
Light-induced proton slip and proton leak at the thylakoid membrane
13
Journal of Plant Physiology 161 (2004) 1325—1337 Light-induced proton slip and proton leak at the thylakoid membrane Michael Richter , Jens Daufenbach, Stefanie Drebing, Verena Vucetic, Duc Tung Nguyen Institut fu ¨r Allgemeine Botanik, Johannes Gutenberg Universita¨t, Saarstr. 21, D-55099 Mainz, Germany Received 9 January 2004; accepted 15 March 2004 Summary A treatment of leaves of Spinacia oleracea L. with light or with the thiol reagent dithiothreitol in the dark led to partly uncoupled thylakoids. After induction in intact leaves, the partial uncoupling was irreversible at the level of isolated thylakoids. We distinguish between uncoupling by proton slip, which means a decrease of the H + /e - ratio due to less efficient proton pumping, and proton leak as defined by enhanced kinetics of proton efflux. Proton slip and proton leak made about equal contributions to the total uncoupling. The enhanced proton efflux kinetics corresponded to reduction of subunit CF 1 -g of the ATP synthase as shown by fluorescence labeling of thylakoid proteins with the sulfhydryl probe 5-iodoacetamido fluorescein. The maximum value of the fraction of reduced CF 1 -g was only 36%, which indicates that in vivo the reduction of CF 1 -g could be limited by fast reoxidation and/or restricted accessibility of CF 1 -g to thioredoxin. Measurements of the ratio ATP/2e indicated that only the uncoupling related to less efficient proton pumping led to a decrease in the ATP yield. r 2004 Elsevier GmbH. All rights reserved. Introduction The driving force of ATP synthesis in the illuminated chloroplast is the chemical potential of protons. This is established in the light by the water splitting reactions of PS II and the vectorial proton pumping across the thylakoid membrane driven by photo- synthetic electron transport. The optimal ATP yield of photosynthetic electron transport is supposed to be 3 molecules of ATP per 4 electrons, which results from a ratio of 3 protons per electron and a ratio of 4 protons used in the synthesis of one molecule of ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS: CF 1 -g; Fluorescence labeling of proteins; H + /e ; Proton leak; Proton slip; Thylakoid uncoupling; Spinacia oleracea 0176-1617/$ - see front matter r 2004 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.03.007 Abbreviations: Chl, chlorophyll; DTT, dithiothreitol; FeCy, ferricyanide; IAF, 5-iodoacetamido-fluorescein; MV, methyl viologen; PPFD, photosynthetically active photon flux density Corresponding author. Tel.: +49-6131-3923166; fax: +49-6131-3923075. E-mail address: [email protected] (M. Richter).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Light-induced proton slip and proton leak at the thylakoid membrane

ARTICLE IN PRESS
Journal of Plant Physiology 161 (2004) 1325—1337
KEYWORDCF1-g;Fluorescenlabeling oH+/e�;Proton leaProton slipThylakoiduncouplingSpinacia o
0176-1617/$ - sdoi:10.1016/j.
AbbreviationPPFD, photosyn�CorrespondE-mail addr
www.elsevier.de/jplph
Light-induced proton slip and proton leak at thethylakoid membrane
Michael Richter�, Jens Daufenbach, Stefanie Drebing, Verena Vucetic,Duc Tung Nguyen
Institut fur Allgemeine Botanik, Johannes Gutenberg Universitat, Saarstr. 21, D-55099 Mainz, Germany
Received 9 January 2004; accepted 15 March 2004
S:
cef proteins;
k;;
;leracea
ee front matter r 20jplph.2004.03.007
s: Chl, chlorophyll; Dthetically active phoing author. Tel.: +49-6ess: [email protected]
SummaryA treatment of leaves of Spinacia oleracea L. with light or with the thiol reagentdithiothreitol in the dark led to partly uncoupled thylakoids. After induction in intactleaves, the partial uncoupling was irreversible at the level of isolated thylakoids. Wedistinguish between uncoupling by proton slip, which means a decrease of the H+/e�-ratio due to less efficient proton pumping, and proton leak as defined by enhancedkinetics of proton efflux. Proton slip and proton leak made about equal contributionsto the total uncoupling. The enhanced proton efflux kinetics corresponded toreduction of subunit CF1-g of the ATP synthase as shown by fluorescence labeling ofthylakoid proteins with the sulfhydryl probe 5-iodoacetamido fluorescein. Themaximum value of the fraction of reduced CF1-g was only 36%, which indicates thatin vivo the reduction of CF1-g could be limited by fast reoxidation and/or restrictedaccessibility of CF1-g to thioredoxin. Measurements of the ratio ATP/2e indicated thatonly the uncoupling related to less efficient proton pumping led to a decrease in theATP yield.r 2004 Elsevier GmbH. All rights reserved.
Introduction
The driving force of ATP synthesis in the illuminatedchloroplast is the chemical potential of protons.This is established in the light by the water splittingreactions of PS II and the vectorial proton pumping
04 Elsevier GmbH. All rights res
TT, dithiothreitol; FeCy, ferricton flux density131-3923166; fax: +49-6131-392ni-mainz.de (M. Richter).
across the thylakoid membrane driven by photo-synthetic electron transport. The optimal ATP yieldof photosynthetic electron transport is supposed tobe 3 molecules of ATP per 4 electrons, which resultsfrom a ratio of 3 protons per electron and a ratio of4 protons used in the synthesis of one molecule of
erved.
yanide; IAF, 5-iodoacetamido-fluorescein; MV, methyl viologen;
3075.

ARTICLE IN PRESS
M. Richter et al.1326
ATP (Berry and Rumberg, 1996). However, thesestoichiometries are expected to vary in response togrowth conditions, which may specifically demandfor enhanced supply with ATP or NADPH. Increasingand decreasing of the ATP yield may be achieved byvariation of the ratio of linear to cyclic electrontransport and, alternatively, by partial uncouplingat the thylakoid membrane. The present workfocuses on a light-induced uncoupling.
Three major uncoupling mechanisms have to beconsidered, which potentially lead to a decrease inthe ATP yield. These are the proton slip and theredox slip, which are both expected to decreasethe H+/e� ratio, and the proton leak, which isexpected to increase the number of protons neededper ATP. A proton slip means a back flow of protons‘‘within’’ a proton pump, if for example, theprotons have to be ejected against a large chemicalpotential of protons. Such a back flow of protons tothe stroma within the cyt b6f complex has beensuggested as a reason for the strong light-induceddecrease in the H+/e� ratio (Berry and Rumberg,1999). A redox slip would occur if an electron wouldpass by a regular coupling site of an electrontransport chain. This is observed in tetramethyl-p-phenylendiamine treated mitochondria (Groen etal., 1990). A proton leak involves the passive backflow of protons through a leaky site of a membrane,which has been observed with photodamagedthylakoids (Ogren and Oquist, 1984; Terashima etal., 1991; Tjus and Andersson, 1993; Curwiel andVan Rensen, 1993).
The research on light effects on the quantitativerelationship between photosynthetic electrontransport and the proton gradient mainly focussedon two sites within the thylakoid membrane,namely, the cyt b6f complex and the couplingfactor.
Cyt b6f complex
Two alternative mechanistic proposals were madeto explain a light-induced variation of the H+/e�
ratio associated with the cyt b6f complex. Schubertet al. (1990) attributed the decrease of the H+/e�
ratio from 3 under low light intensity to 2 understrong light to a redox slip resulting from decreasedQ-cycle activity. This is caused by the decreasedresidence time of electrons on the Rieske-FeS-center, which results from the more oxidized stateof cyt f under strong light. Alternatively, Berry andRumberg (1999) put foreword the hypothesis that ahigh chemical potential of protons induces a protonslip within the cyt b6f complex, which is responsiblefor the lower H+/e� ratio with high light intensity.
ATP synthase
An increase of the proton conductivity through theATP synthase was discussed in connection with: (i) aregulatory adenine nucleotide binding to CF1(Strotmann et al., 1986), (ii) the triggering levelof the DpH for the activation of the ATP synthase(Junge et al., 1970; Schonfeld and Neumann,1977), and (iii) the reduction of disulfide bondsand subsequent alkylation of free cysteine residuesespecially at the subunit CF1-g (Evron and McCarty,2000). Mills and Mitchell (1984) suggested thatreduction of CF1-g increases the fraction of activeATP synthases at a given DpH, possibly due to anenhanced proton flow via the phosphorylating pathrelative to the non-phosphorylating path. Today aclose connection between proton flux through theATP-synthase and CF1-g is already evident from itscentral role in photophosphorylation as a part ofthe rotor, which couples the chemical potential ofprotons to ATP synthesis through its rotatory motionwithin CF1 (Sabbert et al., 1997).
In the present work, we demonstrate that atreatment of leaves at different light intensitiesand a DTT treatment of leaves in the dark led to apartial uncoupling of thylakoids. The uncouplinginvolved a proton slip due to a decrease in theH+/e� ratio as well as a proton leak associated withreduction of CF1-g.
Materials and methods
Growth conditions
Spinach (Spinacia oleracea L.) plants were grownon a nutrient medium described by Robinson(1986). The plants were kept under a light regimeof 11 h at a PPFD of 125 mmolm�2 s�1 and 13 h in thedark. The temperature was 18 1C in the light and15 1C in the dark. The leaves were harvested fromfive- to six-week old plants.
Leaf pretreatment
The leaves were either treated for 45min at thePPFD of the growth light (125 mEm�2 s�1, LLtreatment) or 15min under high light (1000 mEm�2 s�1, HL treatment). For the treatment withdithiothreitol (DTT), the leaves were vacuuminfiltrated in the dark with a solution of 20mMDTT in tap water and subsequently stored for45min in the dark at room temperature. The DTTtreatment did not affect the light-saturated rate ofphotosynthetic oxygen evolution of the leaves,

ARTICLE IN PRESS
Light-induced thylakoid uncoupling 1327
which was between 20 and 25 mmolm�2 s�1 regard-less of whether the leaves were DTT treated or not.The optimal photosynthetic quantum yield ofPhotosystem 2, as expressed by the ratio of variableto maximal chlorophyll fluorescence (Genty et al.,1989), was 0.78 in dark adapted DTT treated and0.8 in untreated leaves. This shows that DTT did notlead to significant dark reduction of the plastoqui-none pool and closure of Photosystem 2 centres.Nevertheless, DTT was proven to penetrate theleaves as it completely inhibited the strong light-induced conversion of violaxanthin to zeaxanthinby the DTT sensitive enzyme violaxanthin de-epoxidase located in the thylakoid lumen space(Yamamoto and Kamite, 1972).
Preparation of thylakoids and labeling ofcyteines of thylakoid proteins with 5-iodoacetamido-fluorescein (IAF)
Thylakoids were prepared from spinach leaves asdescribed earlier (Richter et al., 1990) except thatbovine serum albumine was omitted from thehomogenization medium. The thylakoids werefinally diluted to a chl aþ b content of 1 g l�1 in astorage medium, which was composed of 330mMsorbitol, 10mM NaCl, 5mM MgCl2, 1mM MnCl2,2mM EDTA, 0.5mM KH2PO4, 40mM HEPES/KOH(pH 7.0), and then stored on ice until use. Allpreparation steps were performed in a cold roomunder dim light. The chlorophyll content wasdetermined according to Ziegler and Egle (1965).All experiments described below were performedin parallel using one and the same stock suspensionof thylakoids to avoid influences from differentpreparations.
The treatment of proteins with sulfhydryl specificfluorescence probes is an established approach forthe identification of proteins, which are regulatedby reduction of a disulfide bond (Yano et al., 2001).Ten ml of a stock solution of IAF (50mM in 50mMTricine/NaOH pH 8) were added to 90 ml of storagemedium pH 7 containing isolated thylakoids equiva-lent to 90 mg Chl. The labeling mixture wasincubated for 30min in the dark at room tempera-ture. The labeling was finished by the addition of900 ml ice-cold acetone and subsequent storage at–20 1C for 20min in the dark, which led to theprecipitation of proteins. The precipitate wascollected by centrifugation at 13.000g for 2min.The sediment was washed several times with ice-cold acetone and centrifuged to remove chlorophylland excessive IAF until a colourless supernatant wasachieved. After removing the acetone by a streamof nitrogen gas, 100 ml of denaturing buffer were
added and the samples were heated in boilingwater for 2min. Samples of 20 ml (equivalent to18 mg Chl) were loaded on each lane.
Electrophoresis and western blotting
Proteins were separated on 12% polyacrylamidegels (200� 180� 0.7mm3) at 4 1C using buffersystems according to Laemmli (1970). It is impor-tant to note that all protein samples weredenatured in the presence of 2% DTT to cleave alldisulfide bonds. After the electrophoretic separa-tion of proteins the integrated fluorescence in-tensity of the IAF bound to the proteins wasobtained by densitometric analysis of fluorescencebands using the software Quantity One (BIO RAD,USA).
Western blotting of the gels was performedusing the Trans-Blot semi-dry transfer cell (BIO-RAD, USA) according to the manufacturers instruc-tions and the buffer system according to Towbin etal. (1979). The antiserum against CF1-g was a kindgift of Prof. Berzborn (Bochum, F.R.G.). Theimmunoreactive bands were visualized by colorprecipitation using alkaline phosphatase-conju-gated antibodies.
Photosynthetic electron transport andphotophosphorylation
The photosynthetic electron transport of isolatedthylakoids was measured in an oxygen electrode(DW1, Hansatech, UK) as oxygen consumption withmethyl viologen (MV, 0.1mM). The reaction med-ium contained 0.33M sorbitol, 50mM KCl, and 5mMMgCl2, 40mM HEPES/KOH pH 7.5. This medium isreferred to as the high buffer medium. The DpHacross the thylakoid membrane imposes stronglimitations on the basal electron transport, thatis, the electron transport in the absence of ADP, Pi,and uncoupler. The coupling ratio denotes the ratioof the light-saturated uncoupled electron transportrate with 5mM NH4Cl divided by the light-saturatedbasal electron transport rate of a sample. Theaverage coupling ratio of all thylakoids preparedfrom dark control leaves was 12.571.8, whichindicates a high degree of intactness of thylakoids.A stimulation of the light saturated basal electrontransport was taken as an indicator for uncouplingdue to the pretreatment of leaves. An uncouplingratio (u) is here defined as the quotient of thecoupling ratio of dark control samples divided bythe coupling ratio of samples from light or DTTtreated leaves.

ARTICLE IN PRESS
M. Richter et al.1328
The ATP/2e ratio of samples was determinedwith light-saturated coupled linear electron trans-port (H2O to FeCy). The ATP content was measuredas the increase in absorption at 340 nm afteraddition of hexokinase and glucose-6-phosphatedehydrogenase to the samples (Groen et al., 1990).The electron transport was calculated from the lossof absorption at 420 nm associated with thereduction of FeCy using an � of 1.035 cm�2 mmol�1.
The starting conditions were 1mM ADP, 5mMphosphate and 1mM FeCy in high buffer medium.The photophosphorylation was terminated afterone minute of illumination at a PPFD of 1000 mEm�2 s�1 by the addition of HClO4. After 15minstorage on ice, the samples were neutralized byKOH and kept on ice for 10min. Thereafter, thesuspensions were centrifuged (10min at 4000g) andthe amount of electrons transported determined.Then 1ml of the clear supernatant of the sampleswas added to 1ml of a reaction mixture containing0.15M Tris/HCl pH 7.5, 0.1M glucose, 1mM KCl,1mM MgCl2, 1mM NADP+. Ascorbate (1mM) wasused to reduce residual FeCy. The reaction wasstarted by the addition of hexokinase (14 U/ml) andglucose-6-phosphate dehydrogenase (0.5 U/ml).The ratio of ATP/2e of control samples divided byATP/2e of samples from either HL or DTT treatedleaves was denoted the ATP yield-related uncou-pling ratio (uATP).
Measurement of 9-aminoacridine-fluorescence
The build up of a proton gradient across thylakoidscoupled to basal electron transport was measuredas the quenching of the fluorescence of the diamine9-aminoacridine (9-AA; Schuldiner et al., 1972; Pickand McCarty, 1980).
The 9-AA fluorescence was measured in highbuffer medium with thylakoids at a chl content of30 g l�1, 0.1mM MV as the electron acceptor, and6 mM 9-AA. The measurements were performed witha spectrophotometer (DW-2, SLM AMINCO, USA)operated in the single beam mode. 9-AA fluores-cence was excited at 400 nm using light provided bythe monochromator (bandpass 5 nm). The excitinglight itself had no actinic effects and did not lead tothe build up of a proton gradient. Red actinic lightwas provided by a halogen lamp filtered through ared cut off filter (RG 645, 4mM, Schott, FRG). Itwas led to the sample cuvette at a right angle tothe excitation beam. The intensity of the actiniclight was varied by the use of neutral densityfilters. The photomultiplier was protected againstthe blue excitation light, red actinic light, and
chlorophyll fluorescence by the filter combinationBG 39 (5mm) plus GG 435 (2mm, Schott, F.R.G.)thereby giving a transmission maximum at 500 nm.
Determination of the H+/e�-ratio and thekinetics of proton efflux
The H+/e� ratio with linear electron flow fromwater to FeCy was measured with a pH-micro-electrode (Inlab 423, Mettler Toledo, F.R.G.) asdescribed by Fowler and Kok (1976) in a reactionmedium with low buffer concentration (0.33Msorbitol, 50mM KCl, 5mM MgCl2, 0.3mM tricine/HCl pH 7.0), which is referred to as the low buffermedium. The chlorophyll content was 30mg l�1.With the low buffer medium a PPFD of 300 mEm�2 s�1 was sufficient to saturate the basalelectron transport. During steady-state electrontransport to FeCy, the sum of proton pumping intothe thylakoid lumen and production of protons bywater splitting in the lumen is equal to the protonefflux from the lumen. Because the reduction ofFeCy itself is not accompanied by proton exchangereactions and because only the fraction of theformerly pumped protons is neutralized by OH�
after efflux, the water splitting produces a net gainof two protons per two electrons (see also Fig. 6a).This relation was used to determine the steady-state rate of electron transport to FeCy from theslope of the linear pH decrease in the light (Pw).When the light is switched off, proton pumping andwater splitting both stop immediately. Then theslope of the pH decrease immediately after light off(Pe) represents the rate of proton efflux fromthylakoids during steady-state electron transportand Pe/Pw is a measure for the H+/e� ratio. Thisrelationship is only valid for very small pH changes,where the response of the pH-electrode to light-induced proton uptake into and efflux fromthylakoids can be treated as linear and notlogarithmic (Fowler and Kok, 1976). This wasproven by the addition of aliquots of dilute acidor base, which also proved that the time responseof the system was much faster than the kinetics ofproton efflux from control thylakoids (t1=2 ¼ 12 s).The use of nonactin to suppress a potential light-offmembrane diffusion potential as recommended byBerry and Rumberg (1999) did not change thekinetics and was thus omitted. The data from thepH electrode were digitally converted and sampledat a frequency of 4Hz.
The kinetics of the proton efflux from isolatedthylakoids and the electron transport rate atsaturating light were simultaneously measured inthe low buffer medium using an oxygen electrode

ARTICLE IN PRESS
Light-induced thylakoid uncoupling 1329
vessel, which was modified to fit in the pH micro-electrode. When we focussed on the measurementof the kinetics of proton efflux we used 0.1mM MVas the electron acceptor instead of FeCy. The mainadvantage of MV is that electron uptake is followedby stoichiometric proton uptake, which in turn isequivalent to the release of protons by watersplitting (see Fig. 6a). Therefore, the pH remainsconstant during steady-state electron transport,which makes it easier to determine the initial rateof proton efflux accurately. In addition, all effluxkinetics started at the same pH irrespective of therate and the duration of electron transport. Thechlorophyll content was 30mg l�1 and the PPFD was300 mmolm�2 s�1.
Figure 1. Comparison of basal electron transport (H2O toMV) in high buffer medium with thylakoids isolated fromleaves that had been exposed to different pretreat-ments: HL, 15min 1000mEm�2 s�1, LL, 45min growthlight (125 mEm�2 s�1), DTT, leaves vacuum infiltratedwith 20mM dithiothreitol and incubated in the dark for40min, control, dark adapted leaves. The chl content ofthe suspensions was 30mg l�1 and the concentration ofMV was 0.1mM. The light-saturated uncoupled rate ofoxygen evolution was 375730 mmolmg�1 Chl h�1 with allpretreatments. The coupling ratios of the suspensions,i.e. the ratio of light-saturated uncoupled divided bylight-saturated basal electron transport were 12.5 (con-trol), 6.25 (HL), 6.6 (LL), and 8.9 (DTT). The resultinguncoupling ratios (u) were 2.0 (HL), 1.9 (LL), 1.4 (DTT).The experiments were repeated three times. Thestandard deviation did not exceed 10%.
Results
Thylakoids from light or DTT treated leavesare partly uncoupled
Thylakoids from HL pretreated leaves were par-tially uncoupled, when compared to thylakoidsfrom dark adapted control leaves, as indicated bya lower coupling ratio and higher rates of electrontransport (Fig. 1). The extra oxygen consumption inthe partly uncoupled HL thylakoids was 30 mmolO2mg�1 Chl h�1 with saturating actinic light, whichis equivalent to 20% of the typical light-saturatedoxygen evolution in intact spinach leaves (150 mmolO2mg�1 Chl h�1). The quotient of coupling ratios ofcontrol divided by HL of 2.0 is here denoted as theHL-induced uncoupling ratio (u). The values of u ofa particular sample and respective control re-mained constant over 40min storage of thylakoidsat room temperature in the dark and over evenmore extended periods of storage of the thylakoidson ice (not shown). The HL-induced uncouplingsaturated at a value of u of 2.0 even with prolongedHL treatment (not shown), although this corre-sponds to a relative uncoupling of only 16% of thepotential maximal value of 12.5.
Forty-five minutes exposure of leaves to growthlight (LL) led to a similar uncoupling in thylakoids(u=1.9, Fig. 1), which indicates a saturation ofuncoupling at comparatively low light intensity.The partial uncoupling observed with thylakoidsfrom the DTT treated leaves (u=1.4, Fig. 1) pointedto a mechanism involving the reduction of disulfidebonds. Uncoupling was not induced by reducingconditions in general, as it was not observed inthylakoids from leaves infiltrated with 30mMascorbate in the dark for 40min instead of DTT(not shown).
Thylakoid preparations from light or DTT treatedleaves and thylakoids from control leaves showedthe same rate of light-saturated electron transport,when the samples were completely uncoupled bythe use of NH4Cl. It is therefore clear that theobserved changes in the coupling ratio induced bylight or DTT were due to changes in the basalelectron transport and reflect partial uncoupling,whereas the capacity of the photosynthetic elec-tron transport chain for uncoupled electron trans-port was not affected.
The partial uncoupling made electron transportless efficient in building up a DpH, as shown inFig. 2. The decrease in efficiency was morepronounced after the light pretreatments thanwith the DTT pretreatment and it was also morepronounced with light-saturated electron transportrates than with light-limited rates.
The uncoupling was further characterized usingmethods that distinguish between a reduction in

ARTICLE IN PRESS
Figure 3. Original trace of the pH-electrode signalobtained with light-saturated basal electron transport(H2O to FeCy). The slope of the tangent to the pHdecrease at steady state (Pw) was calculated by linearregression analysis of the last 5 s (20 data points) beforethe light was switched off. It is a measure for the rate ofproton release due to water splitting. The slope of thetangent to the light-off kinetics of the pH (Pe) wascalculated by linear regression analysis of the first 2 s (8data points) after the light was switched off. It is ameasure for the rate of proton efflux at steady state. Thequotient Pe/Pw reflects the ratio H+/e�. The identicaldecreases of the pH after addition of aliquots of diluteKOH and HCl confirm a linear response of the pH
Figure 2. The dependence of the proton gradient on therate of basal electron transport (H2O to MV) at differentactinic light intensities in thylakoids prepared fromleaves pretreated at the conditions described in thelegend to Fig. 1. The proton gradient was calculated fromthe quenching of 9-aminoacridine (9-AA) fluorescence.The concentration of 9-AA was 6 mM. The chlorophyllcontent of the suspensions was 30mg l�1. The data forthe electron transport (H2O to MV) are from Fig. 1 andthe respective quenching of 9-AA fluorescence wasmeasured at an identical PPFD.
M. Richter et al.1330
the H+/e� ratio caused by redox or proton slip andproton leak.
electrode to the concentration of protons within thesmall pH range covered by the signals of light-induced pHchange. The fast responses to the addition of HCl provesthe proper resolution of proton efflux kinetics by the pHelectrode. The chl content of the suspensions was30mg l�1. The PPFD was 300 mEm�2 s�1.
Light and DTT treatment induced a decreasein H+/e�
Fig. 3 shows an original trace of the pH changeassociated with light-saturated linear electrontransport from water to FeCy of thylakoids fromdark control leaves in the low buffer medium. Soonafter the actinic light was switched on, the rate ofproton efflux from the lumen was equal to the rateof proton pumping into the lumen and the lineardecrease of the pH indicated a constant rate ofwater splitting. The steeper decrease immediatelyafter the light was turned off represented theefflux of the protons accumulated in the lumen insteady state, either by proton pumping or watersplitting. As outlined in the Materials and methodssection, the ratio of the rates of proton efflux atlight-off and proton release from water splitting(Pe/Pw) is a measure for the H+/e� ratio. The valueof 2.0 with light-saturated electron transport incontrol thylakoids from dark adapted leaves agreeswell with earlier reports from Fowler and Kok(1976) and Berry and Rumberg (1999). With
thylakoids from HL pretreated leaves this ratiodropped to 1.5 (Table 1). The extent of uncouplingspecifically associated with the decrease in H+/e�
is denoted uH and is calculated according touH=(H
+/e�control)/(H+/e�HL)=1.33. The light-saturated
rate of proton release from water splitting atsteady state (Pw) was stimulated by a factor of 1.9in thylakoids from HL treated leaves compared tothylakoids from dark controls (Table 1). This is ingood agreement to an uncoupling ratio u of 2.0 withMV dependent basal electron transport as obtainedwith identically treated samples in the high buffermedium (Fig. 1), although the absolute electrontransport rates with the low buffer medium werelower by about a factor of three due to the strongerlimitation by the DpH. The value of Pw did not varyin a range of concentrations of FeCy between 1 and
ARTICLE IN PRESS
Table 1. Uncoupling in leaves treated under HL or with DTT in the dark
Uncouplingratio u
PwPw control
Specific uncouplingratio uH
Specific uncouplingratio ue
Ratio ATP=2econtrolATP=2e uATP
HL 2.0 1.9 1.33 1.5 1.44DTT 1.4 1.4 1.2 1.2 1.24
Prior to thylakoid isolation spinach leaves were treated under HL (15min 1000mEm�2 s�1) or leaves were vacuum infiltrated with 20mMDTT in the dark for 40min. Control leaves were dark adapted over night. The membranes prepared from the pretreated leaves werethen used to determine (a) the uncoupling ratio (u), (b) the light-saturated rate of proton release from water splitting (Pw), (c) thespecific uncoupling ratio associated with proton slip (decrease in the ratio H+/e�, uH), (d) specific uncoupling related to proton leak(increased proton efflux from thylakoids, ue), and the specific uncoupling ratio related to a loss in the ATP yield (uATP). The light-saturated rate of oxygen consumption associated with basal electron transport from water to MV in thylakoids from control leaves was30mmolmg�1 chl h�1 in the high buffer medium and about 10mmolmg�1 chl h�1 in the low buffer medium. These rates were usuallyenhanced by at least a factor of 10 after the addition of 5mM of the uncoupler ammonium chloride. The ratio H+/e� of controls withlight-saturated electron transport to FeCy in low buffer medium was 2.0. The ratio ATP/2e of controls with electron transport to FeCyin high buffer medium was 1.3. All measurements were repeated at least three times, standard deviations were less than 10%.
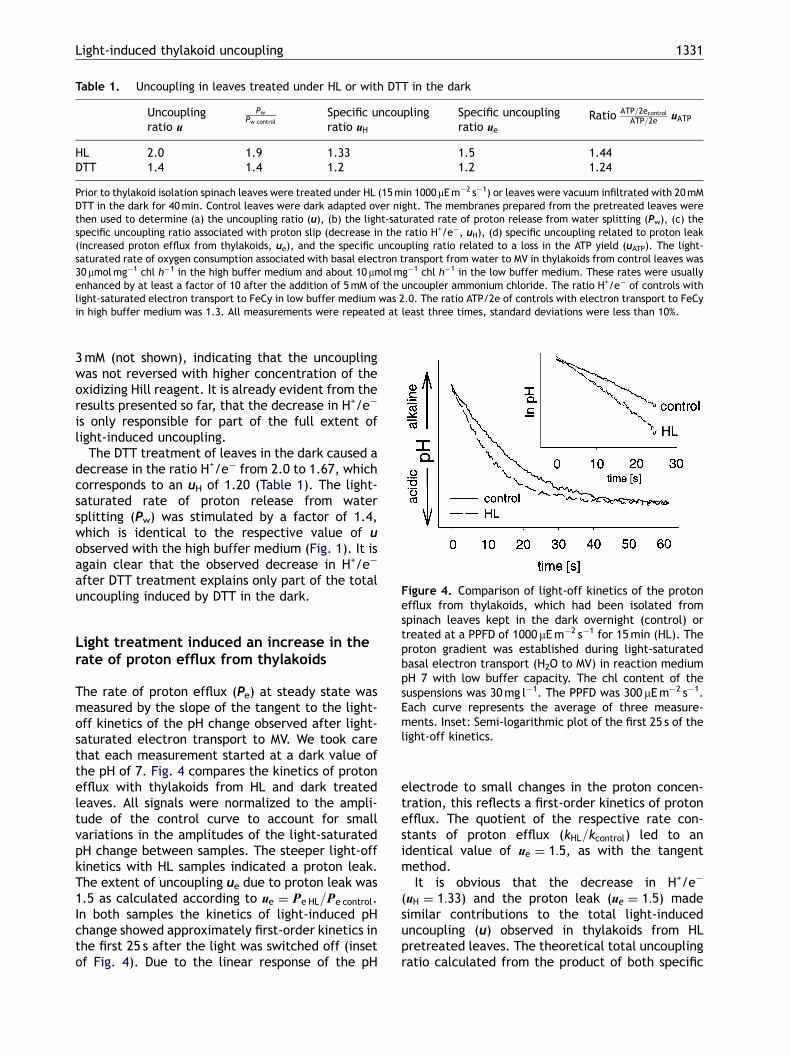
Figure 4. Comparison of light-off kinetics of the proton
Light-induced thylakoid uncoupling 1331
3mM (not shown), indicating that the uncouplingwas not reversed with higher concentration of theoxidizing Hill reagent. It is already evident from theresults presented so far, that the decrease in H+/e�
is only responsible for part of the full extent oflight-induced uncoupling.
The DTT treatment of leaves in the dark caused adecrease in the ratio H+/e� from 2.0 to 1.67, whichcorresponds to an uH of 1.20 (Table 1). The light-saturated rate of proton release from watersplitting (Pw) was stimulated by a factor of 1.4,which is identical to the respective value of uobserved with the high buffer medium (Fig. 1). It isagain clear that the observed decrease in H+/e�
after DTT treatment explains only part of the totaluncoupling induced by DTT in the dark.
efflux from thylakoids, which had been isolated fromspinach leaves kept in the dark overnight (control) ortreated at a PPFD of 1000 mEm�2 s�1 for 15min (HL). Theproton gradient was established during light-saturatedbasal electron transport (H2O to MV) in reaction mediumpH 7 with low buffer capacity. The chl content of thesuspensions was 30mg l�1. The PPFD was 300 mEm�2 s�1.Each curve represents the average of three measure-ments. Inset: Semi-logarithmic plot of the first 25 s of thelight-off kinetics.
Light treatment induced an increase in therate of proton efflux from thylakoids
The rate of proton efflux (Pe) at steady state wasmeasured by the slope of the tangent to the light-off kinetics of the pH change observed after light-saturated electron transport to MV. We took carethat each measurement started at a dark value ofthe pH of 7. Fig. 4 compares the kinetics of protonefflux with thylakoids from HL and dark treatedleaves. All signals were normalized to the ampli-tude of the control curve to account for smallvariations in the amplitudes of the light-saturatedpH change between samples. The steeper light-offkinetics with HL samples indicated a proton leak.The extent of uncoupling ue due to proton leak was1.5 as calculated according to ue ¼ Pe HL=Pe control.In both samples the kinetics of light-induced pHchange showed approximately first-order kinetics inthe first 25 s after the light was switched off (insetof Fig. 4). Due to the linear response of the pH
electrode to small changes in the proton concen-tration, this reflects a first-order kinetics of protonefflux. The quotient of the respective rate con-stants of proton efflux (kHL=kcontrol) led to anidentical value of ue ¼ 1:5, as with the tangentmethod.
It is obvious that the decrease in H+/e�
(uH ¼ 1:33) and the proton leak (ue ¼ 1:5) madesimilar contributions to the total light-induceduncoupling (u) observed in thylakoids from HLpretreated leaves. The theoretical total uncouplingratio calculated from the product of both specific

ARTICLE IN PRESS
M. Richter et al.1332
uncoupling ratios (uH � ue) was 2.0, which corre-sponded well to the uncoupling ratio u of 1.9 asdetermined by the ratio Pw HL=Pw control.
The proton leak was less pronounced in thyla-koids from DTT treated leaves. The efflux specificuncoupling ratio ue was 1.2 (Table 1) as expectedwith regard to the total uncoupling ratio u of 1.4and a value of uH of 1.2.
During the illumination of the thylakoids, wemeasured the light-saturated electron transportfrom water to MV as oxygen consumption. We werethus able to calculate the H+/e ratio by dividingthe efflux rates immediately after light-off bythe corresponding steady-state electron transportrates. We obtained values of 2.3 (control), 1.6 (HL),and 1.8 (DTT). Possibly due to stromal protonconsumption associated with MV reduction (seeFig. 6), the pH gradient and the H+/e� ratios wereslightly higher as compared to the values obtainedwith electron transport to FeCy. But the resultinguncoupling of 1.44 (HL) and 1.28 (DTT) was verysimilar. Therefore, these results were not consid-ered in Table 1.
The ratio of ATP/2e of control thylakoids was 1.3and it decreased to 0.9 and 1.05 in thylakoidsfrom HL and DTT treated leaves. This correspondedto an uncoupling ratio uATP ¼ ½ðATP=2econtrolÞ=ðATP=2eHL;DTTÞ of 1.44 and 1.24 in the respectivepartly uncoupled thylakoids.
Treatment of leaves by light and DTT led topartial reduction of CF1-c
The involvement of disulfide reduction in a thyla-koid protein in partial uncoupling, as suggested bythe DTT experiments, was tested by the use of thesulfhydryl reactive fluorescence probe 5-iodoacet-amidofluorescein (IAF). The identification of thoseproteins carrying a pair of sulfhydryls from thereduction of a disulfide bond would have beenfacilitated by blocking the pre-existing free sulfhy-dryls by a treatment of leaves with the unlabeledsulfhydryl reactive compounds iodoacetamide oriodoacetate prior to illumination or DTT treatment.Unfortunately, this was not possible because of tworeasons. First, the photosynthetic electron trans-port in thylakoids prepared from such pretreatedleaves was strongly impaired. Consequently, thepretreatment not only suppressed the backgroundlabeling of permanently free sulfhydryls, but alsothe thioredoxin dependent reduction of disulfidebonds in the light. A comparison of fluorescencelabeling and electron transport was impossible withsuch material. Second, the blocking at the level of
the intact leaf was not quantitative and showedlarge variation.
Fig. 5a illustrates the fluorescence patterns andthe respective densitometric profiles of thylakoidproteins from dark adapted and HL pretreatedleaves. The major light-induced change is in therange of 34 kDa, where one band in the control laneapparently split into two weaker bands in the HLlane. These bands were both identified as CF1-g byWestern Blotting (Fig. 5b). The lower fluorescenceband is here referred to as g and the upper band asg*. The mobility shift of g* of about 0.8 kDa is inagreement with the binding of two more moleculesof IAF to CF1-g after reduction of its disulfide bond(Cys199 S-S Cys205; Nalin and McCarty, 1984), whichcorresponds to an additional molecular mass of1 kDa. In the HL lane there was one additional weakimmuno band below the g-band. This was possiblydue to a very small amount of CF1-g, which wasunlabeled or carried only one molecule of IAF.
There was only a small amount of g* after 40minDTT treatment of leaves in the dark. With regard tothe reduction of the disulfide bond of CF1-g, theextended DTT treatment in the dark (Fig. 5c) wasapparently less efficient than the shorter treatmentwith HL, which is in strong agreement withKetcham et al. (1984).
Based on the assumption that oxidized andreduced CF1-g contain two and four molecules ofIAF per molecule of protein, respectively, wecalculated the relative amount of reduced CF1-g(CF1-gred) from the fluorescence intensities fg andfg* (Fig. 5) according to CF1-gred=(0.5� fg*)/(0.5� fg*+fg). CF1-gred-values of 0 (control), 0.17(DTT), and 0.36 (HL) and respective uncouplingassociated with proton leak (ue) of 1 (control), 1.2(DTT), and 1.5 (HL) shown in Table 1 follow thesame trend, which points to the possibility that theproton leak is located at CF1 and is associated withreduction of CF1-g.
Our results show that even after the HL treat-ment, which was most effective with regard toreduction of CF1-g and uncoupling, only 36% of CF1-gwas actually in the g*-band. This incompletereduction of CF1-g was probably not a result ofreoxidation of CF1-g in the isolated thylakoids,because uncoupling as well as the binding of IAFto CF1-g were stable despite the presence ofair saturated levels of oxygen and irrespective ofthe time of dark storage before the measurementof uncoupling and addition of label to thylakoids.Our results are in good agreement with earlierresults of Ketcham et al. (1984), who found thateven after a light treatment of thylakoids in thepresence of DTT, only 46% of CF1-g was in thereduced state.

ARTICLE IN PRESS
Figure 5. (a) The fluorescence pattern of thylakoid proteins, which were labeled with the fluorescent sulfhydryl probe5-IAF and then separated by SDS-PAGE. The densitometric profiles of the lanes are shown to the right and left of therespective lane. Protein equivalent to 18 mg chl was loaded on each lane. The thylakoids were isolated from darkadapted spinach leaves (control) or leaves treated at a PPFD of 1000 mEm�2 s�1 for 15min (HL). The position of theunmodulated (CF1-g) and the reduced subunit g of the ATP synthase (CF1-g*) is labeled g and g*. (b) Comparison offluorescence pattern of IAF labeled thylakoid proteins (lane IAF, negative stain) and immuno detection of CF1-g and CF1-g*(lane blot). The figure shows results of a repetition of the experiment shown in 5(a). Only the part of the gel containingthe g-subunit is shown. (c) The appearance of CF1-g* in thylakoids isolated from spinach leaves, which were vacuuminfiltrated with 20mM dithiothreitol and then incubated in the dark for 40min.
Light-induced thylakoid uncoupling 1333
Discussion
It has recently been shown that a DTT treatment ofisolated thylakoids in the dark followed by alkyla-tion with N-ethylmaleimide caused a partial un-coupling. This had been ascribed to reduction andalkylation of CF1-g (Evron and McCarty, 2000). The
present work shows that light or DTT cause also apartial uncoupling of the thylakoid membrane invivo, which involves two different sites andmechanisms. These mechanisms are representedin changes in the uncoupling ratios u, ue, uH, uATP asdefined in the Materials and methods section.Proton leak and proton slip, as represented in the

ARTICLE IN PRESS
M. Richter et al.1334
uncoupling ratios ue and uH, each account for abouthalf of the total uncoupling (u). Our results fromIAF-labeling of thylakoid proteins support a role ofreduction of CF1-g in the mechanism of the protonleak. Reduction of CF1-g and proton leak resembleeach other in that both were saturated at low lightintensity and both were stable during dark storageof thylakoids after induction at the level of intactleaves.
According to accepted views of the function ofCF1-g reduction in activation of the ATP synthase(Junesch and Graber, 1987; Groth and Strotmann,1999) and with regard to its high oxidizing midpointpotential and reported quick reduction kinetics(Quick and Mills, 1986; Kramer et al., 1990), it wassurprising that only about one-third of CF1-g was inthe reduced state even after the strong lighttreatment. We can exclude the possibility thatthe IAF labeling of (thiol modulated or unmodu-lated) CF1-g was not complete. In this case wewould have expected a larger number of immuno-reactive bands, because the labeling led to asignificant shift in the apparent molecular weight.However, we found no additional band with con-trols and only one very weak additional band in thewestern blots of the HL sample. We also excludethe possibility that the extent of IAF labeling wassignificantly affected by reoxidation of CF1-g as theobserved changes in fluorescence pattern de-pended only on the sample pretreatment but noton the time of storage. The stability of the reducedstate of CF1-g in isolated thylakoids under airsaturated oxygen levels points to an in vivoreoxidation mechanism that involves cellular com-pounds like the ‘‘soluble inhibitory factor’’ ovothiolA in Dunaliella salina (Selman-Reimer et al., 1991)or oxidized thioredoxin (Groth and Strotmann,1999). Although the observed low level of reductionof CF1-g during light treatment in vivo could beexplained by fast reoxidation between the end oflight treatment and the start of the thylakoidpreparation, a limited access of thioredoxin to thefast rotating CF1-g as reported by He et al. (2000) inconnection with continuous oxidation is morelikely.
The uncoupling was less pronounced and CF1-gwas less reduced after the DTT treatment in thedark, which possibly relates to the fact that in vivothe thiol-sensitive disulfide bond of CF1-g is lessaccessible to DTT in the absence of a proton motiveforce (McCarty et al., 1972; McCarty and Fagan,1973; Ketcham et al., 1984).
With regard to the uncoupling related todecreased H+/e� our data do not allow a distinctionbetween redox slip (an electron passes by acoupling site) or proton slip (protons released by
protolytic redox reactions near the lumen areactually discharged into the stroma). A redox slipcould be caused by decreased Q-cycle activity asoutlined in Cramer et al. (1996). However, a loss inQ-cycle activity in connection with the oxidizedstate of P-700 (Schubert et al., 1990) is unlikely,because with the saturating light used in ourelectron transport measurements, P-700 is sup-posed to be in the oxidized state and the Q cycleshould then not operate irrespective of the condi-tion of the leaf pretreatment (see Fig. 6). Inaddition, the values of H+/e� between 2.0 and 1.5suggest that the Q cycle did not function withsaturating actinic light in either thylakoid sampleand values below 2.0 would then only be possible ifa fraction of cyt b6f complexes was completelyinactive in proton pumping. For these reasons, aproton slip seems more reasonable. For example,the reduction of a disulfide bond within the cyt b6fcomplex could have induced a structural changesuch that part of the protons were forced to bedischarged back into the stroma instead of intothe thylakoid lumen space (Fig. 6). Such a protonslip has been described for the cyt bc1 complex ofthe respiratory chain as a result of a directionallycontrolled dynamic proton wire (Bruel et al., 1995).This could explain values of H+/e� below 2.0,although we also consider a systematic under-estimation of the H+/e� ratio in the control andpartly uncoupled sample with the applied method.The proton slip reported here differs from anearlier reported proton slip at the cyt b6f complex(Berry and Rumberg, 1999) in that it was indepen-dent from the reduction state of plastoquinone andstable in isolated thylakoids after light-induction inleaves. The proton slip led to enhanced rates ofelectron transport and is therefore to be distin-guished from a recently reported DTT induciblemechanism of down regulation of electron trans-port located at the cyt b6f complex. This wasreversible at the level of thylakoids and wassupposed to protect the photosynthetic apparatusunder light stress conditions (Johnson, 2003).
The extent of uncoupling calculated from therespective ATP yields (uATP) was significantly smal-ler than the uncoupling ratio (u) calculated fromelectron transport rates (Table 1). This points tothe possibility that only one of two observeduncoupling mechanisms causes the decrease inthe ATP yield. We propose that less efficient protonpumping (uH) is the more likely candidate, becausea loss in the ratio H+/e� would inevitably havecaused a corresponding loss in ATP/2e. On thecontrary, proton leak with reduced CF1-g is sup-posed to leave the ATP yield unaffected, as in thepresence of ADP and Pi it is associated with an

ARTICLE IN PRESS
Figure 6. Scheme of the thylakoid membrane and its major complexes involved in light-induced partial uncoupling.The scheme shows the reactions involved in proton translocation in isolated thylakoids associated with electrontransport from water to either FeCy or MV. Part (a) applies to thylakoids from control leaves and part (b) summarizesthe changes in thylakoids with light or DTT induced proton slip and proton leak. The electron transport to FeCy leads toa linear increase in H+ concentration in a sample equivalent to the protons released at the oxygen evolving complex(OEC). This allows the indirect measurement of electron transport from pH measurements. Electron transport to MVconsumes 1 H+ per H+ released at the OEC, and thus it does not lead to a permanent change of the sample pH. Thestoichiometries given in the scheme are based on two electrons transported and a H+ requirement of 4 per ATP. Thevalues do not reflect changes in rates of the respective reactions. For clarity, most of the electron transfer steps areomitted in the scheme. Because the electron transport was measured with saturating light, most of the cyt f isexpected to be in the oxidized state. According to Schubert et al. (1990) this implies, that the Q cycle activity was lowor even zero. Therefore, the reduction of PQ at the PQ reducing site (Q r) and the associated proton uptake are onlyshown in broken lines, and the stoichiometry of pumped protons varies with the assumed Q cycle activity. The dottedlines indicate possible proton wires within the PS 2 and the cyt b6f complex and their direction of proton translocation. Infact the fraction of cyt b6f with a proton slip due to an uncoupling proton wire is unknown and the fraction of CF1 withreduced subunit g is only 36%. The measured ATP yield of 0.9 per two electrons in samples from light treated leaves istherefore higher than the value of 0.5 expected from part (b) of the scheme. It remains unsolved, whether the measuredproton slip was due to a small fraction of cyt b6f with a permanent uncoupling proton wire connecting the PQH2 oxidizingsite (Q 0) to the Q r site or due to alternating connection of the Q 0 site to the uncoupling proton wire and the lumenalproton wire in a larger fraction. The reduction of the disulfide bond in CF1-g causes a partial uncoupling by increasing theproton conductivity of the ATP synthase. This is indicated by a bold line for the efflux through CF0. However, it is supposedthat with coupled conditions the extra proton efflux is associated with extra ATP synthesis and that the intrinsic ATP yieldper H+ of the ATP synthase remains unchanged. This is indicated by bold line for ATP synthesis.
Light-induced thylakoid uncoupling 1335

ARTICLE IN PRESS
M. Richter et al.1336
increased phosphorylating efflux (Groth and Junge,1993; Evron and McCarty, 2000).
The role of the light-induced decrease in theratio H+/e� remains to be solved. Depending onlight intensity, the associated uncoupling wouldlead to significant ‘‘extra electron transport’’ andexcessive generation of reduction potential rela-tive to ATP synthesis. At first glance this could bepotentially harmful, as it could enhance thegeneration of reactive oxygen species and lead todamage of the photosynthetic apparatus (Richter etal., 1990a). On the other hand, partial uncouplingcould lead to enhanced energy storage in the formof malate (Scheibe, 1987), which could be exportedthrough the so-called malate-valve and finallyserve as a substrate for mitochondrial respiration.Since the respiratory chain operates at a higherratio of ATP/2e than the photosynthetic, the exportof reducing equivalents related to partial thylakoiduncoupling could be an alternative way of thechloroplast to supply ‘‘light’’-energy to the cytosol.It is also worth mentioning that plants use a highDpH as a triggering signal for down regulation ofPhotosystem II, when the photosynthetic apparatusis used to its capacity (Richter et al., 1999b; Hortonet al., 2000). The existence of such regulationmay not easily be reconciled with a needfor a comparatively high proton gradient in ATPsynthesis.
Acknowledgements
We gratefully acknowledge the gift of the antibodyagainst the ATP synthase subunit g by Prof.Dr. Berzborn from the University of Bochum, F.R.G.
References
Berry S, Rumberg B. H+/ATP coupling at the unmodulatedCF0CF1-ATP synthase determined by proton fluxmeasurements. Biochim Biophys Acta 1996;1276:51–6.
Berry S, Rumberg B. Proton to electron stoichiometry inelectron transport of spinach thylakoids. BiochimBiophys Acta 1999;1410:248–61.
Bruel C, Manon S, Guerin Martine, Lemesle-Meunier D.Uncoupling of the bc1 complex in S. cerevisiae; Pointmutations affecting the cytochrome b gene bring newinformation about the structural aspect of the protontranslocation. J Bioenergetics Biomembranes 1995;27:527–39.
Cramer WA, Soriano GM, Ponomarev M, Huang D, ZhangH, Martinez SE, Smith JL. Some new structural aspectsand old controversies concerning the cytochrome b6fcomplex of oxygenic photosynthesis. Annu Rev PlantPhysiol Plant Mol Biol 1996;47:477–508.
Curwiel VB, van Rensen JJS. Influence of photoinhibitionon electron transport and photophosphorylationof isolated chloroplasts. Physiol Plant 1993;89:97–102.
Evron Y, McCarty RE. Simultaneous measurement of DpHand electron transport in chloroplast thylakoids by 9-aminoacridine fluorescence. Plant Physiol 2000;124:407–14.
Fowler CF, Kok B. Determination of H+/e� ratios inchloroplasts with flashing light. Biochim Biophys Acta1976;423:510–23.
Genty B, Biantais J-M, Baker NR. The relationshipbetween the quantum yield of photosynthetic elec-tron transport and quenching of chlorophyll fluores-cence. Biochim Biophys Acta 1989;990:87–92.
Groen BH, Berden JA, van Dam K. Differentiationbetween leaks and slips in oxidative phosphorylation.Biochim Biophys Acta 1990;1019:121–7.
Groth G, Junge W. Proton slip of chloroplast ATPase: itsnucleotide dependence, energetic threshold andrelation to an alternating site mechanism of catalysis.Biochemistry 1993;32:8103–11.
Groth G, Strotmann H. New results about structure,function and regulation of the chloroplast ATPsynthase (Cf0CF1). Physiol Plant 1999;106:142–8.
He X, Miginiac-Maslow M, Sigalat C, Eliane K, Haraux F.Mechanism of activation of the chloroplast ATPsynthase. A kinetic study of the thiol modulation ofisolated ATPase and membrane-bound ATP synthasefrom spinach by Escherichia coli thioredoxin. J BiolChem 2000;275:13250–8.
Horton P, Ruban AV, Wentworth M. Allosteric regulation ofthe light-harvesting system of photosystem II. PhilTrans R Soc London B 2000;355:1362–70.
Johnson GN. Thiol regulation of the thylakoid electrontransport chain—a missing link in the regulation ofphotosynthesis. Biochemistry 2003;42:3040–4.
Junesch U, Graber P. Influence of the redox state and theactivation of the chloroplast ATP synthase on proton-transport-coupled ATP synthesis/hydrolysis. BiochimBiophys Acta 1987;893:275–88.
Junge W, Rumberg B, Schroder H. The necessity of anelectric potential difference and its use for photopho-sphorylation in short flash groups. Eur J Biochem1970;14:575–81.
Ketcham SR, Davenport JW, Warncke K, McCarty RE. Roleof the g-subunit of chloroplast coupling factor 1 in thelight-dependent activation of photophosphorylationand ATPase activity by dithiothreitol. J Biol Chem1984;259:7286–93.
Kramer DM, Wise RR, Frederick JR, Alm DM, Hesketh JD,Ort DR, Crofts AR. Regulation of coupling factoractivity to the activities of other thioredoxin-depen-dent chloroplast enzymes. Photosynth Res 1990;26:213–22.
Laemmli UK. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature1970;227:680–5.
McCarty RE, Fagan J. Light-stimulated incorporation ofN-Ethylmaleimide into coupling factor 1 in spinachchloroplasts. Biochemistry 1973;12:1503–7.

ARTICLE IN PRESS
Light-induced thylakoid uncoupling 1337
McCarty RE, Pittman PR, Tsuchiya. Light-dependentinhibition of photophosphorylation by N-Ethylmalei-mide. J Biol Chem 1972;247:3048–51.
Mills JD, Mitchell P. Thiol modulation of the chloroplastprotonmotive ATPase and its effect on photopho-sphorylation. Biochim Biophys Acta 1984;764:93–104.
Nalin CM, McCarty RE. Role of a disulfide bond in the gsubunit in activation of the ATPase of chloroplastcoupling factor 1. J Biol Chem 1984;259:7275–80.
Ogren E, Oquist G. Photoinhibition of photosynthesis inLemna gibba as induced by the interaction betweenlight and temperature. II. Photosynthetic electrontransport. Physiol Plant 1984;62:187–92.
Pick U, McCarty RE. Measurement of membrane DpH..San Pietro A, editor. Methods in Enzymology, Photo-synthesis and Nitrogen Fixation, Vol. 69. New York:Academic Press; 1980. p. 538–46.
Quick WP, Mills JD. Thiol modulation of chloroplastCf0–CF1 in isolated barley chloroplasts and its sig-nificance to regulation of carbon dioxide fixation.Biochim Biophys Acta 1986;851:166–72.
Richter M, Ruhle W, Wild A. Studies on the mechanismof Photosystem II photoinhibition. I. A two stepdegradation of D1-protein. Photosynth Res 1990a;24:229–35.
Richter M, Ruhle W, Wild A. Studies on the mechanism ofPhotosystem II photoinhibition. II. The involvement oftoxic oxygen species. Photosynth Res 1990b;24:237–43.
Richter M, Goss R, Wagner B, Holzwarth AR. Character-ization of the fast and slow reversible components ofnon-photochemical quenching in isolated pea thyla-koids by picosecond time-resolved chlorophyll fluor-escence analysis. Biochemistry 1999;38:12718–26.
Robinson SP. Improved rates of CO2-fixation by intactchloroplasts isolated in media with KCl as osmoticum.Photosynth Res 1986;10:93–100.
Sabbert D, Engelbrecht S, Junge W. Functional and idlingrotatory motion within the F1-ATPase. Proc Natl AcadSci USA 1997;94:4401–5.
Scheibe R. NADP+-malate dehydrogenase in C3-plants:regulation and role of a light-activated enzyme.Physiol Plant 1987;71:393–400.
Schonfeld M, Neumann J. Proton conductance of thethylakoid membrane: modulation by light. FEBS Lett1977;73:51–4.
Schubert K, Liese F, Rumberg B. Analysis of the variabilityof the H/e stoichiometry in spinach chloroplasts..Baltscheffsky M, editor. Current Research in Photo-synthesis, Vol. 3. Dordrecht: Kluwer Academic Publish-ers; 1990. p. 279–82.
Schuldiner S, Rottenberg H, Avron M. Determination ofDpH in chloroplasts. 2. Fluorescent amines as a probefor the determination of DpH in chloroplasts. Eur JBiochem 1972;25:64–70.
Selman-Reimer S, Duhe RJ, Stockman BJ, Selman BR. L-1-N-Methyl-4-Mercaptohistidine disulfide, a potentialendogenous regulator in the redox control of chlor-oplast coupling factor 1 in Dunaliella. J Biol Chem1991;266:182–8.
Strotmann H, Kiefer K, Altvater-Mackensen R. Equilibra-tion of the ATPase reaction of chloroplasts at transi-tion from strong light to weak light. Biochim BiophysActa 1986;850:90–6.
Terashima I, Kashino Y, Katoh S. Exposure of leaves ofCucumis sativus L. to low temperatures in the lightcauses uncoupling of thylakoids. I. Studies with isolatedthylakoids. Plant Cell Physiol 1991;32:1267–74.
Tjus S, Andersson B. Loss of the trans-thylakoid protongradient is an early event during photoinhibitoryillumination of chloroplast preparations. BiochimBiophys Acta 1993;1183:315–22.
Towbin H, Staehelin T, Gordon J. Electrophoretic transferof proteins from polyacrylamide gel to nitrocellulosesheets. Procedure and some applications. Proc NatlAcad Sci USA 1979;76:4350–3.
Yamamoto HY, Kamite L. The effects of dithiothreitol onviolaxanthin de-epoxidation and absorbance changesin the 500-nm region. Biochim Biophys Acta 1972;267:538–43.
Yano H, Wong JH, Lee YM, Cho M-J, Buchanan BB. Astrategy for the identification of proteins targeted bythioredoxin. Proc Natl Acad Sci USA 2001;98:4794–9.
Ziegler R, Egle K. Zur quantitativen Analyse der Chlor-oplastenpigmente. I. Kritische Uberprufung der spek-tralphotometrischen Chlorophyllbestimmung. BeitrBiol Pfl 1965;41:11–37.




















