
Repression of Gonadotropin-Releasing Hormone (GnRH) Gene Expression by Melatonin May Involve...
10
Reproductive Neuroendocrinology Neuroendocrinology 2004;79:63–72 DOI: 10.1159/000076629 Repression of Gonadotropin-Releasing Hormone (GnRH) Gene Expression by Melatonin May Involve Transcription Factors COUP-TFI and C/EBP Beta Binding at the GnRH Enhancer Julia M.A. Gillespie a Deboleena Roy b Hong Cui a Denise D. Belsham a, b, c a Department of Physiology, b Institute for Medical Sciences, University of Toronto, Departments of c Obstetrics and Gynaecology, and Medicine, University of Toronto, and Division of Cellular and Molecular Biology, Toronto General Hospital Research Institute, University Health Network, Toronto, Ont., Canada Received: June 5, 2003 Accepted after revision: November 20, 2003 Denise D. Belsham Department of Physiology, University of Toronto Medical Sciences Building 3137A 1 King’s College Circle, Toronto, Ont., M5S 1A8 (Canada) Tel. +1 416 946 7646, Fax +1 416 978 4940, E-Mail [email protected] ABC Fax + 41 61 306 12 34 E-Mail [email protected] www.karger.com © 2004 S. Karger AG, Basel 0028–3835/04/0792–0063$21.00/0 Accessible online at: www.karger.com/nen Key Words Gonadotropin-releasing hormone W GT1–7 Neurons W Melatonin W Melatonin receptors W Transcription factors Abstract Melatonin activates membrane-bound G-protein-cou- pled receptors mt1 and MT2, but may also bind a family of orphan nuclear receptors, including ROR· and RZRß, representing another potential molecular mechanism of melatonin action. Recently, we demonstrated that mela- tonin downregulates gonadotropin-releasing hormone (GnRH) gene expression in the GT1–7 cell line, specifical- ly at the level of the neuron-specific GnRH gene enhanc- er. In this study, we have examined the region located at –1736/–1728 of the GnRH enhancer shown to be involved in the repression of GnRH by melatonin. This region includes hexameric consensus binding sites for orphan nuclear receptors, including ROR/RZR and COUP-TFI, as well as other defined consensus binding sites for AP-1 and C/EBP. Using electrophoretic mobility shift analysis (EMSA), we have demonstrated that GT1–7 nuclear pro- teins bind specifically to this region of the GnRH enhanc- er to form 4 complexes. EMSA antibody supershift anal- ysis indicates that the transcription factors COUP-TFI and C/EBP beta bind two specific complexes, but ROR·, Oct-1, Pbx-1, c-fos, or c-jun antibodies failed to produce any detectable supershifts. These results provide the first evidence that melatonin may mediate its direct neuroen- docrine control of GnRH gene expression through tran- scription factor binding at specific regions of the GnRH enhancer. Copyright © 2004 S. Karger AG, Basel Introduction The pineal hormone melatonin plays a role in repro- duction in mammals by exerting anti-gonadotropic effects at different levels of the hypothalamic-pituitary-gonadal (HPG) axis [1]. A number of in vitro studies have suggest- ed that the effect of melatonin on this axis occurs through melatonin-mediated modulation of gonadotropin-releas- ing hormone (GnRH)-induced luteinizing hormone (LH) release from gonadotrope cells of the anterior pituitary [2, 3]. Limited in vitro studies on rat brain tissue have sug- gested that melatonin mediates a diurnal modulation of GnRH secretion [4, 5]. Two specific membrane-bound G-protein coupled receptors for melatonin, mt1 [6] and MT2 [7] have been described to mediate the majority of
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Repression of Gonadotropin-Releasing Hormone (GnRH) Gene Expression by Melatonin May Involve...

Reproductive Neuroendocrinology
Neuroendocrinology 2004;79:63–72DOI: 10.1159/000076629
Repression of Gonadotropin-ReleasingHormone (GnRH) Gene Expression by MelatoninMay Involve Transcription Factors COUP-TFIand C/EBP Beta Binding at the GnRH Enhancer
Julia M.A. Gillespiea Deboleena Royb Hong Cuia Denise D. Belshama,b,c
aDepartment of Physiology, bInstitute for Medical Sciences, University of Toronto, Departments of cObstetrics andGynaecology, and Medicine, University of Toronto, and Division of Cellular and Molecular Biology, Toronto GeneralHospital Research Institute, University Health Network, Toronto, Ont., Canada
Received: June 5, 2003Accepted after revision: November 20, 2003
Denise D. BelshamDepartment of Physiology, University of TorontoMedical Sciences Building 3137A1 King’s College Circle, Toronto, Ont., M5S 1A8 (Canada)Tel. +1 416 946 7646, Fax +1 416 978 4940, E-Mail [email protected]
ABCFax + 41 61 306 12 34E-Mail [email protected]
© 2004 S. Karger AG, Basel0028–3835/04/0792–0063$21.00/0
Accessible online at:www.karger.com/nen
Key WordsGonadotropin-releasing hormone W GT1–7 Neurons W
Melatonin W Melatonin receptors W Transcription factors
AbstractMelatonin activates membrane-bound G-protein-cou-pled receptors mt1 and MT2, but may also bind a familyof orphan nuclear receptors, including ROR· and RZRß,representing another potential molecular mechanism ofmelatonin action. Recently, we demonstrated that mela-tonin downregulates gonadotropin-releasing hormone(GnRH) gene expression in the GT1–7 cell line, specifical-ly at the level of the neuron-specific GnRH gene enhanc-er. In this study, we have examined the region located at–1736/–1728 of the GnRH enhancer shown to be involvedin the repression of GnRH by melatonin. This regionincludes hexameric consensus binding sites for orphannuclear receptors, including ROR/RZR and COUP-TFI, aswell as other defined consensus binding sites for AP-1and C/EBP. Using electrophoretic mobility shift analysis(EMSA), we have demonstrated that GT1–7 nuclear pro-teins bind specifically to this region of the GnRH enhanc-er to form 4 complexes. EMSA antibody supershift anal-ysis indicates that the transcription factors COUP-TFI
and C/EBP beta bind two specific complexes, but ROR·,Oct-1, Pbx-1, c-fos, or c-jun antibodies failed to produceany detectable supershifts. These results provide the firstevidence that melatonin may mediate its direct neuroen-docrine control of GnRH gene expression through tran-scription factor binding at specific regions of the GnRHenhancer.
Copyright © 2004 S. Karger AG, Basel
Introduction
The pineal hormone melatonin plays a role in repro-duction in mammals by exerting anti-gonadotropic effectsat different levels of the hypothalamic-pituitary-gonadal(HPG) axis [1]. A number of in vitro studies have suggest-ed that the effect of melatonin on this axis occurs throughmelatonin-mediated modulation of gonadotropin-releas-ing hormone (GnRH)-induced luteinizing hormone (LH)release from gonadotrope cells of the anterior pituitary [2,3]. Limited in vitro studies on rat brain tissue have sug-gested that melatonin mediates a diurnal modulation ofGnRH secretion [4, 5]. Two specific membrane-boundG-protein coupled receptors for melatonin, mt1 [6] andMT2 [7] have been described to mediate the majority of

64 Neuroendocrinology 2004;79:63–72 Gillespie/Roy/Cui/Belsham
melatonin actions. Two orphan nuclear receptors, ROR·and RZRß, both members of the ROR/RZR retinoid-related receptor family that bind ROR responsive ele-ments (ROREs) to alter transcription, have also beenshown to bind melatonin with high affinity [8], therebysuggesting a possible genomic action of melatonin. More-over, there is some evidence that suggests that chickenovalbumin upstream transcription factor (COUP-TF) ho-modimers bind to monomeric ROREs, competing withother bound nuclear receptors to negatively regulate geneexpression [9].
GnRH is the prevailing hormone of the HPG axis andis secreted in a pulsatile manner from the GnRH neuronsof the hypothalamus to control LH and follicle-stimulat-ing hormone (FSH) synthesis and secretion by the anteri-or pituitary. By targeted tumorigenesis, where the SV-40T-antigen (T-Ag) was driven by the rat GnRH 5) regulato-ry region, the murine immortalized GnRH-secretingGT1–7 cell line was generated [10]. This cell model hasbeen used extensively to study cell and molecular analysisof in vitro GnRH neurons. Using the GT1–7 cells the bas-al promoter region and a complex neuron-specific en-hancer within the 5) regulatory region of the GnRH genehave been characterized [11, 12].
Our previous studies have demonstrated that melato-nin downregulates GnRH gene expression in a 24-hourcyclic pattern in GT1–7 cells [13]. Furthermore, melato-nin appears to mediate its effects on GnRH mRNAexpression at the transcriptional level as T-Ag mRNAexpression, which is driven by the rat GnRH 5) regulatoryregion in the GT1–7 cells, mimics that of GnRH follow-ing melatonin treatment [13]. The effect of melatonin onGnRH gene transcription has been localized to 5 specificregions within the well-characterized 300-bp neuron-spe-cific enhancer of the GnRH gene [13]. This indicates thatthe molecular mechanisms responsible for the repressionof GnRH by melatonin are likely due to a complex inter-action of transcription factors at the level of the enhancer.Three of the mapped regions contain binding sites forknown transcription factors, such as Oct-1 [14] andGATA [15], that have previously been linked to basalGnRH gene expression. However, the –1736 to –1723region of the GnRH enhancer was found to contain puta-tive ROREs, therefore was a particularly interesting can-didate for further analysis, since the ROR/RZR orphanreceptors have been postulated to bind melatonin. Fur-ther analysis of the –1736/–1718 sequence reveals con-sensus binding sites for C/EBP and AP-1, which bindsimmediate early gene protein products, such as c-fos andjun family members.
In this study, we have analyzed the –1739/–1718region of the GnRH enhancer and its transcription factorbinding. Herein, we have also shown that multiple GT1–7nuclear proteins bind the –1739 to –1718 region of theGnRH enhancer. Antibody supershift assays indicate thatthe transcription factors COUP-TFI and C/EBP beta canbind to this region. This study is the first to characterizethe downstream transcriptional effects of melatonin tospecific regions within the GnRH enhancer.
Materials and Methods
Cell Culture and ReagentsGT1–7 cells were grown in Dulbecco’s modified Eagle medium
(DMEM) (Gibco-BRL, Burlington, Ont.) supplemented with 10%fetal bovine serum (FBS) (Hyclone Laboratories, Inc., Logan Utah,USA), 4.5 mg/ml glucose and penicillin/streptomycin and main-tained at 37 °C in 5% CO2 as described in [10]. GT1–7 cells weregrown to 80% confluence and treated with 10 nM melatonin for 2 or4 h, without a media change. GT1–7 cells were generally used at pas-sage 16–24. After treatment, the cells were washed and harvested inphosphate-buffered saline (PBS), then pelleted before protein isola-tion or nuclear extract preparation. Melatonin was obtained fromSigma-Aldrich Canada, Ltd. (Oakville, Ont.). The melatonin stocksolutions were prepared in dimethylsulfoxide (DMSO). Pierce BCAProtein Assay Reagent kit was obtained from Pierce Ltd. (Rockford,Ill., USA). Oligonucleotides were prepared by ACGT Corp. (Toron-to, Ont.). COUP-TF I, C/EBP beta, Oct-1, Pbx-1, ROR·, c-fos andc-jun antibodies and IgG were obtained from Santa Cruz Biotechnol-ogy, Inc. (Santa Cruz, Calif., USA).
Electrophoretic Mobility Shift AssaysNuclear extracts were prepared following the method of Lee et al.
[16]. Nuclear protein concentration was determined using the PierceBCA Protein Assay Reagent Kit and samples were stored at –80 °C.EMSA and supershift assays were performed as previously described[14], except that the EMSA reaction mixes were incubated for 15 minat room temperature and then another 20 min at room temperatureafter the addition of 20,000 cpm of the labeled oligonucleotides. Oli-gonucleotides were end labeled by incubation of T4 polynucleotidekinase with [Á-32P]dATP (6,000 Ci/mmol; Perkin Elmer Canada,Woodbridge, Ont.). All labeled probes were purified by electrophore-sis on a 6% polyacrylamide gel. Competition studies were done byadding a 10-fold excess amount of unlabeled oligonucleotide to thereaction mix 15 min before adding the labeled oligonucleotide. In theantibody supershift assays, 2 Ìl of IgG or COUP-TF I (sc-6575X),C/EBP beta (sc-746X), Oct-1 (sc-232X), c-fos (sc-7202), c-jun (sc-44X), Pbx-1 (sc-888X) or ROR· antibodies (sc-6062X) (Santa CruzBiotechnology, Inc.) were added to the reaction mixture and incu-bated at room temperature for 60 min, before adding 20,000 cpm ofprobe for 20 min. The reactions were electrophoresed on a 20-minprerun polyacrylamide gel in 0.25! modified TBE for 3.5 h at 155–175 V. After electrophoresis, the gels were dried and exposed toKodak X-OMAT film for autoradiography.

Analysis of GnRH Repression by Melatonin Neuroendocrinology 2004;79:63–72 65
Western Blot AnalysisTotal protein was isolated from GT1–7 cells by lysis in PBS sup-
plemented with 1% Triton X-100 (Fisher Scientific, Whitby, Ont.)and a 1:100 dilution of Protease Inhibitor Cocktail (Sigma-AldrichCanada, Ltd.). Protein content was measured using Pierce BCA Pro-tein Assay Reagent (Pierce Ltd.). Total protein (20–60 Ìg) fromGT1–7 cells, and mouse ovary was resolved on a 12.5% SDS-poly-acrylamide gel [17] and electrotransferred to Hybond C membranesusing standard procedures (Amersham Life Sciences Inc., Oakville,Ont.). Membranes were incubated in PBST (PBS, pH 7.4, 0.2%Tween-20) with 5% powdered skim milk for 16 h at 4°C with a1:1,000 dilution of polyclonal ROR· or COUP-TFI antibody. Immu-noreactive bands were visualized with peroxidase-labeled secondarygoat antisera at 1:20,000 dilution and enhanced chemiluminescence(ECL; Amersham Life Sciences Inc.), as described by the manufac-turer. Loading and transfer conditions were monitored by reprobingthe blots with an anti-Gß (1:1,000) antibody from Santa Cruz Bio-technology. Densitometry was performed on blots exposed to KodakX-OMAT film using a UMAX Astra 1220 Scanner and the NIHImage software.
Statistical AnalysisComparisons of results between treatments over different times,
were done using one-way analysis of variance (ANOVA). The statisti-cal significance of the results between individual pairs was deter-mined using the Tukey’s multiple comparison test. Data were ana-lyzed using the statistical program package SPSS 10.0 for Windows(University of Toronto site license).
Results
GT1–7 Nuclear Proteins Specifically Bind the –1736to –1728 Region of the GnRH EnhancerIt has previously been reported that the melatonin-
mediated repression of GnRH gene expression can bemapped to 5 specific regions within the GnRH enhancer[13]. These 9-bp regions were elucidated by transfectionanalysis of specific block mutants within the GnRH geneenhancer followed by melatonin treatment [13]. Most ofthe regions contain binding sites for known transcriptionfactors, such as Oct-1 [14] and GATA [15], and have pre-viously been linked to basal GnRH gene expression. How-ever, the –1736 to –1723 region of the GnRH enhancerwas found to contain putative ROREs and therefore aparticularly interesting candidate for further analysis,since the ROR/RZR orphan receptors have been postu-lated to bind melatonin. This region contains several tran-scription factor binding sites, as determined by analysiswith a transcription factor search program (MatchTM,www.gene-regulation.com, BIOBASE Biological Data-bases). Of note, the region also has 100% homology withan AP-1 consensus binding sequence, as well as a GATAconsensus binding region, and a C/EBP binding site. Oli-
Fig. 1. Oligonucleotide sequences used for electrophoretic mobilityshift assays. The sequence of the GnRH enhancer from –1728to –1736 is within a footprinted region [11], and has been linkedto melatonin-mediated repression of GnRH gene expression [13].A The wild-type 22-bp oligonucleotide –1739/–1718, spanning thesequences –1739 to –1718 of the 300 bp GnRH enhancer, was usedas a probe in EMSA. The consensus transcription factor binding se-quences, ROR/RZR nuclear receptor family (RORE; A/GGGTCA),C/EBP (TGAAG/ACA), and AP-1 (ACTNAG), are shown. B Theblock mutant is the sequence linked to melatonin-mediated repres-sion of GnRH mRNA by transient transfections. C An AP-1 consen-sus sequence. D The corresponding AP-1 mutant oligonucleotide.Mutations are shown in bold print.
gonucleotides designed for this region were analyzed forprotein binding (fig. 1). Electrophoretic mobility shift as-says (EMSAs) were performed using nuclear extracts pre-pared from GT1–7 cells treated with melatonin (10 nM,2 or 4 h) or with vehicle alone (control) (fig. 2A). EMSAsdemonstrate that GT1–7 nuclear proteins specificallybind to this region of the enhancer to form four DNA-protein complexes (fig. 2A, complexes 1, 2, 3 and 4; start-ing with the complex with the slowest mobility). Slightdifferences in migration and band appearance were notunusual between individual EMSA gels, depending upon
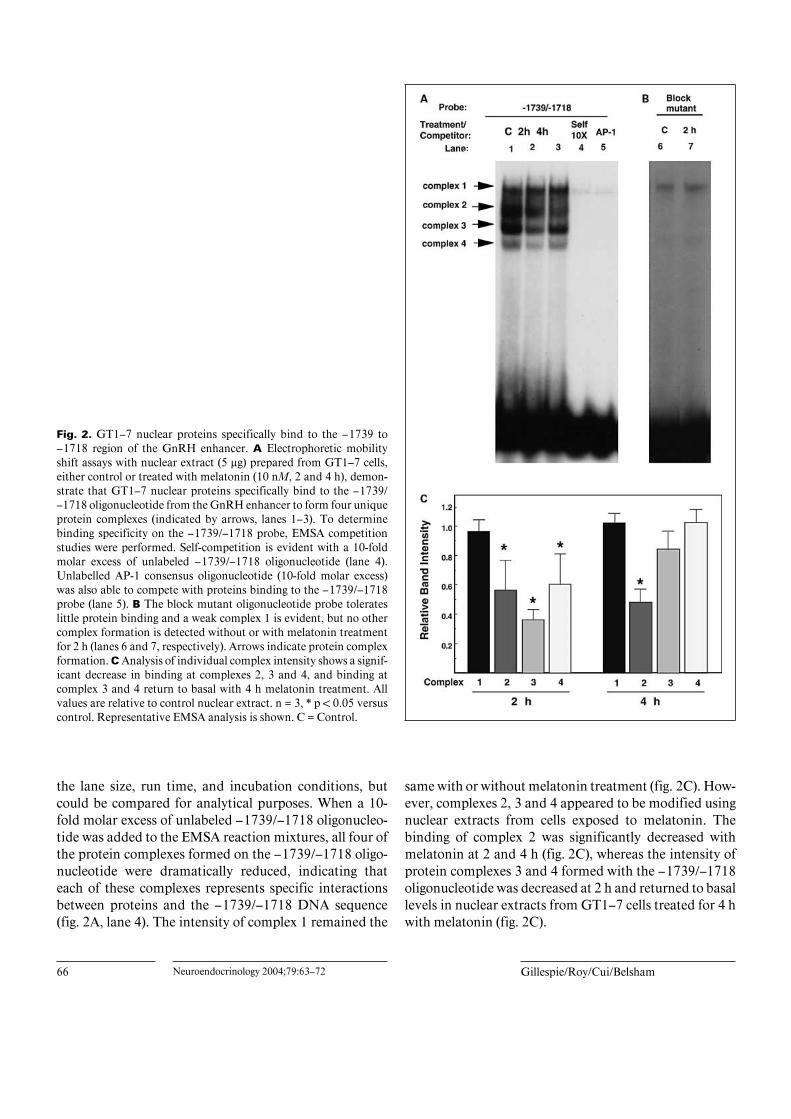
66 Neuroendocrinology 2004;79:63–72 Gillespie/Roy/Cui/Belsham
Fig. 2. GT1–7 nuclear proteins specifically bind to the –1739 to–1718 region of the GnRH enhancer. A Electrophoretic mobilityshift assays with nuclear extract (5 Ìg) prepared from GT1–7 cells,either control or treated with melatonin (10 nM, 2 and 4 h), demon-strate that GT1–7 nuclear proteins specifically bind to the –1739/–1718 oligonucleotide from the GnRH enhancer to form four uniqueprotein complexes (indicated by arrows, lanes 1–3). To determinebinding specificity on the –1739/–1718 probe, EMSA competitionstudies were performed. Self-competition is evident with a 10-foldmolar excess of unlabeled –1739/–1718 oligonucleotide (lane 4).Unlabelled AP-1 consensus oligonucleotide (10-fold molar excess)was also able to compete with proteins binding to the –1739/–1718probe (lane 5). B The block mutant oligonucleotide probe tolerateslittle protein binding and a weak complex 1 is evident, but no othercomplex formation is detected without or with melatonin treatmentfor 2 h (lanes 6 and 7, respectively). Arrows indicate protein complexformation. C Analysis of individual complex intensity shows a signif-icant decrease in binding at complexes 2, 3 and 4, and binding atcomplex 3 and 4 return to basal with 4 h melatonin treatment. Allvalues are relative to control nuclear extract. n = 3, * p ! 0.05 versuscontrol. Representative EMSA analysis is shown. C = Control.
the lane size, run time, and incubation conditions, butcould be compared for analytical purposes. When a 10-fold molar excess of unlabeled –1739/–1718 oligonucleo-tide was added to the EMSA reaction mixtures, all four ofthe protein complexes formed on the –1739/–1718 oligo-nucleotide were dramatically reduced, indicating thateach of these complexes represents specific interactionsbetween proteins and the –1739/–1718 DNA sequence(fig. 2A, lane 4). The intensity of complex 1 remained the
same with or without melatonin treatment (fig. 2C). How-ever, complexes 2, 3 and 4 appeared to be modified usingnuclear extracts from cells exposed to melatonin. Thebinding of complex 2 was significantly decreased withmelatonin at 2 and 4 h (fig. 2C), whereas the intensity ofprotein complexes 3 and 4 formed with the –1739/–1718oligonucleotide was decreased at 2 h and returned to basallevels in nuclear extracts from GT1–7 cells treated for 4 hwith melatonin (fig. 2C).

Analysis of GnRH Repression by Melatonin Neuroendocrinology 2004;79:63–72 67
An oligonucleotide containing the 9-bp block mutantat the –1739/–1718 site found to abolish melatoninrepression in transient transfection analysis [13] was alsoused for EMSA analysis (fig. 1). The 9-bp mutation abol-ished the RORE and C/EBP sites, but not the AP-1 site.When using the block mutant oligonucleotide as a probe,complex 1 is diminished, while complexes 2, 3 and 4 wereabsent (fig. 2B) when compared to the –1739/–1718probe. This suggests that the proteins bound to this regionmay be important for regulation of GnRH transcriptionby melatonin.
Since the –1739/–1718 region also contains an AP-1consensus binding site, we used a commercially availableAP-1 oligonucleotide to analyze the role, if any, of theAP-1 protein complex binding the GnRH enhancer. Un-labeled AP-1 oligonucleotide was able to compete withcomplexes formed on the –1739/–1718 probe (fig. 2A,lane 5). The AP-1 oligonucleotide was used as a probewith GT1–7 nuclear extract. GT1–7 nuclear proteins alsospecifically bind to an AP-1 consensus sequence produc-ing complexes that migrate at the same level as complex 1,and less frequently complex 2, on the –1739/–1718 oligo-nucleotide probe (fig. 3, lanes 1– 4). As expected, the unla-beled AP-1 consensus oligonucleotide was able to self-compete the AP-1 probe (fig. 3, lane 5). Consistent withfigure 2A, unlabeled –1739/–1718 oligonucleotide wasable to compete with the complexes formed by the AP-1consensus sequence probe (fig. 3, lane 7). An oligonucleo-tide with an AP-1 site mutation was used to prove thenecessity of the AP-1 site to complex 1 protein formation.However, quite unexpectedly, unlabeled AP-1 mutantoligonucleotides were also able to compete with the pro-teins formed on the AP-1 probe (fig. 3, lane 6), suggestingthat the AP-1 site is not involved in the complex 1 forma-tion, but instead other region(s) within the oligonucleo-tide were responsible for the EMSA complex formation.
Transcription Factors C/EBP Beta and COUP-TFIInteract with the –1736 to –1728 Sequence on theGnRH EnhancerAs mentioned, the –1739/–1718 oligonucleotide se-
quence contains a number of consensus transcription fac-tor binding sites, including RORE, C/EBP, and AP-1(fig. 1). Unexpectedly, the AP-1 consensus oligonucleo-tide also harbors a C/EBP site, besides the AP-1 site,whereas the AP-1 mutant oligonucleotide only eradicatesthe AP-1 site, not the C/EBP site. The block mutant usedin the original transient transfection analysis [13] is miss-ing the C/EBP and RORE sites, but maintains the AP-1site. This analysis indicates that the C/EBP and RORE
Fig. 3. An AP-1 consensus oligonucleotide is able to bind GT1–7nuclear proteins, but apparently not at the AP-1 binding site. GT1–7cells were treated with 10 nM melatonin for 4 h, and nuclear extractswere prepared. EMSA analysis was performed with 5 Ìg of GT1–7nuclear extract per reaction. An AP-1 consensus oligonucleotide wasused as a probe to determine protein binding. A DNA-protein com-plex is seen migrating at what appears to be the same level as com-plexes 1 and 2 with the –1739/–1718 probe (lanes 1 and 4). Bindingdecreases upon incubation with the nuclear extracts from cellstreated with melatonin (lanes 2 and 3). Unlabelled AP-1 consensusoligonucleotide (10-fold molar excess) was able to compete with pro-teins binding to the –1739/–1718 probe (lane 5), whereas conversely,unlabeled –1739/–1718 oligonucleotide was able to compete for pro-teins binding the AP-1 probe (lane 7). Unexpectedly, the AP-1mutant was able to compete with proteins formed on the AP-1 probe(lane 6). This illustrates that protein binding was specific, and sug-gests that there may be a common transcription factor binding site onboth the –1739/–1718 and AP-1 consensus oligonucleotide se-quences, but not likely the AP-1 site itself. Representative EMSAsare shown, n = 3.
sites are the best candidate binding sites for potentialtranscription factors involved in the repression of GnRHtranscription by melatonin. It has been reported that theorphan nuclear receptor COUP-TFI homodimer can bindto monomeric ROREs [18]. COUP-TFs have also been
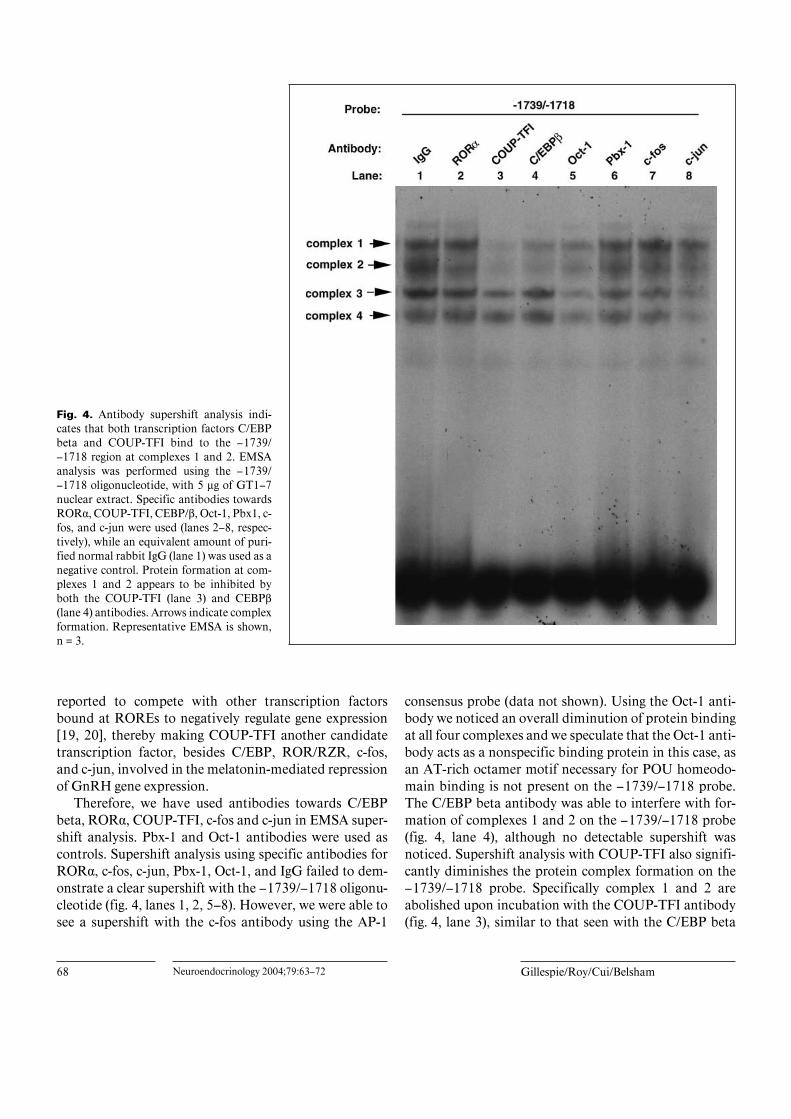
68 Neuroendocrinology 2004;79:63–72 Gillespie/Roy/Cui/Belsham
Fig. 4. Antibody supershift analysis indi-cates that both transcription factors C/EBPbeta and COUP-TFI bind to the –1739/–1718 region at complexes 1 and 2. EMSAanalysis was performed using the –1739/–1718 oligonucleotide, with 5 Ìg of GT1–7nuclear extract. Specific antibodies towardsROR·, COUP-TFI, CEBP/ß, Oct-1, Pbx1, c-fos, and c-jun were used (lanes 2–8, respec-tively), while an equivalent amount of puri-fied normal rabbit IgG (lane 1) was used as anegative control. Protein formation at com-plexes 1 and 2 appears to be inhibited byboth the COUP-TFI (lane 3) and CEBPß(lane 4) antibodies. Arrows indicate complexformation. Representative EMSA is shown,n = 3.
reported to compete with other transcription factorsbound at ROREs to negatively regulate gene expression[19, 20], thereby making COUP-TFI another candidatetranscription factor, besides C/EBP, ROR/RZR, c-fos,and c-jun, involved in the melatonin-mediated repressionof GnRH gene expression.
Therefore, we have used antibodies towards C/EBPbeta, ROR·, COUP-TFI, c-fos and c-jun in EMSA super-shift analysis. Pbx-1 and Oct-1 antibodies were used ascontrols. Supershift analysis using specific antibodies forROR·, c-fos, c-jun, Pbx-1, Oct-1, and IgG failed to dem-onstrate a clear supershift with the –1739/–1718 oligonu-cleotide (fig. 4, lanes 1, 2, 5–8). However, we were able tosee a supershift with the c-fos antibody using the AP-1
consensus probe (data not shown). Using the Oct-1 anti-body we noticed an overall diminution of protein bindingat all four complexes and we speculate that the Oct-1 anti-body acts as a nonspecific binding protein in this case, asan AT-rich octamer motif necessary for POU homeodo-main binding is not present on the –1739/–1718 probe.The C/EBP beta antibody was able to interfere with for-mation of complexes 1 and 2 on the –1739/–1718 probe(fig. 4, lane 4), although no detectable supershift wasnoticed. Supershift analysis with COUP-TFI also signifi-cantly diminishes the protein complex formation on the–1739/–1718 probe. Specifically complex 1 and 2 areabolished upon incubation with the COUP-TFI antibody(fig. 4, lane 3), similar to that seen with the C/EBP beta
Analysis of GnRH Repression by Melatonin Neuroendocrinology 2004;79:63–72 69
antibody. During the EMSA, the antibody is incubatedfor 1 h with the nuclear proteins prior to the addition ofthe probe. Binding of the antibody to the nuclear proteinscould therefore inhibit the binding of the protein to theDNA probe, and as a result no supershift would bedetected.
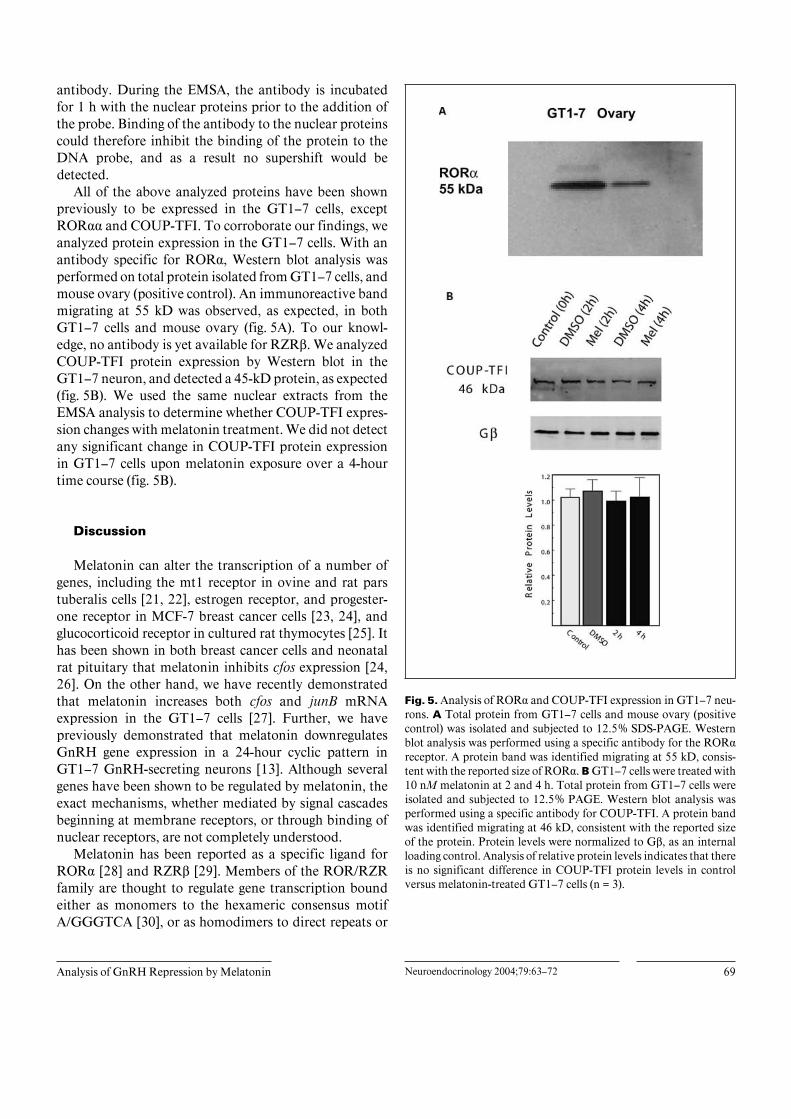
All of the above analyzed proteins have been shownpreviously to be expressed in the GT1–7 cells, exceptROR·· and COUP-TFI. To corroborate our findings, weanalyzed protein expression in the GT1–7 cells. With anantibody specific for ROR·, Western blot analysis wasperformed on total protein isolated from GT1–7 cells, andmouse ovary (positive control). An immunoreactive bandmigrating at 55 kD was observed, as expected, in bothGT1–7 cells and mouse ovary (fig. 5A). To our knowl-edge, no antibody is yet available for RZRß. We analyzedCOUP-TFI protein expression by Western blot in theGT1–7 neuron, and detected a 45-kD protein, as expected(fig. 5B). We used the same nuclear extracts from theEMSA analysis to determine whether COUP-TFI expres-sion changes with melatonin treatment. We did not detectany significant change in COUP-TFI protein expressionin GT1–7 cells upon melatonin exposure over a 4-hourtime course (fig. 5B).
Discussion
Melatonin can alter the transcription of a number ofgenes, including the mt1 receptor in ovine and rat parstuberalis cells [21, 22], estrogen receptor, and progester-one receptor in MCF-7 breast cancer cells [23, 24], andglucocorticoid receptor in cultured rat thymocytes [25]. Ithas been shown in both breast cancer cells and neonatalrat pituitary that melatonin inhibits cfos expression [24,26]. On the other hand, we have recently demonstratedthat melatonin increases both cfos and junB mRNAexpression in the GT1–7 cells [27]. Further, we havepreviously demonstrated that melatonin downregulatesGnRH gene expression in a 24-hour cyclic pattern inGT1–7 GnRH-secreting neurons [13]. Although severalgenes have been shown to be regulated by melatonin, theexact mechanisms, whether mediated by signal cascadesbeginning at membrane receptors, or through binding ofnuclear receptors, are not completely understood.
Melatonin has been reported as a specific ligand forROR· [28] and RZRß [29]. Members of the ROR/RZRfamily are thought to regulate gene transcription boundeither as monomers to the hexameric consensus motifA/GGGTCA [30], or as homodimers to direct repeats or
Fig. 5. Analysis of ROR· and COUP-TFI expression in GT1–7 neu-rons. A Total protein from GT1–7 cells and mouse ovary (positivecontrol) was isolated and subjected to 12.5% SDS-PAGE. Westernblot analysis was performed using a specific antibody for the ROR·receptor. A protein band was identified migrating at 55 kD, consis-tent with the reported size of ROR·. B GT1–7 cells were treated with10 nM melatonin at 2 and 4 h. Total protein from GT1–7 cells wereisolated and subjected to 12.5% PAGE. Western blot analysis wasperformed using a specific antibody for COUP-TFI. A protein bandwas identified migrating at 46 kD, consistent with the reported sizeof the protein. Protein levels were normalized to Gß, as an internalloading control. Analysis of relative protein levels indicates that thereis no significant difference in COUP-TFI protein levels in controlversus melatonin-treated GT1–7 cells (n = 3).

70 Neuroendocrinology 2004;79:63–72 Gillespie/Roy/Cui/Belsham
inverted palindromic consensus binding sites [31]. Todate, melatonin has been shown to downregulate 5-lipox-ygenase gene expression [8] and bone sialoprotein geneexpression [32] directly through ROR/RZR response ele-ments. Some researchers have questioned the validity ofthe melatonin binding studies to ROR/RZR receptorsand further suggest that these receptors may not be thecorrect and/or only nuclear receptors for melatonin [33].
The expression of the ROR· orphan nuclear receptorin GT1–7 neurons raises the question of whether it is alsoinvolved in the melatonin-mediated regulation of GnRHgene expression. The transcriptional changes in GnRH bymelatonin have previously been localized to 5 regionswithin the 300 bp enhancer [13]. Five block mutant con-structs including –1697/–1689, –1736/–1728, –1746/–1738, –1780/–1772 and –1827/–1819 significantly de-creased repression of GnRH gene expression by melato-nin [13]. Each block mutant lies within major footprintsof the 300-bp enhancer as previously determined byDNAse I footprinting [11]. Two of these mutants, –1780/–1772 and –1697/–1689, are located within AT-rich re-gions (AT-a and AT-b, respectively) which have both pre-viously been shown to bind the Oct-1 transcription factor[14]. The –1746/–1738 mutant is located within a foot-printed region previously shown to bind the transcriptionfactor GATA-4 [15]. The –1827/–1819 mutant is alsolocated within a footprinted region; however, the pro-tein(s) bound to this region are yet to be determined. The–1736/–1728 mutant flanks two separate footprints,–1753 to –1734 and –1730 to –1706. Each of these foot-prints has previously been shown to contain regions thatinclude GATA-factor binding sites (GATA-a and GATA-b). However, the GATA factor binding sites do not over-lap the –1736/–1728 block mutant. Upon closer analysisof this region, we noticed the presence of two directrepeats of sequences that are similar (5/6 bp) to the hex-americ consensus binding sequences for members of theROR/RZR nuclear receptor family, located betweenGATA-a and GATA-b. Using supershift analysis, we wereunable to demonstrate that any of the proteins bound tothe –1739/–1718 oligonucleotide is ROR·. Further stud-ies will be undertaken to determine whether the RZRßreceptor, the other nuclear melatonin receptor subtype, isalso present in these protein complexes once the appro-priate antibodies for this orphan nuclear receptor areavailable.
On the other hand, there is some evidence that suggeststhat chicken ovalbumin upstream transcription factor(COUP-TF) homodimers bind to monomeric ROREs[18]. COUP-TFs exist as homodimers and bind DNA
sequences belonging to a wide variety of hormone re-sponse elements (HREs) that are composed of direct,inverted, and everted repeats of the AGGTCA core motif(reviewed in [33]). Because COUP-TFs bind to a numberof HREs, they can act as potent transcriptional repressorsthat antagonize transcriptional activation mediated byother nuclear receptors [18–20, 34, 35]. With EMSAsupershift analysis we have found that COUP-TFI bindsto the –1736/–1728 region of the GnRH enhancer. Wespeculate that COUP-TFI may compete with anothertranscription factor bound at the –1736 to –1728 regionto cause repression of GnRH transcription. The involve-ment of C/EBPß is less clear, but we can speculate that itmay act as a general transcription co-factor. C/EBPß is amember of the basic region-leucine zipper family thatcontrols transcription of a number of genes through pro-tein-protein interactions at the gene level. C/EBPß isexpressed in the GT1–7 cells and has been shown to play arole in the repression of GnRH gene expression by theglutamate/nitric oxide/cGMP signaling pathway by bind-ing a different region of the GnRH neuron-specific en-hancer [36]. Further C/EBP has been linked to the tran-scriptional activity of COUP-TF in a number of studies[34, 37, 38], although these studies did not define theexact role of C/EBP in COUP-TF function. The C/EBPand RORE site overlap at the –1736/–1728 region of theenhancer and may be involved in the switch between bas-al transcriptional activity and repression by melatonin.Further, protein interaction studies must also be done todissect the complex mechanisms involved in the regula-tion of GnRH transcription by melatonin at the level ofthe enhancer.
Additionally, in order to understand the melatonin-induced modulation of protein complex formation on thisregion of the GnRH enhancer, the contribution of mela-tonin-mediated signaling events originating at the level ofthe membrane can not be dismissed. We have previouslyshown that melatonin activates several signal transduc-tion pathways in the GT1–7 neurons, including the ERK1/2 MAPK and protein kinase C pathways linked to therepression of GnRH gene expression [27]. The activationof these signaling pathways by melatonin may result in thephosphorylation of nuclear proteins that may alter theirability to bind DNA [38]. Of interest, a recent report indi-cates that both the ERK 1/2 MAPK and PKC pathwaysmay be involved in the phosphorylation of the COUP-TFI protein [39]. Particularly, PKC-mediated phosphory-lation enhances COUP-TFI affinity for DNA and MAPK-mediated phosphorylation positively regulates the trans-activation function of COUP-TFI, possibly through en-

Analysis of GnRH Repression by Melatonin Neuroendocrinology 2004;79:63–72 71
hancing specific coactivator recruitment. Therefore wepredict that a change in phosphorylation status of the pro-tein upon melatonin stimulation may contribute substan-tially to the repression of GnRH gene expression. Exactlyhow binding at the –1736/–1728 region cooperativelyinteracts with the four other regions within the GnRHenhancer reported to contribute to this process will in-volve detailed studies of protein-protein interactions inorder to delineate the mechanisms involved in melatonin-mediated control of GT1–7 GnRH neuronal function.However, from the studies presented herein, we find thatthe –1736/–1728 region does not bind ROR·, as we origi-nally expected, but instead appears to engage the bindingof C/EBPß and COUP-TFI to regulate GnRH gene ex-pression.
Acknowledgments
We would like to thank Dr. Pamela Mellon, UCSD, for the gener-ous contribution of the GT1–7 cell line. We thank ChristopherMayer for EMSA analysis support, and Dr. Bernardo Yusta for criti-cal reading of the manuscript. This work was supported by a grantfrom the National Sciences and Engineering Research Council ofCanada. D.D.B. is a Canadian Health Research Institute Scholar andCanada Foundation for Innovation Researcher. H.C. was supportedby a Premier’s Research Excellence Award to D.D.B.
References
1 Aleandri V, Spina V, Morini A: The pinealgland and reproduction. Hum Reprod Update1996;2:225–235.
2 Vanecek J, Klein DC: A subpopulation of neo-natal gonadotropin-releasing hormone sensi-tive pituitary cells is responsive to melatonin.Endocrinology 1993;133:360–367.
3 Martin JE, Klein DC: Melatonin inhibition ofthe neonatal pituitary response to luteinizinghormone-releasing factor. Science 1976;191:301–302.
4 Rasmussen DD: Diurnal modulation of rat hy-pothalamic gonadotropin-releasing hormonerelease by melatonin in vitro. J EndocrinolInvest 1993;16:1–7.
5 Malpaux B, Thiery JC, Chemineau P: Melato-nin and the seasonal control of reproduction.Nutr Dev 1999;39:355–366.
6 Reppert SM, Weaver DR, Ebisawa T: Cloningand characterization of a mammalian melato-nin receptor that mediates reproductive andcircadian responses. Neuron 1994;13:1177–1185.
7 Reppert SM, Godson C, Mahle CD, WeaverDR, Slaugenhaupt SA, Gusella JF: Molecularcharacterization of a second melatonin recep-tor expressed in human retina and brain: TheMel1b melatonin receptor. Proc Natl Acad SciUSA 1995;92:8734–8738.
8 Carlberg C, Wiesenberg I: The orphan receptorfamily RZR/ROR, melatonin and 5-lipoxygen-ase: An unexpected relationship. J Pineal Res1995;18:171–178.
9 Wiesenberg I, Missbach M, Carlberg C: Thepotential role of the transcription factor RZR/ROR as a mediator of nuclear melatonin sig-naling. Restor Neurol Neurosci 1998;12:143–150.
10 Mellon PL, Windle JJ, Goldsmith P, Pedula C,Roberts J, Weiner RI: Immortalization of hy-pothalamic GnRH neurons by genetically tar-geted tumorigenesis. Neuron 1990;5:1–10.
11 Whyte DB, Lawson MA, Belsham DD, EralySA, Bond CT, Adelman JP, Mellon PL: A neu-ron-specific enhancer targets expression of thegonadotropin-releasing hormone gene to hypo-thalamic neurosecretory neurons. Mol Endo-crinol 1995;9:467–477.
12 Nelson SB, Lawson MA, Kelley CG, MellonPL: Neuron-specific expression of the rat go-nadotropin-releasing hormone gene is con-ferred by interactions of a defined promoterelement with the enhancer in GT1–7 cells. MolEndocrinol 2000;14:1509–1522.
13 Roy D, Angelini N, Frieda H, Brown GM, Bel-sham DD: Cyclical regulation of GnRH geneexpression in GT1–7 GnRH-secreting neuronsby melatonin. Endocrinology 2001;142:4711–4720.
14 Clark ME, Mellon PL: The POU homeodo-main transcription factor Oct-1 is essential foractivity of the gonadotropin-releasing hormoneneuron-specific enhancer. Mol Cell Biol 1995;15:6169–6177.
15 Lawson MA, Whyte DB, Mellon PL: GATAfactors are essential for activity of the neuron-specific enhancer of the gonadotropin-releasinghormone gene. Mol Cell Biol 1996;16:3596–3605.
16 Lee KAW, Bindereif A, Green MR: A small-scale procedure for preparation of nuclear ex-tracts that support efficient transcription andpre-mRNA splicing. Gene Anal Techn 1988;5:22–31.
17 Laemmli U: Cleavage of structural proteinsduring the assembly of the head of bacterio-phage T4. Nature 1970;227:680–685.
18 Schrader M, Danielsson C, Wiesenberg I, Carl-berg C: Identification of natural monomericresponse elements of the nuclear receptorRZR/ROR: They also bind COUP-TF homo-dimers. J Biol Chem 1996;271:19732–19736.
19 Leng X, Cooney AJ, Tsai SY, Tsai MJ: Molecu-lar mechanisms of COUP-TF-mediated tran-scriptional repression: Evidence for transre-pression and active repression. Mol Cell Biol1996;16:2332–2340.
20 Zeitoun K, Takayama K, Michael MD, BulunSE: Stimulation of aromatase P450 promoter(II) activity in endometriosis and its inhibitionin endometrium are regulated by competitivebinding of steroidogenic factor-1 and chickenovalbumin upstream promoter transcriptionfactor to the same cis-acting element. Mol En-docrinol 1999;13:239–253.
21 Guerrero HY, Gauer F, Schuster C, Pevet P,Masson-Pevet M: Melatonin regulates themRNA expression of the mt(1) melatonin re-ceptor in the rat pars tuberalis. Neuroendocri-nology 2000;71:163–169.
22 Barrett P, MacLean A, Davidson G, MorganPJ: Regulation of the Mel 1a melatonin recep-tor mRNA and protein levels in the ovine parstuberalis: Evidence for a cyclic adenosine 3),5)-monophosphate-independent Mel 1a receptorcoupling and an autoregulatory mechanism ofexpression. Mol Endocrinol 1996;10:892–902.
23 Molis TM, Spriggs LL, Hill SM: Modulation ofestrogen receptor mRNA expression by mela-tonin in MCF-7 human breast cancer cells. MolEndocrinol 1994;8:1681–1690.
24 Molis TM, Spriggs LL, Jupiter Y, Hill SM:Melatonin modulation of estrogen-regulatedproteins, growth factors, and proto-oncogenesin human breast cancer. J Pineal Res 1995;18:93–103.
25 Sainz RM, Mayo JC, Reiter RJ, Antolin I, Este-ban MM, Rodriguez C: Melatonin regulatesglucocorticoid receptor: An answer to its anti-apoptotic action in thymus. FASEB J 1999;13:1547–1556.
26 Sumova A, Vanecek J: Melatonin inhibitsGnRH-induced increase of cFOS immunoreac-tivity in neonatal rat pituitary. J Neuroendocri-nol 1997;9:135–139.

72 Neuroendocrinology 2004;79:63–72 Gillespie/Roy/Cui/Belsham
27 Roy D, Belsham DD: Melatonin receptor acti-vation regulates gonadotropin-releasing hor-mone (GnRH) gene expression and secretion inGT1–7 GnRH neurons: Signal transductionmechanisms. J Biol Chem 2002;277:251–258.
28 Becker-Andre M, Wiesenberg I, Schaeren-Wiemers N, Andre E, Missbach M, Saurat JH,Carlberg C: Pineal gland hormone melatoninbinds and activates an orphan of the nuclearreceptor superfamily. J Biol Chem 1994;269:28531–28534.
29 Wiesenberg I, Missbach M, Kahlen JP, Schra-der M, Carlberg C: Transcriptional activationof the nuclear receptor RZR alpha by the pinealgland hormone melatonin and identification ofCGP 52608 as a synthetic ligand. Nucl AcidsRes 1995;23:327–333.
30 Giguere V, Tini M, Flock G, Ong E, Evans RM,Otulakowski G: Isoform-specific amino-termi-nal domains dictate DNA-binding propertiesof ROR alpha, a novel family of orphan hor-mone nuclear receptors. Genes Dev 1994;8:538–553.
31 Carlberg C, Hooft van Huijsduijnen R, StapleJK, DeLamarter JF, Becker-Andre M: RZRs, anew family of retinoid-related orphan receptorsthat function as both monomers and homodi-mers. Mol Endocrinol 1994;8:757–770.
32 Roth J, Kim B, Lin W, Choi M: Melatonin pro-motes osteoblast differentiation and bone for-mation. J Biol Chem 1999;274:22041–22047.
33 Giguere V: Orphan nuclear receptors: Fromgene to function. Endocr Rev 1999;20:689–725.
34 Lethimonier C, Flouriot G, Kah O, DucouretB: The glucocorticoid receptor represses thepositive autoregulation of the trout estrogenreceptor gene by preventing the enhancer effectof a C/EBPbeta-like protein. Endocrinology2002;143:2961–2974.
35 Achatz G, Holzl B, Speckmayer R, Hauser C,Sandhofer F, Paulweber B: Functional do-mains of the human orphan receptor ARP-1/COUP-TFII involved in active repression andtransrepression. Mol Cell Biol 1997;17:4914–4932.
36 Belsham DD, Mellon PL: Transcription factorsOct-1 and C/EBP b (CCAAT/enhancer-bindingprotein-b) are involved in the glutamate/nitricoxide/cyclic-guanosine 5)-monophosphate-me-diated repression of mediated repression of go-nadotropin-releasing hormone gene expres-sion. Mol Endocrinol 2000;14:212–228.
37 Schwartz C, Catez P, Rohr O, Lecestre D,Aunis D, Schaeffer E: Functional interactionsbetween C/EBP, Sp1, and COUP-TF regulatehuman immunodeficiency virus type 1 genetranscription in human brain cells. J Virol2000;74:65–73.
38 Boulikas T: Phosphorylation of transcriptionfactors and control of the cell cycle. Crit RevEukaryot Gene Expr 1995;5:1–77.
39 Gay F, Barath P, Desbois-Le Peron C, MetivierR, Le Guevel R, Birse D, Salbert G: Multiplephosphorylation events control chicken oval-bumin upstream promoter transcription factorI orphan nuclear receptor activity. Mol Endo-crinol 2002;16:1332–1351.




















