
Up-conversion luminescence in Ho< sup> 3+ and Tm< sup> 3+ co-doped Y< sub> 2 O< sub> 3: Yb< sup> 3+
Upload
independentCategory
view
1download
0
Microbes and Infection 8 (2006) 2130e2137www.elsevier.com/locate/micinf
Original article
Immunogenicity of a recombinant protein containing thePlasmodium vivax vaccine candidate MSP119 and two
human CD4þ T-cell epitopes administered tonon-human primates (Callithrix jacchus jacchus)
Daniela S. Rosa a, Leo K. Iwai b, Fanny Tzelepis a, Daniel Y. Bargieri a,Magda A. Medeiros c, Irene S. Soares d, John Sidney e, Alessandro Sette e, Jorge Kalil b,
Luiz Eugenio Mello f, Edecio Cunha-Neto b, Mauricio M. Rodrigues a,*
a CINTERGEN, Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de S~ao Paulo-Escola Paulista de Medicina,
Rua Botucatu 862, 6th floor, S~ao Paulo, SP 04023-062, Brazilb Laboratorio de Imunologia, Instituto do corac~ao (Incor), Faculdade de Medicina da Universidade de S~ao Paulo, Av. Dr. Eneas de Carvalho Aguiar,
44-Bloco II-9th floor, S~ao Paulo, SP 05403-000, Brazilc Departamento de Ciencias Fisiologicas, Universidade Federal Rural do Rio de Janeiro, BR 464, km 7, Seropedica, RJ 23890-000, Brazil
d Departamento de Analises Clınicas e Toxicologicas, Faculdade de Ciencias Farmaceuticas, Universidade de S~ao Paulo, Av. Prof. Lineu Prestes,
580, Bloco 17, S~ao Paulo, SP 05508-900, Brazile La Jolla Institute for Allergy and Immunology, San Diego, CA 92121, USA
f Departamento de Fisiologia, Universidade Federal de S~ao Paulo-Escola Paulista de Medicina, Rua Botucatu 862, 5th floor,
S~ao Paulo, SP 04023-062, Brazil
Received 10 January 2006; accepted 30 March 2006
Available online 2 June 2006
Abstract
One of the most promising vaccine candidates against the erythrocytic forms of malaria is the 19 kDa C-terminal region of the merozoitesurface protein 1 (MSP119). As part of our studies aimed at the development of a Plasmodium vivax malaria vaccine, we characterized the im-munogenic properties of a new bacterial recombinant protein containing the P. vivax MSP119 and two helper T-cell epitopes, the synthetic uni-versal pan allelic DR epitope (PADRE) and a new internal MSP1 P. vivax epitope (DYDVVYLKPLAGMYK). We found that the recognition ofHis6MSP119-DYDVVYLKPLAGMYK-PADRE was as good as the recognition of His6MSP119 indicating that the presence of the T-cell epitopesPADRE and DYDVVYLKPLAGMYK did not modify the MSP119 epitopes recognized by human IgG. The recombinant protein His6MSP119-DYDVVYLKPLAGMYK-PADRE proved to be highly immunogenic in marmosets (Callithrix jacchus jacchus) when administered in incom-plete Freund’s adjuvant. However, when administered in other adjuvant formulations such as Quil A, CpG ODN 2006 or MPL/TDM, antibodytiters to MSP119 were significantly lower. Among these three adjuvants, Quil A proved to be the most efficient one generating antibody titerssignificantly higher than the others. These results indicated that under the circumstances evaluated, adjuvants were key for the immunogenicityof the recombinant protein His6MSP119-DYDVVYLKPLAGMYK-PADRE.� 2006 Elsevier SAS. All rights reserved.
Keywords: Plasmodium vivax; Recombinant vaccines; Adjuvants
Abbreviations: CFA, complete Freund’s adjuvant; CpG, immunostimulatory CpG motifs; EGF, epidermal growth factor; GST, glutathione S-transferase;
His6MSP119, MSP119 containing a hexa-histidine tag; IFA, incomplete Freund’s adjuvant; MPL/TDM, mono phosphoryl lipid Aþ trehalose dicorynomycolate;
MSP1, merozoite surface protein-1; MSP119, 19 kDa C-terminal region of the MSP1; ODN, oligodeoxynucleotide; PADRE, pan allelic DR epitope; TLR, Toll
like receptor.
* Corresponding author. Tel./fax: þ55 (11) 5571 1095.
E-mail address: [email protected] (M.M. Rodrigues).
1286-4579/$ - see front matter � 2006 Elsevier SAS. All rights reserved.
doi:10.1016/j.micinf.2006.03.012

2131D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
1. Introduction
Plasmodium vivax causes more than 40 million cases ofmalaria every year and an effective vaccine is urgently needed[1]. One of the most promising vaccine candidates against theerythrocytic forms of malaria is the merozoite surface protein1 (MSP1). During the invasion process, a proteolytic cleavagestep releases most of the molecule from the merozoite mem-brane and only a 19 kDa glycosylphosphatidylinositol an-chored fragment of the C-terminus (MSP119) is carried intothe invaded red blood cells [2]. Although the precise biologicalfunction of MSP119 is unknown at present, reverse genetic ex-periments using Plasmodium falciparum provided evidencethat MSP119 is important for parasite survival [3].
Antibodies that recognize MSP119 are potent inhibitors ofmerozoite invasion in vitro, and confer passive immunityagainst rodent malaria infection [4,5]. However, most relevantfor the development of a vaccine against malaria is the factthat active immunization with recombinant proteins based onthe sequence of MSP119 can provide remarkable protectiveimmunity against experimental infection with blood stagesof distinct species of Plasmodium [6e8].
As part of our studies aimed at the development of a Plas-modium vivax malaria vaccine, we are characterizing the im-munogenic properties of several recombinant proteinscontaining the P. vivax MSP119 [9e14]. Bacterial recombinantproteins expressed in the pET vector (His6MSP119) retainednative epitopes being recognized by antibodies of 93.5% ofBrazilian individuals exposed to P. vivax malaria [13]. The ad-dition of a synthetic universal T-cell epitope denominated PanAllelic DR epitope (PADRE) did not modify the MSP119 epi-topes recognized by human naturally acquired IgG [11,13] andimproved the performance of several adjuvant formulationswhen the antigen was administered to a mouse strain, suchas C57BL/6, which develops PADRE-specific helper CD4T cells [14].
In the present study, we generated a new bacterial recombi-nant protein in which we added a new parasite derived putativepromiscuous MSP1 internal helper T-cell epitope to the pro-tein His6MSP119-PADRE, with the rationale of increasingcognate help. Our assumption is that a cognate help may im-prove a human vaccine performance as previously demon-strated in the mouse model of blood stage infection [15].The immunogenic properties of this new recombinant proteindenominated His6MSP119-DYDVVYLKPLAGMYK-PADREwere evaluated by its ability to be recognized by serum IgGantibodies from individuals with patent malarial infectionand upon immunization of non-human primates in the pres-ence of different adjuvant formulations.
2. Materials and methods
2.1. Mapping of a T-cell epitope on the 33 kDa regionof P. vivax MSP1
The amino acid sequence of the 33 kDa region of the C-ter-minal of P. vivax MSP1 was scanned by TEPITOPE algorithm
that can predict epitopes that have a potential ability to bind toone or more of 25 different HLA-DR molecules, by using 25virtual matrices that cover most of the HLA class II peptidebinding specificities in the Caucasian population [16,17].The algorithm provides a scoredthe algebraic sum of the ma-trix values for each peptide positiondto each of the 9-merwindows along the scanned sequence. Nonamers attaininga score above the 3% threshold for a given HLA-DR molecule(a 3% threshold selects sequences with HLA-binding scoresequal to or higher than those of the 3% sequences with highestscores in the TEPITOPE database) are selected by the soft-ware. The algorithm also allows the selection of sequencespredicted to bind simultaneouslydthus, promiscuouslydtoseveral HLA-DR molecules. The sequence within MSP1,DYDVVYLKPLAGMYK, predicted to bind to 24 out of the25 HLA-DR molecules available at TEPITOPE (with the ex-ception of HLA-DR7) of HLA-DR molecules at the 3%threshold was selected.
2.2. Synthetic peptide and binding assay to humanHLA DR molecules
We synthesized the peptide DYDVVYLKPLAGMYK cor-responding to an inner nonamer core selected by TEPITOPEas the multiple HLA-binding motif, with three flanking aminoacids added at both N- and C-terminal ends, to increase the ef-ficiency of in vitro peptide presentation to CD4þ T cells, usingsolid phase technology with the 9-fluorenylmethoxycarbonyl(Fmoc) strategy [18] on an automated benchtop simultaneousmultiple solid-phase peptide synthesizer PSSM8 (Shimadzu,Tokyo, Japan) with Fmoc protected amino acid residues andTGR resin (Novabiochem, San Diego, USA). Therefore, thepeptide was obtained with the C-terminal carboxyl group inamide form. Peptide quality was assessed by Maldi-Tofmass spectometry on a TofSpec-E instrument (Micromass,UK) using a-cyano-4 hydroxy cinnamic acid as the matrix,and its purity was assessed as 90%.
Peptide-binding affinity to purified HLA-DR moleculeswas determined as described previously [19]. The nanomolarconcentration of unlabeled peptide necessary for 50% inhibi-tion of the labeled peptide to the purified HLA-DR molecules(IC50) was used as an approximation of the affinity of interac-tion (KD). The values reported are IC50 nM values.
2.3. Recombinant proteins containing P. vivax MSP119
The recombinant proteins were obtained exactly as de-scribed in reference [12]. We inserted the oligonucleotidescontaining the sequence encoding the 15 amino acids DYDV-VYLKPLAGMYK and the nucleotides to ligate to the twoBamHI site as described for the insertion of the PADRE[12]. This plasmid was used to express the recombinant pro-teins denominated His6MSP119-DYDVVYLKPLAGMYKHis6MSP119-DYDVVYLKPLAGMYK-PADRE. The recom-binant proteins that we used in the present study are listedin Table 1. The expression and purification of the recombinantproteins produced by E. coli transformed with the pET or

2132 D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
pGEX vectors were performed exactly as described [12]. Pu-rified proteins were analyzed by SDS-PAGE and stainedwith Coomassie blue or silver nitrate.
2.4. Immunization of mice with the recombinant proteins
Six to eight week old female C57BL/10.A (H-2a) micewere purchased from Universidade Federal de S~ao Paulo-Escola Paulista de Medicina. Groups of six mice were immu-nized twice, 4 weeks apart, with 10 mg of recombinant proteinin the presence of the indicated adjuvant formulations. Micewere injected subcutaneously (s.c.) in the two hind footpadswith the recombinant antigen emulsified or mixed with adju-vant. A volume of 50 ml was injected into each hind footpad.Four weeks later, each animal received a booster injection ofthe same antigen s.c. at the base of the tail. For these immuni-zations, 10 mg of the indicated recombinant proteins were di-luted in PBS and emulsified in complete or incompleteFreund’s adjuvant (CFA/IFA, 0.05 ml/dose per animal,Sigma). Control mice received only the diluent PBS and theindicated adjuvant. Mice were bled at 4 weeks after the firstor second antigen dose through the tail vein and individual se-rum samples were stored at �20 �C until used.
2.5. Immunization of non-human primateswith the recombinant proteinHis6MSP119-DYDVVYLKPLAGMYK-PADRE
A total of 24 adult (male and female) marmosets (Callithrixjacchus jacchus) weighting between 210 and 400 g were se-lected and randomized into eight groups. According to Brazil-ian law on animal experimentation, the protocol of this studywas reviewed and approved by the institute’s animal care andethical committee. Animals were under the supervision of a vet-erinarian specialized in primatology. Each group contained twomales and one female. Two groups received adjuvant only: (i)Quil A (250 mg/animal per dose), (ii) a mixture of Quil A(250 mg/animal per dose) and monophosphoryl lipid Aþ treha-lose dicorynomycolate (MPL/TDM, 0.3 ml/animal per dose,Sigma). The other six groups received 100 mg of the antigenHis6MSP119-DYDVVYLKPLAGMYK-PADRE in the follow-ing adjuvants: (i) IFA (0.3 ml/animal per dose), (ii) Quil A(250 mg/animal per dose), (iii) CpG ODN 2006 (500 mg/animalper dose, TCGTCGTTTTGTCGTTTTGTCGTT, Coley
Table 1
Recombinant proteins used
Designation Protein expressed
His6-MSP119 MSP119
GST-MSP119a GST-MSP119
His6-MSP119-PADRE MSP119 þ PADREb
His6MSP119-
DYDVVYLKPLAGMYK
MSP119 þ DYDVVYLKPLAGMYK
epitope
His6MSP119-
DYDVVYLKPLAGMYK-PADRE
MSP119 þ DYDVVYLKPLAGMYK
epitopeb þ PADRE
a Amino acids 1616e1704 of P. vivax MSP119 (Belem strain).b PADRE epitope is composed of amino acids AKFVAAWTLKAAA.
Pharmaceuticals), (iv) MPL/TDM (0.3 ml/animal per dose),(v) Quil Aþ CpG ODN 2006, (v) Quil Aþ MPL/TDM.
A total dose of 0.6 ml of antigen/adjuvant formulation wasinjected subcutaneously. This volume was divided into four in-jections of 0.15 ml: two in the inguinal and two in the axillaryregion. The marmosets were immunized a total of four times,at weeks 0, 6, 12 and 18. After anesthesia with ketamin(10e20 mg/kg), blood samples were collected from the femo-ral vein at the indicated days just before each immunization.
During the experiment all animals were observed by trainedpersonnel for the local reactogenicity of each formulation. Weexamined the injection sites for skin warmth, skin erythema,skin edema, muscle induration, ulceration, abscess or other ab-normalities at days 1, 2 and 3 post injection. Systemic toxicity,fever and weight loss were also evaluated on these same days.
2.6. Immunological assays
The detection of human or mouse IgG antibodies againstMSP119 was performed by ELISA as described [9e14]. Anti-bodies to MSP119 in monkey sera were detected by ELISA us-ing goat anti-monkey IgG (Sigma) diluted 1:2000. Each serumwas analyzed in serial dilutions from 1:100 up to 1:1,638,400.The individual titers were considered as the highest dilution ofserum that presented an OD492 higher than 0.1. The results arepresented as the mean of log antibody titers � SD of six miceor three marmosets per group.
2.7. Statistical analysis
One-way ANOVA and Tukey’s honestly significantly dif-ferent (HSD) tests were used to compare the possible differ-ences between the mean values of the different groups.
3. Results
3.1. Expression and purification of recombinant proteinsHis6MSP119-DYDVVYLKPLAGMYK andHis6MSP119-DYDVVYLKPLAGMYK-PADRE
With the aim of boosting cognate help, we used the TEPI-TOPE algorithm to select a peptide sequence in the 33 kDa C-terminal region of P. vivax MSP1 that would most likely bindto multiple HLA-DR molecules. The amino acid sequenceDYDVVYLKPLAGMYK was predicted to be the most pro-miscuous HLA-DR ligand in the 33 kDa region of the P. vivaxMSP1. Synthesis of the corresponding peptide and directbinding assays with the eight most prevalent HLA-DR mole-cules in the general population showed that the peptideDYDVVYLKPLAGMYK bound to HLA-DRB1*0101,DRB1*1101, DRB1*0501, and DRB5*0101 (Table 2).
This epitope was then added to either His6MSP119 orto His6MSP119-PADRE generating the proteins denominatedHis6MSP119-DYDVVYLKPLAGMYK or His6MSP119-DYDVVYLKPLAGMYK-PADRE, respectively (Table 1).Recombinant proteins were expressed in large amounts andpurified from E. coli in aqueous buffer in the absence of

2133D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
detergents. Fig. 1 shows the migration pattern of the purifiedproteins in SDS-PAGE performed under reducing or non re-ducing conditions.
3.2. T-cell helper activity in mice induced by the epitopeDYDVVYLKPLAGMYK
To test whether the epitope DYDVVYLKPLAGMYK couldin fact be recognized by mouse T cells, we immunized differ-ent inbred mice (BALB/c, C57BL/6 and C57BL/10.A) withthe synthetic peptide containing the amino acids DYDV-VYLKPLAGMYK. We found that only C57BL/10.A (H-2a)mice presented specific T-cell proliferative response in vitroto this synthetic peptide (data not shown).
Based on this result, we immunized C57BL/10.A mice withthe recombinant proteins His6MSP119 or His6MSP119-DYDV-VYLKPLAGMYK in the presence of different adjuvant formu-lations. We also used the recombinant protein GST-MSP119 asa positive control. As shown in Table 3, after the first immuniz-ing dose in the presence of CFA, the antibody titers to the epitopeHis6MSP119 of C57BL/10.A mice immunized with
Table 2
Binding values of the peptide DYDVVYLKPLAGMYK to different HLA DR
molecules
HLA-DR molecule IC50 nM values
DR1 (DRB1*0101) 0.73a
DR5 (DRB1*1101) 376a
DR2w2 B1(DRB1*0501) 793a
DR2w2 B2 (DRB5*0101) 0.080a
DR4w4 (DR401) 1636
DR4w15 (DR405) 2053
DR6w19 (DRB1*1302) 5849
DR7 (DRB1*0701) 7294
a IC50 binding values lower than 1000 are significant.
MWkDa
97-
4 5 6
66-
45-
30-
20-
14-
MWkDa 1 2 3
97-
66-
45-
30-
20-
14-
Fig. 1. SDS-PAGE analysis of recombinant proteins of P. vivax MSP119 puri-
fied from E. coli. Proteins were submitted to 12.5% SDS-PAGE under reduc-
ing (left panel) or non-reducing (right panel) conditions and stained with silver
nitrate. Lanes are: 1 and 4, mol. weight standards; 2 and 5, His6MSP119-
DYDVVYLKPLAGMYK; 3 and 6, His6MSP119-DYDVVYLKPLAGMYK-
PADRE.
His6MSP119-DYDVVYLKPLAGMYK was 56.23-fold higherthan animals immunized with the antigen His6MSP119
(P < 0.01). After the second immunizing dose in the presenceof IFA, the titers of mice immunized with His6MSP119-DYDV-VYLKPLAGMYK were still 5.33-fold higher than mice in-jected with His6MSP119 (P < 0.01).
A similar improvement of the antibody immune responsewas observed in C57BL/10.A mice immunized withHis6MSP119-DYDVVYLKPLAGMYK in the presence ofIFA. After the second immunizing dose, antibody titers ofmice immunized with His6MSP119-DYDVVYLKPLAGMYKwere 6.76-fold higher than the titers of mice injected withHis6MSP119 (P < 0.01). Together, these results indicatedthat the epitope DYDVVYLKPLAGMYK can function asa helper T-cell epitope.
3.3. Comparative evaluation of the reactivity of thedifferent recombinant proteins with serum samples fromindividuals with patent P. vivax infection
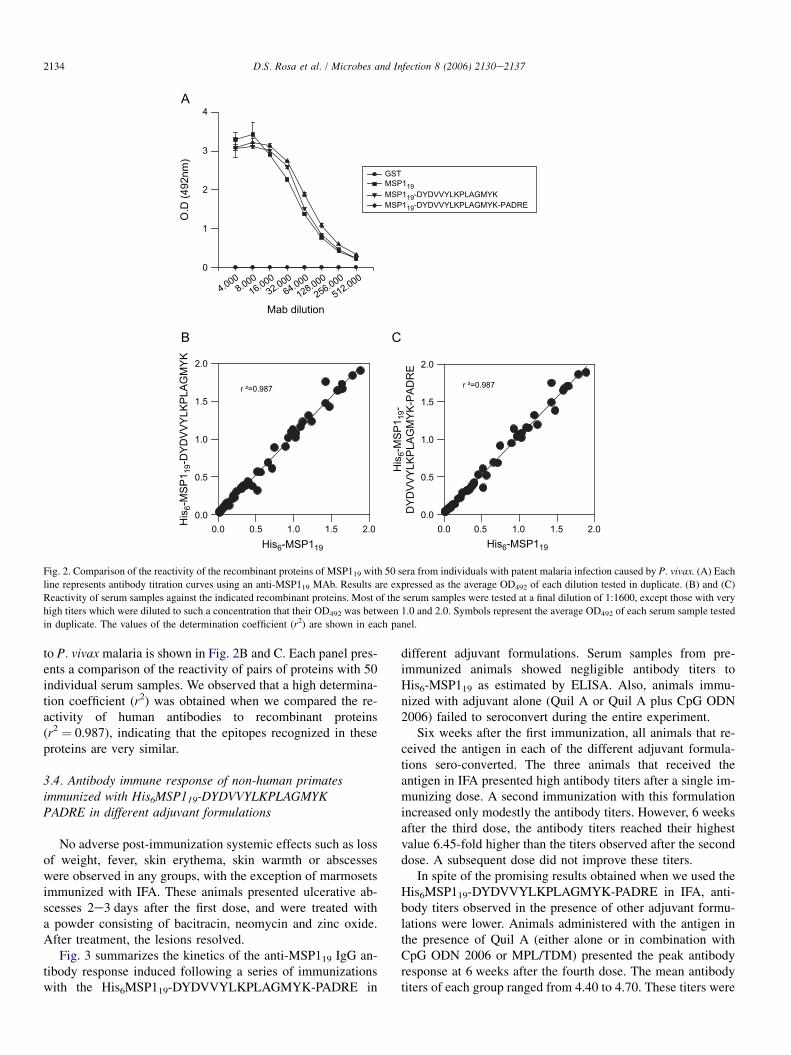
The recognition of the recombinant proteins His6MSP119,His6MSP119-DYDVVYLKPLAGMYK and His6MSP119-DYDVVYLKPLAGMYK-PADRE by a mAb 3F8.A2 specificfor MSP119 was estimated first. This mAb recognizes a confor-mation-dependent epitope [11]. As shown in Fig. 2A, the dif-ferent recombinant proteins were equally well recognized bythe anti-MSP119 MAb.
The comparative analysis of the reactivity of the recombi-nant proteins with sera from 50 individuals naturally exposed
Table 3
Antibody titers of C57BL/10.A mice immunized with the recombinant pro-
teins in different adjuvant formulations
Recombinant antigena Adjuvant Antibody titers
after the first
dose (log)
Antibody titers
after the second
dose (log)
e CFA/IFA <2.0b <2.0b
His6 MSP119 CFA/IFA 3.81 � 0.85b 4.90 � 0.15b
His6MSP119-
DYDVVYLKPLAGMYK
CFA/IFA 5.56 � 0.35b 5.61 � 0.60b
GST- MSP119 CFA/IFA 5.24 � 0.38b 5.16 � 0.39b
His6MSP119 IFA 3.80 � 0.42c 4.55 � 0.21c
His6MSP119-
DYDVVYLKPLAGMYK
IFA 4.40 � 0.24c 5.38 � 0.15c
GST-MSP119 IFA 4.17 � 0.15c 4.63 � 0.29c
a C57BL/10.A mice were immunized as described in Section 2. Results are
expressed as the mean of six mice � SD of log antibody titers detected
4 weeks after the first or second immunizing dose.b The results obtained for the different groups were compared statistically
using one-way ANOVA followed by the Tukey HSD test. After the first or sec-
ond dose, the antibody titers of mice immunized with His6MSP119 were lower
than the mouse group injected with His6MSP119-DYDVVYLKPLAGMYK or
GST-MSP119 (P < 0.01 in both cases). Mice immunized with adjuvant only
had significantly lower antibody titers than mice immunized with the different
recombinant proteins (P < 0.01 in all cases).c After the first dose, antibody titers of mice immunized with the different
recombinant proteins in IFA were not different from each other. After the
second dose, antibody titers of mice immunized with His6MSP119 DYDV-
VYLKPLAGMYK were higher than those immunized with His6MSP119 or
GST- MSP119 (P < 0.01 and P < 0.05 respectively).
2134 D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
O.D
(492
nm)
0
1
2
3
4A
B C
4.0008.000
16.00032.000
64.000
128.000
256.000
512.000
GST MSP119 MSP119-DYDVVYLKPLAGMYKMSP119-DYDVVYLKPLAGMYK-PADRE
Mab dilution
His6-MSP119 His6-MSP119
0.0 0.5 1.0 1.5 2.0 0.0 0.5 1.0 1.5 2.0
His
6-M
SP1 1
9-D
YDVV
YLKP
LAG
MYK
0.0
0.5
1.0
1.5
2.0
0.0
0.5
1.0
1.5
2.0
r ²=0.987H
is6-
MSP
1 19-
DYD
VVYL
KPLA
GM
YK-P
ADR
E
r ²=0.987
Fig. 2. Comparison of the reactivity of the recombinant proteins of MSP119 with 50 sera from individuals with patent malaria infection caused by P. vivax. (A) Each
line represents antibody titration curves using an anti-MSP119 MAb. Results are expressed as the average OD492 of each dilution tested in duplicate. (B) and (C)
Reactivity of serum samples against the indicated recombinant proteins. Most of the serum samples were tested at a final dilution of 1:1600, except those with very
high titers which were diluted to such a concentration that their OD492 was between 1.0 and 2.0. Symbols represent the average OD492 of each serum sample tested
in duplicate. The values of the determination coefficient (r2) are shown in each panel.
to P. vivax malaria is shown in Fig. 2B and C. Each panel pres-ents a comparison of the reactivity of pairs of proteins with 50individual serum samples. We observed that a high determina-tion coefficient (r2) was obtained when we compared the re-activity of human antibodies to recombinant proteins(r2 ¼ 0.987), indicating that the epitopes recognized in theseproteins are very similar.
3.4. Antibody immune response of non-human primatesimmunized with His6MSP119-DYDVVYLKPLAGMYKPADRE in different adjuvant formulations
No adverse post-immunization systemic effects such as lossof weight, fever, skin erythema, skin warmth or abscesseswere observed in any groups, with the exception of marmosetsimmunized with IFA. These animals presented ulcerative ab-scesses 2e3 days after the first dose, and were treated witha powder consisting of bacitracin, neomycin and zinc oxide.After treatment, the lesions resolved.
Fig. 3 summarizes the kinetics of the anti-MSP119 IgG an-tibody response induced following a series of immunizationswith the His6MSP119-DYDVVYLKPLAGMYK-PADRE in
different adjuvant formulations. Serum samples from pre-immunized animals showed negligible antibody titers toHis6-MSP119 as estimated by ELISA. Also, animals immu-nized with adjuvant alone (Quil A or Quil A plus CpG ODN2006) failed to seroconvert during the entire experiment.
Six weeks after the first immunization, all animals that re-ceived the antigen in each of the different adjuvant formula-tions sero-converted. The three animals that received theantigen in IFA presented high antibody titers after a single im-munizing dose. A second immunization with this formulationincreased only modestly the antibody titers. However, 6 weeksafter the third dose, the antibody titers reached their highestvalue 6.45-fold higher than the titers observed after the seconddose. A subsequent dose did not improve these titers.
In spite of the promising results obtained when we used theHis6MSP119-DYDVVYLKPLAGMYK-PADRE in IFA, anti-body titers observed in the presence of other adjuvant formu-lations were lower. Animals administered with the antigen inthe presence of Quil A (either alone or in combination withCpG ODN 2006 or MPL/TDM) presented the peak antibodyresponse at 6 weeks after the fourth dose. The mean antibodytiters of each group ranged from 4.40 to 4.70. These titers were
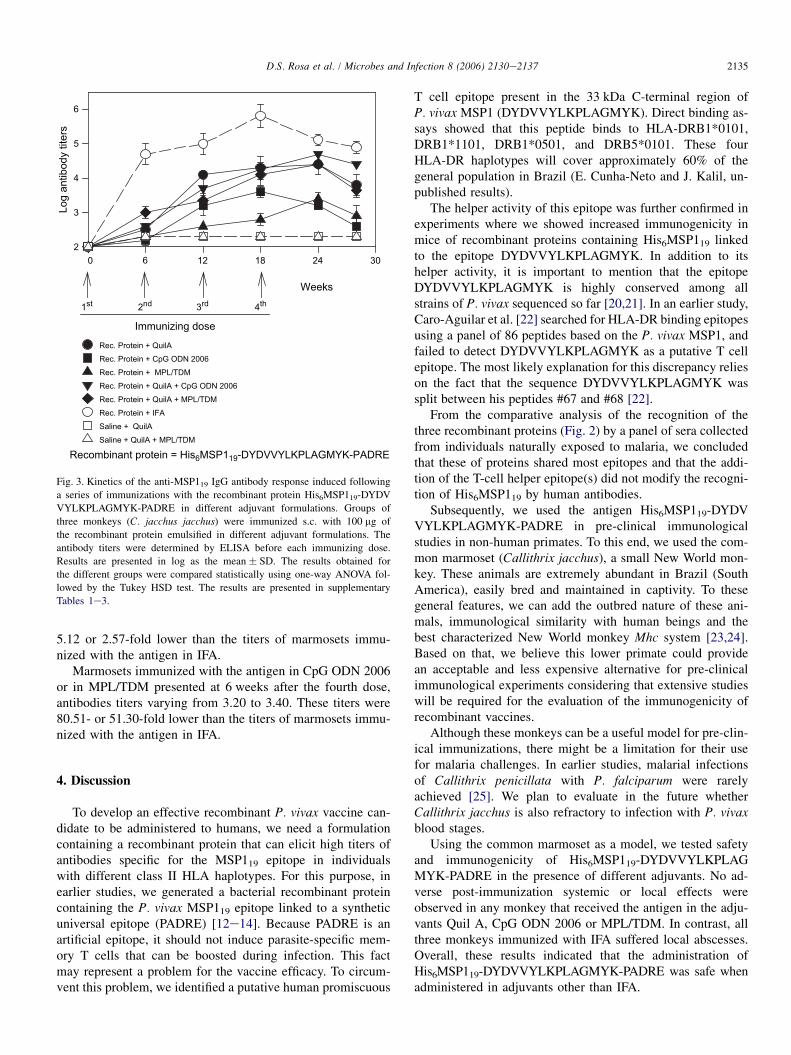
2135D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
5.12 or 2.57-fold lower than the titers of marmosets immu-nized with the antigen in IFA.
Marmosets immunized with the antigen in CpG ODN 2006or in MPL/TDM presented at 6 weeks after the fourth dose,antibodies titers varying from 3.20 to 3.40. These titers were80.51- or 51.30-fold lower than the titers of marmosets immu-nized with the antigen in IFA.
4. Discussion
To develop an effective recombinant P. vivax vaccine can-didate to be administered to humans, we need a formulationcontaining a recombinant protein that can elicit high titers ofantibodies specific for the MSP119 epitope in individualswith different class II HLA haplotypes. For this purpose, inearlier studies, we generated a bacterial recombinant proteincontaining the P. vivax MSP119 epitope linked to a syntheticuniversal epitope (PADRE) [12e14]. Because PADRE is anartificial epitope, it should not induce parasite-specific mem-ory T cells that can be boosted during infection. This factmay represent a problem for the vaccine efficacy. To circum-vent this problem, we identified a putative human promiscuous
Weeks
0 12 18 24 30
Log
antib
ody
titer
s
2
3
4
5
6
Rec. Protein + QuilARec. Protein + CpG ODN 2006Rec. Protein + MPL/TDMRec. Protein + QuilA + CpG ODN 2006 Rec. Protein + QuilA + MPL/TDMRec. Protein + IFASaline + QuilA Saline + QuilA + MPL/TDM
1st 2nd 3rd 4th
Immunizing dose
Recombinant protein = His6MSP119-DYDVVYLKPLAGMYK-PADRE
6
Fig. 3. Kinetics of the anti-MSP119 IgG antibody response induced following
a series of immunizations with the recombinant protein His6MSP119-DYDV
VYLKPLAGMYK-PADRE in different adjuvant formulations. Groups of
three monkeys (C. jacchus jacchus) were immunized s.c. with 100 mg of
the recombinant protein emulsified in different adjuvant formulations. The
antibody titers were determined by ELISA before each immunizing dose.
Results are presented in log as the mean � SD. The results obtained for
the different groups were compared statistically using one-way ANOVA fol-
lowed by the Tukey HSD test. The results are presented in supplementary
Tables 1e3.
T cell epitope present in the 33 kDa C-terminal region ofP. vivax MSP1 (DYDVVYLKPLAGMYK). Direct binding as-says showed that this peptide binds to HLA-DRB1*0101,DRB1*1101, DRB1*0501, and DRB5*0101. These fourHLA-DR haplotypes will cover approximately 60% of thegeneral population in Brazil (E. Cunha-Neto and J. Kalil, un-published results).
The helper activity of this epitope was further confirmed inexperiments where we showed increased immunogenicity inmice of recombinant proteins containing His6MSP119 linkedto the epitope DYDVVYLKPLAGMYK. In addition to itshelper activity, it is important to mention that the epitopeDYDVVYLKPLAGMYK is highly conserved among allstrains of P. vivax sequenced so far [20,21]. In an earlier study,Caro-Aguilar et al. [22] searched for HLA-DR binding epitopesusing a panel of 86 peptides based on the P. vivax MSP1, andfailed to detect DYDVVYLKPLAGMYK as a putative T cellepitope. The most likely explanation for this discrepancy relieson the fact that the sequence DYDVVYLKPLAGMYK wassplit between his peptides #67 and #68 [22].
From the comparative analysis of the recognition of thethree recombinant proteins (Fig. 2) by a panel of sera collectedfrom individuals naturally exposed to malaria, we concludedthat these of proteins shared most epitopes and that the addi-tion of the T-cell helper epitope(s) did not modify the recogni-tion of His6MSP119 by human antibodies.
Subsequently, we used the antigen His6MSP119-DYDVVYLKPLAGMYK-PADRE in pre-clinical immunologicalstudies in non-human primates. To this end, we used the com-mon marmoset (Callithrix jacchus), a small New World mon-key. These animals are extremely abundant in Brazil (SouthAmerica), easily bred and maintained in captivity. To thesegeneral features, we can add the outbred nature of these ani-mals, immunological similarity with human beings and thebest characterized New World monkey Mhc system [23,24].Based on that, we believe this lower primate could providean acceptable and less expensive alternative for pre-clinicalimmunological experiments considering that extensive studieswill be required for the evaluation of the immunogenicity ofrecombinant vaccines.
Although these monkeys can be a useful model for pre-clin-ical immunizations, there might be a limitation for their usefor malaria challenges. In earlier studies, malarial infectionsof Callithrix penicillata with P. falciparum were rarelyachieved [25]. We plan to evaluate in the future whetherCallithrix jacchus is also refractory to infection with P. vivaxblood stages.
Using the common marmoset as a model, we tested safetyand immunogenicity of His6MSP119-DYDVVYLKPLAGMYK-PADRE in the presence of different adjuvants. No ad-verse post-immunization systemic or local effects wereobserved in any monkey that received the antigen in the adju-vants Quil A, CpG ODN 2006 or MPL/TDM. In contrast, allthree monkeys immunized with IFA suffered local abscesses.Overall, these results indicated that the administration ofHis6MSP119-DYDVVYLKPLAGMYK-PADRE was safe whenadministered in adjuvants other than IFA.

2136 D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
In animals vaccinated with His6MSP119-DYDVVYLK-PLAGMYK-PADRE emulsified in IFA, antibody titers werevery high demonstrating that this recombinant protein can behighly immunogenic. IFA is used as a water-in-oil emulsionthat is based on mineral oil and the surfactant mannide mono-oleate. This adjuvant was initially tested in human trials in1950. Its use was stopped after vaccine trials because it causedsevere local reactions in a number of vaccinated individuals.Its mode of action is still unclear, however its potency is cer-tain (reviewed in ref. [26]).
Two of the adjuvants that we used are described as TLR ac-tivators. The MPL or CpG ODN 2006 activates TLR-4 orTLR-9, respectively [27]. Unfortunately, these adjuvants failedto perform at the level of the QuilA or the IFA when used in-dividually. Also, when added to the QuilA, they did not im-prove its adjuvant activity (Fig. 3).
QuilA, a saponin derived from the bark of Chilean tree,Quillaja saponaria, is still being studied on its mechanismsof adjuvanticity [28]. In our trial, it performed better thanMPL or CpG ODN 2006 but still provided lower antibody ti-ters than IFA.
The reason for such difference in the antibody titersamong the animals injected with the distinct adjuvants isnot clear. In BALB/c mice, we described similar findings.Administration of His6MSP119-PADRE in CFA/IFA elicitedhigher antibody titers than the antigen in CpG ODN, MPL/TDM or Quil A [26]. On the other hand, in C57BL/6 mice,the immunogenicity of His6MSP119-PADRE administered inCpG ODN, MPL/TDM or Quil A was similar to that obtainedin CFA/IFA. The improvement was likely due to the fact thatthis mouse strain developed T cells specific to PADRE epi-tope [14]. We concluded that the presence of strong T helperepitopes such as PADRE improved the performance ofweaker adjuvant such as CpG ODN, MPL/TDM or Quil A.Based on the mouse studies, we may speculate that the differ-ences in the antibody titers observed in monkeys immunizedwith the distinct adjuvants could be due to the lack of a strongT cell epitope.
In spite of the fact that the antibody titers obtained with theadjuvant QuilA were not as high as IFA, we considered thatour vaccination trials provided important information. Firstly,we established that the recombinant protein His6MSP119-DYDVVYLKPLAGMYK-PADRE can be highly immuno-genic in primates. Secondly, we developed a new monkeymodel that opens the possibility to explore other adjuvantformulations. Many of these adjuvants are currently beingtested with recombinant proteins based on the MSP1 or otherP. falciparum antigens [29,30]. We may use this informationto select the next group of adjuvants to be tested. Thirdly,the serum generated during these trials can be tested in func-tional assays.
It is important to mention that both T cell epitopes that weintroduced (DYDVVYLKPLAGMYK and PADRE) were de-signed/selected to bind to human HLA-DR molecules. There-fore, it is possible that this recombinant protein may performbetter in humans than in monkeys. Nevertheless, our resultssupport previous studies that concluded that a major challenge
in the development of subunit vaccines for malaria or other in-fectious diseases will be the identification of safe and potentadjuvants capable of inducing immune responses as high asCFA/IFA.
Acknowledgements
This work was supported by grants from FAPESP, TheMillennium Institute for Vaccine Development and Technol-ogy (CNPq - 420067/2005-1) and UNDP/World Bank/WHO/TDR ID 990259. The authors are indebted to Dr J. W. Barn-well (CDC, Atlanta) who provided the mAb anti-MSP119.I.S.S., J.K., L.E.M., E.C.N and M.M.R. were supported by fel-lowships from CNPq. D.S.R., F.T. and D.Y.B. were supportedby fellowships from FAPESP.
References
1 K. Mendis, B.J. Sina, P. Marchesini, R. Carter, The neglected burden of
Plasmodium vivax malaria, Am. J. Trop. Med. Hyg 64 (2001) 97e106.
2 M.J. Blackman, H. Heidrich, S. Donachie, J.S. McBride, A.A. Holder, A
single fragment of a malaria merozoite surface protein remains on the
parasite during red cell invasion and is the target of invasion-inhibiting
antibodies, J. Exp. Med 172 (1990) 379e382.
3 R.A. O’Donnell, A. Saul, A.F. Cowman, B.S. Crabb, Functional conserva-
tion of the malaria vaccine antigen MSP-119 across distantly related
Plasmodium species, Nat. Med 6 (2000) 91e95.
4 R.A. O’Donnell, T.F. de Koning-Ward, R.A. Burt, M. Bockarie,
J.C. Reeder, A.F. Cowman, B.S. Crabb, Antibodies against merozoite sur-
face protein (MSP)-1(19) are a major component of the invasion-inhibitory
response in individuals immune to malaria, J. Exp. Med 193 (2001)
1403e1412.
5 L.M. Spencer Valero, S.A. Ogun, S.L. Fleck, I.T. Ling, T.J. Scott-Finnigan,
M.J. Blackman, A.A. Holder, Passive immunization with antibodies
against three distinct epitopes on Plasmodium yoelii merozoite surface
protein 1 suppresses parasitemia, Infect. Immun. 66 (1998) 3925e3930.
6 T.M. Daly, C.A. Long, A recombinant 15-kilodalton carboxyl-terminal
fragment of Plasmodium yoelii 17XL merozoite surface protein-1 induces
a protective immune response in mice, Infect. Immun. 61 (1993)
2462e2467.
7 K.L.R.L. Perera, S.M. Handunnetti, I. Holm, S. Longacre, K. Mendis,
Baculovirus merozoite surface protein 1 C-terminal recombinant antigens
are highly protective in a natural primate model for human Plasmodium
vivax malaria, Infect. Immun. 66 (1998) 1500e1506.
8 A.W. Stowers, V. Cioce, R.L. Shimp, M. Lawson, G. Hui, O. Muratova,
D.C. Kaslow, R. Robinson, C.A. Long, L.H. Miller, Efficacy of two alter-
nate vaccines based on Plasmodium falciparum merozoite surface protein 1
in an Aotus challenge trial, Infect. Immun. 69 (2001) 1536e1546.
9 I.S. Soares, G. Levitus, J.M. Souza, H.A. del Portillo, M.M. Rodrigues,
Acquired immune responses to the N- and C-terminal regions of Plasmo-
dium vivax merozoite surface protein 1 in individuals exposed to malaria,
Infect. Immun. 65 (1997) 1606e1614.
10 I.S. Soares, M.G. Cunha, M.N. Silva, J.M. Souza, H.A. del Portillo,
M.M. Rodrigues, Longevity of the naturally acquired antibody responses
to the N- and C-terminal regions of Plasmodium vivax MSP1, Am. J.
Trop. Med. Hyg. 60 (1999) 357e363.
11 I.S. Soares, J.W. Barnwell, M.U. Ferreira, M.G. Cunha, J.P. Laurino,
B.A. Castilho, M.M. Rodrigues, A Plasmodium vivax vaccine candidate
displays limited allele polymorphism, which does not restrict recognition
by antibodies, Mol. Med. 5 (1999) 459e470.
12 M.G. Cunha, M.M. Rodrigues, I.S. Soares, Comparison of the immuno-
genic properties of recombinant proteins representing the Plasmodium
vivax vaccine candidate MSP119 expressed in distinct bacterial vectors,
Vaccine 20 (2001) 385e396.

2137D.S. Rosa et al. / Microbes and Infection 8 (2006) 2130e2137
13 M.H. Rodrigues, M.G. Cunha, R.L. Machado, O.C. Ferreira Jr.,
M.M. Rodrigues, I.S. Soares, Serological detection of Plasmodium vivax
malaria using recombinant proteins corresponding to the 19-kDa C-termi-
nal region of the merozoite surface protein-1, Malar. J. 2 (2003) 39.
14 D.S. Rosa, F. Tzelepis, M.G. Cunha, I.S. Soares, M.M. Rodrigues, The pan
HLA DR-binding epitope improves adjuvant-assisted immunization with
a recombinant protein containing a malaria vaccine candidate, Immunol.
Lett. 92 (2004) 259e268.
15 R. Stephens, F.R. Albano, S. Quin, B.J. Pascal, V. Harrison, B. Stockinger,
D. Kioussis, H.U. Weltzien, J. Langhorne, Malaria-specific transgenic
CD4(þ) T cells protect immunodeficient mice from lethal infection and
demonstrate requirement for a protective threshold of antibody production
for parasite clearance, Blood 106 (2005) 1676e1684.
16 T. Sturniolo, E. Bono, J. Ding, L. Raddrizzani, O. Tuereci, U. Sahin,
M. Braxenthaler, F. Gallazzi, M.P. Protti, F. Sinigaglia, J. Hammer, Gener-
ation of tissue-specific and promiscuous HLA ligand databases using DNA
microarrays and virtual HLA class II matrices, Nat. Biotechnol. 17 (1999)
555e561.
17 J. Hammer, C. Belunis, D. Bolin, J. Papadopoulos, R. Walsky, J. Higelin,
W. Danho, F. Sinigaglia, Z.A. Nagy, High-affinity binding of short peptides
to major histocompatibility complex class II molecules by anchor combi-
nations, Proc. Natl. Acad. Sci. U.S.A. 91 (1994) 4456e4460.
18 E. Atherton, R.C. Sheppard, Solid Phase Peptide Synthesis: A Practical
Approach, IRL Press, Oxford, 1989, 152 pp.
19 S. Southwood, J. Sidney, A. Kondo, M.F. del Guercio, E. Appella,
S. Hoffman, R.T. Kubo, R.W. Chesnut, H.M. Grey, A. Sette, Several com-
mon HLA-DR types share largely overlapping peptide binding repertoires,
J. Immunol. 160 (1998) 3363e3673.
20 C. Putaporntip, S. Jongwutiwes, S. Seethamchai, H. Kanbara, K. Tanabe,
Intragenic recombination in the 30 portion of the merozoite surface protein
1 gene of Plasmodium vivax, Mol. Biochem. Parasitol. 109 (2000)
111e119.
21 C. Putaporntip, S. Jongwutiwes, N. Sakihama, M.U. Ferreira, W.G. Kho,
A. Kaneko, H. Kanbara, T. Hattori, K. Tanabe, Mosaic organization and
heterogeneity in frequency of allelic recombination of the Plasmodiumvivax merozoite surface protein-1 locus, Proc. Natl. Acad. Sci. U.S.A. 99
(2002) 16348e16353.
22 I. Caro-Aguilar, A. Rodriguez, J.M. Calvo-Calle, F. Guzman, P. De la
Vega, M.E. Patarroyo, M.R. Galinski, A. Moreno, Plasmodium vivaxpromiscuous T-helper epitopes defined and evaluated as linear peptide
chimera immunogens, Infect. Immun. 70 (2002) 3479e3492.
23 R.E. Bontrop, N. Otting, N.G. de Groot, G.G. Doxiadis, Major histocom-
patibility complex class II polymorphisms in primates, Immunol. Rev. 167
(1999) 339e350.
24 C.P. Genain, S.L. Hauser, Experimental allergic encephalomyelitis in the
New World monkey Callithrix jacchus, Immunol, Rev. 183 (2001)
159e172.
25 E.M. Rocha, L.H. Pereira, V.E. V.E do Rosario, A.U. Krettli, Refractori-
ness of Callithrix penicillata red blood cells to Plasmodium falciparum
in vivo and in vitro, J. Parasitol. 74 (1988) 514e515.
26 L.H. Miller, A. Saul, S. Mahanty, Revisiting Freund’s incomplete adju-
vant for vaccines in the developing world, Trends Parasitol 21 (2005)
412e414.
27 D. van Duin, R. Medzhitov, A.C. Shaw, Triggering TLR signaling in
vaccination, Trends Immunol. 27 (2006) 49e55.
28 M. Singh, D.T. O’Hagan, Recent advances in vaccine adjuvants, Pharm.
Res. 19 (2002) 715e728.
29 S. Pichyangkul, M. Gettayacamin, R.S. Miller, J.A. Lyon, E. Angov,
P. Tongtawe, D.L. Ruble, D.G. Heppner Jr., K.E. Kester, W.R. Ballou,
C.L. Diggs, G. Voss, J.D. Cohen, D.S. Walsh, Pre-clinical evaluation of
the malaria vaccine candidate P. falciparum MSP1(42) formulated with
novel adjuvants or with alum, Vaccine 22 (2004) 3831e3840.
30 L.J. Carvalho, S.G. Oliveira, M. Theisen, F.A. Alves, M.C. Andrade,
G.M. Zanini, M.C. Brigido, C. Oeuvray, M.M. Povoa, J.A. Muniz,
P. Druilhe, C.T. Daniel-Ribeiro, Immunization of Saimiri sciureus
monkeys with Plasmodium falciparum merozoite surface protein-3 and
glutamate-rich protein suggests that protection is related to antibody levels,
Scand. J. Immunol. 59 (2004) 363e372.
Copyright © 2022 FDOKUMEN






![The infrared spectrum of the O⋯H⋯O fragment of H[sub 5]O[sub 2][sup +]: Ab initio classical molecular dynamics and quantum 4D model calculations](https://static.fdokumen.com/doc/165x107/633857c36c6d131be60d0b92/the-infrared-spectrum-of-the-oho-fragment-of-hsub-5osub-2sup-ab-initio.jpg)

![Vibronic and spin-orbit coupling of a d [sup 9] transition-metal ion encapsulated in an icosahedral cage: The (Γ[sub 8]+Γ[sub 9])×(g+2h) Jahn-Teller problem](https://static.fdokumen.com/doc/165x107/634128d48383ee94150239f5/vibronic-and-spin-orbit-coupling-of-a-d-sup-9-transition-metal-ion-encapsulated.jpg)











