
Chloroplast DNA polymorphism reveals geographic structure and introgression in the Quercus...
14
Chloroplast DNA polymorphism reveals geographic structure and introgression in the Quercus crassifolia Quercus crassipes hybrid complex in Mexico E. Tovar-Sa ´ nchez, P. Mussali-Galante, R. Esteban-Jime ´ nez, D. Pin ˜ ero, D.M. Arias, O. Dorado, and K. Oyama Abstract: Quercus crassifolia H. & B. and Quercus crassipes H. & B. are two common temperate forest species in Mex- ico that form hybrids when they occur in mixed (sympatric) areas. In this study, we used chloroplast microsatellite haplo- types to infer phylogeographic patterns onto the genetic structure of populations of both parental species, thus defining probable colonization routes throughout Mexico. Haplotype diversity in seven hybrid zones and four allopatric sites with nonmixed (allopatric) populations of both putative parental species was analyzed. To determine the expansion and coloni- zation routes for the two species, we carried out a nested clade phylogeographic analysis that would allow us to infer the haplotype correspondence to a phylogeographical approach. In spite of sharing many common populations, we determined that there is a different genetic historical colonization for Q. crassipes and for Q. crassifolia. Hybrid populations had the highest levels of genetic variation (Gv), Shannon diversity index (H), and haplotype number (nh) in comparison with non- mixed/allopatric populations of their putative parentals. Furthermore, populations of hybrid zones showed the highest val- ues of genetic differentiation FST, RST . Moreover, both species share four distinctive cpDNA haplotypes, which were most likely acquired by introgression through hybrids located in several populations throughout the Eje Neovolca ´nico area. This last region was confirmed to be a ‘‘hot spot’’ for oak diversity, a place in which different maternal lineages merged. Re ´sume ´: Les Quercus crassifolia H. & B. et Quercus crassipes H. & B. repre ´sentent deux espe `ces communes des fore ˆts tempe ´re ´es du Mexique formant des hybrides lorsqu’elles se retrouvent me ´lange ´es dans des re ´gions sympatriques. Les au- teurs ont utilise ´ les haplotypes des microsatellites chloroplastiques, pour en de ´duire les patrons phyloge ´ne ´tique, sur la base de la structure ge ´ne ´tique des populations des deux espe `ces parentes, et ainsi de ´finir les routes probables de colonisation, pour l’ensemble du Mexique. Ils ont analyse ´ la diversite ´ des haplotypes dans huit zones d’hybridation et quatre sites allo- patriques supportant des populations non me ´lange ´es/allopatriques, pour les deux espe `ces parentes pre ´sume ´es. Afin de de ´- terminer l’expansion et les routes de colonisation pour les deux espe `ces, les auteurs ont conduit une analyse phyloge ´ographique de clades groupe ´s, qui permettrait de de ´duire la correspondance haplotypique, en vue d’une approche phyloge ´ographique. En de ´pit du partage de plusieurs populations communes, on montre qu’il y a une historique ge ´ne ´tique de colonisation diffe ´rente pour le Q. crassipes et le Q. crassifolia. Les populations hybrides posse `dent les plus hauts de- gre ´s de variation ge ´ne ´tique (G v ), d’index de diversite ´ de Shannon (H), et de nombre haplotypique (n h ), comparativement aux populations non me ´lange ´es/allopatriques, des parents pre ´sume ´s. De plus, les populations des zones hybrides montrent les plus grandes valeurs de diffe ´renciation ge ´ne ´tique F ST , R ST . Les deux espe `ces partagent e ´galement quatre haplotypes cpADN, qui ont e ´te ´ probablement acquis par introgression, via des hybrides situe ´s dans plusieurs populations de la re ´gion Eje Neovolcanico. On a de ´ja ` confirme ´ que cette re ´gion constitue un ‘point chaud’ pour la diversite ´ des che ˆnes, un endroit ou ` diffe ´rentes ligne ´es maternelles se sont confondues. Mots-cle ´s : cpSSRs, hybridation, Quercus, phyloge ´ographie, structure ge ´ne ´tique. [Traduit par la Re ´daction] Received 7 December 2006. Published on the NRC Research Press Web site at botany.nrc.ca on 29 February 2008. E. Tovar-Sa ´nchez 1,2 and K. Oyama. Centro de Investigaciones en Ecosistemas, Universidad Nacional Auto ´noma de Me ´xico, Campus Morelia, Antigua Carretera Pa ´tzcuaro No. 8701, Colonia San Jose ´ de la Huerta, Morelia, 58190, Michoaca ´n, Me ´xico. P. Mussali-Galante. Departamento de Biologı ´a Celular y Tisular, Facultad de Medicina, Universidad Nacional Auto ´noma de Me ´xico, Apartado Postal 70-275, D.F. 04510, Me ´xico. R. Esteban-Jime ´nez and D. Pin ˜ ero. Instituto de Ecologı ´a, Universidad Nacional Auto ´noma de Me ´xico, Apartado Postal 70-275, Me ´xico D.F. 04510, Me ´xico. D. Arias and O. Dorado. CEAMISH, Universidad Auto ´noma del Estado de Morelos, Av. Universidad No. 1001, colonia Chamilpa, C.P. 62209, Cuernavaca Morelos, Me ´xico. 1 Corresponding author (e-mail: [email protected]). 2 Present address: CEAMISH, Universidad Auto ´noma del Estado de Morelos, Avenida Universidad No. 1001, Colonia Chamilpa, C.P. 62209, Cuernavaca Morelos, Me ´xico. 228 Botany 86: 228–239 (2008) doi:10.1139/B07-128 # 2008 NRC Canada
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Chloroplast DNA polymorphism reveals geographic structure and introgression in the Quercus...

Chloroplast DNA polymorphism revealsgeographic structure and introgression in theQuercus crassifolia � Quercus crassipes hybridcomplex in Mexico
E. Tovar-Sanchez, P. Mussali-Galante, R. Esteban-Jimenez, D. Pinero, D.M. Arias,O. Dorado, and K. Oyama
Abstract: Quercus crassifolia H. & B. and Quercus crassipes H. & B. are two common temperate forest species in Mex-ico that form hybrids when they occur in mixed (sympatric) areas. In this study, we used chloroplast microsatellite haplo-types to infer phylogeographic patterns onto the genetic structure of populations of both parental species, thus definingprobable colonization routes throughout Mexico. Haplotype diversity in seven hybrid zones and four allopatric sites withnonmixed (allopatric) populations of both putative parental species was analyzed. To determine the expansion and coloni-zation routes for the two species, we carried out a nested clade phylogeographic analysis that would allow us to infer thehaplotype correspondence to a phylogeographical approach. In spite of sharing many common populations, we determinedthat there is a different genetic historical colonization for Q. crassipes and for Q. crassifolia. Hybrid populations had thehighest levels of genetic variation (Gv), Shannon diversity index (H), and haplotype number (nh) in comparison with non-mixed/allopatric populations of their putative parentals. Furthermore, populations of hybrid zones showed the highest val-ues of genetic differentiation FST, RST . Moreover, both species share four distinctive cpDNA haplotypes, which were mostlikely acquired by introgression through hybrids located in several populations throughout the Eje Neovolcanico area. Thislast region was confirmed to be a ‘‘hot spot’’ for oak diversity, a place in which different maternal lineages merged.
Resume : Les Quercus crassifolia H. & B. et Quercus crassipes H. & B. representent deux especes communes des foretstemperees du Mexique formant des hybrides lorsqu’elles se retrouvent melangees dans des regions sympatriques. Les au-teurs ont utilise les haplotypes des microsatellites chloroplastiques, pour en deduire les patrons phylogenetique, sur la basede la structure genetique des populations des deux especes parentes, et ainsi definir les routes probables de colonisation,pour l’ensemble du Mexique. Ils ont analyse la diversite des haplotypes dans huit zones d’hybridation et quatre sites allo-patriques supportant des populations non melangees/allopatriques, pour les deux especes parentes presumees. Afin de de-terminer l’expansion et les routes de colonisation pour les deux especes, les auteurs ont conduit une analysephylogeographique de clades groupes, qui permettrait de deduire la correspondance haplotypique, en vue d’une approchephylogeographique. En depit du partage de plusieurs populations communes, on montre qu’il y a une historique genetiquede colonisation differente pour le Q. crassipes et le Q. crassifolia. Les populations hybrides possedent les plus hauts de-gres de variation genetique (Gv), d’index de diversite de Shannon (H), et de nombre haplotypique (nh), comparativementaux populations non melangees/allopatriques, des parents presumes. De plus, les populations des zones hybrides montrentles plus grandes valeurs de differenciation genetique FST, RST. Les deux especes partagent egalement quatre haplotypescpADN, qui ont ete probablement acquis par introgression, via des hybrides situes dans plusieurs populations de la regionEje Neovolcanico. On a deja confirme que cette region constitue un ‘point chaud’ pour la diversite des chenes, un endroitou differentes lignees maternelles se sont confondues.
Mots-cles : cpSSRs, hybridation, Quercus, phylogeographie, structure genetique.
[Traduit par la Redaction]
Received 7 December 2006. Published on the NRC Research Press Web site at botany.nrc.ca on 29 February 2008.
E. Tovar-Sanchez1,2 and K. Oyama. Centro de Investigaciones en Ecosistemas, Universidad Nacional Autonoma de Mexico, CampusMorelia, Antigua Carretera Patzcuaro No. 8701, Colonia San Jose de la Huerta, Morelia, 58190, Michoacan, Mexico.P. Mussali-Galante. Departamento de Biologıa Celular y Tisular, Facultad de Medicina, Universidad Nacional Autonoma de Mexico,Apartado Postal 70-275, D.F. 04510, Mexico.R. Esteban-Jimenez and D. Pinero. Instituto de Ecologıa, Universidad Nacional Autonoma de Mexico, Apartado Postal 70-275, MexicoD.F. 04510, Mexico.D. Arias and O. Dorado. CEAMISH, Universidad Autonoma del Estado de Morelos, Av. Universidad No. 1001, colonia Chamilpa, C.P.62209, Cuernavaca Morelos, Mexico.
1Corresponding author (e-mail: [email protected]).2Present address: CEAMISH, Universidad Autonoma del Estado de Morelos, Avenida Universidad No. 1001, Colonia Chamilpa, C.P.62209, Cuernavaca Morelos, Mexico.
228
Botany 86: 228–239 (2008) doi:10.1139/B07-128 # 2008 NRC Canada

Introduction
The analysis of spatial subdivision within and amongstspecies at geographical zones of hybridization may providea unique opportunity to understand the mechanisms of re-productive isolation and speciation in nature (Hewitt 1989).Insights into the process, as well as into the spatial patternof natural hybridization, have been greatly developed by theapplication of cline theory (Rieseberg et al. 1999; Barton2001; Brumfield et al. 2001). Character distributions acrosshybrid zones are often geographically coincidental in clineshape and width, suggesting a balance between dispersaland selection within the zone (Haldane 1948; Barton andHewitt 1985).
Nevertheless, detection and analysis of hybridization andintrogression in mixed (sympatric) populations may be prob-lematic. In particular, when extensive introgressive hybrid-ization occurs it is difficult to distinguish from incompletesorting of ancient polymorphisms as an explanation of non-concordant character sets across related taxa (Harrison andBogdanowicz 1997; Goodman et al. 1999; Hare et al.2002). Furthermore, the distribution of potentially hybridiz-ing taxa may be quite fluid in time, which may result inevolutionarily layered patterns of hybridization that obscuredifferentiation between historical and contemporary charac-ter introgression (Arntzen and Wallis 1991). In such instan-ces, historical biogeography and phylogeographic analysismay provide the means to differentiate introgression fromretention of ancient polymorphisms. Nested clade phylogeo-graphic analysis (Templeton and Sing 1993; Templeton etal. 1995) has been useful for establishing statistical associa-tions of both observed phenotypes and geography withgenealogical structure. In general, nested clade phylogeo-graphic analysis has been used to evaluate the cohesion ofevolutionary lineages (Wiens and Penkrot 2002), even whenhybridization is among the potential population processescontributing to the geographic structuring of genetic varia-tion (Templeton 1994; Matos and Schaal 2000).
Cytoplasmic genomes (chloroplast and mitochondrial) canassess the genetic variation and historical gene flow follow-ing hybridization because of their uniparental and asexualnature of inheritance (Whittemore and Schaal 1991). In ad-dition, they show high rates of introgression amongst plantspecies (Rieseberg and Soltis 1991). In oaks, as in mostangiosperm plants, cpDNA is maternally inherited. There-fore, it is only transmitted by seeds (Dumolin-Lapegue etal. 1995). This makes chloroplast-based markers more suitedfor the study of seed dispersal and the geographic structureof genetic diversity (Petit et al. 1997) than nuclear markers.Recent studies have revealed that intraspecific geographicalstructuring of cpDNA in oaks and their related haplotypesoften have similar distribution (Dumolin-Lapegue et al.1997). Furthermore, the extensive sharing of a cytoplasmicvariant among species has a greater relation to the geograph-ical location than to the taxonomic composition of popula-tions (Petit et al. 2002).
In Quercus L. (the oaks; Fagaceae), reproductive isolationamongst closely related species is generally weak (Hardin1975). Species-independent and population-dependent distri-bution of haplotypes have been shown in mixed commun-ities of Quercus species in the white oak group
(Whittemore and Schaal 1991; Petit et al. 1993). Also, asimilar pattern was found in an extensive survey of haplo-type distribution in eight species of European white oaks:significant genetic overlap among the most common specieswas more closely related to geographical location than totaxonomic composition of populations (Petit et al. 2002).
Hybridization phenomena in the genus Quercus is particu-larly important because it is one of the most controversialexamples of the convenience of the ‘‘biological species con-cept’’ (Muir et al. 2001). Oaks have a high frequency of hy-bridization in nature (Palmer 1948; Muller 1952; Tucker1961; Cottam et al. 1982; Jensen et al. 1993; Spellenberg1995; Howard et al. 1997; Ishida et al. 2003) and this is be-lieved to be related to a poor development of sterility bar-riers between Quercus species (Grant 1981; Whittemore andSchaal 1991; Rushton 1993; Bacilieri et al. 1995; Dumolin-Lapegue et al. 1997). This plant group shows complex pat-terns of variation, leading to problems in species identifica-tion (Burger 1975).
The genus Quercus includes over 500 species (Govaertsand Frodin 1998) distributed mainly across the warmer re-gions of the northern hemisphere. Mexico possesses 161species, many of which are endemic (Valencia 2004). Oakforests cover about 4.29% (84 622 km2) of Mexican territory(Flores-Villela and Gerez 1994). Fossil records of Quercus(Daghlian and Crepet 1983) suggest that North Americanflora spread into Mexico during the Oligocene (approxi-mately 35 million years ago), throughout the Sierra MadreOriental, coming from the Appalachian mountains (Martinand Harrell 1957). It is possible that after the migration ofQuercus to Mexico, this group colonized the soils resultingfrom intensive volcanic activity; a characteristic of manyMexican mountains (Miocene) (Graham 1993). This lastscenario may have facilitated the contact between speciesthat were isolated in the Sierra Madre Oriental (SMOr) andin the Sierra Madre Occidental (SMOc), in the mountains ofthe Eje Neovolcanico. The Eje Neovolcanico is the highestmountain range in Mexico (Rzedowski 1978). It is recog-nized as an important center of taxonomic diversity. Nixon(1993) considers it to be a center of diversity for Quercus,as it is for the family Asteraceae, with 370 endemic species(Turner and Nesom 1993). Styles (1993) proposed that sincethese mountains connect the biotas of the SMOc and theSMOr, they are crucial to the evolutionary history of thegenera Pinus and Quercus. The high levels of volcanic ac-tivity in the area gave rise to many microhabitats that al-lowed radiation and speciation of taxa (Challenger 1998),contributing to the high species diversity that is characteris-tic of this region.
Oaks represent a good model for phylogeographical stud-ies in Mexican forests. They represent a key element for oldgrowth forests in the Eje Neovolcanico, richest in its up-lands, since the Miocene (Morley 2001). Oaks are, therefore,present in mixed species communities throughout their geo-graphical distribution range (Aiba and Kitayama 1999; Can-non 2001).
Chloroplast SSRs have been used in many species ofQuercus to study diversity, chloroplast lineages, and theevolutionary history of species (Petit et al. 1993; Bacilieriet al. 1996; Dumolin-Lapegue et al. 1998, 1999; Petit et al.2002).
Tovar-Sanchez et al. 229
# 2008 NRC Canada

In this study, we investigated the genetic variation ofcpDNA, phylogeography, and the interplay between contem-porary versus historical processes, such as gene flow andhaplotype dispersal in the hybrid complex formed by Quer-cus crassifolia H. & B., Quercus crassipes H. & B., andtheir hybrids. This interfertile species pair shows extensiveecological overlap, forming hybrid zones over areas of sym-patry at Eje Neovolcanico and in the southern part of SierraMadre Oriental in central Mexico. Thus, they remain distinctdespite occasional hybridization (Tovar-Sanchez and Oyama2004). However, sometimes sympatric areas do not registerhybrid individuals. In these particular zones the hybrids arerare (between 10 and 17 trees), and they are in a narrowzone amongst well-differentiated taxa (Tovar-Sanchez andOyama 2004).
Phylogenetic relationships estimated from cpDNA haplo-types and a priori taxonomic hypotheses are comparedwithin and between Q. crassifolia and Q. crassipes to assessthe degree of concordance between these data. The permuta-tion tests inherent to nested clade phylogeographic analysisare used to examine the association of haplotype structurewith geography. Phylogeographic patterns of nonconcord-ance are then evaluated within the context of regional bio-geographic history to differentiate, when possible, between
incomplete sorting of ancient polymorphisms and contempo-rary and (or) historical introgressive hybridization. The fol-lowing questions are addressed in this study. (i) What arethe levels of genetic variation in chloroplast types acrossboth the geographical and taxonomic range of both species?(ii) Is the variation geographically structured? (iii) How iscpDNA variation distributed within and among populationsof the Q. crassifolia � Q. crassipes hybrid complex? (iv)What is the extent and direction of the cytoplasmic geneflow between Q. crassifolia and Q. crassipes in the hybridzones? (v) What are the relative contributions of cpDNA tohybrids by parental species?
Materials and methods
Sample collection, DNA extraction, PCR, and cpSSRmarkers
Samples were collected from 290 individuals representing11 populations: two nonmixed (allopatric) populations ofQ. crassifolia in the Sierra Madre Occidental, two nonmixed(allopatric) populations of Q. crassipes in the Sierra MadreOriental, five mixed (sympatric) populations in the Eje Neo-volcanico and two other mixed (sympatric) populations inthe southeast part of the Sierra Madre Oriental (Fig. 1). In
Fig. 1. Map of sampled populations of Quercus crassifolia � Quercus crassipes complex. The mixed stands are represented by numbers:1, Cantera; 2, Canalejas; 3, Tlaxco; 4, Acajete; 5, Esperanza (located at the Eje Neovolcanico); 6, Agua Blanca; 7, Palo Bendito (located atthe Sierra Madre Oriental).
230 Botany Vol. 86, 2008
# 2008 NRC Canada

each mixed (sympatric) population we sampled 10 individu-als per taxa (Q. crassifolia, Q. crassipes and hybrids), and20 individuals, each nonmixed (allopatric) population. Hy-brid individuals were chosen as described by Tovar-Sanchezand Oyama (2004). Total DNA was extracted and purifiedthrough the use of a CTAB-based method, as well as lightsolvents extraction combined with anion-exchange mem-brane purification. This protocol is a modification to theDNeasy Plant Mini Kit (Qiagen, Valencia, Calif.) to removeproteins and other secondary compounds. The DNA concen-tration of each sample was obtained by fluorometric analy-sis, and the DNA quality was estimated by comparing theintensity of bands with known standards of lambda DNA onan agarose gel at 0.8%. Finally, we diluted all DNA samplesto a final concentration of 10 ng/mL.
Chloroplast microsatellites primers were obtained fromWeising and Gardner (1999). We then selected three locifor the genetic analysis (ccmp3, ccmp4, and ccmp41). Theamplification mixture for each sample contained 20 mmol/LTris-HCl, 50 mmol/L KCl, 2 mmol/L MgCl2, 0.13 mmol/Lof each dNTP, 25 mmol/L of primer, 15 ng of total DNA,and 0.8 units of Taq polymerase, in a final volume of25 mL. The polymerase chain reactions (PCR) were carriedout on a PTC-100 Programmable Thermal Controller (MJResearch Inc.) using the following program: 5 min at95 8C, followed by 30 cycles of 1 min at 94 8C, 1 min at50 8C, 30 s at 72 8C, and a final extension of 8 min at72 8C. The annealing temperature was changed for the dif-ferent microsatellite primers: 50 8C for ccmp3, 48 8C forccmp4, and 55 8C for ccmp41, and all annealing cycles re-maining 1 min. Products were resolved on 6% denaturingpolyacrilamide gels (7 mol/L urea) at 60 W constant powerfor 3 h.
Polymorphic fragments were sequenced and analyzed onan ALF automatic sequencer (Pharmacia) at 35 W constantpower for 80–90 min. External molecular weight (50, 100,150, and 250 bp) was used in conjunction with the FragmentManager (version 1.1) conversion software (Pharmacia) tosize the amplified fragments. Initial alignment was per-formed in Sequencher (GeneCodes, Ann Arbor, Mich.) andall base pair changes were verified by visually examiningthe chromatograms. As sequences were incorporated intothe overall alignment, duplicate types were discarded. Onceall sequences were incorporated, the types were exported toGeneDoc (Nicholas and Nicholas 1997) and Mathematica 4(Wolfram 1998). Visual alignment was performed, resultingin a length of 588 base pairs, and informative indels werecoded in a binary fashion. Haplotypes were defined usingARLEQUIN version 2000 (Schneider et al. 2000).
Genetic analysisWe estimated the genetic variation of Q. crassifolia,
Q. crassipes, and derived hybrids using allele frequenciesper locus in each population. Also, we registered in eachtaxon the haplotypic variation, which was estimated usingthe unbiased genetic diversity following the methods of Nei(1987), as well as the global gene diversity for the totalnumber of individuals (GT), number of haplotypes (nh), andShannon diversity index (H). Theta (�) was estimated indi-rectly based on heterozygosity using both mutational models(stepwise and infinite allele) (to an haploid marker, � = 2Nu,
were N is the population size and u is the mutation rate(Schneider et al. 2000). The analysis was performed withthe ARLEQUIN version 2000 (Schneider et al. 2000).
Population structure for Q. crassifolia, Q. crassipes andderived hybrids was estimated using an analysis of molecu-lar variance (AMOVA; Excoffier et al. 1992), to test howsequence variation is partitioned within and among popula-tions at all sites.
Genetic distance among populations was determined byusing Da, for both mutational models (stepwise and infiniteallele) (Nei et al. 1983). The statistical significance of thecorrelation between Da and geographical distance was testedwith a Mantel test (Mantel 1967).
Nested clade phylogeographyc and geographical distanceanalysis
We describe the geographic and genetic structure ofQuercus crassipes and Q. crassifolia using statistical parsi-mony coupled with nested cladistic phylogeographic analy-ses (Templeton et al. 1987; Templeton 1998). Statisticalparsimony was used to estimate cladograms for all uniquehaplotypes (TCS version 1.06; Templeton et al. 1992; Tem-pleton 1998; Clement et al. 2000). Subsequently, the haplo-types that couldn’t be parsimoniously connected (P < 0.05)were inferred via the topology of the maximum parsimonytree and grouped as sister clades at equal nesting levels(Templeton et al. 1992). The nesting methodology used wasdescribed by Templeton et al. (1987) to hierarchically grouphaplotypes until the whole cladogram was incorporated inthe nested structure; uncertain connections remained un-nested and were integrated at higher nesting levels (Tem-pleton and Sing 1993). The position of each haplotype inthe network gave us the possibility to infer the interior statusand when uncertain, we referred to outgroup weights to de-cide the relative position of haplotypes in the unrooted net-work (Castelloe and Templeton 1994).
We tested clades within nested categories against theirgeographic locality, implementing an exact permutation con-tingency analysis (Templeton et al. 1995); this kind of testindicates whether haplotypes are significantly associatedgeographically. We performed a nested geographical dis-tance analysis to test the effects of historical populationevents such as range expansions, colonization, or fragmenta-tion, and compared distances between collection locations toexpected values under the randomized null hypothesis of nogeographical association (Templeton et al. 1995). Also, weformed the null distribution with 10 000 permutations of thenesting clades versus sampling locations (P < 0.05). Bothtests of geographical association were implemented (GeoDisversion 2.0; Posada et al. 2000), and to interpret our resultswe used the Templeton inference key (1998). Lastly, we es-timated gene and haplotype diversity indices (Arlequin ver-sion 2.0; Schneider et al. 2000) to know genetic predictionsof the clades for which we inferred range expansion in thenested clade phylogeographic analysis.
Results
Genetic variationThe three (ccmp3, ccmp4, and ccmp41) cpSSR loci were
polymorphic in 290 individuals of Q. crassifolia,
Tovar-Sanchez et al. 231
# 2008 NRC Canada
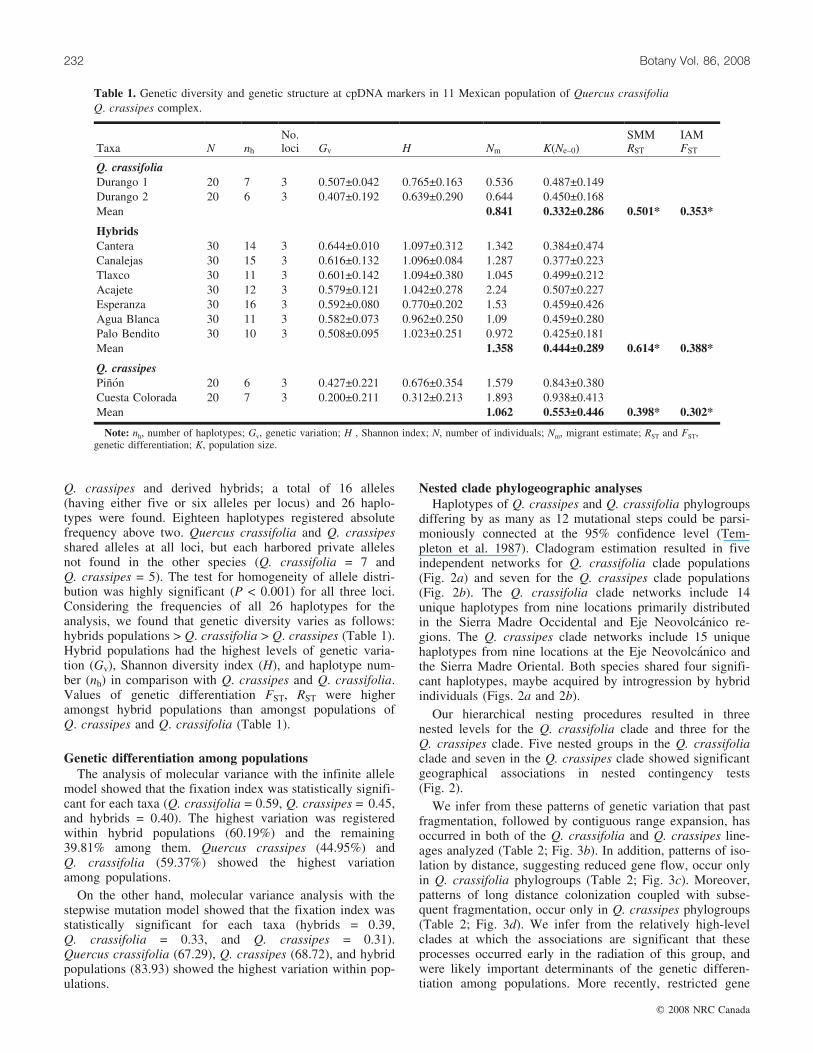
Q. crassipes and derived hybrids; a total of 16 alleles(having either five or six alleles per locus) and 26 haplo-types were found. Eighteen haplotypes registered absolutefrequency above two. Quercus crassifolia and Q. crassipesshared alleles at all loci, but each harbored private allelesnot found in the other species (Q. crassifolia = 7 andQ. crassipes = 5). The test for homogeneity of allele distri-bution was highly significant (P < 0.001) for all three loci.Considering the frequencies of all 26 haplotypes for theanalysis, we found that genetic diversity varies as follows:hybrids populations > Q. crassifolia > Q. crassipes (Table 1).Hybrid populations had the highest levels of genetic varia-tion (Gv), Shannon diversity index (H), and haplotype num-ber (nh) in comparison with Q. crassipes and Q. crassifolia.Values of genetic differentiation FST, RST were higheramongst hybrid populations than amongst populations ofQ. crassipes and Q. crassifolia (Table 1).
Genetic differentiation among populationsThe analysis of molecular variance with the infinite allele
model showed that the fixation index was statistically signifi-cant for each taxa (Q. crassifolia = 0.59, Q. crassipes = 0.45,and hybrids = 0.40). The highest variation was registeredwithin hybrid populations (60.19%) and the remaining39.81% among them. Quercus crassipes (44.95%) andQ. crassifolia (59.37%) showed the highest variationamong populations.
On the other hand, molecular variance analysis with thestepwise mutation model showed that the fixation index wasstatistically significant for each taxa (hybrids = 0.39,Q. crassifolia = 0.33, and Q. crassipes = 0.31).Quercus crassifolia (67.29), Q. crassipes (68.72), and hybridpopulations (83.93) showed the highest variation within pop-ulations.
Nested clade phylogeographic analysesHaplotypes of Q. crassipes and Q. crassifolia phylogroups
differing by as many as 12 mutational steps could be parsi-moniously connected at the 95% confidence level (Tem-pleton et al. 1987). Cladogram estimation resulted in fiveindependent networks for Q. crassifolia clade populations(Fig. 2a) and seven for the Q. crassipes clade populations(Fig. 2b). The Q. crassifolia clade networks include 14unique haplotypes from nine locations primarily distributedin the Sierra Madre Occidental and Eje Neovolcanico re-gions. The Q. crassipes clade networks include 15 uniquehaplotypes from nine locations at the Eje Neovolcanico andthe Sierra Madre Oriental. Both species shared four signifi-cant haplotypes, maybe acquired by introgression by hybridindividuals (Figs. 2a and 2b).
Our hierarchical nesting procedures resulted in threenested levels for the Q. crassifolia clade and three for theQ. crassipes clade. Five nested groups in the Q. crassifoliaclade and seven in the Q. crassipes clade showed significantgeographical associations in nested contingency tests(Fig. 2).
We infer from these patterns of genetic variation that pastfragmentation, followed by contiguous range expansion, hasoccurred in both of the Q. crassifolia and Q. crassipes line-ages analyzed (Table 2; Fig. 3b). In addition, patterns of iso-lation by distance, suggesting reduced gene flow, occur onlyin Q. crassifolia phylogroups (Table 2; Fig. 3c). Moreover,patterns of long distance colonization coupled with subse-quent fragmentation, occur only in Q. crassipes phylogroups(Table 2; Fig. 3d). We infer from the relatively high-levelclades at which the associations are significant that theseprocesses occurred early in the radiation of this group, andwere likely important determinants of the genetic differen-tiation among populations. More recently, restricted gene
Table 1. Genetic diversity and genetic structure at cpDNA markers in 11 Mexican population of Quercus crassifolia �Q. crassipes complex.
Taxa N nh
No.loci Gv H Nm K(Ne–0)
SMMRST
IAMFST
Q. crassifoliaDurango 1 20 7 3 0.507±0.042 0.765±0.163 0.536 0.487±0.149Durango 2 20 6 3 0.407±0.192 0.639±0.290 0.644 0.450±0.168Mean 0.841 0.332±0.286 0.501* 0.353*
HybridsCantera 30 14 3 0.644±0.010 1.097±0.312 1.342 0.384±0.474Canalejas 30 15 3 0.616±0.132 1.096±0.084 1.287 0.377±0.223Tlaxco 30 11 3 0.601±0.142 1.094±0.380 1.045 0.499±0.212Acajete 30 12 3 0.579±0.121 1.042±0.278 2.24 0.507±0.227Esperanza 30 16 3 0.592±0.080 0.770±0.202 1.53 0.459±0.426Agua Blanca 30 11 3 0.582±0.073 0.962±0.250 1.09 0.459±0.280Palo Bendito 30 10 3 0.508±0.095 1.023±0.251 0.972 0.425±0.181Mean 1.358 0.444±0.289 0.614* 0.388*
Q. crassipesPinon 20 6 3 0.427±0.221 0.676±0.354 1.579 0.843±0.380Cuesta Colorada 20 7 3 0.200±0.211 0.312±0.213 1.893 0.938±0.413Mean 1.062 0.553±0.446 0.398* 0.302*
Note: nh, number of haplotypes; Gv, genetic variation; H , Shannon index; N, number of individuals; Nm, migrant estimate; RST and FST,genetic differentiation; K, population size.
232 Botany Vol. 86, 2008
# 2008 NRC Canada

flow and patterns of isolation by distance contributed topopulation structure. The significant processes detected withnested clade phylogeographic analysis and their geographi-cal locations are represented in Fig. 4.
Our haplotype networks illustrate that tip clades are morewidespread geographically than ancient haplotypes. Geneand nucleotide diversity were significantly lower in tipclades (Table 2), supporting the idea that these regions were
Fig. 2. Maximum-parsimony network and nested design for the haplotypes of cpDNA microsatellites from (a) Q. crassifolia and(b) Q. crassipes. Haplotypes are indicated by numbers; in both species three level clades are detected. Line between networks shows thatboth species share four common haplotypes, acquired possibly by introgression through hybrids bi-directionally.
Table 2. Inference chain on the results of nested geographical analyses of three chloroplast DNA data in Q. crassifolia and Q. crassipes inMexico.
Clade Chain of inference Demographic event inferred
Q. crassifolia4_2 1–2-3–5-6–13-YES Past fragmentation followed by range expansion1_1 1–2-3–4-NO Restricted gene flow with isolation by distance9_3 1–2-3–5-6–13-YES Past fragmentation followed by range expansion3_3 1–2-3–5-6–13-YES Past fragmentation followed by range expansion1_4 1–2-3–5-6–13-YES Past fragmentation followed by range expansionTotal cladogram 1–2-3–5-6–13-YES Past fragmentation followed by range expansion
Q. crassipes8_5 1–2-3–5-6–13-YES Past fragmentation followed by range expansion9_6 1–2-3–5-15-NO Past fragmentation and (or) long distance colonization9_3 1–2-3–5-6–13-YES Long distance colonization coupled with subsequent fragmentation5_4 1–2-3–5-6–13-YES Past fragmentation followed by range expansion4_1 1–2-3–5-6–13-YES Long distance colonization coupled with subsequent fragmentationTotal cladogram 1–2-3–5-6–13-YES Long distance colonization coupled with subsequent fragmentation
Tovar-Sanchez et al. 233
# 2008 NRC Canada

colonized through range expansion from ancient types inpost glacial refugia throughout the Sierra Madre Oriental,the Sierra Madre Occidental, and mainly on the Eje Neovol-canico in central Mexico.
For both mutation models (stepwise (SMM) and infiniteallele (IAM)), we found a significant association betweengenetic and geographic distances to Q. crassifolia (Mantel1967; with SMM r = 0.72; P = 0.002 and IAM r = 0.32;P = 0.04) and Q. crassipes (Mantel 1967; with SMM r =0.37; P = 0.03 and IAM r = 0.51; P = 0.003).
Discussion
Genetic structureQuercus crassifolia and Q. crassipes are two related red
oak species that have broad areas of sympatry in the EjeNeovolcanico and in the southeast part of the Sierra MadreOriental in central Mexico where they hybridize in nature(Tovar-Sanchez and Oyama 2004).
The geographic distribution of chloroplast haplotypes inQ. crassifolia and Q. crassipes indicate that in mixed (sym-patric) sites both species do not represent fully isolated genepools, but are actively exchanging chloroplast genomes. De-
spite natural hybridization, Q. crassifolia and Q. crassipesremain morphologically, genetically, and ecologically dis-tinct, as do other pairs of related Quercus species(Kleinschmit et al. 1995; Howard et al. 1997; Bruschi et al.2000; Tomlinson et al. 2000; Ishida et al. 2003). In fact,most pairs of Quercus species that remain distinct, despitehybridization, do so because they are maintained more ef-fectively by ecological divergence than by their genetic in-compatibility (Kleinschmit et al. 1995; Howard et al. 1997;Bruschi et al. 2000; Jiggins and Mallet 2000; Tomlinson etal. 2000). Furthermore, this may be explained by the ‘‘iso-lation by distance’’ phenomenon, as our data suggestthrough the significant correlation of genetic (with infiniteallele model and stepwise mutation model) and geographicaldistances to both species. However, this strong geographicalisolation is not surprising, owing to the fact that seeds areless mobile than pollen (Stanley and Linskens 1974). Forexample, Dow and Ashley (1996) studied American oaks us-ing microsatellite nuclear markers and found high levels oflong distance pollination and more limited acorn dispersal.
The estimated number of migrant chloroplast per genera-tion (Nm), suggest moderate levels of genetic interchangebetween both species (Q. crassifolia = 0.841 and Q. cras-
Fig. 3. Summary of the major inferred historical processes and events that led to the current distribution of Q. crassipes and Q. crassifoliacpDNA haplotypes. The geographic distribution of all significant haplotypes from both species are represented in (a) for all three significantevents described in (b), (c), and (d). Broken lines describe the historical processes related to Q. crassipes species, and solid lines representthe processes corresponding to Q. crassifolia species.
234 Botany Vol. 86, 2008
# 2008 NRC Canada

sipes = 1.062). The migrant estimates increase between spe-cies at the same locality (Nm = 1.358), corresponding to anNm of 2.716 from diploid nuclear locus (Birky et al. 1989),which is higher than other rates of gene flow between popu-lations within many plant species (Govindaraju 1988). Theaforementioned suggests that populations of Q. crassifoliaand Q. crassipes experience a high level of gene flow in thesame geographical area, which is very much greater than thelevel of gene flow from distant conspecific populations. Thehigh level of interspecific gene flow is reasonable, consider-ing the high characterization of morphologically recogniz-able hybrids in the field. Despite the high level ofinterspecific gene flow shown in our study, Q. crassifoliaand Q. crassipes are still differentiated.
In this study, the two estimates of genetic differentiation(FST and RST) were significantly different, both inQ. crassifolia and in Q. crassipes. Over all loci, the estimateof RST was higher than FST, indicating that the two speciesnot only differed in allele frequency distribution, but also inallele size. Such shifts likely occur over longer divergencetimes than changes in frequencies (Slatkin 1995), suggestinghistorically lower levels of introgression. This explains whythe species remain distinct despite occasional hybridization.
The highest estimate of genetic structure was seen be-tween hybrid populations (RST = 0.614 and FST = 0.388) incomparison with Q. crassipes and Q. crassifolia. Also, thepresence of unique haplotypes in hybrid populations sug-gests events of continuous introgression along various gener-ations, with mutation, of generating novel or exclusivevariants (Rieseberg 1995).
The level of differentiation (FST) among populations inQ. crassifolia (0.353) and Q. crassipes (0.302) are relativelysimilar to Q. ilex populations in Morocco (0.33, Belahbib etal. 2001), and the coefficient of differentiation was relativelylower than other North American species: 0.499 for Q. affi-nis and Q. laurina (Gonzalez-Rodrıguez et al. 2004); 0.46for Q. rubra (Magni et al. 2005); and for European oaks,0.856 for Q. petrea; 0.902 for Q. pubescens; 781 for Q. ro-bur; 0.832 for Q. frainetto; 0.946 for Q. faginea; 0.961 forQ. pyrenaica (Petit et al. 2002); and from Morocco, 0.84for Q. suber (Belahbib et al. 2001). Another comparativestudy on cytoplasmic diversity reported that the mean esti-mate of population subdivision measured for 97 plant spe-cies was GST = 0.70; in particular the GST value was 0.73for angiosperm tree species. Our results suggest that thesehybrid zones are promoting an increase in the genetic diver-sity levels (Gv) and the Shannon diversity index (H) in rela-tion to the parental species indicating that there isintrogression in the mixed (sympatric) stands (Raven 1976;Grant 1981). Hybrid populations on the Eje Neovolcanicoshow a unidirectional gradient of genetic diversity, as wellas Shannon index in an east–west direction. The Eje Neo-volcanico, an orographic system that traverses the centralpart of the country in an east–west direction, is consideredin geological terms as the youngest mountain range in Mex-ico. It is hypothesized that its formation process began dur-ing the mid-Tertiary with the western portion formationfollowed by the development of the central and eastern por-tion during the Quaternary–Pliocene (Ferrusquıa-Villafranca1993). In addition, owing to introgressiv hybridization (Rze-dowski 1978), it has been considered to be a center where
new plant species arise. Divergence values (�) suggest thatQ. crassipes is older than Q. crassifolia. However, both spe-cies appear to be older than all the hybrid populations lo-cated at the Eje Neovolcanico and at the southeast part ofthe Sierra Madre Oriental. We suggest that Q. crassifoliacould colonize from the Sierra Madre Occidental to the cen-ter of Mexico through the Eje Neovolcanico, andQ. crassipes colonization is from the Sierra Madre Orientalto the Eje Neovolcanico, where both species overlap, thus,producing hybrid zones. North American flora spread intoMexico during the Oligocene (approximately 35 millionyears ago), throughout the Sierra Madre Oriental, comingfrom the Appalachia mountains (Martin and Harrell 1957),and supported by fossil records of Quercus (Daghlian andCrepet 1983). It is possible to suggest that after the migra-tion of Quercus to Mexico, the oaks could colonize the vol-canic remains originated from an intensive volcanic activity,characteristic of many Mexican mountains (Miocene)(Graham 1993). This last scenario may have facilitated thecontact between species that were isolated in the Sierra Ma-dre Oriental and in the Sierra Madre Occidental, by the EjeNeovolcanico mountains. As a consequence, both popula-tions could have met and diversified.
Clearly, the Eje Neovolcanico was confirmed as a hotspot for diversity of Quercus, where different maternal line-ages merged while exhibiting high levels of diversity. De-spite the differences noted in the way species partitioncpDNA diversity, they extensively share the same haplo-types, even at the local scale. This finding confirms the re-sults of previous studies in Mexico between Q. affinis andQ. laurina (Gonzalez-Rodrıguez et al. 2004) and in Francebetween Q. robur and Q. petrea (Petit et al. 1997) orbetween Q. robur, Q. petrea, and Q. pubescens (Dumolin-Lapegue et al. 1999). It supports the view that introgressionwas not restricted to rare episodes during long periods ofsympatry.
Phylogeographic patternsThe results we obtained show an existence of two equally
monophyletic taxa derived from ancient populations in theSierra Madre Occidental and Sierra Madre Oriental regions.Within this context, we hypothesize that these arose by iso-lation on independent refugia (Figs. 4b, 4c, and 4d), andmixed (sympatric) stand recolonization of most populationsin the Eje Neovolcanico, thus further explaining the high in-traspecific genetic variation found on Q. crassipes andQ. crassifolia.
Within the Q. crassifolia taxa, the first two significantpatterns in our nested clade phylogeographic analysis are ge-netic signatures of contiguous range expansion from SierraMadre Occidental (haplotypes 9_3 and 1_4; populations Du-rango 1 and Durango 2), Eje Neovolcanico (populationsCantera, Canalejas, Tlaxco, Acajete, and Esperanza) and inthe southeast part of the Sierra Madre Oriental (populationsAgua Blanca and Palo Bendito). This pattern is consistentwith the Eje Neovolcanico refugia for the Q. crassifolia hap-lotype with an initial expansion into the Sierra Madre Orien-tal (haplotypes 4_2 and 3_3), followed by an extensiverange expansion southward (after glacial retreat) and north-ward along the Sierra Madre Oriental (Fig. 4b). On the otherhand, our results suggest that Q. crassipes shows fragmenta-
Tovar-Sanchez et al. 235
# 2008 NRC Canada

tion between ancient populations in the Pinon and CuestaColorada (in Sierra Madre Oriental south region) (Fig. 4b),followed by a contiguous range expansion from Sierra Ma-dre Oriental (haplotypes 8_5, 9_6, and 5_4) forward south-east part of the Sierra Madre Oriental (populations AguaBlanca and Palo Bendito) and Eje Neovolcanico (haplotypes8_5, population Esperanza; haplotype 9_6, populations Es-peranza and Acajete; haplotype 5_4, populations Tlaxco,Canalejas, and Cantera) (Fig. 4b).
Alternatively, this base divergence between interior haplo-type 4_1 could indicate isolation in independent refugia onCanalejas population of the Eje Neovolcanico. Followingthis initial divergence, we again see that tip haplotypes havelarger ranges and the genetic signature for a contiguousrange expansion southward (from haplotype 4_1) to the ex-treme southern range of this clade in the Eje Neovolcanico(Fig. 4d). Thus, the geographic patterns of initial divergence,as well as subsequent dispersal for both Q. crassipes andQ. crassifolia taxa, suggest that ancient populations divergedinto allopatry and possibly occupied up to two refugia in thecentral region of Mexico.
Throughout the Plio-Pleistocene, the Eje Neovolcanicomountain chains were dispersal corridors for many taxa, es-pecially when sea levels dropped leaving habitat availablealong the Mexican Gulf and Atlantic Coasts (Auffenbergand Milstead 1965; Swift et al. 1986).
The central highland pre-Pleistocene vicariance hypothe-sis proposed that an east–west vicariant event occurred be-tween isolated populations from the eastern and the interiorhighlands. Our data does not show an east–west geneticstructure pattern within the Q. crassifolia taxa (Fig. 4).Rather, it does show a variation within the Q. crassifoliataxa and is best explained by the central highland dispersalhypothesis that suggests that the Eje Neovolcanico popula-tions, as well as the large and undifferentiated southern
populations, are a result of contiguous range expansion(Fig. 4b).
We find that the significant geographical associations ofhaplotypes in the Eje Neovolcanico occur at relatively lownesting levels (haplotype 1_1; Table 2; Fig. 4c). Judgingfrom the relative depth of differentiation among lineagesthat emerged from the Q. crassifolia taxa refuge, coloniza-tion and differentiation of the Eje Neovolcanico populationsfrom more northwestern populations on the Sierra MadreOccidental may be as recent as the northern range expansionthat occurred after the last glaciation. Our results show con-clusively that dispersal from a refugial population in theeastern Sierra Madre Occidental populations, rather than anancient vicariant event, is the primary mechanism resultingin the close affinities of populations in the eastern and west-ern segments of the parallel main mountain chains (SierraMadre Occidental and Sierra Madre Oriental) on Mexico(Fig. 4). The Q. crassipes taxa is greatly restricted in geo-graphic distribution and is composed of two well-sup-ported population groups that resulted from a pastfragmentation event, possibly a refugial isolation (Fig. 4).First and foremost, one subclade consists of populationsthat extend northward into the Sierra Madre Oriental, re-flecting evidence of genetic isolation by distance. The sec-ond radiation resulted from westward expansion into theEje Neovolcanico region. Nested clade phylogeographicanalysis indicate that, as with the Q. crassifolia taxa, dis-persal from refugial populations was important in shapingcurrent population structure, although the interior cladealso shows evidence of deeper genetic structure owing tofragmentation.
As a result of the high mutation rate in microsatellite loci,these markers could reflect homoplasy. If this phenomenonis prevalent in a population, it would seriously weaken theconclusions drawn by phylogeographyc studies. This occurs
Fig. 4. Geographical representation of third-level clades obtained in the maximum parsimony network of the cpDNA haplotypes of (a) Q.crassifolia and (b) Q. crassipes, superimposed on a map of Mexico. There are five nested haplotypes associated to Q. crassifolia distribution(1_4, 9_3, 1_1, 4_2, and 3_3), and five associated to Q. crassipes distribution (9_6, 4_1, 5_4, 9_3, and 8_5). From these significant andnested haplotypes, only the 9_3 is fixed and shared in both species, maybe because of extensive hybridization (reported previously) andconsequent introgression in Eje Neovolcanico populations.
236 Botany Vol. 86, 2008
# 2008 NRC Canada

when different alleles are identical in size, but not identicalin descent (Estoup et al. 2002). This process is related to theway mutation produces new alleles, a reaction that is ex-pected in most mutation models, except under the infinite al-lele model. Our phylogeographic inferences could not beaccurate if homoplasy is prevalent in cpDNA micrisatelliteloci in the Q. crassipes and Q. crassifolia complex. How-ever, our inferences are reasonably supported owing to thefact that homoplasy does not occur under the infinite allelemodel used in this study. When considering haplotypes (sev-eral loci), as in chloroplast DNA, even though the degree ofhomoplasy is less, it is still significant. Estoup et al. (2002),using simulations, concluded that the large amount of varia-bility at microsatellite loci often largely compensates fortheir homoplasious evolution. The cases in which size ho-moplasy may be a problem include high mutation rates andlarge population sizes. There are no estimates of mutationrate in the Q. crassipes and Q. crassifolia complex withmononucleotide microsatellite. However, Jakobsson and Sall(2007) found a low mutation rate (10–6) in an empiricalstudy of 244 chloroplast mononucleotide microsatellites inArabidopsis thaliana. Also, Navascues and Emerson (2005),using simulations, concluded that with mutation rates lowerthan 10–4, homoplasy did not cause an underestimation, andhaplotypic diversity values for SMM and IAM did not differin all simulations.
AcknowledgementsThe authors thank Susana Valencia, Oscar Rodrıguez,
Mauricio Mora, and Maribel Paniagua for technical assis-tance. This research was supported by a PAEP-UNAM grantand a Consejo Nacional de Ciencia y Tecnologıa scholarshipto E.T.S.
ReferencesAiba, S.-I., and Kitayama, K. 1999. Composition and species diver-
sity in an altitude-substrate matrix of rain forest tree commu-nities on Mount Kinabalu, Borneo. Plant Ecol. 140: 139–157.doi:10.1023/A:1009710618040.
Arntzen, J.W., and Wallis, G.P. 1991. Restricted gene flow in amoving hybrid zone of the newts Triturus cristatus and Triturusmarmoratus in western France. Evolution, 45: 805–826. doi:10.2307/2409691.
Auffenberg, W., and Milstead, W.W. 1965. Reptiles in the Qua-ternary of North America. In The Quaternary of the UnitedStates. Edited by H.E. Wright and D.G. Frey. Princeton Univer-sity Press, Princeton, N.J. pp. 557–567.
Bacilieri, R., Ducousso, A., and Kremer, A. 1995. Genetical, mor-phological, ecological and phenological differentiation betweenQuercus petrea (Matt.) Liebl., and Quercus robur L. in a mixedstand of northwest. Silvae Genet. 44: 1–10.
Bacilieri, R., Ducousso, A., Petit, R.J., and Kremer, A. 1996. Mat-ing system and asymmetric hybridization in a mixed stand ofEuropean oaks. Evolution, 50: 900–908. doi:10.2307/2410861.
Barton, N.H. 2001. The role of hybridization in evolution. Mol.Ecol. 10: 551–568. doi:10.1046/j.1365-294x.2001.01216.x.PMID:11298968.
Barton, N.H., and Hewitt, G.M. 1985. Analysis of hybrid zones.Annu. Rev. Ecol. Syst. 16: 113–148. doi:10.1146/annurev.es.16.110185.000553.
Belahbib, N., Pemonge, M.H., Ouassou, A., Sbay, H., Kremer, A.,and Petit, R.J. 2001. Frequent cytoplasmic exchange between
oak species that are not closely related: Quercus suber andQ. ilex in Morocco. Mol. Ecol. 10: 2003–2012. doi:10.1046/j.0962-1083.2001.01330.x. PMID:11555243.
Birky, C.W., Jr., Fuerst, P., and Maruyama, T. 1989. Organellegene diversity under migration, mutation, and drift: equilibriumexpectations, approach to equilibrium, effects of heteroplasmiccells, and comparison to nuclear genes. Genetics, 121: 613–627.PMID:2714640.
Brumfield, R.T., Jernigan, R.W., McDonald, D.B., and Braun, M.J.2001. Evolutionary implications of divergent clines in an avian(Manacus: Aves) hybrid zone. Evolution, 55: 2070–2087.PMID:11761066.
Bruschi, P., Vendramin, G.G., Bussotti, F., and Grossoni, P. 2000.Morphological and molecular differentiation between Quercuspetrea (Matt.) Liebl., and Quercus pubescens Willd. (Fagaceae)in northern and central Italy. Ann. Bot. (Lond.), 85: 325–333.doi:10.1006/anbo.1999.1046.
Burger, W.C. 1975. The species concept in Quercus. Taxon, 24:45–50. doi:10.2307/1218998.
Cannon, C.H. 2001. Morphological and molecular diversity inLithocarpus (Fagaceae) of Mount Kinabalu. Sabah Parks Nat. J.4: 45–69.
Castelloe, J., and Templeton, A.R. 1994. Root probabilities forintraspecific gene trees under neutral coalescent theory. Mol.Phylogenet. Evol. 3: 102–113. doi:10.1006/mpev.1994.1013.PMID:8075830.
Challenger, A. 1998. Utilizacion y conservacion de los ecosistemasterrestres de Mexico. Pasado, presente, y futuro. Conabio,IBUNAM y Agrupacion Sierra Madre, Mexico.
Clement, M., Posada, D., and Crandall, K.A. 2000. TCS: acomputer program to estimate gene genealogies. Mol. Ecol. 9:1657–1659. doi:10.1046/j.1365-294x.2000.01020.x. PMID:11050560.
Cottam, W.P., Tucker, J.M., and Santamour, F.S. 1982. Oak hybri-dization at the University of Utah. State Arboretum of Utah, SaltLake City, Utah.
Daghlian, C.P., and Crepet, W.L. 1983. The evolutionary signifi-cance of Quercus sect. Erythrobalanus from the Oligocene ofEast Texas. Am. J. Bot. 70: 639–649. doi:10.2307/2443119.
Dow, B.D., and Ashley, M.V. 1996. Microsatellite analysis of seeddispersal and parentage of saplings in bur oak, Quercus macro-carpa. Mol. Ecol. 5: 120–132.
Dumolin-Lapegue, S., Demesure, B., and Petit, R.J. 1995. Inheri-tance of chloroplast and mitochondrial genomes in pedunculateoak investigated with an efficient PCR method. Theor. Appl.Genet. 91: 1253–1256. doi:10.1007/BF00220937.
Dumolin-Lapegue, S., Demesure, B., Fineschi, S., Le Corre, V.,and Petit, R.J. 1997. Phylogeographic structure of white oaksthroughout the European Continent. Genetics, 146: 1475–1487.PMID:9258689.
Dumolin-Lapegue, S., Pemonge, M., and Petit, R.J. 1998. Associa-tion between chloroplast and mitochondrial lineages in oaks.Mol. Biol. Evol. 15: 1321–1331. PMID:9867518.
Dumolin-Lapegue, S., Kremer, A., and Petit, R.J. 1999. Are chlor-oplast and mitochondrial DNA variation species independent inoaks? Evolution, 53: 1406–1413. doi:10.2307/2640887.
Estoup, A., Jarne, P., and Cornuet, J.M. 2002. Homoplasy and mu-tation model at microsattelite loci and their consequences forpopulation genetic analysis. Mol. Ecol. 11: 1591–1604. doi:10.1046/j.1365-294X.2002.01576.x. PMID:12207711.
Excoffier, L., Smouse, P., and Quattro, J. 1992. Analysis of mole-cular variance inferred from metric distance among DNA haplo-types: applications to human mitochondrial DNA restrictiondata. Genetics, 131: 479–491. PMID:1644282.
Tovar-Sanchez et al. 237
# 2008 NRC Canada

Ferrusquıa-Villafranca, I. 1993. Geology of Mexico: a synopsis. InBiological diversity of Mexico: origins and distribution. Editedby T.P. Ramamoorthy, R. Bye, A. Lot, and J. Fa. Oxford Uni-versity Press, New York. pp. 3–107.
Flores-Villela, O., and Gerez, P. 1994. Biodiversidad y conserva-cion en Mexico: vertebrados, vegetacion y uso de suelo. Cona-bio and UNAM, Mexico.
Gonzalez-Rodrıguez, A., Bain, J.F., Golden, J.L., and Oyama, K.2004. Chloroplast DNA variation in the Quercus affinis-Q. laurina complex in Mexico: geographic structure and asso-ciations with nuclear and morphological variation. Mol. Ecol.13: 3467–3476. doi:10.1111/j.1365-294X.2004.02344.x. PMID:15488004.
Goodman, S.J., Barton, N.H., Swanson, G., Abernethy, K., andPemberton, J.M. 1999. Introgression through rare hybridization:a genetic study of a hybrid zone between red and sika deer(genus Cervus) in Argyll, Scotland. Genetics, 152: 355–371.PMID:10224266.
Govaerts, R., and Frodin, D.G. 1998. World checklist and biblio-graphy of Fagales (Betulaceae, Corylaceae, Fagaceae and Tico-dendraceae). Royal Botanic Gardens, Kew, Surrey, UK.
Govindaraju, D.R. 1988. Relationship between dispersal ability andlevels of gene flow in plants. Oikos, 52: 31–35. doi:10.2307/3565978.
Graham, A. 1993. Historical factors and biological diversity inMexico. In Biological diversity of Mexico: origins and distribu-tions. Edited by T.P. Ramamoorthy, R. Bye, A. Lot, and J. Fa.Oxford University Press, New York. pp. 109–127.
Grant, V. 1981. Plant speciation. Columbia University Press, NewYork.
Haldane, J.B.S. 1948. The theory of a cline. J. Genet. 48: 277–284.Hardin, J.W. 1975. Hybridization and introgression in Quercus
alba. J. Arnold Arbor. Harv. Univ. 56: 336–363.Hare, M.P., Cipriano, F., and Palumbi, S.R. 2002. Genetic evidence
on the demography of speciation in allopatric dolphin species.Evolution, 56: 804–816. PMID:12038538.
Harrison, R.G., and Bogdanowicz, S.M. 1997. Patterns of variationand linkage disequilibrium in a field cricket hybrid zone. Evolu-tion, 51: 493–505. doi:10.2307/2411122.
Hewitt, G.M. 1989. The subdivision of species by hybrid zones. InSpeciation and its consequences. Edited by D. Otte andJ.A. Endler. Sinauer, Sunderland, Mass. pp. 85–110.
Howard, D.J., Preszler, R.W., Williams, J., Fenchel, S., andBoecklen, W.J. 1997. How discrete are oaks species? Insightsfrom a hybrid zone between Quercus grisea and Quercus gam-belii. Evolution, 51: 747–755. doi:10.2307/2411151.
Ishida, T.A., Hattori, K., Sato, H., and Kimura, M.T. 2003. Differ-entiation and hybridization between Quercus crispula andQ. dentata (Fagaceae): insights from morphological traits, am-plified fragment length polymorphism markers, and leaf minercomposition. Am. J. Bot. 90: 769–776.
Jakobsson, M., and Sall, T. 2007. Evolution of chloroplast mononu-cleotide microsatellites in Arabidopsis thaliana. Theor. Appl.Genet. 114: 223–235. doi:10.1007/s00122-006-0425-9. PMID:17123063.
Jensen, R.J., Hokanson, S.C., Isebrands, J.G., and Hancock, J.F.1993. Morphometric variation in oaks of the Apostle Islands inWisconsin: evidence of hybridization between Quercus rubraand Q. ellipsoidalis (Fagaceae). Am. J. Bot. 80: 1358–1366.doi:10.2307/2445721.
Jiggins, C.D., and Mallet, J. 2000. Bimodal hybrid zones andspeciation. Trends Ecol. Evol. 15: 250–255. doi:10.1016/S0169-5347(00)01873-5. PMID:10802556.
Kleinschmit, J.R., Bacilieri, G.R., Kremer, A., and Roloff, A. 1995.
Comparison of morphological and genetic traits of penduculateoak (Quercus robur (L.) and sessile oak (Quercus petrea(Matt.)) Liebl.). Silvae Genet. 44: 5–6.
Magni, C.R., Docousso, A., Caron, H., Petit, R.J., and Kremer, A.2005. Chloroplast DNA variation of Quercus rubra L. in NorthAmerica and comparison with Fagaceae. Mol. Ecol. 14: 513–524. doi:10.1111/j.1365-294X.2005.02400.x. PMID:15660942.
Mantel, N. 1967. The detection of disease clustering and ageneralized regression approach. Cancer Res. 27: 209–220.PMID:6018555.
Martin, P.S., and Harrell, B.E. 1957. The Pleistocene history oftemperate biotas in Mexico and eastern United States. Ecology,38: 468–480. doi:10.2307/1929892.
Matos, J.A., and Schaal, B.A. 2000. Chloroplast evolution in thePinus montezumae complex: a coalescent approach to hybridiza-tion. Evolution, 54: 1218–1233. PMID:11005290.
Morley, R.J. 2001. Why are there so many primitive angiospermsin the rain forests of Far East? In Faunal and floral migrationsand evolution in SE Asia–Australasia. Edited by I. Metcalfe,J.M.B. Smith, M. Morwood, and I. Davidson. Balkema Publish-ers, Lisse, Netherlands. pp. 185–200.
Muir, G., Fleming, C.C., and Schlotterer, C. 2001. Three divergentrDNA clusters predate the species divergence in Quercus pet-raea (Matt.) Liebl., and Quercus robur L. Mol. Biol. Evol. 18:112–119. PMID:11158370.
Muller, C. 1952. Ecological control of hybridization in Quercus: afactor in the mechanism of evolution. Evolution, 6: 147–161.doi:10.2307/2405620.
Navascues, M., and Emerson, B.C. 2005. Chloroplast microsatel-lites: measure of genetic diversity and the effect of homoplasy.Mol. Ecol. 14: 1333–1341. doi:10.1111/j.1365-294X.2005.02504.x. PMID:15813774.
Nei, M. 1987. Molecular evolutionary genetics. Columbia Univer-sity Press, New York.
Nei, M., Tajima, F., and Tateto, Y. 1983. Accuracy of estimatedphylogenetic tree from molecular data. II. Gene frequenciesdata. J. Mol. Evol. 19: 153–170. doi:10.1007/BF02300753.PMID:6571220.
Nicholas, K.B., and Nicholas, H.B. 1997. GeneDoc: a tool for edit-ing and annotating multiple sequence alignments. Version 2.6.
Nixon, K.C. 1993. The genus Quercus in Mexico. In Biological Di-versity of Mexico, origins and distributions. Edited byK.C. Nixon. Oxford University Press, New York. pp. 447–458.
Palmer, E.J. 1948. Hybrid oaks of North America. J. Arnold Arbor.Harv. Univ. 29: 1–48.
Petit, R.J., Kremer, A., and Wagner, D.B. 1993. Geographic struc-ture of chloroplast DNA polymorphisms in European oaks.Theor. Appl. Genet. 87: 122–128. doi:10.1007/BF00223755.
Petit, R.J., Pineau, E., Demesure, B., Bacilieri, R., Ducousso, A.,and Kremer, A. 1997. Chloroplast DNA footprint of postglacialrecolonization by oaks. Proc. Natl. Acad. Sci. U.S.A. 94: 9996–10001. doi:10.1073/pnas.94.18.9996. PMID:11038572.
Petit, R.J., Csaikl, U.M., Bordacs, S., Burg, K., Coart, E., Cottrell,J., van Dam, B., Deans, J.D., Dumolin-Lapague, S., Fineschi, S.,Finkeldey, R., Gillies, A., Glaz, I., Goicoechea, P.G., Jensen,J.S., Konig, A.O., Lowe, A.J., Madsen, S.F., Matyas, G., Munro,R.C., Olalde, M., Pemonge, M.H., Popescu, F., Slade, D.,Tabbener, H., Taurchini, D., De Vries, S.G.M., Ziegenhagen,B., and Kremer, A. 2002. Chloroplast DNA variation in Eur-opean white oaks — phylogeography and patterns of diversitybased on data from over 2600 populations. For. Ecol. Manag.156: 5–26. doi:10.1016/S0378-1127(01)00645-4.
Posada, D., Crandall, K.A., and Templeton, A.R. 2000. GeoDis: aprogram for the cladistic nested analysis of the geographical dis-
238 Botany Vol. 86, 2008
# 2008 NRC Canada

tribution of genetic haplotypes. Mol. Ecol. 9: 487–488. doi:10.1046/j.1365-294x.2000.00887.x. PMID:10736051.
Raven, P.H. 1976. Systematics and plant population biology. Syst.Bot. 1: 284–316. doi:10.2307/2418721.
Rieseberg, L.H. 1995. The role of hybridization in evolution: oldwine in new skins. Am. J. Bot. 82: 944–953. doi:10.2307/2445981.
Rieseberg, L.H., and Soltis, D.E. 1991. Phylogenetic consequencesof cytoplasmic gene flow in plants. Evol. Trends Plants, 5: 65–84.
Rieseberg, L.H., Whitton, J., and Gardner, K. 1999. Hybrid zonesand the genetic architecture of a barrier to gene flow betweentwo sunflower species. Genetics, 152: 713–727. PMID:10353912.
Rushton, B.S. 1993. Natural hybridization within the genus Quer-cus L. Ann. Sci. For. 50(Suppl.): 73–90. doi:10.1051/forest:19930707.
Rzedowski, J. 1978. Vegetacion de Mexico. Limusa, Mexico.Schneider, S., Roessli, L., and Excoffier, L. 2000. Arlequin version
2.000: a software for population genetics data analysis.Slatkin, M. 1995. A measure of population subdivision based on
microsatellite allele frequencies. Genetics, 139: 457–462.PMID:7705646.
Spellenberg, R. 1995. On the hybrid nature of Quercusbasaseachicensis (Fagaceae: Sect. Quercus). SIDA Contrib.Bot. 16: 427–437.
Stanley, R.G., and Linskens, H.F. 1974. Pollen: biology, biochem-istry and management. Springer, Berlin, Germany.
Styles, B.T. 1993. El genero Pinus: su panorama en Mexico. InDiversidad Biologica de Mexico. orıgenes y distribucion. Editedby T.P. Ramamoorthy, R. Bye, A. Lot and J. Fa. Instituto deBiologıa, UNAM, Mexico. pp. 385–408.
Swift, C.S., Gilbert, C.R., Bortone, S.A., Burgess, G.H., andYerger, R.W. 1986. Zoogeography of the freshwater fishes ofthe southeastern United States: savannah river to lake Pontchar-train. In The zoogeography of North American freshwater fishes.Edited by C.H. Hocutt and E.O. Wiley. John Wiley and Sons,New York. pp. 213–266.
Templeton, A.R. 1994. The role of molecular genetics in speciationstudies. In Molecular ecology and evolution: approaches and ap-plications. Edited by B. Schierwater, B. Streit, G.P. Wagner, andR. DeSalle. Birkhauser, Basel, Switzerland. pp. 455–477.
Templeton, A.R. 1998. Nested clade analyses of phylogeographicdata: testing hypotheses about gene flow and population history.Mol. Ecol. 7: 381–397. doi:10.1046/j.1365-294x.1998.00308.x.PMID:9627999.
Templeton, A.R., Boerwinkle, E., and Sing, C.F. 1987. A cladisticanalysis of phenotypic associations with haplotypes inferredfrom restriction endonuclease mapping. I. Basic theory and an
analysis of alcohol dehydrogenase activity in Drosophila. Genet-ics, 117: 343–351. PMID:2822535.
Templeton, A.R., Crandall, K.A., and Sing, C.F. 1992. A cladisticanalysis of phenotypic associations with haplotypes inferredfrom restriction endonuclease mapping and DNA sequence data.III. Cladogram estimation. Genetics, 132: 619–633. PMID:1385266.
Templeton, A.R., and Sing, C.F. 1993. A cladistic analysis of phe-notypic associations with haplotypes inferred from restrictionendonuclease mapping. IV. Nested analyses with cladogram un-certainty and recombination. Genetics, 134: 659–669. PMID:8100789.
Templeton, A.R., Routman, E., and Phillips, C.A. 1995. Separatingpopulation structure from population history: a cladistic analysisof the geographical distribution of mitochondrial DNA Haplo-types in the tiger salamander, Ambystoma tigrinum. Genetics,140: 767–782. PMID:7498753.
Tomlinson, P.T., Jensen, R.J., and Hancock, J.F. 2000. Do wholetree silvic characters indicate hybridization in red oak (QuercusSection Lobatae). Am. Midl. Nat. 143: 154–168. doi:10.1674/0003-0031(2000)143[0154:DWTSCI]2.0.CO;2.
Tovar-Sanchez, E., and Oyama, K. 2004. Natural hybridization andhybrid zones between Quercus crassifolia and Quercus crassipes(Fagaceae) in Mexico. Morphological and molecular evidence.Am. J. Bot. 91: 1352–1363.
Tucker, J.M. 1961. Studies in the Quercus undulate complex I. Apreliminary statement. Am. J. Bot. 48: 202–208. doi:10.2307/2439397.
Turner, B.L., and Nesom, G.L. 1993. Biogeografıa, diversidad ysituacion de peligro o amenaza de Asteraceae de Mexico. InDiversidad Biologica de Mexico: orıgenes y distribucion. Editedby T.P. Ramamoorthy, R. Bye, A. Lot, and J. Fa. Instituto deBiologıa, UNAM, Mexico. pp. 545–562.
Valencia, A.S. 2004. Diversidad del genero Quercus (Fagaceae) enMexico. Bol. Soc. Bot. Mex. 75: 33–53.
Weising, K., and Gardner, R. 1999. A set of conserved PCR pri-mers for the analysis of simple sequence repeat polymorphismsin chloroplast genomes of dicotyledonous angiosperms. Gen-ome, 42: 9–19. doi:10.1139/gen-42-1-9. PMID:10207998.
Whittemore, A.T., and Schaal, B.A. 1991. Interspecific gene flowin sympatric oaks. Proc. Natl. Acad. Sci. U.S.A. 88: 2540–2544. doi:10.1073/pnas.88.6.2540. PMID:11607170.
Wiens, J.J., and Penkrot, T.A. 2002. Delimiting species using DNAand morphological variation and discordant species limits inspiny lizards (Sceloporus). Syst. Biol. 51: 69–91. doi:10.1080/106351502753475880. PMID:11943093.
Wolfram, S. 1998. The mathematica book. Version 4.0.1. Cam-bridge University press, Cambridge, UK.
Tovar-Sanchez et al. 239
# 2008 NRC Canada






















