
Fyn binds to and phosphorylates T cell immunoglobulin and mucin domain-1 (Tim1

Developmental and Comparative Immunology 44 (2014) 186–194
Contents lists available at ScienceDirect
Developmental and Comparative Immunology
journal homepage: www.elsevier .com/locate /dc i
Role of goblet cells and mucin layer in protecting maternal IgAin precocious birds
0145-305X/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.dci.2013.12.010
Abbreviations: BALT, bronchus associated lymphoid tissue; BSA, bovine serumalbumin; CFA, complete Freund’s adjuvant; EC, epithelial cells; GALT, gut associatedlymphoid tissue; GG-pIgR, Gallus gallus polymeric Ig receptor; HRP, horse radishperoxidase; IFA, incomplete Freund’s adjuvant; IgA H, IgA heavy chain; MA,maternal antibodies; PBS, phosphate buffered saline; pIgR, polymeric Ig receptor;SC, secretory component; SQPCR, semi quantitative PCR.⇑ Corresponding author.
E-mail address: [email protected] (A. Friedman).1 He is the incumbent of the Ron Barbaro Chair in Veterinary Medicine.
Enav Bar-Shira, Inbal Cohen, Ori Elad, Aharon Friedman ⇑,1
Animal Sciences, R.H. Smith Faculty of Agriculture, Food and Environment, Hebrew University of Jerusalem, PO Box 12, Rehovot 76100, Israel
a r t i c l e i n f o
Article history:Received 14 November 2013Revised 14 December 2013Accepted 15 December 2013Available online 24 December 2013
Keywords:Maternal antibodiesIgAMucosal immunityGoblet cellAdaptive immunity
a b s t r a c t
Immune protection of the gut in early life depends on provision of maternal antibodies, particularly thatof IgA. In precocial birds (in this study Gallus gallus domesticus) the egg provides the only source ofmaternal antibodies, IgA inclusive. The gut-life of IgA in hatchlings is expected to be brief due to antigenbinding and intestinal washout, and maternal IgA is likely to be depleted prior to immune independenceat 7–10 days of age in the domestic fowl. We followed the track of maternal IgA in mucosal surfaces of thefowl and describe for the first time a mechanism that might provide the means to extend the activeperiod of maternal IgA in the gut. Maternal IgA was located in the gut, lung, and cloacal bursa in embryosand hatchlings prior to the appearance of endogenic IgA positive plasma cells (D3 in the bursa or D7 inthe gut and lung); the source of IgA was most probably the yolk, as the plasma was devoid of IgA till D7post-hatch. The levels of maternal IgA decreased with time, but were still easily determined at the onsetof endogenous IgA production following maturation of the adaptive immune system. Persistence ofmaternal IgA in the gut was enabled by goblet cell up-take by a yet un-described mechanism, and itsconsequent release in a mucin-like layer on enterocyte apical surfaces.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction acquiring systemic protection, while IgA serves to flush the diges-
The immune system, and particularly the adaptive immune sys-tem are immature at time of hatch or birth in all birds and mam-mals studied to date. Consequently, immune protection ofhatchlings or neonates against infection is achieved by the activityof the innate immune system and maternally derived antibodies(maternal antibodies – MA) (Friedman et al., 2012). MA serve asa surrogate for adaptive immunity, as the maturation of adaptiveimmunity might take from days to weeks, depending upon the spe-cies (Friedman et al., 2003).
In mammals, different strategies have been developed to pro-vide MA to the neonate. In primates, MA are delivered via the pla-centa (particularly IgG), colostrum (IgA, IgG and some IgM) andmilk (mainly IgA), while in most other mammals MA are deliveredby colostrum and milk (Hurley and Theil, 2011). IgG from the pla-centa and colostrum is absorbed into the vascular system, thus
tive tract, thus neutralizing gut-derived pathogens (Blum, 2006;Mestecky and Russell, 1998; Mestecky et al., 1999).
Precocious birds have no direct contact with parents afterhatch; hence, the only conduit to supply MA is the egg. As preco-cious birds are self-sustainable immediately upon hatch, mecha-nisms that extend maternal immune protection from eggformation till 14 days of life and more (approximately 5 weeks)are of biological interest. It is presumed that mechanisms that ex-tend MA functionality are active in both circulation and gut orrespiratory tract, though they might be adapted to locale. Interest-ingly, in the domestic fowl (Gallus gallus domesticus), IgY (the avianparallel for IgG) is provided in the yolk, while IgA is deposited inthe albumen as the egg (whether fertilized or not) descends themagnum (Friedman et al., 2012; Kaspers et al., 1996). IgY is mostlyabsorbed from the yolk sac into the embryo’s circulation (Kow-alczyk et al., 1985; Kramer and Cho, 1970), while IgA is transportedfrom the albumen to the yolk (embryonic day 12) (Kaspers et al.,1996), but not into the fetal circulation (Rose and Orlans, 1981).The fate of this IgA is presently unclear, but is probably mostimportant for gut and airway protection, as both lumens becomeloaded with yolk prior to hatch (possibly the gut via the yolk sacstalk and oral as well as rectal intake, and airway by oral intake)(Bencina et al., 2005; Kaspers et al., 1996; Rose et al., 1974).
In the domestic fowl and several other species that have beeninvestigated, the supply of maternal IgY and IgA protects the chickfor as long as three weeks post-hatch (Friedman et al., 2012; King

E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194 187
et al., 2010; Kowalczyk et al., 1985). During this period, adaptiveimmunity matures (5–10 days in the gut, 8–14 days systemically)and takes over the burden of immunological protection (Bar-Shiraet al., 2003; Friedman et al., 2012; King et al., 2010; Kowalczyket al., 1985). While the vascular half-life of IgY can explain the per-sistence of MA in the circulation (Garnier et al., 2012), it is pres-ently unclear whether and how maternal IgA in the gut (orrespiratory tract) outlives the presence of yolk, overwhelmed bycommensal bacteria and food-stuff within 24 h.
IgA is a key factor of the mucosal immune system and functionsin host defense by providing an immune exclusion barrier in lumi-nal secretions (Lamm, 2004). Its primary function is to opsonize,thus it blocks interaction between pathogens and their cellular tar-gets (Mestecky et al., 1983; Snoeck et al., 2006). The biology of IgAhas been best studied in mammals (humans and mice) and the fol-lowing description follows these studies unless otherwise stated.IgA is secreted in the gut lamina propria and is actively transportedto the apical epithelial membrane by a unique transport systeminvolving the polymeric Ig receptor (pIgR). Following transport,IgA bound to a fragment of the transporter, termed the secretorycomponent (SC), is secreted and bound by the glycocalyx and mu-cous layers coating the apical membrane of enterocytes (Kim andHo, 2010). The mucous layers contain various antibacterial sub-stances secreted by goblet cells as well as other cells in the epithe-lial barrier (i.e. Paneth cells) (Kim and Ho, 2010). A possiblehomologue to the mammalian pIgR was recently described in thedomestic fowl, the Gallus gallus polymeric Ig receptor (GG-pIgR)(Wieland et al., 2004). While the GG-pIgR gene was expressed inthe liver, intestine and cloacal bursa, receptor mediated transportof IgA through epithelial cells has yet to be described in birds (altri-cial as well as precocial). Furthermore, there is a paucity of datadescribing the temporal development of the IgA secretory systemin the digestive tract as well as the respiratory tract of hatchlings.
To investigate the development of the IgA secretory system ingallus mucosa, as well as the role of maternal IgA in protection ofthe mucosa, we studied the course of IgA appearance in the gutand airway from late embryonic days up to 7 days post hatch. Inparallel we determined GG-pIgR gene expression in intestinaland airway tissue samples as well as in barrier epithelial cells iso-lated from intestine and cloacal bursae of gallus embryos andchicks. Our findings indicate a previously un-described role formaternal IgA protection in the avian gut and ascribe a possible roleto goblet cells in the mechanism of protection.
2. Materials and methods
2.1. Chickens, embryos and chicks
Three parental sources for fertile fowl eggs were used. Rossbroilers and Cobb broiler eggs were obtained from a commercialhatchery (Efrat, Pardes Hana, Israel and Brown and Sons, HodHasharon, Israel, respectively). Lohman layers and cockerels werehoused in individual cages under constant light and temperatureconditions, and fertilized eggs following artificial inseminationwere collected. All eggs were incubated incubated at 37.5 �C and70% relative humidity in a semi automated incubator (D.M.P engi-neering Ltd, Raanana, Israel).
After hatching, chicks were placed in floor pens on wood shav-ings in isolated, disease-free, light and temperature controlledrooms at 32 �C for the first week post-hatch, followed by 28 �C dur-ing the second week. The feed was a commercial starter formu-lated to meet or exceed NRC requirements (Matmor Feed Co.,Ashdod, Israel). Feed and water were supplied ad libitum for theentire experimental period. Experimental procedures were ap-proved by the Animal Care and Welfare Committee of our Institute.
2.2. Immunization
Layers were immunized 5 times intramuscularly against bovineserum albumin (BSA) (Sigma Chemical Co.). Antigen was emulsi-fied in Freund’s complete adjuvant (CFA, Sigma Chemical Co.) forthe first immunization. Subsequent immunizations were per-formed using Freund’s incomplete adjuvant (IFA, Sigma ChemicalCo.). Each injection contained 500 lg antigen, and was deliveredin multiple sites in the pectoralis major.
2.3. Collection of serum, bile and luminal secretions
Serum samples from embryos and chicks were collected fromcentrifuged-clotted blood obtained from the abdominal vein andfrom the jugular or brachial vein, respectively (n = 3–4 specimensfor each time point). Bile samples were collected at the time of sac-rifice by aspirating the contents of the gall bladder (n = 3–4 foreach time point). Samples containing luminal secretions were col-lected by flushing intestinal segments with PBS. Cloacal bursasecretion samples were obtained from bursae that were invertedand incubated in PBS for 6 h at 37 �C. Luminal and cloacal bursasecretions were temporally collected (n = 3–4 for each time point)to determine IgA and IgY levels. All samples were stored at �20 �Cuntil used.
2.4. Antibody determination in serum, bile and luminal secretions byELISA
IgA and IgY antibody in samples were determined using Chick-en IgA or IgY Quantitative ELISA (Bethyl, Montgomery, TX, USA).ELISA was performed according to instructions by the manufac-turer using affinity purified Goat anti Chicken IgA or IgY (Bethyl,Montgomery, TX, USA) for capture. Blocking was performed byBSA (1% in Phosphate Buffer, Kirkegaard and Perry). Detectionwas performed by Goat anti-Chicken IgA HRP or Goat anti-ChickenIgY HRP conjugates (Bethyl, Montgomery, TX, USA). Chicken refer-ence serum (Bethyl, Montgomery, TX, USA) was used to quantitateantibody levels. Bound antibodies were detected by TMB (Kirkeg-aard and Perry). Optical absorbance was determined at 450 nmusing a spectra II ELISA reader (SLT, Salzburg, Austria).
2.5. Histology and immunohistochemistry
Tissue samples were collected and immediately fixed overnightin 3.5% buffered paraformaldehyde solution (0.1 M, pH 7.4) con-taining 0.1% gluteraldehyde (Frutarom, Acre, Israel) at 4 �C. Thesamples were then dehydrated, embedded in paraffin (ThermoShandon, Pittsburgh, PA, USA). For Immunohistochemistry, 2 lm-thick paraffin sections were deparaffinized, rehydrated andwashed in PBS. Endogenous peroxidase was blocked with 0.3%hydrogen peroxide in methanol. Goat anti-Chicken IgA HRP conju-gate and Goat anti-Chicken IgY HRP conjugates (Bethyl, Montgom-ery, TX, USA) were used to detect IgA and IgY respectively in tissuesamples (Goat anti-mouse Ig H&L negative control is shown inESM-1).
For the detection of BSA specific antibodies in chicken embryos,sections were prepared as described above, however after blockingendogenous peroxidase, sections were incubated with BSA (1 lg/ml), rinsed with PBS and then incubated with affinity purifiedBSA specific IgY (1 lg/ml) prepared in our laboratory (accordingto a protocol by Goldring and Coetzer (2003)). Detection was per-formed by Goat anti-Chicken IgY HRP conjugate. The final chromo-genic reaction was carried out using the peroxidase substrate AEC(Zymed labs, San Francisco, CA, USA) at room temperature for30 min in all slides according to the protocol provided by themanufacturer. Sections were counterstained with hematoxylin

188 E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194
(Thermo, Shandon) and the slides were mounted with coverslipsusing GVA mount (Zymed labs, San Francisco, CA, USA). In negativecontrols, the primary antibody was replaced by goat serum to en-sure specificity. Images were obtained by using a light microscope(Olympus) fixed with a DP72 camera (Olympus).
2.6. Small intestinal and bursa epithelial cell (EC) isolation
Small intestine segments and cloacal bursae were removed (atdifferent days pre- and post-hatch; n = 5 for each time point). Bar-rier epithelial cells from small intestine and bursae were isolatedusing a modification of the technique described by Meddingset al. (1990). After gently rinsing with warm PBS plus 1 mM DTT,small intestine segments were filled with citrate buffer (NaCl,96 mM, sodium citrate 27 mM, KH2PO4 5.6 mM and KCl 1.5 mM)adjusted to a pH of 7.3. Both small intestine segments ends weretied and the segments were incubated in PBS for 2 min at roomtemperature. Bursae were inverted and incubated in citrate bufferfor 2 min at room temperature while shaking. Cells were collectedby gentle centrifugation at 300g. Intestinal and bursa epithelialcells sloughed by this procedure were subjected to RNA extraction.The EC identity of the isolated cells was confirmed by histologicalobservation as well as by expression of villin mRNA. Villin is astructural actin-binding protein expressed in the brush border epi-thelial cells as well as in immature epithelial cells and thus servesas a marker for EC (Athman et al., 2002).
2.7. RNA extraction and PCR analysis
All tissue samples collected for RNA extraction were immedi-ately placed in RNALater� (Ambion, life technologies™ Corpora-tion) until extraction. RNA was extracted from tissues and ECusing TRI Reagent™ (Molecular Research Center Inc.) accordingto the manufacturer’s instructions. Contaminating chromosomalDNA was digested with DNase I (RNAse free; 1 U/lg of RNA; Ther-mo Scientific, Fermentas Molecular Biology Tools) for 20 min at37 �C. RNA quality was assessed using Agilent Bioanalyzer. 0.5–1 lg RNA from each sample was reverse transcribed using iScript™
Advanced cDNA Synthesis Kit for RT-qPCR (Bio-Rad) according tothe manufacturers’ protocols. cDNA was amplified by PCR usingSuperScript™ II Rnase H (Invitrogen, life technologies™ Corpora-tion) (for Semi quantitative PCR) or SsoFast™ EvaGreen� Supermix(BioRad) (for real time PCR) and specific primers for the examinedgenes (see Table 1 for details). Primer sequences were designedaccording to GeneBank published sequences. Each primer pairwas calibrated to determine the optimal reaction temperature
Table 1Gene primers.
Gene Primer pair
IgA heavy chain Sense-50GGGCTGGAGTGGGTCGCTGGT-30
Antisense-50-GAGGCGGAGGAGACGATGA -30
GAPDH Sense-50-AAGGGTGGTGCTAAGCGTG-30
Antisense-50-ATGGCATGGACAGTGGTCATAA-30
18S Sense-50-GCGCAAGACGAACTAAAGC-30
Antisense-50-GGTGAGGTTTCCCGT-30
28S Sense-5’-TTACGGTACTTGTTGGCTATCGG-3’Antisense-5’-TTCTCGGAGTCGGGTTGCT-3’
b Actin Sense -5’-CCACAGCTGCCTCTAGCTCTTC-3’Antisense 5’-AGGACTCCATACCCAAGAAAGATG-3’
GG-pIgR Sense- 5’- AAGATGCTGGCCTGTACCT-3’Antisense- 5’- CGGGTCGTAGTGGCAATCAAT-3’
Cytokeratin18 Sense-5’- ATGGGGGTGGTTTGCTGCTGT -3’Antisens-5’- GGGAGCAGAGTTCGGTCTTGT -3
Villin Sense- 5’-TAACCTGGGCGATGTCTTCC-3’Antisense-5’-CCACCCGCCAGACCTCT-3’
* Semi-quantitative PCR.
and cDNA concentration. Expression levels of examined geneswere determined by either semi quantitative (SQ) PCR or RT-PCR.SQ-PCR amplification was calibrated so that the optimal numberof cycles would allow detection of the appropriate mRNA tran-scripts, while keeping amplification for the examined genes inthe log phase (primer dropping method) (Wong et al., 1994). GAP-DH or 18S served as internal standard (present in all tubes of theassay) in all semi quantitative PCR reactions. These PCR reactionswere performed using a Programmable Thermal Controller (MJ re-search INC. Waltham, MA, USA).
RT-PCR was performed using C1000 Thermal Cycler (Bio-Rad).Results were analyzed using Bio Rad CFX manager™ software. Dis-sociation curve analysis was performed at the end of each real timePCR reaction to validate the presence of a single reaction productand lack of primer dimerization. Expression levels of examinedgenes were normalized using at least 2 normalizing genes (18sand 28s).
2.8. Statistical analysis
Statistical analyses was performed with JMP� software (SAS�
Institute Inc.) using Student’s T Test or ANOVA in conjunction withthe Tukey–Kramer method to determine significance of differencesbetween group mean values; comparisons were made between theexperimental group and respective controls.
3. Results
Previous studies had shown that during embryonic develop-ment, IgA was transferred from the albumen via the amniotic fluiddirectly to the embryonic intestine (Hamal et al., 2006; Kasperset al., 1991, 1996; Rose and Orlans, 1981). Consequently, we ex-pected to detect IgA in both intestinal washings as well as alongthe intestinal epithelial barrier. Initially, we measured IgA anti-body levels in washings obtained from digestive tracts of post-hatch chicks, from hatch up to d8 of age; these levels were com-pared to IgA levels in serum and bile. Results in Fig. 1 showdetectable and significant levels of IgA in the intestine of chicksduring the immediate post hatch period (days 0–3 post hatch).IgA levels decreased dramatically thereafter between d4 and d6post hatch. On day 8 post-hatch, IgA levels increased to levels sim-ilar to those observed at hatch. A similar pattern was observed inbiliary IgA; here E18–E20 were included too, to show that IgA lev-els decreased during the immediate pre-hatch period. The sourceof these antibodies was not the blood borne, as the serum was
Expected product length Gene bank accession number
245 bp S40610
206 bp NM204305
292 bp AF173612
101 bp DQ018756.1
132 bp L08165
164 bp AF303371
244 bp BG711525
741 bp* J03781

Fig. 1. Intestinal, serum and bile IgA antibody levels during the first week posthatch. IgA in intestinal secretions, bile and serum (n = 3) was determined byquantitative ELISA. Antibodies were determined in individual birds and embryos,and group averages ± SEM were calculated. The figure portrays the averagedaverage of three different experiments ± SEM. Letters denote significant differencesin IgA levels (p 6 0.05).
E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194 189
devoid of detectable IgA up to day 6-post hatch, increasing dra-matically thereafter (Fig. 1). These results indicate a significantpresence of IgA in the digestive tract (including bile and bursa
cavity) of embryos and hatchlings; this IgA was purported to beof maternal origin.
Next, we determined the presence and location of IgA and IgAsecreting cells in intestinal mucosal tissue. Temporal staining ofduodenal IgA increased with age (Fig. 2): Initially IgA was locatedonly at the mucosal surface (goblet cells and epithelial apical lining– Fig. 2: E19 and D0), and later appeared in plasma cells located inthe lamina propria as well (Fig. 2: D7–D28). The number of gobletcells that stained positive for IgA increased with age. To test if thisobservation was unique to IgA, we stained intestinal sections withgoat polyclonal antibodies specific for IgY (Fig. 3). On day of hatchIgY was limited to intestinal vasculature (not shown), while on day7-post hatch IgY-positive plasma cells were observed in the laminapropria as well (Fig. 3: A–B); while the lamina propria (and submucosa) contained abundant IgY-positive plasma cells, at no timeor location did we observe IgY in the lumen, apical coating or ingoblet cells – these were unique locales for IgA alone (Fig 3C). Thus,both IgA and IgY positive plasma cells are initially detected in thedigestive tract of D7 birds. Goblet cells staining positive for IgAwere located in all gut sections, and appeared to be most abundant,at all ages tested, in the colon (Fig. 4B and C). Careful scrutiny ofgut sections (irrespective of age) revealed numerous goblet cellsdischarging IgA-positive content that spread out on enterocyte sur-faces, thus explaining the presence of IgA on the apical membraneof enterocytes (Fig. 4A, inserts 1–3, B and C). In the embryonic lung(Fig. 5), in the absence of IgA positive plasma cells and in the ab-sence of vascular IgA (arrows, Panel B), IgA was observed in the lu-men of alveoli and on the apical coating of the bronchiolarepithelium (panels A and B, respectively). IgA-positive plasma cellsappeared on D7 of life, and were located in bronchiolar lamina pro-pria (Arrow, Panel C; primarily near primary and secondary bron-chioles). A similar observation was made in the cloacal bursa(Fig. 6): In the late embryonic period (panel A) and on day of hatch(panel B), IgA was identified in the coating overlaying the apicalsurface of bursa epithelium, in the absence of any local IgA-positiveplasma cells or positive intra-tissue staining. Plasma cells positivefor IgA were detected in bursa tissue from d3 post hatch (panels C–F). At this time staining of IgA was apparent throughout the epithe-lial layer; this was indicative of IgA epithelial transport (panels C–E). These results continue to support the possibility that pre-hatchand immediate post-hatch IgA was of maternal origin.
As IgA secreting plasma cells appear in the intestine only at theend of the first week of life (first 3 days of life in the bursa), wesought to confirm that prior to d7, IgA observed in the intestinewas of maternal origin. To directly demonstrate this, hens wereimmunized against BSA till they became hyper-immune (notshown). Then they were inseminated, their eggs collected andincubated. Controls were fertile eggs from BSA negative hens. OnE19 all embryos were sacrificed, intestinal tissue was removedand used for RNA extraction and for immunohistochemistry. Thinsections prepared for immunohistochemistry were examined forthe presence of BSA specific IgA. Results shown in Fig. 7A show thatintestinal tissue obtained from the progeny of BSA immunizedhens did not express IgA H mRNA in their digestive tract. However,the same tissue stained positive for IgA (Fig. 7B). Moreover, the tis-sue obtained from the progeny of BSA immunized dams stainedpositive for anti-BSA IgA (Fig. 7C) while that obtained from theprogeny of non-immune dams was negative for anti-BSA specificIgA (Fig. 7D). The IgA anti-BSA specific staining was observed onthe apical side of the enterocytes as well as in goblet cells similarto the staining pattern observed for total IgA (Fig. 7B and C). Asgoblet cells were negative for IgY antibodies (see Fig. 3), we con-cluded that the staining of goblet cells was due to maternal IgAanti-BSA specific antibodies enclosed within the cytoplasm ofembryonic goblet cells.

Fig. 2. Temporal staining of intestinal IgA. Thin sections (2 lm) were prepared from duodena obtained from chicks at different ages (n = 3 for each age). Goat polyclonalantibodies specific for chicken IgA were used to detect IgA and IgA positive plasma cells (PC) in duodenal sections. In all panels magnification is �20.
Fig. 3. Staining of intestinal IgY and IgY secreting plasma cells. Panel A (�20) demonstrates IgY staining in a large vein located in the adventitia. Panel B (�20) demonstratesIgY plasma cells in the lamina propria, slightly above a villus lymphoid follicle. Panel C (�40) shows numerous IgY-positive plasma cells; enterocytes and goblet cells arenegative for IgY specific staining. Sections were prepared as described in Section 2 and were stained with goat polyclonal antibodies specific for chicken IgY.
Fig. 4. Discharge of goblet-cell mucous containing IgA. Panel A (�20; insets are at �60) demonstrates IgA discharge in the duodenum, and panels B and C (�60) demonstrateIgA discharge in the colon. Sections were prepared and stained as described in Section 2.
Fig. 5. Presence of IgA and IgA-positive plasma cells in lung. Thin sections (2 lm) were prepared from lungs obtained from chicks at different ages (n = 3 for each age). Goatpolyclonal antibodies specific for chicken IgA were used to detect IgA and IgA positive plasma cells (PC) in lung sections. A & B = E19 (�20); Arrows in B indicate blood vessels,C = D7 (�20); Arrow indicates IgA positive plasma cells.
190 E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194
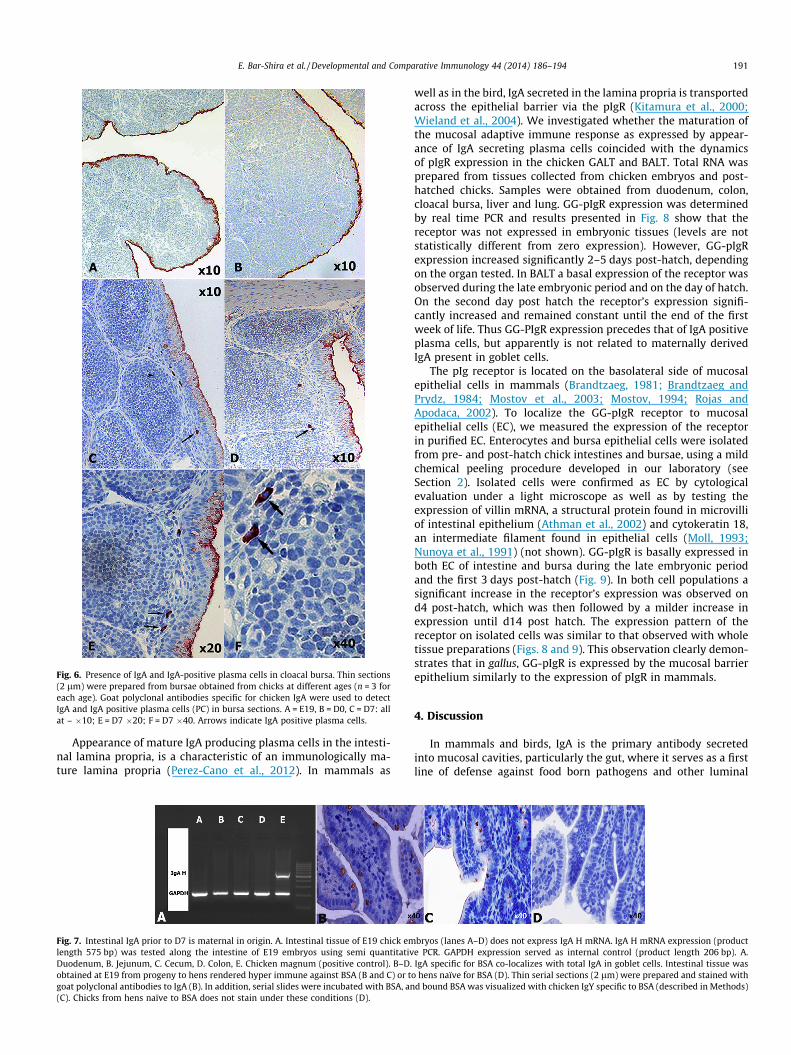
Fig. 6. Presence of IgA and IgA-positive plasma cells in cloacal bursa. Thin sections(2 lm) were prepared from bursae obtained from chicks at different ages (n = 3 foreach age). Goat polyclonal antibodies specific for chicken IgA were used to detectIgA and IgA positive plasma cells (PC) in bursa sections. A = E19, B = D0, C = D7: allat – �10; E = D7 �20; F = D7 �40. Arrows indicate IgA positive plasma cells.
E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194 191
Appearance of mature IgA producing plasma cells in the intesti-nal lamina propria, is a characteristic of an immunologically ma-ture lamina propria (Perez-Cano et al., 2012). In mammals as
Fig. 7. Intestinal IgA prior to D7 is maternal in origin. A. Intestinal tissue of E19 chick emlength 575 bp) was tested along the intestine of E19 embryos using semi quantitativDuodenum, B. Jejunum, C. Cecum, D. Colon, E. Chicken magnum (positive control). B–D.obtained at E19 from progeny to hens rendered hyper immune against BSA (B and C) or tgoat polyclonal antibodies to IgA (B). In addition, serial slides were incubated with BSA, an(C). Chicks from hens naïve to BSA does not stain under these conditions (D).
well as in the bird, IgA secreted in the lamina propria is transportedacross the epithelial barrier via the pIgR (Kitamura et al., 2000;Wieland et al., 2004). We investigated whether the maturation ofthe mucosal adaptive immune response as expressed by appear-ance of IgA secreting plasma cells coincided with the dynamicsof pIgR expression in the chicken GALT and BALT. Total RNA wasprepared from tissues collected from chicken embryos and post-hatched chicks. Samples were obtained from duodenum, colon,cloacal bursa, liver and lung. GG-pIgR expression was determinedby real time PCR and results presented in Fig. 8 show that thereceptor was not expressed in embryonic tissues (levels are notstatistically different from zero expression). However, GG-pIgRexpression increased significantly 2–5 days post-hatch, dependingon the organ tested. In BALT a basal expression of the receptor wasobserved during the late embryonic period and on the day of hatch.On the second day post hatch the receptor’s expression signifi-cantly increased and remained constant until the end of the firstweek of life. Thus GG-PIgR expression precedes that of IgA positiveplasma cells, but apparently is not related to maternally derivedIgA present in goblet cells.
The pIg receptor is located on the basolateral side of mucosalepithelial cells in mammals (Brandtzaeg, 1981; Brandtzaeg andPrydz, 1984; Mostov et al., 2003; Mostov, 1994; Rojas andApodaca, 2002). To localize the GG-pIgR receptor to mucosalepithelial cells (EC), we measured the expression of the receptorin purified EC. Enterocytes and bursa epithelial cells were isolatedfrom pre- and post-hatch chick intestines and bursae, using a mildchemical peeling procedure developed in our laboratory (seeSection 2). Isolated cells were confirmed as EC by cytologicalevaluation under a light microscope as well as by testing theexpression of villin mRNA, a structural protein found in microvilliof intestinal epithelium (Athman et al., 2002) and cytokeratin 18,an intermediate filament found in epithelial cells (Moll, 1993;Nunoya et al., 1991) (not shown). GG-pIgR is basally expressed inboth EC of intestine and bursa during the late embryonic periodand the first 3 days post-hatch (Fig. 9). In both cell populations asignificant increase in the receptor’s expression was observed ond4 post-hatch, which was then followed by a milder increase inexpression until d14 post hatch. The expression pattern of thereceptor on isolated cells was similar to that observed with wholetissue preparations (Figs. 8 and 9). This observation clearly demon-strates that in gallus, GG-pIgR is expressed by the mucosal barrierepithelium similarly to the expression of pIgR in mammals.
4. Discussion
In mammals and birds, IgA is the primary antibody secretedinto mucosal cavities, particularly the gut, where it serves as a firstline of defense against food born pathogens and other luminal
bryos (lanes A–D) does not express IgA H mRNA. IgA H mRNA expression (producte PCR. GAPDH expression served as internal control (product length 206 bp). A.IgA specific for BSA co-localizes with total IgA in goblet cells. Intestinal tissue was
o hens naïve for BSA (D). Thin serial sections (2 lm) were prepared and stained withd bound BSA was visualized with chicken IgY specific to BSA (described in Methods)

Fig. 8. GG-pIgR mRNA expression in GALT, liver and BALT pre- and post-hatch.Tissue samples were obtained from embryos (e) and chicks (d) at different ages(n = 3). Total RNA was extracted and reverse transcribed to cDNA. Transcriptionallevels of GG-pIgR were determined by quantitative real-time PCR. Data areexpressed as mRNA levels normalized to 28S and 18S mRNA levels. GG-pIgRexpression was determined in individual birds (duplicate determinations) andgroup averages ± SEM were calculated. The figure portrays the averaged average ofthree different experiments ± SEM. Letters denote significant differences in GG-pIgRexpression (p 6 0.05).
192 E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194
antigens (Woof and Kerr, 2006). IgA is important for the establish-ment of intestinal immune homeostasis and immune protection, aswell as for the control of autoreactive, proinflammatory, and neo-plastic B cell clones present in both the intestine and the periphery(Cerutti and Rescigno, 2008). In the neonate, and presumably in theavian hatchling, maternal IgA serves identical functions until atime when host-derived IgA has taken over. In the mammal andother altricial organisms, the pre-weaning intake of maternal milkor saliva provides continuous and adequate IgA protection (thoughdifferent mechanisms might apply to altricial birds – see (Pihlajaet al., 2006)). However, in precocial animals such as the domesticfowl, the only source of maternal antibodies, IgA inclusive, is thatprovided in the egg. This ‘‘one time’’ provision needs to protectthe embryo and hatchling till the maturation of the mucosal adap-tive immune response. In this manuscript we undertook to follow
the development of the IgA secretory system in the domestic fowl.Our study focused mainly on the digestive tract and respiratoryairway, and describes two main periods of development: (a) theperiod under maternal antibody protection (late embryonic devel-opment [E18–E19] up to the appearance of host IgA positive plas-ma cells), (b) The period following the maturation of the intestinaladaptive immune response. As we have shown here, the time ofdemarcation, namely the maturation of mucosal immunity, occurs3 (bursa) to 7 (lung and intestine) days post hatch. Thus, in thefowl, maternal IgA is required to protect the mucosa for close toa week post hatch.
The tissue half-life of IgA is yet to be measured, but its intestinalhalf-life is expected to be considerably shorter due to antigen bind-ing as well as intestinal flushing. Consequently, if maternal IgA isrequired to protect the mucosa in the chick, a means to extendthe intestinal half-life of IgA must exist. As we have shown in thismanuscript, maternally derived IgA was indeed directed towardsmucosal surfaces, and was detected as early as E19 (the earliesttime point investigated in this study). Maternal IgA persisted, al-beit at decreasing levels in washings, in the dense mucous coatingof surface EC as well as within goblet cells. We suggest that persis-tence of maternal IgA is made possible due to a unique, and previ-ously un-described, uptake by goblet cells where it serves as areservoir and is concomitantly secreted along with goblet cell mu-cin secretions. Thus, maternal IgA is selectively captured (IgY wasnot detected in mucin or goblet cells – Fig. 3) and confined tothe enterocyte surface, thereby focusing its area of protectionand limiting its loss due to intestinal flushing.
The role of the goblet cell in intestinal innate immunity is a rel-atively novel concept (McDole et al., 2012). In the mammal there isno account, to the best of our knowledge, of IgA positive gobletcells, while in the chicken, a single report by Kincade and Coopernoted IgA positive goblet cells (Kincade and Cooper, 1973). Mucinssecreted by goblet cells create a dynamic physical barrier and pro-vide an area in which growth factors, cytokines, and chemokinesare sequestered (Lievin-Le Moal and Servin, 2006). Mucin trappedsoluble host defense molecules serve to control or deter pathogeninvasion from the underlying mucosal epithelial cells (Moncadaet al., 2003). Hence it is plausible that mucins are able to sequesteror bind luminal IgA. Absorption of IgA by mucin inside goblet cellsprovides a mechanism for slow release into the mucin layer, thusprotecting against rapid depletion after hatching. The mechanismby which IgA enters goblet cells is unknown and requires furtherinvestigation. A possible mechanism is that IgA binds to a cognatereceptor on the goblet cell apical membrane. GG-pIgR is the avianhomologue of pIgR found in mammals (Wieland et al., 2004). Pre-vious studies have shown that this receptor is expressed by intes-tinal tissue of juvenile chickens (Lammers et al., 2010). In thepresent study we have extended these observations to show thatthe receptor is expressed in isolated EC from the gut and cloacalbursa, and that its expression pattern was similar to its expressionin whole tissue preparations. However, our current studies showthat detectable expression of GG-pIgR occurred post-hatch; hence,an alternative mechanism might explain IgA uptake by goblet cellsin the avian gut. Our future research is aimed to decipher gobletcells IgA uptake, and whether the putative receptor it is expressed(at least in the hatchling) on apical membranes.
No IgA was observed in serum during the first week of life, andthis was concomitant with a temporal decrease of intestinal andbile IgA levels during the first week post-hatch. IgA in serum aswell as an increase in intestinal washings followed d8 post-hatch(Fig. 1). The increase in serum IgA levels support the results of Ha-mal et al. who demonstrated an increase in plasma IgA in 7d oldchicks (Hamal et al., 2006). These results also support previouslypublished reports showing that early IgA secreting plasma cells ap-pear in the intestine and spleen about 6 days post-hatch (Lawrence

Fig. 9. GG-pIgR mRNA expression in isolated epithelial cells. Intestinal and bursa EC were isolated by gentle chemical peeling from embryos (e) and chicks (d). Cells collectedfrom 3–5 animals were pooled, total RNA was extracted and reverse transcribed to cDNA. Transcriptional levels of GG-pIgR were determined by quantitative real-time PCR.Data are expressed as normalized mRNA levels to 28S and 18S mRNA levels of duplicate determinations. The figure portrays the averaged average of four differentexperiments ± SEM. Letters denote significant differences in GG-pIgR expression (p 6 0.05).
E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194 193
et al., 1981; Yamamoto et al., 1977). Previous data suggested thatmaturation of adaptive immunity in the respiratory system followsthat of the gut (Fagerland and Arp, 1993; Lawrence et al., 1981). Incontrast, we show here that the appearance of plasma cells in therespiratory system and gut occurs at the same time.
We have shown that maternal IgA persists in the fowl’s gut andrespiratory system until maturation of the host’s IgA response. Thismaturation is primarily observed in the cloacal bursa, and spreadsthereafter to the entire gut and respiratory system as well as thecirculation. The maturation of the avian IgA transporter precedesthat of IgA secretion, and thus it, or an alternative receptor systemmight be connected to the sequestration and protection of
maternal IgA in the mucosal mucin layer and specialized cells suchas the intestinal goblet cell. The possible involvement of the gobletcell in precocial avian immunity is exciting as it implicates this cellin immune related functions similar to those described in themammal (McDole et al., 2012).
Acknowledgements
These studies were supported in part by a grant from the IsraelScience Foundation, and in part by a grant from the Head Scientist– Ministry of Agriculture.

194 E. Bar-Shira et al. / Developmental and Comparative Immunology 44 (2014) 186–194
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.dci.2013.12.010.
References
Athman, R., Louvard, D., Robine, S., 2002. The epithelial cell cytoskeleton andintracellular trafficking. III. How is villin involved in the actin cytoskeletondynamics in intestinal cells? Am. J. Physiol. Gastrointest. Liver Physiol. 283,G496–G502.
Bar-Shira, E., Sklan, D., Friedman, A., 2003. Establishment of immune competence inthe avian GALT during the immediate post-hatch period. Dev. Comp. Immunol.27, 147–157.
Bencina, D., Narat, M., Bidovec, A., Zorman-Rojs, O., 2005. Transfer of maternalimmunoglobulins and antibodies to Mycoplasma gallisepticum and Mycoplasmasynoviae to the allantoic and amniotic fluid of chicken embryos. Avian Pathol.34, 463–472.
Blum, J.W., 2006. Nutritional physiology of neonatal calves. J. Anim. Physiol. Anim.Nutr. 90, 1–11.
Brandtzaeg, P., 1981. Transport models for secretory IgA and secretory IgM. Clin.Exp. Immunol. 44, 221–232.
Brandtzaeg, P., Prydz, H., 1984. Direct evidence for an integrated function of J chainand secretory component in epithelial transport of immunoglobulins. Nature311, 71–73.
Cerutti, A., Rescigno, M., 2008. The biology of intestinal immunoglobulin aresponses. Immunity 28, 740–750.
Fagerland, J.A., Arp, L.H., 1993. Distribution and quantitation of plasma cells, Tlymphocyte subsets, and B lymphocytes in bronchus-associated lymphoidtissue of chickens: age-related differences. Regional Immunol. 5, 28–36.
Friedman, A., Bar-shira, E., Sklan, D., 2003. Ontogeny of gut associated immunecompetence in the chick. World’s Poult. Sci. J. 59, 209–219.
Friedman, A., Elad, O., Cohen, I., Bar Shira, E., 2012. The gut associated lymphoidsystem in the post-hatch chick: dynamics of maternal IgA. Israel J. Vet. Med. 67,75.
Garnier, R., Ramos, R., Staszewski, V., Militão, T., Lobato, E., González-Solís, J.,Boulinier, T., 2012. Maternal antibody persistence: a neglected life-history traitwith implications from albatross conservation to comparative immunology.Proc. R. Soc. B: Biol. Sci. 279, 2033–2041.
Goldring, J.P.D., Coetzer, T.H.T., 2003. Isolation of chicken immunoglobulins (IgY)from egg yolk⁄. Biochem. Mol. Biol. Educ. 31, 185–187.
Hamal, K.R., Burgess, S.C., Pevzner, I.Y., Erf, G.F., 2006. Maternal antibody transferfrom dams to their egg yolks, egg whites, and chicks in meat lines of chickens.Poult. Sci. 85, 1364–1372.
Hurley, W.L., Theil, P.K., 2011. Perspectives on immunoglobulins in colostrum andmilk. Nutrients 3, 442–474.
Kaspers, B., Schranner, I., Losch, U., 1991. Distribution of immunoglobulins duringembryogenesis in the chicken. Zent. Veterinarmed. Reihe A 38, 73–79.
Kaspers, B., Bondl, H., Göbel, T.W.F., 1996. Transfer of IgA from albumen into theyolk sac during embryonic development in the chicken⁄. J. Vet. Med. 43, 225–231.
Kim, Y.S., Ho, S.B., 2010. Intestinal goblet cells and mucins in health and disease:recent insights and progress. Curr. Gastroenterol. Rep. 12, 319–330.
Kincade, P.W., Cooper, M.D., 1973. Immunoglobulin A: site and sequence ofexpression in developing chicks. Science 179, 398–400.
King, M.O., Owen, J.P., Schwabl, H.G., 2010. Are maternal antibodies really thatimportant? Patterns in the immunologic development of altricial passerinehouse sparrows (Passer domesticus). PLoS One 5, e9639.
Kitamura, T., Garofalo, R.P., Kamijo, A., Hammond, D.K., Oka, J.A., Caflisch, C.R.,Shenoy, M., Casola, A., Weigel, P.H., Goldblum, R.M., 2000. Human intestinalepithelial cells express a novel receptor for IgA. J. Immunol. 164, 5029–5034.
Kowalczyk, K., Daiss, J., Halpern, J., Roth, T.F., 1985. Quantitation of maternal-fetalIgG transport in the chicken. Immunology 54, 755–762.
Kramer, T.T., Cho, H.C., 1970. Transfer of immunoglobulins and antibodies in thehen’s egg. Immunology 19, 157–167.
Lamm, M.E., 2004. Immunoglobulins. In: Leonard, J. (Ed.), Encyclopedia ofGastroenterology. Elsevier, New York, pp. 430–431.
Lammers, A., Wieland, W.H., Kruijt, L., Jansma, A., Straetemans, T., Schots, A., denHartog, G., Parmentier, H.K., 2010. Successive immunoglobulin and cytokineexpression in the small intestine of juvenile chicken. Dev. Comp. Immunol. 34,1254–1262.
Lawrence, E.C., Arnaud-Battandier, F., Grayson, J., Koski, I.R., Dooley, N.J., Muchmore,A.V., Blaese, R.M., 1981. Ontogeny of humoral immune function in normalchickens: a comparison of immunoglobulin-secreting cells in bone marrow,spleen, lungs and intestine. Clin. Exp. Immunol. 43, 450–457.
Lievin-Le Moal, V., Servin, A.L., 2006. The front line of enteric host defense againstunwelcome intrusion of harmful microorganisms: mucins, antimicrobialpeptides, and microbiota. Clin. Microbiol. Rev. 19, 315–337.
McDole, J.R., Wheeler, L.W., McDonald, K.G., Wang, B.M., Konjufca, V., Knoop, K.A.,Newberry, R.D., Miller, M.J., 2012. Goblet cells deliver luminal antigen toCD103(+) dendritic cells in the small intestine. Nature 483 (345–U141).
Meddings, J.B., DeSouza, D., Goel, M., Thiesen, S., 1990. Glucose transport andmicrovillus membrane physical properties along the crypt–villus axis of therabbit. J. Clin. Invest. 85, 1099–1107.
Mestecky, J., Russell, M.W., 1998. Passive and active protection against disorders ofthe gut. Vet. Quart. 20 (Suppl. 3), S83–S87.
Mestecky, J., Kutteh, W.H., Brown, T.A., Russell, M.W., Phillips, J.O., Moldoveanu, Z.,Moro, I., Crago, S.S., 1983. Function and biosynthesis of polymeric IgA. Ann. N. Y.Acad. Sci. 409, 292–306.
Mestecky, J., Russell, M.W., Elson, C.O., 1999. Intestinal IgA: novel views on itsfunction in the defence of the largest mucosal surface. Gut 44, 2–5.
Moll, R., 1993. Cytokeratins as markers of differentiation. Expression profiles inepithelia and epithelial tumors. Veroffentlichungen aus der Pathologie 142, 1–197.
Moncada, D.M., Kammanadiminti, S.J., Chadee, K., 2003. Mucin and toll-likereceptors in host defense against intestinal parasites. Trends Parasitol. 19,305–311.
Mostov, K.E., 1994. Transepithelial transport of immunoglobulins. Annu. Rev.Immunol. 12, 63–84.
Mostov, K., Su, T., ter Beest, M., 2003. Polarized epithelial membrane traffic:conservation and plasticity. Nat. Cell Biol. 5, 287–293.
Nunoya, T., Tajima, M., Itakura, C., 1991. Primary culture of chicken bursal plicalepithelium. Res. Vet. Sci. 50, 352–354.
Perez-Cano, F.J., Franch, A., Castellote, C., Castell, M., 2012. The suckling rat as amodel for immunonutrition studies in early life. Clin. Dev. Immunol. 2012, 16–32.
Pihlaja, M., Siitari, H., Alatalo, R.V., 2006. Maternal antibodies in a wild altricial bird:effects on offspring immunity, growth and survival. J. Anim. Ecol. 75, 1154–1164.
Rojas, R., Apodaca, G., 2002. Immunoglobulin transport across polarized epithelialcells. Nat. Rev. Mol. Cell Biol. 3, 944–955.
Rose, M.E., Orlans, E., 1981. Immunoglobulins in the egg, embryo and young chick.Dev. Comp. Immunol. 5, 15–20.
Rose, M.E., Orlans, E., Buttress, N., 1974. Immunoglobulin classes in the hen’s egg:their segregation in yolk and white. Eur. J. Immunol. 4, 521–523.
Snoeck, V., Peters, I.R., Cox, E., 2006. The IgA system: a comparison of structure andfunction in different species. Vet. Res. 37, 455–467.
Wieland, W.H., Orzaez, D., Lammers, A., Parmentier, H.K., Verstegen, M.W., Schots,A., 2004. A functional polymeric immunoglobulin receptor in chicken (Gallusgallus) indicates ancient role of secretory IgA in mucosal immunity. Biochem. J.380, 669–676.
Wong, H., Anderson, W.D., Cheng, T., Riabowol, K.T., 1994. Monitoring mRNAexpression by polymerase chain reaction: the ‘‘primer-dropping’’ method. Anal.Biochem. 223, 251–258.
Woof, J.M., Kerr, M.A., 2006. The function of immunoglobulin A in immunity. J.Pathol. 208, 270–282.
Yamamoto, H., Watanabe, H., Mikami, T., 1977. Distribution of immunoglobulin andsecretory component containing cells in chickens. Am. J. Vet. Res. 38, 1227–1230.
Copyright © 2022 FDOKUMEN




















