
Magnetic and magneto optic properties of substituted polythiophenes
Upload
independentCategory
view
1download
0
Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123
www.elsevier.com/locate/jphotobiol
Role of LHCII organization in the interaction of substituted1,4-anthraquinones with thylakoid membranes
Emilia Apostolova a,*, Sashka Krumova a, Tzvetelina Markova a, Tzvetanka Filipova b,Maria T. Molina c, Ivana Petkanchin d, Stefka G. Taneva a
a Institute of Biophysics, Bulgarian Academy of Sciences, Acad. G. Bonchev Str., B1.21, Sofia 1113, Bulgariab Department of Organic Chemistry, University of Chemical Technology and Metallurgy, Blv. Kl. Ohridski 8, Sofia, Bulgaria
c Insituto de Quımica Medica, C.S.I.C., Calle Juan de la Cierva 3, 28006 Madrid, Spaind Institute of Physical Chemistry, Bulgarian Academy of Sciences, Sofia 1113, Bulgaria
Received 18 June 2004; received in revised form 4 October 2004; accepted 6 October 2004
Available online 2 December 2004
Abstract
The chlorophyll fluorescence, photochemical activity and surface electric properties of thylakoid membranes with different stoi-
chiometry of pigment–protein complexes and organization of the light-harvesting chlorophyll a/b protein complex of photosystem II
(LHCII) were studied in the presence of substituted 1,4-anthraquinones. Data show strong dependence of the quenching of the chlo-
rophyll fluorescence on the structural organization of LHCII. The increase of the LHCII oligomerization, which is associated with
significant reduction of the transmembrane electric charge asymmetry and electric polarizability of the membrane, correlates with
enhanced quenching effect of substituted 1,4-athraquinones. Crucial for the large quinone-induced changes in the membrane electric
dipole moments is the structure of the quinone molecule. The strongest reduction in the values of the dipole moments is observed
after interaction of thylakoids with 3-chloro-9-hydroxy-1,4-anthraquinone (TF33) which has the highest quenching efficiency. The
quinone induced changes in the photochemical activity of photosystem II (PSII) correlate with the total amount of the supramo-
lecular LHCII–PSII complex and depend on the number of substituents in the 1,4-anthraquinone molecule.
� 2004 Elsevier B.V. All rights reserved.
Keywords: Chlorophyll fluorescence; Thylakoid membranes; Light-harvesting chlorophyll a/b protein complex; 1,4-Anthraquinones; Photochemical
activity; Surface electric properties
1. Introduction
The light reactions of photosynthesis are driven by
the excitation energy absorbed by the antenna pigments
and transferred to the reaction centers. High degree oforganization of the pigment–protein complexes is re-
quired for efficient utilization of the solar energy by pho-
tosynthetic organisms. Quinones play an important role
as electron and proton carriers in the electron-transport
1011-1344/$ - see front matter � 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jphotobiol.2004.10.002
* Corresponding author. Tel.: +359 2 979 26 21; fax: +359 2 971 24
93.
E-mail address: [email protected] (E. Apostolova).
system [1,2]. Exogenous quinones are able to compete
with plastoquinone for electrons and quench the fluores-
cence from antenna chlorophyll in chloroplasts, algae
and green bacteria [3–11]. They can be partially or com-
pletely reduced under illumination with loss of theirability to act as quenchers [4]. As an analog of the pho-
tosystem II (PSII) plastoquinone electron acceptor, QB,
substituted quinones compete with QB for a common
binding domain and thereby inhibit the QB function
[12].
In our previous study we found that the quenching of
the chlorophyll fluorescence by exogenous 1,4-anthra-
quinones (1,4-AQ) in thylakoid membranes is due to

116 E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123
interaction of quinones with chlorophyll molecules and
that the quenching efficiency of halogen-substituted
1,4-anthraquinones strongly depends on the position
of the Cl atom [13]. The present investigation is focussed
on the contribution of light-harvesting chlorophyll a/b
protein complex of PSII (LHCII) in the interaction ofthylakoid membranes with 1,4-AQ. The quenching effect
of 1,4-AQ was determined in thylakoid membranes with
different stoichiometry of the chlorophyll–protein com-
plex and structural organization of LHCII. Combining
the fluorescence with the electro-optic data demon-
strates a relationship between the quinone-induced
changes in the chlorophyll fluorescence, the electric
properties and the LHCII amount and its oligomeriza-tion state.
2. Materials and methods
2.1. Substituted 1,4-anthraquinones
In the present study were used 1,4-anthraquinone(QA6) and substituted 1,4-anthraquinones: 9-acetoxy-
1,4-anthraquinone (QA4); 9-hydroxy-1,4-anthraquinone
(QA5); 9-methoxy-1,4-anthraquinone (QA7); 10-chloro-
9-hydroxy-1,4-anthraquinone (TF11); 9-acetoxy-3-bro-
mo-1,4-anthraquinone (TF23); 9-acetoxy-3-chloro-1,4-
anthraquinone (TF29); 3-chloro-9-hydroxy-1,4-anthra-
quinone (TF33); 3-bromo-9-hydroxy-1,4-anthraquinone
(TF34). The substituted 1,4-anthraquinones were syn-thesized as in [13]. The type of the substituent groups,
R3, R9 and R10, for the different substituted 1,4-anthra-
quinones (1,4-AQ) is given in Table 2.
2.2. Thylakoid membranes
Thylakoid membranes were isolated from pea (Pisum
sativum L. cv. Borec and its mutants Coeruleovireus 2/16and Costata 2/133 and P. sativum L. cv. Auralia and its
mutant Chlorotica XV/1422) and barley (Hordeum vulg-
are L. and the chlorophyll b-deficient barley mutant
Chlorina f2) as described in [14] and [15], respectively.
The total chlorophyll concentration was determined by
the method of Lichtentaler [16].
2.3. Chlorophyll fluorescence
Chlorophyll fluorescence at room temperature in the
presence of 1,4-anthraquinone and substituted 1,4-
anthraquinones was measured with Jobin–Yvon JY3
spectrofluorimeter as in [13]. The quinone concentration
was 200 lM. The chlorophyll fluorescence was excited at
480 nm. The thylakoid membranes were suspended in 50
mM HEPES–NaOH (pH 7.5), 0.1 M sucrose, 5 mMNaCl and 20 lM DCMU. The chlorophyll concentra-
tion was 10 lg chl/ml. Before the measurements, thyla-
koid membranes were incubated with 1,4-AQ for 15
min.
The Stern–Volmer quenching constant (KSV) was
used as a characteristic of the quenching effect of the
quinones on the chlorophyll fluorescence [7]:
F m=F 0m � 1 ¼ Ksv½CQ�, ð1Þ
where Fm and F 0m are the fluorescence intensities in ab-
sence and presence of quinone, respectively; CQ is the
quinone concentration.
2.4. Photochemical activity of PS II
The photochemical activity of PSII was measured
polarographically with a Clark-type electrode at 20 �C,with 1,4-benzoquinone as electron acceptor as in [13].
The concentration of 1,4-anthraquinones was 200 lM.
2.5. Electric light scattering
The electric polarizability (ci) and the permanent di-
pole moment (p^) were estimated from the dependence
of the electro-optic effect on the frequency (m) of the ap-plied a.c. field for ellipsoid particles [17]. The electro-op-
tic effect a = (IE�I0)/I0 was defined as a relative change
in the intensity of the scattered light. IE and I0 are the
intensities of the light scattered by the membrane sus-pension when an external electric field of strength E is
applied and without field, respectively. At a low degree
of orientation (i.e., U/kT�1, U is the energy of orienta-
tion, kT is the thermal energy) is expressed by the
equation:
a ¼ AðKl,KdÞI0ðKl,KdÞ
ðb2 þ cÞE2, ð2Þ
where A(Kl,Kd) and I0(Kl,Kd) are optical functions
dependent on the dimensions of the particles and the
geometry of the experimental setup; K = (2p/k 0)sin2(h/2); k 0 is the wavelength of the light in the suspension,
h is the angle of observation (90� in this case); l and d
are the long and short axis of the ellipsoid particles.
b = p^/kT and c = (ci�c^)/kT, where p^ is the permanent
dipole moment arising from the asymmetric charge dis-tribution on the membrane surfaces, i.e., along the par-
ticle symmetry axis; ci and c^ are the electric
polarizabilities along and perpendicular to the mem-
brane surface, respectively; k is the Boltzmann constant
and T is the absolute temperature.
The dimensions of the membrane particles were esti-
mated from the relaxation time of particle disorientation
(at high degree of orientation) after switching off theelectric field as described in [18]. The surface area, S,
of the thylakoid membranes was calculated assuming
that they are prolate ellipsoids with axial ratio 2 [18].
The measuring set-up is described in detail elsewhere
[19]. Electric field pulses were applied to the suspension

E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123 117
in an electro-optical cell (optical horn) with horizontal
platinum electrodes at a fixed distance of 2.6 mm. Elec-
tric pulse generator (Waveteck 185) and power-ampli-
filer (Model PA25, Optimatium, JRL, New York) were
used for low and high intensity electric field pulses,
respectively. The light source was a halogen lamp. Thetransient signal was detected by a photomultiplier
(FQS 35, Zeiss, Yena), the output signal recorded on a
digital storage oscilloscope (Philips PM 3335) and ana-
lyzed with the aid of a personal computer.
The electro-optic measurements were carried out in a
medium containing 0.25 mM sorbitol (pH 5.3–5.8 and
conductivity 4.56 lS/cm). The chlorophyll concentration
was 5 lg chl/ml.The data of the electric dipole moments are mean val-
ues from 4 to 5 independent measurements.
3. Results
3.1. Characteristics of the thylakoid membranes
In this work thylakoid membranes from pea wild
types Borec and Auralia, two mutants of the variety
Borec (Coeruleovireus 2/16 and Costata /133), one mu-
tant of the variety Auralia (Chlorotica XV/1422), barley
wild type and the chlb-deficient mutant of barley Chlo-
rina f2 were studied. The composition and electric prop-
erties of thylakoid membranes from these higher plants
were characterized in our previous investigation [18,20].It was determined that the total amount of both the
LHCII and the supramolecular complex of LHCII–PSII
in thylakoid membranes from Auralia wt and its mutant
Chlorotica XV/1422 is smaller than those from Borec wt
and its mutants Costata 2/133 and Coeruleovireus 2/16
(Table 1; [18,20]). Besides, the organization of LHCII
(i.e., the ratio of oligomeric to monomeric form of
LHCII (LHCIIo/LHCIIm)) varies with the plant type(Table 1; [24,25]). It was also shown that the increase
of the ratio LHCIIo/LHCIIm strictly correlates with
the decrease in the values of membrane electric dipole
moments (electric polarizability (ci) and permanent di-
Table 1
Relative amount of the chlorophyll-protein complexes of pea thylakoid memb
XV/1422; Costata 2/133 and Coeruleovireus 2/16) determined by nondenatura
form; LHCIIm – monomeric form) and PSII (protein containing P680 and a
Plant type LHCII
Chloroticaa XV/1422 33.90 ± 0.29
Auraliaa wt 34.12 ± 0.49
Costatab 2/133 47.03 ± 0.39
Borecb wt 45.86 ± 0.79
Coeruleovireusb 2/16 45.25 ± 1.86
The data are taken from our previous papers [18,20].a Data from [20].b Data from [18].
pole moment (p^)) (data for the electric moments with-
out 1,4-AQ are given in Table 4 and [18,20]).
Chlorina f2 barley mutant is characterized by strongly
reduced pigment content, reduced antenna size of the
PSII and lack of the oligomeric structure of LHCII as
compared to the wild type [15,21].
3.2. Chlorophyll fluorescence of thylakoid membranes
from different plant species in presence of
1,4-anthraqinones
The interaction of 1,4-AQ with thylakoid membranes
from the different plant species results in a strong de-
crease in the intensity of the chlorophyll emission at686 nm without change in the position of the emission
maximum. Stern–Volmer quenching constants (Ksv) for
the 686 nm emission band are shown on Table 2. The
Ksv values of the 1,4-AQ depend on the organization
of thylakoid membranes. KSV for thylakoid membranes
from Auralia wt and its mutant (Chlorotica XV/1422)
are smaller than those from Borec wt and its mutants
(Costata 2/133 and Coeruleovireus 2/16) (Table 2). Theresults clearly show that the quenching effect of 1,4-
AQ is bigger in the membranes containing higher
amount of LHCII (Tables 1 and 2). Similar tendency
is observed for the quenching efficiency of 1,4-AQ in
thylakoids from barley and Chlorina f2 mutant. The
constants (Ksv) of 1,4-AQ for the mutant thylakoids
are smaller than those for barley wild type (Fig. 1).
The increase of the quenching constant with the in-crease of the amount of LHCII is more pronounced
for mono-substituted 1,4-anthraquinone and 3-halo-
gen-9-acetoxy-1,4-anthraquinones than for halogen
substituted 9-hydroxy-1,4-anthraquinones (Table 2).
Comparison of the chlore substituted 9-hydroxy-1,4-
anthraquinones (TF33 and TF11) shows that the posi-
tion of the chlore atom strongly affects the quenching
efficiency of the quinone in all studied thylakoid mem-branes. Independent of the plant type the biggest
quenching effect is found in the presence of 3-chloro-9-
hydroxy-1,4-anthraquinone (TF33). The same is true
for thylakoids from barley wild type and its mutant
ranes from wild types (Auralia and Borec) and the mutants(Chlorotica
ting SDS–PAGE: LHCII (LHCII – total content; LHCIIo – oligomeric
ntenna complex) thylakoid membranes
LHCII + PSII LHCPIIo/LHCPIIm
51.10 ± 0.56 2.45
47.24 ± 0.69 2.82
59.73 ± 0.42 3.34
59.46 ± 0.86 4.57
59.35 ± 1.54 6.62

Table 2
Stern–Volmer quenching constants (KSV) for 686 nm room-temperature chlorophyll fluorescence of thylakoid membranes from Auralia wt, Borec wt
and the mutants Chlorotica XV/1422, Costata 2/133 and Coeruleovireus 2/16 incubated with 1,4-anthraquinone and substituted 1,4-anthraquinones
1,4-AQ Substituent group Chlorotica XV/1422
KSV [M�1]
Auralia wt
KSV [M�1]
Costata 2/133
KSV [M�1]
Borec wt
KSV [M�1]
Coerul. 2/16
KSV [M�1]R3 R9 R10
TF23 Br OCOCH3 H 5.30 · 103 5.15 · 103 6.50 · 103 11.60 · 103 15.90 · 103
TF11 H OH Cl 10.70 · 103 13.90. · 103 15.40 · 103 14.10 · 103 18.00 · 103
TF29 Cl OCOCH3 H 21.70 · 103 23.50 · 103 34.23 · 103 34.20 · 103 32.50 · 103
TF34 Br OH H 42.90 · 103 44.20 · 103 49.70 · 103 48.90 · 103 64.30 · 103
TF33 Cl OH H 121.10 · 103 127.10 · 103 149.37 · 103 180.44 · 103 152.7 · 103
QA7 H OCH3 H 4.40 · 103 5.10 · 103 4.60 · 103 7.14 · 103 8.50 · 103
QA6 H H H 11.40 · 103 10.6 · 103 13.20 · 103 17.00 · 103 25.20 · 103
QA5 H OH H 13.30 · 103 17.90 · 103 24.50 · 103 28.30 · 103 29.60 · 103
QA4 H OCOCH3 H 30.90 · 103 36.70 · 103 46.30 · 103 49.30 · 103 48.40 · 103
R3, R9, R10 are the substituent groups. The data are mean values from 5 to 7 independent experiments. The standard errors are from ±0.03 · 103 to
±0.09 · 103.
2
4
6
8
10
1,4-anthraquinones
TF33
QA4
QA5QA6TF29
KSV
x10-3
[M-1]
Fig. 1. Stern–Volmer quenching constants (KSV) for 1,4-anthraqui-
none, QA6, and the substituted 1,4-anthraquinones, QA4, QA5, TF29
and TF33, in thylakoid membranes from barley wt (�) and Chlorina f2
mutant (�). Fluorescence emission bands: for barley wt at 685 nm and
for Chlorina f2 mutant at 695 nm, respectively. The concentration of
1,4-AQ in the reaction medium is 200 lM. The parameter Fm/F0m
increases linearly with increasing the quinone concentration to 250 lM.
118 E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123
Chlorina f2 (Fig. 1). These results concerning the
quenching efficiency of 1,4-AQ in thylakoid membranes
with different organization from a number of plant types
studied in this work confirm our previous data for an-
other variety of pea [13].
Besides the amount of the LHCII, its organizationalso affects the degree of quenching. Ksv has bigger val-
ues for membranes with higher degree of LHCII oligo-
merization (Table 2). The smallest Ksv values for all
1,4-AQ, with the exception of 1,4-anthraquinone
(QA6) and of 3-bromo-9-acetoxy-1,4-anthraquinone
(TF23), are observed for thylakoid membranes from
the mutant Chlorotica XV/1422, which is characterized
by the lowest oligomer to monomer LHCII ratio. Thesmallest Ksv values for QA6 and TF23 are found for
Auralia wt, which has a slightly bigger LHCIIo/LHCIImratio than that of Chlorotica XV/1422. Therefore, the
higher ratio of oligomeric to monomeric LHCII
(LHCIIo/LHCIIm) is related to bigger quenching effi-
ciency of the quinones (Table 2). The observation that
Ksv has significantly higher value for barley wild type
than for Chlorina f2 thylakoid membranes (Fig. 1) sup-
ports this finding.
3.3. Photochemical activity of thylakoid membranes in the
presence of 1,4-anthraquinones
1,4-Anthraquinones affect significantly the photo-
chemical activity of the thylakoid membranes as well
(Table 3).
In the presence of all mono-substituted 1,4-AQ (QA4,
QA5, QA6, QA7) the photochemical activity (A/A0) of
thylakoid membranes from Borec wild type and its mu-tants (Costata 2/133 and Coeruleovireus 2/16) is bigger
than that of thylakoids from Auralia wild type and
Chlorotica XV/1422 (Table 3). The opposite effect is ob-
served for the di-substituted 1,4-AQ (TF11, TF23,
TF29, TF33, TF34) (Table 3). The strongest inhibition
of the PSII activity was observed in the presence of
chlore substituted 1,4-anthraquinones. Therefore, the
degree of inhibition of the PSII activity by mono- anddi-substituted 1,4-anthraquinones depends on the
amount of the supramolecular complex LHCII–PSII,
which is smaller in thylakoids from Auralia wt and its
mutant than that from Borec wt and its mutants (Table
1). Differences in the structural organization of this
complex might be responsible for the different extent
of contacts between PSII and mono- and di-substituted
1,4-AQ.
3.4. Surface electric properties of thylakoid membranes in
the presence of 1,4-anthraquinones
As mentioned above, there is a correlation between
the LHCIIo/LHCIIm ratio (Table 1) and the values of
the permanent and induced dipole moments (Table 4).

Table 3
Photochemical activities of PS II (A/A0) of pea thylakoid membranes with different organization in presence of 200 lM of 1,4-anthraquionone and
substituted 1,4-anthraquinones
1,4-AQ Substituent group Chlorotica XV/1422 Auralia wt Costata 2/133 Borec wt Coerul. 2/16
R3 R9 R10
TF11 H OH Cl 0.496 0.486 0.453 0.382 0.404
TF33 Cl OH H 0.502 0.493 0.425 0.467 0.451
TF29 Cl OCOCH3 H 0.539 0.569 0.461 0.474 0.485
TF23 Br OCOCH3 H 0.583 0.585 0.598 0.512 0.535
TF34 Br OCOCH3 H 0.605 0.599 0.590 0.545 0.526
QA4 H OCOCH3 H 0.610 0.591 0.686 0.736 0.608
QA5 H OH H 0.657 0.669 0.694 0.848 0.813
QA6 H H H 0.795 0.826 0.833 0.879 0.843
QA7 H OCH3 H 0.835 0.840 0.898 0.896 0.897
A and A0 are the photochemical activities with and without 1,4-AQ, respectively. The data are mean values from 5 to 7 independent experiments. The
standard errors are from ±0.034 to ±0.069.
Table 4
Values of the permanent dipole moment (p^) and the electric polarizability per unit surface area (ci/S) of thylakoid membranes from pea wild types
(Borec and Auralia) and the mutants (Chlorotica XV/1422, Costata 2/133 and Coeruleovireus 2/16) in presence of three selected substituted 1,4-
anthraquinones
1,4-AQ Chlorotica XV/1422 Auralia wt Costata 2/133 Borec wt Coeruleovireus 2/16
p^ · 1024 [Cm]
Without 0.613 0.582 0.446 0.350 0.270
QA5 0.662 0.856 0.691 0.525 0.651
TF11 1.018 0.656 0.571 0.616 0.327
TF33 0.340 0.395 0.268 0.153 0.205
ci/S · 1016 [F]
Without 1.287 0.853 0.730 0.380 0.280
QA5 1.596 2.039 1.905 0.749 0.549
TF11 3.501 0.876 0.947 0.669 0.449
TF33 0.580 0.620 0.504 0.315 0.173
The standard errors are: for p^ – from ±0.031 · 10�24 to ±0.058 · 10�24 and for ci/S are from ±0.072 · 10�16 to ±0.11 · 10�16.
E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123 119
A detailed analysis of the orientational behaviour of
thylakoid membranes in an external electric field before
and after interaction with 1,4-AQ shows strong varia-
tion of p^ and ci (data are shown for the di-substituted1,4-anthraquinones TF11, TF33 and the mono-substi-
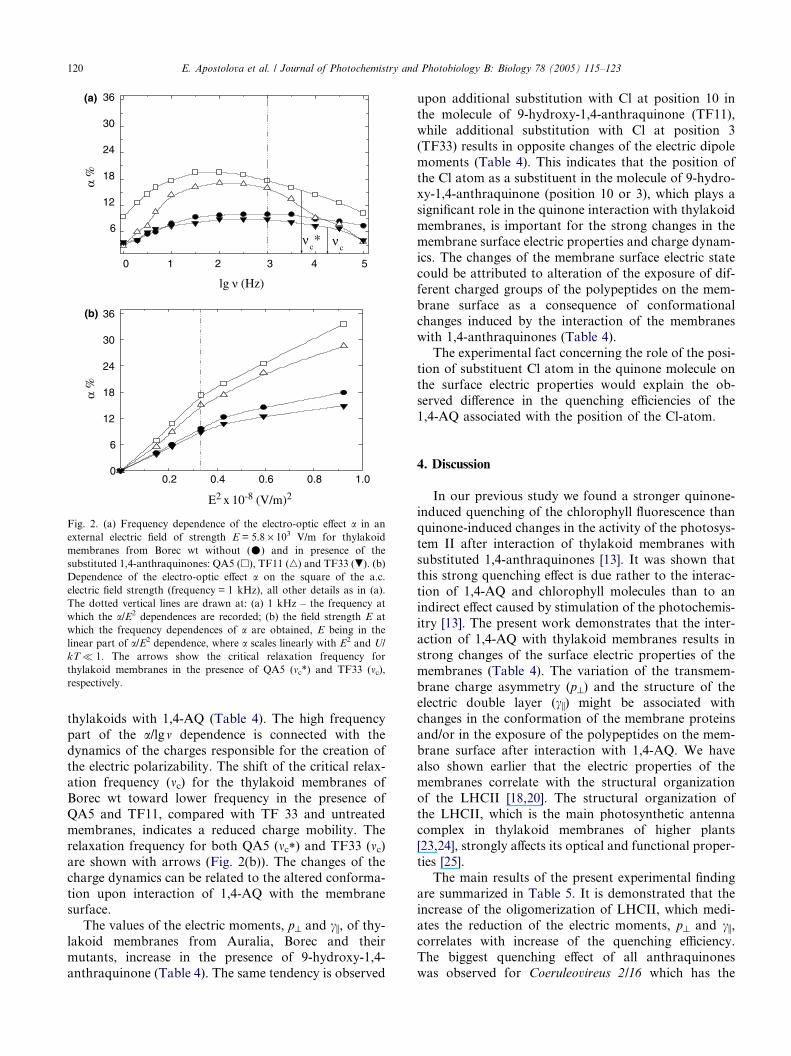
tuted QA5; Fig. 2).
The orientational behaviour of thylakoid membranes
in an external a.c. electric field is determined by their
permanent and induced electric dipole moments (details
on the electro-optic phenomena can be found in Ref.
[17]). The dependence of the electro-optic effect a on
the frequency of the applied a.c. electric pulses, a/lgm,gives information about both electric moments (Fig.
2(a)), as well as on the dynamics of the charges respon-
sible for the creation of the induced dipole moment. At
low frequencies (below the plateau of the frequency
dependence) the effect a decreases with the frequency de-
crease (Fig. 2(a)), which is usually related to the pres-
ence of a permanent dipole moment along the minor
axis of the particles and to orientation dominated bythe permanent dipole [22]. The vertical line shown in
Fig. 2 (a) denotes the frequency at which the a/E2 depen-
dences (Fig. 2(b)) are recorded and that in Fig. 2(b) the
electric field strength at which the frequency depen-
dences of a are obtained (Fig. 2(a)), E being in the linear
part of a/E2 dependence, where a scales linearly with E2
and U/kT�1.The electro-optic effect a of the thylakoid membranes
from Borec wt in presence of the mono-substituted 9-hy-
droxy-1,4-anthraquinone (QA5) is bigger than that
without quinone in the entire frequency range (Fig.
2(a)). On the contrary the magnitude of the effect a in
presence of di-substituted TF33 is reduced (Fig. 2(a)).
The difference in the magnitude of the effect a in the
kHz range, where the orientation of the thylakoid mem-branes is determined by the motion of charge(s) in the
diffuse electric double layer (i.e., by the electric polariz-
ability ci), corresponds to the differences in the initial
slopes of the a/E2 dependence (Fig. 2(b)) between the
thylakoid membranes without and with 1,4-AQ (results
shown for QA5, TF11, TF33; Fig. 2(b)). Therefore, the
interaction of 1,4-AQ with thylakoid membranes leads
to changes in the values of both dipole moments (Table4). This reflects the variation of the transmembrane
charge asymmetry and the ion composition of the dif-
fuse electric double layer related to the interaction of
0 1 2 3 4 5
νc
νc*
α %
α %
lg ν (Hz)
0.2 0.4 0.6 0.8 1.00
6
12
18
24
30
36
6
12
18
24
30
36
E2 x 10-8 (V/m)2
(a)
(b)
Fig. 2. (a) Frequency dependence of the electro-optic effect a in an
external electric field of strength E = 5.8 · 103 V/m for thylakoid
membranes from Borec wt without (�) and in presence of the
substituted 1,4-anthraquinones: QA5 (h), TF11 (n) and TF33 (.). (b)
Dependence of the electro-optic effect a on the square of the a.c.
electric field strength (frequency = 1 kHz), all other details as in (a).
The dotted vertical lines are drawn at: (a) 1 kHz – the frequency at
which the a/E2 dependences are recorded; (b) the field strength E at
which the frequency dependences of a are obtained, E being in the
linear part of a/E2 dependence, where a scales linearly with E2 and U/
kT� 1. The arrows show the critical relaxation frequency for
thylakoid membranes in the presence of QA5 (mc*) and TF33 (mc),respectively.
120 E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123
thylakoids with 1,4-AQ (Table 4). The high frequency
part of the a/lgm dependence is connected with the
dynamics of the charges responsible for the creation of
the electric polarizability. The shift of the critical relax-ation frequency (mc) for the thylakoid membranes of
Borec wt toward lower frequency in the presence of
QA5 and TF11, compared with TF 33 and untreated
membranes, indicates a reduced charge mobility. The
relaxation frequency for both QA5 (mc*) and TF33 (mc)are shown with arrows (Fig. 2(b)). The changes of the
charge dynamics can be related to the altered conforma-
tion upon interaction of 1,4-AQ with the membranesurface.
The values of the electric moments, p^ and ci, of thy-lakoid membranes from Auralia, Borec and their
mutants, increase in the presence of 9-hydroxy-1,4-
anthraquinone (Table 4). The same tendency is observed
upon additional substitution with Cl at position 10 in
the molecule of 9-hydroxy-1,4-anthraquinone (TF11),
while additional substitution with Cl at position 3
(TF33) results in opposite changes of the electric dipole
moments (Table 4). This indicates that the position of
the Cl atom as a substituent in the molecule of 9-hydro-xy-1,4-anthraquinone (position 10 or 3), which plays a
significant role in the quinone interaction with thylakoid
membranes, is important for the strong changes in the
membrane surface electric properties and charge dynam-
ics. The changes of the membrane surface electric state
could be attributed to alteration of the exposure of dif-
ferent charged groups of the polypeptides on the mem-
brane surface as a consequence of conformationalchanges induced by the interaction of the membranes
with 1,4-anthraquinones (Table 4).
The experimental fact concerning the role of the posi-
tion of substituent Cl atom in the quinone molecule on
the surface electric properties would explain the ob-
served difference in the quenching efficiencies of the
1,4-AQ associated with the position of the Cl-atom.
4. Discussion
In our previous study we found a stronger quinone-
induced quenching of the chlorophyll fluorescence than
quinone-induced changes in the activity of the photosys-
tem II after interaction of thylakoid membranes with
substituted 1,4-anthraquinones [13]. It was shown thatthis strong quenching effect is due rather to the interac-
tion of 1,4-AQ and chlorophyll molecules than to an
indirect effect caused by stimulation of the photochemis-
itry [13]. The present work demonstrates that the inter-
action of 1,4-AQ with thylakoid membranes results in
strong changes of the surface electric properties of the
membranes (Table 4). The variation of the transmem-
brane charge asymmetry (p^) and the structure of theelectric double layer (ci) might be associated with
changes in the conformation of the membrane proteins
and/or in the exposure of the polypeptides on the mem-
brane surface after interaction with 1,4-AQ. We have
also shown earlier that the electric properties of the
membranes correlate with the structural organization
of the LHCII [18,20]. The structural organization of
the LHCII, which is the main photosynthetic antennacomplex in thylakoid membranes of higher plants
[23,24], strongly affects its optical and functional proper-
ties [25].
The main results of the present experimental finding
are summarized in Table 5. It is demonstrated that the
increase of the oligomerization of LHCII, which medi-
ates the reduction of the electric moments, p^ and ci,correlates with increase of the quenching efficiency.The biggest quenching effect of all anthraquinones
was observed for Coeruleovireus 2/16 which has the

Table 5
Summary of the effects of 1,4-AQ on the quenching of chlorophyll fluorescence, PSII activity and electric properties (p^ and ci/S) of thylakoidmembranes with different stoichiometry of chlorophyll-protein complexes
Characteristics of thylakoid membranes Parameters
Quenching of
chlorophyll fluorescence
Inhibition of PSII activity p^ and ci/Sa
High amount of LHCII Strong effect No dependence No dependence
Small amount of LHCII Smaller effect No dependence No dependence
High ratio of LHCIIo/LHCIIm Strong effect No dependence No dependence
Small ratio of LHCIIo/LHCIIm Smaller effect No dependence No dependence
High amount of LHCII–PSII No dependence Higher effect for mono-substituted than di-substituted No dependence
Small amount of LHCII–PSII No dependence Smaller effect for mono-substituted than di-substituted No dependence
a The changes of p^ and ci/S depend on the chemical structure of 1,4-AQ.
Fig. 3. Sketch illustrating the correlation between the changes in the
thylakoid membrane electric dipole moments (permanent dipole
moment (p^) and electric polarizability per unit surface area (ci/S))and the extent of fluorescence quenching induced by the interaction of
membranes with 1,4-AQ. A simplified model of thylakoid membrane is
given (only part of the granal thylakoid is drawn, where the PSII and
LHCII complexes are located predominantly). The white and grey
ellipsoids represent the LHCII-oligomers and LHCII-trimers, respec-
tively; the black ellipsoids refer to the PSII-complexes. Other proteins,
pigment–protein complexes and lipid molecules are omitted for
simplicity and clearer representation of the model. The charge
distribution on the membrane surfaces should not be taken as real, it
is used to visualize the changes in charge distribution as a consequence
of the interaction of thylakoid membrane with 1,4-AQ. The changes in
the magnitude of the dipole moments are represented by longer and
shorter arrows, respectively. The scheme emphasizes that the 1,4-AQ-
induced increase and decrease in the dipole moments correlate (i) with
small and high accessibility of the chlorophyll molecules to 1,4-AQ,
and consequently (ii) with high and low fluorescence quenching.
E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123 121
lowestvalues of both the transmembrane charge asym-
metry and the electric polarizability among all the
variety of pea studied [18,20] (without 1,4-AQ) (Table
4). Therefore, the organization of the LHCII and the
electric properties of the membrane are important fac-
tors determining the accessibility of the chlorophyll
molecules to the substituted 1,4-anthraquinones. Dif-
ferences in the packing of the chlorophyll moleculesand in the exposure of polypeptide on the membrane
surface, related to variations in the LHCII organiza-
tion and the electric moments, respectively, might be
responsible for the different degree of the chlorophyll
quenching by substituted 1,4-AQ in thylakoid mem-
branes from different plant species. The relationship
between the variations of the electric dipole moments
and fluorescence quenching is illustrated in Fig. 3,where we have sketched the thylakoid membranes
(an oversimplified model) with the permanent dipole
moment perpendicular to the membrane plane and
the longitudinal electric polarizability. The arrows
show the magnitude of the dipole moments in presence
and without 1,4-AQ.
3-Chloro-9-hydroxy-1,4-anthraquinone (TF33) has
the highest quenching affinity among all substituted1,4-athraquinones and leads to the strongest reduction
of the membrane electric dipole moments (Table 4) for
all samples studied (wild types and mutants). It is to
be noted that in presence of TF33 the smallest values
of p^ and ci are observed for thylakoids from the mutant
Coeruleovireus 2/16 having the biggest quenching con-
stant Ksv (as already mentioned). The good correlation
between the quenching efficiency of 1,4-AQ and theorganization of LHCII, respectively, with the electric
parameters led to the proposal that the interaction and
probable penetration of the anthraquinones within the
membrane bilayer might be controlled by the surface
electric properties of the membrane (Fig. 3). The elec-
tro-optic data support the hypothesis of Karukstis and
Monell [26] that addition and/or removal of quinones
from their interaction site alter the conformation of thatregion. The conformational changes evidently depend
on the structure of the quinone, i.e., on the substituent
and its position in the molecule of 1,4-anthraquinone
(Table 4).
The degree of inhibition of PSII depends on the total
amount of the supramolecular complex LHCII–PSII aswell as on the number of substituting groups in the 1,4-
AQ. The latter reflects different sites of interaction of
mono- and di-substituted 1,4-AQ.
In conclusion, the quenching of the chlorophyll
fluorescence induced by 1,4-AQ strongly depends on
the organization of the thylakoid membranes, particu-
larly on the organization of LHCII, related to the

122 E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123
membrane surface electric properties. This is proved
for thylakoid membranes, characterized with different
stoichiometry of pigment–protein complexes and orga-
nization of the light-harvesting chlorophyll a/b protein
complex of photosystem II, from pea, barley and their
mutants.
5. Abbreviations
1,4-AQ 1,4-anthraquinones
QA4 9-acetoxy-1,4-anthraquinone
QA5 9-hydroxy-1,4-anthraquinone
QA6 1,4-anthraquinoneQA7 9-methoxy-1,4-anthraquinone
TF11 10-chloro-9-hydroxy-1,4-anthraquinone
TF23 9-acetoxy-3-bromo-1,4-anthraquinone
TF33 3-chloro-9-hydroxy-1,4-anthraquinone
TF34 3-bromo-9-hydroxy-1,4-anthraquinone
DCMU 3-(3-4-dichlorophenyl)-1,1-dimethylurea
KSV Stern–Volmer quenching constant
LHCII light-harvesting chlorophyll a/b protein com-plex of photosystem II
LHCIIo LHCII oligomer
LHCIIm LHCII monomer
PSII photosystem II
ci electric polarizability
p^ permanent dipole moment
mc critical relaxation frequency
Acknowledgements
The work was supported by contract K-1301/03 with
Bulgarian National Fund, Science Research. The
authors are grateful to Dr. N. Naydenova from the
Institute of Genetics, Bulgarian Academy of Sciences,for the donation of seeds of pea chlorophyll mutants.
References
[1] B.L. Trumpower (Ed.), Function of Quinones in Energy
Conserving Systems, Academic Press, New York, London,
1982.
[2] N. Fisher, P.R. Rich, A motif for quinone binding sites in
respiratory and photosynthetic systems, J. Mol. Biol. 296 (2000)
1153–1162.
[3] J. Ames, D.C. Fork, Quenching of chlorophyll fluorescence by
quinones in algae and chloroplasts, Biochim. Biophys. Acta 143
(1967) 97–107.
[4] N.G. Bukov, G. Sridhram, E.A. Egorov, R. Carpentier, Interac-
tion of exogenous quinones with membranes of higher plant
chloroplasts: modulation of quinone capacities as photochemical
and non-photochemical quenchers of energy in Photosystem II
during light–dark transitions, Biochim. Biophys. Acta 1604 (2003)
115–123.
[5] N.-U. Frigaard, S. Tokita, K. Matsuura, Exogenous quinones
inhibit photosynthetic electron transfer in Chloroflexus aurantia-
cus by specific quenching of the excited bacteriochlorophyll c
antenna, Biochim. Biophys. Acta 1413 (1999) 108–116.
[6] K.K. Karukstis, S.C. Boegeman, J.A. Fruetel, S.M. Gruber,
M.H. Terris, Multivariate analysis of photosystem II fluorescence
quenching by substituted benzoquinones and naphthoquinones,
Biochim. Biophys. Acta 891 (1987) 256–264.
[7] K.K. Karukstis, S.M. Gruber, J.A. Fruetel, S.C. Boegeman,
Quenching of chlorophyll fluorescence by substituted anthraqui-
nones, Biochim. Biophys. Acta 932 (1988) 84–90.
[8] M. Kitajima, W.L. Butler, Quenching of chlorophyll fluorescence
and primary photochemistry in chloroplasts by dibromothymo-
quinone, Biochim. Biophys. Acta 376 (1975) 105–115.
[9] J.W. Lee, W. Zipfel, T.G. Owens, Quenching of chlorophyll
excited states in photosystem I by quinones. Stern–Volmer
analysis of fluorescence and photochemical yield, J. Lumin. 51
(1992) 79–89.
[10] S. Tokita, N.-U. Frigaard, M. Hirota, K. Shimada, K. Matsuura,
Quenching of bacteriochlorophyll fluorescence in chlorosomes
from Chloroflexus aurantiacus by exogenous quinones, Photo-
chem. Photobiol. 72 (2000) 345–350.
[11] S. Vasil�ev, S. Wiebe, D. Bruce, Non-photochemical quenching of
chlorophyll fluorescence in photosynthesis. 5-Hydroxy-1,4-naph-
thoquinone in spinach thylakoids as a model for antenna based
quenching mechanisms, Biochim. Biophys. Acta 1363 (1998) 147–
156.
[12] K.K. Karukstis, M.A. Berliner, C.J. Jewell, K.T. Kuwata,
Chlorophyll fluorescence measurements to assess the competition
of substituted anthraquinones for the Qb binding site, Biochim.
Biophys. Acta 1020 (1990) 163–168.
[13] E. Apostolova, Tz. Markova, Tz. Filipova, M.T. Molina, S.G.
Taneva, Influence of substituted 1,4-anthraquinones on the
chlorophyll fluorescence and photochemical activity of pea
thylakoid membranes, J. Photochem. Photobiol. B 70 (2003) 75–
80.
[14] K.E. Steinback, J.J. Burke, C.J. Arntzen, Evidence for the role of
surface-exposed segments of the light-harvesting complex in
cation-mediated control of chloroplast structure and function,
Arch. Biochem. Biophys. 195 (1979) 546–557.
[15] A. Dobrikova, A. Ivanov, R.M. Morgan, I. Petkanchin, S.G.
Taneva, Contribution of LHC II complex to the electric properties
of thylakoid membranes: an electric light scattering study of Chl
b-less barley mutant, J. Photochem. Photobiol. B 57 (2000) 33–40.
[16] H.K. Lichtenthaler, Chlorophylls and carotenoids: pigments of
photosynthetic biomembranes, Methods Enzym. 148 (1987) 350–
383.
[17] S.G. Taneva, I. Petkanchin, Surface electric properties of biolog-
ical systems, Trends Photochem. Photobiol. 6 (1999) 113–139.
[18] A. Dobrikova, R.M. Morgan, A. Ivanov, E. Apostolova, I.
Petkanchin, N.P.A Huner, S.G. Taneva, Electric properties of
thylakoid membranes from pea mutants with modified carotenoid
and chlorophyll-protein complexes composition, Photosynth. Res.
65 (2000) 165–174.
[19] S.G. Taneva, M. Busheva, I. Petkanchin, Electric parameters of
Photosystem II particles – electric light scatering study, J.
Photochem. Photobiol. B 10 (1991) 315–327.
[20] A. Dobrikova, A.G. Ivanov, E. Apostolova, N. Naydenova, I.
Petkanchin, S.G. Taneva, Contribution of LHCII complex to the
electric properties of thylakoid membranes, in: N. Gozukirmizi
(Ed.), Plants of the Balkan Peninsula: Into the Next Millenium,
vol. 2, 2001, pp. 75–80.
[21] K.D. Allen, L.A. Staehelin, Resolution of 16 to 20 chlorophyll-
protein complexes using a low ionic strength native green gel
system, Anal. Biochem. 194 (1991) 214–222.
[22] S.P. Stoylov, Colloid Electrooptics, Academic Press, London,
1991.

E. Apostolova et al. / Journal of Photochemistry and Photobiology B: Biology 78 (2005) 115–123 123
[23] S. Jansson, The light-harvesting chlorophyll a/b-binding protein,
Biochim. Biophys. Acta 1184 (1994) 1–19.
[24] H. Paulsen, Chloropyll a/b binding proteins, Photochem. Photo-
biol. 62 (1995) 367–382.
[25] S. Nussberger, J.P. Dekker, W. Kuhlbrandt, B.M. van Bolhuis,
R. van Grondelle, H. van Amerongen, Spectroscopic character-
ization of three different monomeric form of the main chlorophyll
a/b binding protein from chloroplast membranes, Biochemistry 33
(1994) 14775–14783.
[26] K.K. Karukstis, C.R. Monell, Reversal of quinone-induced
chlorophyll fluorescence quenching, Biochim. Biophys. Acta 973
(1989) 124–130.
Copyright © 2022 FDOKUMEN




















