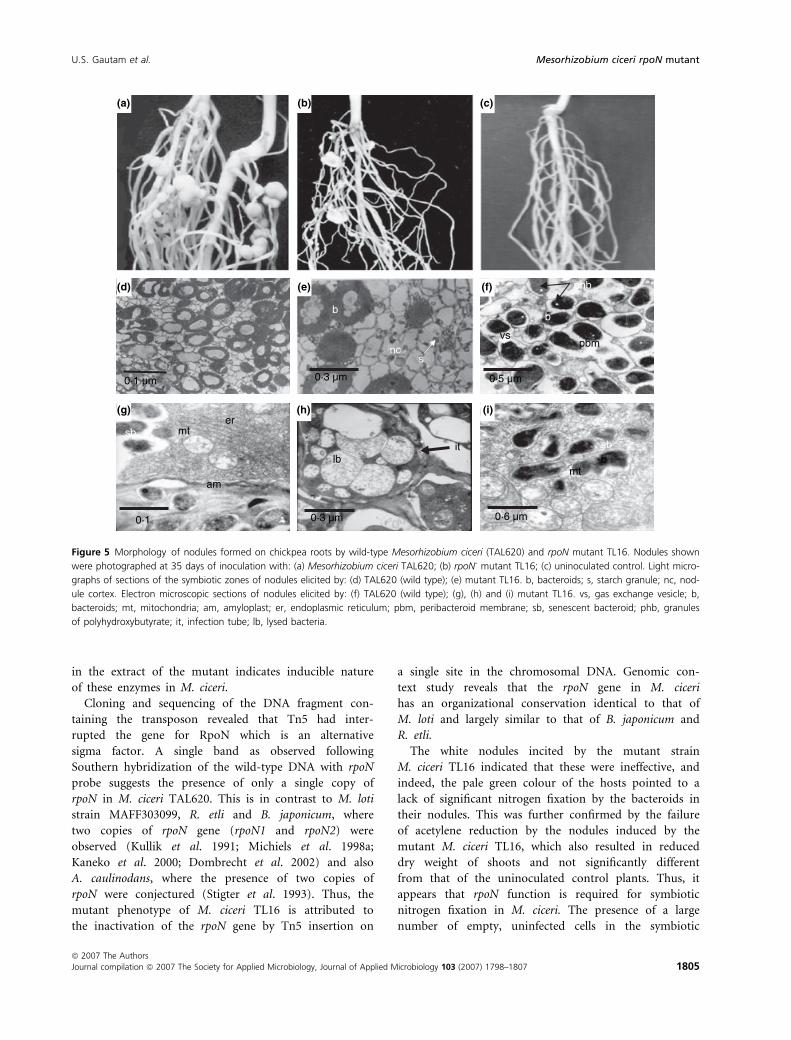
Under nitrogen-deficient condition, nodules induced
by the wild-type strain TAL620 were pink and spherical
(Fig. 5a). In contrast, nodules induced by the mutant
TL16 were small, white and often wrinkled (Fig. 5b).
The control plants without bacterial inoculation, as
expected, had no nodules on their roots (Fig. 5c). Nod-
ules formed by the rpoN mutant M. ciceri TL16 were
examined to determine if the absence of nitrogen fix-
ation was accompanied by any abnormality in nodule
structure. Light microscopic observations revealed that
the nodules contained a large number of empty, un-
infected cells, while only a few infected host cells were
present within a poorly developed symbiotic zone. The
infected cells along with the neighbouring uninfected
cells contained many starch granules near their peripher-
ies as compared with the nodules induced by the wild
type (Fig. 5d,e). Electron microscopic observations
revealed that the nodules induced by the wild type con-
tained mature bacteroids surrounded by a PBM and also
poly-hydroxybutyrate granules. In these nodules, the host
cytoplasm was homogeneous and very electron dense. In
addition, the bacteroids had proliferated to the extent
that most of the host cytoplasm was occupied by them
(Fig. 5f). However, infected cells in nodules induced by
the strain TL16 appeared to degenerate shortly after the
bacteroids were formed. The infection tubes were seen
containing lysed bacteria. Most of the bacteria failed to
transform into bacteroids. Moreover, amyloplasts, mito-
chondria, endoplasmic reticulum and the remnants of
other organelles and membranes were found in the host
cell cytoplasm (Fig. 5 g–i).
Discussion
Cicer arietinum is the third most widely grown grain
legume in the world, and is grown extensively in the
Middle East and many regions of India (van der Maesen
1972). It is a tropical legume and is nodulated by
M. ciceri. To study the genetic basis of Dct in M. ciceri
and its implications in symbiotic performance of the
bacteria, the mutants were raised by Tn5 mutagenesis of
the wild-type strain TAL620. Putative dicarboxylate
mutants were then selected as white colonies on plates
containing glucose and succinate as carbon sources
and 2,3,5-triphenyl tetrazolium chloride. Dicarboxylate
uptake studies with one of the mutants, TL16, con-
firmed the inability of the mutant to transport dicarb-
oxylates, indicating the absence of a functional Dct in
the strain. Although the mutant fails to grow on dicarb-
oxylates, its growth similar to that of the wild type on
other carbon sources, such as pyruvate, glucose, galac-
tose, sucrose, ribose, mannitol, citrate, arabinose or
ribose, indicates that it has a functional tricarboxylic
acid cycle (Bolton et al. 1986). The absence of the activ-
ity of the enzymes that use C4-dicarboxylate as substrate
M (b)kb
10·9
3·2
23·0
4·3
9·4
6·5
M(a)
kb
321 21
Figure 4 Southern hybridization analysis. (a) localization of the site
of Tn5 insertion in Mesorhizobium ciceri mutant strain TL16. Lane 1,
strain TAL620; lane 2, TL16. (b), rpoN containing DNA sequence in
TAL620 (wild type). Lane 1, TL16; lane 2, strain TAL620; and lane 3,
plasmid pUS1.
Mesorhizobium ciceri rpoN mutant U.S. Gautam et al.
1804 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807
ª 2007 The Authors
in the extract of the mutant indicates inducible nature
of these enzymes in M. ciceri.
Cloning and sequencing of the DNA fragment con-
taining the transposon revealed that Tn5 had inter-
rupted the gene for RpoN which is an alternative
sigma factor. A single band as observed following
Southern hybridization of the wild-type DNA with rpoN
probe suggests the presence of only a single copy of
rpoN in M. ciceri TAL620. This is in contrast to M. loti
strain MAFF303099, R. etli and B. japonicum, where
two copies of rpoN gene (rpoN1 and rpoN2) were
observed (Kullik et al. 1991; Michiels et al. 1998a;
Kaneko et al. 2000; Dombrecht et al. 2002) and also
A. caulinodans, where the presence of two copies of
rpoN were conjectured (Stigter et al. 1993). Thus, the
mutant phenotype of M. ciceri TL16 is attributed to
the inactivation of the rpoN gene by Tn5 insertion on
a single site in the chromosomal DNA. Genomic con-
text study reveals that the rpoN gene in M. ciceri
has an organizational conservation identical to that of
M. loti and largely similar to that of B. japonicum and
R. etli.
The white nodules incited by the mutant strain
M. ciceri TL16 indicated that these were ineffective, and
indeed, the pale green colour of the hosts pointed to a
lack of significant nitrogen fixation by the bacteroids in
their nodules. This was further confirmed by the failure
of acetylene reduction by the nodules induced by the
mutant M. ciceri TL16, which also resulted in reduced
dry weight of shoots and not significantly different
from that of the uninoculated control plants. Thus, it
appears that rpoN function is required for symbiotic
nitrogen fixation in M. ciceri. The presence of a large
number of empty, uninfected cells in the symbiotic
0·5 µm
b
pbmvs
ermt
am
sb
lbit
0·1
0·1 µm
phb
sbb
mt
0·6 µm
0·3 µm
0·3 µm
b
ncs
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 5 Morphology of nodules formed on chickpea roots by wild-type Mesorhizobium ciceri (TAL620) and rpoN mutant TL16. Nodules shown
were photographed at 35 days of inoculation with: (a) Mesorhizobium ciceri TAL620; (b) rpoN- mutant TL16; (c) uninoculated control. Light micro-
graphs of sections of the symbiotic zones of nodules elicited by: (d) TAL620 (wild type); (e) mutant TL16. b, bacteroids; s, starch granule; nc, nod-
ule cortex. Electron microscopic sections of nodules elicited by: (f) TAL620 (wild type); (g), (h) and (i) mutant TL16. vs, gas exchange vesicle; b,
bacteroids; mt, mitochondria; am, amyloplast; er, endoplasmic reticulum; pbm, peribacteroid membrane; sb, senescent bacteroid; phb, granules
of polyhydroxybutyrate; it, infection tube; lb, lysed bacteria.
U.S. Gautam et al. Mesorhizobium ciceri rpoN mutant
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 1805

zone of nodules formed by the mutant M. ciceri TL16
and the failure of infecting bacteria to transform into
bacteroids indicate the role of rpoN function in the
maturation of bacteroids and subsequent nitrogen
fixation by them.
Acknowledgements
We are grateful to Prof F.M. Ausubel (Department of
Genetics, Harvard Medical School, Boston, Massachusetts,
USA) for providing the cosmid pLAFR1. We thank
Prof D.H. Figurski (Department of Biology, University of
California, San Diego, La Jolla, USA) for providing the
strain pRK2013. We are grateful to All India Institute of
Medical Sciences, New Delhi, for Electron Microscopic
Facility. We are thankful to the Distributed Information
Sub Center at Institute of Life Sciences, Bhubaneswar,
for making the genetic map of the rpoN gene sequences.
We are grateful to the Dept. of Biotechnology, Govt. of
India, for financial assistance.
References
Albright, L.M., Ronson, C.W., Nixon, B.T. and Ausubel, F.M.
(1989) Identification of a gene linked to Rhizobium meliloti
ntrA whose product is homologous to a family to ATP-
binding proteins. J Bacteriol 171, 1932–1941.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W. and Lipman,
D.J. (1990) Basic local alignment search tool. J Mol Biol
215, 403–410.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J.H.,
Zhang, Z., Miller, W. and Lipman, D.J. (1997) Gapped
BLAST and PSI-BLAST: a new generation of protein
database search program. Nucleic Acids Res 25,
3389–3402.
Bolton, E., Higgisson, B., Harrington, A. and Gara, F.O.
(1986) Dicarboxylic acid transport in Rhizobium meliloti:
isolation of mutants and cloning of dicarboxylic acid
transport genes. Arch Microbiol 144, 142–146.
Bradford, M.M. (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem 72,
248–254.
Ciria, R., Abreu-Goodger, C., Morett, E. and Merino, E.
(2004) GeConT: gene context analysis. Bioinformatics 14,
2307–2308.
Das, S.K. and Mishra, A.K. (1996) Transposon mutagenesis
affecting thiosulfate oxidation in Bosea thiooxidans, a new
chemolithoheterotrophic bacterium. J Bacteriol 178,
3628–3633.
Das, S.K., Gautam, U.S., Chakrabartty, P.K. and Singh, A.
(2006) Characterization of a symbiotically defective serine
auxotroph of Mesorhizobium ciceri. FEMS Microbiol Lett
263, 244–251.
Dixon, R. (1987) Genetic regulation of nitrogen fixation. In
The Nitrogen and Sulphur Cycles Ed. Cole, J.A. and Fergu-
son, S.J. pp. 417–438. Cambridge: Cambridge University
Press (SGM Symposium 42).
Dombrecht, B., Marchal, K., Vanderleyden, J. and Michiels, J.
(2002) Prediction and overview of the RpoN-regulon in
closely related species of the Rhizobiales. Genome Biol 3,
0076.1–0076.11.
Figurski, D.H. and Helinski, D.R. (1979) Replication of an
origin-containing derivative of plasmid RK2 dependent on
a plasmid function provided in trans. Proc Natl Acad Sci
USA 76, 1648–1652.
Finan, T.M., Wood, J.M. and Jordan, D.C. (1983) Symbiotic
properties of C4-dicarboxylic acid transport mutants of
Rhizobium leguminosarum. J Bacteriol 154, 1403–1413.
Friedman, A.M., Long, S.R., Brown, S.E., Buikema, W.J. and
Ausubel, F.M. (1982) Construction of a broad host range
cosmid cloning vector and its use in the genetic analysis of
Rhizobium mutants. Gene 18, 289–296.
Hill, R.L. and Bradshaw, R.A. (1969) Fumarase. Methods Enzy-
mol 13, 91–99.
Jiang, J., Gu, B.H., Albright, L.M. and Nixon, B.T. (1989)
Conservation between coding and regulatory elements of
Rhizobium meliloti and Rhizobium leguminosarum dct
genes. J Bacteriol 171, 5244–5253.
Kaneko, T., Nakamura, Y., Sato, S., Asamizu, E., Kato, T.,
Sasamoto, S., Watanabe, A., Idesawa, K. and 16 other
authors (2000) Complete genome structure of the nitro-
gen-fixing symbiotic bacterium Mesorhizobium loti. DNA
Res 31, 331–338.
Karnovsky, M.J. (1965) A formaldehyde-glutaraldehyde fixative
of high osmolarity for use in electron microscopy. J Cell
Biol 27, 137–141.
Kullik, I., Fritsche, S., Knobel, H., Sanjuan, J., Hennecke, H.
and Fischer, H.M. (1991) Bradyrhizobium japonicum has
two differentially regulated, functional homologs of the
sigma 54 gene (rpoN). J Bacteriol 173, 1125–1138.
Kustu, S., Santero, E., Keener, J., Popham, D. and Weiss, D.
(1989) Expression of sigma 54 (ntrA)-dependent genes is
probably united by a common mechanism. Microbiol Rev
53, 367–376.
Meade, H.M., Long, S.R., Ruvkun, G.B., Brown, S.E. and
Ausubel, F.M. (1982) Physical and genetic characterization
of symbiotic and auxotrophic mutants of Rhizobium melil-
oti induced by transposon Tn5 mutagenesis. J Bacteriol
149, 114–122.
Merrick, M.J. (1993) In a class of its own – the RNA polym-
erase sigma factor r54 (rN). Mol Microbiol 10, 903–909.
Michiels, J., Moris, M., Dombrecht, B., Verreth, C. and Van-
derleyden, J. (1998a) Differential regulation of Rhizobium
etli rpoN2 gene expression during symbiosis and free-living
growth. J Bacteriol 180, 3620–3628.
Michiels, J., van Soom, T., D’hooghe, I., Dombrecht, B.,
Benhassine, T., de Wilde, P. and Vanderleyden, J. (1998b)
The Rhizobium etli rpoN locus: DNA sequence analysis and
Mesorhizobium ciceri rpoN mutant U.S. Gautam et al.
1806 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807
ª 2007 The Authors

phenotypical characterization of rpoN, ptsN, and ptsA
mutants. J Bacteriol 180, 1729–1740.
Miller, J.H. (1972) Experiments in Molecular Genetics. Cold
Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Raychaudhuri, N., Das, S.K. and Chakrabartty, P.K. (2005) Sym-
biotic effectiveness of a siderophore over producing mutant
of Mesorhizobium ciceri. Polish J Microbiol 54, 37–41.
Reynolds, E.S. (1963) The use of lead citrate at high pH as an
electron-opaque stain in electron microscopy. J Cell Biol
17, 208–212.
Ronson, C.W. and Primrose, S.B. (1979) Carbohydrate meta-
bolism in Rhizobium trifolii: identification and symbiotic
properties of mutants. J Gen Microbiol 112, 77–88.
Ronson, C.W., Lyttleton, P. and Robertson, J.G. (1981) C(4)-
dicarboxylate transport mutants of Rhizobium trifolii form
ineffective nodules on Trifolium repens. Proc Natl Acad Sci
USA 78, 4284–4288.
Ronson, C.W., Nixon, B.T., Albright, L.M. and Ausubel, F.M.
(1987) Rhizobium meliloti ntrA (rpoN) gene is required for
diverse metabolic functions. J Bacteriol 169, 2424–2431.
Sambrook, J., Fritsch, E.F. and Maniatis, T.A. (1989) Molecular
Cloning: A Laboratory Manual 2nd edn. Cold Spring
Harbor, NY: Cold Spring Harbor Laboratory Press.
Simon, R. (1984) High frequency mobilization of gram-negat-
ive bacterial replicons by the in vivo constructed Tn5-mob
transposon. Mol Gen Genet 196, 413–420.
Simon, R., Priefer, U. and Puhler, A. (1983) A broad host
range mobilization system for in vivo genetic engineering:
transposon mutagenesis in gram-negative bacteria.
Bio ⁄ Technol 1, 784–790.
Southern, E.M. (1975) Detection of specific sequences among
DNA fragments separated by gel electrophoresis. J Mol Biol
98, 503–517.
Stigter, J., Schneider, M. and de Bruijn, F.J. (1993) Azorhizobi-
um caulinodans nitrogen fixation (nif ⁄ fix) gene regulation:
mutagenesis of the nifA -24 ⁄ -12 promoter element, charac-
terization of a ntrA(rpoN) gene, and derivation of a
model. Mol Plant Microbe Interact 6, 238–252.
Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLU-
STAL W: improving the sensitivity of progressive multiple
sequence alignment through sequence weighting, position
specific gap penalties and weight matrix choice. Nucleic
Acids Res 22, 4673–4680.
Van der Maesen, L.G.J. (1972) Cicer L., a monograph of the
genus with reference to the chickpea (Cicer arietinum L.): its
ecology and cultivation. Medellingen Landbouwhogeshool
Wageningen, Nederland: Publication No. 72-10, Veenmen
and Zonen, Wageningen.
Van Slooten, J.C., Cervantes, E., Broughton, W.J., Wong,
C.H. and Stanley, J. (1990) Sequence and analysis of the
rpoN sigma factor gene of Rhizobium sp. strain
NGR234, a primary coregulator of symbiosis. J Bacteriol
172, 5563–5574.
Veeger, C., Dervartanian, D.V. and Zeylemaker, W.P. (1969)
Succinate dehydrogenase. Methods Enzymol 13, 81–90.
Vincent, J.M. (1970) A Manual for the Practical Study of the
Root Nodule Bacteria. I.B.P. Handbook No. 15. Oxford:
Blackwell.
Yoshida, A. (1969) L-Malate dehydrogenase from Bacillus sub-
tilis. Methods Enzymol 13, 141–145.
U.S. Gautam et al. Mesorhizobium ciceri rpoN mutant
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 1798–1807 1807




















