
Hypothalamic Pro‐GnRH‐GAP, GnRH‐I and GnRH‐II During the Onset of Photorefractoriness in the...
10
Hypothalamic Pro-GnRH-GAP, GnRH-I and GnRH-II During the Onset of Photorefractoriness in the White-Crowned Sparrow ( Zonotrichia leucophrys gambelii ) S. L. Meddle,*§ S. Bush,* P. J. Sharp,† R. P. Millar‡ and J. C. Wingfield§ *Laboratory of Neuroendocrinology, Centre for Integrative Physiology, School of Veterinary Biomedical Sciences, Royal (Dick) School of Veterinary Medicine, Summerhall, Edinburgh, UK. †The Roslin Institute, Roslin, Midlothian, UK. ‡Human Reproductive Sciences Unit, Medical Research Council, University of Edinburgh Chancellor’s Building, Edinburgh, UK. §Department of Biology, University of Washington, Seattle, WA, USA. Key words: birds, brain, gonadotrophin releasing hormone, GnRH, immunohistochemistry, photoperiodism, Pro-GnRH-GAP, seasonal breeding. Abstract Gambel’s white-crowned sparrow is a long distance migrant that undergoes spontaneous gonadal regression as a result of long day exposure. This termination of breeding is caused by the development of photorefractoriness and the birds become insensitive to long days, including continuous light. The present study investigated its possible mechanisms by examining the activity of the gonadotrophin-releasing hor- mone (GnRH) system under different photoperiodic regimes. We investigated the localisation and distri- bution of GnRH-I, its precursor pro-GnRH-GAP and GnRH-II in Gambel’s white-crowned sparrow brain using immunocytochemistry with specific antibodies during photostimulation and the development of photore- fractoriness. The study revealed that photoperiodic treatment, including the onset of photorefractoriness, had no significant effect on the size or number of GnRH-I, pro-GnRH-GAP or GnRH II immunoreactive cells, or the density of the GnRH-I, pro-GnRH-GAP immunoreactive fibres at the median eminence. GnRH-II was not found in the median eminence, suggesting that it does not regulate pituitary gonadotrophin secretion. GnRH-I measurement in hypothalamic extracts by radioimmunoassay did not reveal any significant differ- ence between birds that were photostimulated or in the early stages of photorefractoriness. Furthermore, the action of the excitatory amino acid glutamate agonist N-methyl-D-aspartate on GnRH neurones in photorefractory birds was demonstrated by the significant blockade of luteinising hormone release with a specific GnRH antagonist. Taken together, these results suggest that, in Gambel’s white-crowned sparrow, a decrease in GnRH-I secretion is the initial step for the onset of photorefractoriness and not a decrease in GnRH-I biosynthesis. In most bird species from temperate and high latitudes, seasonal breeding begins during spring when the increase in day length stimulates gonadal maturation (1); this is termed photostimulation. Seasonal breeding is terminated by the development of absolute photorefractoriness, whereby spon- taneous gonadal regression occurs as a result of long day exposure and birds become insensitive to long days, including continuous light (1–3). The phenomenon is characterised by a rapid inhibition of the gonadotrophin-releasing hormone (GnRH) system causing a decrease in luteinising hormone (LH) and follicle stimulating hormone (FSH) release, and thus gonadal regression. It has now been firmly established that the effects of varying day lengths throughout the year on the reproductive system of passerine bird species, is due to the effects on GnRH synthesis and secretion (1, 4–9). Photostimulation causes the rapid release of LH (1, 3, 4, 10–12) and this is closely associated with an increase in hypothalamic GnRH mRNA at least in quail (Coturnix coturnix) and domestic chickens (Gallus domesticus) (13, 14). The second effect of chronic exposure to long days is photorefractoriness and subsequent re-exposure to short days is requisite to allow photosensitivity to resume. The mechanism of absolute photorefractoriness is still a major topic of debate within the field and is yet to be satisfactorily described across all species that exhibit it. Absolute photorefractoriness has mostly been studied in the European starling (Sturnus vulgaris) and the development Correspondence to: Dr Simone L. Meddle, Centre for Integrative Physiology, College of Medicine & Veterinary Medicine, The University of Edinburgh, Hugh Robson Building, George Square, Edinburgh EH8 9XD, UK (e-mail: [email protected]). Journal of Neuroendocrinology, 2006, Vol. 18, 217–226 doi:10.1111/j.1365-2826.2005.01403.x ȑ 2006 The Authors. Journal compilation ȑ 2006 Blackwell Publishing Ltd
Transcript of Hypothalamic Pro‐GnRH‐GAP, GnRH‐I and GnRH‐II During the Onset of Photorefractoriness in the...

Hypothalamic Pro-GnRH-GAP, GnRH-I and GnRH-II During theOnset of Photorefractoriness in the White-Crowned Sparrow(Zonotrichia leucophrys gambelii)
S. L. Meddle,*§ S. Bush,* P. J. Sharp,† R. P. Millar‡ and J. C. Wingfield§*Laboratory of Neuroendocrinology, Centre for Integrative Physiology, School of Veterinary Biomedical Sciences, Royal (Dick) School of VeterinaryMedicine, Summerhall, Edinburgh, UK.†The Roslin Institute, Roslin, Midlothian, UK.‡Human Reproductive Sciences Unit, Medical Research Council, University of Edinburgh Chancellor’s Building, Edinburgh, UK.§Department of Biology, University of Washington, Seattle, WA, USA.
Key words: birds, brain, gonadotrophin releasing hormone, GnRH, immunohistochemistry, photoperiodism, Pro-GnRH-GAP, seasonal breeding.
Abstract
Gambel’s white-crowned sparrow is a long distance migrant that undergoes spontaneous gonadal
regression as a result of long day exposure. This termination of breeding is caused by the development of
photorefractoriness and the birds become insensitive to long days, including continuous light. The present
study investigated its possible mechanisms by examining the activity of the gonadotrophin-releasing hor-
mone (GnRH) system under different photoperiodic regimes. We investigated the localisation and distri-
bution of GnRH-I, its precursor pro-GnRH-GAP and GnRH-II in Gambel’s white-crowned sparrow brain using
immunocytochemistry with specific antibodies during photostimulation and the development of photore-
fractoriness. The study revealed that photoperiodic treatment, including the onset of photorefractoriness,
had no significant effect on the size or number of GnRH-I, pro-GnRH-GAP or GnRH II immunoreactive cells,
or the density of the GnRH-I, pro-GnRH-GAP immunoreactive fibres at the median eminence. GnRH-II was
not found in the median eminence, suggesting that it does not regulate pituitary gonadotrophin secretion.
GnRH-I measurement in hypothalamic extracts by radioimmunoassay did not reveal any significant differ-
ence between birds that were photostimulated or in the early stages of photorefractoriness. Furthermore,
the action of the excitatory amino acid glutamate agonist N-methyl-D-aspartate on GnRH neurones in
photorefractory birds was demonstrated by the significant blockade of luteinising hormone release with a
specific GnRH antagonist. Taken together, these results suggest that, in Gambel’s white-crowned sparrow,
a decrease in GnRH-I secretion is the initial step for the onset of photorefractoriness and not a decrease in
GnRH-I biosynthesis.
In most bird species from temperate and high latitudes,seasonal breeding begins during spring when the increase inday length stimulates gonadal maturation (1); this is termedphotostimulation. Seasonal breeding is terminated by thedevelopment of absolute photorefractoriness, whereby spon-taneous gonadal regression occurs as a result of long dayexposure and birds become insensitive to long days, includingcontinuous light (1–3). The phenomenon is characterised by arapid inhibition of the gonadotrophin-releasing hormone(GnRH) system causing a decrease in luteinising hormone(LH) and follicle stimulating hormone (FSH) release, andthus gonadal regression.It has now been firmly established that the effects of
varying day lengths throughout the year on the reproductive
system of passerine bird species, is due to the effects onGnRH synthesis and secretion (1, 4–9). Photostimulationcauses the rapid release of LH (1, 3, 4, 10–12) and this isclosely associated with an increase in hypothalamic GnRHmRNA at least in quail (Coturnix coturnix) and domesticchickens (Gallus domesticus) (13, 14). The second effect ofchronic exposure to long days is photorefractoriness andsubsequent re-exposure to short days is requisite to allowphotosensitivity to resume. The mechanism of absolutephotorefractoriness is still a major topic of debate withinthe field and is yet to be satisfactorily described across allspecies that exhibit it.
Absolute photorefractoriness has mostly been studied inthe European starling (Sturnus vulgaris) and the development
Correspondence to: Dr Simone L. Meddle, Centre for Integrative Physiology, College of Medicine & Veterinary Medicine, The University of Edinburgh,
Hugh Robson Building, George Square, Edinburgh EH8 9XD, UK (e-mail: [email protected]).
Journal of Neuroendocrinology, 2006, Vol. 18, 217–226 doi:10.1111/j.1365-2826.2005.01403.x
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd

of reproductive photorefractoriness in this species is thoughtto be caused by an inhibition of GnRH release (1, 5, 6).Currently, there is evidence that GnRH synthesis is reduced(9) and, in long-term photorefractory birds, inhibited assynaptic input to GnRH cell bodies is increased (15). Inaddition, following chronic long day exposure, there is asignificant reduction in size and staining density of GnRHperikarya, presumably indicating a decrease in synthesis (8,16). It is assumed that the maintenance of photorefractorinessin this species is also the result of an inhibition of GnRH genetranscription because long-term photorefractory starlingshave over a 100-fold decrease in hypothalamic GnRH peptidecontent (7, 17). Interestingly, hypothalamic GnRH content isnotably lower in photorefractory starlings on long daylengths than in photosensitive birds on short day lengths(4), indicating that hypothalamic GnRH increases during thephotosensitive phase.
Collectively, these findings support a two-step theorywhereby photorefractoriness is initiated by an inhibition ofGnRH release and is maintained by a complete inhibition ofGnRH synthesis in the hypothalamus (5, 6). A clear way totest this theory would be to assess changes in GnRH mRNAexpression. However to date, cloning of the GnRH gene inany avian species that becomes absolutely refractory hasproved elusive. Currently, the GnRH gene has only beencloned in the chicken and quail (13, 18) and these are speciesthat do not become absolutely photorefractory (3, 19). Analternative approach is to measure changes in its precursorpeptide (pro-GnRH-GAP) within the hypothalamus; pro-GnRH-GAP undergoes post-translational processing toproduce a 23 amino-acid signal peptide, GnRH and a 56amino-acid GnRH-associated peptide (GAP) (20). Levels ofpro-GnRH-GAP protein have been shown to be a sensitiveindex of GnRH gene transcription in the rat (21). Previousstudies in starlings have shown that photorefractoriness mayalso be triggered by a reduction in pro-GnRH-GAP synthesiswhich occurs prior to the decrease in hypothalamic GnRHimmunoreactivity (9).
Two forms of GnRH exist in birds; GnRH-I (22) andGnRH-II (23, 24). The recently discovered lamprey GnRH-III has also been described in passerines (25) but is yet to beconfirmed. GnRH-I cell bodies can be immunocytochemicallyidentified in the preoptic area of the hypothalamus, fromwhich fibres project through the brain to terminate in themedian eminence (26). It is widely believed that GnRH-I isthe endogenous releasing hormone for gonadotrophins,owing to its well-documented restricted anatomical locationof fibres in the median eminence (27–30). There is conflictingevidence for whether GnRH-II is found in the medianeminence and the discrepancies may be due to differentantibody sensitivities. GnRH-II has been identified in thequail and chicken median eminence (31, 32) but, in otherstudies, GnRH-II was not found to be present in the medianeminence of the chicken or in the turkey (Meleagris gallopavo)(28, 33). Both GnRH-I and II are potent releasers of LH andFSH (12, 34) in vivo, although there is no evidence thatgonadotrophin release is regulated by GnRH-II under phy-siological conditions. GnRH-II is undetectable in perfusatefrom isolated median eminences (26, 33, 35) but it is releasedfrom quail and turkey hypothalamic explants that lack the
median eminence (29). In addition, only immunisationagainst GnRH-I but not GnRH-II causes complete collapseof the reproductive system (33).
The white-crowned sparrow (Zonotrichia leucophrys gam-belii), is a long distance migrant that undergoes rapidspontaneous gonadal regression as a result of long dayexposure. The mechanism for the induction of absolutephotorefractoriness is still an area of debate but, in Gambel’swhite-crowned sparrow, it appears to be caused by aninhibition or lack of stimulation of GnRH secretion. Injec-tions of N-methyl-D-aspartate (NMDA), a neuroexcitatoryglutamate analogue, cause an increase in LH release regard-less of reproductive state, establishing that GnRH neuronesin reproductively quiescent birds can respond to the stimu-latory action of excitatory amino acids (11).
The present study aimed to further substantiate theevidence that absolute photorefractoriness onset in Gambel’swhite-crowned sparrow is not caused by depleted GnRH-Istores. It has already been established that photorefractori-ness in this species is not caused by the pituitary gland orgonad insensitivity because i.v. injections of GnRH or LHcause a rise in LH and testosterone, respectively (12). Thisspecies was also chosen because it exhibits a truncatedbreeding season. All other immunocytochemical GnRHstudies on passerines have been performed on species thatexhibit a relatively protracted breeding season. Furthermore,these studies have employed antibodies that detect bothGnRH-I and GnRH-II, but there is still no conclusiveevidence to confirm which form is responsible for seasonalbreeding. Thus, one objective of this study was to use specificantibodies for GnRH-I, pro-GnRH-GAP and GnRH-II toimmunocytochemically map the distribution of these forms ofGnRH and, for the first time, evaluate their roles in seasonalbreeding within a single study.
Immunocytochemical studies are a useful means of deter-mining the distribution and estimating the levels of a peptidewithin brain tissue. However, radioimmunoassay (RIA) onbrain tissue provides a more accurate quantifiable means ofmeasuring the hypothalamic content of a peptide. Therefore,we additionally compared the actual hypothalamic content ofGnRH-I between photostimulated and photorefractory birdsusing a specific RIA. Finally, following on from previousfindings of the action of NMDA on LH secretion in thewhite-crowned sparrow (11), we sought to demonstrate thatNMDA has a specific stimulatory action on GnRH release,by blocking its effect with a specific GnRH antagonist.
Materials and methods
Subjects
White-crowned sparrows (Z. leucophrys gambelii) were captured during
autumnal migration in mist nets in Eastern Washington State and held in
outdoor aviaries at the Department of Biology, University of Washington in
Seattle. Prior to the winter solstice, they were anaesthetised with methoxy-
flurane vapour (Metofane; Mallinckrodt Veterinary, Inc., IL, USA) and
laparotomised via a small incision in the body cavity, just behind the ribs, to
determine sex. All males had regressed testes and were housed singly in wire
cages (41 · 39 · 35 cm) and maintained on short days (SD) of 8 h of light
per day (8 : 16 h light/dark cycle at a controlled temperature of 20 �C). Asound proof environmental chamber was designated for each photoperiodic
treatment and only male birds were housed in the chambers. The birds were
218 GnRH and photorefractoriness in the white-crowned sparrow
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

not visually isolated from each other. Water (drinking water and birdbaths)
and bird seed were freely available. The Animal Care and Use Committee at
the University of Washington approved all procedures used in these
experiments.
Effects of photoperiod on pro-GnRH-GAP, GnRH-I and GnRH-II
immunoreactivity in the diencephalon
Male white-crowned sparrows were randomly assigned to one of four
photoperiodic groups (n ¼ 6 in each group). The first group, termed
�photosensitive�, was maintained on an 8 : 16 h light/dark cycle. The second
and third �photostimulated� group experienced either 2 or 5 weeks of long days
(LD) (20 : 4 h light/dark cycle). Birds in the fourth group comprised of
�photorefractory� birds (that were in the early stages of photorefractoriness)
and were subjected to 9 weeks of LD (20 : 4 h light/dark cycle). These groups
were chosen to generate birds in different stages of the reproductive cycle;
birds were transferred to LDs at different times so that all groups could be
sampled on the same calendar day.
Prior to photoperiodic manipulation and at the end of the experiment,
blood samples were collected by wing venepuncture (approximately 100 llwhole blood). Blood was centrifuged and the plasma samples stored at )20 �Cuntil processing for LH measurement. A single assay was performed to
eliminate interassay variation using a micromodification of the RIA originally
devised by Follett et al. (36), as outlined for use in the white-crowned sparrow
in Follett et al. (37). The intra-assay coefficient of variation was 3.5% at
1.52 ng/ml and the level of assay sensitivity or minimum detection limit was
0.05 ng. The results are expressed in terms of ng/ml against a chicken LH
standard (fraction IRC2).
The sparrows were anaesthetised by i.m. injection with an overdose of
250 mg/kg sodium pentobarbitone (Nembutal; Abbott Laboratories, IL,
USA), assessed for moult (evaluated by primary feather loss and re-growth)
and perfused intracardially with 10 ml of heparinised 0.9% saline followed
by 150 ml of modified Zamboni fixative [1.8% paraformaldehyde and
7.5 ml saturated picric acid in 0.01 M phosphate-buffered saline (PBS),
pH 7.4]. Following perfusion, the testes were removed and weighed. Brains
were dissected out of the skull, postfixed in Zamboni fixative at 4 �Covernight, washed in PBS and embedded in gelatine. Coronal 50 lmsections through the whole brain were cut on a vibratome. Alternate free-
floating sections were labelled for pro-GnRH-GAP (code 1947) [for details
and validation of this antibody, see (38)], GnRH-I (code 3/3) or GnRH-II
(code 11/3) [for details and validation of these antibodies, see (33)]. Prior to
the present experiment, the working concentrations of the pro-GnRH-GAP,
GnRH I and GnRH II antibodies were determined for the white-crowned
sparrow brain. Sections of white-crowned sparrow brain were incubated in
primary antibody at the following dilutions: 1 : 100, 1 : 500, 1 : 1000,
1 : 5000, 1 : 10 000 and 1 : 20 000. Decisions on working concentrations
were made based on the signal to background ratio, with optimum staining
considered when cells and fibres were adequately labelled with low
background.
In brief, sections were washed for 2 h in PBS (six changes) containing 0.2%
Triton X-100 (PBST). Endogenous peroxidase activity was blocked by
incubation in 0.3% hydrogen peroxide in PBS (10 min) followed by washing
in PBST (3 · 10 min). The sections were then incubated for 70 h in the
primary rabbit polyclonal antiserum (pro-GnRH-GAP: diluted 1 : 1000;
GnRH I and GnRH II, diluted 1 : 10 000) in 2% normal goat serum for 70 h
at 4 �C. The antibody: antigen complex was visualised using the peroxidase-
antiperoxidase (PAP) technique. Sections were rinsed in PBST and incubated
in goat anti-rabbit immunoglobulins (IgG) diluted at 1 : 100 in PBST for 2 h
at room temperature. Sections were then washed in PBST and then transferred
to the PAP complex diluted at 1 : 100 in PBST for 2 h at room temperature.
Following rinses in PBST and 0.05 M Tris buffer, pH 7.4 (10 min), the
peroxidase was visualised with 3¢,3-diaminobenzidine containing 0.03%
hydrogen peroxide in Tris buffer with the reaction terminated by several
washes in PBS. Serial sections were mounted on gelatine-coated slides and left
to dry overnight. Slides were dehydrated and mounted in DPX mountant
(Aldrich Chemical Company, Inc., WI, USA).
All immunostaining was prevented by incubation of the sections in
nonimmune rabbit serum instead of the antibody. Antibody preabsorption
with 2 lg/ml of the respective synthetic peptide, used to generate the
antibodies, also blocked staining. All working concentrations of antibodies
and solutions used in the labelling of GnRH-I, GnRH-II and its precursor
were optimised prior to the study.
Immunoreactivity quantification and image analysis
All measurements were carried out without the experimenter being aware of
the bird’s photoperiodic treatment. Cell counts were performed with a tally
counter and the cell size and density of fibre staining was measured using
computer-assisted image analysis systems (NIH Image 1.62, Bethesda MD,
USA, or Openlab, Improvision, Coventry, UK). Images of the sections were
captured under bright-field illumination on a light microscope (Carl Zeiss Co.
Ltd., Thornwood, NY, USA) using a · 40 objective lens attached to a digital
camera (Hamamatsu C4742-95; Photonics, Tokyo, Japan) and Apple
Macintosh computer (Apple Computer Inc., Cupertino, CA, USA).
Cell counts
Each brain section from each bird was examined for the presence of pro-
GnRH-GAP, GnRH-I and GnRH-II immunoreactivity. The distribution was
mapped and all labelled cells were counted throughout the brain to allow a
mean calculation of the number of cells counted per treatment group. GnRH-
II cell numbers are expressed as mean values per 50 lm section due to missing
sections from some birds.
Cell size measurement
Sections containing the main population of cell bodies were selected and a
random field of view containing many cells was centred on the computer
screen. The nearest five cells to the centre of the screen that were in focus and
were cut through the centre of the cell were selected and the cell area was
measured and expressed as lm2. Cells covered by other cell bodies, or cells
outside the plane of section and focus, were excluded from the analysis. The
procedure was repeated for two other fields of view, until a total of 15 cells
were measured from each brain. The procedure was repeated for three fields of
view on either side of the third ventricle. Consequently, 15 cells were measured
per side of each section, yielding 30 cells per section. This was repeated in two
other sections, producing 90 measurements per bird from which a mean was
calculated. If some of the brains contained less than 90 immunolabelled
perikarya with clearly defined boundaries, then the mean was taken from as
many cells measured as possible; this was never less than 75 cells.
Optical staining density of fibre tracts and axon terminals in the median
eminence
Image analysis on pro-GnRH-GAP or GnRH-I immunolabelled brains was
carried out on two sections per brain containing medially cut portions through
the median eminence. These sections contained both axon terminals and fibre
tracts. During analysis, a 2000 lm2 region of interest (ROI) was determined
and maintained throughout. For each section chosen, six background
measurements and six axon terminal/fibre tract measurements were obtained.
Readings were obtained by placing the ROI over areas containing high density
staining and background measurements were obtained by placing the ROI
over areas of the median eminence without fibre-tract or axon terminal
staining. The six measurements of each type were then averaged. The mean
background value was subtracted from the mean optical density value to yield
a final specific density value for the section. The density values for the two
sections measured per brain were then averaged to calculate a brain mean.
Effects of photoperiod on GnRH-I in the forebrain and medial basal
hypothalamus of photostimulated and photorefractory birds
Photostimulated (2 weeks, 20 : 4 h LD; n ¼ 7) or photorefractory (12 weeks,
20 : 4 h LD; n ¼ 11) male white-crowned sparrows were anaesthetised with
Metofane, assessed for moult (evaluated by primary feather loss and re-
growth) and then decapitated. Trunk blood was taken from each bird for LH
quantification as in the first experiment. All birds were transferred to a
20 : 4 h light/dark cycle at different times so that all groups could be sampled
on the same calendar day. Whole brains were immediately removed from the
skull and blocks of brain removed from the preoptic area (6 · 6 · 8 mm) and
medial basal hypothalamus (5 · 5 · 7 mm). The whole cerebellum was taken
as a control region. Immediately following dissection each block of brain
tissue was homogenised in 2 ml ice cold 2 M acetic acid and then centrifuged at
3000 r.p.m. for 1 h and 20 min. The supernatant was then aspirated and
stored at )70 �C until chicken GnRH-I was measured in a single assay (run at
three dilutions) (40). The assay sensitivity was 2.6 pg/sample and the results
GnRH and photorefractoriness in the white-crowned sparrow 219
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

are expressed in terms of ng/ml tissue sample against a chicken GnRH-I
standard.
Acute effects of NMDA on LH secretion following GnRH antagonist treatment
Male white-crowned sparrows were randomly assigned to one of three
photoperiodic groups (n ¼ 12 in each group) as in the first experiment. All
birds were transferred to a 20 : 4 h light/dark cycle at different times so that
all groups could be sampled on the same calendar day. Each group was
divided into two treatments (n ¼ 6 in each treatment) and a blood sample was
taken immediately prior to a 20 ll s.c. injection of either vehicle (peanut oil;
Sigma, St Louis, MO, USA) or a specific GnRH antagonist (pGlu-D-Phe-Pro-
Ser-Tyr-D-Phe-Leu-Arg-Pro-Gly-NH2; Sigma) at a dose of 40 lg/kg body
weight. Ninetey minutes following treatment, another blood sample was taken
and all birds were immediately injected with NMDA (Sigma) dissolved in
physiological saline at an s.c. dose of 20 mg/kg body weight (in 20 ll). 20 min
following NMDA injection, a final blood sample was taken. Blood samples
were collected by wing venepuncture (approximately 100 ll whole blood).
Blood was centrifuged and the plasma samples stored at )20 �C until
processing for LH measurement in a single assay as detailed above. All birds
were examined for signs of moult (evaluated by primary feather loss and re-
growth) on the day of experiment.
Statistical analysis
Student’s t-test, one-way ANOVA, Kruskal–Wallis analysis on ranks, two-way
ANOVA, and post-hoc tests (Student–Newman–Keuls and all pairwise multiple
comparison procedure Holm-Sidak method) were used (Sigma Stat software)
as appropriate to compare groups. P < 0.05 was considered statistically
significant. The results are expressed as the group means ± SEM.
Results
Effects of photoperiod on GnRH-I, GnRH-II and pro-GnRH-GAP immunoreactivity in the diencephalon
Birds
Bird mass significantly increased as a result of photostimu-lation and decreased at the onset of photorefractoriness[F(3,21) ¼ 6.51; P < 0.01]. Bird mass increased from29.0 ± 1.5 g on SD to 36.3 ± 2.6 g as a result of sexualmaturity and increased feeding following 2 weeks of LD. Themean mass was significantly lower following 9 weeks of LD(31.0 ± 1.65 g) as a result of body weight stability andgonadal regression (P < 0.05).
Photoperiodic treatment significantly altered paired testi-cular mass (H ¼ 20.09; P < 0.001) (Fig. 1). Photostimula-tion significantly induced a 10-fold increase in gonadal weightfollowing 2 weeks of LD (P < 0.05). Birds exposed to5 weeks of LD underwent rapid testicular maturation,whereas 9 weeks of LD photostimulation caused significanttesticular regression. All birds photostimulated for 9 weeks ofLD were undergoing both primary feather and body moult.
Plasma LH
Prior to photoperiodic treatment, LH levels were low(0.23 ± 0.02 ng/ml; n ¼ 24) and not significantly differentbetween groups [F(3,20) ¼ 2.67; P > 0.05]. Photostimulationsignificantly altered LH levels (H ¼ 17.97; P < 0.001). LHlevels remained low in birds maintained on SD(0.23 ± 0.03 ng/ml) but increased to 1.32 ± 0.08 ng/ml after2 weeks and to 1.96 ± 0.37 ng/ml after 5 weeks of LDphotostimulation. In photorefractory birds (9 weeks of LD)LH levels were significantly decreased to 0.74 ± 0.03 ng/ml(P < 0.05).
Distribution of pro-GnRH-GAP, GnRH-I and GnRH-II
in the diencephalon
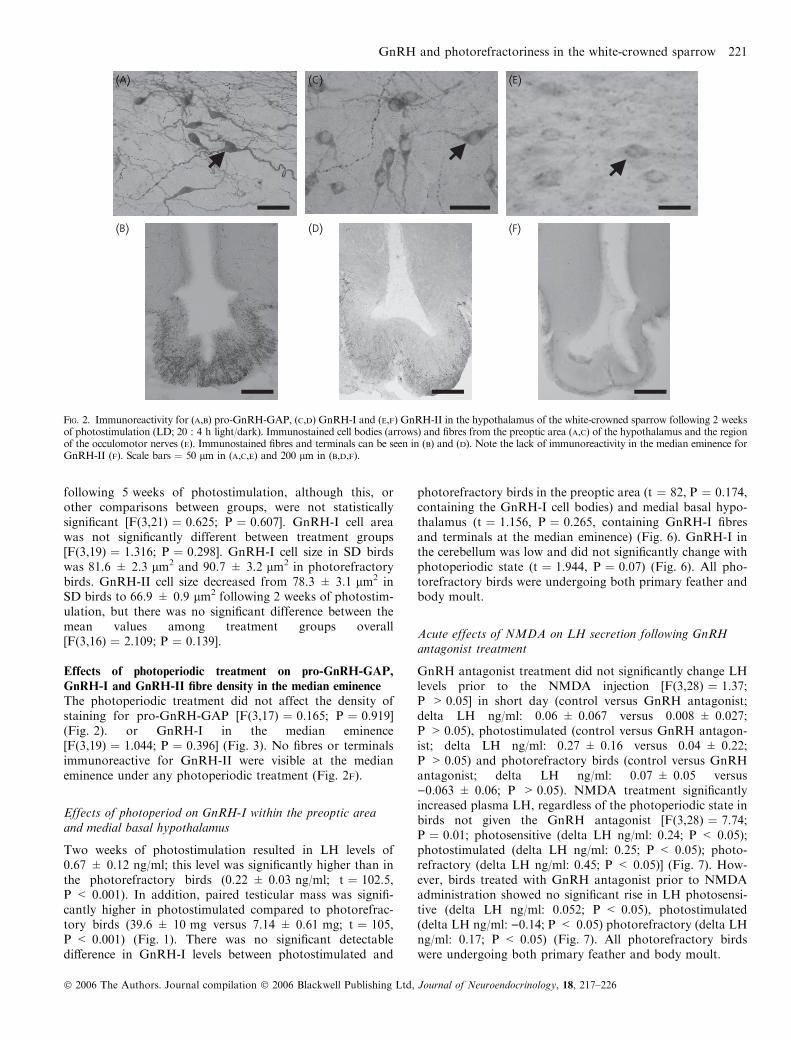
The distributions of GnRH and pro-GnRH-GAP immuno-reactivity were extremely similar and closely matched previ-ous immunocytochemistry studies in the European starling(9). Perikarya were distributed in bilateral bands extendingfrom the preoptic nucleus to septal regions of the brain andthe median eminence was densely innervated with fibresimmunopositive for both GnRH-I and pro-GnRH-GAP(Fig. 2).
GnRH-II containing cell bodies were localised in sectionsincluding and immediately caudal to the anterior com-missure (Fig. 2E) running ventrally and bilaterally fromthe third ventricle. A second population was observed inthe area of the occulomotor complex. No GnRH-IIimmunoreactive fibres were found at the median eminence(Fig. 2F).
Effects of photoperiodic treatment on pro-GnRH-GAP,
GnRH-I and GnRH-II cell number
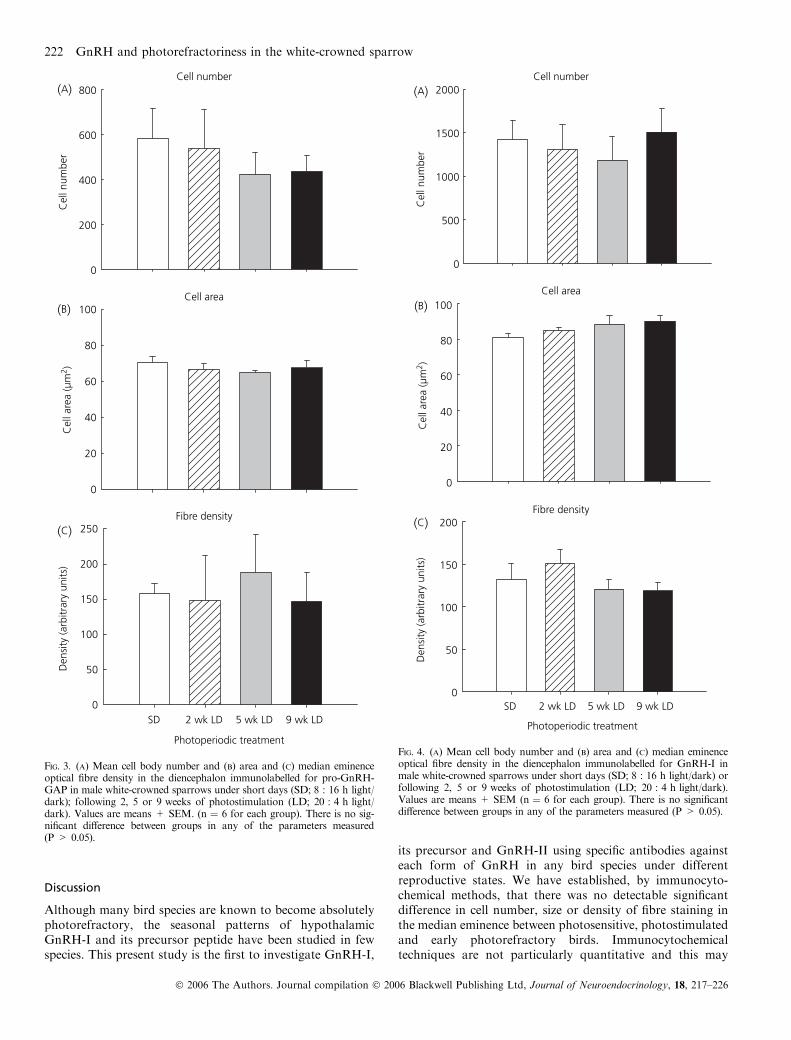
Photoperiod had no effect on the number of pro-GnRH-GAP, GnRH-I or GnRH-II positively labelled cells counted(Figs 3–5). The number of pro-GnRH-GAP cells was notsignificantly different between treatment groups[F(3,21) ¼ 0.411; P ¼ 0.747] and this was also true for thenumber of GnRH-I cells [F(3,20) ¼ 0.290; P ¼ 0.832] andGnRH-II cells per 50 lm section [F(3,11) ¼ 1.145;P ¼ 0.374].
Effects of photoperiodic treatment on pro-GnRH-GAP,
GnRH-I and GnRH-II cell size
Pro-GnRH-GAP, GnRH-I or GnRH-II cell size was notsignificantly altered by photoperiodic treatment (Figs 2–4).The mean size of pro-GnRH-GAP cells ranged from71.7 ± 2.8 lm2 in photosensitive birds to 65.6 ± 1.1 lm2
Weeks of photostimulation
0 2 5 9 12
Test
is m
ass
(mg)
1
3
10
30
100
300
Fig. 1. Paired testicular mass (mg; log scale) in male white-crowned spar-rows under short days (SD; 8 : 16 h light/dark) and following 2, 5, 9 or12 weeks of photostimulation (LD; 20 : 4 h light/dark). Each point repre-sents the mean ± SEM. Photostimulation significantly altered testicularmass (H ¼ 20.09; P < 0.001).
220 GnRH and photorefractoriness in the white-crowned sparrow
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226
following 5 weeks of photostimulation, although this, orother comparisons between groups, were not statisticallysignificant [F(3,21) ¼ 0.625; P ¼ 0.607]. GnRH-I cell areawas not significantly different between treatment groups[F(3,19) ¼ 1.316; P ¼ 0.298]. GnRH-I cell size in SD birdswas 81.6 ± 2.3 lm2 and 90.7 ± 3.2 lm2 in photorefractorybirds. GnRH-II cell size decreased from 78.3 ± 3.1 lm2 inSD birds to 66.9 ± 0.9 lm2 following 2 weeks of photostim-ulation, but there was no significant difference between themean values among treatment groups overall[F(3,16) ¼ 2.109; P ¼ 0.139].
Effects of photoperiodic treatment on pro-GnRH-GAP,
GnRH-I and GnRH-II fibre density in the median eminence
The photoperiodic treatment did not affect the density ofstaining for pro-GnRH-GAP [F(3,17) ¼ 0.165; P ¼ 0.919](Fig. 2). or GnRH-I in the median eminence[F(3,19) ¼ 1.044; P ¼ 0.396] (Fig. 3). No fibres or terminalsimmunoreactive for GnRH-II were visible at the medianeminence under any photoperiodic treatment (Fig. 2F).
Effects of photoperiod on GnRH-I within the preoptic areaand medial basal hypothalamus
Two weeks of photostimulation resulted in LH levels of0.67 ± 0.12 ng/ml; this level was significantly higher than inthe photorefractory birds (0.22 ± 0.03 ng/ml; t ¼ 102.5,P < 0.001). In addition, paired testicular mass was signifi-cantly higher in photostimulated compared to photorefrac-tory birds (39.6 ± 10 mg versus 7.14 ± 0.61 mg; t ¼ 105,P < 0.001) (Fig. 1). There was no significant detectabledifference in GnRH-I levels between photostimulated and
photorefractory birds in the preoptic area (t ¼ 82, P ¼ 0.174,containing the GnRH-I cell bodies) and medial basal hypo-thalamus (t ¼ 1.156, P ¼ 0.265, containing GnRH-I fibresand terminals at the median eminence) (Fig. 6). GnRH-I inthe cerebellum was low and did not significantly change withphotoperiodic state (t ¼ 1.944, P ¼ 0.07) (Fig. 6). All pho-torefractory birds were undergoing both primary feather andbody moult.
Acute effects of NMDA on LH secretion following GnRHantagonist treatment
GnRH antagonist treatment did not significantly change LHlevels prior to the NMDA injection [F(3,28) ¼ 1.37;P >0.05] in short day (control versus GnRH antagonist;delta LH ng/ml: 0.06 ± 0.067 versus 0.008 ± 0.027;P >0.05), photostimulated (control versus GnRH antagon-ist; delta LH ng/ml: 0.27 ± 0.16 versus 0.04 ± 0.22;P >0.05) and photorefractory birds (control versus GnRHantagonist; delta LH ng/ml: 0.07 ± 0.05 versus)0.063 ± 0.06; P >0.05). NMDA treatment significantlyincreased plasma LH, regardless of the photoperiodic state inbirds not given the GnRH antagonist [F(3,28) ¼ 7.74;P ¼ 0.01; photosensitive (delta LH ng/ml: 0.24; P < 0.05);photostimulated (delta LH ng/ml: 0.25; P < 0.05); photo-refractory (delta LH ng/ml: 0.45; P < 0.05)] (Fig. 7). How-ever, birds treated with GnRH antagonist prior to NMDAadministration showed no significant rise in LH photosensi-tive (delta LH ng/ml: 0.052; P < 0.05), photostimulated(delta LH ng/ml: )0.14; P < 0.05) photorefractory (delta LHng/ml: 0.17; P < 0.05) (Fig. 7). All photorefractory birdswere undergoing both primary feather and body moult.
(A)
(B)
(C)
(D)
(E)
(F)
Fig. 2. Immunoreactivity for (A,B) pro-GnRH-GAP, (C,D) GnRH-I and (E,F) GnRH-II in the hypothalamus of the white-crowned sparrow following 2 weeksof photostimulation (LD; 20 : 4 h light/dark). Immunostained cell bodies (arrows) and fibres from the preoptic area (A,C) of the hypothalamus and the regionof the occulomotor nerves (E). Immunostained fibres and terminals can be seen in (B) and (D). Note the lack of immunoreactivity in the median eminence forGnRH-II (F). Scale bars ¼ 50 lm in (A,C,E) and 200 lm in (B,D,F).
GnRH and photorefractoriness in the white-crowned sparrow 221
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226
Discussion
Although many bird species are known to become absolutelyphotorefractory, the seasonal patterns of hypothalamicGnRH-I and its precursor peptide have been studied in fewspecies. This present study is the first to investigate GnRH-I,
its precursor and GnRH-II using specific antibodies againsteach form of GnRH in any bird species under differentreproductive states. We have established, by immunocyto-chemical methods, that there was no detectable significantdifference in cell number, size or density of fibre staining inthe median eminence between photosensitive, photostimulatedand early photorefractory birds. Immunocytochemicaltechniques are not particularly quantitative and this may
Cell area
Cel
l are
a (µ
m2 )
0
20
40
60
80
100
Cell numberC
ell n
umbe
r
0
200
400
600
800(A)
(B)
(C)Fibre density
SD 2 wk LD 5 wk LD 9 wk LD
Den
sity
(arb
itrar
y un
its)
0
50
100
150
200
250
Photoperiodic treatment
Fig. 3. (A) Mean cell body number and (B) area and (C) median eminenceoptical fibre density in the diencephalon immunolabelled for pro-GnRH-GAP in male white-crowned sparrows under short days (SD; 8 : 16 h light/dark); following 2, 5 or 9 weeks of photostimulation (LD; 20 : 4 h light/dark). Values are means + SEM. (n ¼ 6 for each group). There is no sig-nificant difference between groups in any of the parameters measured(P > 0.05).
Cell area
Cel
l are
a (µ
m2 )
0
20
40
60
80
100
Cell number
Cel
l num
ber
0
500
1000
1500
2000
Fibre density
SD 2 wk LD 5 wk LD 9 wk LD
Den
sity
(arb
itrar
y un
its)
0
50
100
150
200
Photoperiodic treatment
(A)
(B)
(C)
Fig. 4. (A) Mean cell body number and (B) area and (C) median eminenceoptical fibre density in the diencephalon immunolabelled for GnRH-I inmale white-crowned sparrows under short days (SD; 8 : 16 h light/dark) orfollowing 2, 5 or 9 weeks of photostimulation (LD; 20 : 4 h light/dark).Values are means + SEM (n ¼ 6 for each group). There is no significantdifference between groups in any of the parameters measured (P > 0.05).
222 GnRH and photorefractoriness in the white-crowned sparrow
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

account for the negative results obtained across measures.However, our findings were further substantiated by a specificGnRH-I RIA that confirmed no significant difference inGnRH-I hypothalamic content between photostimulated andphotorefractory birds. GnRH-I has an established reproduc-tive role in Gambel’s white-crowned sparrow where it elicitsLH release (12, 39) and, as we found in the present study, it islocated in terminals in the median eminence.We investigated whether a termination in GnRH-I gene
expression is the first step in the process of photorefractori-ness in Gambel’s white-crowned sparrow by examiningPro-GnRH-GAP immunoreactivity. The morphology anddistribution of pro-GnRH-GAP neurones were indistinguish-able from those of GnRH-I and similar to that observed inthe European starling (8, 9, 16). However, in contrast to thestarling, there was no significant decrease in pro-GnRH-GAPimmunoreactivity during gonadal regression (9). In starlings,GnRH-I precursor synthesis decreases before GnRH levelsdecline, suggesting that inhibition of GnRH gene expressionis the initial step in the onset of photorefractoriness.
Interestingly, it was noted that cell number was much lowerfor pro-GnRH-GAP (approximately 500) compared toGnRH-I (approximately 1200) (Figs 3 and 4). This is similarto findings in the rat (21) and is probably due to rapidprocessing of pro-GnRH-GAP to GnRH.
The immunohistochemistry results of GnRH-I and itsprecursor pro-GnRH-GAP suggest several possible mecha-nisms for the regulation of GnRH-I secretion in Gambel’swhite-crowned sparrow. During photostimulation, we canassume that there is a concomitant increase in GnRH-Isynthesis with GnRH secretion because there was no obviousincrease in hypothalamic GnRH-I immunoreactivity at thistime. Previous studies in European starlings (4, 7, 8, 16) havedemonstrated a significant increase in GnRH immunoreac-tivity throughout photostimulation, suggesting that synthesisexceeds release in this species. We may interpret our findingsof an absence of changes in GnRH-I in Gamble’s white-crowned sparrow as a consequence of high to maximalGnRH-I level maintenance from sensitivity through tophotorefractoriness. Thus, the process of photostimulationrelies on GnRH-I release and may therefore depend on theremoval of a factor that actively inhibits GnRH-I secretionduring photorefractoriness. Such endogenous factors couldbe opioid peptides because they have been shown to exert aninhibitory influence on GnRH release in the rat (40) and mayalso do so in birds (41; but see also 42).
During gonadal regression, we could not detect any changein the number of GnRH-I labelled cells or density of GnRH-Iimmunoreactivity at the median eminence, providing supportfor an interpretation of a decrease in GnRH-I releaseassociated with a reduction of GnRH-I synthesis. However,to explain our findings of no decreases in pro-GnRH-GAPcell number or size during photorefractoriness onset, weshould consider that other regulatory factors, such as post-translational processing, may play a role in regulating GnRHand its precursor as in the rat (43). Immunocytochemicalstudies in the European starling, house sparrow (Passer
Cell numberC
ell n
umbe
r / 5
0 µm
sec
tion
0
20
40
60
Cell area
SD 2 wk LD 5 wk LD 9 wk LD
Cel
l are
a (µ
m2 )
0
20
40
60
80
100
Photoperiodic treatment
(A)
(B)
Fig. 5. (A) Mean cell body number (B) and area in the diencephalon im-munolabelled for GnRH-II in male white-crowned sparrows under shortdays (SD; 8 : 16 h light/dark); following 2, 5 or 9 weeks of photostimulation(LD; 20 : 4 h light/dark). Values are means + SEM (n ¼ 6 for eachgroup). There is no significant difference between groups in any of theparameters measured (P > 0.05).
Brain region
GnR
H-I
(ng/
ml b
rain
reg
ion)
0
10
20
30
40
50 Photostimulated
Photorefractory
MBHPOA Cerebellum
Fig. 6. Chicken GnRH-I levels in the preoptic area (POA), medial basalhypothalamus (MBH) and cerebellum of photostimulated (n ¼ 7) andphotorefractory (n ¼ 11) white-crowned sparrows as measured by radio-immunoassay. There is no significant difference in levels of GnRH-I betweenthe two groups in any brain region studied (P > 0.05). Values are means+ SEM
GnRH and photorefractoriness in the white-crowned sparrow 223
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

domesticus) and dark-eyed junco (Junco hyemalis) havedemonstrated a reduction in immunoreactive GnRH cellnumber and size in fully photorefractory compared tophotosensitive birds (9, 16, 44, 45). Further work hasindicated that, at least in starlings, the recovery of photosen-sitivity is gradual, beginning with renewed GnRH-I synthesisin the preoptic area of the hypothalamus (7). Nonetheless, ifphotorefractoriness is a consequence of inhibition of GnRH-Isynthesis, then a decrease in hypothalamic GnRH-I should
precede gonadal regression, and this is not the case (46).Therefore, we propose that, at least in white-crownedsparrows and starlings, a decrease in GnRH-I secretion isthe trigger for gonadal regression and a decrease in GnRHsynthesis then follows.
In other species, such as the white-winged crossbill (Loxialeucoptera) and canary (Serinus canaria), GnRH cell number,size and fibre density do not change with reproductive orphotoperiodic condition (47, 48). Furthermore, photorefrac-toriness does not decrease hypothalamic GnRH-I levels in thechicken (49). This provides temporal reproductive flexibility;by this, we mean the capacity to respond to favourableenvironmental conditions. In the wild, the white-wingedcrossbill and canary instigate breeding opportunisticallyregardless of photoperiod (50, 51). This finding is emphasisedby field observations of re-nesting Gambel’s white-crownedsparrows in which photorefractoriness onset may be delayedby as much as 2 weeks (39). This suggests that hypothalamicGnRH is still available for release at this time and otherenvironmental factors other than photoperiod may prolongthe breeding season in this species. Age is another factor thatappears to determine the timing of transition into a photo-refractory stage and, again, the GnRH-I system appearsflexible. Free-living dark eyed juncos that are undergoingtheir first breeding season (SY) terminate breeding and moultearlier than birds that have bred in previous years (ASY).Interestingly, GnRH-I system plasticity remains as there areno significant differences in the GnRH-I system between SYand ASY birds at the end of the breeding season (52).
The present study has ascertained that the hypothalamus ofGambel’s white-crowned sparrow contains GnRH-II,although the question of its specific function is left unan-swered. Other studies in the chicken and turkey, using thesame antibody as we have used in the present study, have alsodocumented an absence of GnRH-II in the median eminence(28, 33). This suggests that GnRH-II is unlikely to be anendogenous secretagogue for LH or FSH and is not involvedin photoperiodic reproductive stimulation or photorefracto-riness. Instead, GnRH-II may be acting as a neurotransmitter(35) to facilitate reproductive behaviour as central infusion ofGnRH-II, but not GnRH-I, into the brain increased sexualbehaviour in female Gambel’s white-crowned sparrows (53)and in musk shrews (54). Nonetheless, GnRH-II is a potentreleaser of LH in birds in vitro and in vivo (12, 55) and wecannot ignore the studies that have visualised GnRH-II in themedian eminence of the Japanese quail and chicken (31, 32).Even so, if GnRH-II exists in the quail and chicken medianeminence, then its function here is yet to be identified.
The explanation for inhibited GnRH secretion duringphotorefractoriness is in dispute, although it is not caused byan increased sensitivity to sex steroid feedback (56). Recentstudies have proposed a possible cause of decreased GnRH-Isecretion during photorefractoriness. A peptide has beenisolated from the avian brain using high-performance liquidchromatography that can inhibit gonadotrophin release fromcultured quail pituitary glands in a dose-dependent manner(57). This peptide has been accordingly termed gonadotro-phin-inhibitory hormone (GnIH) and is located in parvocel-lular neurones of the paraventricular nucleus in the quail andwhite-crowned sparrow (58, 59). Although we are still
Plas
ma
LH (n
g/m
l)
0.0
0.2
0.4
0.6
0.8
1.0 Pre-treatmentNMDA
Photosensitive
*
Plas
ma
LH (n
g/m
l)
0.0
0.5
1.0
1.5
2.0*
Photostimulated
Plas
ma
LH (n
g/m
l)
0.0
0.5
1.0
1.5
Vehicle GnRHantagonist
Photorefractory
*
Fig. 7. Acute effect of a specific gonadotrophin-releasing hormone (GnRH)antagonist s.c. (40 mg/kg) or vehicle pretreatment on plasma luteinisinghormone (LH) concentration following a single s.c. N-methyl-D-aspartate(NMDA) injection (20 mg/kg) in white-crowned sparrows in three differentreproductive conditions. Blood samples were taken immediately prior toGnRH antagonist administration and 20 min following NMDA injection.NMDA was given 90 min following GnRH antagonist administration.Values are means + SEM (n ¼ 6 for each group). *P < 0.05; delta LHfollowing NMDA treatment: comparison between vehicle and GnRH an-tagonist treated sparrows.
224 GnRH and photorefractoriness in the white-crowned sparrow
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

awaiting functional data, GnIH may provide a mechanismwhereby gonadotrophin release is actively inhibited duringphotorefractoriness in the white-crowned sparrow.In a previous study, we proposed that absolute photore-
fractoriness in Gambel’s white-crowned sparrow is driven bya decrease in GnRH-I secretion rather than GnRH-I synthe-sis, because NMDA significantly elevated LH regardless ofphotoperiodic or reproductive condition (11). Here, we haveconfirmed that NMDA most probably exerts its effect directlyon GnRH neurones at a suprapituitary level, as the rise in LHwas completely abolished by a specific GnRH antagonist. Theavailable literature suggests that GnRH antagonists typicallytake a few hours to take effect (60), and this may explain whythere were no significant changes in LH 90 min followingGnRH administration regardless of photoperiodic treatment.These findings provide additional evidence to support thehypothesis that photorefractoriness onset is most likely to bebrought about by changes in GnRH secretion rather thansynthesis. It is unknown whether GnRH synthesis eventuallydecreases in Gambel’s white-crowned sparrow becausechronically photostimulated birds were not examined here.However, bioassays on hypothalamic extracts suggest thatGnRH content does eventually decline in photorefractorymale Gambel’s white-crowned sparrows (12). Further studieson long-term photorefractory Gambel’s white-crowned spar-rows are thus warranted to investigate whether there are keydifferences in the mechanism or dynamics of photorefracto-riness between species. Also, because most studies examiningGnRH in relation to photorefractoriness have been per-formed on birds under laboratory conditions, it is importantthat we consider the GnRH system in the natural environ-ment; this is of relevance because the neuroendocrine systemof wild birds may be dampened under laboratory conditions(61).In summary, the present study has established that the
photoperiodically induced annual breeding cycle of Gambel’swhite-crowned sparrow relies on a withdrawal of the inhibi-tion of GnRH-I secretion during photostimulation and areturn of this active GnRH-I secretion inhibition at the onsetof photorefractoriness. GnRH-II does not appear to beinvolved in the control of gonadal development in this speciesbecause, using a specific GnRH-II antibody, no fibres orterminals were detected in the median eminence.
Acknowledgements
We would like to thank Haruka Wada, Meta Landys, Guy Mackee, Anna
Kim, Eliot Brenowitz, Richard Talbot and Peter Wilson for providing
technical assistance. We would also like to thank the anonymous referees for
their valuable comments on the manuscript. This study was funded by NSF
(IBN-9631350 and OPP-9702594) to J.C.W. P.J.S. was supported by a
BBSRC core strategic grant.
Accepted 29 November 2005
References
1 Dawson A. Photoperiodic control of gonadotrophin-releasing hor-mone secretion in seasonally breeding birds. In: Rao P, Klewer P,eds. Neural Regulation in the Vertebrate Endocrine System. NewYork, New York: Academic/Plenum Publishers; 1999: 141–159.
2 Nicholls TJ, Goldsmith AR, Dawson A. Photorefractoriness in birdsand comparison with mammals. Physiol Rev 1988; 68: 133–176.
3 Sharp PJ. Strategies in avian breeding cycles. Anim Reprod Sci 1996;42: 505–513.
4 Dawson A, Follett BK, Goldsmith AR, Nicholls TJ. Hypothalamicgonadotrophin-releasing hormone and pituitary and plasma FSHand prolactin during photostimulation and photorefractoriness inintact and thyroidectomized starlings (Sturnus vulgaris). J Endocrinol1985; 105: 71–77.
5 Dawson A. Seasonal differences in the secretion of luteinising hor-mone and prolactin in response to N-methyl-DL-aspartate in star-lings (Sturnus vulgaris). J Neuroendocrinol 2005; 17: 105–110.
6 Dawson A, King VM, Bentley GE, Ball GF. Photoperiodic controlof seasonality in birds. J Biol Rhythms 2001; 16: 365–380.
7 Dawson A, Goldsmith AR. Changes in gonadotrophin-releasinghormone (GnRH-I) in the pre-optic area and median eminence ofstarlings (Sturnus vulgaris) during the recovery of photosensitivityand during photostimulation. J Repro Fert 1997; 111: 1–6.
8 GoldsmithAR, IvingsWE, Pearce-Kelly AS, ParryDM, PlowmanG,Nicholls TJ, Follett BK. Photoperiodic control of the development ofthe LHRH neurosecretory system of European starlings (Sturnusvulgaris) during puberty and the onset of photorefractoriness.J Endocrinol 1989; 122: 255–268.
9 Parry DM, Goldsmith AR, Millar RP, Glennie LM. Immunocyto-chemical localization of GnRH precursor in the hypothalamus ofEuropean starlings during sexual maturation and photorefractori-ness. J Neuroendocrinol 1997; 9: 235–243.
10 Meddle SL, Follett BK. Photoperiodically driven changes in Fosexpression within the basal tuberal hypothalamus and medianeminence of Japanese quail. J Neurosci 1992, 1997; 17: 8909–8918.
11 Meddle SL, Maney DL, Wingfield JC. Effects of N-Methyl-D-As-partate on luteinizing hormone release and fos-like immunoreactiv-ity in the male White-crowned sparrow (Zonotrichia leucophrysgambelii). Endocrinology 1999; 140: 5922–5928.
12 Wingfield JC, Farner DS. Endocrinology of reproduction in wildspecies. In: Farner DS, King JR, Parkes KC, eds. Avian Endo-crinology. Academic Press, New York 1993; 9: 164–293.
13 Baines E. Photoperiodic Control of Hypothalamic GnRH GeneExpression in Japanese Quail. PhD Thesis. Edinburgh: University ofEdinburgh; 2001.
14 Dunn IC, Sharp PJ. Photo-induction of hypothalamic gonadotro-phin releasing hormone-I mRNA in the domestic chicken: a role foroestrogen? J Neuroendocrinol 1999; 11: 371–375.
15 Parry DM, Goldsmith AR. Ultrastructural evidence for changes insynaptic input to the hypothalamic luteinizing hormone-releasinghormone neurons in photosensitive and photorefractory starlings.J Neuroendocrinol 1993; 5: 387–395.
16 Foster RG, Plowman G, Goldsmith AR, Follett BK. Immunohist-ochemical demonstration of marked changes in the LHRH system ofphotosensitive and photorefractory European starlings (Sturnusvulgaris). J Endocrinol 1987; 115: 211–220.
17 Dawson A, Talbot RT, Dunn IC, Sharp PJ. Changes in basal hy-pothalamic chicken gonadotrophin-releasing hormone-I and vaso-active intestinal polypeptide associated with a photo-induced cycle ingonadal maturation and prolactin secretion in intact and thyroi-dectomized starling (Sturnus vulgaris). J Neuroendocrinol 2002; 14:533–539.
18 Dunn IC, Beattie KK, Mane D, Sang HM, Talbot RT, Wilson PW,Sharp PJ. Regulation of chicken gonadotrophin-releasing hormone-ImRNA in incubating, nest-deprived and laying bantam hens. Neu-roendocrinology 1996; 63: 504–513.
19 Robinson JE, Follett BK. Photoperiodism in Japanese quail: thetermination of seasonal breeding by photorefractoriness. Proc RoySoc Lon B 1982; 215: 95–116.
20 Seeburg PH, Adelman JP. Characterization of cDNA for precursorof human luteinizing hormone releasing hormone. Nature 1984; 311:666–668.
21 Dutlow CM, Rachman J, Jacobs TW, Millar RP. Prepubertalincreases in gonadotrophin-releasing hormone mRNA, gonadotro-phin-releasing hormone precursor, and subsequent maturation ofprecursor processing in male rats. J Clin Invest 1992; 90: 2496–2501.
22 King JA, Millar RP. Structure of chicken hypothalamic luteinizinghormone releasing hormone I. Structural determination on partiallypurified material. J Biol Chem 1982; 257: 10722–10728.
GnRH and photorefractoriness in the white-crowned sparrow 225
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226

23 King JA, Millar RP. Structure of chicken hypothalamic luteinizinghormone releasing hormone II. Isolation and characterization. J BiolChem 1982; 257: 10729–10732.
24 Miyamoto K, Hasegawa Y, Nomura M, Igarashi M, Kangawa K,Matsuo H. Identification of the second gonadotrophin-releasinghormone in chicken hypothalamus: evidence that gonadotrophinsecretion is probably controlled by two distinct gonadotrophin-releasing hormones in avian species. Proc Natl Acad Sci USA 1984;81: 3874–3878.
25 Bentley GE, Moore IT, Sower SA, Wingfield JC. Evidence for anovel gonadotrophin-releasing hormone in hypothalamic and fore-brain areas in songbirds. Brain Behav Evol 2004; 63: 34–46.
26 Mikami S, Yamada S, Hasegawa Y, Miyamoto K. Localization ofavian LH-RH immunoreactive neurons in the hypothalamus of thedomestic fowl, Gallus domesticus and the Japanese quail, Coturnixcoturnix. Cell Tissue Res 1988; 251: 51–58.
27 Sterling RJ, Sharp PJ. The localisation of LH-RH neurones in thediencephalons of the domestic hen. Cell Tissue Res 1982; 222: 283–298.
28 Millam JR, Faris PL, Youngren OM, El Halawani ME, HartmanBK. Immunohistochemical localization of chicken gonadotrophin-releasing hormone-I and hormone-II (cGnRH-I and cGnRH-II) inturkey hen brain. J Comp Neurol 1993; 333: 68–82.
29 Millam JR, Ottinger MA, Craig-Veit CB, Fan Y, Chaiseha Y, ElHalawani ME. Multiple forms of GnRH are released from perifusedmedial basal hypothalamic area (MBH/POA) explants in birds. GenComp Endocrinol 1998; 111: 95–101.
30 Dunn IC, Millam JR. Gonadotrophin releasing hormone: forms andfunctions in birds. Poultry Avian Biol Rev 1998; 9: 61–85.
31 van Gils J, Absil P, Grauwels L, Moons L, Vandesande F, BalthazartJ. Distribution of luteinizing hormone-releasing hormones I and II(LHRH-I and -II) in the quail and chicken brain as demonstratedwith antibodies directed against synthetic peptides. J Comp Neurol1993; 334: 304–323.
32 Clerens S, D’Hondt E, Berghman LR, Vandesande F, Arckens L.Identification of cGnRH-II in the median eminence of JapaneseQuail (Coturnix coturnix japonica). Gen Comp Endocrinol 2003; 131:48–56.
33 Sharp PJ, Talbot RT, Main GM, Dunn IC, Fraser HM, HuskissonNS. Physiological roles of chicken LHRH-I and -II in the control ofgonadotrophin release in the domestic chicken. J Endocrinol 1990;124: 291–299.
34 Hattori A, Ishii S, Wada M. Effects of two kinds of chicken lutei-nising hormone-releasing hormone (LH-RH), mammalian LH-RH,and its analogs on the release of LH and FSH in Japanese quail andchickens. Gen Comp Endocrinol 1986; 64: 446–455.
35 Katz IA, Millar RP, King JA. Differential regional distribution andrelease of two forms of gonadotrophin-releasing hormone in thechicken brain. Peptides 1990; 11: 443–450.
36 Follett BK, Scanes CG, Cunningham FJ. A radioimmunoassay foravian luteinizing hormone. J Endocrinol 1972; 52: 359–378.
37 Follett BK, Farner DS, Mattocks PW. Luteinizing hormone in theplasma of White-crowned sparrows (Zonotrichia leucophrys gam-belli) during artificial photostimulation. Gen Comp Endocrinol 1975;26: 126–134.
38 Roberts JL, Dutlow CM, Jakubowski M, Blum M, Millar RP.Estradiol stimulates preoptic area-anterior hypothalamic proGnRH-GAP gene expression in ovariectomised rats. Mol Brain Res 1989; 6:127–134.
39 Wingfield JC, Crim JW, Mattocks PW, JrFarner DS. Responses ofphotosensitive and photorefractory male white-crowned sparrows(Zonotrichia leucophyrs gambelii) to synthetic mammalian luteinizinghormone releasing hormone (Syn-LHRH). Biol Reprod 1979; 21:
801–806.40 Bruni JF, Van Vugt DA, Marshall S, Meites J. Effects of naloxone,
morphine and methionine enkephalin on serum prolactin, luteinizinghormone, follicle stimulating hormone, thyroid stimulating hormoneand growth hormone. Life Sci 1977; 21: 461–466.
41 Stansfield SC, Cunningham FJ. Attenuation of endogenous opioidpeptide inhibition of [Gln8] luteinizing hormone-releasing hormonesecretion during sexual maturation in the cockerel. Endocrinology1988; 123: 787–794.
42 Lal P, Sharp PJ, Dunn IC, Talbot RT. Absences of an effect ofnaloxone, an opioid anatagonist, on luteinizing hormone release invivo and luteinizing hormone-releasing hormone I release in vitro inintact, castrated, and food restricted cockerels. Gen Comp Endocrinol1990; 77: 239–245.
43 Kelly MJ, Garrett J, Bosch MA, Roselli CE, Douglass J, AdelmanJP, Ronnekleiv OK. Effects of ovariectomy on GnRH mRNA,proGnRH and GnRH levels in the preoptic hypothalamus of thefemale rat. Neuroendocrinology 1989; 49: 88–97.
44 Hahn TP, Ball GF. Changes in brain GnRH associated with pho-torefractoriness in house sparrows (Passer domesticus). Gen CompEndocrinol 1995; 99: 349–363.
45 Deviche P, Saldanha CJ, Silver R. Changes in brain gonadotrophin-releasing hormone- and vasoactive intestinal polypeptide-like im-munoreactivity accompanying reestablishment of photosensitivity inmale dark-eyed juncos (Junco hyemalis). Gen Comp Endocrinol 2000;117: 8–19.
46 Dawson A. The effects of a single long photoperiod on induction anddissipation of reproductive photorefractoriness in European star-lings. Gen Comp Endocrinol 2001; 121: 316–324.
47 MacDougal-Shackleton SA, Deviche PJ, Crain RD, Ball GF, HahnTP. Seasonal changes in brain GnRH immunoreactivity and song-control nuclei volumes in an opportunistically breeding songbird.Brain Behav Evol 2001; 58: 38–48.
48 Bentley GE, Audage NC, Hanspal EK, Ball GF, Hahn TP. Photo-periodic response of the hypothalamo-pituitary-gonad axis in maleand female canaries, Serinus canaria. J Exp Zool 2003; 296: 143–151.
49 Sharp PJ, Dunn IC, Main GM, Sterling RJ, Talbot RT. Neuroen-docrine control of reduced persistence of egg laying in domestic hens:evidence for the development of photorefractoriness. J Reprod Fert1992; 94: 221–235.
50 Deviche P. Seasonal reproductive pattern of white-winged crossbillsin interior Alaska. J Field Ornithol 1997; 68: 613–621.
51 Leitner S, Van’t Hof TJ, Gahr M. Flexible reproduction in wildcanaries is independent of photoperiod. Gen Comp Endocrinol 2003;130: 102–108.
52 Meddle SL, Wingfield JC, Millar RP, Deviche PJ. Immunocyto-chemical localization of GnRH-I and its precursor in the hypotha-lamus of photorefractory male dark-eyed juncos (Junco hyemalis) ofdifferent year classes. Gen Comp Endocrinol 2005; Nov 5, Epubahead of print.
53 Maney DL, Richardson RD, Wingfield JC. Central administrationof chicken gonadotrophin-releasing hormone-II enhances courtshipbehavior in a female sparrow. Horm Behav 1997; 32: 11–18.
54 Temple JL, Millar RP, Rissman EF. An evolutionarily conservedform of gonadotrophin-releasing hormone coordinates energy andreproductive behavior. Endocrinology 2003; 144: 13–19.
55 Chou HF, Johnson AL, Williams JB. Luteinizing hormone releasingactivity of [Gln8]-LHRH and [His5,Trp7,Tyr8]-LHRH in the cock-erel, in vivo and in vitro. Life Sci 1985; 37: 2459–2465.
56 Wilson FE. Androgen feedback-dependent and -independent controlof photoinduced LH secretion in male tree sparrows (Spizellaarborea). J Endocrinol 1985; 105: 141–152.
57 Tsutsui K, Saigoh E, Ukena K, Teranishi H, Fujisawa Y, Kikuchi M,Ishii S, Sharp PJ. A novel avian hypothalamic peptide inhibitinggonadotrophin release.BiochemBiophysResCom 2000; 275: 661–667.
58 Ukena K, Ubuka T, Tsutsui K. Distribution of a novel avian gon-adotrophin-inhibitory hormone in the quail brain. Cell Tissue Res2003; 312: 73–79.
59 Osugi T, Ukena K, Bentley GE, O’Brien S, Moore IT, Wingfield JC,Tsutsui K. Gonadotrophin-inhibitory hormone in Gambel’s white-crowned sparrow (Zonotrichia leucophrys gambelii): cDNA identifi-cation, transcript localization and functional effects in laboratoryand field experiments. J Endocrinol 2004; 182: 33–42.
60 Murase M, Uemura T, Gao M, Inada M, Funabashi T, Hirahara F.GnRH antagonist-induced down-regulation of the mRNA expres-sion of pituitary receptors: comparisons with GnRH agonist effects.Endocr J 2005; 52: 131–137.
61 Romero LM, Wingfield JC. Alterations in hypothalamic-pituitary-adrenal function associated with captivity in Gambel’s white-crowned sparrows (Zonotrichia leucophrys gambelli). Comp BiochemPhysiol B 1999; 122: 13–20.
226 GnRH and photorefractoriness in the white-crowned sparrow
� 2006 The Authors. Journal compilation � 2006 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 18, 217–226




















