
Response of foliar metabolism in mature trees of< i> Quercus pubescens and< i> Quercus ilex to...
13
Environmental and Experimental Botany 39 (1998) 233 – 245 Response of foliar metabolism in mature trees of Quercus pubescens and Quercus ilex to long-term elevated CO 2 Roberto Tognetti a,b, *, Jon D. Johnson c , Marco Michelozzi d , Antonio Raschi a a Istituto per l’Agrometeorologia e l’Analisi Ambientale applicata all’Agricoltura, Consiglio Nazionale delle Ricerche, Via Caproni 8, Firenze 50145, Italy b Department of Botany, Trinity College, Uni6ersity of Dublin, Dublin 2, Ireland c Intensi6e Forestry Program, Washington State Uni6ersity, Puyallup, WA 98371 -4998, USA d Istituto Miglioramento Genetico delle Piante Forestali, Consiglio Nazionale delle Ricerche, Via Vannucci 13, Firenze 50134, Italy Received 15 October 1997; received in revised form 8 January 1998; accepted 9 January 1998 Abstract Long-term effects on and adaptations of the carbon physiology of long-lived trees exposed to increasing atmospheric levels of CO 2 are unknown. We compared two indigenous Quercus species, Q. ilex and Q. pubescens, growing in a natural CO 2 spring located in central Italy and at a nearby control site. In May, 1995 photosynthetic rate at least doubled when measured with supplemental CO 2 in both species and sites. Dark respiration was much higher at the CO 2 spring site in both species. Foliar sugar and starch concentrations in Q. ilex exhibited significant site and diurnal differences (May and September). In July, 1995 there was little difference in the water potential values of the measured trees at the different sites over the diurnal period. Photosynthetic rate was higher for both species in the CO 2 spring, particularly in the early morning and late afternoon. Mid-day stomatal closure reduced photosynthesis to similar levels. In the morning leaf conductance and transpiration were generally lower in the CO 2 spring trees, contributing to higher instantaneous water use efficiency for both species. Isoprene emission rates were higher in Q. pubescens trees growing in the CO 2 spring. The maximum difference between control and CO 2 spring trees occurred in late afternoon. In contrast, Q. ilex exhibited isoprene emission near background level. Foliage and branch carbon and nitrogen status showed increased concentrations of starch and tannins in Q. ilex and of soluble sugars in Q. pubescens in the elevated CO 2 environment, while nitrogen concentration decreased in both species. Wood gravity increased 6 and 3% in Q. ilex and Q. pubescens, respectively, growing in the CO 2 spring. Q. ilex exhibited afternoon recovery of water potential compared to Q. pubescens which had better night-time recovery. Q. ilex and Q. pubescens exposed to elevated CO 2 for prolonged periods exhibit different mechanisms for dealing with additional reduced carbon and do maintain an altered carbon physiology, even in midst of the region’s characteristic summer drought. © 1998 Elsevier Science B.V. All rights reserved. Keywords: Elevated CO 2 ; Gas exchange; Natural CO 2 springs; Quercus ; Secondary compounds * Corresponding author. Tel.: +39 55 301421/301504; fax: +39 55 308910; e-mail: [email protected].fi.cnr.it S0098-8472/98/$19.00 © 1998 Elsevier Science B.V. All rights reserved. PII S0098-8472(98)00013-6
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Response of foliar metabolism in mature trees of< i> Quercus pubescens and< i> Quercus ilex to...

Environmental and Experimental Botany 39 (1998) 233–245
Response of foliar metabolism in mature trees of Quercuspubescens and Quercus ilex to long-term elevated CO2
Roberto Tognetti a,b,*, Jon D. Johnson c, Marco Michelozzi d, Antonio Raschi a
a Istituto per l’Agrometeorologia e l’Analisi Ambientale applicata all’Agricoltura, Consiglio Nazionale delle Ricerche,Via Caproni 8, Firenze 50145, Italy
b Department of Botany, Trinity College, Uni6ersity of Dublin, Dublin 2, Irelandc Intensi6e Forestry Program, Washington State Uni6ersity, Puyallup, WA 98371-4998, USA
d Istituto Miglioramento Genetico delle Piante Forestali, Consiglio Nazionale delle Ricerche, Via Vannucci 13, Firenze 50134, Italy
Received 15 October 1997; received in revised form 8 January 1998; accepted 9 January 1998
Abstract
Long-term effects on and adaptations of the carbon physiology of long-lived trees exposed to increasingatmospheric levels of CO2 are unknown. We compared two indigenous Quercus species, Q. ilex and Q. pubescens,growing in a natural CO2 spring located in central Italy and at a nearby control site. In May, 1995 photosyntheticrate at least doubled when measured with supplemental CO2 in both species and sites. Dark respiration was muchhigher at the CO2 spring site in both species. Foliar sugar and starch concentrations in Q. ilex exhibited significantsite and diurnal differences (May and September). In July, 1995 there was little difference in the water potential valuesof the measured trees at the different sites over the diurnal period. Photosynthetic rate was higher for both speciesin the CO2 spring, particularly in the early morning and late afternoon. Mid-day stomatal closure reducedphotosynthesis to similar levels. In the morning leaf conductance and transpiration were generally lower in the CO2
spring trees, contributing to higher instantaneous water use efficiency for both species. Isoprene emission rates werehigher in Q. pubescens trees growing in the CO2 spring. The maximum difference between control and CO2 springtrees occurred in late afternoon. In contrast, Q. ilex exhibited isoprene emission near background level. Foliage andbranch carbon and nitrogen status showed increased concentrations of starch and tannins in Q. ilex and of solublesugars in Q. pubescens in the elevated CO2 environment, while nitrogen concentration decreased in both species.Wood gravity increased 6 and 3% in Q. ilex and Q. pubescens, respectively, growing in the CO2 spring. Q. ilexexhibited afternoon recovery of water potential compared to Q. pubescens which had better night-time recovery. Q.ilex and Q. pubescens exposed to elevated CO2 for prolonged periods exhibit different mechanisms for dealing withadditional reduced carbon and do maintain an altered carbon physiology, even in midst of the region’s characteristicsummer drought. © 1998 Elsevier Science B.V. All rights reserved.
Keywords: Elevated CO2; Gas exchange; Natural CO2 springs; Quercus ; Secondary compounds
* Corresponding author. Tel.: +39 55 301421/301504; fax: +39 55 308910; e-mail: [email protected]
S0098-8472/98/$19.00 © 1998 Elsevier Science B.V. All rights reserved.
PII S0098-8472(98)00013-6

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245234
1. Introduction
Elevated concentration of atmospheric CO2 hasgenerally been considered beneficial to manyplants species, although there are a few exceptions(Surano et al., 1986; Houpis et al., 1988). Amongthe benefits are increased photosynthetic rates, atleast in the short-term, as well as increased growth(Eamus and Jarvis, 1989; Amthor, 1995). It hasbeen observed that stomatal conductance declinesas CO2 concentration increases, resulting in de-creased transpiration (Jarvis, 1989). Enhancedrates of assimilation and decreased transpirationleads to an increase in instantaneous water useefficiency (Ceulemans and Mousseau, 1994).
After a long-term exposure to high CO2, accli-mated plants may exhibit decreased photosyn-thetic carbon assimilation (Eamus and Jarvis,1989; Ceulemans and Mousseau, 1994). Severalhypotheses have been proposed to explain thisphenomenon including starch accumulation anddecreased amount or activity of rubisco (Amthor,1995). If plants do not acclimate, they must de-velop mechanisms for dealing with the accumula-tion of reduced carbon, such as increased growth(Amthor, 1995), storage in forms of starch andsugars (Korner and Miglietta, 1994; Chaves et al.,1995) or carbon-based secondary compounds(phenolics and tannins) (Johnson et al., 1997;Lawler et al., 1997) or by volatilization of carboncompounds like isoprene or terpenes (Monson etal., 1991; Johnson et al., 1997; Lawler et al.,1997). The studies on plant-carbon dioxide inter-actions have focused on gas exchange because ofdirect interactions between atmospheric CO2 andits fixation in the leaf. However, changes in tissuechemistry, carbon allocation and nutrient use mayinfluence gas exchange performances and growthon a long-term basis. Secondary plant metabolismmay affect plant productivity in response to risingCO2 concentrations. The allocation of carbon tofoliar tannins has been reported to be a conse-quence of pollution stress (Jordan et al., 1991).The relationship between photosynthesis and iso-prene emission is also affected by CO2 level, andon a carbon basis the rate of isoprene emissionmay account for as much as 15% of the carbonfixed in photosynthesis (Sharkey et al., 1991). In
addition, carbohydrate status and carbon econ-omy of other tree species exposed to CO2 enrich-ment have been found to be affected by reductionin dark respiration (Wullschleger et al., 1992). Theability to sustain increases in growth in high CO2
environments appears, therefore, to be dependenton the interaction between carbon assimilation,carbohydrate status, plant respiration and plant’schemical composition, as well as on the availabil-ity of nutrient resources and limiting factors.
In this study the carbon physiology and diurnalwater relations, gas exchange and isoprene emis-sion responses to long-term elevated CO2 werequantified in two indigenous Quercus species, Q.pubescens and Q. ilex, with contrasting leaflongevity (deciduous versus evergreen, respec-tively) and growing in a naturally CO2-enrichedsite in central Italy (Miglietta et al., 1993; Kornerand Miglietta, 1994). Trees that have been grow-ing for many years in naturally CO2-enrichedatmosphere, such as that around this CO2 spring,provide an unique opportunity to assess responsesand adaptations in natural vegetation exposed toelevated atmospheric CO2 levels for durations andCO2 concentrations impossible to simulate experi-mentally (Jones et al., 1995; van Gardingen et al.,1995). Mature trees at the CO2 spring do notgrow faster than trees at ambient CO2 concentra-tion (Korner and Miglietta, 1994). The initialhypothesis of the study was that long-term differ-ences in CO2 atmosphere availability between theCO2 spring and the control site would be reflectedin altered foliage metabolism, in particular carbonphysiology, of trees at the CO2 spring.
2. Materials and methods
2.1. Plant material and study site
The study was conducted at the natural CO2
spring of Bossoleto, near Rapolano (Siena, centralItaly). Gas is emitted from various vents withinthe crater of the CO2 spring, resulting in a CO2
concentration gradient ranging from 90% (bot-tom) to normal (350 mmol mol−1) along thecrater profile (Miglietta et al., 1993; Korner andMiglietta, 1994; van Gardingen et al., 1995). Mea-

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 235
surements were conducted on adjacent trees ofQuercus pubescens Willd. and Quercus ilex L.(height 6–8 m and DBH 10–12 cm). CO2 concen-trations during sampling at the CO2 spring siteranged in day-time hours from 500 to over 1000mmol mol−1 with rapid fluctuations (for moredetails, see van Gardingen et al., 1995). The H2Sand SO2 concentration in the CO2 spring is verylow and cannot be considered harmful to plants(H. Rennenberg, personal communication). Thecontrol site, 1 km from the CO2 spring site, wascharacterized by similar morphology, light expo-sure, soil nutrient availability, moisture anddepth, plant age and association (van Gardingenet al., 1997). Scaffolding and/or natural rocksallowed access to leaves on branches (2–5 mabove ground level) exposed to direct sunlight(south facing) for similar periods of the day.
2.2. Gas exchange and isoprene emissionmeasurements
Trees of Q. pubescens and Q. ilex of similar sizewere selected for sampling in May, 1995 when thenew flush of leaves had just reached full expansionand the drought stress had not been evident yet,because of spring rainfall. Three trees each of Q.ilex and Q. pubescens per site were selected for themeasurements. On May 14, carbon exchange rateswere measured on four attached leaves (1-year-oldleaves on the evergreen Q. ilex and new fullyexpanded leaves on the deciduous Q. pubescens)on each tree of both species using a portableinfra-red gas analyzer (CIRAS-1, PP System,Hitchin, UK). Samples for photosynthesis (An)were taken in mid-morning (temperature 25°Cand PPFD 1800 mmol m−2 s−1), whereas, darkrespiration was measured at dawn (06:30 h) bycovering the cuvette with aluminum foil. The ref-erence CO2 concentration was set at 600 and 350mmol mol−1 for measurements in the CO2 springand control site, respectively (average day-timeCO2 concentration around the crown of CO2
spring and control trees, respectively). Immedi-ately after measuring photosynthesis at ambientconcentration, photosynthesis was then measuredwith cuvette CO2 concentration adjusted to 2000mmol mol−1 (saturating CO2 concentration and
day-time peak around the crown of CO2 springtrees).
Trees of Q. pubescens and Q. ilex of similar sizewere selected for sampling in July, duringdrought. Ages were determined from stem coresafter the study. The Q. ilex trees were between 16and 17 years old and the Q. pubescens trees werebetween 15 and 25 years old (as evaluated by treerings). Gas exchange rates (photosynthesis, tran-spiration and leaf conductance, gl) were measuredon six attached leaves of the same stage of devel-opment on each tree of the two species (tempera-ture ranged from 25 to 36°C, maximum PPFD1800 mmol m−2 s−1). Instantaneous water useefficiency (WUE) was determined as photosynthe-sis/transpiration ratios. Samples were taken every2 h beginning at 10:00 h (local time) on July 6,1995, and continued until ambient light ap-proached 0 mmol m−2 s−1, about 19:00 h. OnJuly 7, sampling continued at 08:00 and 10:00 h.After sampling each tree at a given site, two smallbranches were detached and measured for xylemwater potential using a Scholander-type pressurechamber (PMS, Corvallis, OR). This period waschosen as representative of marked drought stress.
Coincident with the gas exchange measure-ments in July, six equal-aged leaves from each treewere sampled for isoprene emission. Individualleaves were put into separate 20-ml vials andincubated under full sunlight for 1 h in a shallowwaterbath (Monson and Fall, 1989; Kuzma andFall, 1993). One milliliter of head-space gas waswithdrawn, transported to the laboratory in thedark and on ice, and analyzed by flame ionizationgas chromatography (AutoSystem, Perkin-Elmer,Norwalk, CO). Column conditions were: 15-mfused-silica (3-mm coating) capillary column(Megabore DB-1, J and W Scientific, Folsom,CA); highly purified N2 carrier gas at a flow rateof 5 ml min−1; column temperature of 50°C.Liquid isoprene with 99% purity (Aldrich, Mil-waukee, WI) was used to make a gaseous stan-dard. Isoprene was determined by measuring peakheight with comparison of its retention time withthat of authentic isoprene as described above.Emission rates were expressed on a leaf area basisfollowing photocopying of each leaf and convert-ing from dry mass (DW) to area using standardareas.

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245236
2.3. Carbohydrate, nitrogen and tannin analysis
At dawn (06:30 h, local time) and dusk (19:00h) on May 14, several leaves of similar stage ofdevelopment were cut from three branches har-vested from the same Q. ilex trees describedabove. Leaves were dried immediately in a mi-crowave oven for 3 min (Korner and Miglietta,1994). Later the leaves were oven-dried at 60°Cfor 48 h. Leaves were then ground in a Wiley millfitted with a 20-mesh screen. Approximately 100mg DW of ground tissue was extracted threetimes in boiling 80% ethanol, centrifuged and thesupernatant pooled. The pellet was digested at40°C for 2 h with amyloglucosidase from Rhizo-pus (Sigma, St. Louis, MO) and filtered. Solublesugars and glucose released from starch (totalnon-structural carbohydrate, TNC) werequantified spectrophotometrically following thereaction with anthrone. Nitrogen (N) concentra-tion was determined on 50 mg DW samples by themicro-Kjeldahl method (Black, 1968).
At the end of gas exchange measurements onJuly 7 several small branches of the same stage ofdevelopment (about 30 cm in length) were cutfrom the sample trees, placed in plastic bags, andtransported on ice to the laboratory where theywere stored at 4°C until processing the followingday. At least seven equal-aged leaves per tree wereprocessed for tannins. The next day, major veinswere removed with a razor blade and discarded.The leaves were cut into small pieces; about 1-gsamples FW were immediately ground in 5 ml of70% acetone with a cold tissue homogenizer thencentrifuged at 5000 rpm for 15 min. The superna-tants were stored at 4°C until analysis. Tanninconcentration in the leaf extracts was determinedwith a modified radial diffusion method of Hager-man (1987); a gel containing 1% (w/v) agaroseand 0.05% (w/v) bovine serum albumin (BSA) in50 mM acetic acid and 60 mM ascorbic acidadjusted to pH 5.0 (boiled and stirred, the BSAwas added after cooling the agar to 45°C) wasformed by dispensing about 9.5 ml into standardplastic Petri dishes. Eight 5-mm wells per platewere punched in the gel and two 8-ml aliquots ofeach leaf extract were applied to duplicate wells.The Petri dishes were covered and sealed with
Parafilm and incubated at 30°C for 120 h. Twodiameters at right angles of each diffusion ringwere measured and the areas of the duplicatewells were averaged. Tannin concentration wasdetermined using a standard curve created withpurified tannic acid (Sigma, St. Louis, MO). Theremaining leaves and branches were oven-dried at60°C for 48 h. Leaves and stems were separated,and soluble sugars and glucose released fromstarch as well as nitrogen concentration werequantified by using the methods described above.
The sampling for TNC and N was repeated onSeptember 19, 1995, on both species, when plantsstarted recovering from drought, with the sameprocedure used in May.
2.4. Statistical analysis
The data were subjected to statistical analysisof variance, using the general linear model (GLM)and linear regression (PROCLIN) provided bySAS statistical software. Probability values of lessthan 5% (Duncan’s multiple range test) were con-sidered significant. Analysis of at least duplicatesamples of N and TNC (sugars+starch) in theexperiment carried out in July provided an esti-mate of analytical error (Norby et al., 1986), butbecause leaves and branches were pooled for eachspecies at each site, there is no estimate of plant-to-plant variation, and statistical analysis of thedata is not possible.
3. Results
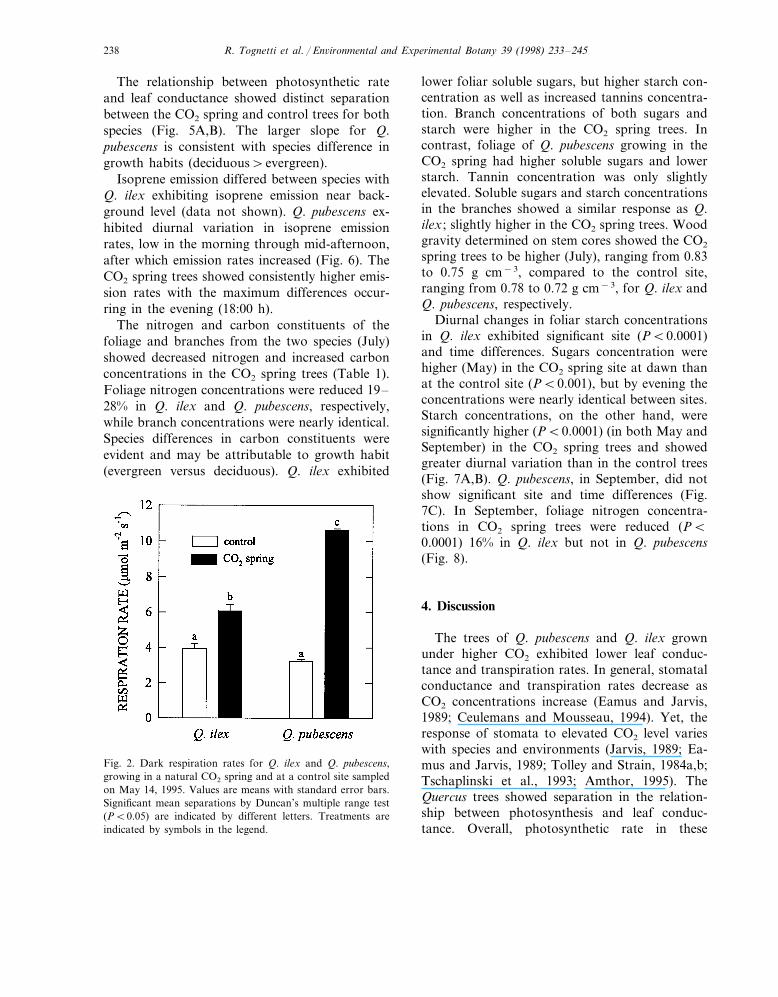
In May Q. pubescens trees did not show anydifference between control and CO2 spring site inphotosynthetic rate measured at ambient CO2 lev-els, whereas Q. ilex trees grown in the CO2 springexhibited nearly double photosynthetic rate values(Fig. 1A). Photosynthetic rate responded posi-tively to supplemental CO2, at least doubling therates for both species and sites. This treatmentexacerbated species differences (Q. ilex alwaysshowing the highest values) but equalized photo-synthetic rate between sites (Fig. 1B). Dark respi-ration was much higher at the CO2 spring site inboth species, about 50% in Q. ilex and 300%
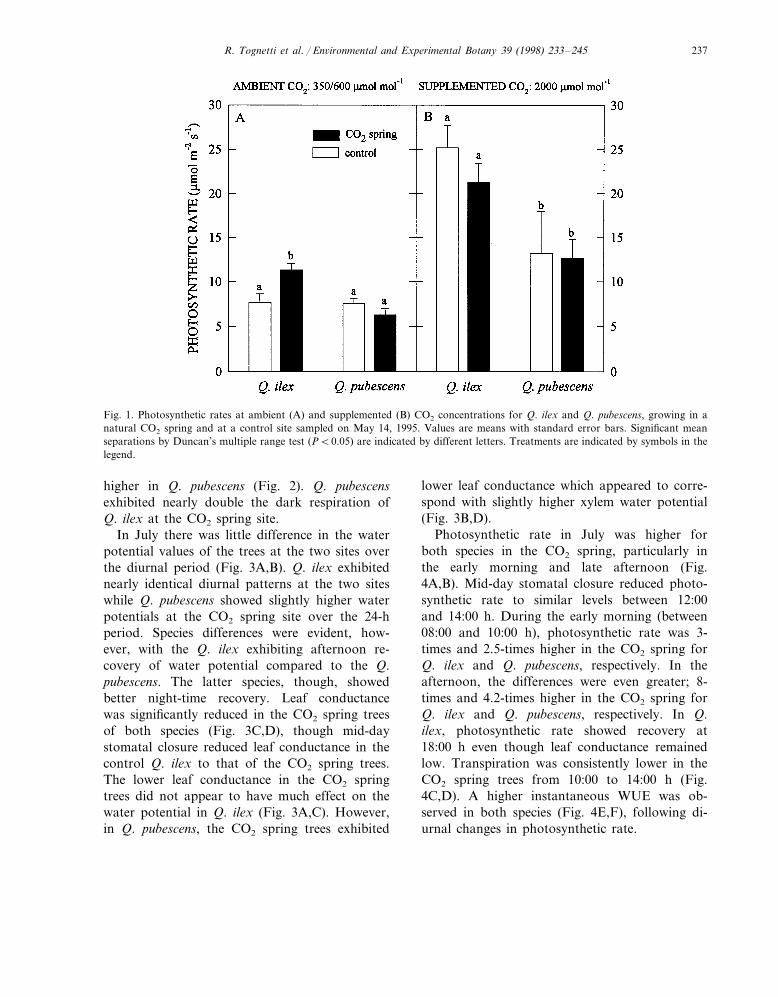
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 237
Fig. 1. Photosynthetic rates at ambient (A) and supplemented (B) CO2 concentrations for Q. ilex and Q. pubescens, growing in anatural CO2 spring and at a control site sampled on May 14, 1995. Values are means with standard error bars. Significant meanseparations by Duncan’s multiple range test (PB0.05) are indicated by different letters. Treatments are indicated by symbols in thelegend.
higher in Q. pubescens (Fig. 2). Q. pubescensexhibited nearly double the dark respiration ofQ. ilex at the CO2 spring site.
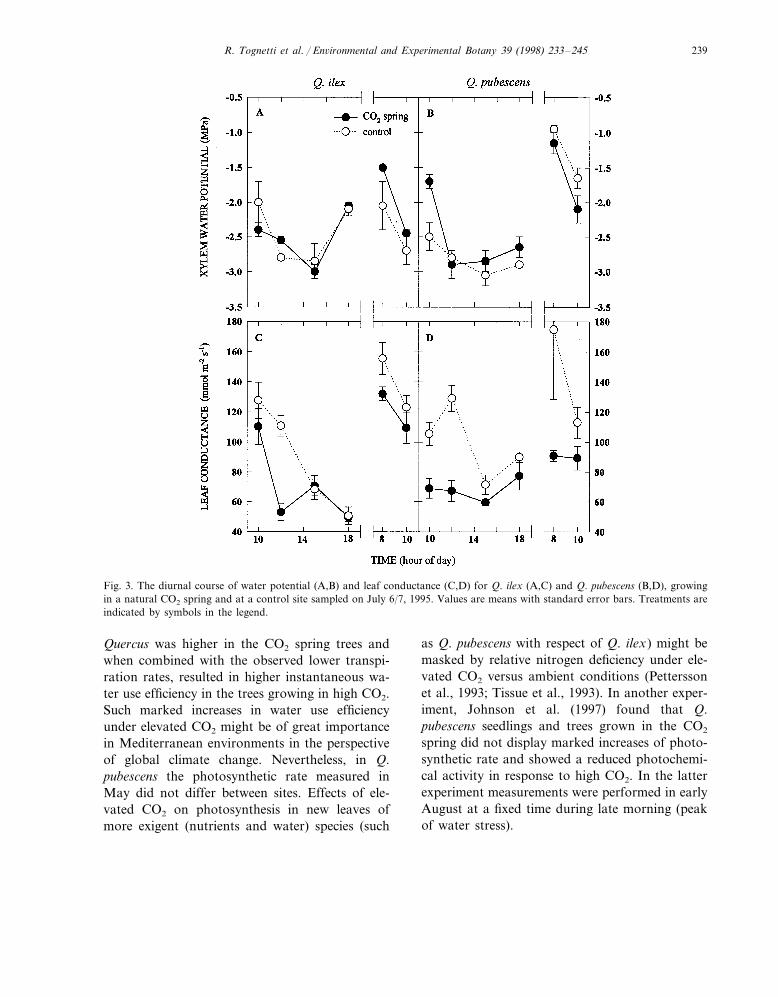
In July there was little difference in the waterpotential values of the trees at the two sites overthe diurnal period (Fig. 3A,B). Q. ilex exhibitednearly identical diurnal patterns at the two siteswhile Q. pubescens showed slightly higher waterpotentials at the CO2 spring site over the 24-hperiod. Species differences were evident, how-ever, with the Q. ilex exhibiting afternoon re-covery of water potential compared to the Q.pubescens. The latter species, though, showedbetter night-time recovery. Leaf conductancewas significantly reduced in the CO2 spring treesof both species (Fig. 3C,D), though mid-daystomatal closure reduced leaf conductance in thecontrol Q. ilex to that of the CO2 spring trees.The lower leaf conductance in the CO2 springtrees did not appear to have much effect on thewater potential in Q. ilex (Fig. 3A,C). However,in Q. pubescens, the CO2 spring trees exhibited
lower leaf conductance which appeared to corre-spond with slightly higher xylem water potential(Fig. 3B,D).
Photosynthetic rate in July was higher forboth species in the CO2 spring, particularly inthe early morning and late afternoon (Fig.4A,B). Mid-day stomatal closure reduced photo-synthetic rate to similar levels between 12:00and 14:00 h. During the early morning (between08:00 and 10:00 h), photosynthetic rate was 3-times and 2.5-times higher in the CO2 spring forQ. ilex and Q. pubescens, respectively. In theafternoon, the differences were even greater; 8-times and 4.2-times higher in the CO2 spring forQ. ilex and Q. pubescens, respectively. In Q.ilex, photosynthetic rate showed recovery at18:00 h even though leaf conductance remainedlow. Transpiration was consistently lower in theCO2 spring trees from 10:00 to 14:00 h (Fig.4C,D). A higher instantaneous WUE was ob-served in both species (Fig. 4E,F), following di-urnal changes in photosynthetic rate.
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245238
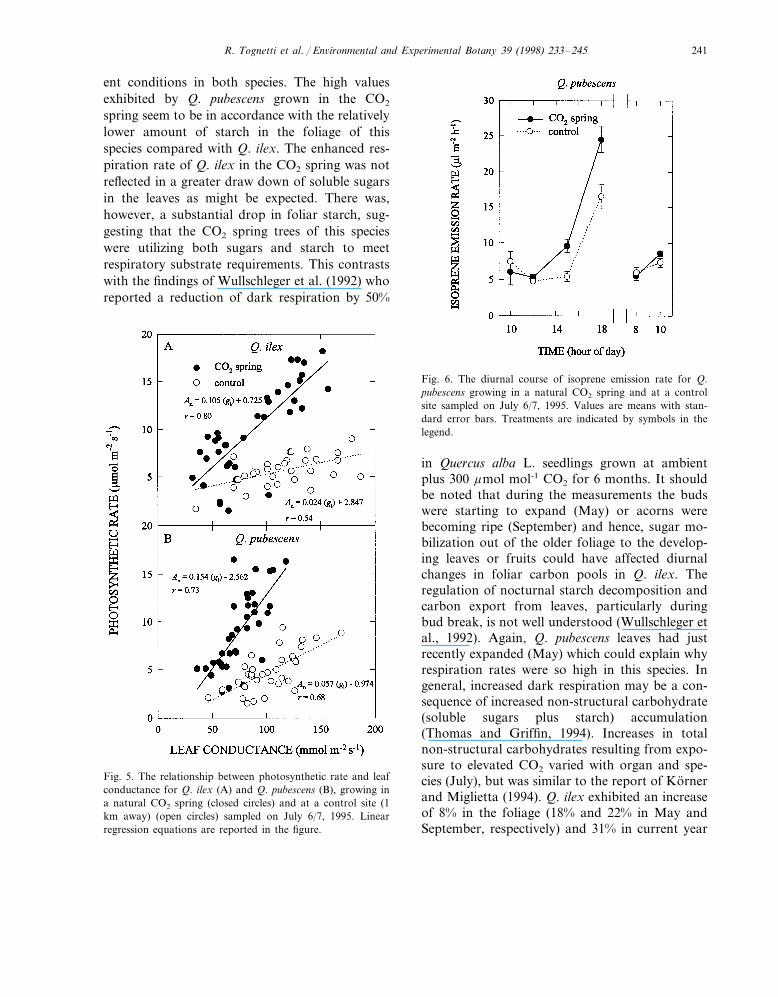
The relationship between photosynthetic rateand leaf conductance showed distinct separationbetween the CO2 spring and control trees for bothspecies (Fig. 5A,B). The larger slope for Q.pubescens is consistent with species difference ingrowth habits (deciduous\evergreen).
Isoprene emission differed between species withQ. ilex exhibiting isoprene emission near back-ground level (data not shown). Q. pubescens ex-hibited diurnal variation in isoprene emissionrates, low in the morning through mid-afternoon,after which emission rates increased (Fig. 6). TheCO2 spring trees showed consistently higher emis-sion rates with the maximum differences occur-ring in the evening (18:00 h).
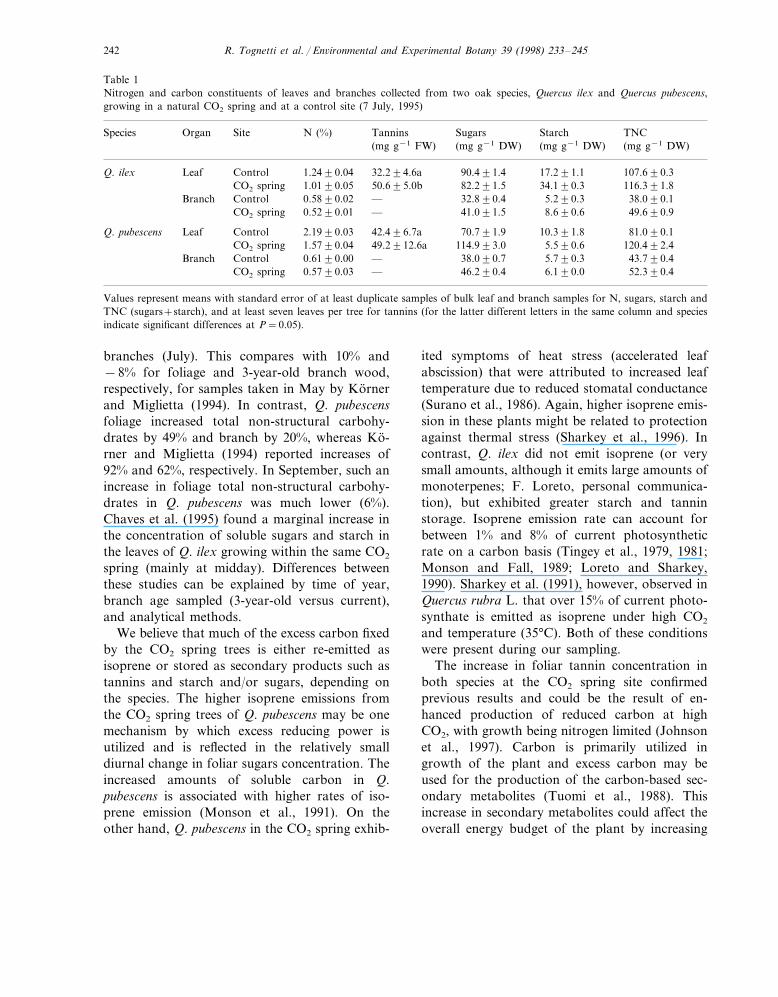
The nitrogen and carbon constituents of thefoliage and branches from the two species (July)showed decreased nitrogen and increased carbonconcentrations in the CO2 spring trees (Table 1).Foliage nitrogen concentrations were reduced 19–28% in Q. ilex and Q. pubescens, respectively,while branch concentrations were nearly identical.Species differences in carbon constituents wereevident and may be attributable to growth habit(evergreen versus deciduous). Q. ilex exhibited
lower foliar soluble sugars, but higher starch con-centration as well as increased tannins concentra-tion. Branch concentrations of both sugars andstarch were higher in the CO2 spring trees. Incontrast, foliage of Q. pubescens growing in theCO2 spring had higher soluble sugars and lowerstarch. Tannin concentration was only slightlyelevated. Soluble sugars and starch concentrationsin the branches showed a similar response as Q.ilex ; slightly higher in the CO2 spring trees. Woodgravity determined on stem cores showed the CO2
spring trees to be higher (July), ranging from 0.83to 0.75 g cm−3, compared to the control site,ranging from 0.78 to 0.72 g cm−3, for Q. ilex andQ. pubescens, respectively.
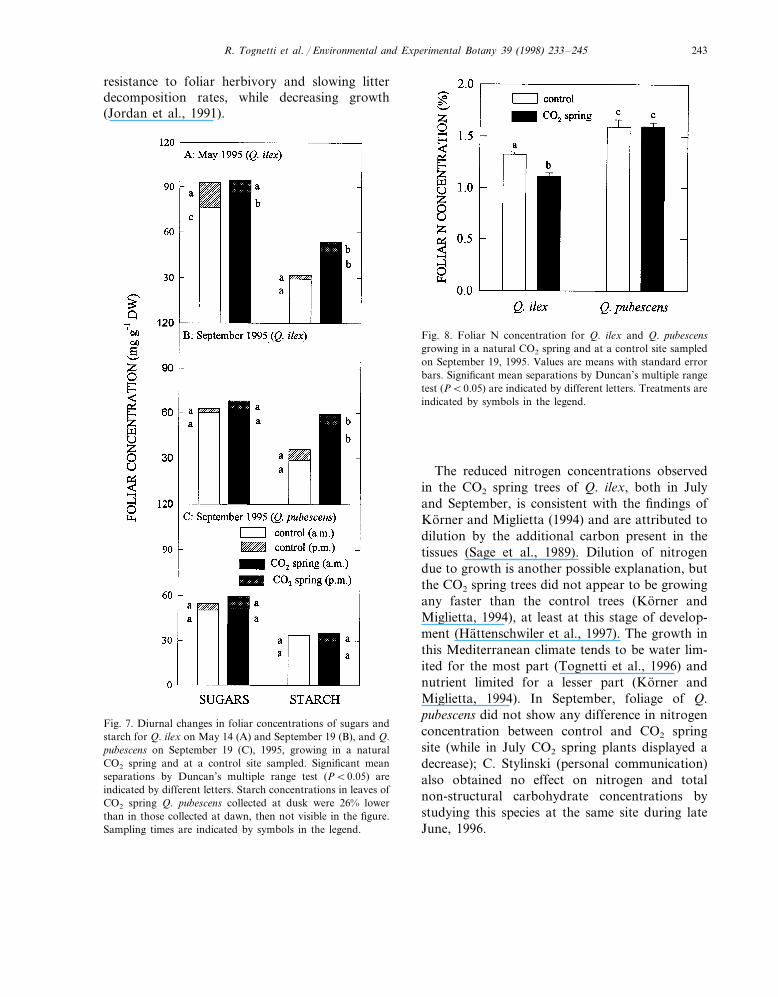
Diurnal changes in foliar starch concentrationsin Q. ilex exhibited significant site (PB0.0001)and time differences. Sugars concentration werehigher (May) in the CO2 spring site at dawn thanat the control site (PB0.001), but by evening theconcentrations were nearly identical between sites.Starch concentrations, on the other hand, weresignificantly higher (PB0.0001) (in both May andSeptember) in the CO2 spring trees and showedgreater diurnal variation than in the control trees(Fig. 7A,B). Q. pubescens, in September, did notshow significant site and time differences (Fig.7C). In September, foliage nitrogen concentra-tions in CO2 spring trees were reduced (PB0.0001) 16% in Q. ilex but not in Q. pubescens(Fig. 8).
4. Discussion
The trees of Q. pubescens and Q. ilex grownunder higher CO2 exhibited lower leaf conduc-tance and transpiration rates. In general, stomatalconductance and transpiration rates decrease asCO2 concentrations increase (Eamus and Jarvis,1989; Ceulemans and Mousseau, 1994). Yet, theresponse of stomata to elevated CO2 level varieswith species and environments (Jarvis, 1989; Ea-mus and Jarvis, 1989; Tolley and Strain, 1984a,b;Tschaplinski et al., 1993; Amthor, 1995). TheQuercus trees showed separation in the relation-ship between photosynthesis and leaf conduc-tance. Overall, photosynthetic rate in these
Fig. 2. Dark respiration rates for Q. ilex and Q. pubescens,growing in a natural CO2 spring and at a control site sampledon May 14, 1995. Values are means with standard error bars.Significant mean separations by Duncan’s multiple range test(PB0.05) are indicated by different letters. Treatments areindicated by symbols in the legend.
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 239
Fig. 3. The diurnal course of water potential (A,B) and leaf conductance (C,D) for Q. ilex (A,C) and Q. pubescens (B,D), growingin a natural CO2 spring and at a control site sampled on July 6/7, 1995. Values are means with standard error bars. Treatments areindicated by symbols in the legend.
Quercus was higher in the CO2 spring trees andwhen combined with the observed lower transpi-ration rates, resulted in higher instantaneous wa-ter use efficiency in the trees growing in high CO2.Such marked increases in water use efficiencyunder elevated CO2 might be of great importancein Mediterranean environments in the perspectiveof global climate change. Nevertheless, in Q.pubescens the photosynthetic rate measured inMay did not differ between sites. Effects of ele-vated CO2 on photosynthesis in new leaves ofmore exigent (nutrients and water) species (such
as Q. pubescens with respect of Q. ilex) might bemasked by relative nitrogen deficiency under ele-vated CO2 versus ambient conditions (Petterssonet al., 1993; Tissue et al., 1993). In another exper-iment, Johnson et al. (1997) found that Q.pubescens seedlings and trees grown in the CO2
spring did not display marked increases of photo-synthetic rate and showed a reduced photochemi-cal activity in response to high CO2. In the latterexperiment measurements were performed in earlyAugust at a fixed time during late morning (peakof water stress).

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245240
Fig. 4. The diurnal course of photosynthetic rate (A,B), transpiration rate (C,D) and instantaneous water use efficiency (WUE) (E,F)for Q. ilex (A,C) and Q. pubescens (B,D), growing in a natural CO2 spring and at a control site sampled on July 6/7, 1995. Valuesare means with standard error bars. Treatments are indicated by symbols in the legend.
The lack of difference in photosynthetic ratebetween CO2 spring and control trees in bothspecies when measured at the same CO2 concen-trations supports the hypothesis of the absence ofdownward photosynthetic acclimation in theseQuercus (Liu and Teskey, 1995). Significant de-pression in photosynthetic rate was not observedin either Quercus species after long-term CO2
exposure, even though Q. ilex showed increasedfoliar starch concentration and Q. pubescens
showed increased foliar sugars concentration(July). It appears that these Quercus do not down-regulate photosynthesis, and thus tend to accumu-late increased assimilate pools. Korner andMiglietta (1994) came to the same conclusionbased on biometric, phenological and chemicalobservations of the same tree species growing inthe Bossoleto CO2 spring.
The rate of dark respiration was higher atelevated CO2 concentrations compared with ambi-
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 241
ent conditions in both species. The high valuesexhibited by Q. pubescens grown in the CO2
spring seem to be in accordance with the relativelylower amount of starch in the foliage of thisspecies compared with Q. ilex. The enhanced res-piration rate of Q. ilex in the CO2 spring was notreflected in a greater draw down of soluble sugarsin the leaves as might be expected. There was,however, a substantial drop in foliar starch, sug-gesting that the CO2 spring trees of this specieswere utilizing both sugars and starch to meetrespiratory substrate requirements. This contrastswith the findings of Wullschleger et al. (1992) whoreported a reduction of dark respiration by 50%
Fig. 6. The diurnal course of isoprene emission rate for Q.pubescens growing in a natural CO2 spring and at a controlsite sampled on July 6/7, 1995. Values are means with stan-dard error bars. Treatments are indicated by symbols in thelegend.
Fig. 5. The relationship between photosynthetic rate and leafconductance for Q. ilex (A) and Q. pubescens (B), growing ina natural CO2 spring (closed circles) and at a control site (1km away) (open circles) sampled on July 6/7, 1995. Linearregression equations are reported in the figure.
in Quercus alba L. seedlings grown at ambientplus 300 mmol mol-1 CO2 for 6 months. It shouldbe noted that during the measurements the budswere starting to expand (May) or acorns werebecoming ripe (September) and hence, sugar mo-bilization out of the older foliage to the develop-ing leaves or fruits could have affected diurnalchanges in foliar carbon pools in Q. ilex. Theregulation of nocturnal starch decomposition andcarbon export from leaves, particularly duringbud break, is not well understood (Wullschleger etal., 1992). Again, Q. pubescens leaves had justrecently expanded (May) which could explain whyrespiration rates were so high in this species. Ingeneral, increased dark respiration may be a con-sequence of increased non-structural carbohydrate(soluble sugars plus starch) accumulation(Thomas and Griffin, 1994). Increases in totalnon-structural carbohydrates resulting from expo-sure to elevated CO2 varied with organ and spe-cies (July), but was similar to the report of Kornerand Miglietta (1994). Q. ilex exhibited an increaseof 8% in the foliage (18% and 22% in May andSeptember, respectively) and 31% in current year
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245242
Table 1Nitrogen and carbon constituents of leaves and branches collected from two oak species, Quercus ilex and Quercus pubescens,growing in a natural CO2 spring and at a control site (7 July, 1995)
Site N (%) TanninsOrganSpecies Sugars Starch TNC(mg g−1 DW)(mg g−1 DW) (mg g−1 DW)(mg g−1 FW)
Control 1.2490.04 32.294.6aQ. ilex 90.491.4Leaf 17.291.1 107.690.3CO2 spring 1.0190.05 50.695.0b 82.291.5 34.190.3 116.391.8Control 0.5890.02 —Branch 32.890.4 5.290.3 38.090.1CO2 spring 0.5290.01 — 41.091.5 8.690.6 49.690.9
Q. pubescens Leaf Control 2.1990.03 42.496.7a 70.791.9 10.391.8 81.090.1CO2 spring 1.5790.04 49.2912.6a 114.993.0 5.590.6 120.492.4Control 0.6190.00Branch — 38.090.7 5.790.3 43.790.4CO2 spring 0.5790.03 — 46.290.4 6.190.0 52.390.4
Values represent means with standard error of at least duplicate samples of bulk leaf and branch samples for N, sugars, starch andTNC (sugars+starch), and at least seven leaves per tree for tannins (for the latter different letters in the same column and speciesindicate significant differences at P=0.05).
branches (July). This compares with 10% and−8% for foliage and 3-year-old branch wood,respectively, for samples taken in May by Kornerand Miglietta (1994). In contrast, Q. pubescensfoliage increased total non-structural carbohy-drates by 49% and branch by 20%, whereas Ko-rner and Miglietta (1994) reported increases of92% and 62%, respectively. In September, such anincrease in foliage total non-structural carbohy-drates in Q. pubescens was much lower (6%).Chaves et al. (1995) found a marginal increase inthe concentration of soluble sugars and starch inthe leaves of Q. ilex growing within the same CO2
spring (mainly at midday). Differences betweenthese studies can be explained by time of year,branch age sampled (3-year-old versus current),and analytical methods.
We believe that much of the excess carbon fixedby the CO2 spring trees is either re-emitted asisoprene or stored as secondary products such astannins and starch and/or sugars, depending onthe species. The higher isoprene emissions fromthe CO2 spring trees of Q. pubescens may be onemechanism by which excess reducing power isutilized and is reflected in the relatively smalldiurnal change in foliar sugars concentration. Theincreased amounts of soluble carbon in Q.pubescens is associated with higher rates of iso-prene emission (Monson et al., 1991). On theother hand, Q. pubescens in the CO2 spring exhib-
ited symptoms of heat stress (accelerated leafabscission) that were attributed to increased leaftemperature due to reduced stomatal conductance(Surano et al., 1986). Again, higher isoprene emis-sion in these plants might be related to protectionagainst thermal stress (Sharkey et al., 1996). Incontrast, Q. ilex did not emit isoprene (or verysmall amounts, although it emits large amounts ofmonoterpenes; F. Loreto, personal communica-tion), but exhibited greater starch and tanninstorage. Isoprene emission rate can account forbetween 1% and 8% of current photosyntheticrate on a carbon basis (Tingey et al., 1979, 1981;Monson and Fall, 1989; Loreto and Sharkey,1990). Sharkey et al. (1991), however, observed inQuercus rubra L. that over 15% of current photo-synthate is emitted as isoprene under high CO2
and temperature (35°C). Both of these conditionswere present during our sampling.
The increase in foliar tannin concentration inboth species at the CO2 spring site confirmedprevious results and could be the result of en-hanced production of reduced carbon at highCO2, with growth being nitrogen limited (Johnsonet al., 1997). Carbon is primarily utilized ingrowth of the plant and excess carbon may beused for the production of the carbon-based sec-ondary metabolites (Tuomi et al., 1988). Thisincrease in secondary metabolites could affect theoverall energy budget of the plant by increasing
R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 243
resistance to foliar herbivory and slowing litterdecomposition rates, while decreasing growth(Jordan et al., 1991).
Fig. 8. Foliar N concentration for Q. ilex and Q. pubescensgrowing in a natural CO2 spring and at a control site sampledon September 19, 1995. Values are means with standard errorbars. Significant mean separations by Duncan’s multiple rangetest (PB0.05) are indicated by different letters. Treatments areindicated by symbols in the legend.
Fig. 7. Diurnal changes in foliar concentrations of sugars andstarch for Q. ilex on May 14 (A) and September 19 (B), and Q.pubescens on September 19 (C), 1995, growing in a naturalCO2 spring and at a control site sampled. Significant meanseparations by Duncan’s multiple range test (PB0.05) areindicated by different letters. Starch concentrations in leaves ofCO2 spring Q. pubescens collected at dusk were 26% lowerthan in those collected at dawn, then not visible in the figure.Sampling times are indicated by symbols in the legend.
The reduced nitrogen concentrations observedin the CO2 spring trees of Q. ilex, both in Julyand September, is consistent with the findings ofKorner and Miglietta (1994) and are attributed todilution by the additional carbon present in thetissues (Sage et al., 1989). Dilution of nitrogendue to growth is another possible explanation, butthe CO2 spring trees did not appear to be growingany faster than the control trees (Korner andMiglietta, 1994), at least at this stage of develop-ment (Hattenschwiler et al., 1997). The growth inthis Mediterranean climate tends to be water lim-ited for the most part (Tognetti et al., 1996) andnutrient limited for a lesser part (Korner andMiglietta, 1994). In September, foliage of Q.pubescens did not show any difference in nitrogenconcentration between control and CO2 springsite (while in July CO2 spring plants displayed adecrease); C. Stylinski (personal communication)also obtained no effect on nitrogen and totalnon-structural carbohydrate concentrations bystudying this species at the same site during lateJune, 1996.

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245244
The diurnal gas exchange pattern suggests thatthe CO2 spring trees take advantage of timeswhen their water balance is more favorable, in theearly morning and late afternoon. This appears tobe a function of stomatal conductance in the earlymorning in both Quercus species and in the after-noon in Q. pubescens. The increase in photosyn-thetic rate in the late afternoon in Q. ilex,however, appears to be due to non-stomatal fac-tors since leaf conductance further decreasedwhile photosynthesis increased. Similar resultswere reported by Tenhunen et al. (1984) for Quer-cus suber L. which increased leaf photosyntheticcapacity and carboxylation efficiency in the after-noon following the recovery of water potential.Diurnal changes in water potential also showedspecies differences. Q. ilex showed afternoon re-covery in water potential as leaf conductancedecreased. Q. pubescens, in contrast, showed littlerecovery of water potential in the afternoon, butdid exhibit recovery during the night. Such speciesdifferences could be related to different roots dis-tribution of the two Quercus species and the abil-ity of Q. ilex to tolerate low water potentials(Tognetti et al., 1996).
In conclusion, Q. pubescens and Q. ilex exposedto elevated CO2 for prolonged periods exhibitdifferent mechanisms for dealing with additionalreduced carbon and do maintain an altered car-bon physiology, even in the midst of the region’scharacteristic summer drought. There is no evi-dence of down-regulation of either photosynthesisor respiration despite the long-term exposure tovery high CO2 concentrations. Diurnal changes inlabile carbon in leaves appear to be affected byelevated CO2. Further research is warranted todetermine the physiological mechanisms that al-low these Mediterranean Quercus to adapt andgrow in high CO2 environments, particularly inlight of their different growth habits.
Acknowledgements
This research was supported by the NationalResearch Council (CNR) of Italy and by E.U.programme ENVIRONMENT, contract EV5V-CT93-0093.
References
Amthor, J.S., 1995. Terrestrial higher-plant response to in-creasing atmospheric (CO2) in relation to global carboncycle. Global Change Biol. 1, 243–274.
Black, C.A., 1968. Methods of Soil Analysis. Part 2: Chemicaland Microbial Properties. Madison: American Society ofAgronomy and American Society for Testing andMaterials.
Ceulemans, R., Mousseau, M., 1994. Effects of elevated atmo-spheric CO2 on woody plants. New Phytol. 127, 425–446.
Chaves, M.M., Pereira, J.S., Cerasoli, S., Clifton-Brown, J.,Miglietta, F., Raschi, A., 1995. Leaf metabolism duringsummer drought in Quercus ilex trees with lifetime expo-sure to elevated CO2. J. Biogeogr. 22, 255–259.
Eamus, D., Jarvis, P.G., 1989. The direct effects of increase inthe global atmospheric CO2 concentration on natural andcommercial temperate trees and forests. Adv. Ecol. Res.19, 1–55.
Hagerman, A.E., 1987. Radial diffusion method for determin-ing tannin in plant extracts. J. Chem. Ecol. 13, 437–449.
Hattenschwiler, S., Miglietta, F., Raschi, A., Korner, C., 1997.Thirty years of in situ tree growth under elevated CO2: amodel for future forest responses. Global Change Biol. 3,463–471.
Houpis, J.L.J., Surano, K.A., Cowles, S., Shinn, J.H., 1988.Chlorophyll and carotenoid concentrations in two varietiesof Pinus ponderosa seedlings subjected to long-term ele-vated carbon dioxide. Tree Physiol. 4, 187–193.
Jarvis, P.G., 1989. Atmospheric carbon dioxide and forests.Phil. Trans. R. Soc. London B324, 369–392.
Johnson, J.D., Michelozzi, M., Tognetti, R., 1997. Carbonphysiology of Quercus pubescens growing in the BossoletoCO2 spring of central Italy. In: Raschi, A., Miglietta, F.,Tognetti, R., van Gardingen, P.R. (Eds.), Plant Responsesto Elevated Carbon Dioxide: Evidence from NaturalSprings. Cambridge University Press, Cambridge, pp. 148–164.
Jones, M.B., Clifton Brown, J., Raschi, A., Miglietta, F., 1995.The effects on Arbutus unedo L. of long-term exposure toelevated CO2. Global Change Biol. 1, 295–302.
Jordan, D.N., Green, T.H., Chappelka, A.H., Lockaby, B.G.,Meldahl, R.S., Gjerstad, D.H., 1991. Response of totaltannins and phenolics in loblolly pine foliage exposed toozone and acid rain. J. Chem. Ecol. 17, 505–513.
Korner, C., Miglietta, F., 1994. Long-term effects of naturallyelevated CO2 on Mediterranean grassland and forest trees.Oecologia 99, 343–351.
Kuzma, J., Fall, R., 1993. Leaf isoprene emission rate isdependent on leaf development and the level of isoprenesynthase. Plant Physiol. 101, 435–440.
Lawler, I.R., Foley, W.J., Woodrow, I.E., Cork, S.J., 1997.The effects of elevated CO2 atmospheres on the nutritionalquality of Eucalyptus foliage and its interaction with soilnutrient and light availability. Oecologia 109, 59–68.
Liu, S., Teskey, R.O., 1995. Responses of foliar gas exchangeto long-term elevated CO2 concentrations in matureloblolly pine trees. Tree Physiol. 15, 351–359.

R. Tognetti et al. / En6ironmental and Experimental Botany 39 (1998) 233–245 245
Loreto, F., Sharkey, T.D., 1990. A gas-exchange study ofphotosynthesis and isoprene emission in Quercus rubra L.Planta 182, 523–531.
Miglietta, F., Raschi, A., Bettarini, I., Resti, R., Selvi, F., 1993.Natural CO2 springs in Italy: a resource for examininglong-term response of vegetation to rising atmospheric CO2
concentrations. Plant Cell Environ. 16, 873–878.Monson, R.K., Fall, R., 1989. Isoprene emission from aspen
leaves. The influence of environment and relation to photo-synthesis and photorespiration. Plant Physiol. 90, 267–274.
Monson, R.K., Guenther, A.B., Fall, R., 1991. Physiologicalreality in relation to ecosystem and global level estimates ofisoprene emission. In: Sharkey, T.D., Holland, E.A.,Mooney, H.A. (Eds.), Trace Gas Emission by Plants.Academic Press, San Diego, pp. 185–205.
Norby, R.J., Pastor, J., Melillo, J.M., 1986. Carbon-nitrogeninteractions in CO2-enriched white oak: physiological andlong-term perspectives. Tree Physiol. 2, 233–241.
Pettersson, R., Mc Donald, A.J.S., Stadenberg, I., 1993. Re-sponse of small birch plants (Betula pendula Roth.) toelevated CO2 and nitrogen supply. Plant Cell Environ. 16,1115–1121.
Sage, R.F., Sharkey, T.D., Seeman, J.R., 1989. Acclimation ofphotosynthesis to elevated CO2 in five C3 species. PlantPhysiol. 89, 590–596.
Sharkey, T.D., Loreto, F., Delwiche, C.F., 1991. High carbondioxide and sun/shade effects on isoprene emission fromoak and aspen tree leaves. Plant Cell Environ. 14, 333–338.
Sharkey, T.D., Singaas, E.R., Vanderveer, P.J., Geron, C.,1996. Field measurements of isoprene emission from treesin response to temperature and light. Tree Physiol. 16,649–654.
Surano, K.A., Daley, P.F., Houpis, J.L.J., Shinn, J.H., Helms,J.A., Palassou, R.J., Costella, M.P., 1986. Growth andphysiological responses of Pinus ponderosa Doug. ex P.Laws. to long-term elevated CO2 concentrations. TreePhysiol. 2, 243–259.
Tenhunen, J.D., Lange, O.L., Gebel, J., Beyschlag, W., Weber,J.A., 1984. Changes in photosynthetic capacity, carboxyla-tion efficiency, and CO2 compensation point associatedwith midday stomatal closure and midday depression of netCO2 exchange of leaves of Quercus suber. Planta 162,193–203.
Thomas, R.B., Griffin, K.L., 1994. Direct and indirect effectsof atmospheric carbon dioxide enrichment on leaf respira-tion of Glycine max (L.) Merr. Plant Physiol. 104, 355–361.
Tingey, D.T., Manning, M., Grothaus, L.C., Burns, W.F.,1979. The influence of light and temperature in isopreneemission rates from live oaks. Physiol. Plant 47, 112–118.
Tingey, D.T., Evans, R., Gumpertez, M., 1981. Effects ofenvironmental conditions on isoprene emission from liveoak. Planta 152, 565–570.
Tissue, D.T., Thomas, R.B., Strain, B.R., 1993. Long-termeffects of elevated CO2 and nutrients on photosynthesis andrubisco in loblolly pine seedlings. Plant Cell Environ. 16,859–865.
Tognetti, R., Giovannelli, A., Longobucco, A., Miglietta, F.,Raschi, A., 1996. Water relations of oak species growing inthe natural CO2 spring of Rapolano (central Italy). Ann.Sci. For. 53, 475–485.
Tolley, L.C., Strain, B.R., 1984a. Effects of CO2 enrichment ongrowth of Liquidambar styraciflua and Pinus taeda seedlingsunder different irradiance levels. Can. J. For. Res. 14,343–350.
Tolley, L.C., Strain, B.R., 1984b. Effects of CO2 enrichmentand water stress on growth of Liquidambar styraciflua andPinus taeda seedlings. Can. J. Bot. 62, 2135–2139.
Tschaplinski, T.J., Norby, R.J., Wullschleger, S.D., 1993. Re-sponses of loblolly pine seedlings to elevated CO2 andfluctuating water supply. Tree Physiol. 13, 283–296.
Tuomi, J., Niemela, P., Chapin III, F.S., Bryant, J.P., Siren, S.,1988. Defensive responses of trees in relation to theircarbon/nutrient balance. In: Mattson, W.J., Levieux, J.,Bernard-Dagan, C. (Eds.), Mechanisms of Woody PlantDefenses Against Insects: Search for Pattern. Springer-Ver-lag, New York, pp. 52–72.
van Gardingen, P.R., Grace, J., Harkness, D.H., Miglietta, F.,Raschi, A., 1995. Carbon dioxide emissions at an Italianmineral spring: measurements of average CO2 concentra-tion and air temperature. Agr. For. Meteorol. 73, 17–27.
van Gardingen, P.R., Grace, J., Jeffree, C.E., Byari, S.H.,Raschi, A., Bettarini, I., 1997. Long-term effects of en-hanced CO2 concentrations on gas exchange: research op-portunities using CO2 springs. In: Raschi, A., Miglietta, F.,Tognetti, R., van Gardingen, P.R. (Eds.), Plant Responsesto Elevated Carbon Dioxide: Evidence from NaturalSprings. Cambridge University Press, Cambridge, pp. 69–86.
Wullschleger, S.D., Norby, R.J., Hendrix, D.L., 1992. Carbonexchange rates, chlorophyll content, and carbohydratestatus of two forest tree species exposed to carbon dioxideenrichment. Tree Physiol. 10, 21–31.
.





![Synthesis and Characterization of LiFePO[sub 4] and LiTi[sub 0.01]Fe[sub 0.99]PO[sub 4] Cathode Materials](https://static.fdokumen.com/doc/165x107/631dae063dc6529d5d079742/synthesis-and-characterization-of-lifeposub-4-and-litisub-001fesub-099posub.jpg)

![Structural and thermoelectric properties of Bi[sub 2]Sr[sub 2]Co[sub 2]O[sub y] thin films on LaAlO[sub 3] (100) and fused silica substrates](https://static.fdokumen.com/doc/165x107/634467196cfb3d406409325f/structural-and-thermoelectric-properties-of-bisub-2srsub-2cosub-2osub-y.jpg)





![Small anisotropy, weak thermal fluctuations, and high field superconductivity in Co-doped iron pnictide Ba(Fe[sub 1−x]Co[sub x])[sub 2]As[sub 2]](https://static.fdokumen.com/doc/165x107/633cc1de7000aa173d0630af/small-anisotropy-weak-thermal-fluctuations-and-high-field-superconductivity-in.jpg)






