
Kashmir Flycatcher Pale Rock Sparrow Ernst Schäfer - Indian ...
Upload
independentCategory
view
2download
0
Journal of AnimalEcology 0888\57\ 517Ð526
Þ 0888 BritishEcological Society
517
Spatial and temporal variation in demography of a house
sparrow metapopulation
BERNT!ERIK S$THER$\ THOR HARALD RINGSBY$\OšYVIND BAKKE% and ERLING JOHAN SOLBERG&$Department of Zoology and %Department of Mathematical Sciences\ Norwegian University of Science andTechnology\ N!6380 Trondheim\ Norway^ and &Norwegian Institute for Nature Research\ Tungasletta 1\ N!6374Trondheim\ Norway
Summary
0[ In order to compare spatial and temporal variation in the demography of a meta!population of house sparrows living on four islands o} the coast of Northern Norway\we computed the asymptotic population growth rate "l# for three transitions betweenyears\ using a projection matrix model[1[ The mean growth rate of the metapopulation was positive "l�0=94#[2[ Large variation was found in space and time in the asymptotic growth rate[ Eventhough the standard deviations of the estimates were large\ l was signi_cantly greaterthan 0 on one island in 1 out of 2 years[ In contrast\ l signi_cantly less than 0 occurredon an island on _ve occasions[3[ Decomposition of the magnitude of the spatial and temporal components of l
showed that one year "0884# had a particularly negative impact on l[ In contrast\ oneisland had a great positive e}ect[4[ These di}erences in l were most closely related to variation in the juvenile survivalrate and in the fecundity rate\ which\ in turn\ was closely related to variation both inspace and time in the proportion of the eggs that produced a ~edgling[5[ As expected from the large spatio!temporal variation in l\ the sensitivities and theelasticities of l to variation in the di}erent elements in the projection!matrix alsodi}ered both in space and time[ When l was small\ it was most in~uenced by variationin adult survival rate[ In contrast\ for large l\ it was most sensitive to variation in therecruitment rate[6[ We suggest that conditions during the breeding season determine whether a popu!lation acts as a source or sink in this temperate small passerine metapopulation\ andthat the ratio of juveniles to adults can be used to characterize populations as sourcesor sinks[7[ These results demonstrate large variation in space and time in the demography ofa small passerine metapopulation[ These di}erences result from a combination ofregional e}ects on the whole metapopulation\ and factors that occur at one particularisland in a single year[
Key!words] demography\ elasticity\ house sparrow\ metapopulation\ populationgrowth rate\ sensitivity[
Journal of Animal Ecology "0888# 57\ 517Ð526
Introduction
The spatial structure of populations can be cat!egorized schematically into some main types "Har!rison + Taylor 0886#[ In the classical models of Levins"0858\ 0869#\ metapopulations are considered to com!prise a number of subpopulations living in patches of
equal quality with no local dynamics[ This simplemodel has been found to describe the spatial organ!ization of many populations quite well "see examplesin Hanski 0886#[ In particular\ such Levins!type mod!els have been successful in describing\ at least quali!tatively\ the dynamics of several spatially structuredbutter~y populations "Thomas + Hanski 0886#[ In

518
B[!E[ S%ther et al[
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
contrast to this classical metapopulation structure\environmental heterogeneity in space and time is likelyto a}ect population dynamics[ Pulliam "0877#described this type of spatial organization as a sourceÐsink system[ Here both the local and global dynamicsare a}ected by the population ~uctuations in someparticular patches where the growth rate is positive[These source populations form sources for emigrationof individuals to other patches\ which would go extinctin the absence of immigration[ This type of spatialmodel has been assumed to describe the spatial organ!ization of vertebrates better than the classical Levins!type metapopulation model "Pulliam + Danielson0880^ Harrison + Taylor 0886#[
Theoretical studies have suggested that di}erencesin the type of spatial variation in demography havegreat impact on both the local and global dynamicsof a metapopulation "see reviews in Noon + McKel!vey 0885 and Gyllenberg\ Hanski + Hastings 0886#[ Inthe idealized Levins!type of metapopulation "Levins0869#\ an equilibrium exists where the fraction ofoccupied patches is equal to the di}erence between themigration and extinction rate divided by the migrationrate\ assuming that each patch has the same prob!ability of being colonized[ However\ at the local scale\extinction is certain when time goes to in_nity[ Whenthese models were extended to include colonizationand extinction rates that were allowed to change withhabitat occupancy\ two equilibrium points wereidenti_ed "Hanski + Gyllenberg 0882#[ A stable equi!librium existed only when a relatively high proportionof the available patches was occupied[ This suggeststhat securing viable metapopulations requires that alarge proportion of the suitable habitat in the area isoccupied "Hanski\ Moilanen + Gyllenberg 0885#[ Thepresence of both an unstable and stable equilibriumpoint was recently con_rmed by Lande\ Engen +S%ther "0887# and S%ther\ Engen + Lande "0888# in_nite metapopulation models including local dynam!ics and stochastic e}ects when the migration rate perindividual was low[ However\ these analyses showedthat these equilibria were strongly a}ected by changesin the local dynamics[ Thus\ local di}erences in thedemography of the population may have largedynamical consequences at the global metapopulationlevel[
Very few studies have been able to provide dataon spatial variation in demography\ although somestudies have recorded spatial variation in particularlife history traits "e[g[ Dhondt et al[ 0889^ Blondel\Pradel + Lebreton 0881#[ However\ based on a studyof colour!ringed individuals on small islands\ Smithet al[ "0885# found strong support for a sourceÐsinktype of spatial structure "Pulliam 0877# in a song spar!row "Melospiza melodia# metapopulation[
This paper presents data on spatial and temporalvariation in several demographic variables in a meta!population of house sparrow "Passer domesticus# atfour islands o} the coast of northern Norway[ These
data are used to calculate the population growth ratel of these subpopulations by means of a projectionmatrix model "Caswell 0878# in order to compare howl varies in time and space in this metapopulation[ Inparticular\ we want to examine the presence of sourcepopulations\ i[e[ whether l is consistently greater than0 in some populations[
Methods
STUDY AREA
This study was conducted during the years 0882Ð86on four islands "Gj%ro�y\ Hestmanno�y\ Indre Kvaro�yand Ytre Kvaro�y\ see Fig[ 0# located o} the coast ofHelgeland in northern Norway "55>N\02>E#[ Theseislands are part of a larger archipelago including atotal of 06 islands where a large proportion of thehouse sparrows has been banded "for a map of thelocation of the islands\ see Ringsby 0887#[ The land!scape comprises a mixture of agricultural land\ heath!land and mountains[ The islands di}er in size from¼1 km1 "Indre Kvaro�y# to 02 km1 "Hestmanno�y#[ Onall islands the sparrows are living close to people\especially in association with cattle farms[ The climateis oceanic but although at the Arctic Circle\ the islandsare usually snow!free for much of the winter[
Although the house sparrow is considered as a rela!tively sedentary species "Summers!Smith 0877#\ thereis interchange of\ especially\ juveniles between theislands "A[ Altwegg et al[\ unpublished data#[ Everyyear\ at least 1Ð2 immigrants from neighbourhoodislands were recorded on each island[ Similarly\ somejuveniles also move away from their birth island tosettle[ Thus\ this justi_es the use of the concept meta!population as de_ned by Hanski + Simberlo} "0886#because the populations do not ~uctuate completelyindependently of each other[
DATA COLLECTION
Since the spring of 0882\ a large proportion of thestationary house sparrow population has been cap!
10 km
Arctic Circle
Ytre Kvarøy
Indre Kvarøy
Hestmannøy
Gjærøy
Fig[ 0[ Map of the study area[

529
Demography of ahouse sparrowmetapopulation
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
tured by intensive mist!netting in the period MayÐAugust and has been individually colour!ringed[
During the breeding season the reproductive suc!cess of these individuals "number of ~edglings pro!duced per season# was determined by inspecting thenests at regular intervals[ On the islands included inthe present study\ most nests were located inside barnsand cowsheds[ In our study area the house sparrowlays between one and three clutches during the breed!ing season\ which lasts from May to early August"Ringsby\ S%ther + Solberg 0887#[ When the nestlingswere about 3Ð4 days old\ they were individuallybanded with an aluminium ring[ The breeding successwas estimated as the number of nestlings close to~edging present at the last visit[ All ~edglings wereindividually colour!ringed[
The population size was censused by counting thesparrows just before the onset of breeding[ Becausethe house sparrows live in close contact with humansettlements\ the population estimates became quiteaccurate[ On three of the islands "Hestmanno�y\ YtreKvaro�y and Gj%ro�y# the proportion of ringed birdswas so large "×89)# and the capture e.ciency sohigh that the number of di}erent birds captured dur!ing the breeding season was assumed to give an evenbetter estimate of the population size than the censusdata[ However\ census data were used as populationestimates at Indre Kvaro�y\ where a lower proportionof individuals was banded "¼69)#[ The largest aver!age population size was found at Hestmanno�y\whereas the greatest population ~uctuations occurredat Indre Kvaro�y "Table 0#[
ESTIMATES OF MORTALITY RATES
In order to estimate the survival rate from data basedon colour!ringed individuals\ estimates were obtainedof the probability of recapturing a bird that was pre!sent in the population "Lebreton et al[ 0881#[ For eachbird a vector of _ve indicator variables for each yearfrom 0882 to 0886 were obtained[ The indicator was9 if the bird was not recorded "observed or captured#in a given year and 0 if it was[ In addition\ we recorded\if possible\ whether the bird was hatched that year[Individuals that dispersed from an island were re!
Table 0[ The annual variation in the estimated breedingpopulation size of the house sparrow at the di}erent islands"see locations in Fig[ 0#
Year
Island 0882 0883 0884 0885
Gj%ro�y 23 36 23 30Indre Kvaro�y 22 30 50 37Ytre Kvaro�y 26 22 27 14Hestmanno�y 64 67 71 57
corded as dead[ Immigrants were included among theadults[
Denoting the years 0\ 1\ 2\ 3 and 4\ the parametersestimated were juvenile pi and adult survival rate qi
from year i − 0 to i\ and recapture rate ri for the yearsi � 1\ 2\ 3[ For i � 4 "0886#\ survival and recapturerates were confounded because there is no way ofdiscriminating between death and failure to recapture[Instead we estimated the product of survival andrecapture rates p4 r4 and q4 r4 for juveniles and adults\respectively[
Estimation procedures for these kind of parametersare well known from the literature "e[g[ see Lebretonet al[ 0881#[ Assume that a bird was observed and alivein year k − 0[ Then there are three possible events foryear k and the subsequent years] "i# The bird wasrecorded "captured or observed# and alive in year k^"ii# the bird was not recorded in year k\ but was re!corded and alive in a subsequent year^ or "iii# the birdwas not recorded in year k or any subsequent year[Clearly\ in the _rst two cases the bird was alive\whereas in the third case either it was dead or it wasalive but remained unrecorded[
Let ak denote the likelihood "the probability of anobservation as a function of the parameters# for a birdfrom year k to year k ¦ 0\ both years inclusive\ giventhat it was alive in year k − 0[ Letting a5 � 0\ ak is thende_ned recursively by]
ak � 0 − qk ¦ qk"0 − rk#ak¦0 eqn 0a
if the bird was not recorded in year k or any sub!sequent year\
ak � qk"0 − rk#ak¦0 eqn 0b
if the bird was not recorded in year k but was recordedin a subsequent year\ or
ak � qkrkak¦0 eqn 0c
if the bird was recorded in year k\ for k � f ¦ 0\[ [ [\ 4\where f is the _rst year the bird was captured\ f � 0\1\ 2 or 3[ If the bird was hatched in year f\ qf¦0 shouldbe replaced by pf¦0[
Now af¦0 is the likelihood function for one bird"because it was alive at year f#\ and the total likelihoodis the product of the likelihoods for each bird[Maximum likelihood estimates were obtained byapplying the downhill simplex method "Press et al[0878# to the logarithm of the likelihood function[ Toforce the parameters into the interval ð9\0Ł\ they wererepresented as the inverse logit transformation ðtakingx to 0:"0 ¦ e−x#Ł of another number in the max!imization process[
The 00 parameters were estimated for each of thefour study islands[
ESTIMATION OF POPULATION GROWTH RATE
For a given year and island\ let B denote the numberof female "assuming an equal sex ratio# o}spring pro!

520
B[!E[ S%ther et al[
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
duced per female\ and p and q the survival rate ofjuveniles and adults\ respectively[ Denoting by N thepopulation size immediately before hatching\ theexpected number of ~edglings becomes BN[ Theexpected number of birds before hatching the nextyear is the sum of the number of surviving ~edglings\BpN\ and the number of surviving adults\ qN\ that is"Bp ¦ q#N[ Thus\ the growth rate is l � Bp ¦ q[ Thegrowth rate was estimated for years 0882Ð83\ 0883Ð84 and 0884Ð85[
The growth rate can also be seen as the dominanteigenvalue of a 1 × 1 projection matrix "Caswell0878#[ Let x denote the number of juveniles just beforehatching and y the number of older birds[ Then thenumber of juveniles and adults the following year areBpx ¦ Bpy and qx ¦ qy\ respectively[ This cor!responds to the projection matrix]
P � 0Bpq
Bpq 1 eqn 1
Its dominant eigenvalue is l � Bp ¦ q[Various methods of assessing errors in estimates
of di}erent demographic traits "including populationgrowth rates# have been proposed "e[g[ Lande 0877^Alvarez!Buylla + Slatkin 0882\ 0883#[ Because weconstructed a computer program to estimate the sur!vival and recapture rate "see above#\ it was easy torepeat the computations for bootstrap samples[ Inaddition\ this method avoids assuming any speci_cdistribution for the estimators[ For each island andyear\ following Efron + Tibshirani "0882#\ 199 boot!strap samples were made by drawing individuals "withreplacement# from the original data set until theyattained the same size as the original sample[
For each bootstrap sample a bootstrap replicate ofthe parameter estimate was obtained\ and the stan!dard deviation was estimated by the standardestimator\ zðS ui
1 − "S ui#1:199Ł:199\ where the ui|sare the bootstrap replications of a parameter[
DECOMPOSING TREATMENT EFFECTS
In order to analyse how the population growth rate isin~uenced by island and time "Horvitz\ Schemske +Caswell 0886#\ we express the population growth rateof island i at year t as]
l"it# � l"[[# ¦ bi ¦ gt ¦ "bg#it\ eqn 2
i � 0\ 1\ 2\ 3 and t � 0\ 1\ 2\ where l"[[# is the growthrate calculated from the grand mean of all the matr!ices\ and bi\ gt and "bg#it are island\ time and interactione}ects\ respectively[ Then]
bi � l"i[# − l"[[# eqn 3
gt � l"[t# − l"[[# eqn 4
"bg#it � l"it# − bi − gt − l"[[#\ eqn 5
where l"i[# and l"[t# are the growth rates calculated from
the mean over years of the matrices for island i\ andfrom the mean over islands of the matrices for year t\respectively[
STABLE AGE DISTRIBUTION\ SENSITIVITY AND
ELASTICITY
The stable age distribution\ that is the long!term pro!portion of individuals in each age class if the pro!jection matrix is constant\ is given by the eigenvectorthat corresponds to the eigenvalue l\
A � 0Bpq 1[ eqn 6
Denoting by ajk the element in row j and column kin the projection matrix P\ j\ k � 0\ 1\ the sensitivityof l to change in ajk is sjk � 1l:1ajk "Caswell 0878#[For our projection matrix\ the matrix of sensitivitiesis]
S �0
Bp ¦ q 0BpBp
qq1 eqn 7
The elasticity of l to a change in ajk "de Kroon et al[0875# is
1 ln l
1 ln ajk
�1 ln l
1l
1l
1ajk>1 ln ajk
1ajk
�0l
sjk>0ajk
�ajksjk
l[
For P\ the matrix of elasticities is]
E �0
"Bp ¦ q#1 0"Bp#1
BpqBpq
q1 1[ eqn 8
The sum of elasticities is 0\ which is a general result"Caswell 0878#[
We also consider sensitivities and elasticities tochanges in the parameters directly instead of indirectlythrough matrix entries] 1l:1B � p\ 1l:1p � B\1l:1q � 0\ 1 ln l:1 ln B � 1 ln l:1 ln p � Bp:l and1 ln l:1 ln q � q:l[
Results
Large temporal and spatial variation was found inseveral reproductive traits[ The number of ~edglingsproduced per pair per season "Table 1# di}ered sig!ni_cantly among islands "P ³ 9=9990# and years"P ³ 9=94#\ in addition to a signi_cant interactionbetween island and year "P ³ 9=904#[ This suggeststhat there is noncorrelated variation in fecundityamong years and islands "see also Ringsby et al[ 0887#[Similarly\ the mean number of lost eggs or ~edglingsper pair per season showed similar variation"P ³ 9=902\ P ³ 9=900 and P ³ 9=905 for island\ yearand island × year interaction\ respectively#[ Therewas no signi_cant "P × 9=0# spatial or temporal vari!ation either in mean clutch size\ in the number ofclutches laid or in the mean total number of eggs laidper season[ The variation within the metapopulation

521
Demography of ahouse sparrowmetapopulation
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
Table 1[ Annual variation in mean "2SD# reproductive success "number of ~edglings per female#\ the juvenile and adultsurvival rate in the four study populations "see Fig[ 0#[ The survival rates and standard deviations were estimated using themaximum likelihood estimation procedures described in the text[ The _gures in brackets represent the sample size of females
Island
Demographic variable Year Gj%ro�y Indre Kvaro�y Ytre Kvaro�y Hestmanno�y
Reproductive success 0882 1=778 2 0=505 "8# 5=222 2 1=776 "2# 2=253 2 1=831 "00# 2=931 2 1=921 "13#0883 1=603 2 1=088 "03# 4=999 2 9=843 "01# 4=599 2 1=646 "09# 1=999 2 1=966 "28#0884 1=952 2 1=937 "05# 2=764 2 1=920 "7# 1=556 2 1=311 "5# 1=715 2 0=569 "12#
Juvenile survival rate 0882 9=0318 2 9=9590 9=4677 2 9=0196 9=1485 2 9=9829 9=0793 2 9=93120883 9=0128 2 9=9407 9=2305 2 9=9762 9=1377 2 9=9431 9=1331 2 9=93460884 9=0205 2 9=9445 9=0993 2 9=9704 9=0189 2 9=9596 9=9658 2 9=9170
Adult survival rate 0882 9=5987 2 9=9714 9=6466 2 9=9783 9=4682 2 9=0993 9=5759 2 9=95720883 9=4285 2 9=9863 9=7144 2 9=9807 9=4131 2 9=0099 9=4268 2 9=95460884 9=4556 2 9=9743 9=4300 2 9=9734 9=4603 2 9=9733 9=3824 2 9=9596
in the number of ~edglings produced per year per pairwas therefore related to the proportion of the eggs ornestlings that was lost before ~edging[
There was large annual and spatial variation injuvenile survival rates\ ranging from 9=9658 atHestmanno�y in 0884 to 9=4677 at Indre Kvaro�y in0882 "Table 1#[ These di}erences may be related toannual and spatial variation in ~edgling size "Ringsbyet al[ 0887#[ In contrast\ less variation was found inthe adult survival rate[ Elsewhere "Ringsby et al[\ inpress#\ we found\ with a larger data set including eightislands and using SURGE "Lebreton et al[ 0881# thatboth island and an island × year interaction explaineda signi_cant proportion of the variance in the adultsurvival rate[ Similarly\ in a smaller subset of _veislands\ a similar analysis of the variation in juvenile
Fig[ 1[ Annual variation in the asymptotic population growth rate "l# estimated by the projection matrix model "equations 1\2# of the house sparrow populations at Gj%ro�y "ž#\ Indre Kvaro�y "Ž#\ Ytre Kvaro�y "R# and Hestmanno�y "T#[ The dottedline indicates l � 0[
survival rate showed a signi_cant e}ect of year and\again\ an island × year interaction[
The mean growth rate of the metapopulation waslÞ� 0=94[ However\ the di}erences among years andislands in the demographic traits "Table 1# generatedlarge variation in space and time in l "Fig[ 1#[ On oneisland "Indre Kvaro�y#\ l was signi_cantly greater than0 "P ³ 9=90# in 1 years[ In contrast\ l was signi_cantless than 0 during the two last years on Gj%ro�y andHestmanno�y "P ³ 9=94 in 0883 and P ³ 9=90 in 0884for both islands#\ and on Ytre Kvaro�y in 0884"P ³ 9=94#[
Decomposing the magnitude of the temporal andspatial components of the variation in lÞ "Fig[ 2a\b#\assuming a _xed e}ect of island and year using equa!tions 3\ 4\ 5\ showed that the year 0884 had the largest

522
B[!E[ S%ther et al[
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
Fig[ 2[ Deviations in the mean growth rate of the house spar!row metapopulation that is a result of "a# an island e}ect\and "b# a year e}ect\ assuming _xed e}ects of island and year"see equations 3\ 4\ 5\ 6 for details of estimation#[ The islandsare] 0\ Gj%ro�y^ 1\ Indre Kvaro�y^ 2\ Ytre Kvaro�y^ and 3\Hestmanno�y[
negative e}ect on lÞ "Fig[ 2b#[ In contrast\ the islandIndre Kvaro�y had the largest main positive _xed e}ect"Fig[ 2a#[ An examination of the magnitude of theisland!year interaction "Fig[ 3# showed that the tem!poral variation of the e}ects on lÞ di}ered among theislands[ At Indre Kvaro�y a particularly large l wasfound in 0882\ whereas 0884 had a negative e}ect ofapproximately the same magnitude "Fig[ 3#[ Incontrast\ at Gj%ro�y the positive e}ect on l increasedduring the study period because the values of all threedemographic variables remained relatively similaracross years on this island "Table 1#[
As expected from the large temporal variation in l
"Figs 1\ 2\ 3#\ standing age!distribution in most yearsdi}ered signi_cantly from that expected "as de_ned byequation 6# if the asymptotic growth rate was realized"Table 2#[ On Indre Kvaro�y\ the proportion of juv!eniles in the population was less than expected\
Fig[ 3[ Deviations in the mean growth rate of the house spar!row metapopulation that is due to an island × year inter!action\ assuming _xed e}ects of island and year "see equa!tions 2\ 3\ 4\ 5 for details of estimation#[
whereas on Ytre Kvaro�y and Hestmanno�y "in 0882and 0883# more juveniles were recorded than expected[
There was large temporal and spatial variation inthe elasticities ejk and sensitivities sjk of l to the di}er!ent matrix elements ajk "Table 3#[ On two islands"Gj%ro�y and Hestmanno�y#\ s00 "and s10# was less thans01 "and s11#[ Accordingly\ the largest elasticity on theseislands was e11[ Thus\ on this island variation in adultsurvival rate had the greatest impact on l[ On theother two islands\ large annual variation was foundin the di}erent elasticities and sensitivities^ forinstance\ on one island "Indre Kvaro�y#\ the con!tribution of e11 to l increased from 7=4) in 0882 to43=0) in 0884 "Table 3#[
These variations in the di}erent elasticities werestrongly related to variation in l[ The elasticity ofl to a change in adult survival "eq# increased withdecreasing l "Fig[ 4#[ In contrast\ the reverse trend wasrecorded for the elasticity of the fecundity rate "eB#where the highest elasticity was found in the popu!lations with largest l "Table 3#[
Discussion
This study demonstrates considerable variationamong the subpopulations\ both in space and timein the asymptotic population growth rate within thismetapopulation of house sparrow[
Even though the sample sizes often were small"Table 1#\ resulting in large standard deviations in theestimates\ l was in several cases signi_cantly di}erentfrom 0 "Fig[ 1#[ On one of the islands "Gj%ro�y# l ³ 0

523
Demography of ahouse sparrowmetapopulation
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
Table 2[ Spatial and temporal variation in the observed and expected "see text# age!distribution in the house sparrow populationjust before onset of the breeding season[ P denotes the signi_cance of the deviation from the expected age distribution\ aspredicted from equation 6[ Juv � birds that were born the previous breeding season\ and remained on the island of birth\whereas Ad � birds older than 0 year
Year
0882 0883 0884Age
Island distribution Juv Ad P Juv Ad P Juv Ad P
Gj%ro�y Observed 04 08 ³9=90 7 08 ×9=94 7 13 ×9=94Expected 7=4 14=4 5=4 19=4 5=0 14=8
Indre Kvaro�y Observed 8 03 ³9=90 09 18 ³9=90 2 21 ³9=90Expected 05=2 5=6 08=8 08=0 8=0 14=8
Ytre Kvaro�y Observed 02 06 ³9=90 06 06 ³9=990 5 03 ³9=90Expected 5=9 13=9 5=9 17=9 1=1 06=7
Hestmanno�y Observed 08 13 ³9=94 18 27 ³9=94 7 34 ×9=94Expected 01=4 29=4 19=7 35=1 8=4 32=4
Table 3[ Spatial and temporal variation in the elasticities "eij# and sensitivities "sij# of the growth rate l of the house sparrowpopulations to a change in the element ajk in the 1 × 1 Leslie!matrix A "see de_nition in the text#\ where j represents the rowand k represents the column[ eB and eq are the elasticity of the population growth rate to a change in the fecundity rate andthe adult survival rate\ respectively
Island
Variable Year Gj%ro�y Indre Kvaro�y Ytre Kvaro�y Hestmanno�y
s00 � s10 � eB 0882 9=1418 9=6964 9=3187 9=17460883 9=1265 9=4974 9=4695 9=20110884 9=0821 9=1533 9=1203 9=0793
s01 � s11 � eq 0882 9=6360 9=1814 9=4691 9=60320883 9=6513 9=3804 9=3183 9=57670884 9=7957 9=6245 9=6575 9=7085
e00 0882 9=9539 9=4995 9=0736 9=97050883 9=9453 9=1475 9=2145 9=98640884 9=9262 9=9588 9=9424 9=9215
e01 � e10 0882 9=0778 9=1958 9=1340 9=19300883 9=0700 9=1388 9=1349 9=10360884 9=0448 9=0834 9=0667 9=0368
e11 0882 9=4471 9=9744 9=2141 9=40920883 9=4702 9=1305 9=0733 9=36290884 9=5498 9=4300 9=4897 9=5606
in all 2 years\ suggesting that this population could\according to Pulliam "0877#\ be considered as a sink\dependent on immigration to persist[ However\ ouranalyses showed that none of the islands could func!tion as permanent sources for this colonizationbecause l ³ 0 in all populations in the last study year"Figs 1\ 3#[ Because many values of l were close to 0"Fig[ 1#\ the status of the populations is likely tochange from year to year[ Unfortunately\ because theinterchange of both juvenile and adult house sparrowsamong the islands was small\ the sample sizes arecurrently too small to examine whether islands withl ³ 0 actually had a higher immigration rate of indi!viduals from islands in the neighbourhood than didislands with l × 0[ Analyses of the dispersal patternamong 03 islands\ including the four study islands\
showed large annual variation in dispersal rates\ inde!pendent of spatial variation in l "A[ Altwegg et al[\unpublished data#[ Other passerine studies\ however\have documented that immigration rates in some yearmay be so large that rapid population declines arecounteracted "Stacey + Taper 0881^ Smith et al[ 0885#[
In principle\ the asymptotic growth rate l associ!ated with the matrix in equation 0 can only be used topredict future population size in a constant environ!ment[ The large deviations "Table 2# of the recordedand stable age distribution "i[e[ the steady state pro!portion of individuals in each age!class that occurswhen the asymptotic growth rate is realized if theprojection matrix is constant# suggest l will not beparticularly useful for predicting annual variation inthe size of the house sparrow populations at these
524
B[!E[ S%ther et al[
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
Fig[ 4[ The elasticity of the population growth rate l of thehouse sparrow populations to a change in adult survival rate"eq# in relation to l[ The symbols denote the di}erent islands]Gj%ro�y "ž#\ Indre Kvaro�y "Ž#\ Ytre Kvaro�y "R# andHestmanno�y "T#[
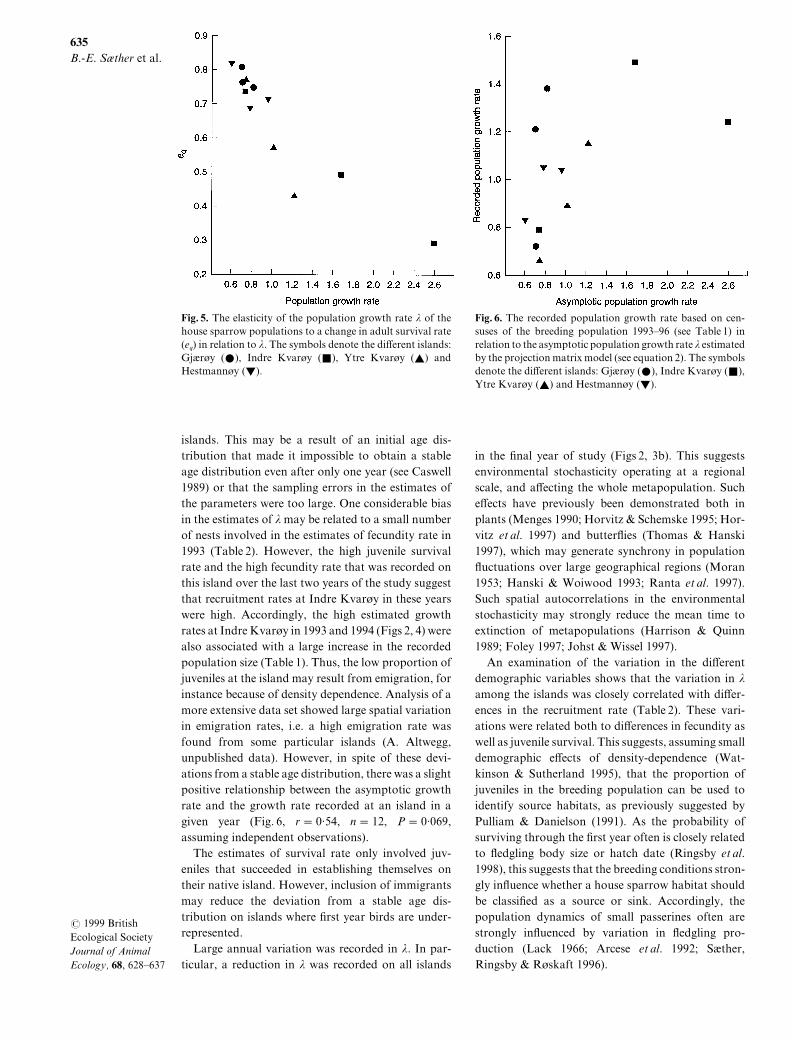
islands[ This may be a result of an initial age dis!tribution that made it impossible to obtain a stableage distribution even after only one year "see Caswell0878# or that the sampling errors in the estimates ofthe parameters were too large[ One considerable biasin the estimates of l may be related to a small numberof nests involved in the estimates of fecundity rate in0882 "Table 1#[ However\ the high juvenile survivalrate and the high fecundity rate that was recorded onthis island over the last two years of the study suggestthat recruitment rates at Indre Kvaro�y in these yearswere high[ Accordingly\ the high estimated growthrates at Indre Kvaro�y in 0882 and 0883 "Figs 1\ 3# werealso associated with a large increase in the recordedpopulation size "Table 0#[ Thus\ the low proportion ofjuveniles at the island may result from emigration\ forinstance because of density dependence[ Analysis of amore extensive data set showed large spatial variationin emigration rates\ i[e[ a high emigration rate wasfound from some particular islands "A[ Altwegg\unpublished data#[ However\ in spite of these devi!ations from a stable age distribution\ there was a slightpositive relationship between the asymptotic growthrate and the growth rate recorded at an island in agiven year "Fig[ 5\ r � 9=43\ n � 01\ P � 9=958\assuming independent observations#[
The estimates of survival rate only involved juv!eniles that succeeded in establishing themselves ontheir native island[ However\ inclusion of immigrantsmay reduce the deviation from a stable age dis!tribution on islands where _rst year birds are under!represented[
Large annual variation was recorded in l[ In par!ticular\ a reduction in l was recorded on all islands
Fig[ 5[ The recorded population growth rate based on cen!suses of the breeding population 0882Ð85 "see Table 0# inrelation to the asymptotic population growth rate l estimatedby the projection matrix model "see equation 1#[ The symbolsdenote the di}erent islands] Gj%ro�y "ž#\ Indre Kvaro�y "Ž#\Ytre Kvaro�y "R# and Hestmanno�y "T#[
in the _nal year of study "Figs 1\ 2b#[ This suggestsenvironmental stochasticity operating at a regionalscale\ and a}ecting the whole metapopulation[ Suche}ects have previously been demonstrated both inplants "Menges 0889^ Horvitz + Schemske 0884^ Hor!vitz et al[ 0886# and butter~ies "Thomas + Hanski0886#\ which may generate synchrony in population~uctuations over large geographical regions "Moran0842^ Hanski + Woiwood 0882^ Ranta et al[ 0886#[Such spatial autocorrelations in the environmentalstochasticity may strongly reduce the mean time toextinction of metapopulations "Harrison + Quinn0878^ Foley 0886^ Johst + Wissel 0886#[
An examination of the variation in the di}erentdemographic variables shows that the variation in l
among the islands was closely correlated with di}er!ences in the recruitment rate "Table 1#[ These vari!ations were related both to di}erences in fecundity aswell as juvenile survival[ This suggests\ assuming smalldemographic e}ects of density!dependence "Wat!kinson + Sutherland 0884#\ that the proportion ofjuveniles in the breeding population can be used toidentify source habitats\ as previously suggested byPulliam + Danielson "0880#[ As the probability ofsurviving through the _rst year often is closely relatedto ~edgling body size or hatch date "Ringsby et al[0887#\ this suggests that the breeding conditions stron!gly in~uence whether a house sparrow habitat shouldbe classi_ed as a source or sink[ Accordingly\ thepopulation dynamics of small passerines often arestrongly in~uenced by variation in ~edgling pro!duction "Lack 0855^ Arcese et al[ 0881^ S%ther\Ringsby + Ro�skaft 0885#[

525
Demography of ahouse sparrowmetapopulation
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
Spatial and temporal variation in the recruitmentrate can either result from a ~uctuating environmentor the presence of density dependence\ or result froman interaction between these two mechanisms[ Cur!rently we do not have data to analyse their e}ectson recruitment\ because estimates of environmentalstochasticity may require long!term "×09 years# dataseries "S%ther et al[ 0887#[ However\ _rst!year survivalis closely related both to ~edgling body size and hatchdate "Ringsby et al[ 0887#[ Furthermore\ the pro!portion of eggs lost during the breeding period a}ectsthe fecundity rate "Table 1#[ Studies of other pass!erines have shown that both temporal "Slagsvold 0865^van Noordwijk\ McCleery + Perrins 0884# and spatialvariation in the date of egg laying "Nager + vanNoordwijk 0884# may be related to di}erences in spr!ing temperature[ This suggests that climatic con!ditions during the breeding season a}ect the recruit!ment rate in these populations[ However\ the decreasein l at Indre Kvaro�y during the study period "Figs 1\3# was associated with a large increase in populationsize "Table 0#\ which suggests a density!dependentreduction in l[
Whatever the proximate mechanisms\ the analysesof elasticities and sensitivities "Table 3# suggest thate}ect on l of a perturbation of an element ajk willdepend on whether the subpopulation is a source orsink[ When l ³ 0\ the recruitment rate is small and achange in adult survival rate will have a greater e}ectthan in source populations[ Thus\ the viability of suchsink populations will be strongly dependent on vari!ation in the adult survival rate "Fig[ 4#[ In contrast\ athigh growth rates\ variation in fecundity or _rst!yearsurvival contributed more to l[ This suggests thatestimating the number of juveniles per adult may bea general approach to identifying source and sinkhabitats\ at least in such relatively short!lived species[
Acknowledgements
This study was _nanced by grants from the ResearchCouncil of Norway and the Norwegian Directoratefor Nature Management[ We are grateful to R[Altwegg\ T[ Berge\ S[ Krogstad\ N[M[ Pedersen\ K[Solbakken\ H[ Sta}\ B[ Staven and I[ Stewart for helpwith the _eldwork[ The comments from J[N[M Smithand E[ Ranta greatly improved the manuscript[
References
Alvarez!Buylla\ E[R[ + Slatkin\ M[ "0882# Finding con!_dence limits on population growth rates] Monte Carlotest of a simple analytic method[ Oikos\ 57\ 162Ð171[
Alvarez!Buylla\ E[R[ + Slatkin\ M[ "0883# Finding con!_dence limits on population growth rates] three real exam!ples revisited[ Ecology\ 64\ 144Ð159[
Arcese\ P[\ Smith\ J[N[M[\ Hochachka\ W[M[\ Rogers\ C[M[+ Ludwig\ D[ "0881# Stability\ regulation\ and the deter!mination of abundance in an insular Song Sparrow popu!lation[ Ecology\ 62\ 794Ð711[
Blondel\ J[\ Pradel\ R[ + Lebreton\ J[!D[ "0881# Low fec!undity insular blue tits do not survive better as adults thanhigh fecundity mainland ones[ Journal of Animal Ecology\50\ 194Ð102[
Caswell\ H[ "0878# Matrix Population Models[ SinauerAssociates\ Sunderland\ MA[
Dhondt\ A[A[\ Adriansen\ F[\ Matthysen\ E[ + Kempenaers\B[ "0889# Nonadaptive clutch sizes in tits[ Nature\ 237\612Ð614[
Efron\ B[ + Tibshirani\ R[J[ "0882# An Introduction to theBootstrap[ Chapman + Hall\ New York[
Foley\ P[ "0886# Extinction models for local populations[Metapopulation Biology "eds I[ A[ Hanski + M[ E[ Gilpin#\pp[ 104Ð135[ Academic Press\ San Diego[
Gyllenberg\ M[\ Hanski\ I[A[ + Hastings\ A[ "0886# Struc!tured metapopulation models Metapopulation Biology "edsI[ A[ Hanski + M[ E[ Gilpin#\ pp[ 82Ð011[ Academic Press\San Diego[
Hanski\ I[A[ "0886# Metapopulation dynamics] from con!cepts and observations to predictive models[ Meta!population Biology "eds I[ A[ Hanski + M[ E[ Gilpin#\ pp[58Ð80[ Academic Press\ San Diego[
Hanski\ I[A[ + Gyllenberg\ M[ "0882# Two general meta!population models and the core!satellite hypothesis[ Amer!ican Naturalist\ 031\ 06Ð30[
Hanski\ I[A[ + Simberlo}\ D[ "0886# The metapopulationapproach\ its history\ conceptual domain\ and applicationto conservation[ Metapopulation Biology "eds I[ A[ Hanski+ M[ E[ Gilpin#\ pp[ 4Ð15[ Academic Press\ San Diego[
Hanski\ I[A[ + Woiwood\ I[P[ "0882# Spatial synchrony inthe dynamics of moth and aphid populations[ Journal ofAnimal Ecology\ 51\ 545Ð577[
Hanski\ I[A[\ Moilanen\ A[ + Gyllenberg\ M[ "0885# Mini!mum viable metapopulation size[ American Naturalist\036\ 416Ð430[
Harrison\ S[ + Quinn\ J[F[ "0878# Correlated environmentsand the persistence of metapopulations[ Oikos\ 45\ 182Ð187[
Harrison\ S[ + Taylor\ A[D[ "0886# Metapopulation dynam!ics[ Metapopulation Biology "eds I[ A[ Hanski + M[ E[Gilpin#\ pp[ 16Ð31[ Academic Press\ San Diego[
Horvitz\ C[C[ + Schemske\ D[W[ "0884# Spatiotemporalvariation in demographic transitions of a tropical under!storey herb] projection matrix analysis[ Ecological Mono!graphs\ 54\ 044Ð081[
Horvitz\ C[C[\ Schemske\ D[W[ + Caswell\ H[ "0886# Therelative {importance| of life!history stages to populationgrowth] prospective and retrospective analyses Structured!Population Models in Marine\ Terrestrial and FreshwaterSystems "eds S[ Tuljapurkar + H[ Caswell#\ pp[ 136Ð160[Chapman + Hall\ New York[
Johst\ K[ + Wissel\ C[ "0886# Extinction risk in a temporallycorrelated ~uctuating environment[ Theoretical PopulationBiology\ 41\ 80Ð099[
de Kroon\ H[J[\ Plaiser\ A[\ van Groenendael\ J[ + Caswell\H[ "0875# Elasticity] The relative contribution of demo!graphic parameters to population growth rate[ Ecology\56\ 0316Ð0320[
Lack\ D[ "0855# Population Studies of Birds[ Clarendon Press\Oxford[
Lande\ R[ "0877# Demographic models of the northern spot!ted owl "Strix occidental caurina#[ Oecologia\ 64\ 590Ð596[
Lande\ R[\ Engen\ S[ + S%ther\ B[!E[ "0887# Extinction timesin _nite metapopulation models with stochastic localdynamics[ Oikos\ 72\ 272Ð278[
Lebreton\ J[!D[\ Burnham\ K[P[\ Clobert\ J[ + Anderson\D[R[ "0881# Modeling survival and testing biologicalhypotheses using marked animals] a uni_ed approach withcase studies[ Ecological Monographs\ 51\ 56Ð007[
Levins\ R[ "0858# Some demographic and genetic conse!quences of environmental heterogeneity for biological con!

526
B[!E[ S%ther et al[
Þ 0888 BritishEcological SocietyJournal of AnimalEcology\ 57\ 517Ð526
trol[ Bulletin of the Entomological Society of America\ 04\126Ð139[
Levins\ R[ "0869# Extinction[ Some Mathematical Problemsin Biology "ed[ M[ Gerstenhaber#\ pp[ 64Ð096[ AmericanMathematical Society\ Providence\ RI[
Menges\ E[S[ "0889# Population viability analysis for anendangered plant[ Conservation Biology\ 3\ 41Ð51[
Moran\ P[A[P[ "0842# The statistical analysis of the Canadianlynx cycle[ II[ Synchronization and meteorology[ Aus!tralian Journal of Zoology\ 0\ 180Ð187[
Nager\ R[G[ + van Noordwijk\ A[J[ "0884# Proximate andultimate aspects of phenotypic plasticity in timing of greattit breeding in a heterogeneous environment[ AmericanNaturalist\ 035\ 343Ð363[
Noon\ B[R[ + McKelvey\ K[S[ "0885# A common frameworkfor conservation planning] linking individual and meta!population models[ Metapopulations and Wildlife Con!servation "ed[ D[ R[ McCullough#\ pp[ 028Ð054[ IslandPress\ Washington[
van Noordwijk\ A[J[\ McCleery\ R[H[ + Perrins\ C[M[ "0884#Selection for the timing of great tit breeding in relationto caterpillar growth and temperature[ Journal of AnimalEcology\ 53\ 340Ð347[
Press\ W[H[\ Flannery\ B[P[\ Teukolsky\ S[A[ + Vetterling\W[T[ "0878# Numerical recipes in Pascal[ Cambridge Uni!versity Press\ Cambridge[
Pulliam\ H[R[ "0877# Sources\ sinks\ and population regu!lation[ American Naturalist\ 021\ 541Ð550[
Pulliam\ H[R[ + Danielson\ B[J[ "0880# Sources\ sinks\ andhabitat selection] a landscape perspective on populationdynamics[ American Naturalist\ 026\ S49ÐS55[
Ranta\ E[\ Kaitala\ V[\ Lindstro�m\ J[ + Helle\ E[ "0886#Moran e}ect and synchrony in population dynamics[Oikos\ 67\ 025Ð031[
Ringsby\ T[H[ "0887# Variation in space and time] the biologyof a house sparrow metapopulation[ Dr[ scient[ thesis\NTNU\ Trondheim[
Ringsby\ T[H[\ S%ther\ B[!E[ + Solberg\ E[J[ "0887# Factorsa}ecting juvenile survival in House Sparrow[ Journal ofAvian Biology 18\ 130Ð136[
Ringsby\ T[H[\ S%ther\ B[!E[\ Altwegg\ R[ + Solberg\ E[J["0888# Temporal and spatial variation in survival ratesof a house sparrow\ Passer domesticus\ metapopulation[Oikos\ 74\ in press[
S%ther\ B[!E[\ Ringsby\ T[H[ + Ro�skaft\ E[ "0885# Life his!tory variation\ population processes and priorities in spec!ies conservation] towards a reunion of research paradigms[Oikos\ 66\ 106Ð115[
S%ther\ B[!E[\ Engen\ S[\ Islam\ A[\ McCleery\ R[ + Perrins\C[ "0887# Environmental stochasticity and extinction riskin a population of a small songbird\ the great tit[ AmericanNaturalist\ 040\ 330Ð349[
S%ther\ B[!E[\ Engen\ S[ + Lande\ R[ "0888# Finite meta!populations models with density dependent migration andstochastic local dynamics[ Proceedings of the RoyalSociety] Biological Sciences\ 155\ 002Ð007[
Slagsvold\ T[ "0865# Annual and geographical variation inthe time of breeding of the Great Tit Parus major and thePied Flycatcher Ficedula hypoleuca in relation to environ!mental phenology and spring temperature[ Ornis Scan!dinavica\ 6\ 016Ð034[
Smith\ J[N[M[\ Taitt\ M[J[\ Rogers\ C[M[\ Arcese\ P[\ Keller\L[F[\ Cassidy\ A[L[E[V[ + Hochachka\ W[M[ "0885# Ametapopulation approach to the population biology of theSong Sparrow Melospiza melodia[ Ibis\ 027\ 019Ð017[
Stacey\ P[B[ + Taper\ M[ "0881# Environmental variationand the persistence of small populations[ Ecological Appli!cations\ 1\ 07Ð18[
Summers!Smith\ D[ "0877# The Sparrows[ T[ + A[D[ Poyser\Calton[
Thomas\ C[D[ + Hanski\ I[A[ "0886# Butter~y meta!populations[ Metapopulation Biology "eds I[ A[ Hanski +M[ E[ Gilpin#\ pp[ 248Ð275[ Academic Press\ San Diego[
Watkinson\ A[R[ + Sutherland\ W[J[ "0884# Sources\ sinksand pseudo!sinks[ Journal of Animal Ecology\ 53\ 015Ð029[
Received 5 February 0887^ revision received 05 September0887
Copyright © 2022 FDOKUMEN




















